【推荐下载】关于半夏种质资源的随机扩增多态性DNA技术分析
随机扩增多态性DNA分析机械通气性肺炎大肠杆菌的多样性

Di v e r s i t y o f Es c h e r i c h i a c o l i i n me c ha n i c a l v e n t i l a t e d p n e u mo n i a
b y r a nd o m a mp l i ie f d p o l ym o r p hi c DNA
B 【 s 阳 性 检 出率 高 , 应 引起 临 床 的 重 视 。
3 - 内酰胺 酶 ; 基 因分型 [ 中 图分 类号 ] R 5 6 3 . 1 [ 文 献标 识码 】 A 【 文章 编 号 ] l 6 7 2 - 7 3 4 7 ( 2 0 0 7 ) 0 2 . 0 3 5 5 . 0 4
t e d p n e u mo n i a nd a t o e x p l o r e he t me c h a n i s ms f o r d r u g es r i s t a n c e.M e t h o d s RAPD— P CR wa s u s e d t o a l l - ly a z e he t in f g e r p r i n in t g o f2 0 s t r a i n s o fEs c h e r i c h i a c o l i .ES BL s we e r d e t e c t e d b y d o u b l e d i s k t e s t .Re s ul t s Twe n t y c l i n i c l a s t r a i n s o f Es c h e r i c h i a l c o l i we e r d i v i d e d i n t o 1 0 t y p e s b y RAP D.T h e p o s i t i v e r a t e f o ES BL s wa s 65. 0% . S e v e n t e e n s t r a i n s h a d t h e s a me l i n e i n 1 4 47 b p. Co n c l u s i o n T h e g e n e t y p e o f E s c h e r i c h i a c o l i n me c h a ni c l v a e n t i l a t d e p n e m o u ia n i s d i v e si r ie f d. Pa t i e n t s a r e ma i n l y i n f e c t e d i n h o s p i — t ls a .T h e d e t ct e i o n at r e o f ES BL s i s h J ,wh i c h s h o u l d b e p a i d mu c h a t t e n t i o n t o.
【国家自然科学基金】_随机扩增多态性dna(rapd)_基金支持热词逐年推荐_【万方软件创新助手】_20140730
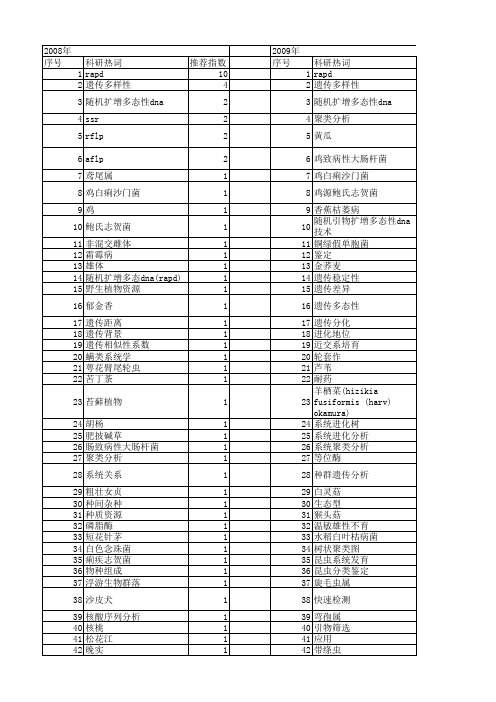
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
推荐指数 14 3 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
科研热词 推荐指数 rapd 10 遗传多样性 4 随机扩增多态性dna 2 ssr 2 rflp 2 aflp 2 鸢尾属 1 鸡白痢沙门菌 1 鸡 1 鲍氏志贺菌 1 非混交雌体 1 霜霉病 1 雄体 1 随机扩增多态dna(rapd) 1 野生植物资源 1 郁金香 1 遗传距离 1 遗传背景 1 遗传相似性系数 1 螨类系统学 1 萼花臂尾轮虫 1 苦丁荼 1 苔藓植物 1 胡杨 1 肥披碱草 1 肠致病性大肠杆菌 1 聚类分析 1 系统关系 1 粗壮女贞 1 种间杂种 1 种质资源 1 磷脂酶 1 短花针茅 1 白色念珠菌 1 痢疾志贺菌 1 物种组成 1 浮游生物群落 1 沙皮犬 1 核酸序列分析 1 核桃 1 松花江 1 晚实 1 早实 1 新疆 1 抗病基因 1 差异性 1 多元回归 1 基因组dna 1 地理隔离 1 地理种群 1 加拿大披碱草 1 人参 1
半夏育种实验报告

一、实验背景半夏(学名:Pinellia ternata),又称三步跳、地栗子等,是天南星科植物,其干燥块茎入药,具有显著的药用价值。
半夏在我国传统医学中被广泛使用,具有治疗伤寒寒热、心下坚、喉咽肿痛、头眩、胸张、咳逆、肠鸣、止汗等功效。
然而,随着半夏种植面积的扩大和市场需求的增长,半夏的种源混杂、品质退化、病虫害严重等问题日益突出,严重影响了半夏药材的质量和临床用药的安全。
因此,开展半夏育种实验,培育优质、高产、抗病虫害的半夏新品种,对于保障半夏药材的质量和供应具有重要意义。
二、实验目的1. 通过半夏育种实验,筛选出优质、高产、抗病虫害的半夏新品种。
2. 探究半夏的遗传规律,为半夏遗传育种提供理论依据。
3. 优化半夏种植技术,提高半夏产量和品质。
三、实验材料与方法1. 实验材料实验材料包括半夏种子、半夏块茎、生长调节剂、农药等。
2. 实验方法(1)半夏种子处理将半夏种子浸泡在5%的氢氧化钠溶液中,处理时间为15分钟,然后用清水冲洗干净。
(2)半夏播种选择肥沃、排水良好的砂质土壤作为播种地,按照行距20~25厘米、株距2~5厘米进行播种,亩用种量60公斤左右。
(3)半夏田间管理① 水分管理:半夏喜湿润,怕干旱,生长期间要保持土壤湿润。
在干旱季节,可适当增加灌溉次数。
② 肥料管理:在播种前,每亩施入腐熟农家细肥或土杂肥1500~2000公斤、过磷酸钙20~25公斤。
生长期间,根据植株生长情况,适时追肥。
③ 病虫害防治:根据病虫害发生情况,选用高效、低毒、低残留的农药进行防治。
(4)半夏育种方法① 杂交育种:采用半夏与不同品种的半夏进行杂交,筛选出优良杂交种。
② 选择育种:根据半夏的产量、品质、抗病虫害能力等性状,选择优良单株进行繁殖。
③ 基因工程育种:利用分子生物学技术,将优良基因导入半夏,培育抗病虫害、高产、优质的新品种。
四、实验结果与分析1. 半夏种子发芽率实验结果显示,经过氢氧化钠溶液处理的半夏种子发芽率显著提高,发芽率可达90%以上。
随机扩增多态性DNA标记技术及其在药用植物研究中的应用

随机扩增多态性DNA标记技术及其在药用植物研究中的应用作者:王晓慧汤晓闯杨恩秀姜程曦董建勇黄志锋【摘要】随机扩增多态性DNA(RAPD)技术是近年来广泛应用的分子标记技术之一,以PCR反应为基础,其特点是快速简便、易操作、成本较低,DNA 需要量少、无需放射性分析,也不会污染等。
该文简述了RAPD技术的原理及优缺点以及其在药用植物遗传多样性分析、药材鉴别和DNA指纹图谱的构建、亲缘关系及系统学研究、药材的道地性等方面的应用,并展望了其发展前景。
【关键词】随机扩增多态性DNA 遗传多样性药材鉴别道地性分子标记(Molecular marker)是以个体间遗传物质内核苷酸序列变异为基础的遗传标记,是DNA水平遗传多态性的直接反映,以其快速、准确、所提供的信息量大、不受环境影响等特点,已被广泛应用于遗传学评价、目的基因定位和遗传图谱的构建等领域。
随机扩增多态性DNA(RAPD)技术是近年来广泛应用的分子标记技术之一,以PCR反应为基础,其特点是快速简便、易操作、成本较低、DNA需要量少、无需放射性分析也不会污染等[1]。
1 RAPD技术的原理RAPD ( random amplified polymorphic DNA )是 1990 年美国杜邦公司科学家 J. G. K. Williams 和加利福尼亚生物研究所 J. Welsh 领导的两个小组几乎同时发展起来的一项新的分子标记技术。
Williams 称之为 RAPD,Welsh 称之为 AP-PCR(arbitrary primer PCR)。
其建立在 PCR 技术基础上,是以任意序列的寡核苷酸单链 ( 通常为10个碱基,AP-PCR 则为20~30个碱基) 为引物,对所研究的基因组 DNA 进行随机扩增。
RAPD 所用的一系列引物的 DNA 序列各不相同,但对于任一引物,它同基因组 DNA 序列有特定的结合位点。
这些特定的结合位点在基因组某些区域内的分布如符合 PCR 扩增的反应条件,即在一定范围内模板 DNA 上有与引物互补的反相重复序列时,就可扩增出此范围的 DNA 片段。
利用随机扩增多态性DNA技术分析刺五加的遗传多样性

利用随机扩增多态性DNA技术分析刺五加的遗传多样性【摘要】目的从DNA水平对收集的4个产地的刺五加进行遗传多样性分析,为更好地开发利用刺五加资源奠定基础。
方法随机扩增多态性DNA(RAPD)技术分析遗传多样性,UPGMA进行聚类分析。
结果10个随机引物扩增得到72条片段,多态性位点比率为88.89%。
不同产地间的遗传相似系数在0.469 3~0.785 1之间,均值为0.629 3,同一产地不同居群间的遗传相似系数在0.563 0~0.954 2之间,均值为0.764 4。
不同产地刺五加之间的遗传多样性高于同一产地不同居群的个体,其中产地穆棱的刺五加与产地和龙、本溪、尚志之间的相似性较低,单成一支。
产地之间的相似性与地理距离呈不完全相关性。
结论刺五加种群内部具有较高的遗传多样性,利用RAPD技术分析刺五加的遗传多样性是可行的。
【关键词】刺五加遗传多样性扩增多态性DNAAbstract:Objective In order to develop and utilize the resource of Eleutherococcus senticosus ,the genetic persity of Eleutherococcus senticosus from four habitats were analyzed by RAPD analysis. MethodsRAPD was used to analyze the genetic similarity,then cluster analysis was carried out by UPGMA method.ResultsTen random primers generated 72 bands (rate of polymorphic bands was 88.89%). The genetic similarity index among different habitats were 0.469 3~0.785 1 (the mean value was 0.629 3), while the genetic similarity index among different populations of Eleutherococcus senticosus from the samehabitat were 0.563 0~0.954 2 (the mean value was 0.764 4). The cluster analysis showed that the genetic persity of Eleutherococcus senticosus from different habitats was higher than that of Eleutherococcus senticosus from different populations grown in the same habitat and the similarity of Eleutherococcus senticosus from Muling separated with that from other habitats from Helong, Benxi and Shangzhi. The results also showed that the similarity among habitats was partly in agreement with the geographic distance. ConclusionThe genetic persity were high in the population of Eleutherococcus senticosus,so it is feasible to analyze the genetic persity of Eleutherococcus senticosus from differenthabitats by RAPD.Key words:Eleutherococcus senticosus; Genetic persity; RAPD刺五加Eleutherococcus senticosus是我国医药珍品,根、茎、叶均可入药。
随机扩增多态性DNA标记技术

