Pixantrone_dimaleate_HNMR_12806_MedChemExpress
中国肾移植受者巨细胞病毒感染临床诊疗指南(2023_版)

· 指南与共识·中国肾移植受者巨细胞病毒感染临床诊疗指南(2023版)中华医学会器官移植学分会 中国医师协会器官移植医师分会 中国医疗保健国际交流促进会肾脏移植学分会 【摘要】 近几年在实体器官移植(SOT )受者巨细胞病毒(CMV )感染诊疗领域,无论是诊断方法还是新型抗CMV 药物都有了一些新的进展,对CMV 感染的诊治产生了积极的影响。
为了进一步规范中国肾移植术后CMV 感染的管理,中华医学会器官移植学分会组织了国内多个学科相关领域专家,参考《中国实体器官移植受者巨细胞病毒感染诊疗指南(2016版)》和国内外已发表的最新文献和指南,制定了《中国肾移植受者巨细胞病毒感染诊疗指南(2023版)》,新版指南更新了CMV 流行病学,CMV 感染的危险因素和普遍性预防的研究进展,新增CMV 感染定义,细化CMV 血症和CMV 病的诊断标准,并对新型抗CMV 药物进行了介绍。
【关键词】 肾移植;实体器官移植;巨细胞病毒;感染;病毒血症;巨细胞病毒病;普遍性预防;抢先治疗【中图分类号】 R617, R373 【文献标志码】 A 【文章编号】 1674-7445(2024)03-0001-20Clinical diagnosis and treatment guidelines for cytomegalovirus infection in kidney transplant recipients in China (2023edition) Branch of Organ Transplantation of Chinese Medical Association, Branch of Organ Transplantation Physician of Chinese Medical Doctor Association, Branch of Kidney Transplantation of China International Exchange and Promotive Association for Medical and Health Care. *The First Affiliated Hospital of Xi 'an Jiaotong University , Xi 'an 710061, China Correspondingauthors:DingXiaoming,Email:***************.cnXueWujun,Email:******************.cn【Abstract 】 In recent years, there have been significant advances in the diagnosis and treatment of cytomegalovirus (CMV) infection in solid organ transplant (SOT) recipients, including diagnostic method and anti-CMV drugs. These advancements have had a positive impact on the management of CMV infection in SOT recipients. To further standardize the management of CMV infection after kidney transplantation in China, Branch of Organ Transplantation of Chinese Medical Association organized a multidisciplinary group of experts in relevant fields. They referred to the ‘Diagnosis and Treatment Guidelines for Cytomegalovirus Infection in Solid Organ Transplant Recipients in China (2016 edition)’ and the latest published literature and guidelines, resulting in the development of the ‘Clinical Diagnosis and Treatment Guidelines for Cytomegalovirus Infection in Kidney Transplant Recipients in China (2023 edition)’. The updated guideline includes CMV epidemiology, research progress on the risk factors and universal prevention of CMV infection, the definition for CMV infection, detailed diagnostic criteria for CMV viremia and CMV disease, as well as an introduction to new anti-CMV drugs.【Key words 】 Kidney transplantation; Solid organ transplantation; Cytomegalovirus; Infection; Viremia;Cytomegalovirus disease; Universal prevention; Preemptive therapyDOI: 10.3969/j.issn.1674-7445.2024096基金项目:国家自然科学基金(82370802、82170766、82270789、81970646);陕西省卫生健康肾脏移植科研创新平台(2023PT-06)执笔作者单位: 710061 西安,西安交通大学第一附属医院(丁小明);首都医科大学附属北京友谊医院(林俊);首都医科大学附属北京朝阳医院(胡小鹏);复旦大学附属中山医院(戎瑞明);西安交通大学第一附属医院(郑瑾)通信作者:丁小明,Email :***************.cn ;薛武军,Email :******************.cn第 15 卷 第 3 期器官移植Vol. 15 No.3 2024 年 5 月Organ Transplantation May 2024 巨细胞病毒(cytomegalovirus,CMV)是一种全球传播广泛的β-疱疹病毒,原发感染之后在体内会呈潜伏状态,当人体的免疫功能下降时病毒会被再激活。
Nipro 产品数据册:单用途翼型肾脏细胞注射器说明书

H O S P I T A L P R O D U C T WING CATHSTERILE, SINGLE-USE WINGED CATHETER WITH INJECTION PORT FOR IV INFUSIONPRODUCT DATA SHEET2PRODUCT COMPLIANCE• CE-marked, Class IIa Medical Devices, Rule 7, MDD 93/42/EEC, UMDNS 10727• Complies with the following norms, directives, and regulations: ―ISO 594-1: 1986 ―EN 980:2008 ―EN 1041:2008 ―EN 1707:1996 ―ISO 7864:2016―ISO 9626:2016 ―EN ISO 10555-1:2013 ―EN ISO 10993-1/4/5/7/11 ―ISO 10993-10:2010 ―EN ISO 11135-1:2014―EN ISO 11607-1:2009 /AMD 2014 ―EN ISO 13485:2016 ―EN ISO 14971: 2012• Labels contain 12 languagesMANUFACTURING DETAILSLegal manufacturer: Nipro Corporation Country of origin: ThailandSTERILIZATION AND SHELF LIFESterilizationSingle-use only Indicator stickerShelf lifeEtO (Ethylene oxide)Each outer box (or shipping carton) contains a chemical indicator that indicates sterility. This indicator comes in the form of a [blue] sticker that turns[red] when sterilized.5 yearsWING CATHSTERILE, SINGLE-USE WINGED CATHETER WITH INJECTION PORT FOR IV INFUSION• Ultra sharp three-beveled needle to minimize discomfort • Radiopaque• Injection port with universal luer fitting • Flexible wings for easy grip• Color-coded wings and hub to indicate gauge• Siliconization of needle minimizes penetration and gliding force• T ransparent ETFE catheter tube for quick visualization of blood in the flashback chamber to facilitate insertion • Flashback in hub means the needle is correctly in the vein • Flashback in catheter means the catheter is correctly in the vein • Latex-free, DEHP-free, PVC-free • Available in 16-18-20-22-24 G• IV infusion duration: hours to several days • For use by healthcare professionals onlyClass IIa Medical DevicesRule 7MDD 93/42/EEC UMDNS 1072701233MATERIALS USEDDEHP-free Latex-free PVC-free PRODUCT RANGE OVERVIEWPACKAGING DETAILSLanguages12 languages on inner and outer box:English (EN), French (FR), Dutch (NL), German (DE), Spanish (ES), Italian (IT), Portuguese (PT), Greek (EL), Swedish (SV), Danish (DA), Norwegian (NO), and Finnish (FI)GlueMaterial: Acrylic resinNeedle (cannula)Material:Stainless steel SUS-304Needle hub Material:Polycarbonate (PC)Catheter hubMaterial: Polypropylene (PP)Port capMaterial: Polyethylene (PE)Catheter tubeMaterial: Ethyltetrafluoroethylene (ETFE)Cutting angleThree-beveledLuer capMaterial: Polypropylene (PP)Cap connectorMaterial: Polypropylene (PP)Caulking pinMaterial: Stainless steel SUS-304One-way valveMaterial: Silicone tubeLubricantMaterial: Silicone PolydimethylsiloxaneMembrane filterMaterial: Random micro glass filterVent fittingMaterial: Polypropylene (PP)Catheter wingMaterial: Polypropylene (PP)Nipro Medical Europe : European Headquarters, Blokhuisstraat 42, 2800 Mechelen, Belgium T: +32 (0)15 263 500 | F: +32 (0)15 263 510 |***********************| D a t a s h e e t - W i n g _C a t h - T H A - 27.N o v 2019Label detailsTransport conditionsClosed and dryStorage conditions Open the packaging only immediately before use to guarantee sterility.。
THE INTERNATIONAL JOURNAL OF MEDICAL ROBOTICS AND COMPUTER ASSISTED SURGERY Int J Med Robot

Introduction
Computer-assisted surgery (CAS) is a methodology that translates into accurate and reliable image-to-surgical space guidance. Neurosurgery is a very complex procedure and the surgeon has to integrate multi-modal data to produce an optimal surgical plan. Often the lesion of interest is surrounded by vital structures, such as the motor cortex, temporal cortex, vision and audio sensors, etc., and has irregular configurations. Slight damage to such eloquent brain structures can severely impair the patient (1,2). CASMIL, an imageguided neurosurgery toolkit, is being developed to produce optimum plans resulting in minimally invasive surgeries. This system has many innovative features needed by neurosurgeons that are not available in other academic and commercial systems. CASMIL is an integration of various vital modules, such as rigid and non-rigid co-registration (image–image, image–atlas and
开启片剂完整性的窗户(中英文对照)

开启片剂完整性的窗户日本东芝公司,剑桥大学摘要:由日本东芝公司和剑桥大学合作成立的公司向《医药技术》解释了FDA支持的技术如何在不损坏片剂的情况下测定其完整性。
太赫脉冲成像的一个应用是检查肠溶制剂的完整性,以确保它们在到达肠溶之前不会溶解。
关键词:片剂完整性,太赫脉冲成像。
能够检测片剂的结构完整性和化学成分而无需将它们打碎的一种技术,已经通过了概念验证阶段,正在进行法规申请。
由英国私募Teraview公司研发并且以太赫光(介于无线电波和光波之间)为基础。
该成像技术为配方研发和质量控制中的湿溶出试验提供了一个更好的选择。
该技术还可以缩短新产品的研发时间,并且根据厂商的情况,随时间推移甚至可能发展成为一个用于制药生产线的实时片剂检测系统。
TPI技术通过发射太赫射线绘制出片剂和涂层厚度的三维差异图谱,在有结构或化学变化时太赫射线被反射回。
反射脉冲的时间延迟累加成该片剂的三维图像。
该系统使用太赫发射极,采用一个机器臂捡起片剂并且使其通过太赫光束,用一个扫描仪收集反射光并且建成三维图像(见图)。
技术研发太赫技术发源于二十世纪九十年代中期13本东芝公司位于英国的东芝欧洲研究中心,该中心与剑桥大学的物理学系有着密切的联系。
日本东芝公司当时正在研究新一代的半导体,研究的副产品是发现了这些半导体实际上是太赫光非常好的发射源和检测器。
二十世纪九十年代后期,日本东芝公司授权研究小组寻求该技术可能的应用,包括成像和化学传感光谱学,并与葛兰素史克和辉瑞以及其它公司建立了关系,以探讨其在制药业的应用。
虽然早期的结果表明该技术有前景,但日本东芝公司却不愿深入研究下去,原因是此应用与日本东芝公司在消费电子行业的任何业务兴趣都没有交叉。
这一决定的结果是研究中心的首席执行官DonArnone和剑桥桥大学物理学系的教授Michael Pepper先生于2001年成立了Teraview公司一作为研究中心的子公司。
TPI imaga 2000是第一个商品化太赫成像系统,该系统经优化用于成品片剂及其核心完整性和性能的无破坏检测。
Differentiation of Human Pluripotent Stem Cells into Retinal Cells

