蛋白质的合成加工
分泌蛋白的合成加工和运输过程

分泌蛋白的合成加工和运输过程
分泌蛋白的合成、加工和运输是一个复杂的过程,涉及多个细胞器和分子机制。
以下是一般的分泌蛋白合成加工和运输的过程概述:
1.合成过程:
o合成:分泌蛋白的合成发生在细胞的核内,由核糖体通过蛋白质合成过程进行。
合成的蛋白质称为前
蛋白。
o信号肽:在合成过程中,蛋白质序列中可能存在一个信号肽序列,该序列指示着该蛋白质是一个分泌
蛋白。
信号肽将帮助定位蛋白质到正确的位置。
2.加工过程:
o初始加工:在合成过程结束后,前蛋白将进入内质网(ER)。
在ER中,前蛋白将经历一系列的初始加
工步骤,包括信号肽的剪切和糖基化。
o终末加工:从ER中,蛋白质将进一步进入高尔基体,然后进入高尔基体的囊泡以进行成熟和终末加工。
该过程可能包括糖基化、脱糖基化、剪切等多种修
饰方式。
3.运输和存储过程:
o高尔基体到细胞膜:成熟的蛋白质囊泡从高尔基体进入细胞膜的分泌途径。
这些囊泡将与细胞膜融合,
释放蛋白质到细胞外。
o分泌颗粒:某些蛋白质可能在高尔基体中被包裹形成分泌颗粒,存储在细胞内。
这些颗粒在需要时,
可以通过融合细胞膜释放蛋白质。
4.分泌:
o定向分泌:某些蛋白质需要特定的信号序列来定向到特定的细胞膜区域,例如突触前膜和上皮细胞表
面。
o不定向分泌:其他蛋白质可能没有特定的定向信号,将被均匀地分泌到细胞膜上。
整个过程涉及到多个细胞器、蛋白质修饰和转运机制。
它的精细调控确保了分泌蛋白的准确合成和传递,使其可以发挥正常的功能。
蛋白质合成后的加工及转运

The signal-recog整n理it课io件n particle (SRP)
14
③转移通道:存在与内质网膜上的跨膜通道。
④。 SRP受体(SPR receptor),是膜的整合蛋白, 为异二聚体蛋白,存在于内质网上,可与SRP特异结合。
⑤停止转移序列(stop transfer sequence),肽链上的 一段特殊序列,与转移通道蛋白亲合力很高,能阻止肽 链继续进入内质网腔。
第五节 蛋白质合成后的加 工及转运
整理课件
1
本节内容:
一、蛋白质合成后的细胞定位;
二、蛋白质合成后的转运;
三、蛋白质合成后的加工及修饰;
整理课件
2
一、蛋白质合成后的细胞定位:
1、蛋白质是在细胞中游离的核糖体上或者是在糙面内 质网上的核糖体上合成的。
2、蛋白质合成后需要运转到特定的位点起作用:
(1)、内质网驻留蛋白、高尔基体驻留蛋白质、溶酶 体蛋白质、分泌蛋白质、膜蛋白等这些蛋白是由位于 糙面内质网上的核糖体合成的。然后进入内质网腔或 内质网膜。
输入内质网
-Leu-Ala-Leu-Lys-Leu-Ala-Gly-Leu-AspIle-
+H3N-Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-PheLys-Pro-Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-LeuLeu-
-Ser-Lys-Leu-COO-
整理课件
34
(四)、叶绿体的蛋白质转运
转运到基质的前体蛋白具有典型的N端序列。转运到 叶绿体内膜和类囊体膜的前体蛋白含有两个N端信号序 列,第一个被切除后,暴露出第二个信号序列,将蛋白 导向内膜或类囊体膜。
第十一章 蛋白质的生物合成及加工修饰

第十一章蛋白质的生物合成及加工修饰(The Biosynthesis and transport of protein)在不同的蛋白质分子中,氨基酸有着特定的排列顺序,这种特定的排列顺序不是随机的,而是严格按照蛋白质的编码基因中的碱基排列顺序决定的。
基因的遗传信息在转录过程中从DNA转移到mRNA,再由mRNA将这种遗传信息表达为蛋白质中氨基酸顺序的过程叫做翻译。
翻译的过程也就是蛋白质分子生物合成的过程,在此过程中需要200多种生物大分子参加,其中包括核糖体、mRNA、tRNA及多种蛋白质因子。
第一节参与蛋白质生物合成的物质一、合成原料自然界由mRNA编码的氨基酸共有20种,只有这些氨基酸能够作为蛋白质生物合成的直接原料。
某些蛋白质分子还含有羟脯氨酸、羟赖氨酸、γ-羧基谷氨酸等,这些特殊氨基酸是在肽链合成后的加工修饰过程中形成的。
