蛟河林业局林分生长量(率)的变化研究
吉林蛟河不同演替阶段针阔混交林土壤健康评价1)

吉林蛟河不同演替阶段针阔混交林土壤健康评价1)吴金卓;蔡小溪;林文树【摘要】应用层次分析法、隶属度函数和森林土壤健康指数法,对吉林省蛟河市林业实验区管理局内4种不同演替阶段的森林土壤健康进行评价;主要选取5个物理性指标、8个化学性指标、1个生物指标建立评价体系,使用隶属度函数进行无量纲化处理,应用层次分析法求出各指标的权重值,得出森林土壤的健康指数。
结果表明:森林土壤健康等级降幂排序为中龄林、成熟林、老龄林、近熟林;相关性分析可知,土壤健康指数,与全氮、有效磷、速效钾呈极显著相关性,与水解氮、有机质呈显著相关。
%We used analytical hierarchy process , standard scoring functions and forest soil health index to evaluate the health of forest soil at four sucessional stages in Jiaohe Management Bureau of Forestry Experimental Area in Jilin Province .We se-lected five indexes of physical properties , eight indexes of chemical properties and one biological indicator to establish the assessment indicator system .The standard scoring functions were used to process dimensionless , and the analytical hierar-chy process was used to calculate the index weight , and finally the health index of forest soil was obtained .The descending orderof forest soil health level was middle-aged forest, mature forest, old-growth forest, and near-mature forest.By the correlation analysis , the correlations between health index of forest soil and total nitrogen , available phosphorus , and avail-able potassium were highly significant , and health index was significantly correlated with available nitrogen and organic matter.【期刊名称】《东北林业大学学报》【年(卷),期】2015(000)006【总页数】5页(P78-82)【关键词】针阔混交林;森林土壤;土壤健康指数;隶属度函数;层次分析法【作者】吴金卓;蔡小溪;林文树【作者单位】东北林业大学,哈尔滨,150040;东北林业大学,哈尔滨,150040;东北林业大学,哈尔滨,150040【正文语种】中文【中图分类】S7142针对森林生长与结构特征,利用合适的土壤健康评价指标对森林土壤进行调查与评价,在不断演变的情况下探索出促进森林土壤健康发展的管理措施与方向,最终实现森林健康的目标,是全世界林业工作者面临的新课题[1]。
加强林业资源管理 实现林区可持续发展

L i n y e y u a n y i加强林业资源管理实现林区可持续发展高圣超当前,在我国一些地区存在着林业资源过度开发的现象,这使得这些区域的土地沙漠化现象较为严重。
基于此,我国相关职能部门需要加强林业资源的管理,要采取一定的措施,以实现林区的可持续发展。
一、我国林业资源管理中所存在的主要问题分析1、林权改革难度较大在十八大之后,党和政府进一步明确了要对农村集体林权制度进行改革。
一方面需要将集体的林地经营权交给农户,另一方面要将林木的所有权交给农户。
此外,要确立农民的主体地位,以便将集体林权制度改革落实下去。
但是,在改革发展的过程中,因为技术以及资金的相对缺乏,再加上一些林权过度集中以及其他方面的问题,导致林业的经营发展受到了影响。
因此,农民育林的积极性受到了较大打击,这影响了我国林业的良好发展。
2、补偿制度不够规范为了推动我国林业的发展,我国政府部门制定了相应的补偿制度,例如生态公益林补偿制度。
而补偿资金主要有以下几种用途:(1)用于公益林专职管护人员的劳务费;(2)用于集体或者个人的补偿费用;(3)用于管护区的补植苗木费;(4)用于林木抚育费等等。
在我国林业的发展中,生态公益林补偿制度在其中发挥着巨大的作用。
但是,我国一些地区的补偿资金存在着管理不够规范的现象,这影响了林区的可持续发展。
综合来看,生态公益林补偿有以下几方面的影响:首先,因为补偿政策的宣传力度不足,使得许多林农缺乏补偿意识。
其次,因为补偿资金较少,所以很难满足农户的期望,从而打击了他们的积极性。
3、砍伐现象较为普遍当前,在我国一些地区,存在着乱砍滥伐的现象。
众所周知,乱砍滥伐会破坏当地的生态平衡,还影响森林资源的长远发展。
因此,林业管理部门需要加强对林地用途的管理,要杜绝乱砍滥伐的现象发生。
二、林业资源的管理策略分析1、加强对生态系统的保护要想加强对林业资源的管理,则需要我国林业部门需要进行生态系统的保护,要尽可能提高当地的森林覆盖率。
吉林蛟河针阔混交林树木生长的空间关联格局

吉林蛟河针阔混交林树木生长的空间关联格局郝珉辉;张忠辉;赵珊珊;张春雨;赵秀海【摘要】以吉林蛟河21.12hm2(660m×320m)针阔混交林样地为对象,利用2009年和2014年森林生长观测数据,研究树木生长的空间自相关格局及其生境影响机制.在样地生境型划分结果的基础上,采用Ripley's L(r)函数分析不同生境型中树木种群空间分布特征;利用标记相关函数分析不同生境型中树木生长特征的空间关联格局.研究结果表明:(1)红松(生境型3∶1-5m)、蒙古栎(生境型3:1-3m)、胡桃楸(生境型2:1-2m;生境型3:1-7m)、黄檗(生境型2∶1-3m;生境型4:1-5m)、水曲柳(生境型3∶1-2m;生境型4:1-2m)、瘤枝卫矛(生境型2∶1-15m)在特定生境和空间尺度上呈随机分布,但空间格局仍以聚集性分布为主;其余10个物种则在全部0-30m尺度上呈聚集分布.(2)标记相关函数分析显示春榆、毛榛、色木槭、瘤枝卫矛和千金榆的径向生长至少在一个生境中表现出正相关格局;暴马丁香、胡桃楸、裂叶榆、瘤枝卫矛、水曲柳、紫椴、糠椴、毛榛、色木槭和白牛槭的径向生长至少在一个生境中表现出负相关格局;红松、黄檗、蒙古栎和簇毛槭的径向生长在全部尺度上均未检测到显著的空间关联格局.因此,不同树种径向生长的空间自相关特征不同,树种生长特征的空间关联格局具有明显的生境依赖性.%This study aimed to determine the spatial autocorrelation patterns of tree growth and the mechanisms influencing habitat heterogeneity.A dataset of observed forest growth was used,with data between 2009 and 2014 from a 21.12 hm2 stem-mapped mixed forest plot (temperate coniferous and broadleaved forest) in Jiaohe,Jilin Province.Based on habitat type classifications,Ripley's L(r) function was adopted to evaluate the spatial distribution characteristics of tree locations.A mark correlation function(MCF) was used to evaluate the spatial autocorrelation characteristics of tree gro wth.Our results showed that (1) Pinus koraiensis (habitat type 3 ∶ 1-5 m),Quercus mongolica (habitat type 3 ∶ 1-3 m),Juglans mandshurica (habitat type 2 ∶ 1-2 m;habitat type 3 ∶ 1-7 m),Phellodendron amurense (habitat type 2 ∶ 1-3 m;habitat type 4∶1-5 m),Fraxinus mandshurica (habitat type 3∶1-2 m;habitat type 4∶1-2 m),and Euonymus verrucosus (habitat type 2∶1-15 m) were randomly distributed at particular scales and for certain habitat types.Nonetheless,they mainly showed clumped distribution patterns.For ten other species,the spatial distribution of tree positions showed an aggregated pattern at a scale of 0-30 m.(2) The mark correlation function analysis indicated that the radial growth of Ulmus davidiana var.japonica,Corylus mandshurica,Acer mono,E.verrucosus,and Carpinus cordata were positively autocorrelated for at least one of the habitat types.In contrast,the radial growth of Syringa reticulatavar.amurensis,J.mandshurica,Ulmuslaciniata,E.verrucosus,F.mandshurica,Tilia amurensis,Tilia mandshurica,C.mandshurica,A.mono,and Acer mandshuricum were negatively autocorrelated at least in one of habitat types.Other species did not show any significant autocorrelations at any of the studied scales.Our results indicate that different species had different spatial autocorrelation characteristics of radial growth,as well as significantly different habitat dependencies.【期刊名称】《生态学报》【年(卷),期】2017(037)006【总页数】9页(P1922-1930)【关键词】径向生长;生境差异;竞争作用;生境依赖性;Ripley's L(r)函数【作者】郝珉辉;张忠辉;赵珊珊;张春雨;赵秀海【作者单位】北京林业大学森林资源与生态系统过程北京市重点实验室,北京100083;吉林省林业科学研究院,长春130033;吉林省林业科学研究院,长春130033;北京林业大学森林资源与生态系统过程北京市重点实验室,北京 100083;北京林业大学森林资源与生态系统过程北京市重点实验室,北京 100083【正文语种】中文森林中普遍存在着种内和种间相互作用,林木个体之间的相互作用可大体分为正向作用和负向作用[1- 2]。
2004-2014年10年间蒙古栎胸径生长规律研究