随机扩增多态性DNA标记技术参考周延清2005.生物实验室系列---DNA分子标记技术在植物研究中的应用. [北京]化学工业出版社P79-130一、引言随机扩增多态性DNA标记技术,检测出核酸碱基序列的变异,并且将这种变异以遗传标记的方式予以应用,使植物遗传学许多方面的研究产生了革命性的变化。
最早检测到的DNA水平的变化是限制性片段长度多态性(RFLP)。
但是,在过去一段时间,聚合酶链反应(Polymerase chain reaction, PCR)极大的影响了分子生物学几乎所有领域,而且基本程序被改进后,可以开发出多种检测核苷酸水平差异的方法。
不过,这些方法大多需要预先知道DNA片段序列的一些信息,以便根据已知的信息设计合成目的的DNA序列两端的引物,通过PCR选择性地扩增目的的DNA。
1990年,Williams等发表了一种检测核苷酸序列多态性的新方法,这种方法以PCR为基础,可不必预先知道DNA序列的信息。
随后,随机扩增多态性DNA技术(randomly amplified polymorphic DNA,RAPD)广泛地应用于植物和其它领域的研究中。
RAPD标记技术由于操作简便、快速、省时、省力、DNA用量少,迅速受到人们重视,并在农、林、医及植物和微生物学的各个领域中得到广泛应用,在基因定位与分离、连锁和系统演化等各方面取得了很大的进展。
二、RAPD标记技术的概念和原理任何生物种都具有特定顺序和结构的遗传物质---------DNA。
由于在生物进化过程中选择性的不同。
生物基因组DNA的不同区域表现出高度保守或高度变异的现象,具有不同的遗传多样性。
随机扩增多态性DNA(RAPD)标记技术是通过分析遗传物质DNA经过PCR扩增的多态性来诊断生物体内在基因排布与外在性状表现的规律的技术,由于片段被引物选择性地扩增,扩增了的片段能在凝胶上清晰地显现出来,这样就可以通过同种引物扩条带的多态性反映出模板的多态性,RAPD只需要一个引物,长度为10个核苷酸左右,引物顺序是随机的,因而可以在对被检测对象无任何分子生物学资料的情况下对其基因组进行分析。
半夏的研究进展

半夏的研究进展一、本文概述半夏,作为中医药学中的一种重要药材,历史悠久,应用广泛。
其独特的药理作用及在多种疾病治疗中的显著疗效,使得半夏的研究一直备受关注。
本文旨在系统梳理和综述半夏的研究进展,以期为半夏的深入研究及临床应用提供参考。
文章将从半夏的化学成分、药理作用、临床应用以及现代研究技术等方面进行全面介绍,并对未来的研究方向进行展望。
通过对半夏的深入研究,有望为传统中医药的发展和创新提供新的思路和方法。
二、半夏的化学成分与药理作用半夏,作为一种传统中药材,具有悠久的历史和广泛的应用。
其独特的化学成分和药理作用一直是中医药研究的热点。
近年来,随着科学技术的进步,对半夏的化学成分和药理作用的研究也取得了显著的进展。
半夏含有多种复杂的化学成分,主要包括生物碱类、多糖类、皂苷类、挥发油等。
其中,生物碱类成分是半夏的主要药效成分,如半夏碱、次半夏碱等。
半夏还含有多种微量元素和氨基酸,这些成分共同构成了半夏独特的药理作用基础。
半夏具有广泛的药理作用,主要包括镇咳祛痰、止吐抗晕、抗炎抗氧化等。
在镇咳祛痰方面,半夏能够通过刺激呼吸道黏膜,增加分泌物的排出,从而有效缓解咳嗽和痰多症状。
在止吐抗晕方面,半夏能够调节中枢神经系统,抑制恶心和呕吐的反射,对于晕车、晕船等晕眩症状有良好的缓解作用。
半夏还具有显著的抗炎抗氧化作用,能够减轻炎症反应,保护细胞免受氧化损伤。
近年来,随着对半夏药理作用的深入研究,还发现半夏具有抗肿瘤、抗心血管疾病等新的药理作用。
研究表明,半夏中的某些成分能够抑制肿瘤细胞的增殖和转移,对于某些癌症的治疗具有一定的辅助作用。
半夏还能够调节心血管功能,降低血压、改善心肌缺血等,对于心血管疾病的预防和治疗具有一定的意义。
半夏作为一种传统中药材,其化学成分和药理作用的研究取得了显著的进展。
未来,随着科学技术的不断发展,相信对半夏的研究将更加深入,为中医药的发展和人类健康做出更大的贡献。
三、半夏的现代研究与应用近年来,随着科学技术的不断进步,半夏的研究和应用也取得了显著的进展。
贵州野生半夏资源遗传多样性的ISSR分析

图 2 不 同 分布 区样 品 的 IS S R扩 增 (9 ) 89 电泳 图谱
13 引物 筛选与 P R扩增 . C 以表 型差异较 大 的柳 叶半 夏 、 叶半 夏 和 珍珠 半 桃
距离 , S S 在 P S软件上 对遗传 距离 矩阵 和地理距 离矩 阵
进 行相关 性分 析 。
夏 为模 板进 行筛选 , 1 0条 引物 中共筛选 出 l 从 0 6条多 态 性好 、 带 清 晰 的引 物 , 退火 温 度 和序 列 见 表 1 条 其 。
较大 , 分布在 3 . % ~ 0 3 之 间 , 8O 6 .% 平均 为 5 . % , 3 4 高 于 5 % 的 样 品 占全部 贵 州样 品的 2 5 分 布 于全 省各 5 /,
为 了确保 扩增谱 带 的可 靠性 , 每个 引 物 重 复扩 增
2次 , 将重 复 出现 ቤተ መጻሕፍቲ ባይዱ带记 为 1 无 带 记 为 0 排 除 模糊 不 , , 清 的带 和无法 准确标 识 的带 , 计供 试 材 料 的 多态 性 统 数 据 。应 用 P P E E vr o . 2 软 件在 假 定样 品 O G N ( es n1 3 ) i 处于 H ry ib r 平 衡状 态下 , 算 6 ad We eg n 计 0个 样 品 的基
2 2 不 同样 品 的多态性 位点百 分率 .
2 / 0 L反应体 系 , 内含 1 ufr15m o LMg1, ×bf ,. m l C2 e /
2 0 N P , a N 聚合 酶 ( 京 天 根 ) 5 Ⅱ L d T s 1U T qD A 北 , 0 8 I L引物 ( . n / 上海 生工 ) 2 g模 板 D A ,0n N 。反 应程 序为 : 9 4℃预 变性 5 n 9 ;4℃变 性 3 退火 4 i, mi 0S, 5r n a 7 2℃延 伸 2mi ,5个 循环 ;2℃延 伸 7r n 保 n 3 7 i。4c a C I 存 。P R扩增 反应 产物 用 15 的琼脂糖凝 胶 ( E C .% 含 B
远志种质资源遗传多样性随机扩增多态性DNA分析(一)

远志种质资源遗传多样性随机扩增多态性DNA分析(一)【摘要】目的用随机扩增多态性DNA(RAPD)技术分析远志PolygalatenuifoliaWilld.及卵叶远志P.sibiricaL.的遗传多样性。
方法反应总体积25μl,内含10×PCRbuffer2.5μl,dNTPs2.0μl,MgCl22.0μl,引物1.0μl,模板2ng,Taq酶0.3μl。
扩增程序为:94℃预变性4min,然后进行45循环:94℃变性15s,36℃复性1min,72℃延伸1.3min,循环结束后72℃延伸4min。
结果10个引物共检测到154个位点,平均每个引物扩增9.24条带,其中148条带表现为多态性,占总带数的94.3%;距离系数为24时,11个样品明显聚为远志居群和卵叶远志居群两类。
结论远志与卵叶远志具有丰富的遗传多样性,遗传聚类与其地理分布有一定的相关性。
【关键词】远志;卵叶远志;遗传多样性;随机扩增多态性DNAAbstract:ObjectiveToanalyzegeneticdiversityofPolygalatenuifoliaWilldandP.sibiricaL.withRAPD-PC R.MethodsTheRAPD-PCRreactionsyst emwasconstructedwith2.5μl10×PCRbuffer,2.0μldNTPs,2.0μl MgCl2,1.0μlprimer,2ngtemplateand0.3μlTaqwithtotalreactionvolumeof25μl.PCRamplifyingseque nceswereasfollows:Initialdenaturatingfor4minat94℃,thendenaturatingfor15sat94℃,annealingfor 1minat36℃andextensionfor1.3minat72℃.After45timescycle,keepingextensionfor4min.Results15 4polymorphismstrandsweredetectedwith10primers,amongwhich148strandswereofpolymorphis m.Clusteranalysisindicatedthatallsamplescouldbeclusteredintotwopopulationswithdistancecoeffic ientof24.ConclusionPolygalatenuifoliaWilldandP.sibiricaL.haverichgeneticdiversity.Thegeneticclust eringhassomerelevancewithgeographicaldistributionofP.tenuifoliaWilldandP.sibiricaL. Keywords:PolygalatenuifoliaWilld.;PolygalasibiricaL.;Geneticdiversity;RAPDPCR远志来源于远志科(Polygalaceae)植物远志Polygalatenuifoliawilld或卵叶远志P.sibiricaL.的干燥根,临床应用广泛。
利用随机扩增多态性DNA技术分析刺五加的遗传多样性

LS IH NM DCN N A E I E IAR SA C 0 O .9N . IHZ E E IIEA DM T RAM DC E E R H2 8V L 1 O6 0
时珍 国医国药 20 0 8年第 1 9卷第 6期
eo o c ss n i s s f m u a i t w r n lz d b AP a ay i.M e h d r c c u e t o u r f rh bt s e e a ay e y R D n ss c o o a l t o s RAP s u e n y et e g n t i lr D wa s d t a a z h e ei smi i o l c a
t , e ls r n y i w s a ido t yU G to . s l e n o r es e e td 2 b n s(ae f oy o hc y t ncut a s a cr e u b P MA me d Reut T nr d m p i r g nr e a d rt o lm r i h e al s r h s a m a 7 p p
行 的。
关 键词 : 刺五加 ; 遗传 多样性 ; 扩增 多态性 D A N
中图分 类号 :5 7 ¥6
文献 标 识码 : A
文章 编 号 :0 80 0 (0 8 0 .3 5 2 10 -8 5 2 0 )6 10 - 0
Ge e i v r iy o e t r c c uss n i o u y RAPD n te Di e st fEl u he o o c e tc s s b Ana y i l ss
X N h o bn I G Z a — i ,L 0 F n — u A e g y n ,WU P n I u y n , H N H i o g e g ,L — a S E a — n Y l
半夏不同种质资源主要农艺性状与产量的相关及通径分析

半夏不同种质资源主要农艺性状与产量的相关及通径分析
柳 敏,王彩云,李恒谦,周茂嫦,成忠均,张翔宇 ∗
( 毕节市中药研究所,贵州毕节 551700)
摘要 以 21 份不同来源地半夏种质资源为研究对象,对各种质资源株高、茎粗、分茎数等主要农艺性状进行相关性和通径分析,得出影
positively with the plant height, stem diameter, tillering number, fibrous roots number, tuber hundred grain weight, bulbil hundred grain
weight and the distance from tuber to bulbil. The direct path coefficient of the effect degree of each agronomic trait on the yield of P. ternata
文章编号 0517-6611(2023)22-0038-04
doi:10.3969 / j.issn.0517-6611.2023.22.011
开放科学(资源服务)标识码(OSID):
Correlation and Path Analysis on Main Agronomic Characters and Yield of Pinellia ternata Germplasm Resources
夏高产品种引种选育,对打造半夏区域品牌具有重要意义。
种质资源,但未开展其他叶型的相关研究。 笔者将收集的 21
份不同半夏种质资源为材料,通过主要农艺性状的相关性和
中国野生半夏的遗传多样性和遗传结构研究

