DNA甲基化的特征 PPT
合集下载
甲基化表观遗传学PPT

That’s That s
all!
度甲基化的部分DNA序列被大的非甲基化DNA序列所分隔 开
4.难以被DNA修复系统所识别: CG→TG是可遗传的
2.DNA的甲基化的过程 的甲基化的过程
3.DNA甲基化的检测 甲基化的检测
1.传统实验方法 Methylation-sensitive restriction enzymes Methylation-specific enzyme McrBC 2.现代方法 MeDIP: methylated DNA immunoprecipitation assay MBD:methylation binding domain 3.DNA甲基化位点的确定:Bisulfite genomic sequencing
(二)真核生物的DNA甲基转移酶 真核生物的 甲基转移酶
1. 哺乳动物 DNMT1,DNMT3A,DNMT3B,DNMT3L, 哺乳动物: DNMT2 2. 拟南芥:DRM2, MET1,DNMT2, CMT3 拟南芥: , 3. 粗糙脉孢菌 粗糙脉孢菌(Neurospora crassa): DIM2,dim-5, RID
1.DNA的甲基化 的甲基化 1. DNA甲基化的模式:
(1) 线虫:无甲基化的胞嘧啶 (2) 果蝇:极少量的甲基化胞嘧啶,识别模式CpT (主要) vs. CpG (极少) (3) 哺乳动物:CpG (~70%)或者CpNpG
2.CpG– Cytosine phosphate Guanine 3. 镶嵌的甲基化(mosaic methylation): 基因组中,高
(a) EGCG: 绿茶中的聚苯复合物 ) 小分子,阻遏Dnmt的活性中心 (b)RG108: 小分子,阻遏 ) 的活性中心 (c) 4-amino-benzoic acid衍生: 衍生: 衍生 Procainamide (d) Psammaplins (e) 寡聚核苷酸机理(如图) 抑制剂的抑制机理(如图)
DNA甲基化

表观遗传学
第二章
DNA甲基化
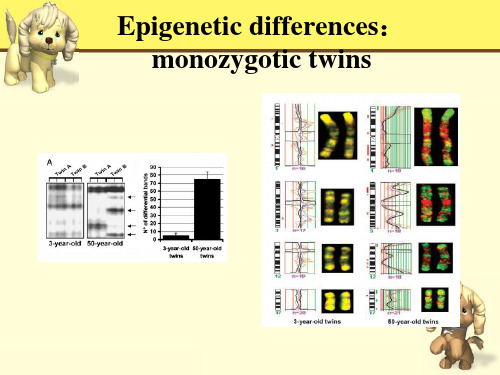
Epigenetic differences: monozygotic twins
简单地把DNA甲基化理解为“一把锁”,凡是被 DNA甲基化标记的部分,大都是需要被“尘封”“监 禁”的基因,比如基因组的“捣蛋鬼”—转座子,就 是被甲基化这把“锁”管制着,失去管制或管制不严, 这些“捣蛋鬼”会在基因组里跳来跳去,把基因组搞 得一团糟,会引起很多问题,如肿瘤、精神疾病等。
CpG岛的甲基化(1)
1. CpG岛:富含CpG区域,长度500~1000bp,GC含量超 过55% 2. 非随机出现:~60%的编码基因的5’UTR区域(转录起 始区域)含有CpG岛。 3. CpG的含量: (1) CG出现的期望值(百分比):1/16 = 6.25% (2) 观察值:很少(~1%) (3) 原因:CG具有很高的突变率 4. C-T转换率是其他碱基对转换率的10-40倍以上 5. 例如:人类肿瘤细胞中的p53基因,50%的点突变都发 生在CpG上。
C––> 5mC ––> T?
Deamination 去氨基化反应
基因组甲基化的特点:
可逆性——许多甲基化位点可以根据细胞活性的要 求重新甲基化或去甲基化; 组织特异性——不同的组织细胞具有不同的甲基化 模式,为基因表达设定程序。
异常的甲基化可分为高甲基化和低甲基化 ,前 者指正常组织中不发生甲基化的位点被甲基化 ,后 者是指在正常组织中发生甲基化的位点去甲基化。
DNA甲基转移酶:系统发育分析
哺乳动物的DNA甲基转移酶
DNMT1: maintenance methyltransferases DNMT3A & DNMT3B: de novo methyltransferases–胚胎移植过程中高表达
第二章
DNA甲基化
Epigenetic differences: monozygotic twins
简单地把DNA甲基化理解为“一把锁”,凡是被 DNA甲基化标记的部分,大都是需要被“尘封”“监 禁”的基因,比如基因组的“捣蛋鬼”—转座子,就 是被甲基化这把“锁”管制着,失去管制或管制不严, 这些“捣蛋鬼”会在基因组里跳来跳去,把基因组搞 得一团糟,会引起很多问题,如肿瘤、精神疾病等。
CpG岛的甲基化(1)
1. CpG岛:富含CpG区域,长度500~1000bp,GC含量超 过55% 2. 非随机出现:~60%的编码基因的5’UTR区域(转录起 始区域)含有CpG岛。 3. CpG的含量: (1) CG出现的期望值(百分比):1/16 = 6.25% (2) 观察值:很少(~1%) (3) 原因:CG具有很高的突变率 4. C-T转换率是其他碱基对转换率的10-40倍以上 5. 例如:人类肿瘤细胞中的p53基因,50%的点突变都发 生在CpG上。
C––> 5mC ––> T?