【 中 图分 类 号 】 s 7 【 文 献标识 码】 A 【 文章编 号】 1 0 0 3 — 1 6 5 0( 2 0 1 6 ) 0 2 — 0 1 3 4 — 0 2
蒙 古栋 ( 学名: Qu e r c u s mo n g o l i c a F i s c h . e x L e d e b ) 为落 叶乔 木 ,高达 3 0 m。小 枝无 毛 ,叶倒 卵 形或 倒 卵状 长椭 圆形 , 长 7 - 1 9 c m, 先端短钝尖 , 基 部楔圆或耳状 , 粗 钝齿 7 一 l O 对, 幼叶沿 脉疏被毛 , 老叶近 无毛 , 侧脉 7 一 l l 对 】 。森林生态系统是 陆地 上 面积最大 、 分 布最广 、 组成 结构最复 杂 、 物质 资源最丰 富的生态 系统 , 它也是 自然 界功能 最完善的资 源库和生物基因库 。因此 , 森林生态系统对地球 上的生 物 , 特 别是 人类的生存有着至关 重 要的作用 。 林分直径分布 指林分 内各种大小直径的林木按径阶 分布状态 , 对于林 分的材 种结构 、 直径分布 、 断面积 、 材 积以及这 些 因子 生长 的研 究有重要 意义 , 是编制林分 出材量表 、 林 分生长 过程表 或收 获量表的基础 ,同时也是评估林分经济利 用价值和 经济效 益 , 预测未来林分直径结构 , 研究和推断相关 因子的结构 规律 , 制定 、 检查营林技术措施效果的依据 。 1 研 究 地 自然 概 况 蛟 河市位于吉林省东部 , 蛟河市东靠教化 市 , 南与桦甸市毗 邻, 西与永 吉县 、 吉林市 区隔送 花江相望 , 北接舒 兰市 、 五 常县 , 辖区面 积 6 0 5 0 平方千米 [ 4 1 , 地 理坐标为 东经 1 2 6  ̄ 4 4 ’ 一 1 2 8  ̄ 0 0 ’ , 北 纬4 3 。 l 2 ’ ~ 4 4  ̄ 0 9 ’ 之 间[ 5 1 , 全 局含有 巨大的林 产品资 源 , 是 吉林 省 较大的林业资源库。 1 . 1 样地设置 2 0 0 4 年在蛟河 市辖区天南林场 内选择 5 3 年 大小的天然 蒙 古栎幼林龄 作为研 究对象 , 设置 2 4 . 4 9 m× 2 4 . 4 9 m大小 的正方形 样地一块 , 样地垂直投影面积为 0 . 0 6 h m 2 。对样地 内样木胸径 ≥ 5 c m的林木进行编号 , 同时记录样地 中心点 的 G P S 值, 最后对样 地 内每株检 尺样木根据垂直投影 ,在正方形记录纸 上按 照编号 依次 点点位并做样木标注 , 以备样地复位之 用[ 6 1 。 1 . 2 数据采集 2 0 0 4 年和 2 0 1 4 年分别根据样地内样 木号进行检查 , 检查记 录采用每木 真值检尺 , 记录保 留一位小数。 1 . 3 研究方法 根据样地 内样木胸径分布特征对样木 前期胸径进行分段 处 理, 分段区间采用 2 c m—个区分段 , 共分为 1 2 个 区分段 , 根据分段 结果和样木生长量进行统计分析, 结果见图 l 。根据样木生长量绘 制生长量回归曲线 , 结果见图 2 。根据生长基数( 前期样木胸径 ) 和 生长量相互关系协 同样木生长量回归曲线共同研究样木胸径与生 长量分布规律 , 进而研究该 地区蒙古栎幼龄林生长规律 。 1 . 4 数据处理与分析 数 据 统计 分析 采 用 e x c e l 数 据 库软 件 ,图 像可 视化 采 用 S i g ma p l o t l O . 0软件。 2 研 究结 果 根据 统计结果可知 , 该研究 区域 内( 0 . 0 6 h m ) 两期 同时存活 的样 木共计 1 2 6株 , 其 中蒙 古栎 1 2 5 株, 椴树 l 株, 样木 前期平 均胸径 D 1 = 1 2 . 8 c m, 样木后期 平均胸 径 = 1 4 . O c m, 样木净生长 量3 - = 1 . 2 c m, 样 木年均生长量约 为 0 . 1 2 c m, 没参 与统 计的样木共 3 株, 其中1 株为 “ 负” 增长样木 , 根 据现地调查 该样 木现地生长 情况良好 , 暂 定为胸径错测木 , 另外 2 株为新进阶的样木 , 2 株均
吉林省蛟河林业实验区管理局天然林胸径及生长量研究
2017 年第 11 期(下半月)Nong Min Zhi Fu Zhi You 农民致富之友95科研◎林业科学吉林省蛟河林业实验区管理局天然林胸径及生长量研究陈连栋1 潘丽铭2 王万峰1林业行业作为我国的传统行业具有悠久的历史,在改革开放初期,全国开展家园大建设的过程中,林业作为主要的木材提供场所,为新中国的建设作出了巨大的贡献,与此同时也带来了过度砍伐和破坏严重的后果,随着我国经济的高速发展和人民生活水平的提供,人类对森林的休闲娱乐功能提出了更高的要求,鉴于此,很多科研工作者和有识之士提出了恢复森林的科学命题,国家更是出台了一些列天然林保护政策,为森林各项功能的恢复提供了必要条件,因此天然林研究再一次成为了很多科研工作者的热点。
直径结构是森林结构最重要的组成部分,其不仅是树高、断面积和材积等结构的基础,而且也是估算森林蓄积量,指导森林可持续经营的主要依据,因此本文主要对蛟河地区天然林的直径结构和生长量进行初步调查研究。
1 研究地自然概况吉林省蛟河林业实验区管理局,是长白山余脉的尾部地区,地理坐标为东经127°35′-127°51′,北纬43°51′-44°05′,辖区总经营面积31823公顷,多以林业用地为主,森林覆盖率近87%,具有优越的地理环境和丰富的动植物资源,很多东北地区的特有物种在研究区内均有分布,是搞林业研究的专家学者们趋之若鹜之所。
在“东北地区天然林全面禁伐”项目实施以来,该区内的林农在没有木材生产的前提下,丰富的林下资源如蘑菇、核桃、大榛子等林产品便是该辖区林农的主要经济来源,受到绝大多数林产品爱好者的青睐。
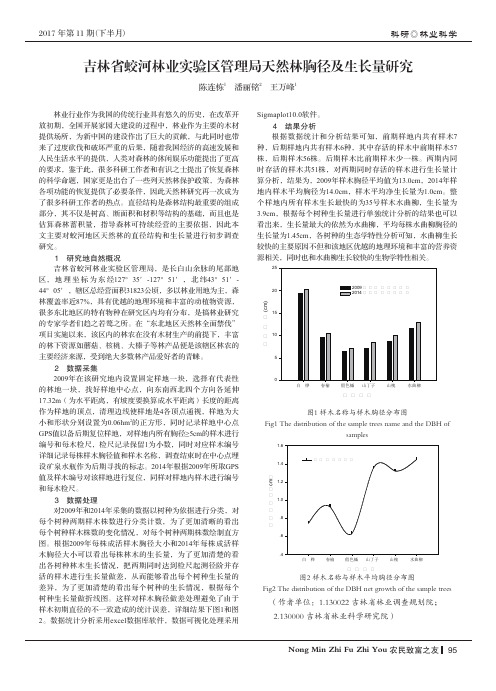
2 数据采集2009年在该研究地内设置固定样地一块,选择有代表性的林地一块,找好样地中心点,向东南西北四个方向各延伸17.32m (为水平距离,有坡度要换算成水平距离)长度的距离作为样地的顶点,清理边线使样地是4各顶点通视,样地为大小和形状分别设置为0.06hm 2的正方形,同时记录样地中心点GPS 值以备后期复位样地,对样地内所有胸径≥5cm 的样木进行编号和每木检尺,检尺记录保留1为小数,同时对应样木编号详细记录每株样木胸径值和样木名称,调查结束时在中心点埋设矿泉水瓶作为后期寻找的标志。
林政资源管理存在的问题与对策

林业园艺Lin ye yu a n yi 1吉林省蛟河市的林业资源吉林省蛟河市处于吉林省东部,总面积六千两百三十五平方千米,水流域面积为两千四百二十六平方千米,有较为丰富的水资源。
吉林省蛟河市各国有林场经营面积为6821175亩,达到总面积的66%以上,属于吉林省主要林区之一。
其中林业用地3689805亩,有林地面积3435364.35亩,森林总蓄积量2719.35万立方米。
林地面积按起源划分,天然林3111337.80亩,人工林324026.55亩;按功能划分,生态公益林3486409亩(其中国家级公益林3362792亩,省级公益林123617亩)商品林68671.05亩(其中天然林45293.40亩)。
吉林省蛟河市主要种植树种为红松、黄菠萝、水曲柳、桦树及杨柳等47种。
2林政资源管理存在的问题林业作为社会公益事业的重要组成部分,能够供给人们生活所需的林产品,还能够建设生态环境。
林业对人们的生活产生不可忽视的良性影响,人们对于建设生态林业的重视,有利于加强林政资源管理,有利于建设良好的生态环境。
虽然相关林业管理部门对于林业建设工作较为重视,但在实际操作过程中还存在较多的问题。
2.1林政资源管理建设力度不足林政资源管理工作建设力度不足,主要是因为没有完善的法律法规作为基础,也没有完善的管理制度作为约束,导致工作人员在完成工作时没有相关的标准作为参考,严重影响了本地区林政资源管理工作的顺利开展。
专业人员参与工作较少,会影响到本地区的现代林业建设。
部分地区的林政资源管理工作使用的设备与方式较为落后,也会影响到林政资源管理工作的开展,严重影响了本地区的生态文明建设。
2.2相关法制意识较为薄弱本地区林政资源管理工作的宣传存在一些漏洞,相关的法律法规建设也不完善,使得林业保护意识宣传不足,无法营造良好的舆论环境。
在完成管理工作过程中,管理人员对于相关法律法规的宣传也存在一定的问题,严重影响了人们对于生态建设的重要性认识,也会影响到当地的林政资源管理工作的顺利开展。
吉林省蛟河林业实验区管理局森林资源现状及可持续经营对策

吉林省蛟河林业实验区管理局森林资源现状及可持续经营对策王辉;秦树林
【期刊名称】《林业勘查设计》
【年(卷),期】2012(000)002
【摘要】通过对吉林省蛟河林业实验区管理局2011年森林资源数据的统计分析,并结合当地森林资源特点及经营管理存在的问题,从加强森林资源管理和优化森林资源结构等方面提出了可持续发展对策.
【总页数】4页(P2-5)
【作者】王辉;秦树林
【作者单位】吉林省林业调查规划院;吉林省林业调查规划院
【正文语种】中文
【相关文献】
1.吉林省蛟河林业实验区管理局组织开展"庆七一党建活动日"活动 [J], 邳田辉
2.吉林省蛟河林业实验区管理局天然林分布情况 [J], 宋彩民;郑军;王峰洁
3.浅谈蛟河林业实验区管理局森林抚育现状及经营建议 [J], 王大勇;周纯
4.双辽市林业局森林资源现状及可持续经营对策 [J], 秦树林;郑军;王磊
5.吉林省蛟河林业实验区管理局 [J],
因版权原因,仅展示原文概要,查看原文内容请购买。
吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应