87M.A. Hayat (ed.), Stem Cells and Cancer Stem Cells, Volume 6,DOI 10.1007/978-94-007-2993-3_9, © Springer Science+Business Media B.V . 20129A bstractRetinal and macular degeneration disorders are characterized by a progressive loss of photoreceptors, which causes visual impairment and blindness. In some cases, the visual loss is caused by dysfunction, degen-eration and loss of underlying retinal pigment epithelial (RPE) cells and the subsequent death of photoreceptors. The grim reality is that there is no successful treatment for most of these blindness disorders. Cell therapy aimed at replenishing the degenerating cells is considered a potential ther-apeutic approach that may delay, halt or perhaps even reverse degenera-tion, as well as improve retinal function and prevent blindness in the aforementioned conditions. Human embryonic stem cells (hESC) and induced pluripotent stem cells (iPSCs) may serve as an unlimited donor source of photoreceptors and RPE cells for transplantation into degenerat-ing retinas and for retinal disease modeling.I ntroductionThe vertebrate eyes form as bilateral evaginations of the forebrain, called optic vesicles (Martínez-Morales et al. 2004 ; Fig. 9.1a ). During develop-ment, the optic vesicles begin to invaginate to form a cup-shaped structure, the optic cup. The inner, thicker neural layer of the optic cup differ-entiates into the neural retina, and the outer, thin-ner pigmented layer forms the retinal pigmentepithelium (RPE). At the early developmental stages, the neuroepithelial cells that compose the optic vesicle are morphologically and molecu-larly identical and are all able to give rise to neu-ral retina and RPE. Exogenous signals coming from the adjacent tissues, including factors from the fi broblast growth factor (FGF) and transform-ing growth factor beta (TGF b ) families, dictate the fate of these cells. The mature vertebrate ret-ina is comprised of six types of neurons and one type of glia (the Müller glia). These seven cell types constitute three nuclear layers: retinal gan-glion cells in the ganglion cell layer (GCL); the horizontal, bipolar and amacrine interneurons, and Müller glial cells in the inner nuclear layer (INL); and rod and cone photoreceptors in the outer nuclear layer (ONL; Harada et al. 2007;M . I delson • B . R eubinoff (*)T he Hadassah Human Embryonic Stem Cell Research Center, The Goldyne Savad Institute of Gene Therapy & The Department of Obstetrics and Gynecology , H adassah University Medical Center ,E in Kerem 12000 ,J erusalem 91120 ,I srael e -mail: b enjaminr@ekmd.huji.ac.il D ifferentiation of HumanPluripotent Stem Cells into Retinal Cells Masha Idelson and Benjamin Reubinoff88M. Idelson and B. ReubinoffFig. 9.1b ). The photoreceptor cells capture lightphotons and transform their energy into electrical signals by a mechanism called phototransduction. The visual pigment which is utilized in this process is located on membranal discs in the outer seg-ments of photoreceptors. The outer segments are continuously renewed: the old discs are shed and new disks form. When the photoreceptors absorb light, they send the signal through the retinal interneurons to the ganglion cells which transmit the electrical impulse to the brain by their axons forming the optic nerve. Rods are responsible for night vision, whereas cones are responsible for color vision and detecting fi ne details. The macula is a small part of the retina which is rich in cones and responsible for detailed central vision.R PE cells that compose the outer layer of the optic cup are pigmented cuboidal cells which lie between the neural retina and the choriocapil-laris, which include the blood vessels supplying the retina. The multiple villi on their apical side are in direct contact with the outer segments ofextraocular mesenchymeabneural retinalensoptic nerveoptic cupsurface ectodermRPEFGFoptic vesiclechoroidBM RPE cone ONLINL GCLlightHC BC MC ACONrod F ig. 9.1 D evelopment and structural arrangement of the retina. ( a ) Schematic representation of retinal development including the transition from optic vesicle to optic cup and retinal patterning. ( b ) Schematic diagram of retinal cells arrangement and connections. A bbreviations :A C amacrinecell, B C bipolar cell, B M Bruch’s membrane, G CL gan-glion cell layer, H C horizontal cell, I NL inner nuclear layer, M C Müller cell, O N optic nerve, O NL outer nuclear layer89 9 Differentiation of Human Pluripotent Stem Cells into Retinal Cellsthe photoreceptor cells; on their basal side, the RPE is in contact with the underlying basal mem-brane, termed Bruch’s membrane that separates the RPE from the choroid. These cells play cru-cial roles in the maintenance and function of the retina and its photoreceptors. As a layer of pig-mented cells, the RPE absorbs the stray light that was not absorbed by the photoreceptors. The RPE cells form a blood–retinal barrier due to decreased permeability of their junctions. The RPE cells transport ions, water, and metabolic end products from the retina to the bloodstream. They are involved in supplying the neural retina with nutrients from the bloodstream, such as glu-cose, retinol, and fatty acids. Another important function of the RPE is the phagocytosis of shed photoreceptor outer segments. After the outer segments are digested, essential substances such as retinal are recycled. Retinal is also recycled and returned to photoreceptors by the process known as the visual cycle. The precise functioning of the RPE is essential for visual performance. Failure of one of these functions can lead to degeneration of the retinal photoreceptors, vision impairment and blindness.T here are many inherited and age-related eye disorders that cause degeneration of the retina as a consequence of loss of photoreceptor cells. Retinal and macular degeneration disorders can be divided into two main groups. The fi rst group primarily affects the photoreceptors and involves the majority of cases of retinitis pigmentosa. In the second group, the primary damage is to the adjacent RPE cells, and as a consequence of this damage, the photoreceptors degenerate. This group includes age-related macular degeneration, Stargardt’s macular dystrophy, a subtype of Leber’s congenital amaurosis in which RPE65 is mutated, Best’s disease and some cases of retini-tis pigmentosa, as well.W ith regard to retinitis pigmentosa (RP), it is a group of inherited retinal degeneration diseases that are caused, as mentioned above, by a primary progressive loss of rod and cone photoreceptors, followed by a subsequent degeneration of RPE (Hartong et al. 2006). The disease affects approxi-mately 1.5 million patients worldwide and is the most common cause of blindness in people under 70 years of age in the western world. The disease can be characterized by retinal pigment deposits visible on the fundus examination. In most cases, the disease primarily affects rods. At later stages of the disease, the degeneration of cones takes place. As a consequence of disease progression, the patients’ night vision is reduced. Patients initially lose peripheral vision while retaining central vision (a visual status termed “tunnel vision”). In advanced cases, central vision is also lost, commonly at about 60 years of age. The disease affects about 1 in 4,000. The inheritance can be autosomal-recessive, autosomal-dominant or X-linked (in ~50–60%, 30–40%, and 5–15% of cases, respectively). Mutations in more than 140 genes have been iden-tifi ed as causing RP (Hartong et al. 2006).Among these genes are those involved in phototransduc-tion, like rhodopsin, the a- and b- subunits of phos-phodiesterase, the a- and b- subunits of Rod cGMP gated channel and arrestin. The additional muta-tions were found in genes encoding structural pro-teins, like peripherin, rod outer segment protein and fascin. They were also found in transcription factors involved in photoreceptors’ development such as Crx and Nrl, and in other genes, whose products are involved in signaling, cell-cell interac-tion and trafficking of intracellular proteins. Currently, there is no effective cure for RP. Treatment with vitamin A palmitate, omega-3 fatty acids and other nutrients may somewhat slow the rate of the disease progression in many cases. Reduction in exposure to light was also shown to decrease the rate of retinal degeneration.A mong the group of retinal degenerations that are caused by primary loss of RPE cells or their function, age-related macular degeneration (AMD) is the most frequent condition and the leading cause of visual disability in the western world (Cook et al. 2008).Among people over 75 years of age, 25–30% are affected by AMD, with progressive central visual loss that leads to blindness in 6–8%. The retinal degeneration pri-marily involves the macula. The dry form of AMD is initiated by hyperplasia of the RPE and formation of drusen deposits, consisting of meta-bolic end products underneath the RPE or within the Bruch’s membrane. It may gradually progress into the advanced stage of geographic atrophy90M. Idelson and B. Reubinoff with degeneration of RPE and photoreceptorsover large areas of the macula causing central visual loss. Ten percent of dry AMD patients will progress to neovascular (wet) AMD, with blood vessels sprouting through the Bruch’s membrane with subsequent intraocular leakage and/or