glutamate receptor-like channel3.3
新型广谱杀虫药沙罗拉纳在国外伴侣动物体外寄生虫防治中的应用

952021年第38卷第4期动物体外寄生虫严重危害我国畜牧业和宠物业的发展。
而抗寄生虫药滥用导致耐药性问题日益突出。
此外,有机氯等杀虫药物,在杀虫的同时,会广泛残留在环境中,造成环境污染;有机磷、拟除虫菊酯等药物毒性较高,使用不恰当易造成动物药物中毒。
因此,开发新型、高效、安全的杀虫药物,对防治动物外寄生虫具有十分重要的意义[1]。
新型广谱杀虫药沙罗拉纳在国外伴侣动物体外寄生虫防治中的应用徐燕婷,杨光友(四川农业大学动物医学院,四川成都 611130)摘 要:沙罗拉纳(sarolaner )属异恶唑啉类化合物,是一种新型广谱杀虫药。
沙罗拉纳已被证实通过特异性阻断昆虫γ-氨基丁酸受体和谷氨酸门控氯离子通道而显示出抗寄生虫活性。
本文综述了近年来沙罗拉纳在国外伴侣动物体外寄生虫防治中的开发及临床应用情况,旨在为该药的深入研究和国内开发应用提供参考。
近年来,该药主要用于犬猫等伴侣动物体外寄生虫的临床预防和治疗,对蜱类、蚤类、螨类等体外寄生虫有高效的杀灭作用。
与其他杀虫药相比,该药在分子结构、作用位点和选择性等方面有显著差异,具有杀虫活性强、治疗剂量低、安全性高等优点。
此外,该药通过与多种药物联合应用,可提高治疗效果,扩大治疗范围。
就国外研究来看,沙罗拉纳是一种安全广谱的高效杀虫剂,拥有广阔的应用前景。
关键词:沙罗拉纳;异恶唑啉;伴侣动物;体外寄生虫;杀虫剂;研究进展中图分类号:S855.9+1 文献标识码:A 文章编号:1005-944X (2021)04-0095-07DOI :10.3969/j.issn.1005-944X.2021.04.020 开放科学(资源服务)标识码(OSID ):Application of New Type of Sarolaner in Ectoparasite Control inCompanion Animals in Other CountriesXu Yanting ,Yang Guangyou(College of Veterinary Medicine ,Sichuan Agricultural University ,Chengdu ,Sichuang 611130,China )Abstract :Sarolaner ,a kind of isoxazoline compounds ,is a new broad-spectrum insecticide ,which could show antiparasitic activity through specifically blocking insect γ-aminobutyric acid (GABA )receptor and glutamate-gated chloride channel. In the paper ,the development and clinical application of Sarolaner in prevention and control of ectoparasites in companion animals in other countries were summarized to provide references for further study ,development and application in China. In recent years ,the drug had been mainly used for clinical prevention and treatment of ectoparasites in companion animals including dogs and cats ,which was highly effective for eradicating ectoparasites such as ticks ,fleas and mites. The drug ,with the advantages of strong insecticidal activity ,low therapeutic dose and high safety ,was greatly different from other insecticides in molecular structure ,action site and selectivity ,etc. In addition ,the coverage could be expanded with high effectiveness by combined application of the drug and others. As it was studied in other countries ,Sarolaner was a safe ,broad-spectrum and highly effective insecticide with promising prospects.Key words :sarolaner ;isoxazoline ;companion animal ;ectoparasite ;insecticide ;advance in study 收稿日期:2020-11-19 修回日期:2020-12-07基金项目:国家林业局林业科学技术研究项目(2018-03)通信作者:杨光友。
缺血性脑损伤与瞬时受体电位M通道的研究进展

缺血性脑损伤与瞬时受体电位M通道的研究进展缺血性脑血管病是临床常见病、多发病,其发病机制复杂。
钙超载在缺血性脑损伤中起重要作用。
瞬时受体电位M通道(transient receptor potential melastatin,TRPM)是位于细胞膜上的一类重要的非选择性阳离子通道超家族,对钙离子有较高的通透性,在缺血性脑损伤中起重要作用,对TRPM通道的研究将成为治疗缺血性脑损伤新的靶点。
本文就胞内钙离子超载在缺血性脑损伤中的作用、TRPM通道及其參与的缺血性脑损伤的机制予以综述。
标签:脑缺血;钙超载;瞬时受体电位M通道随着世界人口老龄化速度的加快,缺血性脑血管病的发病率逐年升高,现已成为威胁人类生命的最主要疾病之一,因其发病率、致残率和病死率高,给个人、家庭和社会带来巨大的精神压力和沉重的经济负担。
近年来研究发现,脑缺血和缺血后再灌能引起一系列病理和生化的变化,主要表现为脑能量耗竭,兴奋性氨基酸释放增加,胞内钙离子(intracellular calcium concentration,[Ca2+]i)超载,氧自由基产生增加,细胞坏死和凋亡等[1]。
其中,胞内钙离子超载被认为是脑缺血诱导神经元损伤的主要机制。
除了NMDA型谷氨酸受体和电压门控性钙通道以外,瞬时受体电位通道(transient receptor potential channels,TRP)因其对阳离子的通透性的特点[2],该通道家族在缺血性脑损伤中的作用受到越来越多的关注,并成为研究的热点之一。
1 [Ca2+]i与缺血脑损伤在生理情况下,细胞外钙离子(calcium,Ca)浓度为1~2mmol/L,细胞内则小于200nmol/L,两者相差10000多倍。
生理情况下,Ca2+主要由Ca2+通道和Na+-Ca2+交换两种途径进入胞内,胞内的Ca2+由钙泵泵出细胞外。
细胞内Ca2+处于一个动态平衡的状态,这种钙稳态对于维持细胞正常的生理功能有重要的意义[3]。
基于重症支气管哮喘差异表达基因及其治疗中药筛选的生物信息学分析

第 50 卷第 2 期2024年 3 月吉林大学学报(医学版)Journal of Jilin University(Medicine Edition)Vol.50 No.2Mar.2024DOI:10.13481/j.1671‐587X.20240214基于重症支气管哮喘差异表达基因及其治疗中药筛选的生物信息学分析陈丽平1,2, 韩立1, 卞华1, 庞立业1(1. 南阳理工学院河南省张仲景方药与免疫调节重点实验室,河南南阳473004;2. 河南中医药大学呼吸疾病中医药防治省部共建协同创新中心,河南郑州450046)[摘要]目的目的:通过生物信息学方法探讨重症支气管哮喘[简称重症哮喘(SA)]的差异表达基因,分析其作用机制,并筛选潜在具有治疗作用的中药及活性成分。
方法:在高通量基因表达(GEO)数据库中选取GSE136587和GSE158752数据集,利用R软件对数据集进行差异分析获得差异表达基因,并进行蛋白-蛋白相互作用(PPI)网络分析,筛选核心基因,寻找关键通路和枢纽基因。
最后将核心基因提交至Coremine数据库筛选具有潜在治疗作用的中药,并通过《中华医典》检索相关中药方剂。
结果结果:共筛选出466个差异表达基因。
通过STRING平台构建PPI网络共筛选包括25 kDa 突触关联蛋白(SNAP25)、谷氨酸离子型受体2(GRIA2)、轴突蛋白1(NRXN1)、钾电压门控通道亚家族A成员1(KCNA1)、突触囊泡蛋白 1(SYT1)和嗜铬蛋白A(CHGA)等核心靶点25个。
基因本体(GO)功能富集显示SA的生物学过程与细胞趋化性和白细胞迁移等有重要关系,京都基因与基因组百科全书(KEGG)富集的通路主要涉及骨髓白细胞迁移、白细胞趋化性、细胞趋化性、白细胞迁移、对外部刺激反应的正向调节和骨髓白细胞活化等信号通路。
采用网络药理学方法基于核心靶点筛选得到具有潜在治疗SA作用的中药367种,其中人参、水牛角、全蝎和黄芪等中药涉及多个核心靶点,与SA具有高度相关性,在《中华医典》中检索具有高度相关性的中药,共得到17个潜在具有治疗效果的中药方剂。
谷氨酸循环及谷氨酸兴奋性毒性
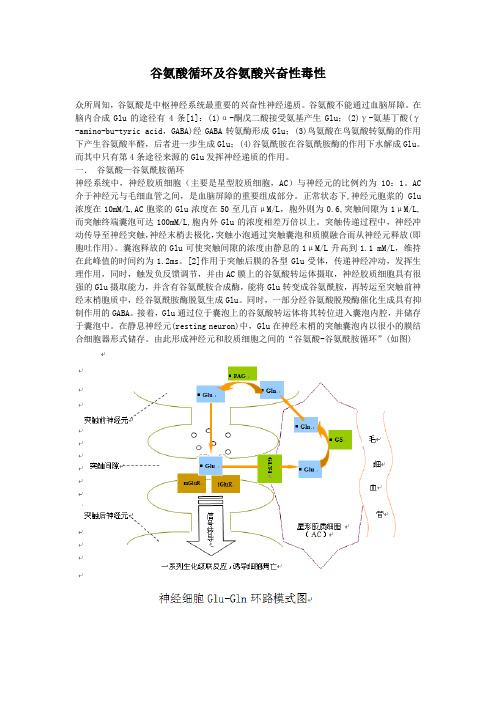
谷氨酸循环及谷氨酸兴奋性毒性众所周知,谷氨酸是中枢神经系统最重要的兴奋性神经递质。
谷氨酸不能通过血脑屏障。
在脑内合成Glu的途径有4条[1]:(1)α-酮戊二酸接受氨基产生Glu;(2)γ-氨基丁酸(γ-amino-bu-tyric acid,GABA)经GABA转氨酶形成Glu;(3)鸟氨酸在鸟氨酸转氨酶的作用下产生谷氨酸半醛,后者进一步生成Glu;(4)谷氨酰胺在谷氨酰胺酶的作用下水解成Glu。
而其中只有第4条途径来源的Glu发挥神经递质的作用。
一.谷氨酸—谷氨酰胺循环神经系统中,神经胶质细胞(主要是星型胶质细胞,AC)与神经元的比例约为10:1。
AC 介于神经元与毛细血管之间,是血脑屏障的重要组成部分。
正常状态下,神经元胞浆的Glu 浓度在10mM/L,AC胞浆的Glu浓度在50至几百μM/L,胞外则为0.6,突触间隙为1μM/L,而突触终端囊泡可达100mM/L,胞内外Glu的浓度相差万倍以上。
突触传递过程中,神经冲动传导至神经突触,神经末梢去极化,突触小泡通过突触囊泡和质膜融合而从神经元释放(即胞吐作用)。
囊泡释放的Glu可使突触间隙的浓度由静息的1μM/L升高到1.1 mM/L,维持在此峰值的时间约为1.2ms。
[2]作用于突触后膜的各型Glu受体,传递神经冲动,发挥生理作用,同时,触发负反馈调节,并由AC膜上的谷氨酸转运体摄取,神经胶质细胞具有很强的Glu摄取能力,并含有谷氨酰胺合成酶,能将Glu转变成谷氨酰胺,再转运至突触前神经末梢胞质中,经谷氨酰胺酶脱氨生成Glu。
同时,一部分经谷氨酸脱羧酶催化生成具有抑制作用的GABA。
接着,Glu通过位于囊泡上的谷氨酸转运体将其转位进入囊泡内腔,并储存于囊泡中。
在静息神经元(resting neuron)中,Glu在神经末梢的突触囊泡内以很小的膜结合细胞器形式储存。
由此形成神经元和胶质细胞之间的“谷氨酸-谷氨酰胺循环”(如图)二.谷氨酸受体GluR分为亲离子型受体和代谢型受体(mGluR)。
全麻机制研究——细胞内钙离子信号通路研究进展