下图给出部分特殊氨基酸分子式:二、mRNA是合成蛋白质的直接模板蛋白质是在胞质中合成的,而编码蛋白质的信息载体DNA 却在细胞核内,所以必定有一种中间物质用来传递DNA 上的信息,实验证明:mRNA 是遗传信息的传递者,是蛋白质生物合成过程中直接指令氨基酸掺入的模板,因此得名信使RNA。
原核细胞中每种mRNA分子常带有多个功能相关蛋白质的编码信息,以一种多顺反子的形式排列,在翻译过程中可同时合成几种蛋白质;而真核细胞中,每种mRNA一般只带有一种蛋白质编码信息,是单顺反子的形式。
mRNA以它分子中的核苷酸排列顺序携带从DNA传递来的遗传信息,作为蛋白质生物合成的直接模板,决定蛋白质分子中的氨基酸排列顺序。
不同的蛋白质有各自不同的mRNA,mRNA除含有编码区外,两端还有非编码区。
非编码区对于mRNA的模板活性是必需的,特别是5'端非编码区在蛋白质合成中被认为是与核糖体结合的部位。
mRNA特点:短命原核:半衰期几秒-几分钟真核:半衰期数小时。
功能是蛋白质合成的模板,多肽链氨基酸排列顺序就取决于mRNA 的核苷酸的排列顺序。
蛋白质加工和合成的过程

蛋白质加工和合成的过程
蛋白质合成的过程:在细胞核中进行转录在核糖体上进行翻译内质网加工高尔基体再加工。
所需的细胞器有:细胞核、核糖体、内质网、高尔基体、线粒体。
1、转录过程以四种核糖核苷酸为原料,以DNA分子的一条链为模板,在DNA解旋酶、RNA 聚合酶的作用下消耗能量,合成RNA,在细胞核中进行;
2、翻译过程以氨基酸为原料,以转录过程产生的mRNA为模板,在酶的作用下,消耗能量产生多肽链,多肽链经过折叠加工后形成具有特定功能的蛋白质,在细胞质的核糖体上进行;
3、内质网进行粗加工;
4、高尔基体进行再加工形成成熟的蛋白质。
过程中需要线粒体提供能量。
【创新设计】高中生物 212蛋白质的合成与运输课件 中图必修1

2.核糖体 (1)功能:核糖体是 蛋白质 的合成场所。 (2)分布:无论 真核 细胞还是 原核 细胞,甚至 线粒体 和
叶绿体 中,都有核糖体存在。 (3)成分:由 核糖核酸(RNA) 和 蛋白质 组成。 (4)结构:由 大亚基 和 小亚基 组成,蛋白质合成时,大、 小亚基结合在一起,组成核糖体。
3.修饰加工 (1)场所:内质网、 高尔基体 和 细胞质基质 。 (2)含义:为新生肽链添加上 糖链 、 甲基 或 羟基 并对 其 剪切和折叠 等。 (3)意义:对维持和调节 蛋白质 活动、进而成为 有功能 的蛋白 质有重要意义。 [思维激活1] 参与分泌蛋白的合成与分泌过程的细胞结构有哪些? 提示 核糖体、内质网、高尔基体、线粒体、细胞膜。
(3)研究的手段为同位素标记法。
1.下列物质中属于分泌蛋白的是( ) ①胃蛋白酶 ②抗体 ③氧化酶 ④性激素 ⑤胰岛素 A.①②③ B.②③④ C.③④⑤ D.①②⑤ 解析 氧化酶是胞内蛋白,性激素是脂质类物质,不是蛋白质。 答案 D
2.下列关于蛋白质的修饰的叙述中,不正确的是( ) A.蛋白质在加工与修饰前,已具有一定的活性 B.内质网、高尔基体和细胞质基质均能对新生肽链进行加 工修饰 C.蛋白质的修饰主要是指为新生肽链添加上糖链、甲基或 者羟基并对其剪切和折叠等 D.蛋白质的修饰对维持和调节蛋白质的活性,进而成为有 功能的蛋白质有着重要意义 解析 多肽链在核糖体上合成后,必须经过内质网、高尔基 体加工修饰才能形成空间结构,成为有一定活性的蛋白质。ቤተ መጻሕፍቲ ባይዱ答案 A
答案 (1)1线粒体、6叶绿体 (2)放射性同位素标记法 (3)胰液(消化酶)(或胰岛素或胰高血糖素) (4)内质网膜和高尔基体膜、高尔基体膜和细胞膜 (5)合成和分泌油脂不需要核糖体参与
蛋白质的合成与运输

)
【解析】 内质网是脂质合成的场所,所以合成固醇类激素的分泌细胞内质网 应该比较发达。分泌蛋白的修饰和加工是由内质网和高尔基体共同完成。生物膜之 间可通过具膜小泡的转移进行相互转化。
【答案】 B
5.如图表示用含 3H 标记的氨基酸培养液培养某细胞过程中蛋白质的合成和分泌 示意图,则该细胞中出现 3H 的部位依次为( )
【答案】 B