bleed-ing, accelerating the loss of central vision. While the complicating neovascularization can be treated with anti-VEGF agents, currently there is no effective treatment to halt RPE and photore-ceptor degeneration and the grim reality is that many patients eventually lose their sight (Cook et al. 2008).S targardt’s macular dystrophy (SMD) is the most common form of inherited macular dystro-phy affecting children (Walia and Fishman 2009). The disease is symptomatically similar to AMD. The prevalence of SMD is about 1 in 10,000 chil-dren. The disease involves progressive central visual loss and atrophy of the RPE beneath the macula following accumulation of lipofuscin in RPE cells, which is suggested to consist of non-degradable material, derived from ingested pho-toreceptor outer segments. The inheritance is predominantly autosomal recessive, although an autosomal dominant form has also been described. The mutation in the ABCA4 gene was found to be a most common cause of SMD. The product of the ABCA4 gene is involved in energy transport to and from photoreceptors. The mutated protein cannot perform its transport function and, as a result, photoreceptor cells degenerate and vision is impaired. Currently, there is no effective treat-ment for SMD.C ell therapy to replenish the degenerating cells appears as a promising therapeutic modality that may potentially halt disease progression in the various retinal and macular degeneration dis-orders caused by loss and dysfunction of RPE cells and photoreceptors (da Cruz et al. 2007).I n this chapter we will discuss the potential of human pluripotent cells which includes human embryonic stem cells (hESC) and induced pluripotent stem cells (iPSCs), to gen-erate various types of retinal cells that could be used for transplantation therapy of retinal degen-eration disorders and disease modeling for drug discovery. C ell Therapy of Retinal and Macular DegenerationsT he eye is an attractive organ for cell therapy as it is easily accessible for transplantation and for simple monitoring of graft survival and potential complications by direct fundoscopic visualiza-tion. Anatomically, it is a relatively confi ned organ limiting the potential of unwanted extra-ocular ectopic cell distribution, and a low number of cells are required to replenish the damaged cells. The eye is also one of the immune privi-leged sites of the body.T he concept of replacing dysfunctional or degenerated retina by transplantation has been developing ever since the fi rst retina-to-retina transplant in 1986 (Turner and Blair 1986).In most studies, primary retinal immature (fetal) tissue has been used as donor material. It was demonstrated that such transplants can survive, differentiate, and even establish connections with the host retina to a limited degree (Ghosh et al. 1999). The subretinal transplantation of healthy RPE has some advantages over neural retinal transplantation, as it concerns only one cell type that is not involved in neural networking. Transplantation of RPE has been studied exten-sively in animal models (Lund et al. 2001).The most commonly used animal model of retinal degeneration is the Royal College of Surgeons (RCS) rat model, in which primary dysfunction of the RPE occurs as a result of a mutation in the receptor tyrosine kinase gene M ertk(D’Cruz et al. 2000). This leads to impaired phagocytosis of shed photoreceptor outer segments, with sec-ondary degeneration and progressive loss of pho-toreceptors within the fi rst months of life. It was reported that rat and human RPE cells rescued photoreceptor cells from degeneration when transplanted into the subretinal space of RCS rats (Li and Turner 1988; Coffey et al. 2002).The ability of transplanted RPE cells to restore retinal structure and function has been demonstrated in clinical trials. In humans, autologous transplanta-tions of peripheral RPE as well as macular trans-locations onto more peripheral RPE provide a proof that positioning the macula above relatively91 9 Differentiation of Human Pluripotent Stem Cells into Retinal Cellshealthier RPE cells can improve visual functionin AMD patients (Binder et al. 2004; da Cruz et al. 2007). Nevertheless, the surgical procedures for autologous grafting are challenging and are often accompanied by signifi cant complications. In addition, autologous RPE transplants may carry the same genetic background, environmen-tal toxic and aging-related effects that may have led to macular RPE failure and the development of AMD in the patient. It is also problematic to use autologous cells when all the RPE cells are damaged. Cell sources that can be used for such therapy include allogeneic fetal and adult RPE (Weisz et al. 1999; Binder et al. 2004; da Cruz et al. 2007). However, the use of fetal or adult retinal tissues for transplantation is severely lim-ited by ethical considerations and practical prob-lems in obtaining sufficient tissue supply. The search for a cell source to replace autologous RPE such as immortalized cell lines, umbilical cord-derived cells as well as bone marrow-derived stem cells continues.T he derivation of hESCs more than a decade ago has raised immense interest in the potential clinical use of the cells for regeneration (Thomson et al. 1998; Reubinoff et al. 2000).Along the years, signifi cant progress has been made towards the use of hESCs in clinical trials.T he other promising source of cells for transplantation therapy is iPSCs that are simi-lar to hESCs in their stemness characteristics and pluripotency. These cells could be gener-ated from different human somatic cells by transduction of four defi ned transcription fac-tors: Oct3/4, Sox2, Klf4, and c-Myc (Takahashi et al. 2007).G eneration of RPE and neural retina from hESCs and iPSC has numerous advantages, as it can be done from pathogen-free cell lines under good manufacturing practice (GMP) conditions with minimal variation among batches. Such cells can be characterized extensively prior to preclinical studies or for clinical applications, and an unlimited numbers of donor cells can be generated from them. In the following para-graphs, strategies for induction of differentiation of hESCs and iPSCs towards RPE and neural retina fate are reviewed. D ifferentiation into Retinal Pigment EpitheliumI t was reported for the fi rst time in mice and pri-mates that the differentiation of ES cells into RPE could be induced by co-culture with PA6 stromal cells (Kawasaki et al. 2002; Haruta et al. 2004). The resulting cells had polygonal epithelial mor-phology and extensive pigmentation. The cells expressed the markers that are characteristic of RPE. They developed typical ultrastructures and exhibited some functions of RPE. The differenti-ation of hESC into RPE was first reported by Klimanskaya et al. (2004).According to their protocol, hESCs underwent spontaneous differ-entiation by overgrowth on mouse embryonic fibroblasts (MEF), in feeder-free conditions or, alternatively, as embryoid bodies (EBs) in com-bination with withdrawal of bFGF from the medium. The yield of the formation of RPE cells after 4–8 weeks of spontaneous differentiation was relatively low; for example,<1% of EBs con-tained pigmented cells at this stage. However, after 6–9 months in culture, all the EBs contained pigmented cells. The areas of pigmented cells could be further isolated mechanically and prop-agated by passaging as RPE lines. Klimanskaya and colleges characterized the hESC-derived RPE cells by transcriptomics and demonstrated their higher similarity to primary RPE tissue than to human RPE lines D407 and ARPE-19. The low yield of spontaneously differentiating RPE cells was improved by induction of differentia-tion with Wnt and Nodal antagonists, Dkk1 and LeftyA, respectively, the factors that are sug-gested to promote retinal differentiation. This treatment gave rise to pigmented cells within 38% of the hESC colonies after 8 weeks (Osakada et al. 2008). Immunostaining with the ZO-1 anti-body showed that by day 120, hESC-derived pig-mented cells formed tight junctions (about 35% of total cells). We showed that differentiation toward the neural and further toward the RPE fate could be augmented by vitamin B3 (nicotin-amide; Idelson et al. 2009).We further showed that Activin A, in the presence of nicotinamide, effi ciently induces and augments differentiation92M. Idelson and B. Reubinoffinto RPE cells. This is in line with the presumed role of Activin A in RPE development i n vivo .In the embryo, extraocular mesenchyme-secreted members of the TGF b superfamily are thought to direct the differentiation of the optic vesicle into RPE (Fuhrmann et al. 2000).Under our culture conditions, when the cells were grown in suspen-sion as free-fl oating clusters, within 4 weeks of differentiation, 51% of the clusters contained pigmented areas and about 10% of the cells within the clusters were pigmented. When we modifi ed the differentiation conditions to includea stage of monolayer culture growth, the yield of the RPE-like pigmented cells was signifi cantly improved and 33% of the cells were pigmented after 6 weeks of differentiation. The derivation of RPE from hESCs and iPSCs without any external factor supplementation was also demonstrated by other groups (Vugler et al. 2008 ; Meyer et al. 2009 ; Buchholz et al. 2009).T he hESC-derived RPE cells were extensively characterized, including demonstration, both at the mRNA and the protein levels, of the expres-sion of RPE-specifi c markers, such as RPE65, CRALBP, Bestrophin, Tyrosinase, PEDF, PMEL17, LRAT, isoforms of MiTF abundant in RPE, and others. The cells expressed markers of tight junctions that join the adjacent RPE cells: ZO-1, occludin and claudin-1 (Vugler et al. 2008 ) . Electron microscopic analysis revealed that the hESC-derived RPE cells showed features characteristic of RPE. The cells were highly polarized with the nuclei located more basally, and the cytoplasm with the mitochondria and melanin granules of different maturity more api-cally. A formation of basal membrane was observed on the basal surface of the RPE cell. Similar to putative RPE, the hESC-derived RPE basal membrane was shown to be composed of extracellular matrix proteins, collagen IV , lami-nin and fi bronectin (Vugler et al.2008).The appearance of apical microvilli was demonstrated at the apical surface of the RPE. The presence of tight and gap junctions on the apical borders of the RPE cells was also confi rmed by electron microscopy. O ne of the most important functions of RPE cells i n vivo is phagocytosis of shed photoreceptor outer segments, as part of the continuous renewal process of rods and cones. The hESC-derived RPE cells demonstrated the ability to phagocyto-size latex beads or purifi ed photoreceptor outer segments, confi rming that these cells are func-tionali n vitro . It may be concluded from all these studies that human pluripotent stem cells have a potential to give rise to pigmented cells exhibiting the morphology, marker expression and functionof authentic RPE.D ifferentiation into Retinal Progenitors and Photoreceptors O ur group showed, for the fi rst time, the potential of highly enriched cultures of hESC-derived neu-ral precursors (NPs) to differentiate towards the neural retina fate (Banin et al. 2006).We demon-strated that the NPs expressed transcripts of key regulatory genes of anterior brain and retinal development. After spontaneous differentiation i n vitro , the NPs gave rise to progeny expressing markers of retinal progenitors and photoreceptor development, though this was uncommon and cells expressing markers of mature photorecep-tors were not observed. We showed that after transplantation into rat eyes, differentiation into cells expressing specifi c markers of mature photoreceptors occurred only after subretinal transplantation (between the host RPE and pho-toreceptor layer) suggesting that this specifi c microenvironment provided signals, yet unde-fi ned, that were required to support differentia-tion into the photoreceptoral lineage.P rogress towards controlling and inducing the differentiation of hESCs into retinal progenitors and neurons i n vitro was reported in the study of Lamba et al. ( 2006).They treated hESC-derived EBs for 3 days with a combination of factors,including Noggin, an inhibitor of BMP signaling, Dkk1, a secreted antagonist of the Wnt signaling pathway and insulin-like growth factor 1 (IGF-1), which is known to promote retinal progenitor dif-ferentiation. The cultivation of EBs with these factors was followed by differentiation on Matrigel or laminin for an additional 3 weeks in the presence of the combination of the three93 9 Differentiation of Human Pluripotent Stem Cells into Retinal Cellsfactors together with bFGF. Under these culture conditions, the majority of the cells developed the characteristics of retinal progenitors and expressed the specifi c markers Pax6 and Chx10 (82% and 86% of the cells, respectively). The authors showed that after further differentiation, the cells expressed markers of photoreceptor development Crx and Nrl (12% and 5.75%, respectively). About 12% of the cells expressed also HuC/D, the marker of amacrine and ganglion cells. The expression of markers of the other sub-types of retinal neurons was demonstrated, as well. However, only very few cells (<0.01%) expressed markers of mature photoreceptors, blue opsin and rhodopsin. The abundance of cells expressing markers of photoreceptors could be accelerated by co-culture with retinal explants, especially when the explants originated from mice bearing a mutation that causes retinal degeneration.T o better characterize the phenotype of retinal cells obtained with this differentiation protocol, a microarray-based analysis comparing human retina to the hESC-derived retinal cells was per-formed (Lamba and Reh 2011).It was demon-strated that gene expression in hESC-derived retinal cells was highly correlated to that in the human fetal retina. In addition, 1% of the genes that were highly expressed in the hESC-derived cultures could be attributed to RPE and ciliary epithelium differentiation.A n alternative protocol for the derivation of retinal progenitors and photoreceptors was pro-posed by Osakada et al. (2008).Similar to the protocol for the derivation of RPE cells, they used serum-free fl oating cultures in combination with the Dkk1 and LeftyA. After 20 days of cul-ture in suspension, the cells were replated on poly-D-lysine/laminin/fi bronectin-coated slides. Osakada and co-authors demonstrated that on day 35 in culture, about 16% of colonies were positive for retinal progenitor markers Rx and Pax6. Differentiation towards photoreceptor fate was augmented in the presence of N2 by treat-ment with retinoic acid and taurine, which are known inducers of rod fate differentiation. Under these conditions, after an extended culture period of 170 days, about 20% of total cells were positive for Crx, an early photoreceptor marker. On day 200, about 8.5% of the cells expressed the mature rod photoreceptor marker, rhodopsin, as well as cone photoreceptor markers, red/green and blue opsins (8.9% and 9.4%, respectively).A n alternative approach was proposed by the same group based on the use of small molecules. In this method, the chemical inhibitors CKI-7 and SB-431542 that inhibit Wnt and Activin A signaling, respectively, and Y-27632, the Rho-associated kinase inhibitor, which prevents disso-ciation-induced cell death, were used. These molecules were shown to mimic the effects of Dkk1 and LeftyA (Osakada et al. 2009).This strategy, which doesn’t involve the use of recom-binant proteins which are produced in animal or E scherichia coli cells, is more favorable for the gen-eration of cells for future transplantation therapy.I n another study that was published by Meyer et al .(2009), after initial differentiation in sus-pension for 6 days, the aggregates were allowed to attach to laminin–coated culture dishes. After further differentiation as adherent cultures, neu-roepithelial rosettes were formed, which were mechanically isolated and subsequently culti-vated as neurospheres. The authors didn’t use any soluble factors; moreover, they showed that under these conditions, the cells expressed endogenous Dkk1 and Noggin. They also demonstrated that in concordance with the role of bFGF in retinal specifi cation, the inhibition of endogenous FGF-signaling abolished retinal differentiation. Under their differentiation protocol, by day 16, more than 95% of the cells expressed the retinal pro-genitor markers, Pax6 and Rx. The authors dem-onstrated that by day 80 of differentiation, about 19% of all neurospheres contained Crx+ cells and within these Crx+ neurospheres, 63% of all cells express Crx and 46.4% of the cells expressed mature markers, such as recoverin and cone opsin.I n all of the above studies, differentiated cells expressing the retinal markers were obtained; however, the cells were not organized in a three-dimensional retinal structure. In a paper recently published by Eiraku et al. (2011),the authors cul-tured free-fl oating aggregates of mouse ES cells in serum-free medium in the presence of base-ment membrane matrix, Matrigel, that could also94M. Idelson and B. Reubinoffbe substituted