全麻机制研究——细胞内钙离子信号通路研究进展常晶;陆菡;于布为【摘要】全身麻醉药物的分子作用机制及细胞作用机制虽不明确,但是其对神经递质释放的影响已经达到共识:一方面全身麻醉药物分子与离子通道蛋白或突触蛋白直接作用可以改变神经递质的释放,另一方面也可能通过改变细胞内信号通路而影响神经递质的释放.作为细胞内的重要信使,钙离子浓度([Ca2+]i)受到钙内流系统、钙外流系统及内质网等多方面调节,而存在于这些调节通路的蛋白受体大多数被证明是全身麻醉药物的潜在作用位点.细胞内钙离子最终影响细胞兴奋性和神经递质的释放的重要作用,使得全身麻醉药物调节 [Ca2+]i成为研究全麻机制的重要方面.%There is a broad consensus about anesthetics acting on neurotransmitter release in central nervous system although the molecular and cellular mechanisms of general anesthetics are still elusive. Changes in neurotransmitter release may relate to direct interaction of the anesthetic molecule with an ion channel protein or synaptic protein, but they can also be a consequence of alterations in intracellular signaling. Calcium is one of the most important messengers in cells and its intracellular concentration may be modulated by calcium influx system, calcium efflux system and endoplasmic reticulum which are affected by several agents including general anesthetics. The role of intracellular calcium in coupling neuronal excitation to the release of neurotransmitters into the synaptic cleft makes general anesthetic modulation of the intracellular calcium concentration ( [ Ca2+ ] )an important potential mechanism of action for these agents.【期刊名称】《中国药理学通报》【年(卷),期】2013(029)001【总页数】4页(P11-14)【关键词】钙离子信号;细胞内钙离子浓度;钙内流;钙外流系统;细胞内钙离子储存;全麻机制【作者】常晶;陆菡;于布为【作者单位】上海交通大学医学院附属瑞金医院麻醉科,上海,200025;上海交通大学医学院附属瑞金医院麻醉科,上海,200025;上海交通大学医学院附属瑞金医院麻醉科,上海,200025【正文语种】中文【中图分类】R-05;R284.1;R614.2;R971.2随着全身麻醉药物的发展,全身麻醉由于其安全、舒适、无痛,已在临床中得到广泛应用。
谷氨酸受体拮抗剂功能原理

谷氨酸受体拮抗剂功能原理英文回答:Glutamate receptor antagonists are a class of drugs that work by blocking the action of glutamate receptors in the brain. Glutamate is the primary excitatory neurotransmitter in the central nervous system and plays a crucial role in various physiological processes, including learning, memory, and synaptic plasticity. However, excessive glutamate activity can lead to neurotoxicity and is associated with various neurological disorders, such as epilepsy, Alzheimer's disease, and stroke.By antagonizing glutamate receptors, these drugs inhibit the binding of glutamate to its receptors, thereby reducing the excitatory effects of glutamate on neuronal activity. This helps to restore the balance between excitatory and inhibitory neurotransmission in the brain. There are several types of glutamate receptors, including NMDA receptors, AMPA receptors, and kainate receptors.Glutamate receptor antagonists can target specific subtypes of these receptors or act on multiple receptor subtypes.One example of a glutamate receptor antagonist is memantine, which is commonly used in the treatment of Alzheimer's disease. Memantine selectively blocks NMDA receptors, which are involved in learning and memory processes. By inhibiting the overactivation of NMDA receptors, memantine helps to improve cognitive function and reduce the symptoms of Alzheimer's disease.Another example is ketamine, which is known for its use as an anesthetic. Ketamine is a non-competitive antagonist of NMDA receptors and has been found to have rapid antidepressant effects in individuals with treatment-resistant depression. The exact mechanism of ketamine's antidepressant action is still under investigation, but it is believed to involve the modulation of glutamatesignaling and synaptic plasticity.Glutamate receptor antagonists can also have potential side effects. For example, blocking glutamate receptors canimpair normal synaptic transmission and affect cognitive function. Additionally, excessive inhibition of glutamate receptors can lead to sedation, dizziness, and other central nervous system side effects. Therefore, the use of glutamate receptor antagonists requires careful monitoring and dosage adjustment to minimize these adverse effects.Overall, glutamate receptor antagonists play a crucial role in the treatment of various neurological disorders by modulating glutamate signaling in the brain. Their ability to restore the balance between excitatory and inhibitory neurotransmission offers therapeutic potential for conditions such as Alzheimer's disease, epilepsy, and depression.中文回答:谷氨酸受体拮抗剂是一类通过阻断大脑中的谷氨酸受体来发挥作用的药物。
谷氨酸受体拮抗剂对培养大鼠海马神经元活力的影响

[文章编号]1006-2440(2012)06-0548-04谷氨酸是一种兴奋性氨基酸,主要有N-甲基-D-天冬氨酸(NMDA )和AMPA 两种亚型。
以往研究表明它对神经元具有兴奋性毒性作用[1],人们常用其受体拮抗剂来阻断这种毒性作用[2]。
但近年研究发现,谷氨酸的受体拮抗剂虽阻断了谷氨酸的毒性,但自身似乎也对细胞有一定的毒性,这种作用还不为人们所熟知[3]。
本研究分别使用NMDA 和AMPA 的拮抗剂作用于培养海马神经元,旨在探讨不同受体拮抗剂对神经元的影响。
1材料与方法1.1材料(1)动物:孕18天SD 胎鼠30只由南通大学动物实验中心提供。
(2)主要试剂:DMEM/F12培养液、Neurobasal 无血清神经培养基、B27、0.5%胰蛋白酶(Gibco 公司),台盼蓝(trypan )、四甲基偶氮唑盐(MTT )、酸化异丙醇、卓西平马来酸盐(MK-801)、艾芬地尔(ifenprodil )、CGP-37849、L-701.324、NBQX 和HA-966(上海宝曼生物公司);TUNEL 试剂盒(德国罗氏公司);多聚赖氨酸、阿糖胞苷(Sigma 公司)。
*[作者简介]李爱红,女,汉族,生于1970年7月,江苏如皋人,硕士,副主任医师。
研究方向:颅脑损伤及脑保护治疗。
通信作者:郭爱松,E-mail:guoaisong@谷氨酸受体拮抗剂对培养大鼠海马神经元活力的影响李爱红1*,郭爱松2,陈鑫1(南通大学附属医院1神经内科;2康复医学科,江苏226001)[摘要]目的:观察N-甲基-D-天冬氨酸受体拮抗剂卓西平马来酸盐、艾芬地尔、CGP-37849、L-701.324、HA-966和AMPA 受体拮抗剂NBQX 分别对培养大鼠海马神经元细胞活力的影响。
方法:取孕18天的SD 胎鼠海马神经元细胞原代培养,培养的细胞在加入上述药物后,在培养第2天和第7天行台盼蓝染色观察细胞存活情况,四甲基偶氮唑盐法检测细胞存活率,TUNEL 法检测细胞凋亡。
阿米替林改善手脚麻木的作用原理