2.为了研究酵母菌细胞内蛋白质的合成, 研究人员在其培养基中添加 3H 标记的 亮氨酸后,测得与合成和分泌乳蛋白相关的一些细胞器上放射性强度的变化曲线如 下图中甲,有关的生物膜面积变化如图乙,其相关结构关系如图丙,则下列有关说 法不正确的是( )
A.图丙中首先可观察到 3H 标记的细胞器是③ B.能在图丙中④上观察到 3H 标记表明可能有分泌蛋白合成 C.图甲中 c 曲线所指的细胞结构是高尔基体 D.图乙中 d 曲线表示的细胞结构是内质网
提示:原核细胞除细胞膜外无其他膜结构,因此没有内膜系统。
[思维升华] 生物膜的种类、结构和功能联系
细胞膜:单层膜,细胞的外界膜 核膜:双层膜,具核孔,细胞核的外界膜 生物膜 双层膜结构:叶绿体、线粒体 (1) 的种类 细胞器膜单层膜结构:内质网、高尔基 体、液泡等
1.关于蛋白质的分选和运输,下列说法不正确的是( A.蛋白质合成后,一般在其氨基酸序列中含有分选信号 B.游离的核糖体合成的蛋白质中无分选信号
)
C.核糖体在细胞中的存在部位不同,也影响蛋白质的去向 D.蛋白质只有被准确地运输到相应的部位才能执行特定的功能
【解析】 无论是在内质网的核糖体上还是游离的核糖体上合成的蛋白质,一 般其氨基酸序列中均含分选信号,决定着蛋白质的去向和最终定位,而核糖体在细 胞中的存在部位不同,也影响蛋白质的去向。
蛋白质的合成过程

6、蛋白质激酶参与真核细胞蛋白质合成的调节:在真核细胞中,蛋白质激酶可以催 化起始因子eIF2的磷酸化。而eIF2的作用是将Met-tRNAMet运送至40S核糖体亚基上, eIF2被磷酸化后就难以再投入下一轮的起始作用。所以蛋白质合成受到抑制。若使其 恢复其功能,必须解除其磷酸化,这由专一的磷酸酶来催化。
30S复合物形成:
AUG
小亚基
AUG
IF3
IF3
fMet
IF3
GTP、IF1、 IF2 fMet-tRNAf
5
UAC AUG
GTP、IF1、IF2
70S复合物的形成:
A位点
fMet
5
P位点
fMet
5
UAC AUG
GTP、IF1、IF2
UAC AUG
+ 50S核糖体
GTP、IF1、IF2
GDP+Pi、IF1、IF2
和释放、肽链的折叠和加工处理。
(一)氨基酸的活化与转运
氨基酸在掺入肽链之前必须活化(activition)以获得额外的能量。活化反应是在 氨酰-tRNA合成酶(aminoacyl-tRNA synthetase)催化下进行的。活化了的氨基酸与 tRNA形成氨酰-tRNA。这一反应可在可溶性细胞质内完成。活化反应分两步进行: 1、氨基酸-AMP-酶复合物的形成:反应如下: ATP+氨基酸+酶——→氨基酸-AMP-酶+PPi 反应需要Mg2+或Mn2+,并且是ATP水解释放能量供复合物的形成。在复合物中,氨基 酸的羧基通过酸酐键与AMP上的5’-磷酸基相连接,形成高能酸酐键,从而使氨基酸的 羧基得到活化。 2、氨基酸从复合物上面转移到相应的tRNA上面: 氨基酸-AMP-酶+PPi ——→氨酰-tRNA+AMP+酶
第8章 蛋白质生物合成和翻译后加工

第8章蛋白质生物合成和翻译后加工一、学习要点本章涉及的基本概念较多,如翻译、核酶、翻译的起始因子、延长因子、释放因子、三联体密码、密码的简并性、密码的摆动性、SD序列、信号肽等。
主要理解和掌握蛋白质合成的分子基础、遗传密码的基本特性、多肽链生物合成的一般过程、确保翻译准确性的机制、翻译后加工和定向运输等内容。
1、蛋白质合成的分子基础:与蛋白质合成有关的RNA有:mRNA、tRNA和rRNA。
mRNA 作为蛋白质多肽链合成的直接模板。
tRNA在蛋白质合成主要起接头作用,① tRNA的 3′端- CCA序列是氨基酸接受位点(氨基酸接受臂);②DHU环是识别氨酰tRNA合成酶的位点;③TψC环是核糖体的识别位点;④反密码子环上的反密码子作用是mRNA的密码子。
rRNA主要是与蛋白质结合形成核糖体,作为蛋白质合成的场所。
2、遗传密码。
mRNA上决定一个特定氨基酸的三个相邻的核苷酸称为遗传密码.又叫三联体密码或密码子。
其特点主要有:1)无标点符号的;2)简并性(degeneracy),除Met (AUG )和Trp(UGG )外,每个氨基酸都有一个以上的密码子,这种现象称为密码的简并,把编码相同氨基酸的密码子称同义密码子。