Deamination 去氨基化反应
基因组甲基化的特点:
可逆性——许多甲基化位点可以根据细胞活性的要 求重新甲基化或去甲基化; 组织特异性——不同的组织细胞具有不同的甲基化 模式,为基因表达设定程序。
异常的甲基化可分为高甲基化和低甲基化 ,前 者指正常组织中不发生甲基化的位点被甲基化 ,后 者是指在正常组织中发生甲基化的位点去甲基化。
DNA甲基转移酶:系统发育分析
哺乳动物的DNA甲基转移酶
DNMT1: maintenance methyltransferases DNMT3A & DNMT3B: de novo methyltransferases–胚胎移植过程中高表达
DNA甲基化与CpG岛

DNA甲基化与CpG岛
• 引言 • DNA甲基化的功能 • CpG岛的特性与分布 • DNA甲基化与CpG岛的关系 • DNA甲基化与CpG岛的研究方法 • DNA甲基化与CpG岛的前景与展望
01
引言
DNA甲基化的定义
DNA甲基化是指在DNA序列中,CpG位点的胞嘧啶被甲基所 修饰的过程。这种修饰是一种重要的表观遗传学标记,对基 因表达和细胞分化等生物学过程具有重要影响。
甲基化与失活
在X染色体失活过程中,DNA甲基化在X染色体上广泛发生,导致相关基因沉默 和X染色体整体失活。这种甲基化模式有助于维持X染色体失活的稳定性和遗传性 。
03
CpG岛的特性与分布
CpG岛的识别标准
01
CpG密度高
CpG岛内CpG位点的密度显著高于 周围序列。
启动子关联
CpG岛通常与基因的启动子区域相 关联。
该方法使用特异性抗体富集甲基化的DNA片段, 然后进行高通量测序,以识别甲基化位点。
该方法使用甲基化结合蛋白(MBD蛋白)富集甲 基化的DNA片段,然后进行高通量测序,以识别 甲基化的CpG岛。
06
DNA甲基化与CpG岛的前景与展望
在疾病诊断和治疗中的应用
肿瘤甲基化检测
通过检测肿瘤组织中DNA的甲基化状态 ,有助于肿瘤的早期诊断和预后评估。
在基因组印记和X染色体失活过程中, DNA甲基化起到关键作用,通过甲基 化特定基因或基因组区域,使这些基 因或区域沉默,不参与基因表达。
基因组印记
印记基因
基因组印记是指某些基因在不同细胞类型中的表达存在差异,这些差异由DNA 甲基化水平决定。印记基因通常在发育过程中由父本或母本来源的等位基因选 择性表达。
亚硫酸氢盐测序
• 引言 • DNA甲基化的功能 • CpG岛的特性与分布 • DNA甲基化与CpG岛的关系 • DNA甲基化与CpG岛的研究方法 • DNA甲基化与CpG岛的前景与展望
01
引言
DNA甲基化的定义
DNA甲基化是指在DNA序列中,CpG位点的胞嘧啶被甲基所 修饰的过程。这种修饰是一种重要的表观遗传学标记,对基 因表达和细胞分化等生物学过程具有重要影响。
甲基化与失活
在X染色体失活过程中,DNA甲基化在X染色体上广泛发生,导致相关基因沉默 和X染色体整体失活。这种甲基化模式有助于维持X染色体失活的稳定性和遗传性 。
03
CpG岛的特性与分布
CpG岛的识别标准
01
CpG密度高
CpG岛内CpG位点的密度显著高于 周围序列。
启动子关联
CpG岛通常与基因的启动子区域相 关联。
该方法使用特异性抗体富集甲基化的DNA片段, 然后进行高通量测序,以识别甲基化位点。
该方法使用甲基化结合蛋白(MBD蛋白)富集甲 基化的DNA片段,然后进行高通量测序,以识别 甲基化的CpG岛。
06
DNA甲基化与CpG岛的前景与展望
在疾病诊断和治疗中的应用
肿瘤甲基化检测
通过检测肿瘤组织中DNA的甲基化状态 ,有助于肿瘤的早期诊断和预后评估。
在基因组印记和X染色体失活过程中, DNA甲基化起到关键作用,通过甲基 化特定基因或基因组区域,使这些基 因或区域沉默,不参与基因表达。
基因组印记
印记基因
基因组印记是指某些基因在不同细胞类型中的表达存在差异,这些差异由DNA 甲基化水平决定。印记基因通常在发育过程中由父本或母本来源的等位基因选 择性表达。
亚硫酸氢盐测序
表观遗传学-DNA甲基化

DEAMINATION
Deamination: 去氨基化反应
An enzyme to remove it from DNA: uracil-Nglycosylase.