阿米替林改善手脚麻木的作用原理阿米替林是一种抗癫痫和镇痛药物,它的作用原理主要涉及与神经传递物质的相互作用以及中枢神经系统的抑制作用。
首先,阿米替林与钠离子通道的相互作用起到了重要作用。
在神经细胞膜上,存在着多种类型的钠离子通道,其中导致迅速神经冲动传导的钠离子通道(Voltage-Gated Sodium Channel, VGSC)在神经冲动传递过程中起到了至关重要的作用。
阿米替林被认为是一种钠离子通道拮抗剂,它通过减少钠离子通道的流通而减缓或阻断了电位的维持和传导,从而抑制了异常的神经冲动产生和传递。
其次,阿米替林的药理效应可能与其具有的抗胆碱能活性有关。
截至目前,阿米替林是否通过抑制胆碱酯酶来干预胆碱能神经递质的作用仍存在争议。
但是,研究发现,阿米替林和其活性代谢物能够选择性地和神经元中的乙酰胆碱受体(Acetylcholine Receptor, AChR)结合,从而抑制乙酰胆碱的作用。
这也部分解释了阿米替林对于一些神经系统疾病,如帕金森病和阵发性震颤病等的治疗作用。
此外,阿米替林还具有对谷氨酸递质系统的调节作用。
谷氨酸是中枢神经系统中主要的兴奋性神经递质之一,它通过激活谷氨酸受体(Glutamate Receptor)而产生其作用。
研究发现,阿米替林抑制了某些亚型的谷氨酸受体的功能,从而减少了兴奋性信号的传递。
这一作用可能有助于阿米替林减缓或改善一些神经系统疾病的症状,如神经痛和帕金森病等。
此外,阿米替林还具有一定的GABA能系统影响。
GABA(γ-氨基丁酸)是中枢神经系统中主要的抑制性神经递质之一,它通过刺激GABA受体而产生其作用。
阿米替林被认为通过抑制GABA转氨酶(GABA Transaminase)来增加GABA 的含量,从而增强GABA能系统的抑制作用。
这可以改善一些过度兴奋性疾病的症状,如癫痫等。
总结起来,阿米替林对手脚麻木的作用主要涉及与钠离子通道、胆碱能神经递质及其受体、谷氨酸递质系统以及GABA能系统的相互作用。
昆虫离子型受体及其功能研究进展