其生物学意义在于减少有害突变,对生物物种的稳定有一定作用。
3) 遗传密码具有的变偶性(摆动性),密码子第1位、第2位碱基配对是严格的,第3位碱基可以有一定的变动,这一现象称为变偶性或摆动性(wobble)。
4)64组密码子中61组编码氨基酸,UAA、UAG和UGA为终止密码子(或无义密码子), AUG为起始密码子。
5)遗传密码的通用性和变异性,通用性是指各种低等和高等生物,包括病毒、原核以及真核生物,基本上共用一套遗传密码,反应了生物世界的统一性;目前已知线粒DNA(mtDNA)的编码方式与通常遗传密码子有所不同。
3 多肽链合成的机理。
(1)多肽链的延伸方向。
在翻译时,核糖体沿mRNA从5’→3’方向进行翻译,多肽链延伸的方向为氨基端→羧基端(与多肽的人工合成不同,在多肽的人工合成中,肽链从羧基端→氨基端)。
第八章:蛋白质的合成与加工

3. fMet-tRNAfMet被起始因子IF2和一分子的GTP一同带到小亚基 的P位
密码虽有简并性,但在特定的物种中,基因并非平均地使用 每个密码子,这种现象称为密码子偏倚(codon bias)。 如亮氨酸可由6个密码子编码(TTA、TTG、CTT、CTC、 CTA和CTG)。但是在人类基因中,大多被CTG编码,而且 几乎不被TTA或CTA编码。密码子偏倚的生物学原因并不清 楚,但不同种属的生物有不同的偏好。
(3)密码子具有普遍性
高等生物和低等生物在很大程度上共用一套密码子,说明核 酸中的核苷酸序列与蛋白质中的氨基酸序列之间对应关系的 遗传密码在进化的早期就已确定下来了。但是,也存在一些 例外的情况。
例外1:在支原体中,终止密码子UGA被用来编码Trp,而 UGG仍是Trp的密码子,但用得很少。
ቤተ መጻሕፍቲ ባይዱ
例外2:在一些纤毛虫(单细胞原生动物)种,终止密码子 UAA、UAG被用来编码谷氨酰胺。
2. 核糖体小亚基与IF3附着于mRNA上的核糖体结合位点上
30S亚基在mRNA上的结合位置,正好使起始密码子置于30S 亚基P位上。而在30S亚基的A位上结合有IF1,其作用是防止 tRNA在翻译的起始阶段结合到A位上
IF3在上一轮蛋白质合成即将结束时开始与小亚基结合,并 协助70S核糖体解离成大、小亚基。由此可看出,IF3具有双 重功能:一方面它与30S亚基结合后,阻止30S亚基与50S亚 基重新结合成70S核糖体,使30S亚基处于游离状态;另一 方面它还能辅助30S亚基与mRNA结合,30S亚基也必须有 IF3的参与才可能与mRNA形成复合体。
7.2.2 tRNA的三级结构
通过X-衍射来研究几种 酵母的tRNA晶体,发现 各种tRNA 存在共同的三 维结构。每一tRNA的三 叶草型二级结构都要折叠 成致密的倒L型三级结构。
第二章 蛋白质的合成、转运、加工与修饰

顺反子: 顺反子 : 编码一种多肽链并连同起始信号和终止 信号在内的DNA区段。 区段。 信号在内的 区段 单顺反子mRNA:编码一种多肽链的mRNA分子。 :编码一种多肽链的 分子。 单顺反子 分子 多顺反子mRNA: 编码数种不同多肽链的同一条 : 多顺反子 mRNA分子。多见于原核生物。 分子。 分子 多见于原核生物。 反义链/有意义链 ( ) 模板链 双链DNA分子中 模板链: 反义链 有意义链/(-)链/模板链:双链 有意义链 分子中 被转录成RNA转录本的链。 转录本的链。 被转录成 转录本的链 正义链/无意义链 ( ) 正义链 无意义链/(+)链 无意义链
(S) )
SD 序 列 / 核 糖 体 结 合 位 点 ( ribosomal binding site , RBS) : 原核细胞 的翻译起始密码子AUG的上游 ) 原核细胞mRNA的翻译起始密码子 的翻译起始密码子 的上游 相距8~ 个核苷酸处有一段由 个核苷酸处有一段由4~ 个核苷酸组成的富含 相距 ~13个核苷酸处有一段由 ~6个核苷酸组成的富含 嘌呤的序列, 为核心, 嘌呤的序列 , 以 5’-AGGA-3’为核心,它与核糖体小亚基 为核心 上的16S-rRNA 的近 末端处的一段短序列互补。 的近3’末端处的一段短序列互补 末端处的一段短序列互补。 上的 Kozak序列 Kozak序列:a favorable context for efficient eukaryotic 序列: translation initiation(PuNNATGPu)。(S) ( ) ) 典型的Poly(A)加尾信号:AATAAA。(S) 加尾信号: 典型的 加尾信号 。 ) cDNA 末 端 快 速 扩 增 法 ( rapid amplification of cDNA ends, RACE)(S) , ) )