Epigenetics, 2008-2009, Semester 1, USTC
DNA甲基化的检测
1. 传统实验方法
Methylation-sensitive restriction enzymes Methylation-specific enzyme McrBC
DNA甲基化抑制基因转录的机制
Long-term silencing
Epigenetics, 2008-2009, Semester 1, USTC
直接干扰机制 (1)
Epigenetics, 2008-2009, Semester 1, USTC
直接干扰机制 (2)
Epigenetics, 2008-2009, Semester 1, USTC
1. 转座子的甲基化 2. 人类基因组~45%的区域是转座子 3. 高度甲基化,细胞中90%的甲基化CpG位于转 座子中 4. 转座子的活性:对机体非常有害 5. 宿主防御模型:抑制转座子的活性
Epigenetics, 2008-2009, Semester 1, USTC
转座子导致的PEV
CpG岛的甲基化 (1)
1. CpG岛:富含CpG区域,长度500~1000bp,GC 含量超过55% 2. 非随机出现:~60%的编码基因的5’UTR区域 (转录起始区域)含有CpG岛。 3. CpG的含量:
(1) CG出现的期望值(百分比):1/16 = 6.25% (2) 观察值:很少(~1%) (3) 原因:CG具有很高的突变率
生物学DNA甲基化
Epigenetic differences: monozygotic twins
简单地把DNA甲基化理解为“一把锁”,凡是被 DNA甲基化标记的部分,大都是需要被“尘封”“监 禁”的基因,比如基因组的“捣蛋鬼”—转座子,就 是被甲基化这把“锁”管制着,失去管制或管制不严, 这些“捣蛋鬼”会在基因组里跳来跳去,把基因组搞 得一团糟,会引起很多问题,如肿瘤、精神疾病等。
生在CpG上。
CpG岛的甲基化(2)
1. CpG在重复片段以及基因的3’UTR区域能够发生 甲基化
2. CpG岛通常不被甲基化 3. 哺乳动物中:26,000-45,000 CpG岛。常分布在持
家基因和一些组织表达特异性基因的启动子区域
4. CpG岛:可以被HpaII酶(C|CGG)切成小片段,因 此也叫HTF岛
Bisulfite Modification
测序: 非甲基化的C将被测序成T,反链为A
结合重亚硫酸盐的限制性内切酶法 (combined bisulfiterestriction analysis,
酵母与果蝇基因组中未能检测到任何甲 基化CpG,这两种生物并不依赖DNA甲基化 的方式来控制基因活性,它们采用其它的机 制来达到同一目的。
脊椎动物与高等植物普遍利用DNA甲基 化作为重要的调控机制。
DNA甲基化的分子机理
指在 DNA 甲基转移酶 (DNMTs)的作用下, 以 S -腺苷甲硫氨酸 ( SAM )为甲基供体,将 甲基添加在 DNA分子中的碱基上。
2.胞嘧啶甲基化后产生5-甲基化胞嘧啶能够自发的脱氨基形成 胸腺嘧啶(Thymine):5mC -> T
3. 哺乳动物中,~1%的DNA碱基能够发生甲基化修饰
4. DNA甲基化的分布:
简单地把DNA甲基化理解为“一把锁”,凡是被 DNA甲基化标记的部分,大都是需要被“尘封”“监 禁”的基因,比如基因组的“捣蛋鬼”—转座子,就 是被甲基化这把“锁”管制着,失去管制或管制不严, 这些“捣蛋鬼”会在基因组里跳来跳去,把基因组搞 得一团糟,会引起很多问题,如肿瘤、精神疾病等。
生在CpG上。
CpG岛的甲基化(2)
1. CpG在重复片段以及基因的3’UTR区域能够发生 甲基化
2. CpG岛通常不被甲基化 3. 哺乳动物中:26,000-45,000 CpG岛。常分布在持
家基因和一些组织表达特异性基因的启动子区域