㊀Guihaia㊀Apr.2024ꎬ44(4):766-776http://www.guihaia-journal.comDOI:10.11931/guihaia.gxzw202207008吴敏ꎬ吴诗琪ꎬ潘凤ꎬ等ꎬ2024.中国野生半夏的遗传多样性和遗传结构研究[J].广西植物ꎬ44(4):766-776.WUMꎬWUSQꎬPANFꎬetal.ꎬ2024.GeneticdiversityandgeneticstructureofwildPinelliaternata(Araceae)inChina[J].Guihaiaꎬ44(4):766-776.中国野生半夏的遗传多样性和遗传结构研究吴㊀敏ꎬ吴诗琪ꎬ潘㊀凤ꎬ石㊀甜ꎬ赵㊀财∗(贵州大学生命科学学院/农业生物工程研究院ꎬ山地植物资源保护与种质创新教育部重点实验室ꎬ山地生态与农业生物工程协同创新中心ꎬ贵阳550025)摘㊀要:第四纪气候波动以及地理和环境隔离深刻地影响了现代植物的遗传多样性㊁遗传结构和地理分布格局ꎮ该研究采用分子谱系地理学的研究方法对药用植物半夏19个居群共212个个体的3个叶绿体片段psbK ̄psbI㊁atpF ̄atpH和trnL ̄F进行分析ꎬ探究半夏的遗传多样性㊁遗传结构㊁地理分布格局模式及成因ꎬ并探讨其居群历史动态ꎮ结果表明:(1)半夏总单倍型多样性Hd为0.882ꎬ总核苷酸多样性π为1.23ˑ10 ̄3ꎬ在物种水平上表现出较高的遗传多样性ꎮ(2)分子方差分析(AMOVA)结果显示ꎬ半夏遗传变异主要发生在居群间ꎬ显著的遗传分化(FST=0.909ꎬP<0.001)和较低的种群内遗传多样性(HS=0.134)ꎻ种群间遗传分化系数NST=0.913>GST=0.855(0.01<P<0.05)ꎬ表明叶绿体单倍型具有明显的谱系地理结构ꎮ(3)中性检验结果显示ꎬTajima sD值㊁FuandLi sD值以及FuandLi sF∗值均为不显著正值ꎬFu sFs值为不显著负值且失配分析曲线呈双峰ꎬ表明半夏居群整体没有经历过扩张事件ꎮ(4)单倍型地理分布显示ꎬ西南地区和中-东部地区具有单倍型多样性较高ꎬ并存在特有单倍型ꎬ故推测第四纪冰期时在这两个区域存在冰期避难所ꎮ总之ꎬ通过3个叶绿体基因对不同区域半夏的分析ꎬ阐明了其遗传多样性㊁遗传结构和地理分布格局ꎬ为半夏优良种源的分子筛选和保护提出了科学的建议和保护策略ꎮ关键词:半夏ꎬ地理分布格局ꎬ遗传多样性ꎬ遗传结构ꎬ避难所中图分类号:Q948㊀㊀文献标识码:A㊀㊀文章编号:1000 ̄3142(2024)04 ̄0766 ̄11GeneticdiversityandgeneticstructureofwildPinelliaternata(Araceae)inChinaWUMinꎬWUShiqiꎬPANFengꎬSHITianꎬZHAOCai∗(KeyLaboratoryofPlantResourceConservationandGermplasmInnovationinMountainousRegion(MinistryofEducation)ꎬCollaborativeInnovationCenterforMountainEcology&Agro ̄Bioengineering(CICMEAB)ꎬCollegeofLifeSciences/InstituteofAgro ̄bioengineeringꎬGuizhouUniversityꎬGuiyang550025ꎬChina)收稿日期:2023-06-18㊀接受日期:2023-07-20基金项目:贵州科技支撑计划项目(黔科合支撑[2019]2451 ̄2号)ꎻ贵州省教育厅教育部重点实验室开放课题(黔教合KY字[2019]033)ꎻ贵州省生物学国内一流建设学科开放基金(GNYL[2017]009)ꎻ贵州省中药材现代产业技术体系建设项目(GZCYTX ̄02)ꎻ贵州省梵净山地区生物多样性保护与利用重点实验室开放课题基金([2020]2003)ꎮ第一作者:吴敏(1996 )ꎬ硕士研究生ꎬ主要从事植物系统与进化生物学研究ꎬ(E ̄mail)2495649994@qq.comꎮ∗通信作者:赵财ꎬ博士ꎬ副教授ꎬ主要从事植物系统与进化㊁植物多样性及保护方向研究ꎬ(E ̄mail)zhaocai_11@163.comꎮAbstract:Duetoglobalclimatechange(especiallytherepeatedfluctuationoftheQuaternaryclimate)andenvironmentalisolationhaveaprofoundimpactonthegeneticdiversityꎬgeneticstructureꎬgeographicaldistributionpatternandpopulationhistoricaldynamicsofmodernplants.Weusedtheresearchmethodofmolecularpedigreegeographytoanalyzethethreenon ̄codingchloroplastfragmentspsbK ̄psbIꎬatpF ̄atpHandtrnL ̄Fof212individualsfrom19populationsinChinaofPinelliaternataꎬaAraceaePinelliaperennialherbwithhighvalueofmedicinꎬtoexplorethegeneticdiversityꎬgeneticstructureꎬgeographicaldistributionpatternanditscausesꎬandalsoexplorethehistoricaldynamicsofitspopulation.Theresultswereasfollows:(1)Thetotalhaplotypediversity(Hd)ofP.ternatawas0.882andthetotalnucleotidediversity(π)was1.23ˑ10 ̄3ꎬwhichmeanshighgeneticdiversityatthespecieslevel.(2)Theresultsofmolecularanalysisofvariance(AMOVA)showedthatgeneticvariationofP.ternataoccurredmainlyin89.27%ofpopulationsꎬwithsignificantgeneticdifferentiation(FST=0.909ꎬP<0.001)andlowerwithinpopulationgeneticdiversity(HS=0.134).Thecoefficientofgeneticdifferentiationamongpopulationswas0.855and0.913respectivelyꎬNST>GST(0.01<P<0.05)indicatedthatthechloroplasthaplotypeshadobviousmolecularlineageandgeographicalstructure.(3)ThecentraltestresultsshowedthatTajima sDvalueꎬFuandLi sDvalueandFuandLi sF∗valuewereinsignificantlypositivevalue.Fu sFsvaluewasinsignificantlynegativevalue.AndthemismatchanalysiscurveshoweddoublepeaksꎬindicatingthattheP.ternatapopulationshadnotexperiencedexpansionevents.(4)Thegeographicaldistributionofhaplotypeshowedthatthesouthwestandcentraleasternregionswerecharacterizedbyhigherhaplotypediversityꎬandtherewereuniquehaplotype.ForthisreasonꎬitisspeculatedthattherewereiceagesheltersinthesetworegionsduringtheQuaternaryiceage.InawordꎬthroughtheanalysisofthreechloroplastgenesofP.ternataindifferentregionsꎬwehaveclarifieditsgeneticdiversityꎬgeneticstructureandgeographicaldistributionpattern.InadditionꎬwehaveputforwardscientificsuggestionsandprotectionstrategiesformolecularscreeningandprotectionofexcellentprovenancesofP.ternata.Keywords:Pinelliaternataꎬgeographicaldistributionpatternꎬgeneticdiversityꎬgeneticstructureꎬglacialrefugia㊀㊀全球气候变化ꎬ尤其第四纪以来的气候波动和环境变化ꎬ深刻地影响了现代植物的遗传多样性㊁遗传结构和地理分布格局(Harrisonetal.ꎬ2001ꎻHewittꎬ1996ꎬ2004ꎻBennett&Provanꎬ2008)ꎮ在欧洲和北美洲ꎬ植物谱系地理学的研究得到了广泛的开展ꎬ揭示了植物现存种群的遗传结构和演化历史进程(Hickersonetal.ꎬ2010ꎻQiuetal.ꎬ2011ꎻLiuetal.ꎬ2012ꎻYeetal.ꎬ2017)ꎮ冰期来临时ꎬ广泛发育的大陆冰川使得高纬度地区的喜温植物退缩到南方ꎬ形成 冰期避难所 (Hewittꎬ2004)ꎻ相反ꎬ间冰期或冰后期气候变暖ꎬ冰川消退ꎬ避难所的植物迁移回到高纬度地区(Hewittꎬ1996ꎬ2004ꎻSoltisetal.ꎬ2006ꎻShaferetal.ꎬ2010)ꎮ然而ꎬ全球气候变化和环境波动对我国植物的遗传多样性㊁遗传结构㊁谱系地理分布模式和种群动态历史的影响迥异于欧洲和北美地区(Qiuetal.ꎬ2011ꎻLiuetal.ꎬ2012ꎻMengetal.ꎬ2015ꎻWangetal.ꎬ2017)ꎬ表现在以下几个方面: (1)中国境内由于未受大规模冰川覆盖ꎬ冰期到来时ꎬ气温降低ꎬ植物在分布区发生范围性收缩ꎬ存在多个冰期避难所ꎻ(2)间冰期或冰后期ꎬ由于气候变暖ꎬ存活于冰期避难所的个体发生范围性扩张ꎻ(3)青藏高原隆起引起一系列山脉㊁河流隔离ꎬ季风引起的干湿交替㊁干旱㊁第四纪冰期-间冰期交替等是植物谱系地理格局形成的主要因素ꎮ但这些研究主要集中在青藏高原㊁西南㊁秦岭及临近地区ꎬ并且主要以木本植物为研究对象ꎬ而对其他区域ꎬ以及草本植物开展类似的研究相对较少ꎮ植物叶绿体DNA(chloroplastDNAꎬcpDNA)为闭环双链DNAꎬ与核基因组DNA相比ꎬ叶绿体DNA具有分子量小㊁多拷贝和结构简单的特点且不同的cpDNA片段间很难发生重组行为ꎬ故在植物居群研究中的遗传分化水平较高ꎬ基因流较低ꎬ能够更明确地阐述物种的遗传变异和谱系地理格局(Aviseetal.ꎬ1987ꎻAviseꎬ1998ꎬ2000ꎻSudhiretal.ꎬ2016)ꎮ因此ꎬ越来越多的研究开始基于多个叶绿体基因开展植物的遗传多样性和遗传结构ꎬ谱系地理分布格局和种群历史动态研究(赵玉芬ꎬ2022)ꎮ半夏(Pinelliaternata)为天南星科(Araceae)7674期吴敏等:中国野生半夏的遗传多样性和遗传结构研究半夏属(Pinellia)多年生草本植物ꎬ别名守田㊁地文等ꎬ民间又称麻芋头㊁天落星㊁无心菜等ꎬ是临床常用中药ꎬ生长在海拔2500m以下潮湿㊁温暖㊁荫蔽㊁疏松的砂质土壤中ꎬ为东亚特有种ꎮ在我国除内蒙古㊁青海㊁新疆和西藏外各省区均有野生半夏的分布(李玉先等ꎬ2004ꎻ杜鹃ꎬ2006ꎻ张君毅ꎬ2007ꎻ陈黎明等ꎬ2020)ꎮ由于地理距离的原因ꎬ种群和表型的分化随着时间的推移而发生ꎬ导致其遗传多样性和遗传结构受到影响ꎮ同时由于广泛的商业开发以及保护措施的缺乏ꎬ导致半夏野生资源的数量急剧下降ꎬ因此ꎬ应对其野生资源做好收集和遗传背景分析等方面的工作ꎮ随着半夏利用范围的不断扩展ꎬ有关研究也不断丰富ꎮ然而ꎬ研究主要集中在化学成分㊁毒性以及药理作用等方面(李哲等ꎬ2021)ꎮ少数学者对半夏的系统发育㊁亲缘演化关系等方面进行了研究ꎮ例如ꎬ赵月梅和李筱玲(2016)利用matK+rbcL联合序列分析半夏属植物的系统发育亲缘关系ꎬ表明matK+rbcL联合序列可以作为DNA条形码鉴别半夏属物种ꎻ张君毅(2007)用ITS序列分析我国不同地区的半夏的序列差异及其与其地理分布和外部形态的相关性ꎬ并表明半夏rDNA变异与其地理分布相关ꎬ以及潘凤等(2021)用ITS序列揭示半夏的群体遗传结构和遗传多样性ꎻ张景等(2021)利用筛选出的SSR引物分析不同居群间半夏的遗传多样性ꎻ郑丹书等(2013)利用叶绿体非编码区(psbK ̄psbI和atpF ̄atpH)对半夏及近缘种进行序列分析ꎬ得到较丰富的变异位点ꎬ但仍未基于多个叶绿体基因全面开展半夏遗传多样性和遗传结构㊁谱系地理分布格局模式和种群历史动态研究ꎮ半夏作为一个草本㊁广布种ꎬ生态适应强ꎬ跨越了不同气候带和多个生物多样性研究热点区域ꎬ是探究草本植物和广布种遗传结构ꎬ谱系地理分布模式和种群历史动态研究的理想材料ꎮ本研究采用植物谱系地理学的研究方法ꎬ基于3个叶绿体基因psbK ̄psbI㊁atpF ̄atpH和trnL ̄F对华东㊁华中㊁西北㊁西南等地区的15个省ꎬ19个自然居群共212个野生材料进行分析ꎬ探究药用植物半夏的遗传多样性和遗传结构ꎬ谱系地理分布模式和种群历史动态ꎬ旨在揭示半夏不同居群间和居群内的遗传变异㊁单倍型地理分布格局ꎬ推断其在第四纪冰期的避难所ꎬ并为半夏优良种源的分子筛选和保护提出科学的建议和保护策略ꎮ本研究为半夏资源保护㊁利用和分子辅助育种提供重要参考和依据ꎮ同时ꎬ也为进一步探讨中国植物区系演化及物种多样性形成提供一定的理论基础ꎮ1㊀材料与方法1.1实验材料半夏材料于2015年至2020年采集和保存ꎬ覆盖了贵州㊁浙江㊁甘肃和四川等15个省区ꎬ横跨中国南北方ꎬ并呈现华东㊁华中㊁西北㊁西南等不同方位和区域自然分地理布区ꎬ共采集到19个自然居群共212个个体ꎬ位置信息采用全球定位系统测量(表1)ꎮ野外采集半夏的新鲜叶片后立即用硅胶干燥保存ꎬ用于总DNA提取ꎮ所采集的凭证标本存放于贵州大学生命科学学院植物标本室ꎮ1.2DNA提取及扩增测序采用新型植物基因组DNA提取试剂盒(离心柱型)从硅胶干燥的叶片中提取半夏总DNAꎮ引物参考已经发表半夏的序列及相关报道(郑丹书等ꎬ2013)ꎬ筛选出3对叶绿体基因片段psbK ̄psbI㊁atpF ̄atpH和trnL ̄Fꎬ并由生工生物工程(上海)股份有限公司合成引物ꎮ引物序列及PCR扩增程序见表2ꎮPCR扩增反应体系均为25μLꎬ2ˑTaqPCRMasterMix12.5μLꎬ包括ddH2O8.5μLꎬ正向引物1μLꎬ反向引物1μLꎬDNA模板2μLꎮ采用1%琼脂糖凝胶电泳法检测总DNA和PCR产物ꎬ电泳结束后将凝胶取出放置于凝胶成像仪中进行观察结果ꎬ将符合标准的产物送往生工生物工程(上海)股份有限公司进行纯化及测序ꎮ1.3数据分析利用MEGA7.0软件(Sudhiretal.ꎬ2016)对序列进行多序列的比对分析ꎬ校正错误碱基并对序列进行对位排序ꎬ并进行人工校正且除去两端序列引物区域ꎮ利用PhyloSuite软件(Zhangetal.ꎬ2020)对所得序列进行拼接ꎮ运用DnaSP6.0软件统计单倍型数量(Librado&Rozasꎬ2009)ꎬ计算各居群单倍型多样性(Hd)㊁核苷酸多样性(π)㊁Tajima sD值㊁FuandLi sF∗值㊁FuandLi sD值以及Fu sFs值ꎬ并进行失配分析以检测是否经历扩张事件(Tajimaꎬ1989ꎻFuꎬ1997)ꎮ利用PERMUTCpSSR2.0计算总遗传多样性(HT)㊁居群内平均遗传多样性(HS)及遗传分化系数(GST和NST)ꎬ并利用Network构建单倍型网络图(Bandelt867广㊀西㊀植㊀物44卷表1㊀半夏居群地理分布信息Table1㊀GeographicaldistributioninformationofpopulationsofPinelliaternata居群编号Populationcode个体数Numberofindividuals采集地Location经度Longitude纬度Latitude凭证标本号VoucherspecimenNo.C114辽宁丹东DandongꎬLiaoning124ʎ21ᶄ15ᵡE40ʎ00ᶄ47ᵡNWubx1C215湖北宜昌YichangꎬHubei111ʎ56ᶄ29ᵡE30ʎ55ᶄ14ᵡNWubx2C316贵州黔东南QiandongnanꎬGuizhou108ʎ44ᶄ51ᵡE26ʎ23ᶄ50ᵡNWubx3C411甘肃天水TianshuiꎬGansu103ʎ52ᶄ15ᵡE36ʎ04ᶄ40ᵡNWubx4C56云南昭通ZhaotongꎬYunnan104ʎ19ᶄ12ᵡE27ʎ49ᶄ56ᵡNWubx5C615云南昭通ZhaotongꎬYunnan103ʎ22ᶄ28ᵡE26ʎ34ᶄ25ᵡNWubx6C715安徽池州ChizhouꎬAnhui116ʎ49ᶄ45ᵡE29ʎ41ᶄ22ᵡNWubx7C815江苏南通NantongꎬJiangsu120ʎ36ᶄ20ᵡE32ʎ10ᶄ24ᵡNWubx8C94贵州开阳KaiyangꎬGuizhou107ʎ25ᶄ32ᵡE27ʎ17ᶄ58ᵡNWubx9C1015山东青岛QingdaoꎬShandong119ʎ58ᶄ57ᵡE36ʎ05ᶄ12ᵡNWubx10C1114湖南邵阳ShaoyangꎬHunan110ʎ56ᶄ46ᵡE27ʎ21ᶄ58ᵡNWubx11C127湖南常德ChangdeꎬHunan111ʎ26ᶄ09ᵡE29ʎ19ᶄ22ᵡNWubx12C1315浙江义乌YiwuꎬZhejiang120ʎ07ᶄ24ᵡE29ʎ13ᶄ18ᵡNWubx13C144贵州贵阳GuiyangꎬGuizhou106ʎ25ᶄ04ᵡE26ʎ22ᶄ57ᵡNWubx14C154陕西商洛ShangluoꎬShaanxi109ʎ54ᶄ36ᵡE33ʎ53ᶄ59ᵡNWubx15C163河北石家庄ShijiazhuangꎬHebei114ʎ34ᶄ19ᵡE38ʎ00ᶄ44ᵡNWubx16C1715四川绵阳MianyangꎬSichuan104ʎ51ᶄ13ᵡE31ʎ22ᶄ53ᵡNWubx17C1813宁夏固原GuyuanꎬNingxia111ʎ47ᶄ11ᵡE32ʎ55ᶄ48ᵡNWubx18C1911江西上饶ShangraoꎬJiangxi119ʎ58ᶄ57ᵡE36ʎ05ᶄ12ᵡNWubx19表2㊀PCR引物信息及反应程序Table2㊀PrimerandprotocolsofPCR引物名称Sequencename引物序列(5ᶄ ̄3ᶄ)Primersequence(5ᶄ ̄3ᶄ)PCR反应程序PCRprotocolpsbK ̄psbIpsbKF:TTAGCCTTTGTTTGGCAAGpsbIR:AGAGTTTGAGAGTAAGCAT94ħꎬ5minꎻ40ˑ(94ħꎬ30sꎻ57.2ħꎬ30sꎻ72ħꎬ40s)ꎻ72ħꎬ5minatpF ̄atpHatpFF:ACTCGCACACACTCCCTTTCCatpHR:GCTTTTATGGAAGCTTTAACAAT94ħꎬ5minꎻ35ˑ(94ħꎬ30sꎻ50ħꎬ30sꎻ72ħꎬ40s)ꎻ72ħꎬ5mintrnL ̄FtrnL:CGAAATCGGTAGACGCTACGtrnF:ATTTGAACTGGTGACACGAG94ħꎬ5minꎻ35ˑ(94ħꎬ30sꎻ54ħꎬ30sꎻ72ħꎬ1min)ꎻ72ħꎬ10minetal.ꎬ1999)ꎮ利用Arlequin软件包(Exoffier&Lischerꎬ2010ꎻ路雪林ꎬ2018)中的分子方差分析(AMOVA)分析群体遗传结构以及检测居群内和居群间遗传变异ꎬ并计算遗传分化系数(FST)和基因流(Nm)ꎬ进一步揭示居群的分化程度ꎮ用ArcGIS10.2软件绘制居群单倍型地理分布图ꎮ2㊀结果与分析2.1序列变异和单倍型多样性psbK ̄psbI+atpF ̄atpH+trnL ̄F联合序列成功测序212个个体ꎬ比对校正后的序列长度为19479674期吴敏等:中国野生半夏的遗传多样性和遗传结构研究表3㊀半夏居群内单倍型分布及多态性信息Table3㊀HaplotypedistributionandpolymorphismofhaplotypesinpopulationofPinelliaternata居群编号PopulationcodecpDNAnπ(ˑ10 ̄3)HdH单倍型(样本数)Haplotypes(numberofsamples)C114001H7(14)C215001H10(15)C316001H10(16)C411001H10(11)C56001H11(6)C6150.550.53332H10(7)ꎬH11(8)C7150.380.51403H10(4)ꎬH11(1)ꎬH12(10)C815001H13(15)C94001H14(4)C1015001H8(15)C1114001H1(14)C127001H1(7)C13150.490.47602H2(5)ꎬH3(10)C1440.510.50002H4(3)ꎬH5(1)C154001H2(4)C163001H2(3)C17150.270.53332H2(7)ꎬH6(8)C1813001H9(13)C1911001H2(11)㊀注:n.测序个体数ꎻπ.核苷酸多样性ꎻHd.单倍型多样性指数ꎻH.单倍型种类数目ꎮ㊀Note:n.Numberofsequenceindividualsꎻπ.NucleotidediversityꎻHd.DiversityofhaplotypesꎻH.Numberofhaplotypespecies.bpꎬ包含11个变异位点ꎬG+C含量为31.80%ꎮ将这3个序列片段提交至GeneBankꎬ序列号分别为OL310546-OL310559㊁OL310532-OL310545和OL310560-OL310573ꎮ如表3所示ꎬ只有西南地区的云南昭通(C6)㊁贵州贵阳(C14)㊁四川绵阳(C17)和华东部的安徽池州(C7)以及浙江义乌(C13)具有多个单倍型ꎬ其余居群都只有1种单倍型ꎮ运用DnaSP软件得出半夏总单倍型多样性Hd为0.8820ꎬ总核苷酸多样性π为1.23ˑ10 ̄3ꎻ其中ꎬ云南昭通(C6)和四川绵阳(C17)的单倍型多样性最高(0.5333)ꎬ其次是安徽池州(C7)以及浙江义乌(C13)ꎮ总体来看ꎬ西南地区和华东地区的居群遗传多样性水平较高ꎮ2.2单倍型分布用DnaSP软件对psbK ̄psbI+atpF ̄atpH+trnL ̄F联合序列进行了单倍型检测(不考虑插入和缺失)ꎬ共得14个单倍型(H1-H14)(图1)ꎮ其中ꎬ单倍型H10的个体数为53个ꎬ出现的频率最高ꎬH2和H10分布范围最广ꎬ单倍型H10在湖北宜昌(C2)㊁贵州黔东南(C3)㊁甘肃天水(C4)㊁云南昭通(C6)及安徽池州(C7)均有分布ꎻ单倍型H2分布在浙江义乌(C13)㊁陕西商洛(C15)㊁河北石家庄(C16)㊁四川绵阳(C17)和江西上饶(C19)ꎮ除单倍型H1㊁H2㊁H10和H11存在多个居群中以外ꎬ其余单倍型均为特有单倍型ꎮ单倍型H3只分布在浙江义乌(C13)ꎻ单倍型H4和H5只分布在贵州贵阳(C14)ꎻ单倍型H6只分布在四川绵阳(C17)ꎻ辽宁丹东(C1)仅存在特有单倍型H7ꎻ山东青岛(C10)仅有特有单倍型H8ꎻ单倍型H9仅存在宁夏固原(C18)ꎻ单倍型H12仅分布在安徽池州(C7)ꎻ江苏南通(C8)仅存在特有单倍型H13ꎻ贵州开阳(C9)只有特有单倍型H14ꎮ而安徽池州(C7)拥有最多的单倍型种类(H10㊁H11和H12)ꎮ根据半夏叶绿体基因的单倍型网络分析结果显示ꎬ以H10为中心分别衍生出其他单倍型ꎬ推测H10为祖先单倍型(图2)ꎮ2.3失配分析与中性检验使用DnaSP软件对其序列进行中性检验和失配分析ꎬ中性检验不显著ꎮ中性检验得出较为保守的Tajima sD统计值为0.67883(P>0.10)ꎻFuandLi sD值为1.38007(0.05<P<0.10)ꎻFuandLi sF∗值为1.34421(P>0.10)ꎻFu sFs值为-1.304ꎬTajima sD为正值ꎬ表明半夏群体可能经历瓶颈效应ꎮ失配分布图呈现为双峰ꎬ期望值与观测值相悖ꎬ违背种群扩张模型ꎬ中性检验结果也没有检测到种群扩张信号ꎬ这说明群体历史平稳ꎬ整体没有经历过扩张事件(图3)ꎮ2.4遗传多样性与群体遗传结构利用PERMUT进行遗传多样性分析ꎬ其总遗传多样性HT为0.882ꎬ种群内平均遗传多样性HS077广㊀西㊀植㊀物44卷饼状图表示每个居群中单倍型的频率ꎮThepiechartshowsthefrequencyofhaplotypesineachpopulation.图1㊀半夏19个居群单倍型地理分布图Fig.1㊀Haplotypegeographicaldistributionofhaplotypesof19populationsofPinelliaternata图2㊀半夏种群单倍型网络图Fig.2㊀HaplotypenetworkofPinelliaternatapopulations为0.134ꎻ种群间遗传分化系数GST为0.855㊁NST为0.913ꎮ种群间遗传分化系数均为NST>GST(0.01<虚线代表预期分布ꎬ实线代表实际分布ꎮDottedlinerepresentsthedistributionsexpectedforexpandingandsolidlinerepresentstheobservedmismatchdistribution.图3㊀半夏种群错配分析Fig.3㊀MismatchanalysisofPinelliaternatapopulationsP<0.05)ꎬ表明在群体水平上ꎬ在同一群体中出现亲缘关系相近的不同单倍型ꎬ叶绿体单倍型具有1774期吴敏等:中国野生半夏的遗传多样性和遗传结构研究表4㊀半夏种群的分子方差分析Table4㊀Analysisofmolecularvariance(AMOVA)inPinelliaternatapopulations变异来源Sourceofvariation自由度df总方差Sumofsquares变异成分Variancecomponents变异百分比Percentageofvariation(%)遗传分化系数Coefficientofgeneticdifferentiation(FST)居群间Amongpopulations18226.7721.1221489.27∗0.909居群内Withinpopulations19426.1670.1348810.73∗总体Total212252.9391.25702㊀注:∗表示显著性差异(P<0.05)ꎮ㊀Note:∗indicatesignificantdifferences(P<0.05).明显的分子谱系地理结构ꎮ分子方差分析(AMOVA)结果显示遗传变异主要发生在居群间ꎬ在19个半夏居群中ꎬ居群间遗传变异占总变异的89.27%ꎬ而仅有10.73%的变异来自居群内(表4)ꎮ遗传分化系数FST为0.909(P<0.001)ꎬ1000次重复的显著性检验ꎬFST>0.25ꎬ说明半夏群体之间的遗传分化程度已经达到显著水平ꎬ存在明显的隔离ꎮ假设该片段变异处于漂变-迁移平衡ꎬ则基于FST值估算出物种水平上居群间的平均基因流值(Nm)为0.02ꎬ说明半夏居群间基因流较小ꎮ3㊀讨论与结论3.1遗传多样性居群遗传多样性与其生存环境紧密相关ꎮ居群或物种生存环境的许多因素直接或间接地影响遗传多样性(蒋福娟ꎬ2017ꎻ洪霞等ꎬ2021)ꎮ一般而言ꎬ广布种比狭域种具有更高的遗传多样性(Hamrickꎬ1992)ꎮ半夏分布范围广ꎬ不同地理环境下存在不同的遗传和形态变异ꎮ我们在调查采样时ꎬ也观察到不同生态型的半夏ꎬ在植株大小㊁叶形和佛焰苞都有不同表现ꎬ这与我们的实验数据所推测的结果吻合ꎮ通过对半夏19个天然群体212个个体的3个cpDNA片段进行分析ꎬ结果揭示半夏总单倍型多样性为Hd=0.8820ꎬ核苷酸多样性为π=1.23ˑ10 ̄3ꎬ高于Petit等(2005)报道的170个物种叶绿体遗传变异的平均值0.67ꎮ张景等(2021)采用SSR分子标记研究17个半夏居群发现其Nei基因多样性指数(h)平均为1.03ꎬ潘凤等(2021)采用ITS序列分析20个半夏居群得到单倍型多样性Hd为0.8596ꎮ本研究结果与上述研究结果一致ꎬ均表明半夏在物种水平上具较高的遗传多样性ꎮ产生这一结果ꎬ一方面是自然环境的选择作用ꎮ研究发现物种的分布范围与遗传多样性紧密相关ꎬ不同分布区或生境地形地貌复杂㊁气候变化多样ꎬ生境的差别导致存在一定的地理隔离ꎬ使居群在形态㊁生理㊁遗传㊁生态习性等方面出现较大的分化(王爱华等ꎬ2011ꎻ潘凤等ꎬ2021)ꎮ另一方面可能是基因突变ꎬ研究发现半夏兼具有性和无性两种繁育系统ꎬ无性生殖占优势ꎬ其原因是半夏雄配子高度不育(王祖秀等ꎬ2000)ꎮ而在有性生殖较为困难的植物中基因突变是导致物种遗传变异的主要因素(王爱华等ꎬ2011)ꎬ这一特点可能使其居群间具有较高的遗传多样性ꎮ同时ꎬ半夏具有复杂的基因库ꎬ在漫长的历史进化过程和世代交替中可能积累了丰富的叶绿体遗传变异ꎬ这为基因突变提供了丰富的物质基础ꎮ3.2地理分布格局和种群历史动态植物种群的地理分布格局模式和遗传结构受到地质历史气候变化㊁生境异质性以及种子基因交流程度等诸多因素的影响(刘家奇等ꎬ2021)ꎮ本研究发现半夏具有明显的谱系地理结构(NST=0.913>GST=0.855ꎬ0.01<P<0.05)ꎬ居群间存在显著的遗传分化(FST=0.909ꎬP<0.001)ꎬ说明居群间基因交流不频繁ꎬ存在显著的地理隔离或环境异质性ꎮ基于FST值估算出物种水平上居群间的平均基因流Nm=0.02ꎬ当Nm<1时ꎬ基因流不足以抵制居群内因遗传漂变而引起的居群分化(Slatkinꎬ1985)ꎮ因此ꎬ我们推断地理隔离(或者环境异质性)和遗传漂变是导致半夏居群间遗传分化的主要因素ꎮ半夏居群间基因流低ꎬ主要有以下两个277广㊀西㊀植㊀物44卷原因ꎮ(1)与半夏的繁育方式(种子繁殖㊁块茎繁殖和珠芽繁殖)有关(张晓芸等ꎬ2019)ꎮ其中ꎬ珠芽对半夏的繁殖非常重要ꎬ珠芽的大小和数量与半夏块茎的产量密切相关(Zhangetal.ꎬ2013)ꎮ半夏的珠芽㊁子块茎和果实均在母株附近ꎬ缺乏有效的传播方式ꎬ是开拓性较差的物种(顾得兴和郭巧生ꎬ1990)ꎬ导致居群间基因流低ꎬ居群间的遗传分化较高ꎮ(2)由于地理隔离和环境因素的影响使得半夏生境出现片段化ꎬ致使不同居群或不同区域半夏群体间基因交流被隔断或减弱ꎮ中性检验结果Tajima sD和Fu sFs均不显著ꎬ表明半夏种群符合中性进化模型ꎮ失配分布图呈现为双峰ꎬ期望值与观测值相悖ꎬ表明半夏没有经历近期大范围扩张ꎬ这与潘凤等(2021)认为半夏发生过扩张的结果相反ꎬ这可能与叶绿体DNA的遗传方式有关ꎬ叶绿体DNA遗传属于母系遗传ꎬ基因流大部分是通过种子来传播(Hu&Liꎬ2002)ꎬ而核基因为双亲遗传ꎬ种子流和花粉流对居群间基因流均有贡献ꎮ相比花粉传播ꎬ种子传播有一定限制ꎬ导致结果不一致ꎮ单倍型地理分布显示ꎬ存在多个相对分离的分布区ꎬ各分布区都存在各自特有单倍型和主要单倍型ꎮ研究表明ꎬ一般位于网络结构图的中间位置㊁频率较高㊁地理分布广泛的单倍型更为古老(Freelandetal.ꎬ2012ꎻ莫忠妹等ꎬ2022)ꎮ本研究中单倍型H10频率较高㊁地理分布广泛ꎬ位于网络中心位置ꎬ推测其为最古老的单倍型ꎮ其中ꎬ安徽池州(C7)不仅拥有较高的单倍型多样性ꎬ还拥有最多的单倍型种类ꎬ这与半夏起源于华东地区ꎬ沿长江向周围扩散的结论相吻合(李恒ꎬ1996)ꎮ19个居群中有14个居群都只有一种单倍型ꎬ这可能与群体内变异程度以及生殖方式有极大关系ꎬ对于主要以营养生殖的半夏来说ꎬ很可能在一个群体内只有一个基因型ꎬ即只有一个无性系ꎮ冰期避难所通常指冰期尤其是历史进程中的末次盛冰期ꎬ气候和地势条件产生巨大变动ꎬ导致生物物种进行地理位置上大规模的转移后为植物提供生存领域(Hafferꎬ1969ꎻ赵晶晶等ꎬ2021)ꎮ从冰川避难所理论得知ꎬ遗传多样性高㊁古老单倍型和地方性单倍型较多的地区可能是该物种在冰期时的潜在避难所(Favreetal.ꎬ2010)ꎮ由于西南地区的云南昭通(C6)㊁贵州贵阳(C14)㊁四川绵阳(C17)以及华东地区的安徽池州(C7)㊁浙江义乌(C13)均具有较高的遗传多样性ꎬ并含有古老的单倍型和特有单倍型ꎮ因此ꎬ半夏在第四纪冰期时存在至少两个以上的冰期避难所ꎮ我们推测半夏在第四纪冰期时可能在中东部和西南地区存在其冰期避难所ꎬ与本课题组对薤白的研究结果一致(莫忠妹等ꎬ2019ꎻ石甜等ꎬ2021)ꎬ这可能与它们的分布和习性相似有关ꎮ本研究开展半夏遗传多样性ꎬ谱系地理模式和种群历史动态分析ꎬ采样覆盖了贵州㊁浙江㊁甘肃和四川等15个省区ꎬ横跨中国南北方ꎬ并呈现华东㊁华中㊁西北㊁西南等不同方位和区域自然地理分布ꎬ呈现了该植物在中国的整个地理分布情况ꎮ尽管我们没有收集到日本和朝韩半岛的材料ꎬ但我们的研究结论与本课题组曾对与半夏地理分布相似的薤白㊁大叶柴胡草本植物谱系地理和遗传多样性的研究和前人对分布横夸中国西南地区和中东部银杏(Ginkgobiloba)(Weietal.ꎬ2008ꎻ樊晓霞ꎬ2014)㊁枫香树(Liquidambarformosana)(孙荣喜ꎬ2017)谱系地理模式的研究结论有相似性和一致性ꎬ因此ꎬ我们的实验样品采样点的选择是可行的ꎬ得出的结论是可靠的ꎮ3.3药用植物半夏的优良种源分子筛选与保护策略半夏块茎入药ꎬ有燥湿化痰㊁降逆止呕㊁清痞散结之功能ꎬ是最常用的中药材之一ꎬ在临床上多用于抗肿瘤㊁抗生育㊁抗痉挛㊁调血脂㊁祛痰㊁消肿和治疗冠心病(杜娟ꎬ2006ꎻ王爱华等ꎬ2012)ꎮ近年来ꎬ随着农田化肥㊁化学农药(如除草剂)的过度使用和资源的无序采挖ꎬ农田㊁荒山荒地不断被开垦利用ꎬ半夏野生生长环境遭到破坏ꎬ致使半夏野生资源日趋枯竭ꎮ另外ꎬ国内外对半夏的需求量与日俱增ꎬ尤其是全球升温ꎬ肺疾增多ꎬ商品市场需求量日益增加ꎮ因此ꎬ选育优良种源或培育新品种成为半夏生产发展的紧迫现实需求ꎮ此外ꎬ由于连作障碍导致病虫害严重㊁产品质量降低㊁产量下降㊁价格升高㊁新品种数量少(或无)等问题(安艳等ꎬ2018)ꎮ目前生产中ꎬ发现极少数块茎为红色的半夏品种具有高产㊁高抗性等特点ꎬ但是却没有一套完整的选育方法(张景等ꎬ2021)ꎮ研究表明半夏惧怕阳光直射ꎬ适度遮阴可促进其生长发育ꎬ而过度遮阴则会使植株长势差ꎬ甚至死亡(常琴ꎬ2022)ꎮ在高温(ȡ32ħ)或温度过低的冬季均会引起半夏倒苗或休眠ꎬ进而影响半夏产量3774期吴敏等:中国野生半夏的遗传多样性和遗传结构研究(张明等ꎬ2004)ꎮ因此选育方向为耐强光高温㊁耐低温阴湿㊁耐连茬等ꎬ旨在提取半夏资源中的优异新种质ꎮ选育样本通过生物学特性对比㊁抗性鉴定㊁产量分析㊁繁殖特性研究等不同指标筛选出了优良株系ꎬ再通过资源圃基础材料的田间表现ꎬ以生长势㊁单株叶片数㊁抗病性㊁珠芽着生位置及能力㊁块茎形状及生产力等几个主要指标为目标值的筛选ꎬ选出优良目标株系ꎮ常琴(2022)将半夏产量性状与细胞学特征关联分析ꎬ发现基因组小的半夏种质似乎更利于株芽形成及地下块茎增殖ꎮ本研究从分子水平上发现云南昭通(C6)㊁贵州贵阳(C14)㊁四川绵阳(C17)以及华东地区的安徽池州(C7)㊁浙江义乌(C13)均具有较高的遗传多样性ꎬ属于野生优良种源分布地ꎮ除了对半夏选育优良品种或培育新品种外ꎬ还应对其野生资源采取保护措施ꎬ对上述遗传多样性较高的地区采取就地保护或迁地保护ꎮ半夏由于分布广ꎬ野生数量少ꎬ应该主要采用迁地保护为主ꎬ如建立药用植物半夏种质资源圃(库)等ꎬ重点保护具有较高遗传多样性的居群ꎮ对拥有独特的单倍型的居群ꎬ如辽宁丹东(C1)㊁安徽池州(C7)㊁江苏南通(C8)㊁贵州开阳(C9)㊁山东青岛(C10)㊁浙江义乌(C13)㊁贵州贵阳(C14)㊁四川绵阳(C17)㊁宁夏固原(C18)等也应给予优先保护ꎮ同时ꎬ在制定迁地保护策略时ꎬ拥有特殊性状的个体也应当尽量纳入保护范围ꎮ参考文献:ANYꎬYANGDꎬLIXꎬetal.ꎬ2018.StudyontheobstacleeffectandphysiologicalmechanismofPinelliaternatecontinuouscropping[J].JNWAgricꎬ27(7):1017-1022.[安艳ꎬ杨丹ꎬ李鑫ꎬ等ꎬ2018.半夏连作障碍效应及生理机制研究[J].西北农业学报ꎬ27(7):1017-1022.]AVISEJCꎬARNOLDJꎬBALLRMꎬetal.ꎬ1987.IntraspecificphylogeographyꎻthemitochondrialDNAbridgebetweenpopulationgeneticsandsystematics[J].AnnRevEcolSystꎬ18(1):489-522.AVISEJCꎬ1998.Thehistoryandpurviewofphylogeography:apersonalreflection[J].MolEcolꎬ7(4):371-379.AVISEJCꎬ2000.Phylogeography:Thehistoryandformationofspecies[M].CambridgeꎬMassachusettsꎬLondonꎬEngland:HarvardUniversityPress:1-464.BANDELTHJꎬFORSETERPꎬROHLAꎬ1999.Median ̄joiningnetworksforinferringintraspecificphylogenies[J].MolBiolEvolꎬ16(1):37-48.BENNETTKDꎬPROVANJꎬ2008.Whatdowemeanby refugia [J].QuatSciRevꎬ27:2449-2455.CHANGQꎬ2022.GeneticdiversityanalysisandexcellentgermplasmselectionofPinelliaternate[D].Guiyang:GuizhouUniversity.[常琴ꎬ2022.半夏遗传多样性分析与优异种质筛选[D].贵阳:贵州大学.]CHENLMꎬHEZGꎬHANRLꎬ2020.AdvancesingermplasmresourcesofPinelliaternata[J].HeilongjiangAgricSciꎬ(2):131-135.[陈黎明ꎬ何志贵ꎬ韩蕊莲ꎬ2020.半夏种质资源研究进展[J].黑龙江农业科学ꎬ(2):131-135.]DUJꎬ2006.ComparativestudyoncomprehensivecharactersofPinelliaternataindifferentpopulations[D].Beijing:CapitalNormalUniversity:1-79.[杜娟ꎬ2006.不同居群半夏综合性状的比较研究[D].北京:首都师范大学:1-79.]EXOFFIERLꎬLISCHERHELꎬ2010.Arlequinsuitever3.5:anewseriesofprogramstoperformpopulationgeneticsanalysesunderLinuxandWindows[J].MolEcolNotesꎬ10(3):564-567.FANXXꎬ2014.TheresearchonthevarietyofgeneticstructureofGinkgobilobabasedoncpDNAhypolytypes[J].NatSciJHarbinNormUnivꎬ30(4):104-107.[樊晓霞ꎬ2014.基于叶绿体DNA单倍型的银杏遗传多样性格局研究[J].哈尔滨师范大学自然科学学报ꎬ30(4):104-107.]FAVREAꎬYUANYMꎬKUPFERPꎬetal.ꎬ2010.PhylogenyofsubtribeGentianinae(Gentianaceae):biogeographicinferencesdespitelimitationsintemporalcalibrationpoints[J].Taxonꎬ59(6):1701-1711.FREELANDJRꎬKIRKHꎬPETERSENSDꎬ2012.Phylogeography[M].2nded.NewJersey:JohnWiley&SonsInc.ꎬWiley ̄Blackwell:225-269.FUYXꎬ1997.Statisticaltestsofneutralityofmutationsagainstpopulationgrowthꎬhitchhikingandbackgroundselection[J].Geneticsꎬ147(2):915-925.GUDXꎬGUOQSꎬ1990.AstudyonthebiologicalcharacteristicsofPinelliaternatapopulations[J].JNanjingAgricUnivꎬ13(2):11-16.[顾得兴ꎬ郭巧生ꎬ1990.半夏群体生物学特性的研究[J].南京农业大学学报ꎬ13(2):11-16.]HAFFERJꎬ1969.Speciationinamazonianforestbirds[J].Scienceꎬ165(3889):131-137.HARRISONSꎬYUGꎬTAKAHARAHꎬetal.ꎬ2001.Palaeovegetation.DiversityoftemperateplantsineastAsia[J].Natureꎬ413(6852):129-130.HAMRICKJLꎬ1992.Factorsinfluencinglevelsofgeneticdiversityinwoodyplantspecies[J].NewForꎬ6(1/2/3/4):95-124.HEWITTGMꎬ1996.Somegeneticconsequencesoficeagesꎬandtheirroleindivergenceandspeciation[J].BiolJLinnSocꎬ58(3):247-276.477广㊀西㊀植㊀物44卷。
半夏内生真菌的多样性及其药化研究