第41卷第7期2021年4月生态学报ACTAECOLOGICASINICAVol.41,No.7Apr.,2021基金项目:国家自然科学基金项目(31971650,31600480)收稿日期:2020⁃06⁃05;㊀㊀网络出版日期:2021⁃01⁃28∗通讯作者Correspondingauthor.E⁃mail:zcy_0520@163.comDOI:10.5846/stxb202006051462夏梦洁,王晓霞,郝珉辉,赵秀海,张春雨.吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应.生态学报,2021,41(7):2794⁃2802.XiaMJ,WangXX,HaoMH,ZhaoXH,ZhangCY.Distributionpatternoffunctionaltraitsanditsresponsetotopographicfactorsinaconiferandbroad⁃leavedmixedforestinJiaohe,Jilinprovince.ActaEcologicaSinica,2021,41(7):2794⁃2802.吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应夏梦洁1,王晓霞2,郝珉辉1,赵秀海1,张春雨1,∗1北京林业大学国家林业和草原局森林经营工程技术研究中心,北京㊀1000832吉林省林业勘察设计研究院,长春㊀130022摘要:森林群落的构建过程及其内在机制是生态学研究的热点问题㊂植物功能性状是指能够代表植物的生活史策略,反映植物对环境变化响应的一系列植物属性㊂通过植物功能性状的分布格局及其对环境因素的响应有助于推测群落的构建过程及其内在作用机制㊂以吉林蛟河21.12hm2温带针阔混交林样地为研究对象,采集并测量了样地内34种木本植物的6种不同的功能性状㊂以20mˑ20m的样方为研究单元,通过计算平均成对性状距离指数(meanpairwisetraitdistance;PW)和平均最近邻体性状距离指数(meannearestneighbortraitdistance;NN)来探讨群落中单个性状和综合性状的分布格局㊂同时结合地形因子采用回归分析探讨功能性状的分布格局对局域生境变化的响应㊂基于PW的结果显示:单个性状中除叶面积外,其余性状的分布格局均为聚集分布多于离散分布;基于NN的结果显示:除叶面积和最大树高外,其余性状的分布格局为聚集分布多于离散分布㊂此外,由6种单个性状组成的综合性状的分布格局同样为聚集分布多于离散分布㊂基于回归分析的结果显示:森林群落中功能性状的分布格局受到海拔㊁坡度和坡向等因素的显著影响,而凹凸度的影响则不显著㊂研究结果表明包括环境过滤和生物相互作用的非随机过程能够影响温带针阔混交林的群落构建过程,中性过程对该区域群落构建过程的影响不显著㊂关键词:功能性状;功能性状结构;群落构建机制;环境因素Distributionpatternoffunctionaltraitsanditsresponsetotopographicfactorsinaconiferandbroad⁃leavedmixedforestinJiaohe,JilinprovinceXIAMengjie1,WANGXiaoxia2,HAOMinhui1,ZHAOXiuhai1,ZHANGChunyu1,∗1ResearchCenterofForestManagementEngineeringofStateForestryandGrasslandAdministration,BeijingForestryUniversity,Beijing100083,China2ForestrySurveyandDesignInstituteofJilinProvince,Changchun130022,ChinaAbstract:Understandingthemechanismandprocessofcommunityassemblyisakeyissueinecologicalresearch.Functionaltraits,whichrepresentplantphysiological,morphological,andphenologicalattributes,havedecisiveeffectsonplantgrowth,survival,reproductionanddispersal.Examiningthespatialdistributionpatternofplantfunctionaltraitsanditsresponsetolocalenvironmentalvariationishelpfultospeculatetheunderlyingprocessesandmechanismsthatregulatecommunityassembly.Inthisstudy,weexploredtheunderlyingmechanismsofcommunityassemblyusingthedatasetfroma21.12hm2temperateconiferandbroad⁃leavedmixedforestplotinJiaohe,NortheastChina.Fourtopographicattributes,includingelevation,convexity,slope,andaspect,weremeasuredaslocalenvironmentalfactors.Sixfunctionaltraits,namelyleafarea,specificleafarea,leafcarboncontent,leafnitrogencontent,leafcarbontonitrogenratio,andmaximumtreeheight,werecollectedandmeasuredforall34woodyspeciesthatpresentinthistemperateforestplot.Twofunctionalstructureindices,includingthemeanpair-wisetraitdistance(PW)andthemeannearestneighbortraitdistance(NN),wereusedtoexplorethespatialdistributionpatternofplanttraits.Furthermore,regressionanalyseswereusedtoinvestigatetheresponseoftraitpatternstoenvironmentalvariation.BasedontheresultsofPW,wefoundthat,exceptfortheleafareatraits,theremainingtraitsmainlyshowedtheclumpedspatialpatterns.TheresultsbasedonNNshowedthat,exceptfortheleafareaandmaximumtreeheight,theremainingtraitspresentedtheclumpedspatialpatterns.Theresultsofregressionanalysesshowedthatthedistributionpatternsoffunctionaltraitsweresignificantlyaffectedbyelevation,slope,andaspect.Incontrast,convexityhadnosignificanteffectontraitdistributioninthistemperateforestcommunity.Ourresultsindicatethatthenon⁃randomprocessesincludingenvironmentalfilteringandbiologicalinteractionsplayedimportantrolesinregulatingforestcommunityassembly,whiletheroleofneutralprocesswasnotsignificantinthistemperateconiferandbroad⁃leavedmixedforest.KeyWords:functionaltraits;functionaltraitstructures;communityassembly;environmentalvariables群落构建机制与构建过程是生态学研究的热点问题,生态位理论和中性理论是解释群落构建过程的两个主要理论㊂生态位理论认为生态位完全相同的物种不能共存于群落中,环境过滤和生物间相互作用是影响生物多样性格局的两个重要过程[1⁃2]㊂而中性理论则认为群落中的物种多样性是由长时间物种的随机迁入迁出和生态漂移形成的[3⁃5]㊂扩散限制是中性理论的重要推论,即随着空间距离的增加,群落的相似性会降低,因此扩散限制也是影响群落构建的重要过程㊂植物功能性状是指能够影响植物本身的生理㊁生态学过程,反映植物对环境变化响应的一系列功能属性,因此常被用来探讨群落的构建机制与构建过程[6⁃7]㊂例如Freschet等在不同的森林类型中利用两种叶片性状即叶氮含量和比叶面积验证了生物和非生物因子共同驱动群落的构建[8]㊂有研究显示不同性状在相同空间尺度下的分布格局不同,例如:比叶面积在局域尺度下呈聚集分布时,而种子大小则表现为离散分布,表明不同功能性状对不同生态过程的响应不同[9⁃11]㊂此外,近年来生态学家试图将植物性状的分布格局与特定生境条件相联系[12],这对于森林群落的经营管理和多样性保护具有十分重要的意义㊂将群落功能性状的分布与局域尺度的环境因子联系起来,有助于检验出不同环境因子对性状分布格局的影响,并推测相应的生态学过程㊂本研究通过植物的叶面积㊁比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比和最大树高6类主要功能性状,以21.12hm2大面积森林固定监测样地群落调查数据为基础,分析了20mˑ20m空间尺度下的群落功能性状分布格局,并评估生态位过程和中性过程在吉林蛟河针阔混交林群落构建机制中的相对重要性㊂本研究同时结合样地内海拔高度㊁坡度㊁坡向和凹凸度四种地形因子,探讨不同地形因子对森林群落中性状分布的影响㊂研究旨在揭示吉林蛟河温带针阔混交林中功能性状的分布格局及其对生境变化的响应,并推测其背后的群落构建机制和构建过程㊂1㊀研究方法1.1㊀研究区概况与样地设置本研究区位于吉林省蛟河林业试验区管理局,地理位置为43ʎ57.897ᶄ 43ʎ58.263ᶄN,127ʎ42.789ᶄ 127ʎ43.310ᶄE㊂该气候属于受季风影响的温带大陆性气候,年均温为3.8ħ,最热月为7月,平均气温21.7ħ,最冷月为1月,平均气温-18.6ħ,年均降水量695.9mm㊂该地区林分类型为典型的次生针阔混交林,土壤类型为棕色森林土,土层厚度为20 100cm[13]㊂2009年建立面积为21.12hm2(660mˑ320m)的固定森林监测样地,样地最低海拔为425.3m,最高海拔为525.8m,相对最大高差为100.5m,地势变化较大㊂样地群落调查过程中,记录了样地内所有胸径(DBH)大于1cm的木本植物的物种名称㊁胸径㊁树高㊁冠幅和相对位置坐标,并挂牌标记进行长期监测㊂样地内主要树种有胡桃楸(Juglansmandshurica)㊁紫椴(Tiliaamurensis)㊁蒙古栎5972㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀(Quercusmongolica)㊁色木槭(Acermono)和红松(Pinuskoraiensis)等㊂图1㊀样地地形示意图Fig.1㊀Schematicdiagramoftheforestplot1.2㊀地形因子测量地形因子可以综合反映样地内土壤养分和水分状况,并在一定程度解释群落内植物的生长状况[14],本研究中我们采用海拔高度㊁凹凸度㊁坡度和坡向这四个地形因子记为环境因子㊂将整个研究区域划分为528个20mˑ20m的样方,采用Harm等人的计算方法计算地形因子[15]㊂具体计算过程为:每个样方4个顶点海拔值的平均值为该样方的平均海拔高度;目标样方的海拔减去与之相邻的8个样方的海拔平均值为该样方的凹凸度,其中,山脊为正值,洼地为负值;分别连接每个样方的3个顶点形成4个三角形平面,每个平面与该样方投影夹角的平均值为目标样方的坡度;4个平面与正北方向的角度平均值为目标样方的坡向[15⁃17]㊂基于该样地,已开展的相关研究表明,森林中不同的树种具有不同的生境偏好,树种的生长和分布都会受到地形因子的显著影响[13,18⁃19],并且有研究表明环境因子对树种分布的影响可能是通过物种所具有的功能性状而发挥作用[16]㊂但地形因子如何影响树种功能性状的分布格局目前仍然不十分清楚,有开展必要进一步的研究㊂1.3㊀功能性状样品的采集与测定本研究选择样地内胸径(DBH)大于5cm的乔木和大灌木树种作为研究对象,共计34个种㊂于2017年生长季采集并测量了样地内34个树种的6类关键功能性状数据,包括:叶面积㊁比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比和最大树高㊂叶片性状反映植物的生活史策略及其对环境条件的响应[20],叶面积反映植物对光的捕获能力,比叶面积反映植物在生长过程中的资源获取能力;叶氮含量表示植物在叶经济型谱上的位置,反映植物的资源获取能力[6];叶碳含量是植物生长过程中维持碳平衡的重要参数[21];叶碳氮比反映林木的碳氮代谢情况以及营养利用率㊂最大树高作为茎干性状的代表,反映植物光学生态位和综合竞争能力[16,22]㊂功能性状样品的采集和测定均参照Cornelissen等人的要求进行,常见种的采样株数在30株以上,部分稀有种的采样株数在10株以上[23];最大树高为样地内每个树种最高的前5%个体的平均值[17]㊂1.4㊀构建功能性状树在获取叶面积㊁比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比和最大树高的性状数据,首先对原始性状数据进行对数转化,然后通过主成分分析获取能够代表所有功能性状的主成分,选取其中综合解释量在90%以上的主成分作为综合功能性状因子,通过计算欧氏距离生成性状距离矩阵,采用最远距离法对性状距离矩阵进行系统聚类,生成功能性状聚类树[24]㊂1.5㊀计算功能性状分布格局本文计算了分别基于6种单个性状和一种综合性状的性状聚类,采用平均成对性状距离指数(meanpairwisetraitdistance;PW)和平均最近邻体性状距离指数(meannearestneighbortraitdistance;NN)计算样地内每个20mˑ20m的样方的功能性状分布格局[25⁃26]㊂这两种算法均需要通过性状聚类树随机转换实现,利用观察到的PW值和NN值量化群落性状分布结构,保持物种数量及个体数不变,将聚类树的末端树种随机转换999次,计算零模型下的功能性状分布,采用随机分布结果将观测值标准化,使我们获得标准条件下的PW和NN值即S.E.S.PW和S.E.S.NN,计算S.E.S.PW和S.E.S.NN的公式如下:S.E.S.PW=-1ˑ(PWact-mean(PWrand))/sd(PWrand)S.E.S.NN=-1ˑ(NNact-mean(NNrand))/sd(NNrand)式中,PWact代表实际观察到的平均成对性状距离,mean(PWrand)代表零模型随机转换999次物种后的成对性状距离模拟值的平均值,sd(PWrand)代表零模型模拟值的标准差;同理,NNact表示实际观察到的群落平均最近6972㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀邻体性状距离,mean(NNrand)代表零模型随机转换999次物种后的最近邻体性状距离模拟值的平均值,sd(NNrand)代表零模型模拟值的标准差㊂S.E.S.PW(或S.E.S.NN)值大于零代表功能性状呈聚集分布格局,小于零代表功能性状呈离散分布格局,等于零代表功能性状随机分布㊂1.6㊀功能性状分布对地形变化的响应本研究采用海拔高度㊁坡度㊁坡向和凹凸度这4个地形因子为环境变量,结合组合功能性状的分布格局,运用逐步回归的方法分析不同地形因子对综合功能性状格局的解释程度,探讨影响样地内群落构建的主要驱动因子㊂本研究相关计算均在统计软件R.3.5.1中完成,分别采用了 vegan ㊁ picante 软件包[27⁃28]㊂2㊀结果2.1㊀植物功能性状树本研究采用最远距离法进行系统聚类得到功能性状聚类树(图2),与直接反映植物进化关系的系统发育树不同,功能性状树主要反映了植物生态功能与生态策略之间的相似性(即性状与性状之间的相似性)㊂例如:本研究中山杨㊁白桦都属于先锋树种,具有较大的比叶面积㊁较高的叶氮含量和较低的木质密度,因此在功能性状树上的位置相近;色木槭㊁拧筋槭㊁糠椴㊁紫椴等属于硬阔树种,其叶片为单叶,具有较小的叶面积和较高的木质密度,因此在功能性状树上的位置相近;水曲柳㊁花曲柳和黄檗同属于硬阔树种,但其叶片为复叶,具有较大的叶面积和较高的木质密度,因此在功能性状树上的位置相近;而水榆花楸㊁稠李㊁山丁子㊁青楷槭㊁簇毛槭㊁千金榆㊁鼠李㊁黑樱桃㊁毛榛和花楸等灌木或小乔木则聚为一类,这一类树种的最大特征为其最大树高较小,是样地内典型的亚林层树种㊂此外,还有部分树种以其独特的性状特征单独划分为一类,例如胡桃楸(复叶树种)以其极大的叶面积,怀槐(豆科固氮树种)以其极高的叶氮含量等特征单独聚为一类,反映了此类树种在样地内独特的生态地位㊂2.2㊀植物功能性状分布格局基于平均成对性状距离(PW)的研究结果显示(表1;图3):单个性状中,叶面积的分布格局为离散分布(即图3中蓝色区域)多于聚集分布(即图3中红色区域),在全部528个样方中,有432个样方(81.8%)呈现为离散分布;除叶面积外,其余性状的分布格局均为聚集分布多于离散分布㊂基于平均最近邻体性状距离(NN)的研究结果显示(表1;图4):单个性状中,叶面积和最大树高分布格局为离散分布多于聚集分布,在全部528个样方中,分别有79.9%和60.0%的样方呈现为离散分布,而其余性状的分布格局为聚集分布多于离散分布(表1;图4)㊂总体而言基于平均最近邻体性状距离和平均成对性状距离的研究结果具有较高的一致性㊂此外,由6种单个性状组成的综合性状因子的分布格局同样为表现为聚集分布多于离散分布(表1;图5)㊂表1㊀性状呈聚集分布和离散分布的样方比例/%Table1㊀ProportionofaggregationanddispersionpatternsS.E.S.PWS.E.S.NN结构聚集(显著聚集)Aggregation(significant)结构分散(显著分散)Dispersion(significant)结构聚集(显著聚集)Aggregation(significant)结构分散(显著分散)Dispersion(significant)叶面积Leafarea18.2(1.5)81.8(25.6)20.1(0.8)79.9(22.7)比叶面积Specificleafarea98.1(27.3)1.9(0.0)73.5(25.2)26.5(0.0)叶氮含量Leafnitrogen92.8(7.8)7.2(0.0)89.0(4.9)11.0(0.0)叶碳含量Leafcarbon69.3(3.8)30.7(0.2)76.7(8.3)23.3(1.1)叶碳氮比Carbontonitrogenratio81.1(10.0)18.9(0.4)86.9(6.6)13.1(0.4)最大树高Maximumtreeheight61.4(9.7)38.6(5.3)40.0(4.5)60.0(23.3)综合性状因子Alltraits48.9(4.4)51.1(0.8)60.4(4.0)39.6(0.6)㊀㊀S.E.S.NN和S.E.S.PW分别表示PW和NN标准化效应值,括号数值内表示性状分布呈显著聚集或显著离散(P<0.05)的样方比例2.3㊀环境变量对功能性状分布格局的影响本研究利用综合性状因子计算了局域环境变量,包括海拔高度(ELE)㊁坡度(SLO)㊁坡向(ASP)和凹凸度7972㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀8972㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀图2㊀综合性状聚类树Fig.2㊀Clusteringtreebasedonfunctionaltraits(CON)对功能性状分布格局的相对解释程度㊂研究基于逐步回归的算法,结果发现局域环境变量对S.E.S.PW的解释量为37.4%,对S.E.S.NN的解释量为23.6%,其具体公式为:S.E.S.PW=0.380ELE+0.366SLO+0.088ASP;S.E.S.NN=0.306ELE+0.288SLO+0.074ASP,结果显示在已知的环境因子中,海拔高度㊁坡度和坡向对群落功能性状分布格局的影响显著,而凹凸度则对功能性状分布无显著影响,被排斥在回归模型之外㊂群落功能性状分布格局与局域环境变量显著相关,表明环境过滤在该研究区域内发挥着的重要作用㊂3㊀讨论3.1㊀平均成对性状距离指数和平均最近邻体性状距离指数平均成对性状距离指数(PW)和平均最近邻体性状距离指数(NN)都可以用来探讨群落功能性状的分布格局,这两种方法都是基于距离矩阵来量化群落功能性状的分布格局[29]㊂不同的是PW侧重于对整体群落的分析,是分析性状分布格局的基础[25],它用于衡量群落中所有物种的平均相关性[10],侧重于整个群体的平均值;而NN指数则是测量亲缘关系最近物种对之间的相关性[30],显示的是群落功能性状聚类树的终端性状分布格局㊂有研究认为这两种方法都具有检测环境过滤的作用,而NN指数更易检测相似性限制带来的影响[30]㊂PW指数和NN指数计算同一区域的性状分布格局,结果出现偏差,可能是性状在样地区域内分布不均造成的,本研究结果显示最大树高在不同计算方法下的结果存在差异,可能与该性状在样方内的分布不均匀有关[31]㊂图3㊀植物功能性状分布格局(基于平均成对性状距离法)Fig.3㊀Distributionpatternsofplantfunctionaltraits(basedonthemeanpairwisetraitdistance)红色表示性状聚集分布,蓝色表示性状离散分布,颜色越深表示性状聚集或离散分布的程度越明显图4㊀植物功能性状分布格局(基于平均最近邻体性状距离法)Fig.4㊀Distributionpatternsofplantfunctionaltraits(basedonthemeannearestneighbortraitdistance)红色表示性状聚集分布,蓝色表示性状离散分布,颜色越深表示性状聚集或离散分布的程度越明显9972㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀图5㊀综合性状因子的分布格局Fig.5㊀Distributionpatternsbasedonallsixtraits红色表示性状结构聚集,蓝色表示性状结构离散,颜色越深表示性状的聚集或离散程度越明显3.2㊀功能性状分布格局与群落构建机制已开展的相关研究表明,森林中不同的树种具有不同的生境偏好,树种的生长和分布都会受到地形因子的显著影响[13,18⁃19],并且有研究表明环境因子对树种分布的影响可能是通过物种所具有的功能性状而发挥作用[16]㊂因此,生态学家通过量化群落中功能性状的分布格局来推断影响群落构建的生态学机制㊂不同的功能性状反映了植物不同的生理㊁生态特征和生活史策略,因此其在群落中的分布格局以及其对生境变化的响应也会存在差异㊂环境过滤和相似性限制可以通过作用于不同类型的功能性状来影响群落中物种和性状的分布格局[32]㊂研究显示,环境会过滤掉不适生的性状,导致适生性状呈聚集状态[33],而离散的性状分布则是由功能性状之间的相似性限制造成,其与生态位分化有关㊂一般认为物种所具有的性状越相似,物种间的竞争就越激烈,而竞争导致某些具有相似性状的物种占据不同的生态位,使物种或性状的分布呈离散格局[34]㊂在此基础上,有研究认为与生产力相关的功能性状在小尺度上是聚集的,尤其是与叶的相对生长速率,叶片寿命和叶片的营养物质相关的性状[35]㊂本研究显示叶碳含量㊁叶氮含量和叶碳氮比等直接反映叶片营养物质的性状以及反映植物生长过程的比叶面积均呈显著聚集状态,综合性状结构同样呈显著聚集分布格局,与前人的研究结果较为一致[36⁃37]㊂研究显示竞争型性状在群落内容易呈离散分布的结构[26],而叶面积呈显著离散分布,可能与研究区域的林分类型有关㊂此外,最大树高在两种不同运算下结果有差异,该研究结果与Swenson等的研究类似,可能是由于最大树高在不同样方内分布不均,导致PW和NN计算结果不同[31]㊂本研究结果显示叶氮含量㊁叶碳含量和比叶面积等性状在群落中呈聚集分布,反映了环境过滤在群落构建中的重要作用,该结果与温带和热带地区的研究结果类似[9],但组合性状分布结构显示,虽然呈显著聚集结构的样方较多,但样地中也有很多有离散趋势的样方,这表明除环境过滤外,生物之间的相互作用对该地区的群落构建也有影响,非随机过程是影响该地区群落构建的主要驱动因子[38]㊂3.3㊀地形因子与群落功能性状分布本文从综合功能性状的角度,结合局域的地形因子,利用不同的功能性状结构指数,对两种计算结果回归分析显示,地形因子对性状分布格局影响显著㊂有研究指出,在环境条件较差的区域,例如低海拔干旱和高海拔霜冻地区,由于环境过滤作用的影响,群落的功能性状结构通常呈聚集分布,而在环境条件适宜的低海拔地区,生物间的竞争作用强烈,而环境过滤的作用并不明显,因此群落的功能性状结构多呈离散分布[39]㊂性状分布呈现的这种差异既来自生态位保守的影响,也是性状对每个特定的环境条件的适应性反应[40]㊂在我们的研究区域内,如图5综合性状分布格局显示,样地东部和西北部海拔较高处的功能性状格局多呈聚集分布,样地中部多呈离散分布,从图1的地形图中可以看出,聚集分布多处于上坡带,海拔高度和地面凹凸度均有较大幅度的改变,而离散分布多处于地势低平㊁环境条件较为温和的区域㊂凹凸度㊁海拔高度㊁坡度和坡向这四种地形因子与综合性状结构逐步回归分析显示,海拔和坡度坡向对群落性状分布具有显著影响,凹凸度的影响很小,基本忽略不计,而在Liu等人[16]的研究中,发现海拔高度和地0082㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀形凹凸度对性状分布的影响最大,研究结果的差异可能与研究地的地形条件不同有关㊂有研究指出,海拔较高的区域,温度和大气压降低,太阳辐射增加,可以影响植物的生长进程[41]㊂样地地形图显示,整块样地的地形变化差异显著,将样地一分为二,明显东部地区海拔高,西部地区海拔趋于平缓,样地生境存在明显的异质性,这种生境异质性很大程度上影响了资源的利用情况[42],并进一步影响了功能性状的分布格局㊂在这种地形条件下,坡度能够显著影响到植物对土壤水分㊁养分以及光照等资源的获取与利用,从而对植物的性状进行选择和过滤,进而影响了群落的物种和性状分布格局㊂4㊀结论本研究从植物功能性状的角度出发,结合环境因子,探讨影响蛟河地区群落构建的主要驱动机制㊂研究表明,不同性状的分布格局不同,比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比在研究区域内呈明显的聚集分布,表示环境过滤对物种的功能性状分布格局具有重要作用;叶面积呈明显的离散分布,表示相似性限制的重要作用;最大树高在不同计算方法下的分布结果有差异㊂研究显示,环境因子对性状分布具有显著影响,其中海拔高度的影响最为显著,表明环境过滤的重要作用㊂综上所述,本研究认为非随机过程是蛟河针阔混交林群落构建的主要驱动因子,环境过滤和生物之间的相似性限制是群落内物种和功能性状分布格局的主要决定因素㊂参考文献(References):[1]㊀WeiherE,ClarkeGDP,KeddyPA.Communityassemblyrules,morphologicaldispersion,andthecoexistenceofplantspecies.Oikos,1998,81(2):309⁃322.[2]㊀SiefertA,RavenscroftC,WeiserMD,SwensonNG.Functionalbeta⁃diversitypatternsrevealdeterministiccommunityassemblyprocessesineasternNorthAmericantrees.GlobalEcologyandBiogeography,2013,22(6):682⁃691.[3]㊀EtienneRS,AlonsoD.Adispersal⁃limitedsamplingtheoryforspeciesandalleles.EcologyLetters,2005,8(11):1147⁃1156.[4]㊀RosindellJ,HubbellSP,EtienneRS.Theunifiedneutraltheoryofbiodiversityandbiogeographyatageten.Trendsinecology&evolution,2011,26(7):340⁃348.[5]㊀柴永福,岳明.植物群落构建机制研究进展.生态学报,2016,36(15):4557⁃4572.[6]㊀周道玮.植物功能生态学研究进展.生态学报,2009,29(10):5644⁃5655.[7]㊀孟婷婷,倪健,王国宏.植物功能性状与环境和生态系统功能.植物生态学报,2007,31(1):150⁃165.[8]㊀FreschetGT,DiasATC,AckerlyDD,AertsR,VanBodegomPM,CornwellWK,DongM,KurokawaH,LiuGF,OnipchenkoVG,OrdoñezJC,PeltzerDA,RichardsonSJ,ShidakovII,SoudzilovskaiaNA,TaoJP,CornelissenJHC.Globaltocommunityscaledifferencesintheprevalenceofconvergentoverdivergentleaftraitdistributionsinplantassemblages.GlobalEcologyandBiogeography,2011,20(5):755⁃765.[9]㊀SwensonNG,EnquistBJ.Ecologicalandevolutionarydeterminantsofakeyplantfunctionaltrait:wooddensityanditscommunity⁃widevariationacrosslatitudeandelevation.AmericanJournalofBotany,2007,94(3):451⁃459.[10]㊀WebbCO.Exploringthephylogeneticstructureofecologicalcommunities:anexampleforrainforesttrees.AmericanNaturalist,2000,156(2):145⁃155.[11]㊀房帅,原作强,蔺菲,叶吉,郝占庆,王绪高.长白山阔叶红松林木本植物系统发育与功能性状结构.科学通报,2014,59(24):2342⁃2348.[12]㊀AndersonMJ,CristTO,ChaseJM,VellendM,InouyeBD,FreestoneAL,SandersNJ,CornellHV,ComitaLS,DaviesKF,HarrisonSP,KraftNJB,StegenJC,SwensonNG.Navigatingthemultiplemeaningsofβdiversity:aroadmapforthepracticingecologist.EcologyLetters,2011,14(1):19⁃28.[13]㊀郝珉辉,张忠辉,赵珊珊,张春雨,赵秀海.吉林蛟河针阔混交林树木生长的空间关联格局.生态学报,2017,37(6):1922⁃1930.[14]㊀WestobyM,FalsterDS,MolesAT,VeskPA,WrightIJ.Plantecologicalstrategies:someleadingdimensionsofvariationbetweenspecies.AnnualReviewofEcologyandSystematics,2002,33:125⁃159.[15]㊀HarmsKE,ConditR,HubbellSP,FosterRB.Habitatassociationsoftreesandshrubsina50⁃haneotropicalforestplot.JournalofEcology,2001,89(6):947⁃959.[16]㊀LiuJJ,TanYH,SlikJWF.Topographyrelatedhabitatassociationsoftreespeciestraits,compositionanddiversityinaChinesetropicalforest.ForestEcologyandManagement,2014,330:75⁃81.1082㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀2082㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀[17]㊀郝珉辉,李晓宇,夏梦洁,等.抚育采伐对蛟河次生针阔混交林功能结构和谱系结构的影响.林业科学,2018,54(5):1⁃9.[18]㊀ZhangCY,ZhaoYZ,ZhaoXH,VonGadowK.Species⁃habitatassociationsinanortherntemperateforestinChina.SilvaFennica,2012,46(4):501⁃519.[19]㊀郝珉辉,张忠辉,赵珊珊,赵秀海,叶尔江㊃拜克吐尔汉,张春雨.吉林蛟河针阔混交林树木生长与生境的关联性.生态学报,2017,37(10):3437⁃3444.[20]㊀WrightIJ,ReichPB,WestobyM,AckerlyDD,BaruchZ,BongersF,Cavender⁃BaresJ,ChapinT,CornelissenJHC,DiemerM,FlexasJ,GarnierE,GroomPK,GuliasJ,HikosakaK,LamontBB,LeeT,LeeW,LuskC,MidgleyJJ,NavasML,NiinemetsÜ,OleksynJ,OsadaN,PoorterH,PootP,PriorL,PyankovVI,RoumetC,ThomasSC,TjoelkerMG,VeneklaasEJ,VillarR.Theworldwideleafeconomicsspectrum.Nature,2004,428(6985):821⁃827.[21]㊀BoernerREJ.Foliarnutrientdynamicsandnutrientuseefficiencyoffourdeciduoustreespeciesinrelationtositefertility.JournalofAppliedEcology,1984,21(3):1029⁃1040.[22]㊀MolesAT,WartonDI,WarmanL,SwensonNG,LaffanSW,ZanneAE,PitmanA,HemmingsFA,LeishmanMR.Globalpatternsinplantheight.JournalofEcology,2009,97(5):923⁃932.[23]㊀CornelissenJHC,LavorelS,GarnierE,DíazS,BuchmannN,GurvichDE,ReichPB,TerSteegeH,MorganHD,VanDerHeijdenMGA,PausasJG,PoorterH.Ahandbookofprotocolsforstandardisedandeasymeasurementofplantfunctionaltraitsworldwide.AustralianJournalofBotany,2003,51(4):335⁃380.[24]㊀PetcheyOL,GastonKJ.Functionaldiversity(FD),speciesrichnessandcommunitycomposition.EcologyLetters,2002,5(3):402⁃411.[25]㊀LiuXJ,SwensonNG,ZhangJL,MaKP.Theenvironmentandspace,notphylogeny,determinetraitdispersioninasubtropicalforest.FunctionalEcology,2013,27(1):264⁃272.[26]㊀WeiherE,KeddyPA.Assemblyrules,nullmodels,andtraitdispersion:newquestionsfromoldpatterns.Oikos,1995,74(1):159⁃164.[27]㊀OksanenJ,GuillaumeBF,KindtR,LegendreP,MinchinPR,OᶄHaraRB,SimpsonGL,SolymosP,StevensMHH,WagnerH.Vegan:communityecologypackage.Rpackageversion2.0⁃9,2013,2(9):1⁃295.http://vegan.r⁃forge.r⁃project.org.[28]㊀KembelSW,CowanPD,HelmusMR,CornwellWK,MorlonH,AckerlyDD,BlombergSP,WebbCO.Picante:Rtoolsforintegratingphylogeniesandecology.Bioinformatics,2010,26(11):1463⁃1464.[29]㊀SwensonNG,EnquistBJ.Opposingassemblymechanismsinaneotropicaldryforest:implicationsforphylogeneticandfunctionalcommunityecology.Ecology,2009,90(8):2161⁃2170.[30]㊀KraftNJB,CornwellWK,WebbCO,AckerlyDD.Traitevolution,communityassembly,andthephylogeneticstructureofecologicalcommunities.TheAmericanNaturalist,2007,170(2):271⁃283.[31]㊀SwensonNG,EricksonDL,MiXC,BourgNA,Forero⁃MontañaJ,GeXJ,HoweR,LakeJK,LiuXJ,MaKP,PeiNC.Phylogeneticandfunctionalalphaandbetadiversityintemperateandtropicaltreecommunities.Ecology,2012,93(sp8):S112⁃125.[32]㊀IngramT,ShurinJB.Trait⁃basedassemblyandphylogeneticstructureinnortheastpacificrockfishassemblages.Ecology,2009,90(9):2444⁃2453.[33]㊀ÅgrenGI,FagerströmT.Limitingdissimilarityinplants:randomnesspreventsexclusionofspecieswithsimilarcompetitiveabilities.Oikos,1984,43(3):369⁃375.[34]㊀MacArthurRH.Populationecologyofsomewarblersofnortheasternconiferousforests.Ecology,1958,39(4):599⁃619.[35]㊀黄建雄,郑凤英,米湘成.不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响.植物生态学报,2010,34(3):309⁃315.[36]㊀ReichPB,UhlC,WaltersMB,EllsworthDS.Leaflifespanasadeterminantofleafstructureandfunctionamong23Amazoniantreespecies.Oecologia,1991,86(1):16⁃24.[37]㊀ReichPB,WaltersMB,EllsworthDS.Fromtropicstotundra:globalconvergenceinplantfunctioning.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1997,94(25):13730⁃13734.[38]㊀LakeJK,OstlingA.Commenton"Functionaltraitsandniche⁃basedtreecommunityassemblyinanAmazonianforest".Science,2009,324(5930):1015⁃1015.[39]㊀KlugeJ,KesslerM,KlugeJ.Phylogeneticdiversity,traitdiversityandniches:speciesassemblyoffernsalongatropicalelevationalgradient.JournalofBiogeography,2011,38(2):394⁃405.[40]㊀Diniz⁃FilhoJAF,BiniLM,RodríguezMÁ,RangelTFLVB,HawkinsBA.Seeingtheforestforthetrees:partitioningecologicalandphylogeneticcomponentsofBergmannᶄsruleinEuropeanCarnivora.Ecography,2007,30(4):598⁃608.[41]㊀KörnerC.Theuseof altitude inecologicalresearch.TrendsinEcology&Evolution,2007,22(11):569⁃574.[42]㊀GoldbergD,NovoplanskyA.Ontherelativeimportanceofcompetitioninunproductiveenvironments.JournalofEcology,1997,85(4):409⁃418.。
吉林蛟河阔叶红松林样地种-面积关系