4. CpG岛:可以被HpaII酶(C|CGG)切成小片段,因 此也叫HTF岛
Bisulfite Modification
测序: 非甲基化的C将被测序成T,反链为A
结合重亚硫酸盐的限制性内切酶法 (combined bisulfiterestriction analysis,
酵母与果蝇基因组中未能检测到任何甲 基化CpG,这两种生物并不依赖DNA甲基化 的方式来控制基因活性,它们采用其它的机 制来达到同一目的。
脊椎动物与高等植物普遍利用DNA甲基 化作为重要的调控机制。
DNA甲基化的分子机理
指在 DNA 甲基转移酶 (DNMTs)的作用下, 以 S -腺苷甲硫氨酸 ( SAM )为甲基供体,将 甲基添加在 DNA分子中的碱基上。
2.胞嘧啶甲基化后产生5-甲基化胞嘧啶能够自发的脱氨基形成 胸腺嘧啶(Thymine):5mC -> T
3. 哺乳动物中,~1%的DNA碱基能够发生甲基化修饰
4. DNA甲基化的分布:
甲基化与肿瘤(共38张PPT)

雌激素、孕酮、雄激素和泌乳刺激素受体异常 甲基化发生在乳腺和子宫肿瘤中,可能导致这些 癌细胞不对此类激素发生应答。表现出视黄酸受 体β2和细胞视黄醇结合蛋白Ⅰ的启动子过甲基化的肿 瘤可能废止类维生素A的分化作用。
细胞因子信号
几条细胞因子通路负调控中的发现均暗示细胞因 子信号抑制剂〔SOCS〕家族蛋白的存在,特别是 转录活性转录体Jak/STAT通路的受体相关酪氨酸/ 信号变化器和激活剂。在人类癌症中, SOCS-1和 SOCS-3经历了甲基化相关的沉默。
Gene expression is affected by nearby DNA sequences called promoters, which often include CpG islands. When MeCP2 binds to the CpG island in a promoter (bottom), associated proteins act to prevent expression of the gene.
组蛋白甲基化功能
组蛋白甲基化与异染色质形成 组蛋白甲基化与基因印记
组蛋白甲基化与X染色体失活
组蛋白甲基化与异染色质形成
SUV39H1和SUV39H2甲基转移酶对异染色质的形成具有 重要作用。将两个基因突变后,H3K9的甲基化会减少一 半,这样出生的小鼠有丝分裂中染色体的别离有缺陷。在 裂殖酵母中H3K9位点的甲基化可以将常染色质和异染色 质在特定区域分开。在形成异染色质的过程中,Su〔var〕 3-9通过与异染色质蛋白1〔HP1〕相互作用来控制蛋白质 的定位。
〔3〕DNA甲基化通过改变染色质结构,抑制基因表达。 最近,有两项在大鼠7号染色体中的研究都发现组蛋白甲基化可能是除了DNA甲基化以外维持基因印记的另一种重要途径。 A、 CpG岛主要位于基因的启动子区,局部位于基因的第一个外显子区; 几条细胞因子通路负调控中的发现均暗示细胞因子信号抑制剂〔SOCS〕家族蛋白的存在,特别是转录活性转录体Jak/STAT通路的受体相关酪氨酸 /信号变化器和激活剂。
细胞因子信号
几条细胞因子通路负调控中的发现均暗示细胞因 子信号抑制剂〔SOCS〕家族蛋白的存在,特别是 转录活性转录体Jak/STAT通路的受体相关酪氨酸/ 信号变化器和激活剂。在人类癌症中, SOCS-1和 SOCS-3经历了甲基化相关的沉默。
Gene expression is affected by nearby DNA sequences called promoters, which often include CpG islands. When MeCP2 binds to the CpG island in a promoter (bottom), associated proteins act to prevent expression of the gene.