with a combination of laminin, entactine and Nodal. Using a mouse reporter ES cell line, in which green fl uorescent protein (GFP) is knocked in at the Rx locus, the authors showed that Rx-GFP+ epithelial vesicles were evaginated from the aggregates after 7 days of differentiation under these conditions. On days 8–10, the Rx-GFP+ vesicles changed their shape and formed optic cup-like structures. The inner layer of these structures expressed markers of the neural retina whereas the outer layer expressed markers of RPE. The authors demonstrated that differen-tiation into RPE required the presence of the adjacent neuroectodermal epithelium as a source of diffusible inducing factors. In contrast, the differentiation into neural retina did not require tissue interactions, possibly because of the intrinsic inhibition of the Wnt-signaling pathway. Eiraku and colleagues showed that the retinal architecture, which was formed within the optic vesicle-like structures, was comparable to the native developing neural retina.R ecently, optic vesicle-like structures were also derived from hESCs and iPSCs using the protocol described above, which is based on iso-lating the neural rosette-containing colonies and culturing them in suspension (Meyer et al. 2011). The cells within the structures expressed the markers of retinal progenitors, and after differen-tiation gave rise to different retinal cell types. It was shown that the ability of optic vesicle-like structures to adopt RPE fate could be modulated by Activin A supplementation. The production of these three-dimensional retinal structures opens new avenues for studying retinal development in normal and pathological conditions.T ransplantation of Pluripotent Stem Cell-Derived Retinal CellsA key step towards future clinical transplanta-tions of hESC-derived RPE and neural retina is to show proof of their therapeutic potential i n vivo. Various animal models of retinal degeneration have been used to evaluate the therapeutic effect of transplanted retinal cells. Human ESC-derived RPE cells were transplanted subretinally to the degenerated eyes of RCS rats. Transplantation of the hESC-derived RPE cells between the RPE and the photoreceptor layer rescued retinal struc-ture and function (Lund et al. 2006; Vugler et al. 2008; Idelson et al. 2009; Lu et al. 2009).The subretinally engrafted hESC-derived RPE cells salvaged photoreceptors in proximity to the grafts as was shown by the measurement of the thick-ness of the ONL, the layer of photoreceptor nuclei, which is an important monitor of photore-ceptor cell survival. The ONL thickness was significantly increased in transplanted eyes in comparison to the degenerated non-treated eyes.I n order to evaluate the functional effect of transplanted cells i n vivo, the electroretinography (ERG) that directly measures the electrical activ-ity of the outer (a-wave) and inner (b-wave) retina in response to light stimulation was used. It was demonstrated that after transplantation of hESC-derived RPE, ERG recordings revealed a signifi -cant preservation of retinal function in the treated eyes as compared to control untreated eyes (Lund et al. 2006; Idelson et al. 2009).The visual func-tion of the animals was also estimated by an optomotor test, which monitors the animal’s refl exive head movements in response to a rotat-ing drum with fi xed stripes. Animals transplanted with hESC-derived RPE showed signifi cantly better visual performance in comparison to con-trol animals (Lund et al. 2006; Lu et al. 2009). The presence of rhodopsin, a major component of photoreceptor outer segments, within the sub-retinaly transplanted pigmented cells suggested that they could perform phagocytosis i n vivo (Vugler et al. 2008; Idelson et al. 2009).B ridging the gap between basic research and initial clinical trials requires immense resources to ensure safety and efficacy. Human ESC-derived RPE cell lines were generated using a current Good Manufacturing Practices (cGMP)-compliant cellular manufacturing process (Lu et al. 2009). Long-term studies analyzing safety and efficacy of transplantation of these GMP-compliant hESC-derived RPE cells revealed that the subretinally transplanted cells survived for a period of up to 220 days and provided prolonged functional improvement for up to 70 days after transplantation. The potential of the hESC-derived。
基于SE_注意力CycleGAN_的蓝印花布单纹样自动生成

理论与方法丝绸JOURNALOFSILK基于SE注意力CycleGAN的蓝印花布单纹样自动生成SinglepatternautomaticgenerationofbluecalicobasedonSEattentionCycleGAN冉二飞1ꎬ2ꎬ贾小军1ꎬ2ꎬ喻擎苍1ꎬ谢㊀昊1ꎬ2ꎬ陈卫彪2ꎬ3(1.浙江理工大学计算机科学与技术学院(人工智能学院)ꎬ杭州310018ꎻ2.嘉兴学院信息科学与工程学院ꎬ浙江嘉兴314001ꎻ3.浙江师范大学数学与计算机科学学院ꎬ浙江金华321004)摘要:根据蓝印花布纹样的风格特征ꎬ文章提出一种端到端的蓝印花布纹样自动生成方法ꎬ实现简笔画图像向蓝印花布单纹样的自动迁移ꎮ针对蓝印花布的抽象风格和小数据集问题ꎬ重新构造CycleGAN生成网络中的编码器和解码器ꎬ使用SE(squeezeandexcitation)注意力模块和残差模块与原始的卷积模块串联ꎬ提高特征提取能力和网络学习能力ꎮ同时减少生成网络中转换器的残差块层数ꎬ降低过拟合ꎮ实验结果表明ꎬ基于SE注意力CycleGAN网络方法自动生成的蓝印花布新纹样主观性上更贴合原始风格ꎬ与原图更加接近ꎬ有助于蓝印花布的数字化传承和创新ꎮ关键词:蓝印花布ꎻSE注意力ꎻ风格迁移ꎻCycleGANꎻ单纹样ꎻ半监督学习ꎻ图像生成中图分类号:TS941.2ꎻTP391.7㊀㊀㊀㊀文献标志码:A㊀㊀㊀㊀文章编号:10017003(2024)01003107DOI:10.3969/j.issn.1001 ̄7003.2024.01.004收稿日期:20230525ꎻ修回日期:20231201作者简介:冉二飞(1998)ꎬ男ꎬ硕士研究生ꎬ研究方向为计算机视觉与图像处理ꎮ通信作者:贾小军ꎬ教授ꎬxjjiad@sina.comꎮ㊀㊀蓝印花布源于唐宋ꎬ盛于明清ꎬ是中国传统的民间手工艺品ꎮ作为首批列入国家级非物质文化遗产名录的民间传统工艺ꎬ其使用简单淳朴的蓝白两色ꎬ创造出绚丽多姿的艺术世界ꎬ因其纹样设计风格特征鲜明㊁ 线断意连 而闻名于世ꎮ目前对蓝印花布的研究ꎬ大都是从纹样的视觉语义寓意ꎬ历史发展和制作工艺角度进行[1 ̄3]ꎬ而对于纹样的自动生成技术研究略显单薄ꎬ且往往不够简单实用[4 ̄5]ꎮ随着深度学习的发展ꎬ尤其是图像风格迁移研究的进展ꎬ提供了扩展蓝印花布纹样的新思路ꎬ即将其他风格的简单图像转化为蓝印花布风格的纹样ꎬ从而大大提高蓝印花布设计的效率ꎬ有利于创新蓝印花布纹样ꎮ图像的风格迁移是两个不同域中图像的转换ꎬ具体来说就是提供一种风格图像ꎬ将任意一张图像转化为这种风格ꎬ并尽可能保留原图像的内容ꎮ如今实现风格迁移的方法多种多样ꎬ大致分为基于神经网络的图像风格迁移和基于对抗生成网络的图像风格迁移ꎮGatys等[6]首先将深度学习运用在风格迁移任务上ꎬ利用Gram矩阵将图像表示为内容和风格两部分ꎬ通过图像重建使内容图的Gram矩阵逼近风格图的Gram矩阵ꎮ这种方法可以生成风格图像ꎬ但是收敛速度慢ꎬ渲染时间长ꎮWang等[7]为了减少伪影ꎬ引入了相似性损失函数ꎬ添加了一个后处理细化步骤ꎮ其方法可以稳定地对摄影作品的图像进行风格转换ꎮLuan等[8]约束卷积神经网络从输入到输出的变换ꎬ使其表示为颜色空间中的局部仿射ꎬ实现了如天气㊁季节等多种场景的艺术风格转换ꎬ但是风格迁移的效率低下ꎬ效果一般ꎮJohnson等[9]使用在ImageNet上预训练的VGGNet简化损失函数计算过程ꎬ使效率得到较大提升ꎬ但该方法需要构造复杂的损失函数ꎮGoodfellow等[10]提出了生成对抗网络理论(GAN)ꎬGAN为图像风格转换提供了新的思路ꎬ掀起了新的研究热潮ꎮMirza等[11]提出了带有条件约束的cGAN(conditionalGAN)ꎬ该模型通过对输入图像额外增加一个条件标签ꎬ来引导模型生成方向ꎮIsola等[12]提出了Pix2Pix算法模型ꎬ通过图像作为输入来进行图像风格转换ꎬ而不是传统的噪音ꎬ大大提升了生成图像的可控性ꎮcGAN和Pix2Pix都需要配对的数据集ꎬ但是在很多情况下ꎬ并没有完美的成对数据集ꎮZhu等[13]提出了CycleGAN算法模型ꎬ摆脱了配对训练数据集的限制要求ꎬ使用半监督的方式实现不同风格之间的图像转换ꎬ如将马转化为斑马㊁春夏秋冬的转换㊁油画和真实图像的转换等ꎬ但CycleGAN在几何形状改变方面表现不佳ꎮChen等[14]提出CartoonGANꎬ用于将现实图像转化为漫画风格ꎮ该模型在CycleGAN的基础上针对卡通图像加入边缘对抗损失ꎬ使生成的图像具有漫画图像一样的清晰边缘ꎬ但在处理现实人脸到漫画人脸这类形变较大的转换时效果不佳ꎮ以上模型大多只对于图像纹理与色调的风格进行转换ꎬ忽略了几何形变方面的风格转换ꎮ蓝印花布的风格有很强的抽象特性ꎬ为了稳定生成蓝印花布的纹样ꎬ针对上述图像风格化存在问题ꎬ本文使用SE注意力机制改进CycleGAN模型来13Vol.61㊀No.1SinglepatternautomaticgenerationofbluecalicobasedonSEattentionCycleGAN实现蓝印花布纹样图像的风格迁移ꎮ对生成网络中的编码器和解码器进行重构ꎬ提升CycleGAN模型几何形变方面的能力ꎬ使得生成结果贴近蓝印花布纹样的抽象风格ꎮ1㊀CycleGAN原理GAN模型由生成网络和判别网络组成ꎬ其目标是让生成网络的模型学会一种映射ꎬ使得原始域的数据分布拟合目标域的数据分布ꎮ在训练过程中ꎬ生成网络需要生成伪造的样本使判别网络判断为真ꎬ而判别网络则要尽力判别输入样本是真实样本还是生成的伪造样本ꎬ两个神经网络在这个对抗中不断优化ꎬ最后网络模型能够输出接近样本分布的数据ꎮGAN的优化目标可以看成是一个极大极小博弈ꎬ在训练最后的生成网络和判别网络之间实现纳什均衡ꎬ使得生成网络能够生成接近样本数据分布特征的目标数据ꎮ其目标方程[10]为:minG㊀maxDV(DꎬG)=Ex~pdata[log(D(x))]+Ez~pz[log(1-D(G(z)))](1)式中:G表示生成网络ꎬD表示判别网络ꎬPdata表示真实样本x的分布ꎬPz表示输入的噪声z的分布ꎬV表示损失函数ꎬE表示数学期望ꎬG(z)表示生成网络G根据噪声z生成的假图像ꎬD(x)和D(G(z))分别表示判别器D判断真实样本x和假图像G(z)是真实样本分布的概率ꎮGAN模型在跨域图像风格转换任务上存在一个缺陷ꎬ生成网络可能会把原始域映射到目标域上的子集ꎬ甚至有可能将原始域全部映射到一张图像上ꎬ而判别网络只关注生成图像是否属于目标域ꎬ所以仅通过单独的对抗损失ꎬ无法达到将原始域映射到目标域的结果ꎮ对于这个问题ꎬCycleGAN没有采用像Pix2Pix算法使用严格配对数据集的做法ꎬ而是使用循环一致性损失解决这一问题ꎮCycleGAN模型通过第一个生成网络G(x2y)ꎬ将输入的X域图像转换成Y域ꎬ然后通过第二个生成网络F(y2x)转换回来ꎬ将原始域中的数据经过两次转换后ꎬ转换回来的图像应与原始输入尽量相同ꎮ同样地ꎬ对于Y域的图像通过F(y2x)和G(x2y)重新生成伪造的Y域图像与原始输入的Y域图像进行比较ꎮ通过这种方式解决了X域可能都映射到Y域同一张图像的情况ꎮCycleGAN模型有两个生成网络和两个判别网络ꎬ其中两个生成对抗网络中的生成网络共享权重ꎮCycleGAN模型在结构上像一个环形网络ꎬ从X域向Y域转换的GAN网络结构如图1所示ꎮCycleGAN循环一致性损失函数计算方法[13]为:Lcyc(GꎬF)=Ex~pdata(x)F(G(x)-x)1+Ey~pdata(y)F(G(y)-y)1(2)对抗损失由两部分组成ꎬ分别对应两个单独的GAN模型:LGAN(GꎬDyꎬXꎬY)=Ey~pdata(y)[logDy(y)]+Ex~pdata(x)[log(1-Dy(G(x)))](3)图1㊀CycleGAN网络结构Fig.1㊀CycleGANnetworkstructure㊀㊀LGAN(FꎬDxꎬYꎬX)=Ex~pdata(x)[logDx(x)]+Ey~pdata(y)[log(1-Dx(F(y)))](4)整个环形GAN网络的总损失[13]计算方法为:L(GꎬFꎬDxꎬDy)=LGAN(GꎬDyꎬXꎬY)+LGAN(FꎬDxꎬYꎬX)+λLcyc(GꎬF)(5)式中:X㊁Y分别代表两个数据域ꎬx㊁y为两个数据域中的样本数据ꎬG为从X到Y的映射函数ꎬF为从Y到X的映射函数ꎬDx㊁Dy为判别网络ꎬλ为控制循环一致损失函数的权重ꎮ2㊀改进的CycleGAN网络结构CycleGAN网络实现了无配对图像集之间的风格迁移ꎬ但泛化能力较弱ꎬ当训练图像与测试图像之间差距较大时ꎬ迁移效果不佳ꎮ直接使用CycleGAN进行简笔画向蓝印花布风格纹样转化生成的结果不理想ꎮ针对这一问题对原始CycleGAN的生成网络结构进行改进ꎬCycleGAN的生成网络结构由编码器㊁残差结构和解码器组成ꎮ编码器与解码器都是由3个卷积模块组成ꎬ每个模块包含一个卷积层㊁一个实例正则化层及一个Relu激活函数ꎮ为了增强CycleGAN网络提取特征的能力ꎬ本文提出了使用残差块结构加上原始卷积模块和SE注意力模块取代原卷积模块的方法ꎮ2.1㊀注意力机制注意力机制(attentionmechanism)是解决信息超载问题的一种资源分配方案ꎬ当计算资源有限时ꎬ可以把计算资源分配给更重要的任务ꎮ在神经网络学习过程中ꎬ参数越多模型所存储的信息量就越大ꎬ模型的表达能力也越强ꎬ但这会带来信息过载的问题ꎮ引入注意力机制ꎬ可以在众多的输入信息中聚焦于对当前任务更为关键的信息ꎬ降低对其他信息的关注度ꎬ提高任务处理的效率和准确性ꎮSE(squeezeandexcitation)模块[15]在通道维度增加注意力机制ꎬ通过一系列变换操作得到一个权重矩阵ꎬ对原特征进行重构来得到更重要的特征信息ꎬ关键步骤为压缩(squeeze)和激发(excitation)ꎮ通过自动学习的方式ꎬ获取特征通道的重要程度ꎬ以此为每个特征通道赋予不同的权重值ꎬ从而提升对当前任务有用的特征图的通道利用率ꎬ并抑制对当前任务影响不大的通道ꎮSE模块的结构[15]如图2所示ꎮ23第61卷㊀第1期基于SE注意力CycleGAN的蓝印花布单纹样自动生成图2㊀特征压缩与激发模块结构Fig.2㊀Featurecompressionandexcitationmodulestructure㊀㊀实现通道注意力的过程分为压缩(squeeze)ꎬ激发(excitation)和调节(scale)三个步骤ꎮ压缩阶段ꎬ使用全局池化(globalpooling)操作ꎬ将HˑWˑC的特征图沿着通道方向转化成为1ˑ1ˑC的特征ꎬ即图2中的Fsq过程将每一个二维的特征图变成了一个具有整体卷积感受野的实数Zcꎬ其计算方法为:Zc=Fsq(uc)=1HˑWðHi=1ðWj=1uc(iꎬj)(6)式中:H㊁W为输入特征图的高和宽ꎬuc(iꎬj)为在i㊁j位置上的特征值ꎮ激发过程的操作Fex( ꎬW)是通过两个全连接层来捕捉通道的内部依赖程度参数得到特征通道的裁剪系数ꎬ其计算方法为:s=Fex(zꎬW)=σ(W2δ(W1z))(7)式中:W1和W2为两个全连接层学习参数ꎬδ为Relu函数ꎬσ为Sigmoid函数ꎬz为上一步求得的1ˑ1ˑC的特征向量ꎮ最后是调节权重ꎬ将s视为每个通道的重要程度ꎬ通过Fscale(.