贵州大学2009届硕士研究生学位论文半夏内生真菌的多样性及其药化研究学科专业:微生物学研究方向:真菌分子系统学导师:康冀川研究生:苏昊中国﹒贵州﹒贵阳2009年6月2009Master Thesis of Guizhou UniversityStudies on the diversity and pharmaceutical chemistry of endophytic fungi in Pinellia ternataSpeciality:MicrobiologyField:Molecular Systemics of FungiSupervisor:Kang JichuanGraduate:Su HaoGuiyang Guizhou ChinaJune2009摘要 (1)1Abstract Abstract (3)3第一章前言 (5)51植物内生真菌研究进展 (5)1.1植物内生真菌的概念 (5)1.2药用植物内生真菌研究进展 (5)1.2.3存在的问题 (7)1.2.4展望 (8)2半夏研究进展 (8)2.1半夏简介 (8)2.2半夏生态及在我国的分布 (9)2.3半夏化学成分和药理作用研究 (9)3植物内生真菌的分类鉴定 (14)3.1传统的分类鉴定方法 (14)3.2分子生物学鉴定方法 (14)4课题研究内容 (17)5本研究的目的和意义 (17)第二章半夏内生真菌的分离 (18)181材料与方法 (18)181.2仪器和试剂 (18)1.2.1用具 (18)1.2.2试剂 (19)1.2.3主要设备 (19)1.3分离培养基 (19)1.3.1马铃薯葡萄糖琼脂培养基 (19)1.3.2抗生素用法与用量 (19)1.4内生真菌分离纯化方法 (19)1.4.1样品表面消毒及预处理 (19)1.4.2内生真菌的分离与纯化 (20)1.5保存方法 (20)2结果与分析 (21)212.1消毒剂及消毒时间的确定 (21)2.2半夏内生真菌的分离结果 (22)3小结与讨论 (23)233.1内生真菌的分离 (23)3.2内生真菌分离方法的有效性 (23)第三章半夏内生真菌的分类鉴定 (23)23第一节半夏内生真菌的形态学鉴定 (24)241材料和方法 (24)241.1菌株及培养基 (24)1.2试剂及仪器 (24)1.3内生真菌的形态学鉴定方法 (24)1.3.1培养性状的观察要点 (24)1.3.3标本的制作 (25)1.3.4显微摄影照片 (25)1.3.5菌种的鉴定 (25) (2525)2结果与分析.................................................................................................................................2.1部分内生真菌形态描述 (25)27 (27)第二节半夏内生真菌的分子生物学鉴定................................................................................... (2727)1材料与方法.................................................................................................................................1.1实验菌株及培养 (27)1.2仪器和试剂 (27)1.2.1所用器具 (27)1.2.2主要设备 (28)1.2.3试剂 (28)1.3基因组DNA的提取及纯化 (28)1.3.1试剂配制 (28)1.3.2基因组DNA的提取 (29)1.4DNA纯度的检测 (30)1.4.1紫外光吸收检测 (30)1.4.2琼脂糖凝胶电泳检测 (30)1.5多聚酶链式反应(PCR扩增) (30)1.5.1本实验所用引物 (30)1.5.2扩增条件 (31)1.5.3扩增产物检测 (31)1.6扩增产物的纯化及序列的测定 (31)1.7内生真菌的分子鉴定及系统发生关系分析 (31)2结果与分析................................................................................................................................. (3232)2.1ITS区段的PCR扩增 (32)2.2PCR扩增产物的序列测定 (32)2.3在GenBank上的搜索(Blast)结果 (33)2.4半夏内生真菌的分子生物学鉴定及系统发生关系分析 (34)2.4.1基于ITS序列的分析 (34)2.4.2通过rDNA ITS序列分析所揭示的半夏内生真菌多样性 (34)2.4.3半夏内生真菌的分布 (37)40第三节小结与讨论....................................................................................................................... (40) (4040)1半夏内生真菌的多样性及分布.................................................................................................2分子生物学技术与内生真菌分类............................................................................................. (4141)42第四章半夏内生真菌的产盐酸麻黄碱的初步研究................................................................... (42) (4242)1材料和方法.................................................................................................................................1.1仪器、药材和试剂 (42)1.1.1仪器及用品 (42)1.1.2试剂 (42)1.1.4试验菌株 (42)1.2方法 (43)1.2.2标准品的配制 (43)1.2.3待测样品的制备 (43)1.2.4TLC法检测 (43)1.2.5HPLC色谱条件 (43)1.2.6HPLC法计算盐酸麻黄碱的含量 (44) (4444)2结果与分析.................................................................................................................................2.1TLC法检测盐酸麻黄碱 (44)2.2标准品及样品的HPLC检测图谱与分析 (44) (4747)3小结与讨论.................................................................................................................................48 (48)致谢.........................................................................................................................................49参考文献......................................................................................................................................... (49)57 (57)附录.........................................................................................................................................60 (60)图版.........................................................................................................................................63原创性声明............................................................................................................................. (63)摘要植物内生真菌是指在某一时期生活在健康植物体内但对寄主植物组织并不引起明显侵染的真菌。
种质资源鉴定的一种分子遗传技术标准:随机扩增多态性DNA(RAPD)