蛋白质合成,加工与运输

• 示例:类囊体膜蛋白 Lhcb1
① 前体Lhcb1(在胞质中) 外膜
转运肽,cpRSP54
跨叶绿体内
Hsp70, GTP
② 蛋白N-端进入基质,转运肽即被水解
③ 第一个滞留片段(-Glu-X-X-His-X-Arg-)使Lhcb1停留 在
类囊体膜上。其中的His和Glu/Ar叶g对绿与体叶绿素和叶黄
TOM TIM23
白进入内膜或膜间隙
跨膜后被切除,含 疏水性的停止转移 序列,蛋白被安插 到内膜。
内 TOM 膜 TIM23
结构类似于N端信号
序列,但位于蛋白 内 TOM
质内部。
膜 TIM23
为线粒体代谢物的 转运蛋白,如腺苷 转位酶,具有多个 内部信号序列和停 止转移序列,形成 多次跨膜蛋白。
内 TOM 膜 TIM22
4. C-端与结构蛋白相连部位为富含Ala的片段,易于形成 -sheet,是信号肽酶的识别和切割位点。
5. 信号肽不一定位于蛋白的N-末端。如卵清蛋白的信号 肽
位于中部。 6. 某些膜蛋白的信号肽在跨膜之后不被水解掉。Cyt P45
2. 信号肽引导的蛋白跨内质网膜过程:
• 属于边翻译边运输过程:识别 停泊 跨膜 水解
二 激素与激素原:
1 概念:如胰岛素原( 81aa)
类胰蛋白酶 切除C肽(30aa)
类羧肽酶B
Arg 60
成熟胰岛素(51aa) Lys59
A肽
COOH
• 原肽(propeptide): 其两侧含有成对碱性aa。 C肽
• 含原肽的蛋白叫原蛋白
2 原肽的功能:
H2N B肽
Arg 32Arg 31
《蛋白质合成后加工》PPT课件

现以哺乳动物的胰岛素为例说明这种分泌过程。胰岛素由 51个氨基酸残基组成,但胰岛素mRNA的翻译产和在兔网织红细 胞无细胞翻译体系中为86个氨基酸残基,称为胰岛素原,在麦 胚无细胞翻译系统中为110个氨基酸残基组成的前胰岛素原, 后来证明,在前胰岛素原的N末端有一段富含疏水氨基酸的肽 段做为信号肽,使前胰岛素原能穿越内质网膜进入内质网内腔, 在内腔壁上信号肽被水解。所以在哺乳动物细胞内,当多肽链 合成完成时,前胰岛素原已成为胰岛素原。然后胰岛素原被运 到高尔基复合体,切去C肽成为成熟的胰岛素,最终排出胞外。 像真核细胞的前清蛋白,免疫球白轻链,催乳素等都有相似的 分必方式 。
切除 连接
图2-1 蛋白质自我剪接
1994年,Perler等将蛋白质自我剪接中被切除的肽段命名 为蛋白内含肽(intein),也称蛋白内含子或内蛋白子,将位 于蛋白内含肽两侧、切除部位再连接形成的产生蛋白称为蛋白 外显肽(estein),又称蛋白外显子或外蛋白子。
蛋白内含肽的基因不是单独的开放阅读框,它插入在蛋白 外显肽的基因中,和内含子的区别在于它可以和蛋白外显肽的 基因一起表达,而不是在mRNA阶段被切除。
某些蛋白氨基端有一段15~30个氨基酸残基的顺序,这段顺 序称信号肽,与蛋白质传送到特定的细胞器、膜,或分泌出细胞 外有关。信号肽在运送过程中通常由专一的酶催化而水解除去。
2.氨基酸残基的修饰 (1)甲基化修饰
某些蛋白质中的赖氨酸残基需要甲基化,某些谷氨酸残基 的羧基也要甲基化,以除去负电荷。 (2)羧化修饰
2.真核生物蛋白质分泌
真核生物不但有细胞核、细胞质和细胞膜,而且还有许多膜 性结构的细胞器,在细胞须内合成的蛋白质怎样的到达细胞的不 同部位呢?了解比较清楚的是分泌性蛋白质的转运。
蛋白质的加工

protein through.
翻译后易位
氨基酸侧链修饰作用包括磷酸化(如核糖体蛋白质)、糖基化(如各种糖蛋白)、甲基化(如组蛋白、肌肉蛋白质)、乙基化(如组蛋白)、羟
基化(如胶原蛋白)和羧基化等。
2 二硫键形成
1.