ꎬ.)逐通道加权到先前的特征图上ꎬ完成通道维度上的注意力机制计算ꎮFscale(scꎬuc)=scuc(8)式中:sc为第C个通道的裁剪系数ꎬuc为第C个通道的特征图ꎮ2.2㊀网络结构CylceGAN算法模型是一个循环迭代的网络ꎬ包括两个生成网络G(x2y)㊁F(y2x)及两个判别网络Dx㊁Dyꎮ在本文研究中ꎬX代表简笔画数据域ꎬY代表蓝印花布纹样数据域ꎮ两个生成网络分别生成简笔画图像和蓝印花布图像ꎮ生成网络通过学习两个图像域之间的映射函数ꎬ最终生成网络G(x2y)能够使一个简笔画图像转化后获得蓝印花布纹样的风格特点ꎮ2.2.1㊀生成网络生成网络主要由编码器㊁转换器和解码器组成ꎮ本文采用残差块结构㊁原始卷积模块和SE注意力模块对原CycleGAN模型中编码器和解码器进行改进ꎬ并削减转化器中残差块的数量ꎮ对于新卷积模块(Se ̄ConvLayer)ꎬ使用残差模块(ResidualBlock)及原始卷积模块(Conv3ˑ3stride=2padding=1ꎻInstanceNormꎻRelu)加SE注意力模块(SeBlock)取代原始卷积模块ꎬ如图3(a)所示ꎮ对于新反卷积模块(SeDeConvLayer)ꎬ使用SE注意力模块(SeBlock)及原始反卷积模块(Upsamplescale_factor=2ꎻConv3ˑ3stride=1padding=1ꎻInstanceNormꎻRelu)加残差模块(ResidualBlock)取代原始反卷积模块ꎬ如图3(b)所示ꎮ图3㊀改进的新模块Fig.3㊀Newimprovedmodules改进的整个生成网络结构如图4所示ꎮ首先将输入的256ˑ256灰度图像送入一个padding为1ꎬ卷积核大小为3ˑ3的卷积IniConvLayer中进行初始化ꎬ将通道数扩展为16ꎮ而后将输出送入到SeConvLayer1中进行特征提取ꎬ并在SeBlock中根据式(6~8)进行特征图权重的分配ꎬ得到32个128ˑ128的特征向量ꎮ对于得到的特征向量ꎬSeConvLayer2再次进行特征提取和特征图权重的分配ꎬ最终编码器提取出64个尺寸为64ˑ64的特征向量ꎮ由4个残差模块组成的转换器组合提取不同的特征ꎬ将输入转化为目标域的特征向量ꎮ每个残差块中包含2个卷积层㊁2个实例正则化层和1个Relu激活层ꎮ解码器为编码器的逆过程ꎬ采用与新卷积模块类似的2个新反卷积层SeDeConvLayer1和SeDeConvLayer2还原图像的低级特征ꎬ得到一个16ˑ256ˑ256的特征向量ꎬ最后通过一个1ˑ1卷积层FinConvLayer还原成目标域的图像ꎬ之后将还原的图像输入判别网络判断ꎬ根据式(5)计算梯度ꎬ更新生成网络参数ꎮ33Vol.61㊀No.1SinglepatternautomaticgenerationofbluecalicobasedonSEattentionCycleGAN图4㊀改进的CycleGAN的生成网络结构Fig.4㊀ImprovedCycleGANgenerativenetworkstructure2.2.2㊀判别网络判别网络的作用为判别生成网络产生的伪样本是否属于目标数据域ꎬ促使生成网络生成更加难以分辨真伪的样本ꎬ其网络结构如图5所示ꎮ本实验的判别网络使用PatchGan[10]判别网络ꎬ由5个卷积模块组成ꎬ前四个模块用于抽取输入图像的特征ꎬ最后一个卷积模块得到一张特征图ꎬ此时特征图上每一个点都有着70ˑ70的感受野ꎬ代表了判别网络对于其代表的70ˑ70图像块的判别结果ꎮ最后将所有图像块进行判别之后得到的平均结果作为这张输入图像的判别结果ꎬ随后根据式(5)计算梯度ꎬ更新判别网络参数ꎮ图5㊀CycleGAN判别网络结构Fig.5㊀CycleGANdiscriminatornetworkstructure3㊀实验和结果分析3.1㊀环境与数据集实验平台处理器为Intel(R)Xeon(R)Platinum8350CCPU@2.60GHzꎬ显卡型号为NAVIDRTXA5000ꎬ24GB显存ꎬ操作系统为Ubuntu18.04ꎬ使用PyTorch深度学习框架ꎬ版本为1.8.1ꎬCuda11.1ꎮ基于改进CycleGAN的风格迁移算法分别选用412张动物简笔画和216张蓝印花布单纹样图作为实验样本数据集ꎮ训练过程中不同数据集间没有指定配对关系ꎬ其中简笔画数据集通过互联网收集ꎬ蓝印花布单纹样从已有的蓝印花布图像中截取ꎬ两个数据集图像大小统一调整为256ˑ256像素ꎮ部分数据集图像如图6所示ꎮ3.2㊀训练过程设置循环一致性损失超参数λ值为10ꎬ批量训练样本数量(batchsize)为1ꎬ训练总轮次(epoch)为400ꎬ采用adam优化器ꎬ初始学习率(lr)为0.0002ꎬ在100个epoch后使用指数图6㊀训练时的部分数据集Fig.6㊀Partialdatasetduringtraining衰减策略对学习率进行动态调整ꎮ训练过程中的部分实验数据如图7所示ꎮ模型训练50个epoch时转换效果如图7第二行所示ꎬ此时图像尚没有形成明显的蓝印花布风格ꎬ原始形状保留较为完整ꎬ但此时生成的图像颜色已经变为黑底白纹ꎬ并且组成图像内容的线条已经转化为大小不同的白色块ꎬ有了向蓝印花布风格转化的趋势ꎮ模型训练100个epoch时转换效果如图7第三行所示ꎬ此时产生的图像仍显得零乱ꎬ产生的色块也较多ꎬ不符合蓝印花布抽象精炼的特点ꎬ但是相比50个epoch时产生的色块少了许多ꎬ初步形成了蓝印花布的风格ꎮ模型训练400个epoch时转换效果如图7第四行所示ꎬ此时生成的图像抓住了原始图像中的关键信息ꎬ并将其融合进了蓝印花布的风格里ꎬ图案边界更加清晰ꎬ主体模糊程度明显减弱ꎬ整体风格更为凝实㊁简练ꎬ并在不妨碍整体布局的情况下ꎬ对于过长的线条或弯曲弧度较大的造型产生了合适的断线ꎬ比较符合蓝印花布 线断意连 的特点ꎮ43第61卷㊀第1期基于SE注意力CycleGAN的蓝印花布单纹样自动生成图7㊀训练过程的转换效果Fig.7㊀Conversioneffectoftrainingprocess㊀㊀训练过程损失如图8所示ꎬ在GAN模型训练过程中由于生成网络和判别网络的对抗ꎬ在训练一段时间达到稳定期后ꎬ生成网络和判别网络的损失都应该在一个小区间内波动ꎬ而不会有明显的持续上升或者下降趋势ꎮ在图8中ꎬ前200个epoch的学习率相对较大ꎬ总损失total_loss曲线震荡明显ꎬ200个epoch后随着学习率的降低和训练达到稳定期ꎬtotal_loss在较小的区间波动ꎮ在400个epoch时产生的图像已经有了蓝印花布纹样的风格并且网络已稳定ꎮ图8㊀训练过程total_loss变化Fig.8㊀Total_losschangesduringtraining3.3㊀网络对比为了验证所提出网络的优越性ꎬ本文将提出的方法与对于同样基于对偶思想的CycleGAN㊁DualGAN[16]㊁DiscoGAN[17]进行对比ꎬ结果如图9所示ꎮ在图9中ꎬ从上到下依次为原图㊁CycleGAN算法㊁DualGAN算法㊁DiscoGAN算法与SE ̄CycleGAN(ours)算法生成的结果ꎮ对于DualGAN和DiscoGAN来说ꎬ其生成网络采用Unet结构ꎬ保留了大量原始的细节ꎬ在灰度图中仍能隐约看到原始的图像ꎬ但是在高度抽象化的蓝印花布风格中ꎬ这些细节反而使得生成效果大大降低ꎬ并且图像中没有产生合适断线ꎬ整体结构也更加混乱ꎮCycleGAN虽然表现出断线ꎬ但是断线后形成的色块过于紧凑ꎬ没有呈现蓝印花布的风格ꎮSE ̄CylceGAN(ours)明显提升了图像的抽象化程度ꎬ相对于其他网络色块少而精炼ꎬ主体更加突出ꎬ线条分布合理ꎬ 线断意连 风格明显ꎬ表现最优ꎮ图9㊀不同网络生成结果Fig.9㊀Generatedresultsofdifferentnetworks3.4㊀注意力机制对比为了直观展示使用不同注意力机制的风格迁移效果ꎬ本文将SE注意力机制替换为CBAM注意力机制[18]和ECA注意力机制[19]并进行对比ꎬ结果如图10所示ꎮ在图10中ꎬ从上到下依次为原图㊁CycleGAN算法㊁CBAM ̄CycleGAN算法㊁ECA ̄CycleGAN算法与SE ̄CycleGAN(ours)算法生成的结果ꎮ其中CBAM ̄CycleGAN对于原图像的细节保留过多ꎬ断线产生的位置不合理ꎬ转化后形成的风格比起SE ̄CycleGAN(ours)有较大的不足ꎬ在小猫图像和狐狸图像上较为明显ꎬ且CBAM ̄CycleGAN明显产生了更多的噪点ꎮ而ECA ̄CycleGAN对于原图内容的把握明显缺失ꎬ色块排列杂乱无章ꎬ没有明确的主题或焦点ꎬ难以捕捉到图片想要表达的核心信息ꎬ甚至生成了如第二列蜜蜂图所示的意义不明的图像ꎮSE ̄CycleGAN(ours)生成的效果最好ꎬ色块更加写意ꎬ色块边缘更加柔和ꎬ如牛的耳朵细看之下宛如花瓣ꎬ并且断线的位置恰到好处ꎬ符合蓝印花布 线断意连 的风格ꎬ在整体上结构清晰ꎬ有更高的艺术价值ꎮ实验结果表明ꎬ相较于其他算法ꎬ使用SE注意力机制的CycleGAN算法在生成蓝印花布图样上有明显的优势ꎮ图10㊀使用不同注意力的改进CycleGAN网络生成结果Fig.10㊀ImprovedCycleGANnetworkgeneratedresultsusingdifferentattentionmethods53Vol.61㊀No.1SinglepatternautomaticgenerationofbluecalicobasedonSEattentionCycleGAN4㊀结㊀论作为国家非物质文化遗产的蓝印花布ꎬ对其进行数字化传承和创新有重要的价值和意义ꎮ本文提出了一种基于SE注意力机制的改进CycleGAN的端到端网络SE ̄CycleGANꎮ通过SE注意力模块和残差模块重新塑造的生成网络提高了原始CycleGAN的几何变形能力ꎬ相对于其他网络ꎬ图像的内容与蓝印花布的风格有更好的融合ꎬ产生的噪点也较少ꎬ并且新提出的网络本身有小样本训练ꎬ无需配对数据集的特点ꎬ可以很好地迁移到其他类似纹样的生成ꎮ但是因为数据集和分辨率的限制ꎬ目前生成的纹样仍较为简单ꎮ而且ꎬ受到笔画和图像信息密集程度的影响较大ꎬ难以生成更加复杂㊁内容丰富多样的纹样ꎮ接下来将在多类㊁复杂的蓝印花布纹样自动生成方面开展研究工作ꎮ«丝绸»官网下载㊀中国知网下载参考文献:[1]侯莉莉ꎬ须秋洁.蓝印花布纹样设计之点画技法探析[J].美术教育研究ꎬ2022(15):50 ̄52.HOULLꎬXUQJ.Analysisofstipplingtechniquesinbluecalicopatterndesign[J].ArtEducationResearchꎬ2022(15):50 ̄52.[2]金晓伟.蓝印花布图案的视觉语言研究[J].美术观察ꎬ2022(8):73 ̄74.JINXW.Studyonthevisuallanguageofbluecalico[J].ArtObservationꎬ2022(8):73 ̄74.[3]杜威.蓝印花布艺术传承研究[J].西部皮革ꎬ2022ꎬ44(21):54 ̄56.DUW.Studyontheinheritanceofbluecalicofabricact[J].WestLeatherꎬ2022ꎬ44(21):54 ̄56.[4]贾小军ꎬ叶利华ꎬ邓洪涛ꎬ等.基于卷积神经网络的蓝印花布纹样基元分类[J].纺织学报ꎬ2020ꎬ41(1):110 ̄117.JIAXJꎬYELHꎬDENGHTꎬetal.Elementsclassificationofveinpatternsusingconvolutionalneuralnetworksforbluecalico[J].JournalofTextileResearchꎬ2020ꎬ41(1):110 ̄117.[5]贾小军ꎬ邓洪涛ꎬ滕姿ꎬ等.应用轮廓线拟合提取蓝印花布图案基元[J].纺织学报ꎬ2018ꎬ39(8):150 ̄157.JIAXJꎬDENGHTꎬTENGZꎬetal.Extractionofimageelementsforbluecalicobasedoncontourfitting[J].JournalofTextileResearchꎬ2018ꎬ39(8):150 ̄157.[6]GATYSLꎬECKERAꎬBETHGEM.Imagestyletransferusingconvolutionalneuralnetworks[C]//ProceedingsoftheIEEEConferenceonComputerVisionandPatternRecognition(CVPR).LasVegas:IEEEꎬ2016.[7]WANGLꎬWANGZꎬYANGXꎬetal.Photographicstyletransfer[J].TheVisualComputerꎬ2020ꎬ36(2):317 ̄331.[8]LUANFꎬPARISSꎬSHECHTMANEꎬetal.Deepphotostyletransfer[C]//ProceedingsoftheIEEEConferenceonComputerVisionandPatternRecognition(CVPR).HawaiiState:IEEEꎬ2017.[9]JOHNSONJꎬALAHIAꎬLIFF.Perceptuallossesforreal ̄timestyletransferandsuper ̄resolution[C]//Europeanconferenceoncomputervision(ECCV).Cham:Springerꎬ2016:694 ̄711.[10]GOODFELLOWIꎬPOUGET ̄ABADIEJꎬMIRZAMꎬetal.Generativeadversarialnetworks[J].CommunicationsoftheACMꎬ2020ꎬ63(11):139 ̄144.[11]MIRZAMꎬOSINDEROS.Conditionalgenerativeadversarialnets[J].ComputerScienceꎬ2014:2672 ̄2680.[12]ISOLAPꎬZHUJYꎬZHOUTꎬetal.Image ̄to ̄imagetranslationwithconditionaladversarialnetworks[C]//ProceedingsoftheIEEEConferenceonComputerVisionandPatternRecognition(CVPR).HawaiiState:IEEEꎬ2017.[13]ZHUJYꎬPARKTꎬISOLAPꎬetal.Unpairedimage ̄to ̄imagetranslationusingcycle ̄consistentadversarialnetworks[C]//ProceedingsoftheIEEEInternationalConferenceonComputerVision(ICCV).HawaiiState:IEEEꎬ2017.[14]CHENYꎬLAIYKꎬLIUYJ.Cartoongan:Generativeadversarialnetworksforphotocartoonization[C]//ProceedingsoftheIEEEConferenceonComputerVisionandPatternRecognition(CVPR).SaltLakeCity:IEEEꎬ2018.[15]HUJꎬSHENLꎬSUNG.Squeeze ̄and ̄excitationnetworks[C]//ProceedingsoftheIEEEConferenceonComputerVisionandPatternRecognition(CVPR).SaltLakeCity:IEEEꎬ2018.[16]YIZꎬZHANGHꎬTANPꎬetal.Dualgan:Unsupervisedduallearningforimage ̄to ̄imagetranslation[C]//ProceedingsoftheIEEEInternationalConferenceonComputerVision(ICCV).HawaiiState:IEEEꎬ2017.[17]KIMTꎬCHAMꎬKIMHꎬetal.Learningtodiscovercross ̄domainrelationswithgenerativeadversarialnetworks[C]//InternationalConferenceonMachineLearning.Sydney:arXive ̄printsꎬ2017.[18]WOOSꎬPARKJꎬLEEJYꎬetal.Cbam:Convolutionalblockattentionmodule[C]//ProceedingsoftheEuropeanConferenceonComputerVision(ECCV).Berlin:SpringerScienceꎬ2018.