种质资源鉴定的一种分子遗传技术标准:随机扩增多态性
DNA(RAPD)
Lowe.,AJ;刘旭
【期刊名称】《生物技术通报》
【年(卷),期】1997(000)005
【摘要】RAPD(随机扩增多态性DNA)变异越来越多地被用于种质样品的鉴定。
由于在某些条件下,RAPD结果重性较差,限制这一技术的应用。
本文综合了一些文献中引起片段变异的主要原因,分析了减少这些不需要变异的方法。
由此提出了一个含有全部描述符的项目表,它包括RAPD必不可少的条件和再现植物材料RAPD结构的必须核对的程序。
此标准程序和描述形式有促进类似材料在不同实验室所做结果进行比较,而且允许RAPD结果作
【总页数】5页(P37-41)
【作者】Lowe.,AJ;刘旭
【作者单位】不详;不详
【正文语种】中文
【中图分类】S324
【相关文献】
1.草地早熟禾种质资源遗传多样性的RAPD鉴定与分类研究 [J], 高丽娜;陈雅君;高丽娟;申凤娟
2.利用RAPD标记鉴定草地早熟禾种质资源的遗传多样性 [J], 涂明月;李杰;何亚
丽;李醒;李俊;袁晓君
3.红麻种质资源遗传多样性和分子鉴定技术研究Ⅱ.红麻种质资源的遗传多样性和亲缘关系的AFLP分析 [J], 程舟;杨晓伶;卢宝荣;鲛岛一彦;陈家宽
4.垂枝杉种质资源的RAPD鉴定及遗传多样性研究 [J], 宋丛文;许业洲;胡兴宜;管兰华
5.黄瓜种质资源遗传多样性的RAPD鉴定与分类研究 [J], 李锡香;朱德蔚;杜永臣;张广平;沈镝
因版权原因,仅展示原文概要,查看原文内容请购买。
半夏研究进展的论文