3 信号肽与蛋白质转运的关系
蛋白质插入或穿过生物膜的过程称为蛋白质易位(protein translocation)。
点最近那个氨基酸常带很短侧链(Ala或Gly )
An N-terminal signal sequence is hydrophobic
The signal sequence of bovine growth hormone consists of the N-terminal amino acids and has a central highly hydrophobic region, preceded or flanked by regions containing polar amino acids.
3 特定氨基酸化学修饰
1.
Proteins enter the nucleus by passage through very large nuclear pores.
The leader sequence of yeast cytochrome c ⑩蛋白质有序降解机理。
Proteins can enter the ER-Golgi pathway only by associating with the endoplasmic reticulum while they are being
2 蛋白质易位
2.1 概 述 2.2 共翻译易位 2.3 翻译ck
2.1 概 述
第五节蛋白质合成后的加工及转运课件

在蛋白质合成过程中,信号识别颗粒与新生蛋白质结合,引导蛋白 质向内质网定位。
作用
确保新生蛋白质正确地转运到内质网,进行进一步的加工和修饰。
跨膜运
跨膜运输
是指蛋白质通过生物膜的运输过 程,包括通过细胞膜、线粒体膜 、叶绿体膜等。
运输方式
包括主动运输和被动运输,其中 主动运输需要消耗能量,而被动 运输则不需要。
多肽链在核糖体上合成的同时,通过 信号肽的引导,进入内质网腔或跨膜 转运至高尔基体、溶酶体和细胞骨架 。
03
膜泡运输
通过形成囊泡的方式,将已经折叠好 的蛋白质从一个膜结构转运到另一个 膜结构。例如,从内质网到高尔基体 ,或从高尔基体到溶酶体。
PART 04
蛋白质合成后的加工和转 运的调节
蛋白质合成后的加工的调节
蛋白质的乙酰化
在蛋白质合成后,某些赖氨酸残基可被乙酰化,从而调节 蛋白质的稳定性。这一过程由乙酰转移酶催化。
蛋白质转运的调节
01
核孔复合体的调节
核孔复合体是细胞核膜上的转运孔道,可选择性地将蛋白质从细胞质转
运到细胞核内或从细胞核转运到细胞质。核孔复合体的转运活性受到多
种因素的调节。
02
囊泡转运的调节
2023-2026
ONE
KEEP VIEW
蛋白质合成后的加工 及转运课件
REPORTING
CATALOGUE
目 录
• 蛋白质合成后的加工 • 蛋白质的转运 • 蛋白质分选的信号和途径 • 蛋白质合成后的加工和转运的调节 • 蛋白质合成后的加工及转运异常与疾病的关系
PART 01
蛋白质合成后的加工
蛋白质二硫键的形成
在蛋白质合成后,某些氨基酸残基需要经过氧化形成二硫 键,以稳定蛋白质的高级结构。这一过程由特异的二硫键 异构酶催化。
蛋白质加工——精选推荐
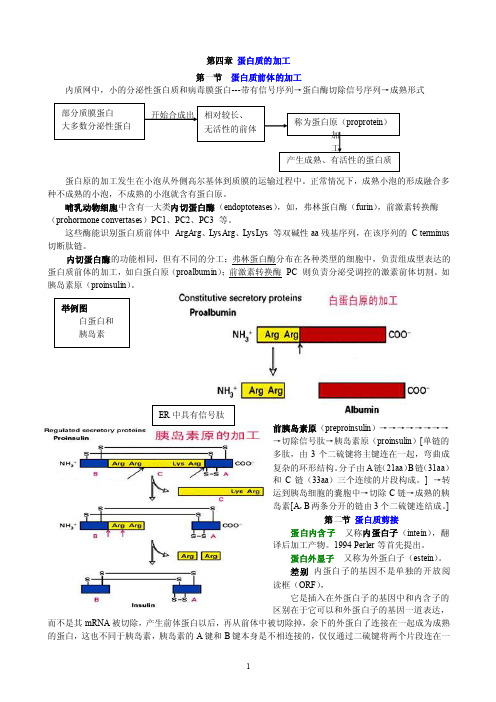
第四章 蛋白质的加工第一节 蛋白质前体的加工内质网中,小的分泌性蛋白质和病毒膜蛋白---带有信号序列→蛋白酶切除信号序列→成熟形式蛋白原的加工发生在小泡从外侧高尔基体到质膜的运输过程中。
正常情况下,成熟小泡的形成融合多种不成熟的小泡,不成熟的小泡就含有蛋白原。
哺乳动物细胞中含有一大类内切蛋白酶(endoptoteases ),如,弗林蛋白酶(furin),前激素转换酶(prohormone convertases )PC1、PC2、PC3 等。
这些酶能识别蛋白质前体中 ArgArg 、LysArg 、LysLys 等双碱性aa 残基序列,在该序列的 C terminus 切断肽链。