[19]WANGQLꎬWUBGꎬZHUPFꎬetal.ECA ̄Net:Efficientchannelattentionfordeepconvolutionalneuralnetworks[C]//Proceedingsofthe2020IEEE/CVFConferenceonComputerVisionandPatternRecognition(CVPR).Seattle:IEEEꎬ2020.63第61卷㊀第1期基于SE注意力CycleGAN的蓝印花布单纹样自动生成SinglepatternautomaticgenerationofbluecalicobasedonSEattentionCycleGANRANErfei12JIAXiaojun12YUQingcang1XIEHao12CHENWeibiao231.SchoolofComputerScienceandTechnologySchoolofArtificialIntelligence ZhejiangSci ̄TechUniversity Hangzhou310018China 2.CollegeofInformationScienceandEngineering JiaxingUniversity Jiaxing314001China 3.CollegeofMathematicsandComputerScience ZhejiangNormalUniversity Jinhua321004ChinaAbstract BluecalicoisatraditionalcraftprintinganddyeingproductinChinawithalonghistory.Itisasimpleandprimitiveblueandwhitefabricthatreflectspeople spreferencesandembodiestheirlongingforahappylifeandsimpleaesthetictaste.Itisfamousforitsuniquepatterndesignstyleandbrokenlines.However thelackofanalgorithmfortheautomaticgenerationofbluecalicopatternshashinderedtheinnovativeresearchofpatterns.Forthisreason anend ̄to ̄endautomaticgenerationmethodofthebluecalicopatternwasproposedtorealizetheautomaticconversionfromasimplestrokeimagetoasinglebluecalicopattern.OursolutionbelongstoCycleGAN ̄basedmethods whicharepopulartostylizeimagesinartisticformssuchaspainting.CycleGANisagenerativeadversarialnetworkbasedonduallearning overcomingthethelimitationsofrequiringcorrespondingdatasetsofothermethods butitisslightlyinsufficientforgeometricshapechanges.Giventheabstractstyleandsmalldatasetofbluecalico thecoderanddecoderintheCycleGANgenerationnetworkarereconstructed andtheSEsqueezeandexception attentionmoduleandresidualmoduleareconnectedinserieswiththeoriginalconvolutionmoduletoimprovetheabilityoffeatureextractionandnetworklearning.SEattentionmechanismgivesdifferentweightstodifferentpositionsofimagesfromtheperspectiveofthechanneldomain withthenetworkfocusingonkeyinformation.Atthesametime reducingthenumberofresidualblocklayersoftheconverterinthegenerativenetworktoreduceover ̄fittingisimplemented.Besides tocarryoutthisexperiment wehavemadethedatasetofasinglepatternofbluecalicoandthedatasetofsimplestrokes.Duetothelackofpreviousresearchonthisissueandthelackofappropriatemeasurementindicatorsforsuchexperiments thestudyusedthegeneratedimageforvisualcomparison.Intheexperiment toprovethesuperiorityoftheproposedalgorithm wecomparedouralgorithmwiththeoriginalCycleGANandotheralgorithmsbasedonthedualidealikeDualGANandDiscoGAN.Theexperimentalresultsshowthatourproposedalgorithmcaneffectivelycombinethecontentofsimplestrokeswiththestyleofbluecalico whiletheoriginalCycleGANalgorithmhasinsufficientgeometricdeformationability andtheothertwoalgorithmssavetoomuchoriginalinformation whichdoesnotconformtotheconcisecharacteristicsofbluecalico.Atthesametime thestudyalsocomparedtheeffectofusingdifferentattentionmechanisms andcomparedtheECAefficientchannelattention mechanismwithCBAMconvolutionalblockattentionmodule mechanism.Amongvariousattentionmechanisms theSEattentionmechanismstillhasthebesteffect.TheimagegeneratedbyusingECAattentionlacksthecontentoftheoriginalimage theimagestyleconversiongeneratedbyusingECAattentionisnotenough.Besides comparedwiththeothertwomethods theimagegeneratedbyusingtheSEattentionmechanismhassmoothercolorblockedgesandlessnoise.Thereasonforthegapcausedbyusingdifferentattentionmechanismsmaybethatdifferentattentionmechanismsincreasethelearningabilityofgeneratorstodifferentdegrees andsomemakeitdifficultforgeneratorsanddiscriminatorstoformeffectivecompetition.Throughtheaboveexperiments thebluecalicoautomaticallygeneratedbythemethodbasedontheSEattentionCycleGANnetworkisclosertotheoriginalstyle.Throughthisalgorithm thedesignprocessofthebluecalicocanbesimplified whichisbeneficialtothedigitalinheritanceandinnovationofthebluecalico.Asanationalintangibleculturalheritage thebluecalicoisofimportantvalueandsignificance.Inthispaper anend ̄to ̄endnetworkSE ̄CycleGANbasedontheSEattentionmechanismisproposed.Comparedwithothernetworks thecontentoftheimageisbetterintegratedwiththestyleofthebluecalico.Theproposednetworkdoesnotneedmatchingdatasets soitcanbewellmigratedtothegenerationofothersimilarpatterns.Nevertheless duetothelimitationofthedatasetandresolution thepatternsgeneratedatpresentarestillrelativelysimple.Thenextresearchworkwillfocusonstudyingthemethodofgeneratingmulti ̄classandcomplexbluecalico.Keywords bluecalico SEattention styletransfer CycleGAN singlepattern semi ̄supervisedlearning imagegeneration73。
F-STOP 对照图片,病毒免疫咖啡草坛说明书

F -S T O P ™ F o r P i c t u r e -P e r f e c t , D i s e a s e -F r e e T u r f g r a s s 12770• Provides Systemic Prevention And Control Of Turfgrass Diseases• Prevents Over 15 Major Lawn Diseases • For Use On All Types Of Home Lawns • One Application Protects For Up To 4 WeeksKEEP OUT OF REACH OF CHILDRENCAUTIONBUYER ASSUMES ALL RISKS OF USE, STORAGE OR HANDLING OF THIS MATERIALNOT IN STRICT ACCORDANCE WITH DIRECTIONS GIVEN HEREWITH.NET WEIGHT 10 LBS. (4.53 KG)ACTIVE INGREDIENT:myclobutanil: a-butyl-a-(4-chlorophenyl)-1-H-1,2,4 triazole-propanenitrile: ...............0.39%OTHER INGREDIENTS: .............................................................................................99.61%TOTAL .....................................................................................................................100.00%This product contains 0.195 lb.. of myclobutanil per 50 lb. bag.C o v e r s U p T o 2,500 S q . F t .F-STOP ™ For Picture-Perfect, Disease-Free TurfgrassF -S T O P ™F o r P i c t u r e -P e r f e c t , D i s e a s e -F r e e T u r f g r a s sF-STOP ™For Picture-Perfect, Disease-Free TurfgrassPRECAUTIONARY STATEMENTSHAZARDS TO HUMANS AND DOMESTIC ANIMALSCauses Moderate Eye Irritation.Avoid contact with eyes or clothing. Wash thoroughly with soap and water after handling and before eating, drinking, chewing gum, using tobacco, or using the toilet.Notice: Read the entire label. Use only according to label directions. Before using this product, read “Warranty Disclaimer,” “Inherent Risks of Use,” and “Limitation of Remedies” at end of Directions for Use. If terms are unacceptable, return at once unopened.In case of emergency endangering health or the environment involving this product,call 1-800-992-5994Agricultural Chemical: Do not ship or store with food, feeds, drugs, or clothing.FIRST AIDIf in eyes: Hold eye open and rinse slowly and gently with water for 15-20 minutes. Remove contact lenses, if present, after the first 5 minutes, then continue rinsing eye. Call a Poison Control Center or doctor for treatment advice. Have the product container or label with you when calling a Poison Control Center or doctor, or going for treatment. You may also contact 1-800-992-5994 for emergency treatment information.ENVIRONMENTAL HAZARDSThis pesticide is toxic to fish. To protect the environment, do not allow pesticide to enter or run off into storm drains, drainage ditches, gutters or surface waters. Applying this product in calm weather when rain is not predicted for the next 24 hours will help ensure that wind or rain does not blow or wash pesticide off the treatment area. Sweeping any product that lands on a driveway, sidewalk, or street, back onto the treated area of the lawn or garden will help to prevent runoff to water bodies or drainage systems.DIRECTIONS FOR USEIt is a violation of Federal law to use this product in a manner inconsistent with its labeling. Read all directions carefully before applying this product.Not for use on turfgrass being grown for sale or other commercial use as sod, or for commercial seed productions, or for research purposes.STORAGE AND DISPOSALDo not contaminate water, food or feed by storage and disposal.PESTICIDE STORAGE: Keep away from fire and sparks. Store in a cool, dry, well-ventilated area. CONTAINER HANDLING: Nonrefillable container. Do not reuse or refill this container.If empty: Place in trash or offer for recycling if available.If partly filled: Call your local solid waste agency for disposal instructions. Never place unused product down any indoor or outdoor drain.HOW THIS PRODUCT WORKSferti•lome® F-STOP™is a systemic, protectant and curative fungicide for the control of listed diseases in established home lawns and ornamental turfgrass. Optimum disease control is achieved when this product is applied to established turfgrass in a regularly scheduled preventative program. Use this product in conjunction with turf management practices that promote good plant health and optimum disease control. The key to selecting a fungicide is the proper diagnosis of the organism causing the disease. Diagnostic kits, extension experts, or other identification methods should be used when developing disease control strategies.HOW TO APPLYApply ferti•lome® F-STOP™ uniformly over the turfgrass area using a properly calibrated drop or rotary-type spreader designed to apply granules. Before each application, calibrate the spreader according to the equipment manufacturer’s directions. Check frequently to make sure equipment is operating properly and distributing product uniformly. A more uniform application may be achieved by spreading half of the required amount of product over the area and then applying the remaining half in swaths at a right angle to the first. Avoid skips and excessive overlaps during application. Avoid the use of spreaders that apply this product in narrow rows or concentrated bands.Wash hands with soap and water promptly after use.Do not allow people or pets to contact treated area until after product dust has settled into the turfgrass, or if watered in, after the turfgrass surface in the treated area has dried.WHEN TO APPLYReduce the