半夏研究进展的论文半夏研究进展的论文摘要从植株形态变异研究、遗传多样性研究、有效成分研究、驯化栽培等方面阐述了半夏的研究进展,以供参考。
关键词半夏植株形态变异遗传多样性有效成分驯化栽培半夏(pinellia ternata(thunb.)breit.)又名麻芋头、麻芋子、天落星和野芋头等,为天南星科半夏属多年生草本植物,广泛分布于我国长江流域以及东北、华北等地区[1]。
半夏是我国重要的传统中药材,也是重要的出口产品。
随着半夏利用范围的不断扩展和市场价格的攀升,有关研究也较为活跃,同时也存在很多亟需解决的问题。
1半夏植株形态变异研究具有丰富的形态变异是半夏属植物的一个重要特点。
生长于不同环境或同一生态环境的不同居群甚至同一居群的不同个体间,各种器官(包括根、球茎、珠芽、叶片、叶柄等)在形态上都存在着丰富的变异类型。
其中以叶、球茎和珠芽的变异更明显。
对叶和球茎的变异,较早的文献中已有较多报道。
近年来,半夏珠芽变异引起研究者的重视。
半夏珠芽变异主要表现在珠芽数量、珠芽着生位置和芽眼数量等方面。
据赵忠堂等[2]研究,不同类型半夏形成的珠芽数量差异较大,以线形叶型半夏的珠芽数量最多,芍药叶型半夏的珠芽数量最少。
半夏珠芽的多少与种茎大小呈正相关,即同一类型的半夏,种茎越大,形成的珠芽数越多,珠芽在产量中所占比例也越大。
还有一类无论是叶柄还是叶端均不结珠芽的“新居群”,经移栽试验和原分布地考证,该类半夏遗传性状稳定。
半夏珠芽正常着生位置在叶柄下部内侧,但有的半夏在叶端也可着生1枚珠芽。
a等报道,半夏种内各群居普遍存在着染色体的复合多倍现象,不同群居的染色体数目为2n=28、72、96、104、108、115、116、118、128。
染色体数目的丰富变异决定了半夏的遗传多样性。
开展半夏遗传多样性研究,对于半夏资源的保护和合理开发利用具有十分重要的意义[5],但其研究的难点在于找到有效的分析方法。
近年来,借助同工酶技术进行半夏遗传多样性分析的工作日渐增多。
南充道地药材半夏的遗传多样性分析