内切蛋白酶的功能相同,但有不同的分工:弗林蛋白酶分布在各种类型的细胞中,负责组成型表达的蛋白质前体的加工,如白蛋白原(proalbumin );前激素转换酶 PC 则负责分泌受调控的激素前体切割。
如胰岛素原(proinsulin )。
前胰岛素原(preproinsulin )→→→→→→→→→切除信号肽→胰岛素原(proinsulin )[单链的多肽,由3个二硫键将主键连在一起,弯曲成复杂的环形结构。
分子由A 链(21aa )B 链(31aa )和C 链(33aa )三个连续的片段构成。
] →转运到胰岛细胞的囊胞中→切除C 链→成熟的胰岛素[A ,B 两条分开的链由3个二硫键连结成。
]第二节 蛋白质剪接 蛋白内含子 又称内蛋白子(intein ),翻译后加工产物。
1994 Perler 等首先提出。
蛋白外显子 又称为外蛋白子(estein )。
差别 内蛋白子的基因不是单独的开放阅读框(ORF ),它是插入在外蛋白子的基因中和内含子的区别在于它可以和外蛋白子的基因一道表达,而不是其mRNA 被切除,产生前体蛋白以后,再从前体中被切除掉,余下的外蛋白了连接在一起成为成熟的蛋白,这也不同于胰岛素,胰岛素的A 键和B 键本身是不相连接的,仅仅通过二硫键将两个片段连在一起。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
综述细胞内的蛋白质合成、加工、修饰、分选与运输方式及其生物学意义。
蛋白质是生命活动的主要承担者,是构成细胞和生物体结构的重要物质,在生物体及细胞的生命活动中发挥重大作用。
1.许多蛋白质是构成细胞和生物体结构的重要物质,称为结构蛋白。
2.细胞内的化学反应离不开酶得催化,绝大多数酶都是蛋白质。
3.有些蛋白质具有运输载体的功能。
(血红蛋白运输氧)
4.有些蛋白质起信息传递的作用,能够调节机体的生命活动。
(如,胰岛素)
5.有些蛋白质有免疫功能,人体的抗体是蛋白质,可以帮助人体抵御病菌和病毒等抗原的侵害。
1 蛋白质的合成
蛋白质的生物合成过程实质上是基因表达的一个过程,它包括转录和翻译。
即把mRNA 分子中的碱基排列顺序转变为蛋白质或多肽链中的氨基酸排列顺序的过程,可分为起始、延长和终止3个阶段,分别由不同的起始因子、延伸因子和终止因子(释放因子)参与。
细胞中的蛋白质都是在核糖体上合成的,并都是起始于细胞质基质之中。
2 蛋白质的加工与修饰
许多新生肽要经过一种或几种共价键修饰,这种修饰可以在正延伸着的肽链中进行。
一般情况下,翻译后修饰一是为了功能上的需要,另一种情况是折叠成天然构象的需要。
在粗面内质网合成并进入内质网腔的蛋白质发生的主要化学修饰作用有糖基化、羟基化、酰基化和二硫键的形成。
而在细胞质基质中发生蛋白质修饰的类型主要有辅酶或辅基与酶的共价结合、磷酸化和去磷酸化、糖基化、甲基化、酰基化等。
蛋白质的修饰加工主要包括:
切除加工:包括切除N-端甲硫氨酸、信号肽序列和切除部分肽段,将无活性的前体转变成活性形式。
(包含信号肽的胰岛素前体称为前胰岛素原,去掉信号肽的胰岛素的前体称为胰岛素原),进一步切除称为C链的肽段后才能形成活性形式的胰岛素)
糖基化:糖基化主要发生在内质网和高尔基体中。
粗面内质网上合成的大多数蛋白在都发生了糖基化。
主要作用是促进蛋白质在成熟过程中折叠成正确构象,增加蛋白质的稳定性,有N-连接的糖基化和O-连接的糖基化之分。
(重点)
羟基化:最常见的是内质网上合成的跨膜蛋白在通过内质网和高尔基体的转运过程中发生的,它由不同的酶来催化,把软脂酸链共价地连接在某些跨膜蛋白的暴露在细胞质基质中的结构域。
磷酸化与去磷酸化:蛋白磷酸化与去磷酸化参与代谢调控和信号转导以及蛋白与蛋白之间的相互作用。
(PDGF受体的酪氨酸残基经过自身磷酸化后才与细胞质定位蛋白质结合。
)
亲脂修饰:最常见的亲脂修饰是酰化和异戊二烯化。
蛋白质亲脂修饰后可以改变膜结合能力和特定的蛋白与蛋白之间的相互作用。
N-豆蔻酰化(豆蔻酸以酰酰氨键形式共价连在肽链N 端的残基上)能增加特定G蛋白的α亚基对膜结合的β、γ亚基的亲和力。