http : ! www. insect. cry. cndol : 10.16380/j.kcxb. 2020.11.01211 月 November2020, 63(11): 1399 -1410昆虫学报ACTAENTOMOLOGICASINICA昆虫离子型受体及其功能研究进展郭金梦,董双林(南京农业大学植物保护学院,南京210095)摘要:昆虫是地球上最大的动物类群,在长期的进化过程中形成了复杂的感受系统,其中以化学感 受(嗅觉和味觉)最为重要&气味受体(odoanl orpyr , OR )和味觉受体(gustatoo oceptor , GR ) 分别在嗅觉和味觉中起关键作用,但近年发现离子型受体#ionotropic receptor, IR)也在一些化学感 受中发挥作用,有些IR 还参与温度和湿度等非化学感受& IR 家族由离子型谷氨酸受体# Pwotopbglutamate receptor, iRluR)进化而来,具有典型的3个跨膜结构域;与OR 相似,每个IR 至少需要一个共受体(ca-receptor )才能发挥功能,现已发现4个IR 共受体(IR8a , IT25a , IR76b 和IR93a );两个或多个IR 可以共表达于同一个嗅觉神经元,IR 间的不同组合导致神经元不同的气味反应谱&一种昆虫通常含有几个到上百个不等的IR,其中部分IR 为触角特异表达,但绝大多数IR 的功能 及其作用机制仍不清楚&本文综述了 IR 的结构和进化以及昆虫IR 基因鉴定、表达和功能等方面 的研究进展,以期为深入研究昆虫IR 的功能、作用机制及在害虫防治中的应用提供参考&关键词: (化学感受; 型受体;感器;嗅觉;味觉中图分类号:Q966 文献标识码:A 文章编号:0454-6296(2020)11-1399-12Research prgrss in ionotropic rcentor and their functions in insectsGUO Jin-Meng , DONG Shuang-Fin * * ( Colleye of Plant Protection , Nanjing AgTcultural University , Nanjing 210095, China )基金项目:国家自然科学基金项目(31672350, 31872300)作者简介:郭金梦,女,1992年4月生!山东泰安人,博士研究生!研究方向为昆虫生理生化与分子生物学! E-mail : ********************* 通讯作者 Corresponding aulCer , E-mail : slkong@ njau. cdu. cn收稿日期 Received : 2020-04-26 % 接受日期 Accepted : 2020-07-W8Abstract : As the laraest goup in the animal kingdom , insects have evolved campleo sensoo systems in osponso to various environmental stimuli , in which the chemosensation ( olfaction and gustation ) is the most important. The odorant receptors ( ORs) and gustatoo aceptors ( GRs) play a key ole in olfaction and gustation , eespectieee.Howeeee , in eecentXeaes , ithasbeen eound thationoteopiceeceptoes ( IRs ) are also involved in chemosensation as well as non-chemosensation such as sensation of temperature andhumidity.TheIR eamieyiseeoeeed eeom ionoteopicgeutamateeeceptoes ( iGeuRs ) which contain theee typical transmembrane domains. Similar to ORs , each IR needs to be cacxpossed with al lust one of the four identified ca-receptors(IR8a ,IR25a , IR76b a nd IR93a ) for its nomial function. Two or moreIRs con be cacxpossed in a same ollctoo neuron , and diReont IR combinations lexd to their diOeont responso profiles in neurons. One insect species usuVly contains several to over one hundred IRs, withsome IRs showing antennae-specibc expression pVterns , but most IRs remaining unknown in the functionand the mechanism of action . In this vticm , the resexoh advances in the structuo and evolution of IRs , and theidentieication , eipee s ion and eunction oeIR genesin insectsweeeeeeiewed , hopingtopeoeide some references for the further studies on the function and mechanisms of action of IRs , and the potentialuso of insect IRs as taraels in pest control.Key W o C s : Insect ; chemosensation % ionotropic receptor ; sensilla % olfaction %gustation1400昆虫学扌艮Ada Entomologicp Sinio63昆虫化学感受(包括味觉和嗅觉)在昆虫觅食、寻找配偶和选择产卵部位以及躲避天敌等行为中起到重要的作用(Hansson and SVnsmyr,2011)(已知的昆虫化学感受相关蛋白主要包括气味结合蛋白(odorant bindiny protein,OBP)、气味受体(odo/nl receptor,OR))化学感受蛋白(chemosensoy patein, CSP))味觉受体(yustatoy receptor,GR))感觉神经蛋白(sensoe neueon membeane peotein, SNMP))离子型受体(ionotropic receptor,IR)和气味降解酶(odorant deyradiny enzymo,ODE)等(Eyun ei a;,2017)oIR子受(ionoteopicgeutamate receptor,ibluR)家族演化而来,是一类保守的配体门控离子通道(liyand-gated ion channel),在配体分子与其结合后通道打开,导致膜内外离子的流动从而产生膜电位。
神经科学专业英语词汇
神经科学专业英语词汇摘要:神经科学是一门研究神经系统的结构、功能、发育、遗传、生化、生理、药理和病理的学科,涉及多个学科领域,如生物学、心理学、计算机科学、医学等。
本文列举了神经科学专业课所涉及的专业英语词汇,以供有兴趣的读者参考。
1. 神经科学导论中文英文神经科学neuroscience神经系统nervous system神经元neuron神经胶质细胞glial cell突触synapse神经递质neurotransmitter膜电位membrane potential动作电位action potential离子通道ion channel神经回路neural circuit神经网络neural network神经可塑性neural plasticity神经发育neural development神经退行性疾病neurodegenerative disease认知神经科学cognitive neuroscience行为神经科学behavioral neuroscience计算神经科学computational neuroscience分子神经科学molecular neuroscience细胞神经科学cellular neuroscience发育神经科学developmental neuroscience系统神经科学systems neuroscience感觉神经科学sensory neuroscience运动神经科学motor neuroscience情感神经科学affective neuroscience社会神经科学social neuroscience临床神经科学clinical neuroscience神经解剖学neuroanatomy神经生理学neurophysiology神经化学neurochemistry神经药理学neuropharmacology神经内分泌学neuroendocrinology神经免疫学neuroimmunology神经遗传学neurogenetics2. 神经元和神经胶质细胞中文英文多极神经元multipolar neuron双极神经元bipolar neuron单极神经元(假单极)unipolar neuron (pseudounipolar)轴突(轴索)axon (axis)轴突起始段axon hillock轴突终末axon terminal轴突分支axon collateral轴突鞘axon sheath髓鞘myelin sheath髓鞘间隙(诺氏结)node of Ranvier神经节ganglion神经纤维束nerve fiber神经nerve神经元突起neurite树突dendrite树突棘dendritic spine神经元体(细胞体)soma (cell body)细胞核nucleus细胞质cytoplasm细胞膜cell membrane核糖体ribosome内质网endoplasmic reticulum高尔基体Golgi apparatus线粒体mitochondrion微管microtubule微丝microfilament神经丝(神经原纤维)neurofilament (neurofibril)肌动蛋白actin微管相关蛋白(MAP)microtubule-associated protein (MAP)泰瑞酶(tau蛋白)tau protein (tau)神经纤维缠结(神经原纤维缠结)neurofibrillary tangle (neurofibril tangle)胶质细胞前体(祖细胞)glial progenitor (precursor) cell星形胶质细胞astrocyte沟通型星形胶质细胞protoplasmic astrocyte纤维型星形胶质细胞fibrous astrocyte水肿型星形胶质细胞reactive astrocyte胶质瘢痕glial scar脉络丛上皮细胞choroid plexus epithelial cell中文英文血脑屏障blood-brain barrier胶质限制膜glia limitans脑脊液cerebrospinal fluid微胶质细胞microglia髓鞘形成胶质细胞myelinating glial cell中枢髓鞘形成细胞(少突胶质细胞)central myelinating cell (oligodendrocyte)周围髓鞘形成细胞(雪旺氏细胞)peripheral myelinating cell (Schwann cell)雷迪克氏细胞(卫星细胞)satellite cell (enteric glial cell)神经干细胞neural stem cell神经前体细胞(祖细胞)neural progenitor (precursor) cell神经分化neural differentiation神经迁移neural migration3. 突触和神经递质中文英文突触synapse突触前膜presynaptic membrane突触后膜postsynaptic membrane突触间隙synaptic cleft突触小泡synaptic vesicle突触囊泡synaptic vesicle突触囊泡融合synaptic vesicle fusion突触囊泡回收synaptic vesicle recycling突触囊泡转运synaptic vesicle transport突触囊泡动力学synaptic vesicle dynamics突触递质释放synaptic transmitter release突触递质回收synaptic transmitter uptake突触递质降解synaptic transmitter degradation突触递质受体synaptic transmitter receptor突触递质效应synaptic transmitter effect神经递质neurotransmitter经典神经递质classical neurotransmitter非经典神经递质non-classical neurotransmitter小分子神经递质small molecule neurotransmitter大分子神经递质(神经肽)large molecule neurotransmitter (neuropeptide)乙酰胆碱(ACh)acetylcholine (ACh)乙酰胆碱酯酶(AChE)acetylcholinesterase (AChE)乙酰胆碱受体(AChR)acetylcholine receptor (AChR)胆碱能神经元(ACh神经元)cholinergic neuron (ACh neuron)胆碱能受体cholinergic receptor离子型胆碱能受体(nAChR)ionotropic cholinergic receptor (nAChR)代谢型胆碱能受体(mAChR)metabotropic cholinergic receptor (mAChR)神经肌肉接头neuromuscular junction副交感神经系统parasympathetic nervous system交感神经系统sympathetic nervous system胆碱能神经递质系统cholinergic neurotransmitter system多巴胺(DA)dopamine (DA)多巴胺合成酶(TH)dopamine synthase (TH)多巴胺转运蛋白(DAT)dopamine transporter (DAT)多巴胺受体(DR)dopamine receptor (DR)多巴胺能神经元(DA神经元)dopaminergic neuron (DA neuron)多巴胺能神经递质系统dopaminergic neurotransmitter system基底神经节basal ganglia黑质substantia nigra纹状体striatum前额叶皮层prefrontal cortex奖赏系统reward system去甲肾上腺素(NE)norepinephrine (NE)去甲肾上腺素合成酶(DBH)norepinephrine synthase (DBH)去甲肾上腺素转运蛋白(NET)norepinephrine transporter (NET)去甲肾上腺素受体(AR)adrenergic receptor (AR)去甲肾上腺素能神经元(NE神经元)noradrenergic neuron (NE neuron)去甲肾上腺素能神经递质系统noradrenergic neurotransmitter system蓝斑locus coeruleus脑干brainstem脑血管cerebral blood vessel应激反应stress response血清素(5-HT)serotonin (5-HT)血清素合成酶(TPH)serotonin synthase (TPH)血清素转运蛋白(SERT)serotonin transporter (SERT)血清素受体(5-HTR)serotonin receptor (5-HTR)血清素能神经元(5-HT神经元)serotonergic neuron (5-HT neuron)血清素能神经递质系统serotonergic neurotransmitter system神经管raphe nuclei杏仁核amygdala海马hippocampus松果体pineal gland节律调节circadian rhythm regulation情绪调节emotional regulation记忆形成memory formation抑郁症depression谷氨酸(Glu)glutamate (Glu)谷氨酸合成酶(GAD)glutamate synthase (GAD)谷氨酸转运蛋白(GLT)glutamate transporter (GLT)谷氨酸受体(GluR)glutamate receptor (GluR)谷氨酸能神经元(Glu神经元)glutamatergic neuron (Glu neuron)谷氨酸能神经递质系统glutamatergic neurotransmitter system离子型谷氨酸受体(iGluR)ionotropic glutamate receptor (iGluR)代谢型谷氨酸受体(mGluR)metabotropic glutamate receptor (mGluR) NMDA受体NMDA receptorAMPA受体AMPA receptorKA受体KA receptor长时程增强(LTP)long-term potentiation (LTP)长时程抑制(LTD)long-term depression (LTD)神经元突触可塑性synaptic plasticity学习和记忆learning and memory兴奋性毒性(凋亡)excitotoxicity (apoptosis)脑缺血cerebral ischemia脑损伤brain injury神经退行性疾病neurodegenerative diseaseγ-氨基丁酸(GABA)gamma-aminobutyric acid (GABA)GABA合成酶(GAD)GABA synthase (GAD)GABA转运蛋白(GAT)GABA transporter (GAT)GABA受体(GABAR)GABA receptor (GABAR)GABA能神经元(GABA神经元)GABAergic neuron (GABA neuron)GABA能神经递质系统GABAergic neurotransmitter system离子型GABA受体(GABAA和GABAC)ionotropic GABA receptor (GABAA and GABAC)代谢型GABA受体(GABAB)metabotropic GABA receptor (GABAB)抑制性突触传递inhibitory synaptic transmission抑制性突触后电位(IPSP)inhibitory postsynaptic potential (IPSP)突触抑制synaptic inhibition神经元抑制neuronal inhibition脑内抑制平衡inhibitory balance in the brain神经元兴奋性调节neuronal excitability regulation神经元同步化neuronal synchronization神经振荡neural oscillation4. 神经元膜电位和动作电位中文英文神经元膜电位neuronal membrane potential静息膜电位resting membrane potential膜阻抗membrane impedance膜电容membrane capacitance膜电阻membrane resistance膜导电membrane conductance电流current电压voltage电阻resistance导电conductance电路circuit欧姆定律Ohm's law电池battery电源power source电位差potential difference电容capacitance电荷charge电场electric field离子ion阳离子cation阴离子anion离子浓度ion concentration离子梯度ion gradient离子平衡ion equilibrium离子通道ion channel通透性permeability选择性selectivity电化学梯度electrochemical gradient动力学平衡dynamic equilibrium考克斯-福斯特方程(Nernst方程)Nernst equation (Goldman-Hodgkin-Katz equation)霍奇金-赫胥黎方程(GHK方程)Goldman-Hodgkin-Katz equation (GHK equation)电流-电压关系current-voltage relationship伊凡诺夫-马克斯韦尔方程(IV方程)I-V equation (Ivanov-Maxwell equation)离子通道类型ion channel type电压门控离子通道voltage-gated ion channel配体门控离子通道ligand-gated ion channel机械门控离子通道mechanosensitive ion channel热门控离子通道thermosensitive ion channel光门控离子通道light-gated ion channel离子通道结构ion channel structure离子通道亚基ion channel subunit离子通道孔域ion channel pore domain离子通道电压传感器ion channel voltage sensor离子通道配体结合位点ion channel ligand binding site离子通道状态ion channel state开放状态open state关闭状态closed state不应期refractory period激活activation失活inactivation钠离子通道sodium ion channel钾离子通道potassium ion channel钙离子通道calcium ion channel氯离子通道chloride ion channel动作电位action potential动作电位阈值action potential threshold动作电位上升相action potential rising phase动作电位峰值action potential peak动作电位下降相action potential falling phase动作电位超极化相(余极化相)action potential hyperpolarization phase (afterhyperpolarization phase)动作电位传播action potential propagation电缆理论cable theory电缆方程cable equation空间常数space constant时间常数time constant电流源current source电流汇current sink电流注入current injection电流刺激current stimulation电极electrode钳制技术clamp technique电压钳制voltage clamp电流钳制current clamp轴突初始段axon initial segment节间区internode region髓鞘myelin髓鞘形成myelination髓鞘退化demyelination髓鞘病变myelinopathy多发性硬化症(MS)multiple sclerosis (MS)传导速度conduction velocity盐跃式传导saltatory conduction节段性传导segmental conduction节段性抑制segmental inhibition节段性增强segmental facilitation节段性可塑性segmental plasticity5. 神经元突触传递和突触可塑性中文英文突触传递synaptic transmission突触前神经元presynaptic neuron突触后神经元postsynaptic neuron突触前释放presynaptic release突触后反应postsynaptic response突触后电位(PSP)postsynaptic potential (PSP)兴奋性突触后电位(EPSP)excitatory postsynaptic potential (EPSP)抑制性突触后电位(IPSP)inhibitory postsynaptic potential (IPSP)突触整合synaptic integration空间整合spatial summation时间整合temporal summation神经元输出neuronal output神经元输入-输出函数neuronal input-output function神经元输入阻抗neuronal input impedance神经元输出阻抗neuronal output impedance突触可塑性synaptic plasticity突触强度synaptic strength突触增强synaptic potentiation突触抑制synaptic depression短时程可塑性short-term plasticity长时程可塑性long-term plasticity短时程突触增强(STP)short-term potentiation (STP)短时程突触抑制(STD)short-term depression (STD)长时程突触增强(LTP)long-term potentiation (LTP)长时程突触抑制(LTD)long-term depression (LTD)突触前可塑性presynaptic plasticity突触后可塑性postsynaptic plasticityHebb学习规则Hebbian learning rule反Hebb学习规则anti-Hebbian learning rule突触修剪synaptic pruning6. 神经元网络和神经编码神经元网络neuronal network神经元群neuronal population神经元集合neuronal ensemble神经元回路neuronal circuit神经元模块neuronal module神经元连接neuronal connection突触连接synaptic connection电隙连接(电耦合)gap junction (electrical coupling)化学突触(化学耦合)chemical synapse (chemical coupling)突触权重synaptic weight突触矩阵synaptic matrix连接拓扑connection topology连接图connectivity graph连接概率connection probability连接密度connection density神经元活动neuronal activity神经元放电neuronal firing神经元放电频率neuronal firing rate神经元放电模式neuronal firing pattern神经元放电相位neuronal firing phase神经元放电序列neuronal firing sequence神经元放电同步neuronal firing synchronization神经元放电相关性neuronal firing correlation神经元放电耦合neuronal firing coupling神经元放电编码neuronal firing coding神经编码neural coding速率编码rate coding时序编码temporal coding相位编码phase coding空间编码spatial coding模式编码pattern coding群体编码population coding稀疏编码sparse coding分布式编码distributed coding局部化编码localized coding神经解码neural decoding逆向重建inverse reconstruction信息论information theory信息量information content信息熵information entropy互信息mutual information信噪比signal-to-noise ratio噪声抑制noise suppression信号增强signal enhancement信号处理signal processing7. 神经元网络模型和神经元网络仿真中文英文神经元网络模型neuronal network model神经元模型neuron model突触模型synapse model电路模型circuit model网络拓扑模型network topology model网络动力学模型network dynamics model网络功能模型network function model生物实际性(生物合理性)biological realism (biological plausibility)计算效率computational efficiency数学分析mathematical analysis数值仿真numerical simulation仿真软件simulation software仿真平台simulation platform仿真语言simulation language仿真脚本simulation script神经元网络仿真neuronal network simulation仿真结果simulation result仿真分析simulation analysis仿真验证simulation validation仿真优化simulation optimization仿真可视化simulation visualization仿真数据simulation data仿真参数simulation parameter仿真变量simulation variable仿真常量simulation constant仿真函数simulation function仿真对象simulation object仿真事件simulation eventHodgkin-Huxley模型(HH模型)Hodgkin-Huxley model (HH model) FitzHugh-Nagumo模型(FHN模型)FitzHugh-Nagumo model (FHN model)莱克-范德波尔模型(LIF模型)Leaky integrate-and-fire model (LIF model)伊兹克维奇模型(Izhikevich模型)Izhikevich model (Izhikevich model)中文英文NEURON软件NEURON software GENESIS软件GENESIS software NEST软件NEST software Brian软件Brian software PyNN软件PyNN software NEURON语言NEURON language HOC语言HOC language Python语言Python language MATLAB语言MATLAB language C语言 C languageC++语言C++ language Java语言Java languageR语言R language。
谷氨酸受体拮抗剂功能原理