组蛋白甲基化功能
组蛋白甲基化与异染色质形成 组蛋白甲基化与基因印记
组蛋白甲基化与X染色体失活
组蛋白甲基化与异染色质形成
SUV39H1和SUV39H2甲基转移酶对异染色质的形成具有 重要作用。将两个基因突变后,H3K9的甲基化会减少一 半,这样出生的小鼠有丝分裂中染色体的别离有缺陷。在 裂殖酵母中H3K9位点的甲基化可以将常染色质和异染色 质在特定区域分开。在形成异染色质的过程中,Su〔var〕 3-9通过与异染色质蛋白1〔HP1〕相互作用来控制蛋白质 的定位。
〔3〕DNA甲基化通过改变染色质结构,抑制基因表达。 最近,有两项在大鼠7号染色体中的研究都发现组蛋白甲基化可能是除了DNA甲基化以外维持基因印记的另一种重要途径。 A、 CpG岛主要位于基因的启动子区,局部位于基因的第一个外显子区; 几条细胞因子通路负调控中的发现均暗示细胞因子信号抑制剂〔SOCS〕家族蛋白的存在,特别是转录活性转录体Jak/STAT通路的受体相关酪氨酸 /信号变化器和激活剂。
DNA甲基化检测技术ppt课件

MS-HRM检测方法及步骤
4. DNA样本的甲基化修饰:
可使用FFPE Tissue Kit (QIAGEN) 对DNA甲基化修饰及回收;
3. PCR扩增:
a. 上样体系 b. PCR-HRM反应程序
MS-HRM检测方法及步骤
a. 上样体系:
反应体系组分(20μL) 10×PCR Buffer(15 mM mgcl2)
癌
与DNA 修复与转录激活有关 周期素依赖性蛋白激酶抑制剂
乳腺癌、卵巢癌 GIT 、头与颈部瘤、NHL、肺癌
钙/钙调素-依赖的丝氨酸/苏氨酸磷酸化酶; 凋亡抑制 肺癌
E-cadherin ER GSTP1 hMLH1
MGMT P15
增强增殖、侵袭与转移 激素抵抗 失去对致癌物活性代谢产物的解毒作用 缺损DNA错配修复,基因点突变
DNA甲基化的形式:
DNA甲基化主要形成5-mC和少量的N6-甲基嘌呤(N6-mA)及7-甲基鸟嘌 呤(7-mG)
CpG 岛(CpG Island)
在基因组的某些区域中,通常是基因的启动子区域,5’端非翻译 区和第一个外显子区,CpG序列密度非常高,超过均值5倍以上,成为 鸟嘌呤和胞嘧啶的富集区,称之为CpG岛(CpG Islands, CGIs)。
DNA甲基化与肿瘤的关系
MGMT基因在许多肿瘤中被认为是抗肿瘤药物治疗的预测标记。MGMT启 动子肿瘤特异性甲基化,可以抑制MGMT蛋白的活性,从而使得肿瘤细胞对 烷化类的抗肿瘤药物敏感,因而被广泛用于肿瘤化疗治疗。
Figure: Kaplan–Meier Estimates of Overall Survival, According to MGMT Promoter Methylation Status.
24-第10章 基因组表观遗传-DNA甲基化

抑制基因表达 Dnmts(DNA甲基化酶)打靶CpG位点甲基化DNA的胞嘧啶。 甲基化DNA被甲基化结合蛋白MBD识别,然后招募组蛋白修饰酶如组蛋 白脱乙酰化酶(histone deacetylases ,HDAC)使组蛋白去乙酰基化,同 时 也可招募组蛋白甲基转移酶( histone methyltrans –ferases ,HMT)使 组 蛋白甲基化。 随后招募染色质重建复合物SUV39H1,促使染色质收缩 沉默。
受精胚与体细胞胚的发育差异
哺乳动物体细胞
胚胎与受精卵胚
胎发育存在许多
差异. 这些差异主 要表现在体细胞
胚与受精胚基因
组的程序化. 受精 卵的雄核与雌核
基因组DNA在去 甲化的程序上存
在时间差. 雄核 基因组DNA去甲 基化先于雌核基
因组DNA.基因组 DNA甲基化程序 的差异会导致胎
发育异常.
谢谢!
siRNA
介 导
DNA
甲基化
植物染色质沉默的调控机制 siRNA介导的转座子和重复 序 列甲基化的可能模式。由Pol IV和Pol V产生的siRNA 转 移到AGO4。AGO4然后与DNA甲基化酶等因子互作在靶 DNA位置甲基化。DRM2:DOMAINSREARRANGED METHYLTRANSFERASE 2,重新甲基化酶。 Nat Rev Genet .11:204-220,2010
ቤተ መጻሕፍቲ ባይዱ
分化
5-氮脱氧胞嘧啶是胞嘧啶的类似物, 当加入的5-氮脱氧胞嘧啶添加到培养 的小鼠细胞中,在DNA进入复制时, 5-氮脱氧胞嘧啶可取代胞嘧啶。在下 一轮复制时可消除原有的DNA甲基化, 促使基因表达。 Hpa II是一个可 以识别-CCGG-的DNA限制性内切酶,但对甲基化的-C5mCGG-敏感,可 从HpaII的酶切条带的变化可以探知基因组DNA甲基化的状态。培养的小 鼠 细胞在5-氮脱氧胞嘧啶处理后可检测到HpaII酶切带型的改变。
受精胚与体细胞胚的发育差异
哺乳动物体细胞
胚胎与受精卵胚
胎发育存在许多
差异. 这些差异主 要表现在体细胞
胚与受精胚基因
组的程序化. 受精 卵的雄核与雌核
基因组DNA在去 甲化的程序上存
在时间差. 雄核 基因组DNA去甲 基化先于雌核基
因组DNA.基因组 DNA甲基化程序 的差异会导致胎
发育异常.