南充道地药材半夏的遗传多样性分析张秀云;涂颖;喻萍;王含彦【期刊名称】《西南师范大学学报(自然科学版)》【年(卷),期】2012(037)011【摘要】目的:对南充道地药材半夏的遗传多样性进行研究,为半夏的保护利用提供参考.方法:采用随机扩增多态DNA(RAPD)方法对60份半夏基因组进行扩增,所获数据通过NTSYS-pc v2.1、POPGENE v1.31及Arlequin3.0进行分析.结果:从43条引物中筛选出22条条带清晰且重复性好的引物用于实验和统计分析,共扩增出88个位点,多态性位点占79.5%,表现出较高的遗传多样性.分子方差分析表明各来源地内材料间差异构成了遗传变异的主要来源.聚类分析显示,所有供试半夏可明显聚为三类,其中栽培种分布较集中.组间遗传差异分析表明大部分来源地间差异不显著,组间基因流动较频繁.结论:南充地区的半夏资源具有较高的遗传多样性;材料间遗传距离的远近与其地理来源有一定相关性,野生种的遗传背景较栽培种更为丰富,说明人为的引种、筛选、栽培已对半夏资源的多样性造成了影响.因此,应尽快建立半夏种质资源库,通过形态学、细胞学及分子生物学技术进行系统分类,为保护利用半夏资源提供理论依据和物质基础.%This paper aims to have a profound knowledge of the genetic diversity of Pinellia ternate of Nanchong. By means of 22 random primers selected to study on 60 P. ternate from eight districts, including 49 wild accessions and 11 cultivars. The application result is record with 0 or 1 and the data calculated with NTSYS-pc v2. 1, POPGENE v1. 31 and Arlequin 3. 0. The result shows that the selected 22 random primers used in PCR amplification to detected 88 bands, amongwhich 79. 5 percent is polymorphic. The results of cluster analysis show that all the accessions researched in this study are classified into three categories. And the collection of cultivars has more narrow genetic background than the wild types. Most genetic difference among groups is not significant, which suggests being relative with frequently gene flowing a-mong groups. The conclusion is that the genetic variation is relatively high in S. baicalensis, and the genetic similarity among the accessions is related to the geographic origination. And the artificial choice would reduce the diversity of P. ternate germplasm. Therefore, it is necessary to protect the P. ternata resources by building the germplasm pool.【总页数】7页(P94-100)【作者】张秀云;涂颖;喻萍;王含彦【作者单位】川北医学院生物化学教研室,川北医学院应用技术研究所,四川南充637007;川北医学院临床医学系2008级29班,四川南充637007;川北医学院医学影像系2009级7班,四川南充637007;川北医学院生物化学教研室,川北医学院应用技术研究所,四川南充637007【正文语种】中文【中图分类】R931.2【相关文献】1.道地药材荆半夏的指纹图谱研究 [J], 卢志刚;唐本遂;石银珍;张逸;何秀唐;胡芳2.荆门道地药材荆半夏产业发展现状与高产栽培技术 [J], 刘菊;张元虎;王艾鸿;夏振华;杨雪;郑应莉;贺小芹;李娟娟3.道地药材西和半夏高效栽培技术 [J], 孙连虎;潘水站;王义存;魏莉霞4.南充地区半夏资源调查及与省外半夏形态特征的比较 [J], 白权;李敏;贾敏如;孙琪华5.毛细管气相色谱法测定南阳道地药材唐半夏的有机氯农药残留 [J], 郭万周;贾荷丽;杨媛媛因版权原因,仅展示原文概要,查看原文内容请购买。
随机扩增多态DNA技术在天麻遗传多样性分析中的应用

随机扩增多态DNA技术在天麻遗传多样性分析中的应用王德信【期刊名称】《中国农学通报》【年(卷),期】2010(26)4【摘要】以乌天麻、黄天麻和绿天麻3种变型为研究对象,通过RAPD分析研究天麻遗传多样性,从而为天麻种源的筛选与鉴定及品种选育提供科学依据。
采用改良的CTAB法提取天麻样品DNA,筛选出重复性好的10条随机引物,对130个样品的基因组DNA进行扩增,建立RAPD分析图谱。
扩增结果:54条表现为特异性带,多态性变异率高达65.1%,说明天麻变异程度较大。
聚类分析表明:栽培群体乌天麻是1个高度混杂的群体,虽然个体的表现型基本相同,但从遗传物质的组成来看,基因型不一致。
研究表明天麻具有丰富的遗传多样性,其中乌天麻遗传多样性最丰富,变异复杂。
为鉴定优质天麻品种、天麻种质资源合理有效利用和进行遗传育种,提供了可借鉴的科学依据。
【总页数】6页(P19-24)【关键词】天麻;RAPD分析;遗传多样性【作者】王德信【作者单位】山东菏泽学院资源与环境系【正文语种】中文【中图分类】S326【相关文献】1.随机扩增多态性DNA技术对湘产蛇莓种质资源的遗传多样性分析Δ [J], 刘湘丹;高昱;张亚利;蔡嘉洛;童巧珍;张聪子2.随机扩增多态DNA技术在遗传育种中的应用 [J], 张鹏;解玉红;周东坡3.用随机扩增多态DNA (RAPD)技术分析野生与笼养绿孔雀种群的遗传多样性 [J], 柯亚永; 常弘; 张国萍4.随机引物扩增DNA多态性技术在分析金黄色葡萄球菌和肺炎杆菌多态性中的应用 [J], 高大威;马佳毓;郭晓奎;赵月辉5.利用随机扩增多态性DNA技术分析刺五加的遗传多样性 [J], 邢朝斌;劳凤云;吴鹏;李玉彦;沈海龙因版权原因,仅展示原文概要,查看原文内容请购买。
随机扩增DNA多态性技术在粘质沙雷菌分型研究中的应用