谷氨酸受体拮抗剂功能原理英文回答:Glutamate receptor antagonists are a class of drugsthat work by blocking the action of glutamate receptors in the brain. Glutamate is the most abundant excitatory neurotransmitter in the central nervous system and plays a crucial role in various brain functions, including learning, memory, and synaptic plasticity.By antagonizing glutamate receptors, these drugs can inhibit the effects of glutamate and reduce its excitatory activity. This can be beneficial in certain conditionswhere excessive glutamate activity is thought to contribute to pathological processes, such as epilepsy, neurodegenerative disorders, and chronic pain.For example, in the treatment of epilepsy, excessive glutamate release can lead to overexcitation of neurons and the generation of seizures. Glutamate receptor antagonists,such as memantine, can block the effects of glutamate and help prevent the occurrence of seizures.In neurodegenerative disorders like Alzheimer's disease, there is evidence of excessive glutamate release and excitotoxicity, which can contribute to neuronal damage and cognitive decline. Glutamate receptor antagonists, such as riluzole, can help reduce glutamate-mediated excitotoxicity and potentially slow down the progression of the disease.In chronic pain conditions, glutamate receptors in the spinal cord play a crucial role in transmitting pain signals. By blocking these receptors, drugs like ketamine can help alleviate pain and provide relief to patients.Overall, the functional principle of glutamate receptor antagonists lies in their ability to block the action of glutamate and reduce its excitatory effects. This can have therapeutic benefits in conditions where excessiveglutamate activity is involved.中文回答:谷氨酸受体拮抗剂是一类通过阻断大脑中的谷氨酸受体作用的药物。
谷氨酸受体iGluR_抑制剂_激动剂