吉林蛟河阔叶红松林样地种-面积关系陈桂莲;谭凌照;范春雨;张新娜;张春雨;赵秀海【摘要】种-面积关系是群落生态学的核心问题之一,是生物多样性尺度转换的重要依据.利用吉林蛟河阔叶红松林30 hm2的样地数据,采用随机取样与巢式取样方法,分别在10、20、30 hm2尺度上建立对数模型(Logarithmic function)、幂函数模型(Power function)和逻辑斯蒂模型(Logistic function)拟合局域种-面积关系,并利用赤池信息准则(AIC)进行拟合结果优度检验.结果表明,取样方法对种-面积关系的构建有显著影响,随机取样优于巢式取样.种-面积关系的构建与尺度(取样上限)密切相关:在小尺度上(10 hm2),对数模型与逻辑斯蒂模型拟合效果优于幂函数模型;在中尺度和大尺度上(20、30 hm2),相对于对数模型和幂函数模型,逻辑斯蒂模型能更好地拟合阔叶红松林的种-面积关系.据AIC值可知,随机取样下的逻辑斯蒂模型拟合效果最好,是拟合30 hm2阔叶红松林样地种-面积关系的最适模型.因此研究时需要根据区域森林群落的实际情况选择种-面积模型.%The species-area relationship (SAR) is a core component of community ecology,and is an important basis for biological diversity scaling.The SAR is used to describe community types and can solve many ecological problems,such as the determination of minimum sampling areas in a community.Therefore,it is of great importance to diversity conservation.Recently,a number of studies have demonstrated substantial uncertainties in selecting the best SAR model for a data set.In the present study,a 30-hm2 permanent forest plot was established in a broad-leaved Korean pine forest in Jiaohe,Jilin Province,China.All trees with diameters at breast height (DBH) ≥ 1 cm were tagged and the height,DBH,and crown diameter of these trees weremeasured and recorded.We established a logarithmic model,a power function model,and a logistic model using the 30-hm2 sample plot to simulate the SAR of a broad-leaved Korean pine forest.We examined how SARs simulated by logarithmic,power function,and logistic models differed after random sampling or nested sampling methods had been used to collect data,and how this difference was affected by sampling scales (broad,moderate,and fine scales).The Akaike Information Criterion (AIC) value was used to compare the goodness-of-fit for each SAR model.The results showed that the sampling method had a significant influence on the SAR,and that the goodness-of-fit for random sampling was better than that for nested sampling at all sampling scales.The establishment of a species-area relationship was closely related to the sampling scales,and the logarithm and logistic models were superior to the power function model at the fine scale (10 hm2).At the moderate and large scales (20 hm2 and 30 hm2,respectively),the logistic model better fitted the species-area relationship for broad-leaved Korean pine forest than did the logarithm and the power function models..A comparison of the different models showed that the logistic model with random sampling produced an optimal fit for the species-area relationship within the 30 hm2 broad-leaved Korean pine sample area (AIC =76.91),and that the appropriate minimum sampling area was 10 hm2.We concluded that both sampling scale and sampling method had significant influences on the SAR.The scale effect on the SAR is closely related to the community species distribution pattern,and the impacts may result from habitat heterogeneity andsuccessional stage.Habitat heterogeneity and community succession stage might have influenced the number of regional species and species composition,and these different species distribution patterns were reflected in the different SAR curves.Therefore,in practical applications,the variation in the actual community structure and environments within the sampling area should be fully considered.Further work needs to consider the actual situation of the local forest community to simulate the species-area relationship models【期刊名称】《生态学报》【年(卷),期】2017(037)014【总页数】8页(P4770-4777)【关键词】种-面积关系;拟合优度;取样方法;取样尺度【作者】陈桂莲;谭凌照;范春雨;张新娜;张春雨;赵秀海【作者单位】北京林业大学林学院,北京100083;森林资源生态系统过程北京市重点实验室,北京 100083;北京林业大学林学院,北京100083;森林资源生态系统过程北京市重点实验室,北京 100083;北京林业大学林学院,北京100083;森林资源生态系统过程北京市重点实验室,北京 100083;北京林业大学林学院,北京100083;森林资源生态系统过程北京市重点实验室,北京 100083;北京林业大学林学院,北京100083;森林资源生态系统过程北京市重点实验室,北京 100083;北京林业大学林学院,北京100083;森林资源生态系统过程北京市重点实验室,北京 100083【正文语种】中文种-面积关系描述了物种随取样面积的增加而变化的规律,是群落生态学研究的核心问题之一[1]。
草冠净在幼林除草中的应用研究