谢谢!
siRNA
介 导
DNA
甲基化
植物染色质沉默的调控机制 siRNA介导的转座子和重复 序 列甲基化的可能模式。由Pol IV和Pol V产生的siRNA 转 移到AGO4。AGO4然后与DNA甲基化酶等因子互作在靶 DNA位置甲基化。DRM2:DOMAINSREARRANGED METHYLTRANSFERASE 2,重新甲基化酶。 Nat Rev Genet .11:204-220,2010
ቤተ መጻሕፍቲ ባይዱ
分化
5-氮脱氧胞嘧啶是胞嘧啶的类似物, 当加入的5-氮脱氧胞嘧啶添加到培养 的小鼠细胞中,在DNA进入复制时, 5-氮脱氧胞嘧啶可取代胞嘧啶。在下 一轮复制时可消除原有的DNA甲基化, 促使基因表达。 Hpa II是一个可 以识别-CCGG-的DNA限制性内切酶,但对甲基化的-C5mCGG-敏感,可 从HpaII的酶切条带的变化可以探知基因组DNA甲基化的状态。培养的小 鼠 细胞在5-氮脱氧胞嘧啶处理后可检测到HpaII酶切带型的改变。
DNA甲基化
Dnmt3a & Dnmt3b
表达特异性: Dnmt3a: ubiquitous Dnmt3b: 前额,眼睛
Dnmt3a & Dnmt3b
对哺乳动物的发育至关重要
三、DNA去甲基化
1. DNA去甲基化 (DNA demethylation): 5甲基 胞嘧啶 (5mC) 替代成胞嘧啶的过程 2. 两种方式
MET1 DNMT2
CMT3 DIM2 RID
拟南芥 拟南芥
拟南芥 粗糙脉孢菌 粗糙脉孢菌
Maintenance (CpG) 未知
Maintenance (CpNpG) de novo (CpN) RIP 必须
哺乳动物DNA甲基化途径
启动子的甲基化
(a) 未知蛋白质因子X失 去,DNMT结合,促使 启动子甲基化 (b) 聚合酶失去, DNMT结合->甲基化 (c) 某些转录因子招募 DNMT到启动子区域, 促使甲基化 (d) 组蛋白甲基转移酶 (HMT)招募DNMT…
直接干扰机制 (2)
间接机制
The methyl-CpGbinding proteins MeCP1 and MeCP2能 够与甲基化的DNA结合 MeCP2能够招募Sin3a, HDACs,形成复合物, 阻遏转录
二、DNA甲基转移酶
哺乳动物的DNA甲基转移酶
DNMT1: maintenance methyltransferases DNMT3A & DNMT3B: de novo methyltransferases – 胚胎移植过程中高表达
4. C-T转换率是其他碱基对转换率的10-40倍以上 5. 例:人类肿瘤细胞中的p53基因,50%的点突 变都发生在CpG上。
DNA甲基化

• 组蛋白H3 N端尾部作用 酿酒酵母研究系统,在本身不存在甲基化的酵母 基因组上建立DNA甲基化谱式,组蛋白H3 N端尾 部对于DNA甲基化起着不可或缺的作用。
进一步研究发现,辅助因子Dnmt3能通过其PHD结构域 与第四位赖氨酸未甲基化的组蛋白H3发生相互作用,进 而招募DNA甲基转移酶Dnmt3a到靶位点发生起始性DNA 甲基化。
Thank You!