iGluRIonotropic glutamate receptorsiGluR (ionotropic glutamate receptor) is a ligand-gated ion channelthat is activated by the neurotransmitter glutamate. iGluR are integralmembrane proteins compose of four large subunits that form acentral ion channel pore. Sequence similarity among all knownglutamate receptor subunits, including the AMPA, kainate, NMDA,and δ receptors.AMPA receptors are the main charge carriers during basaltransmission, permitting influx of sodium ions to depolarise thepostsynaptic membrane. NMDA receptors are blocked by magnesiumions and therefore only permit ion flux following prior depolarisation.This enables them to act as coincidence detectors for synaptic plasticity. Calcium influx through NMDA receptors leads to persistent modifications in the strength of synaptic transmission.iGluR Inhibitors & Modulators(S)-(-)-5-Fluorowillardiine is a potent and specific AMPAR agonist.(S)-(-)-5-Fluorowillardiine Hcl is a potent and specific AMPAR agonist (S)-Willardiine is a potent agonist of AMPA/kainate receptors with24-Hydroxycholesterol is a natural sterol, which serves as a positive,N-Methyl-d-Aspartate (NMDA) receptorsRmodulator7-Chlorokynurenic acid is a selective antagonist at the glycinecomplexintoreuptake of glutamate7-Chlorokynurenic acid sodium salt is a selective antagonist at theN-methyl-D-aspartate receptorreuptake of glutamate antagonist and anCat. No.: HY-16713Cat. No.: HY-16713A Cat. No.: HY-12499Cat. No.: HY-N2370 Cat. No.: HY-B2121Cat. No.: HY-100811 Cat. No.: HY-100811A Cat. No.: HY-15073CFM-2 is a selective non-competitive AMPAR antagonist.CIQ is a subunit-selective potentiator of NMDA receptors containing CMPDA is a positive allosteric modulator of AMPA receptors with EC50s of 45.4 ± 4.2 nM/63.4 ± 5.6 nM for GluA2i/GluA2o receptor.receptor antagonist.ion-channel antagonist receptor/ion channel site to produce a Coluracetam(MKC-231) is a new choline uptake enhancerCX546 is a selective positive AMPAR modulator; the prototypicalCat. No.: HY-102053Cat. No.: HY-12503Cat. No.: HY-18699Cat. No.: HY-12508Cat. No.: HY-15066Cat. No.: HY-101809Cat. No.: HY-17553Cat. No.: HY-12505Dynorphin A (1-10) an endogenous opioid neuropeptide, binds to . Dynorphin A (1-10) of 42.0 μM.Eliprodil(SL-820715) is a non-competitive NR2B-NMDA receptor antagonist(IC50=1 uM), less potent for NR2A- and NR2C-containing receptor positive Felbamate (FBM) is a potent nonsedative anticonvulsant whose Felbamate hydrate (FBM) is a potent nonsedative anticonvulsant Flupirtine(D 9998) is a selective neuronal potassium channel opener Flupirtine Maleate(D 9998) is a selective neuronal potassium channelopener that also has NMDA receptor antagonist propertiesCat. No.: HY-P1594Cat. No.: HY-12881Cat. No.: HY-10937Cat. No.: HY-B0184Cat. No.: HY-B0184ACat. No.: HY-17001ACat. No.: HY-17001Cat. No.: HY-100785NMDAα-amino-3-hydroxy-5-methylisoxazole-4-propionic acid ( )AMPAIbotenic acid has agonist activity at both the N-methyl-D-aspartate ( ) and trans-ACPD or metabolotropic quisqualate (Q )m IC87201, an inhibitor of PSD95-nNOS protein-protein interactions,-dependent NO and cGMP formation.Ifenprodil tartrate is a novel N-methyl-D-aspartate (NMDA) receptor antagonist that selectively inhibits receptors containing the NR2Bagonistbeta-adrenergic receptors of 13.65 μΜ and 3.48 μΜ for myometrial and placcntal beta-adrenergic receptor, respectively. Isoxsuprine hydrochloride isKynurenic acid, an endogenous tryptophan metabolite, is a broad-spectrum antagonist targeting <b >NMDA, glutamate, α7 nicotinic acetylcholine receptor. Kynurenic acid is also a selective Kynurenic acid, an endogenous tryptophan metabolite, is a broad-spectrum antagonist targeting <b >NMDA, glutamate, α7 nicotinic acetylcholine receptor. Kynurenic acid is also a selectiveCat. No.: HY-19243Cat. No.: HY-103228 Cat. No.: HY-N2311Cat. No.: HY-100457 Cat. No.: HY-12882A Cat. No.: HY-B1270 Cat. No.: HY-100806Cat. No.: HY-107512LY451395(Mibampator) is a potent and highly selective potentiator MDL 105519 is a potent and selective antagonist of glycine binding NMDA receptor H]glycine binding in vitro and in Meclofenoxate hydrochloride, an ester of dimethylethanolamine (DMAE) and 4-chlorophenoxyacetic acid (pCPA), has been shown to improve memory, have a mentally stimulating effect, and improve Memantine, an amantadine derivative with low to moderate-affinity for NMDA receptors, inhibit CYP2B6 and CYP2D6 with Ki of 0Mephenesin is an NMDA receptor antagonist, is a centrally acting MRZ 2-514 is an antagonist of the strychnine-insensitive modulatory Naspm (1-Naphthyl acetyl spermine), a synthetic analogue of Joro ) receptors Cat. No.: HY-10934Cat. No.: HY-15085Cat. No.: HY-16312Cat. No.: HY-17555Cat. No.: HY-B0365ACat. No.: HY-B1283Cat. No.: HY-101620Cat. No.: HY-12506)NMDA mimicking theaction of glutamate, the neurotransmitter which normally acts at that NMDA-IN-1 is a potent and NR2B-selective NMDA antagonist with Ki of 0.85 nM; NR2B Ca2+ influx IC50 is 9.7 nM; no activities on NR2A,NR2C, NR2D, hERG-channel and α1-adrenergic receptor.Noopept (GVS-111) is a medication promoted and prescribed in NT 13 (TPPT) is a tetrapeptide having the amino acid sequence L-threonyl-L-prolyl-L-prolyl-L-threonine amide. NT 13 is a partial ) agonist used in the study receptor positive allosteric modulator.Orphenadrine citrate is a NMDA receptor antagonist with Ki of 6.0N-methyl-D-aspartate (NMDA) receptor antagonist with Ki of 6Cat. No.: HY-106408Cat. No.: HY-17551Cat. No.: HY-12962Cat. No.: HY-17456Cat. No.: HY-P7060Cat. No.: HY-101216Cat. No.: HY-B0369ACat. No.: HY-B1126PEPA is an allosteric modulator of AMPA receptors; binds to theGluA2o and GluA3o LBDs and can be utilized as an indicator ofreceptorAMPARs of 263 and 296 nM,gamma-aminobutyric acid (GABA), used in treatment of a wide rangeProcyclidine hydrochloride is a potent anti-cholinergic agent, and is QNZ46 is a NR2C/NR2D-selective NMDA receptor non-competitiveantagonist (IC50 values are 3, 6, 229, and >300, >300 μM for NR2D,Rapastinel (GLYX-13) is an N-methyl-D-aspartate receptor (NMDAR) modulator that has characteristics of a glycine site partial agonist.modulator with glycine-site partial agonist properties and currently in a phase II clinical development program as an adjunctive therapy for majorCat. No.: HY-12509Cat. No.: HY-14451 Cat. No.: HY-104020A Cat. No.: HY-B0585 Cat. No.: HY-B1487Cat. No.: HY-15703 Cat. No.: HY-16728Cat. No.: HY-16728BRo 25-6981 Maleate is a potent and selective activity-dependentblocker of NMDA receptors containing the NR2B subunit. IC50 values are 0.009 and 52 μM for cloned receptor subunit combinations , with potent SDZ 220-581 is a potent, competitive antagonist at the NMDASDZ 220-581 ammonium salt is a potent, competitive antagonist at SDZ 220-581 Hcl is a potent, competitive antagonist at the NMDASunifiram (DM-235) is a piperazine derived ampakine-like drug which has nootropic effects in animal studies with significantly higher SYM 2206 is a novel, potent, non-competitive AMPA receptor Talampanel is a potent and selective AMPA-receptor antagonist, is a Cat. No.: HY-13993ACat. No.: HY-101600Cat. No.: HY-13059Cat. No.: HY-13059ACat. No.: HY-13059BCat. No.: HY-17550Cat. No.: HY-18689Cat. No.: HY-15079antagonist NMDAof 10 -240455 is a potent and selective N-methyl D-aspartate ( )NMDAYM 872 is a selective antagonist of the glutamate receptor subtype,α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA)ZK200775(Fanapanel; MPQX) is a highly selective AMPA/kainate antagonist with little activity against NMDA; have Ki values of 3.2 nM, 100 nM, and 8.5 μM against quisqualate, kainate, and NMDA,ZK200775 hydrate(Fanapanel; MPQX) is a highly selectiveAMPA/kainate antagonist with little activity against NMDA; have Ki values of 3.2 nM, 100 nM, and 8.5 μM against quisqualate, kainate,ZL006 is a potent inhibitor of nNOS/PSD-95 interaction, and inhibitsHY-W018061Cat. No.: HY-19391 Cat. No.: HY-15072Cat. No.: HY-15069 Cat. No.: HY-15069A Cat. No.: HY-100456。
2020年高考天津卷第14题的深度分析及习题选编策略

2021年第5期2020年高考天津卷第14题的深度分析及习题选编策略广东省中山市第二中学(528429)许停高飞燕摘要对2020年高考生物学天津卷第14题从素材来源、命题立意、解题思维等角度进行了深度 分析,提出了高三生物学教学中习题选编的启示。
关键词高考试题;深度分析;习题选编文章编号 1005 -2259(2021)5 -007丨-04高考的核心功能是立德树人、服务选材和引导 教学。
高考试题是良好的教学素材,对其进行深度 分析,有利于领会命题立意、厘清命题思维、把握命 题倾向,对教学具有重要的反拨和引导作用。
2020 年高考生物学天津卷第14题(以下简称“第14 题”)颇具代表性,笔者尝试对其进行深度分析,以期总结高三生物学教学中的习题选编策略。
1原题赏析例题神经细胞间的突触联系往往非常复杂。
2.3构建模型活动,彰显科学思维模型是对研究对象的简化和概括,是科学研究 中对复杂生命现象的简单化描述方法。
台湾地区 会考试题中经常以实物或图画的形式呈现模型,能 形象直观地呈现生物对象的特征。
模型建构是科 学思维的重要维度之一,能反映考生在复杂情境中 运用重要概念、原理、规律和生物学科思想方法解决 问题的能力。
在新的模型情境中,考生通过解读构 建模型,能准确地提取关键信息进行图文转换,阐明 模型的本质特征,从而揭示生命现象的本质和规律。
例5图4a为植物体内物质运输示意图,图4b为叶片气孔状态示意图。
白天植物进行旺盛的 蒸腾作用时,有关体内水分运输方向(甲或乙)及 叶片气孔状态(丙或丁),下列关于组合正确的是(D)A•甲,丙 B.甲,丁 C.乙,丙 D.乙,丁评析本题以绿色植物白天进行蒸腾作用为载体,用图文并茂的方式呈现情境,考查的重要概念图1为大鼠视网膜局部神经细胞间的突触示意图。
涉及物质运输、蒸腾作用和气孔的关闭。
试题要求考生从图文信息中准确判断水分在茎中的运输方 向,并与气孔状态模型联结起来,从而建构宏观水平水分在植物体内运输和微观水平气孔关闭的二 级模型,考查考生高层次模型建构能力,深化其结 构与功能相适应的生命观念。
TRPM2通道与炎症反应