草灌净在红松幼林除草中的应用何金山1张焕新2(1.吉林市龙潭区杨木林场,吉林吉林 132010 2.蛟河市林业局,吉林蛟河132500)摘要:幼林除草是造林绿化工作中的重要环节,是促进苗木生长的主要措施之一。
本文根据多年的使用经验,论述了草灌净在红松幼林除草的优点、用量及时间。
幼林除草是造林绿化工作中的重要环节,是改善造林地环境,促进苗木生长的主要措施之一。
多年来,我们一直采用人工除草措施进行幼林抚育,为造林苗木的正常生长做了大量的工作,但是人工除草存在着一些不足之处。
一是除草费用相对较高;二是除草过程中易伤害到苗木。
为此从2002年开始我们使用草灌净(一种除草剂)对红松造林地进行除草,现将几年来使用草灌净在红松造林幼林除草的应用情况总结如下:一、成活率及生长量对比从2003年至2007年我们连续做了5年的对比实验。
在立地条件相同的同一块红松造林地,一部分采用草灌净锄草,另一部分按标准采用人工锄草(5年7次),对苗木的成活率和年生长量进行对比。
从调查结果看,使用药剂除草的成活率平均高出人工除草的3%以上,高度年生长量两者几乎相等,看不出差别,这说明,草灌净除草对红松苗木正常生长没有影响。
二、草灌净除草的优点1、除草成本低按照《营造林技术规程》,红松幼林除草一般是5年7次(2、2、1、1、1),按当前使用人工费用,每公顷除草一次需300元,七次则需要2100元。
使用草灌净除草,需用喷洒5次,即每年一次,每次每公顷需草灌净2000毫升,5次共需要10000毫升。
按当前价值看,草灌净每公顷需要费用800元,喷洒药剂人工费用每公顷500元(每次100元,共5次),使用草灌净锄草,每公顷总计需要费用1300元,相对人工除草,每公顷可节约800 元,节约费用比率达38%。
2、除草效果好采用人工除草方式除草,不能从根本上解决草的生长,杂草不但阻挡了苗木的通风和透光,而且还同苗木争夺土地的营养成分,从而影响了苗木的正常生长;另外,人工锄草或多或少都可能损伤到苗木(一般伤苗率为3%-5%),造成苗木的死亡或影响苗木生长。
距离和密度制约对吉林蛟河阔叶红松林幼苗生长的影响