– 呈现上述现象的基因称为印记基因(imprinted gene), 它又可分为父系印记基因(父源印记,母源表达)和母系 印记基因(母源印记,父源表达)。
DNA甲基化与基因印记
• 印记基因发生的机制尚待深人研究,但一般认为主要由于 来自双亲等位基因被甲基化而导致沉默,即DNA CpG岛 的胞嘧啶5’位置上被加上甲基。
• 甲基化与X染色体失活
• 在雌性哺乳动物,剂量补偿是通过一条X染色体的失活来实现的,由X染 色体失活中心(Xic)控制。Xic区域有两个与X染色体失活相关的基因Xist 和Tsix,两者的动态表达在X染色体失活中起着重要作用,X染色体失活 前,Xist和Tsix是共同表达的,X染色体失活开始后,Tsit停止表达。
DNA甲基化与基因印记
H3K9 甲基化与转录基因沉默 在组蛋白尾部众多的赖氨酸残基甲基化中,H3K9甲基化是基因转录沉默的标 志。
最近Weinberg等 发现针对EF1 A启动子siRNA双链和反义链均能通过组蛋白 H3K9甲基化介导基因沉默。甲基化的H3K9可募集HP1到染色质,HP1与多 种转录抑制因子结合,抑制基因转录。
• 哺乳动物中,这种反义调节可能代表着较为普遍的长距离转录调控机制 ,但调节Xist和Tsix的表达的详细机制仍不清楚。通过对失活和有活性的 X染色体DNA甲基化状态的研究发现,在失活的染色体上,大部分基因 的CpG是甲基化的,在活化的染色体是非甲基化的。活化染色体与非活 化染色体甲基化状态的不同只代表着甲基化在X染色体失活中的一种晚 发现象,它对失活状态的维持具有重要作用,但没有始动作用。
进一步研究发现,辅助因子Dnmt3能通过其PHD结构域 与第四位赖氨酸未甲基化的组蛋白H3发生相互作用,进 而招募DNA甲基转移酶Dnmt3a到靶位点发生起始性DNA 甲基化。
Thank You!
– 呈现上述现象的基因称为印记基因(imprinted gene), 它又可分为父系印记基因(父源印记,母源表达)和母系 印记基因(母源印记,父源表达)。
DNA甲基化与基因印记
• 印记基因发生的机制尚待深人研究,但一般认为主要由于 来自双亲等位基因被甲基化而导致沉默,即DNA CpG岛 的胞嘧啶5’位置上被加上甲基。
• 甲基化与X染色体失活
• 在雌性哺乳动物,剂量补偿是通过一条X染色体的失活来实现的,由X染 色体失活中心(Xic)控制。Xic区域有两个与X染色体失活相关的基因Xist 和Tsix,两者的动态表达在X染色体失活中起着重要作用,X染色体失活 前,Xist和Tsix是共同表达的,X染色体失活开始后,Tsit停止表达。
DNA甲基化与基因印记
H3K9 甲基化与转录基因沉默 在组蛋白尾部众多的赖氨酸残基甲基化中,H3K9甲基化是基因转录沉默的标 志。
最近Weinberg等 发现针对EF1 A启动子siRNA双链和反义链均能通过组蛋白 H3K9甲基化介导基因沉默。甲基化的H3K9可募集HP1到染色质,HP1与多 种转录抑制因子结合,抑制基因转录。
• 哺乳动物中,这种反义调节可能代表着较为普遍的长距离转录调控机制 ,但调节Xist和Tsix的表达的详细机制仍不清楚。通过对失活和有活性的 X染色体DNA甲基化状态的研究发现,在失活的染色体上,大部分基因 的CpG是甲基化的,在活化的染色体是非甲基化的。活化染色体与非活 化染色体甲基化状态的不同只代表着甲基化在X染色体失活中的一种晚 发现象,它对失活状态的维持具有重要作用,但没有始动作用。
解读DNA甲基化

演讲结束!