interval between applications of this product when conditions are favorable for disease development. Unless otherwise specified, when disease pressure is high or when used as a curative, use higher rates of ferti•lome® F-STOP™ and shorter application interval. Under light to moderate disease pressure apply this product at the low use rate and/or longer application intervals. To avoid pick-up, lightly irrigate treated areas soon after application. On short cut bentgrass (1/2 inch or less) when temperatures are above 80°F, apply only to dry foliage.HOW MUCH TO APPLYOptimum disease control is achieved when ferti•lome® F-STOP™ is applied in a preventative disease control program at a rate of 4 lb per 1,000 sq. ft. See the following table for specific application rates for various diseases. Under any circumstances, do not apply more than 46 lb of this product per1,000 sq. ft. per year.SPREADER GUIDEONE BAG WILL COVER UP TO 2,500 SQUARE FEETSPREADER SPREADER SETTINGS Scotts®/Republic Accugreen (Drop) 4 1/4Scotts®/Republic Speedy Green (Broadcast) 3 3/4 ferti•lome® /EarthWay Ev-N-Spred (Broadcast)14TERMS AND CONDITION OF USEIf terms of the following Warranty Disclaimer, Inherent Risks of Use, and Limitation of Remedies are not acceptable, return unopened package at once to the Seller for a full refund of purchase price paid. Otherwise, use by the Buyer or any other user constitutes acceptance of the terms under Warranty Disclaimer, Inherent Risks of Use, and Limitations of Remedies.WARRANTY DISCLAIMERSeller warrants that this product conforms to the chemical description of the label and is reasonably fit for the purposes stated on the label when used in strict accordance with the directions, subjectto the Inherent Risks set forth below. Seller MAKES NO OTHER EXPRESS OR IMPLIED WARRANTY OF MERCHANTABILITY OR FITNESS FOR A PARTICULAR PURPOSE OR ANY OTHER EXPRESS OR IMPLIED WARRANTY.INHERENT RISKS OF USEIt is impossible to eliminate all risks associated with use of this product. Plant injury, lack of performance, or other unintended consequences may result because of such factors as use of the product contrary to label instructions (including conditions noted on the label, such as unfavorable temperature, soil conditions, etc.), abnormal conditions (such as excessive rainfall, drought, tornadoes, hurricanes), presence of other materials, the manner of application, or other factors, allof which are beyond the control of the Seller. To the extent allowed by law, all such risks shall be assumed by the Buyer.LIMITATION OF REMEDIESTo the extent consistent with applicable law, the exclusive remedy for losses or damages resulting from this product (including claims based on contract, negligence, strict liability, or other legal theories), shall be limited to, at Seller’s election, one of the following:1. Refund of purchase price paid by Buyer or user for product bought, or2. Replacement of amount of product used.To the extent allowed by law, Seller shall not be liable for the losses of damages resulting from the handling or use of this product unless Seller is promptly notified of such loss or damage in writing. In no case shall Seller be liable for consequential or incidental damages or losses.The terms of the Warranty Disclaimer and Inherent Risks of Use above and this Limitation of Remedies cannot be varied by any written or verbal statements or agreements. No employee or sales agent of Seller or the Seller is authorized to vary or exceed the terms of the Warranty Disclaimer or this Limitation of Remedies in any manner.Disease ferti•lome® F-STOP™(lb/1,000 sq ft)ApplicationInterval/Timing(Days)Directions RestrictionsAnthracnose Red Thread Septoria Leaf Spot 414 - 21Apply when conditions are favorable for disease development.Do not apply more than 46pounds of product per 1,000sq. ft. per year.For Nassau and Suffolkcounties in New York State,do not apply more than 11.5lb of this product per 1,000sq ft per year.Brown Patch14Begin applications when conditions are favorable for disease development andb efore disease symptoms are apparentCopper SpotZonate Leaf SpotApply when conditions are favorable for disease development.Crown Rot Leaf Spot Melting-Out Apply when conditions are favorable for disease development. For crown rot, water in with 3 to 4 gallons of water per 1,000 sq. ft. to increase penetration to crown a nd roots.Dollar Spot14 - 28Apply when conditions are favorable for disease development. Make no morethan 3 consecutive applications for Dollar Spot control before rotating to aregistered fungicide with a diiferent mode of actionFusarium Blight14 - 21Apply when conditions are favorable for disease development.Fusarium Patch(Pink Snow Mold)21 - 28Apply when conditions are favorable for disease development.Gray Leafspot14Apply prior to snow cover.Leaf Spot Apply in the fall after turfgrass enters dormancy and/or in the spring prior to theinitiation of growth.Necrotic Ring Spot Spring: 28Make applications on a preventative basis in early to mid-spring.Necrotic Ring Spot Fall: 28Make 2 applications beginning in August before the turfgrass goes dormant.Powdery MildewRusts14 - 28Apply when conditions are favorable for disease development.Summer Patch Begin applications in the spring when conditions are favorable for disease de-velopment. Make 2 to 4 applications depending on recommendations from localTurfgrass Extension Experts.Water in with at least 3 to 4 gallons of water per 1,000 sq. ft. to increase penetra-tion to crown and roots.Manufactured by:230 FM 87 • BONHAM, TEXAS 75418EPA Reg. No. 62719-461-7401 EPA Est. No. 7401-TX-01Visit Us At: Product Questions? 855-270-477612770-0515-TP。
Medical image extraction apparatus, medical image
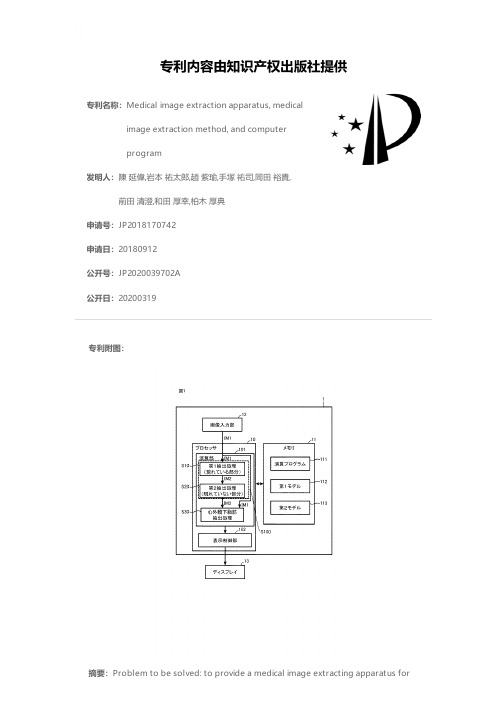
专利名称:Medical image extraction apparatus, medicalimage extraction method, and computerprogram发明人:陳 延偉,岩本 祐太郎,趙 紫瑜,手塚 祐司,岡田 裕貴,前田 清澄,和田 厚幸,柏木 厚典申请号:JP2018170742申请日:20180912公开号:JP2020039702A公开日:20200319专利内容由知识产权出版社提供专利附图:摘要:Problem to be solved: to provide a medical image extracting apparatus forautomatically extracting an image of an internal structure of a living body from a photographing image. Medical image extractor 1Taken from IM1A first portion A1 which can be recognized on a photographing image andThe second part that cannot be recognized on the photographing image andA processor 10 for carrying out image extraction processing for extracting the biological structure a comprisingImage extraction processingA first extraction processing S10 for extracting a first portion from a taken image andA second extraction processing S10 for generating an interpolated body structure is shown in which an image representing the extracted first portion is interpolated.Diagram申请人:学校法人立命館,社会医療法人誠光会地址:京都府京都市中京区西ノ京東栂尾町8番地,滋賀県草津市矢橋町1660国籍:JP,JP代理人:坂本 寛更多信息请下载全文后查看。
儿童神经母细胞瘤的临床分析

另1例胰腺双肾多囊肿,右肾肿块,腹膜后多发淋 巴结肿大,右巨肾及肿瘤切除,术后CT示腹主动 脉旁淋巴结明显肿大,胰头受侵,左肾较多囊肿, 不排除肿瘤侵犯。经OPEC化疗后左肾区肿块缩小, 1月后增大,B超示左肾实性占位肾门淋巴结增大, 予多个化疗方案,肿块反复增大,对化疗敏感,加 强化疗后准备自体干细胞移植,家人决定暂观察, 至今随诊7年未复发。
治疗转归
失访1例,治疗中放弃4例,治疗中感染死亡1例, 复发9例。
复发后6例放弃治疗,1例放疗后失访,1例感染死 亡,1例Ⅳ期患儿手术后复发,维甲酸治疗痊愈。 此患儿1月发病,以双下肢活动障碍就诊,右肝浸 润,MR示右肾后方腰椎旁肿物,边界不清,沿椎 间孔入椎管内致脊髓圆锥受压侵犯肌肉。手术后 MR见右肾门结节,NSE增高,予维甲酸1月后复查 肿瘤侵犯椎板椎管内,随诊自服维甲酸,现痊愈。
讨论:临床表现与诊断
局限性原发性肿瘤的手术切除能提供最好的治愈机 会。36例患儿中仍以腹部包块为首发表现者最多, 发现较早,分期早,即使分期较差者亦无骨髓转移。
重视特征性的熊猫眼,面部肿物、上睑下垂、头外 伤后双眼肿胀失明者亦需及时做影像检查或肿物活 检
原发脑膜浸润 ?
Kushner BH, Kramer K, Modak S, et al
18例治疗患儿中治愈病例5例:⑴3例为Ⅰ 期,其中2例手术切除,1例家人拒绝手术, 行化疗后,手术探查少量黄色炎性渗出腹水, 未见癌细胞。
治疗转归
2例为Ⅳ期,其中1例骨髓、骨转移,予2OPEC/ 2PECA化疗,自服中药,手术切除原发灶,半年后 原发灶、BM复发,CTX+卡铂,再次手术清扫, 化疗+放疗,痊愈。
想到神经母细胞瘤的可能性
经X线检查、B超、NSE、尿VMA定性(必要时做定量)检查 以及骨髓穿刺检查等,90%病例可以不经组织切片而做出明确 诊断。
【精品】设计与自然设计与自然的功能关系PPT资料

第五章 设计与自然的功 能关系
大自然是世界上最好的设计师,自然的进化从整体 到局部都显示出天工出神入化的杰作。生物界有着种类 繁多的动植物及其他生命存在,它们在漫长的进化过程 中,为了求得生存与发展,逐渐具备了适应自然界变化 的本领。人类生活在自然界中,也学会了与周围的生物 友好相处,这些生物所具有的各种奇异能力,吸引着人 们去想象、去模仿。人类运用其观察、思维和设计的能 力,通过对生物的模仿和初步的创造性劳动,制造出简 单的生产工具,用来增强自己与自然界斗争的本领和生 存能力。
仿生学力图找到研究对象的生物原理,通过对生物 的感知,形成对生物体的感性认识;从功能出发,对照 生物原型进行定性的分析。
仿生设计的最初起源
004_Cloaca No. 003 / 004 / 005_ 仿生灯具 . 001 / 002_ 骨椅 . 002 / 003_ 榨汁器 草图 . 从视觉的角度而言,仿生设计学侧重研究生物体的视觉器官对图像的识别、对视觉信号的分析与处理,以及相应的视觉流程,研究的结果可以广泛应用于产品设计、视觉传达设计 和公共环境设计之中。 003 / 004 / 005_ 仿生灯具 .
Fig.002_ 骨骼椅子. 荷兰. Joris Laarman 设计.2006
在工程与产品设计的领域,自然形态所具有的功能, 天然地带有经济实用的原则,从另一方面启发设计为人 类使用的功能产品。从这个意义上讲,功能化的自然形 态有着异常的魅力。
仿生设计学在仿生学和设计学的基础上发展起来, 并且与许多自然和理工学科如生物学、控制论、信息论、 人机学、心理学、材料学、色彩学等相联系。仿生设计 应当成为设计领域的一个重要方式。
001_Cloaca No. 菲利普·斯塔克设计. 生物界有着种类繁多的动植物及其他生命存在,它们在漫长的进化过程中,为了求得生存与发展,逐渐具备了适应自然界变化的本领。 第五章 设计与自然的功能关系 生物界有着种类繁多的动植物及其他生命存在,它们在漫长的进化过程中,为了求得生存与发展,逐渐具备了适应自然界变化的本领。 生物界有着种类繁多的动植物及其他生命存在,它们在漫长的进化过程中,为了求得生存与发展,逐渐具备了适应自然界变化的本领。 Pascal Mourgue 设计 003 / 004_ 可充填坐垫的骨椅. 001 / 002 / 003_ 壁上仿生机器人.