TRPM2通道与炎症反应①索婷姣蒋丽娜赵自刚②(河北北方学院微循环研究所,张家口075000)中图分类号R392R364.5文献标志码A文章编号1000-484X(2022)12-1523-05[摘要]瞬时感受器电位M2型(TRPM2)通道是一种非谷氨酸依赖性离子通道,是瞬时受体电位(TRP)通道超家族成员之一。
不同致病因素作用于各种免疫细胞,引起TRPM2通道开放,导致阳离子内流,尤其是Ca2+,进而调节诸多信号通路,引起炎症反应的发生发展。
本文聚焦TRPM2通道在炎症反应中的免疫学作用,期望为靶向TRPM2通道决策炎症相关疾病的治疗措施拓展新的思路。
[关键词]瞬时感受器电位M2型通道;Ca2+;炎症反应;免疫细胞TRPM2channel and inflammationSUO Tingjiao,JIANG Lina,ZHAO Zigang.Institute of Microcirculation,Hebei North University,Zhangjiakou 075000,China[Abstract]Transient receptor potential M2(TRPM2)channel,a glutamate-independent ion channel,is a member of the tran⁃sient receptor potential(TRP)channel superfamily.Different pathogenic factors act on all kinds of immune cells,causing the opening of TRPM2channels,resulting in cation influx,especially Ca2+,which in turn regulates many signaling pathways and participates in development of inflammatory responses.The article focuses on the immunological roles of TRPM2channels during inflammation,and provides a new way for targeting TRPM2channels to determine therapeutic strategies for inflammation-related diseases.[Key words]Transient receptor potential M2channel;Ca2+;Inflammation;Immune cell瞬时受体电位(transient receptor potential,TRP)通道由700~2000个氨基酸残基组成,位于细胞膜或胞内细胞器膜上的一类重要的非选择性阳离子通道,在研究果蝇光反应异常突变时发现。
胰高血糖素

主要用途
胰高血糖素是促进分解代谢的激素。它促进肝糖原分解和糖异生的作用很强,使血糖明显升高;促进脂肪分 解和脂肪酸氧化;加速氨基酸进入肝细胞,为糖异生提供原料。血糖浓度亦是调节胰高血糖素分泌的主要因素。 血糖降低,胰高血糖素分泌增多,反之则减少;胰岛素可通过降低血糖而间接促进胰高血糖素分泌,也可通过旁 分泌方式,直接作用于邻近A细胞,抑制其分泌;交感神经促进胰高血糖素分泌,迷走神经则抑制其分泌。
胰高血糖素
脊椎动物胰脏的胰岛α细胞分泌的激素
01 基本信息
03 主要作用
目录
02 组织结构 04 分泌调节
05 主要用途
07 分泌机制
目录
06 药品说明
胰高血糖素(glucagon)亦称胰增血糖素或抗胰岛素或胰岛素B。它是伴随胰岛素由脊椎动物胰脏的胰岛α 细胞分泌的一种激素。与胰岛素相对抗,起着增加血糖的作用。
谢谢观看
药品说明
药品名称:升血糖素、胰高血糖素、高血糖素
英文名称:Glucagon
适应症:该品现主要用于低血糖症,在一时不能口服或静注葡萄糖时非常有用。不过,通常低血糖时仍应首 选葡萄糖。近来亦用于心源性休克。
用量用法:肌注、皮下注射或静注,用于低血糖症,每次0.5mg~1.0mg,5分钟左右即可见效。如20分钟仍 不见效,则应尽快应用葡萄糖。用于心源性休克,连续静脉输注,每小时1~12mg。
胰高血糖素(升糖素)是一种由胰脏胰岛α-细胞分泌的激素,由29个氨基酸组成直链多肽,分子量为3485 道尔顿。胰高血糖素的第一级结构是:NH2-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-LeuAsp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp-Leu-Met-Asn-Thr-COOH。
谷氨酸glu适配体序列

谷氨酸glu适配体序列Glutamate (Glu) is one of the most abundant excitatory neurotransmitters in the central nervous system. It plays a crucial role in various physiological processes, including learning, memory, and synaptic plasticity. As a result, understanding the structure and function of glutamate receptors, the proteins that bind to glutamate, is of great importance in neuroscience research. One key aspect of this research is the identification and characterization of glutamate receptor ligands, or compounds that bind to the receptor and modulate its activity. These ligands, also known as glutamate receptor agonists or antagonists, can be natural or synthetic compounds and are widely used in both basic research and drug development.The sequence of the glutamate receptor ligand-binding domain, or the part of the receptor that directly interacts with glutamate and other ligands, is of particular interest to researchers. This sequence, often referred to as the glutamate receptor ligand-binding domain sequence or simplythe glutamate receptor sequence, is crucial for understanding the molecular mechanisms of ligand binding and receptor activation. By studying this sequence, researchers can gain insights into the structure-function relationships of glutamate receptors and develop new ligands with improved selectivity and efficacy.In recent years, advances in molecular biology and bioinformatics have greatly facilitated the identification and analysis of glutamate receptor sequences. High-throughput sequencing technologies, such as next-generation sequencing, have enabled the rapid and cost-effective determination of glutamate receptor sequences from various species, including humans, rodents, and non-human primates. These sequences can then be analyzed using bioinformatics tools to identify conserved regions, predict protein structures, and design experiments to study the function of specific domains.The glutamate receptor ligand-binding domain sequenceis also of interest in the field of drug discovery. By understanding the specific amino acid residues andstructural motifs that are critical for ligand binding, researchers can design and screen small molecules or biologics that modulate glutamate receptor activity. This approach has led to the development of several drugs that target glutamate receptors, including the N-methyl-D-aspartate (NMDA) receptor, for the treatment ofneurological and psychiatric disorders. For example, memantine, a non-competitive NMDA receptor antagonist, is used to treat Alzheimer's disease, while ketamine, a rapid-acting antidepressant, acts as an NMDA receptor antagonist.Furthermore, the glutamate receptor ligand-binding domain sequence is important for understanding the evolutionary history of glutamate receptors. By comparing the sequences of glutamate receptors from different species, researchers can gain insights into the molecular changesthat have occurred during evolution and identify conserved and divergent features of these receptors. This comparative approach can provide valuable information about the functional diversity of glutamate receptors and their roles in different organisms.In conclusion, the glutamate receptor ligand-binding domain sequence is a key element in the study of glutamate receptors and their ligands. By analyzing this sequence, researchers can gain insights into the molecular mechanisms of ligand binding, develop new drugs for neurological and psychiatric disorders, and explore the evolutionary history of glutamate receptors. The continued study of the glutamate receptor sequence is essential for advancing our understanding of the brain and developing novel therapeutic interventions for neurological and psychiatric conditions.。
神经科学研究中的光遗传学

神经科学研究中的光遗传学光遗传学是一种研究光感受器如何控制生物体行为和生理过程的方法。
它利用遗传工程技术,将光感受器蛋白基因导入到目标细胞中,从而能够通过光的输入直接操纵细胞活动。
在神经科学研究中,光遗传学已经成为一种强有力的工具,为我们理解神经信号传导和神经网络行为提供了新的途径。
由于神经元是神经网络的基本单元,因此神经元发放的电信号非常重要。
在神经系统的基础研究中,科学家们一直在探索如何通过特定的刺激来操纵神经元活动。
早期研究主要使用电刺激、化学刺激等手段来激活或抑制神经元。
这些方法通常无法针对特定类型的神经元活动,并且也受到信号传播的限制。
光遗传学则提供了一种直接、非侵入性的操纵神经元的方法。
它利用一些特殊的蛋白质,如光敏离子通道和离子泵,这些蛋白质是从一些盲色动物中发现的,它们具有高灵敏度、高选择性和快速响应的表型。
根据这些蛋白的功能不同,光遗传学被分为两大类:光激活类和光抑制类。
光激活类蛋白质主要是光敏离子通道,如嘌呤核苷酸敏感的钠通道(channelrhodopsin,ChR2)和谷氨酸离子通道(glutamatereceptor ion channel,GluCl),这些蛋白质在接受光信号后可以将离子通道打开,从而引发神经元的兴奋性。
光抑制类蛋白质则主要是光敏离子泵,如大飞蛾蛋白(Halorhodopsin,NpHR),它可以在接受光信号时将离子通道关闭,从而抑制神经元的兴奋性。
光遗传学在神经科学研究中的应用非常广泛。
例如,在研究学习与记忆时,科学家可以在小鼠海马中表达ChR2蛋白,接着通过光刺激控制海马神经元的兴奋性,模拟学习和遗忘的过程。
在探究神经退行性病变时,科学家可以使用NpHR蛋白抑制神经元的兴奋性,从而减轻神经元衰竭带来的影响。
光遗传学的应用不仅限于实验室的基础研究,它也有着广泛的应用前景,在神经系统疾病的临床治疗方面也有很大潜力。
例如,利用光遗传学可以在脑区域中激活或抑制特定类型的神经元,从而缓解癫痫、精神分裂症等神经系统疾病的症状。
拟南芥AtGLRs基因家族的研究