随机扩增DNA多态性技术在粘质沙雷菌分型研究中的应用侯晓娜;张传树;傅炜昕;杨婧
【期刊名称】《中国感染控制杂志》
【年(卷),期】2004(003)003
【摘要】目的应用随机扩增DNA多态性(RAPD)技术对医院感染的粘质沙雷菌进行基因分型.方法对心外科ICU病房5例术后发热患者血液中分离出的粘质沙雷菌,用VITEK细菌全自动分析仪进行系统生化鉴定,K-B纸片扩散法进行药敏试验,用2条不同的随机引物进行RAPD基因分型.结果 5株粘质沙雷菌药敏谱全部一致,2条随机引物中一条引物未能分型,另一条引物可分为2个基因型.其中,4株菌扩增出相同带谱,为同基因型;1株菌有独特的带谱为另一种基因型.结论本次调查的5株粘质沙雷菌有2种RAPD基因型,其中4株为相同基因型,证实存在感染流行.
【总页数】3页(P205-207)
【作者】侯晓娜;张传树;傅炜昕;杨婧
【作者单位】沈阳军区总医院,辽宁,沈阳,110016;大连医科大学检验医学院,辽宁,大连,116012;沈阳军区总医院,辽宁,沈阳,110016;沈阳军区总医院,辽宁,沈阳,110016【正文语种】中文
【中图分类】R372
【相关文献】
1.黏质沙雷菌随机扩增DNA多态性DNA分型 [J], 李华
2.黏质沙雷菌随机扩增多态性DNA法基因分型及意义 [J], 叶明亮;邱方;刘士君
3.肺炎克雷伯菌AmpC酶基因分型和随机扩增DNA多态性研究 [J], 向玉
4.随机扩增DNA多态性技术在肠球菌流行病学分型中的应用 [J], 桂炳东;胡龙华
5.随机引物扩增技术对变形菌DNA多态性分型研究 [J], 马清光;赵月辉
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
关于半夏种质资源的随机扩增多态性DNA 技术分析2012-12-24【编者按】:医药论文是科技论文的一种是用来进行医药科学研究和描述研究成果的论说性文章。
论文网为您提供医药论文范文参考,以及论文写作指导和格式排版要求,解决您在论文写作中的难题。
关于半夏种质资源的随机扩增多态性DNA 技术分析作者:杨俊宝,朱秀志,罗成科,李俊,魏会廷,彭正松【摘要】目的分析半夏种质资源在分子水平上的遗传多样性。
方法应用随机扩增多态性DNA 技术(RAPD)对21 份半夏种质资源进行检测。
结果29 个引物共得到162 条扩增DNA 片段,其中123 条(75.9%)的片段具多态性,揭示了半夏居群间较丰富的遗传多样性。
利用UPGMA 法进行聚类分析表明,所有材料可划分为3 类,根据RAPD 遗传相似系数划分的类群同地理分布有一定关系。
结论半夏种质资源在分子水平上确实存在较大遗传差异。
RAPD 标记可作为构建半夏DNA 指纹图谱的有效工具。
【关键词】半夏;随机扩增多态性DNA 技术;遗传多样性;DNA 指纹图谱Abstract:Objective To discuss the genetic diversity of the germplasm resources ofPinellia ternata on molecular level. Methods The genetic diversity of P. ternata 21 differentpopulations were tested by RAPD markers. Results A total of 162 products were amplified by29 10-mer arbitrary primers, 123 (75.9%) products were found to be polymorphic. The resultsrevealed the abundant genetic diversity in P. ternata 21 different populations. The clusteranalysis using UPGMA method showed that 21 populations were classified as 3 groups, andthe genetic diversity in P. ternata based on RAPD GS was correlated with geographicdistribution. Conclusion There actually existed much genetic diversity on molecular levelamong the germplasm resources of P. ternata. RAPD markers could be effective tools toconstruct DNA fingerprintings of P. ternata.Key words:Pinellia ternata;RAPD;genetic diversity;DNA fingerprintings此间的遗传关系尚不清楚。
因此,有必要运用分子生物学方面的新技术手段加强其种质资源遗传多样性研究,并开展半夏野生种质资源的保护工作,从而为进一步选育优良种质2提供理论指导和依据。
随机扩增多态性DNA 技术(random amplified polymorphism DNA,RAPD)是以PCR 为基础的快速简便的分子标记技术,因其费用不高,DNA 用量少、不需预知研究对象的基因组序列、PCR 引物无种属限制等优点,目前已应用于人参[2]、石斛[3]等药用植物的遗传多样性研究。
本研究选取了21 个野生半夏居群进行了RAPD-PCR 扩增分析,构建遗传聚类树状图对半夏居群间的遗传关系进行了研究,旨在探讨其居群间的遗传关系,为半夏种质资源的保护、合理开发利用和新品种选育提供有价值的资料,也为半夏药材DNA指纹图谱的构建奠定一定的基础。
1 材料与方法1.1 药材采自四川、重庆及陕西等地不同地点,共21 份。
来源详见表1。
1.2 DNA 的提取取每一居群10~15 不同单株新鲜叶片约0.3 g,经消毒和洗净后,放在乳钵中加入600L 氯仿于室温下研磨成匀浆,然后加入1.5 mL SDS 抽提液[100 mmoL/L Tris-HCl(pH 8.0);100 mmol/L EDTA(pH 8.0);0.5 mol/L NaCl;1.5%(W/V) SDS]混匀。
将1.2 mL 匀浆转移至1.5 mL 离心管,室温( 15 ℃)静置10 min。
室温下12 000 r/min 离心10 min,取上层水相于另一干净1.5 mL 离心管,加入等体积氯仿-异戊醇(24∶1)进行抽提,重复抽提2~3 次,直到界面清晰为止。
取上清液加入到500 L 预冷(4 ℃)无水乙醇中沉淀DNA,此时可见絮状DNA 沉淀下来。
去上清液,沉淀经70%乙醇洗涤3 次,空气中干燥后用TE 溶解备用。
用紫外分光光度计检测样品DNA 浓度,通过计算,稀释到终浓度为50 ng/ L 备用。
1.3 RAPD 反应用Sangon 公司生产的S 系列的100 个十聚体随机引物对材料进行扩增筛选,从中共筛选出谱带清晰的引物29 个用于试验和统计分析,反应在BIO-RAD 的MyCyclerTM PCR 仪上进行。
反应总体积25 L,内含1 PCR buffer,1.5 mmol/L MgCl2,1 U Taq 酶,0.25 mol/L 引物;100 ng 模板DNA,dNTPs 各0.2 mmol/L。
扩增前94 ℃预变性3 min,每循环在94 ℃变性1 min,36 ℃退火1 min,72 ℃延伸2 min,共45 个循环,完成最后一个循环后,在72 ℃保温10 min,然后在4 ℃保存。
扩增产物在1.5%琼脂糖凝胶上电泳分离。
溴化乙锭染色,凝胶成像仪上观察照像、记录。
1.4 统计学方法RAPD 扩增产物按条带有无分别赋值,有带记为1,无带记为0。
利用软件POPGENE处理得到遗传相似系数(Genetic Similarities, GS)[4]。
利用GS 按不加权成对群算术平均法(unweighted pair group method with arithmetic average, UPGMA)进行聚类。
统计分析在NTSYS pc[5]软件系统下进行。
表1 供试的半夏来源(略)2 结果与讨论2.1 扩增片段多态性RAPD 扩增结果表明:29 个引物中有28 个(96.6%)引物的扩增产物具多态性。
图1为引物S20 的扩增结果。
29 个引物共得到162 条扩增DNA 片段,平均每个引物可获得5.6 个DNA 片段, 其中123 条具有多态性,占75.9%。
表明半夏种质资源多态性水平较高,充分体现了半夏不同居群间的遗传多样性。
2.2 遗传相似系数半夏21 个不同居群的遗传相似系数值见表2。
从中可见,半夏不同居群间的GS 分布较广,范围为0.592 6~0.895 1,说明其遗传背景较复杂,表明其遗传多样性相对较丰富。
PT0002 居群的半夏与PT0009 居群的半夏相似系数最大,高达0.895 1,说明它们的亲缘关系最近。
另外,PT0002 居群的半夏与PT0054 居群的半夏相似系数最小,为0.592 6,说明它们的亲缘关系较远。
表2 21 个半夏居群遗传相似系数值(略)2.3 供试材料的聚类分析结果tips:感谢大家的阅读,本文由我司收集整编。
仅供参阅!2012-12-24【编者按】:医药论文是科技论文的一种是用来进行医药科学研究和描述研究成果的论说性文章。
论文网为您提供医药论文范文参考,以及论文写作指导和格式排版要求,解决您在论文写作中的难题。
浅谈中药的特点及其药学服务作者:李忠林,李小川,宋文惠,徐智【关键词】中药;药学服务随着医院药学从传统的药品供应模式向以患者为中心的药学技术服务模式的转变,以合理用药为核心的临床药学工作逐步开展起来,药学服务便成为热门话题见诸报刊,然而, 涉及内容基本上都是有关西药药学服务。
为此,笔者就中药的特点及其临床药师服务作一探讨。
1 影响中药疗效的因素较多,决定了中药应用需要更广泛的药学服务影响中药疗效的因素有诸多方面,除了生物和环境因素外,中药的品种、产地、采收、储藏、炮制、配伍、剂型、剂量、煎服等本身因素则是影响其疗效的主要方面。
1.1 保证中药饮片质量,提高临床疗效1.1.1 适时采收因不同药用植物的根、茎、叶、花、果实、种子或全草都有一定的生长期和成熟期, 而其有效成分含量的高低,将随其不同入药部位和植物各部分的不同生长期而异,故采药时间应随着中药的品种和入药部位的不同而有所区别。
如丹参的有效成分丹参酮ⅡA、丹参酮Ⅰ的含量,第4 季度收获的丹参比其他季节收获者高2~3 倍;薄荷在开花盛期采收者,挥发油含量最高;青蒿中所含抗癌成分青蒿素在7-8 月花前盛叶期含量高达0.6%, 开花后含量逐渐下降[1]。
作为中药师可指导药农适时采收,从源头上把握药材质量。
1.1.2 合理贮藏中药贮存、保管条件直接影响其质量。
不同的贮藏时间、温度、湿度和方法对药物所含成分有明显影响。
有人曾做过实验,三棵针中的小檗碱随贮存时间的延长其含量逐渐降低,贮存3 年在见光、避光两种条件下,小檗碱含量分别降低54.1%和39.88%。
又如刺五加贮存时间超过3 年或在高温(40~60 ℃)、高湿(相对湿度74%以上)、日光照射等条件下贮存6 个月,其中所含丁香苷几乎丧失殆尽[1]。
中药颗粒剂﹑散剂易吸潮发粘, 注射剂的澄明度易受温度﹑pH 值的影响,故要指导中药保管人员结合每种药物特点合理贮藏,防止中药发霉、变色、虫蛀、泛油等,保证中药质量,从而发挥其应有疗效,减少不良反应的产生。
1.2 协助医师针对病情选择恰当的中药这一点是临床药师发挥药学服务的关键所在,要求药师必须具备较扎实的中医药知识和一定的临床经验,笔者认为,可从以下几点协助医师,提高临床治疗效果。
1.2.1 必须按照中医药理论辨证用药科学技术的发展促进了多学科的渗透、交叉,用现代药学理论来研究传统中药也取得了可喜成绩。
然而,与易理解的西药理论相比,中医药理论博深难明,以至于有人对中医持怀疑态度,也出现了用西医药理论指导中药应用的误区。
现代医药学和传统医药学两者建立的基础不同,理论体系、思维模式都存在差异,临床中药师应坚持宣传,提高他们对中医药理论的认识。
应用中药只有在中医药理论的指导下辨证施治,才能取得应有的治疗效果。