距离和密度制约对吉林蛟河阔叶红松林幼苗生长的影响
吉林蛟河阔叶红松林是中国东北地区重要的生态系统之一,阔叶红松林幼苗的生长状况对于该地区的生态环境保护和可持续发展有着至关重要的作用。
在阔叶红松林幼苗生长过程中,距离和密度制约因素对其生长发育起着重要的影响,下文将对此进行探讨。
首先,距离制约对于幼苗的生长影响很大。
由于距离影响了幼苗之间的竞争和相互作用,合理的距离安排可以促进其生长发育。
在阔叶红松林幼苗的生长过程中,苗木相互之间的距离过于接近会大大限制其生长发育,因为它们需要争夺土壤养分、水分和光线。
过于紧密的种植会使得苗木之间存在极大的竞争,以至于受到抑制。
但是,过度的间距也会对幼苗的生长造成不良的影响。
因为过大的间距会导致土壤水分和其他资源的散失,使得幼苗的生长发育受到限制,难以实现良好的生长状态。
其次,密度制约也是影响幼苗生长的重要因素。
幼苗的密度会直接影响它们之间的相互作用和竞争状况,进而影响其生长速率和生长状态。
阔叶红松林幼苗的种植密度过高会使得整个林区内的生态环境不平衡,导致不良的生长效果。
因此,合理的密度安排可以改善苗木的生长状况,提高其生长发育的质量与速率。
在这方面,可以通过适当调整栽植密度、减少植物种类的数量等方式来解决这个问题。
总体而言,距离和密度制约因素对于吉林蛟河阔叶红松林幼苗的生长发育有着很大的影响。
采取恰当的距离和密度安排方法,可以改善幼苗的生长质量和速率,同时提升阔叶红松林的生态
环境。
这样可以确保吉林蛟河阔叶红松林获得可持续生态发展,成为一个更加繁荣的地区。
观念一转天地新——蛟河市林业转型发展走笔