甲基化:通过甲基转移酶进行。
天冬氨酸的甲基化能促进已破坏蛋白的修复或降解,在2,3-二磷酸核酮糖羧化酶(rihilose-2,3-biosphosphate carboxylase)、钙调蛋白(calmodulin)、组氨酸(histone)、某些核糖体蛋白和细胞色素C中都有甲基化的赖氨酸残基。
二硫键形成:二硫键通常只发现于分泌蛋白(如胰岛素)和某些膜蛋白中,在细胞质中由于有各种还原性物质,所以细胞质蛋白没有二硫键。
因为内质网腔是一个非还原性环境,所以粗糙内质网上的新生肽只暂时形成二硫键。
当新生肽进入内质网腔时,一些肽链可能会按氨基酸次序依次暂时形成二硫键,但最终会通过交换二硫键位置的形式形成正确的结构,内质网中可能还有一种二硫键异构酶催化该过程。
3 蛋白质的分选和转运
绝大多数的蛋白都是在细胞质基质游离核糖体上或者粗面内质网膜结合核糖体上合成的。
由于蛋白质发挥结构或功能作用的部位几乎遍布细胞各个膜区或组分,因此必然存在不同的机制确保蛋白质分选、转运至细胞的特定部位。
通过不同的途径转运到细胞的特定部位成为蛋白质分选或定向转运。
信号假说的确认为细胞内蛋白质的定向转运研究指明了方向,除了信号肽外,人们发现了一系列蛋白质分选序列,统称信号序列以及信号斑,而根据信号序列运输方向的不同分为三种类型,即入核信号、引导肽和信号肽。
入核信号指导核蛋白的运输,引导肽指导线粒体、叶绿体和过氧化物酶体蛋白的运输,信号肽则指导内膜系统的蛋白质运输。
蛋白质的分选大体可分为:
1 翻译后转运途径:也就是合成后转运。
在细胞质基质游离核糖体上完成多肽链的合成后,其中一些蛋白质不带分选信号,就留在细胞质基质中,成为可溶性驻留蛋白和支架蛋白,构成自身的结构成分;带有分选信号的蛋白质,按期分选信号的种类被分别转运到细胞的不同部位,如细胞核、线粒体、过氧化物酶体等。
2 共翻译转运途径:也就是边合成边转运。
蛋白质在细胞质基质的游离核糖体上开始合成后,由信号肽引导转移至粗面内质网,即在核糖体上多肽链开始合成不久,在N-末端形成的信号肽引导核糖体附着到内质网膜上,信号肽传入内质网腔并继续其合成,新合成的多肽链可游离于内质网腔成为可溶性蛋白,也可插入内质网膜成为跨膜蛋白。
以这种方式合成的蛋白质除一部分留在内质网外,大部分经高尔基体加工包装运至溶酶体、细胞质膜或分泌到细胞外。
(重点)
根据蛋白质分选的转运方式或机制看,蛋白质分选的转运可分为蛋白质的跨膜转运、门控转运和膜泡运输以及细胞质基质中的蛋白转运等。
其分选指令存在于多肽自身。
其中蛋白质在细胞质基质与细胞核之间的运输通过核孔复合体进行,属于门控运输。
进入细胞核的蛋白质是细胞核包括染色质的结构基础之一,并且作为一些调控因子和酶参与细胞核内基因转录等活动。
蛋白质从细胞质基质进入内质网、线粒体和过氧化物酶体等有膜的细胞器是通过跨膜转运的方式进行的。
进入内质网的蛋白质可分为(1)向细胞外分泌的蛋白质,包括胰腺细胞分泌的酶、浆细胞分泌的抗体。
小肠被细胞分泌的黏蛋白。
内分泌腺分泌的多肽类激素和胞外基质成分等。
这类蛋白质常以分泌泡的形式通过细胞的胞吐作用运输到细胞外。
(2)膜整合蛋白,构成细胞质膜上的膜蛋白及内质网、高尔基体和溶酶体膜上的膜蛋白。
(3)构成内膜系统细胞其中的可溶性驻留蛋白。
有些驻留蛋白需要与其他细胞组分严格隔离,如溶酶体与植物液泡中的酸性水解酶类,内质网、高尔基体和胞内体中固有的蛋白质以及其他有重要生物活性的蛋白质,在合成后进入内质网,便于与其他细胞组分进一步区分,也有利于对它们的加工与活化。
进入线粒体的蛋白,作为结构蛋白参与线粒体内外膜的构成,作为各种氧化酶、代谢酶参与线粒体的各项生物功能,包括氧化代谢、呼吸链中电子的传递和质子的转移以及氧化磷酸化过程。
进入外膜系统的蛋白,除了参与生物膜的基本构建以为,构成了细胞膜上的各种生物泵,参与物质运输。
同时,细胞膜、核膜等受体蛋白,参与着生物的信号传导过程。
蛋白质通过不同类型的转运小泡从粗面内质网合成部位转运至高尔基体,进而分选转运到细胞的不同部位,是膜泡运输的典型代表。