中国测序论坛
18
中国测序论坛
中国测序论坛
3、DNA甲基化与表观遗传学
• 基因组印迹的分子机理与甲基化密切相关。DNA 甲基化模式的维持对基因印迹在亲子代之间的遗 传是必须的。 • 生长发育过程中,雌性哺乳类动物细胞中的两条 X染色体其中一条失去活性的现象。 X染色体上存 在一个与X染色体失活有密切联系的核心部位。 核心区命名位X染色体失活中心。失活的染色体 上DNA序列都呈高度甲基化,导致绝大多数基因 转录处于关闭状态。
肿瘤的去甲基化治疗
中国测序论坛
DNA甲基化程度依赖于DNMT活性。正常甲
基化模式的建立需要DNMT1和DNMT3的共同作用, DNMT1是DNMT3启动CpG核苷酸从头甲基化的保证,而 DNMT3则使甲基化水平稳定在正常需要水平(去甲基化
)
—— 抑制DNMT活性药物是治疗肿瘤的新希望
中国测序论坛
DNA甲基化的生物学功能
1. 2. 3. 4. 5. DNA甲基化与遗传物质的稳定性 DNA甲基化与基因表达调控 DNA甲基化与表观遗传学 DNA甲基化与胚胎发育 DNA甲基化与肿瘤
中国测序论坛
1、DNA甲基化与遗传物质的稳定性
a) 研究证明细菌DNA复制起始与DNA甲基化以及DNA 与细菌质膜的相互作用有关。DNA便甲基化作为一 种标签决定了复制起始点与细胞膜的结合,控制了 复制起始,使得DNA复制与细胞分裂保持一致。 b) DNA错配修复(mismatch repair)作为细胞增殖过 程中纠正DNA复制错误的重要手段,对保证DNA复 制的忠实性与基因组的稳定性起重要作用。复制后 双链DNA在短期内(数分钟)保持半甲基化状态, 错配修复系统从而能够区分“旧链”与“新链’”, 为校正新链中掺入的错误碱基提供了理想的分子标 记。
相关主题
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
DNA甲基化的特征
一.DNA甲基化与羟甲基化
1.胞嘧啶 2.第五个C原子上 3.甲基或羟甲基的修饰
二.DNA甲基化分布
1. DNA甲基化发生在CpG dinucleotides
低频率! 42% GC ,
在人基因组中的频率为1%,而含有 理论值上为(0.21 * 0.21 = 4.41
%)
ቤተ መጻሕፍቲ ባይዱ
2. CpG island
Gene: CpG transcription start sites (TSS), cis-
regulatory elements
四. DNMTs-功能
大家应该也有点累了,稍作休息
大家有疑问的,可以询问和交流
四. DNMTs-结构
五.TET家族蛋白-功能 (5mC
5hmC)
五.TET蛋白-结构
I.不被甲基化
II.长度为500-2000bp
III.常分布在看家基因和特异性基因启动子区
二.DNA甲基化的分布
1 基因内甲基化抑制转录延伸,但与基因表达为正相关。 2 CpG island 不被甲基化可能与使得基因维持可以转录状态有关
二.DNA甲基化分布的特殊情况
1.在非CpG环境下的低水平甲基化 (功能未知)
ES cells, oocytes, and brain (不发生DNA复制)
2.启动子中CpG island 发生甲基化
imprinted and pluripotency-related genes (长期转录
沉默)
三.DNA羟甲基化的分布
cell: ES cells(0.03%) , Purkinje neurons(0.59%)
Demethylation :
imprinting loci, gene bodies, intergenic regions, mobile elements, CpG island promoters
意义: DNA甲基化使得减数分裂 相关的基因沉默,去甲基 化可以使其恢复功能。
八.Global demethylation in erythropoiesis
六.Reprogramming of DNA methylation during preimplantation development
第一次卵裂前
几次卵裂后
不发生Reprogramming of DNA methylation:
七.Reprogramming of DNA methylation in gametes
一.DNA甲基化与羟甲基化
1.胞嘧啶 2.第五个C原子上 3.甲基或羟甲基的修饰
二.DNA甲基化分布
1. DNA甲基化发生在CpG dinucleotides
低频率! 42% GC ,
在人基因组中的频率为1%,而含有 理论值上为(0.21 * 0.21 = 4.41
%)
ቤተ መጻሕፍቲ ባይዱ
2. CpG island
Gene: CpG transcription start sites (TSS), cis-
regulatory elements
四. DNMTs-功能
大家应该也有点累了,稍作休息
大家有疑问的,可以询问和交流
四. DNMTs-结构
五.TET家族蛋白-功能 (5mC
5hmC)
五.TET蛋白-结构
I.不被甲基化
II.长度为500-2000bp
III.常分布在看家基因和特异性基因启动子区
二.DNA甲基化的分布
1 基因内甲基化抑制转录延伸,但与基因表达为正相关。 2 CpG island 不被甲基化可能与使得基因维持可以转录状态有关
二.DNA甲基化分布的特殊情况
1.在非CpG环境下的低水平甲基化 (功能未知)
ES cells, oocytes, and brain (不发生DNA复制)
2.启动子中CpG island 发生甲基化
imprinted and pluripotency-related genes (长期转录
沉默)
三.DNA羟甲基化的分布
cell: ES cells(0.03%) , Purkinje neurons(0.59%)
Demethylation :
imprinting loci, gene bodies, intergenic regions, mobile elements, CpG island promoters
意义: DNA甲基化使得减数分裂 相关的基因沉默,去甲基 化可以使其恢复功能。
八.Global demethylation in erythropoiesis
六.Reprogramming of DNA methylation during preimplantation development
第一次卵裂前
几次卵裂后
不发生Reprogramming of DNA methylation:
七.Reprogramming of DNA methylation in gametes