观念一转天地新——蛟河市林业转型发展走笔作者:暂无来源:《新长征·党建版》 2015年第7期王向新蛟河市位于吉林省东部,长白山西麓,松花湖畔。
全市林业经营总面积45.56 万公顷,活立木总蓄积量2849 万立方米,森林覆盖率58.5%,是吉林省重点林区之一。
林业经济曾是蛟河市的支柱产业,为地方经济发展做出了很大贡献。
然而,在世纪之交,蛟河林业遇到了重大危机,由于多年的过量无序采伐,资源枯竭,经济濒临崩溃,年最高亏损额达2000 余万元。
据统计,2000 年蛟河林业局总负债2.1亿元,60%以上的林业职工下岗,成为全市社会不稳定因素。
面对困境,蛟河林业局党委没有被困难吓倒,他们积极“转观念、调结构、促升级”,牢固树立生态安全和生态文明的战略思想,在经营理念和发展战略上勇于创新,着力打造法制、民生、效能、廉政、创新林业,不断加快林业经济的发展,使全市林业系统走出了低谷,实现了林业经济新的突破。
转变思路方有出路“观念决定出路,思想决定效益。
”面对资源匮乏、生态恶化、债台高筑、人心涣散等难题,蛟河市林业局党委认识到,必须改变经营思想,用市场杠杆撬动债务巨石,把发展的主业从木材生产转到生态建设上来,大力实施封山育林,绿化秀美山川,向生态要效益,用市场调结构。
2005 年,他们在提出跳出山门变林业经济为林区经济的同时,开始林外创业,加速推进传统林业向现代林业转变。
一个名为“五五三二一”的庞大工程开始在全市实施:开展退耕还林、“三北四期”防护林、重点公益林、速生丰产林、松花江流域治理5 项工程。
按照《中共中央、国务院关于加快林业发展的决定》提出跨行业、跨行政、跨地域的方针,成立了吉林省内第一家跨行政、跨行业、跨地域的场圃联合体。
经吉林省人民政府批准,通过企业改革成立注册资金6000 万元的吉林省拉法山国家森林公园股份有限公司,吸纳域内19 个国有场圃为公司股东,职工参股。
相继组建5 个林业分公司,即森华木业公司、天林石材开发分公司、森林旅游分公司、森林经营分公司、森龙经贸开发分公司。
抚育采伐对吉林蛟河针阔混交林幼苗更新的影响

抚育采伐对吉林蛟河针阔混交林幼苗更新的影响吉林蛟河针阔混交林是我国东北地区重要的森林资源之一,近年来受到人类活动的影响,其生态环境已经发生了不可逆转的变化。
在众多的人类活动中,抚育采伐是对该林区影响较大的一种活动,它不仅会对成林的树木产生直接的影响,同时也会对林下幼苗的更新产生一定的影响。
对于林下幼苗的更新,抚育采伐主要有以下几个方面的影响:1. 光照利用率提高:抚育采伐主要是对成林的树木的干预,目的是为了让光线更好地照射到林地内部,使得整个林分能得到足够的光照以生长。
这也为林下幼苗的生长提供了充足光照的保障。
研究发现,抚育采伐后林下的光照强度能够得到提高,林下草本植物和灌木的生长状况得到了较大的改善,这种改变也对于幼苗的更新产生了一定的帮助。
2. 土壤改良:抚育采伐后,伐出的树木可以为林区提供一定的凋落物,这对于林下幼苗的生长以及土壤中的营养物质供应都是有益的。
另外,伐出的树木也可以减少林分的密度,这可以降低林下土壤湿度,避免因长期的潮湿而导致的幼苗死亡。
3. 退化土地的转化:针阔混交林的幼苗更新能力一般,林分的老化和退化也使得林下幼苗的更新变得更加困难。
而抚育采伐能够通过清除枯枝败叶和树干,以及加强种类及年龄结构的调整,促进了退化土地向优良的森林土地转化,增加了幼苗更新的可能性。
4. 生物多样性的保护:抚育采伐对保护针阔混交林的生物多样性也有着积极的影响。
抚育采伐能够减少过度的开发,保持部分林分的完整性,保护了其中的种类多样性;同时,抚育采伐也为保护野生动物提供了栖息和生长的环境,促进了该地区的生态平衡。
因此,抚育采伐虽然会对成林的树木产生一定的影响,但也为该地区的生态环境和幼苗更新提供了诸多的改善。
权衡各方面的利弊,适度的抚育采伐可以成为实现可持续发展的一种有效手段。
林分生长量测定

直接用野外测得的资料整列直径生长量有下 列问题: ①所测得的胸径生长量2L,实际上是去皮胸径 生长量 ②带皮胸径d是期末(t)时的胸径,应与胸径生 n t 长量相对应的期中( )时带皮胸径。 2
二、林分生长的特点(2)
林分的生长发育可分为四个阶段: (一)幼龄林阶段:在此阶段由于林木间尚未发生 竞争,自然枯损量接近于零。所以林分的总蓄积 是在不断增加。 (二)中龄林阶段:发生自然稀疏现象,但林分蓄 积正的生长量仍大于自然枯损量,因而林分蓄积 量仍在增加。 (三)近熟林阶段: 随着竞争的增剧自然稀疏急速 增加,此时林分蓄积的正生长量等于自然枯损量, 反映出林分蓄积量停滞不前。 (四)成、过熟林阶段: 林分蓄积正的生长量小于 枯损量,反映林分蓄积量在下降。
第一节 林分生长量的概念及种类
• 林分生长通常是指它的蓄积的生长量而言,它 是由组成林分的树木材积消长的累积。 • 林分生长过程与树木生长过程截然不同,树木 生长过程属于“纯生”型;而林分生长过程, 由于森林存在自然稀疏现象,所以属“生灭 型”。
• 显然林分生长模型要比树木生长模型复杂的多。
一、森林自然稀疏现象
• 这种方法主要利用胸径的过去定期生长量间接推算蓄积生 长量,并用来预估未来林分蓄积生长量。
• 一次调查法要求:预估期不宜太长、林分林木株数不变。
• 一次调查法不能估计林分枯损量和采伐量。
• 包括:材积差法、林分表法、一元材积指数法、双因素法、 近似生长率法和单木生长率法。
一、材积差法
• 材积差法:将一元材积表中胸径每差lcm的材积 差数,作为现实林分中林木胸径每生长lcm所引 起的材积生长量,利用一次测得的各径阶的直径 生长量和株数分布序列,从而推算林分蓄积生长 量 的方法。 • 应用此法必须具备两个前提条件:一是要有经过 检验而适用的一元材积表;二是要求待测林分期 初与期末的树高曲线无显著差异。 • 用材积差法的步骤(1)胸径生长量的测定和整 列;(2)各径阶株数分布;(3)应用一元材积 表计算蓄积生长量。
吉林蛟河针阔混交林12个树种生物量分配规律

吉林蛟河针阔混交林12个树种生物量分配规律何怀江;叶尔江.拜克吐尔汉;张春雨;左强;邳田辉;高海涛【期刊名称】《北京林业大学学报》【年(卷),期】2016(38)4【摘要】研究生物量分配是了解植物结构与功能的有效手段,对陆地森林生态系统碳循环研究起着重要作用。
本文以吉林省蛟河林业实验区管理局天然次生混交林内12个优势树种为研究对象,探讨了各树种生物量器官(叶、枝、干、根)分配特征及其与个体大小的关系。
结果表明:1)12个树种各器官的相对生长遵循异速生长理论,相对生长关系并不一致。
枝与干(1.091~1.254)、枝与根(1.012~1.158)、根与干(1.015~1.202)以及地下与地上部分(0.991~1.070)近于等速生长,叶与枝(0.655~0.757)、叶与干(0.777~0.931)和叶与根(0.718~0.859)呈现为异速生长。
2)12个树种各器官生物量分配遵循异速生长分配理论,叶、枝、干和根生物量分配比例的范围依次为1.80%~6.54%、13.87%~27.09%、51.12%~65.03%和15.76%~25.52%,各器官生物量分配比例的均值大小表现为:干(57.09%)〉根(21.46%)〉枝(18.59%)〉叶(2.86%)。
根茎比(R/S)范围为0.189~0.355,均值为0.279。
3)各器官生物量分配比例以及R/S均与树种有关,不同树种各器官生物量分配比例以及树种间R/S存在显著差异(P0.05),其他各器官分配比例均与个体大小呈显著相关关系(P〈0.05)。
具体表现为随个体增大,叶和干生物量分配比例显著降低、枝生物量分配比例显著增加(P〈0.05)的趋势。
研究表明:植物各器官在其生长过程中并非都是等速生长,异速生长广泛存在于各器官的生长过程中,同时各器官的生物量分配遵循异速生长分配理论。
为了获得更多的空间和营养,植物在生长过程中遵循最优化分配理论,将更多的资源分配给有利于提高自身竞争力的器官,以达到具有更强竞争力和生产力的目的。
蛟河市林业局蒙古栎纯林生长量初探

蛟河市林业局蒙古栎纯林生长量初探吴慧书;郭彦林【摘要】对吉林省蛟河市辖区蒙古栎纯林十年间生长变化情况进行研究,结果表明该地区蒙古栎中龄林十年间单株平均生长量约为0.059cm,生长量总体服从正态分布,生物量积累缓慢;胸径主要分布区间为5cm~25cm,胸径为10cm~ 15cm的蒙古栎样木共计85株,占样木总数45%以上,为样地内主要样木,样木胸径总体也呈正态分布,和以往研究结果一致.【期刊名称】《林业勘查设计》【年(卷),期】2016(000)002【总页数】2页(P64-65)【关键词】蒙古栎;生长量;胸径【作者】吴慧书;郭彦林【作者单位】吉林省林业调查规划院;吉林省林业调查规划院【正文语种】中文蒙古栎(Quercus mongolica?Fisch.ex Ledeb)属壳斗科、栎属,落叶乔木,高达30m,树皮灰褐色,深纵裂。
蒙古栎为栎中抗逆性较强的一个种,主要分布于我国东北和华北地区,是东北夏绿阔叶林的重要建群种[1]。
地球上天然森林面积日趋减少,木材和林副产品供应也日益紧张,人类生存环境面临严重威胁。
因此对蒙古栎等天然生态系统生物量和生产力的研究意义重大。
1.1 研究地自然概况蛟河市地处吉林省中东部长白山西麓,位于长吉图开发开放先导区腹地和中心节点,东与敦化市相邻,西隔松花湖与吉林市、永吉县相望,南与桦甸市接壤,北与舒兰市、黑龙江省五常市毗连,全市幅员面积6050km2。
蛟河林业局地处松辽平原向长白山过渡地带,地理坐标为东经126°44′~128°00′,北纬43°12′~44°09′之间。
全局林木种类繁多,森林资源丰富,蕴藏有丰富的林产品资源,是吉林省较大的林业资源宝库之一[2]。
研究地位于蛟河市林业局天南林场26林班1小班,样地内平均坡度为26°,土壤质地为暗棕壤,样地中心点地理坐标为东经126°96′、北纬43°82′。
吉林省农业直接补贴效率分析及对策

整体上看 ,我国农业保护与支持路径 比
较单一, 完全是对农业 、 对农民的直接补贴 , 没有通过农业 政策性金融行为 实施 间接补
贴 ,财政资金的支农效率并没有充分发挥 。
2 直接补贴效应分析
无论是 “ 四项直接补贴” 还是 “ , 六小工
・7 ・ 2
此 ,用财政直接补贴提高农 民的收入 ,最
形 OP B 2面积之差 。 2Q
形成的净收益下降幅度 ( 高峰 、王学镇、羊
文辉 ,2 0 。 0 4)
对农 民实行直接补贴 ,即不符合 WT O 相关规则 ,也不符合农业保护国际惯例 , 最
后 必将 造成 巨大的财 政赤字 ,从 长远来 看 ,
/ s
、 / A
不适合我国的国情。对农民实行直接补贴 ,
行 为 的惩罚 力度 ,地方 政府 应 积极支 持和协 助农 发行 打击恶 意逃废 银行 债务 的行 为 。 三是 建立贷 款风 险补偿 机 制 ,发 展规模 大 、信誉 好 的担 保机 构 。可 以采取政 府 出资
4 强化 “ . 1 政府与农发行的合作机制”
农 发行作 为唯一 的农业 政策性 银行 ,以 贯彻 、配合政 府农业社 会经 济政 策或意 图为 宗 旨 ,是政府 在市 场上 的延伸 。 因此 ,农发 行与政 府 的合 作 ,能够促进 农业 生产结 构 的
农村中小学教育和农村 医疗体系实施了财政
投入 ,依靠 省财政 实施 了 “ 变 肉”工程牧 粮
Q l Q 2 Q
业小区补贴 ,依靠省财政和地方政府的财政 力量对农民实施了农业保险补贴和新农合医
疗保 险补贴 ,另外 ,还有少 许 的机 电产 品下
乡补贴 。
吉林省蛟河地区天然林林木株数与直径结构研究

2017 年第 10 期(下半月)农民致富之友 Nong Min Zhi Fu Zhi You110科研◎林业科学吉林省蛟河地区天然林林木株数与直径结构研究潘丽铭1 宋彩民2 王峰洁2在蛟河辖区内布设样地一块,对样地内样木进行调查研究,结果表明样地内共有样木60株,其中白桦最多为19株,约占样木总数的30%,水曲柳次之15株,其余样木种类依次为假色槭、春榆、山槐、杂木、山丁子,根据样木种类对样木胸径值进行分类统计,统计结果表明该研究地内样木胸径均值最大的样木组为白桦,白桦的胸径均值为20.3cm ,其它样木组胸径均值从大到小依次为山丁子、山槐、水曲柳、春榆、假色槭。
随着科学技术的日益发展和成熟,中国的改革成果遍布世界,“一带一路”的新思维正在改变着人类生存的每一个角落,人类正在进入世界经济高速发展的另一个新时期,物质水平日益丰富的同时精神生活的追求也成了人类更加迫切的愿望,自然保护区、森林公园、城市公园如雨后春笋般因需而出,而且各大公园人满为患,为忙碌的现代人群提供了游憩休闲、散步谈心的理想场所,森林中清新的空气、超高的负离子含量也为游人的身心健康做出了巨大贡献。
因此,森林的各项指标研究也逐渐成为很多科研工作者的重要趋势,天然林的直径结构是最基本的林分结构,不仅因为林分直径便于测定,而且因为它直接影响树木的树高、干形、材积、材种及树冠等因子的变化,能为许多森林经营技术及测树制表提供理论依据,受到了很多科研工作者的青睐,但主要以城市和自然保护区为主,本文以天然林为研究对象对天然林直径结构进行研究和分析,旨在为天然林在林业领域的研究提供基础。
1 研究地自然概况本次研究地位于吉林省蛟河林业实验区管理局,研究地中心点地理坐标为东经:127°36′00′′,北纬:43°56′00′′,研究地周边环境优美,森林资源丰富,大多以天然次生林为主,由于研究地内交通不便人烟稀少,且经吉林省批准长期作为吉林省北华大学林学院多学科林业学生的实习基地,因此植被保护较为完整,人为破坏较少,是科研工作的理想场所。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
03%,净 生长 率分别 为 20%和 2 5 .6 .8 . %,净 0
增率分别为 0 4 . %和 0 2 5 . %。 4
表 2 蛟河 林业局 林分 各优 势树种 ( )消长率 表 组
平 均 红松林
12 .9 59 .7 00 .4
.
云杉林 落叶松林 臭松林 水 曲柳林 胡桃楸林 椴树林
当代生态农业 2 1 年第 3 期 01 、4
蛟 河林 业局 林 分 生长 量 ( ) 变化 研 究 率 的
辛成群 张义涛
( 吉林 省蛟 河市林 业局 ,蛟 河 12 0 ) 3 50
摘要 : 根据某林业局 19 年和 20 97 07年两期地理信息 系统属性数据库及报表资源,局级 固定
样 地 的复查数 据 ,将全局 森林 生长量 ( ) 体统 计分析 ,得 出森林资 源的林 分 生长量 ( ) 率 整 率
6
表 1展示 , 出结 论如下 : 龄组 来看 , 得 从 幼龄林 长 势 比较 快 , 均总 生长率 为 6 年 .l 3 %,
长率 为 26 %,净增 率为 1 1 .6 . %;近 、成 过 5 熟林 各个 生长 率非 常接 近 ,年 均总 生长率 分
1
(
J
保留生长率为 3 2 进界生长率为 21%, . %, 9 . 1
分 生 长量 ( ) 率 指标 的准确性 和科学 性 。 经 过上 述一 系列 的调查和 比较 ,本 文将
技术上保证数据分析应用的综合性,信息利
用充 分 。利用 地 理信息 系统特有 的空 间分析
功能 ,将同一地区不同时段的资源环境数据
进行 空 间分 析操作 ,并通 过分析 研究林 分生
大学 出版社. 0 . 2 7 0 8
学, 0 () 2 89 0 [ 陈颂 东. 5] 日本农业保护的经验值得借 鉴[】 Jl
财 经科 学, 0 () 2 82 0
[ 高峰, 学镇, 2] 王 羊文辉6 ・
当代生态农业
2 1 年第 3 期 01 、4
3
资 源 变 化分 析 [ . 蒙 古 林 业 调 查 设 J内 】
计 .0 11:4~3 2 0 () 3 5
[ 林媚珍, 2] 张镜铿. 海南岛森林 资源 的动态变 化 及 其 可 持 续 发 展 [ . 态 科 J 生 】
学 . 0 , () 2 9 2 01 4: ~8 0 9 8
的 变化 的 结 论 。
关键词 :林 分
生长量 ( ) 率
统计分析 变化
作者简 介 :辛成群 ( 92 )中级工程 师 ,主要 从 事林 业经 营管理 等工作 16 一
本研究利用地理信息系统把森林资源数
据通过 空 间与属性 的分类进 行统 一管理 ,在
局 级 固定 样地 的复查 数据 ,绘制 统计 分析 图 表 及地 理信 息系统成 果 图对 汪 清林 业 局 的森 林 资 源 现 状 及 动态 变 化 情 况 进 行 了 分 析研 究 。除无非林 地外 ,参加 消长 量计算 的样地 6 4块 、样 木 662株 ,保证 了本研 究 的林 9 99
长量 ( )变化 的规律 。 率
根据蛟 河林 业局 19 7年和 20 两期 9 07年 地理 信息 系统属 性数据 库及报 表资源 ,结合 19 年和 20 年 二类调 查时设 置 的 19 块 7 9 07 40
、 。 / ’、 / ‘、
‘ 一 、
_ I,
全局森林生长量 ( ) 率 整体统计出来( 见表 1
版 )19 , () 5 ,9 92 4 : 7~3 2 2 3 9
47 %, 曲柳林 的总生 长率 为 49 %,云杉 .5 水 .8 林 的 总生长率 为 37% ,胡 桃楸林 的总 生长 . 6 率为 2 3 . %,椴 树林 的总生 长率 为 29%, 4 . 2 柞树林 的总生 长率为 25%,榆树 林 的总生 . 4 长率 为 5 4 . %,色树林 的总生 长 率为 42 %, 3 . 7 枫桦林 的总生 长率为 26%, 白桦林 的总生 . 3 长率 为 2 4 . %,杨树林 的总 生长率 为 3 6 7 . %, 2
的理论分析【 . J 农业经济 问题,048 ] 2 0()
[ ]何广 文. 3 对农村政策金 融改革的理性思考
【. J 农业经济问题,043 】 2 0 ()
[ 4]贾万 军, 王寒菊.百信农 民资金互助 社金
融 运 作 效 率 分 析 [ . 徽 农 业 科 J 安 ]
参考文献
[ ]高 鸿业. 1 西方经济学[ . 京:中国人 民 M] 北
其 次是人工 落 叶松 林 和红松林 ,年均 总生长 率 分别为 68%和 60%;同时 ,云杉林 的 .1 . 7
总生 长 率 为 37 % ,臭松 林 的 总 生 长 率 为 . 6
[] 3 邱扬, 马正岩, 屯. 张金 山西森林 资源 的动态
变化 分 析 【 . J 山西 大 学 学 报 ( ] 自然 科 学
净生 长率 为 57 %,净增 率为 49%;其 次 .8 .7
别为 2 0 . %和 27%,保 留生长率分 别为 7 . 3
20 %和 1 7 ,进 界 生 长 率 为 01%和 .1 .% 5 .8
是中龄林 ,年均总生长率为 3 7 . %,保留生 2 长率为 2 9 . %,进界生长率为 0 3 6 . %,净生 3
O1 .4
19 .9 34 .4
13 .6
13 - 15 .4 4 1 .5
柞树林
榆树林
・77 ・
当代生态农业
2 1 年第 3 期 01 、4
从 表 2中可 以得 出结 论 如下 :从 优势树 种 ( ) 看 ,人 工樟 子松林 和人工 云杉林 组 来
年 均总生长 率最大 , 别为 95 %和 89 %; 分 . 7 .9
和表 2 ,用来 分 析森林 资源 的增长情 况 。 )
、 /●、 / 。、 / ’ 、 —
, 、 , 、 / ’ 、 / ’ 、
h-一,
’ 、
/ ’、
/ 。 、 、 — ,’ 、
,
理 的 比价 ,充分 考虑调 整种植 结构 的需 要 。 从长 期来看 ,保 护价应 实现 由保 护粮食 品种 到保 护优 良品种 的转变 ,促进 粮食 品质结构 的调整 。