拟南芥AtGLRs基因家族的研究叶放;陈庆;朱世华【摘要】AtGLRs是拟南芥中类似动物iGluRs的受体家族,在植物的钙离子信号传导和转运、光信号传导、碳氮代谢调节、ABA生物合成、胁迫响应、根的生长等方面起着重要作用,文章对AtGLRs基因功能的最新研究进展进行了综述.【期刊名称】《宁波大学学报(理工版)》【年(卷),期】2011(024)001【总页数】4页(P14-17)【关键词】拟南芥;iGluR;AtGLR;谷氨酸【作者】叶放;陈庆;朱世华【作者单位】宁波大学生命科学与生物工程学院,浙江宁波31521l;宁波大学生命科学与生物工程学院,浙江宁波31521l;宁波大学科学技术学院,浙江宁波315211【正文语种】中文【中图分类】Q344+.4L-谷氨酸是动物的一种重要兴奋性神经递质,在中枢神经系统中, 离子通道型谷氨酸受体(Ionotropic Glutamate Receptors, iGluRs)作为谷氨酸门控的离子通道, 介导了大部分的神经兴奋传递[1].研究发现, 在没有神经系统的植物中, 如拟南芥、烟草、豌豆、玉米和水稻中也有编码类似受体(Glutamte Receptors, GLRs)的基因存在, 其中拟南芥存在由20个谷氨酸受体基因组成的基因家族, 植物中的此类基因吸引了一些学者的研究, 笔者对AtGLRs基因功能研究的成果和进展进行了综述.1998年在拟南芥中首次发现了与哺乳动物iGluRs高度同源的拟南芥谷氨酸受体(A. thaliana Glutamate Receptors, AtGLRs)基因[2], 其后的研究发现了一个含20个AtGLRs的基因家族[3], 这些基因可以划分为 3个簇(Cluster)或亚族(Subfamily),用RT-PCR分析根、叶、花和果4个器官中20个基因的表达, 发现20个基因中有5个仅在根部特异性表达, 这5个AtGLRs基因都属于第2亚族; 剩余 15个基因除了在根中表达外, 在其他器官中也有表达[4]. 单一细胞质mRNA RT-PCR的结果显示: AtGLRs基因协同表达的种类和数量在同一个体的不同组织(表皮和叶肉)间差异较小, 而在不同个体的相同组织(表皮和叶肉)间却存在较大差异. 说明AtGLR基因家族成员间可能在功能上存在冗余[5].对植物 GLRs氨基酸序列的分析发现, 植物GLRs能够形成类似动物离子通道型谷氨酸受体的离子通道结构, 它们的跨膜结构域和配体结合结构域氨基酸序列与iGluRs高度同源: 含6个保守结构域, 包括4个跨膜结构域: M1~M4以及2个配体结合结构域GlnH1(S1)和GlnH2(S2)[6-7], 这些谷氨酸受体的结构相似, 由 4~5个亚单位聚合形成离子通道, 每个亚单位由一个基因编码. 相同亚单位聚合形成同源多聚体离子通道, 不同亚单位聚合成异源多聚体通道, 对不同离子的通透性因亚基的组成而异[6].植物GLRs与动物iGluRs之间的分离可能早于动物iGluRs的分化, 因为植物GLRs与动物3种iGluRs (NMDA/AMPA/KA受体)不同, 单独为一簇[4]. GLRs、iGluRs有2个与细菌周质结合蛋白同源的结构域, 作为激动剂结合位点: 其中一个属于赖氨酸/精氨酸/鸟氨酸受体(LAOBP)家族; 另一个则是N末端结构域(NTD), 属于亮氨酸/异亮氨酸/缬氨酸受体(LIVBP)家族, LIVBP结构域的功能为激动剂变构结合和促进亚基装配成寡聚蛋白[8]. 在动植物谷氨酸受体分化前, 一次重组事件将GLRs LIVBP域同G蛋白偶联受体7个跨膜结构域融合,产生了原始subC-GPCRs,而iGLRs基因通过GLRs基因的点突变, 选择进化而来. 因此与动物iGluRs LIVBP 域相比, 植物GLRs LIVBP域同subC-GPCRs LIVBP域的联系更为密切, 推测远古的植物 GLR基因是iGLRs和subC-GPCRs基因的共同祖先[9].4.1 钙离子通道和钙离子转运4.1.1 钙离子通道根尖细胞在外源谷氨酸处理下会产生快速的细胞膜去极化, 同时伴随胞质内 Ca2+浓度峰值出现; 用质膜Ca2+通道抑制剂La3+处理后, 谷氨酸就不能引起胞质Ca2+浓度的增加和细胞膜的去极化;用Ca2+螯合剂EGTA预处理根尖, 谷氨酸同样不能引起胞质Ca2+浓度的增加, 说明谷氨酸能触发Ca2+通过质膜流入胞内从而导致胞质 Ca2+浓度的显著变化[10]. 动物iGluRs拮抗剂DNQX (6,7 dinotropuinoxaline 2,3 (1H, 4H) dione)可以阻断子叶中谷氨酸(甘氨酸)引起的胞质Ca2+浓度增加[11]. 进一步利用突变体glr3.3进行的电生理实验发现, 谷氨酸不能激发突变体 glr3.3的细胞膜去极化和胞质 Ca2+浓度峰值出现, 因此推测AtGLRs参与了植物的钙离子调控[12]. 通过与大鼠GluR1和GluR6的融合实验, 发现AtGLR1.1和AtGLR1.4具有对Na+、K+和 Ca2+选择性通透的核心结构域[13]. 过度表达AtGLR3.1基因会削弱 Ca2+诱导的气孔关闭, 说明与 Ca2+调控的气孔关闭运动相关[14]. 因此, 推测AtGLRs具有谷氨酸门控的钙离子通道功能. 4.1.2 钙离子转运通过RT-PCR和mRNA印迹方法发现AtGLR3.2基因的转录产物主要集中在分裂旺盛的组织和较为成熟的胚轴、叶和花芽的维管组织中, 推测可能与钙离子转运相关[15].AtGLR3.2的过度表达使植株产生了缺钙症状,导致根尖坏死和叶尖坏疽. 但转基因植株的总钙量同野生型相比差异并不显著, 经过量 Ca2+处理挽救后, 钙缺乏症状消失, 植株恢复正常. 表明过度表达的 AtGLR3.2并不影响 Ca2+的吸收和积累, 而是导致 Ca2+的利用率降低才产生缺钙症状. AtGLR3.2启动子融合GUS转化拟南芥, GUS基因在根和茎的维管组织中表达, 在导管邻近细胞中的表达尤其强烈, 推测AtGLR3.2编码的亚单位参与了钙离子转运[16]. AtGLR1.3和AtGLR3.3启动子融合GUS转化拟南芥, AtGLR1.3主要在根尖皮层表达, 而 AtGLR3.3主要在植株维管组织表达, 因为植物吸收土壤中的矿物质等离子与根尖的皮层组织相关而维管组织是植物运输矿物质的通道, 暗示它们的功能可能分别与 Ca2+的吸收和转运相关[17]. 此后, 在小萝卜中过度表达 RsGluR也导致与过度表达AtGLR3.2相似的表型[18].4.2 光信号传导光培养条件下, 生长在含 DNQX平板上的拟南芥幼苗其下胚轴长度是对照长度的1.5~2倍, 同时 DNQX削弱了幼苗的叶绿素合成, 这些效应是光特异性的, 在黑暗条件下无效. 说明DNQX至少能够部分阻断光诱导的幼苗下胚轴伸长抑制和叶绿素合成[2]. BMAA (β-Methylamino-L-alanine, 一种动物 iGluR激动剂), 在光照条件下可以抑制拟南芥幼苗根的生长和子叶张开, 并刺激下胚轴伸长2~3倍. 与DNQX处理相似, 这种效应同样是光特异性的. 同时, 该效应可以被加入的外源谷氨酸消除[19], 推测AtGLRs基因在植物体内可能起到一种光信号传导的作用.4.3 碳氮代谢的调节和ABA关系在缺N (仅含3%蔗糖)培养基中, 反义AtGLR1.1 (antiAtGLR1.1)拟南芥植株与野生型对照, 其种子萌发受到抑制. 在外加N后, antiAtGLR1.1种子的萌发恢复正常, 说明不同的C/N比能够影响种子的萌发, 高 C/N 比抑制了 antiAtGLR1.1的种子萌发. ABA在碳氮代谢和种子萌发中起到重要作用, 在antiAtGLR1.1株系中, 与碳氮代谢相关的HXK1 (己糖激酶I)等多个关键酶转录降低; 而ABA1转录水平提高, 导致植株体内 ABA 的积累, 经测定antiAtGLR1.1株系的内源ABA浓度较野生型要高8倍, 说明AtGLR1.1参与了C/N的代谢, 并通过对ABA合成的调控来控制种子萌发[20-21].4.4 环境胁迫对拟南芥AtGLR3.4基因启动子融合GUS转基因植株提供外力刺激, 发现机械外力(如损伤、穿刺)能够诱导 AtGLR3.4表达量增加[22]. 冷胁迫能够诱导拟南芥细胞膜快速去极化和细胞外 Ca2+流入,导致胞质Ca2+浓度快速增加[23]. 通过定量RT-PCR分析, 发现冷处理 5min, 再放回正常环境 15min之后, AtGLR3.4基因转录水平达到最高, 但此效应能被质膜 Ca2+通道抑制剂 La3+阻断, 用一定浓度ABA 对实验材料进行处理, ABA并不影响AtGLR3.4基因的转录[22]. 推测AtGLR3.4基因在环境胁迫快速响应机制中的作用是基于Ca2+信号, 而与ABA无关.4.5 氨基酸信号的协同作用甘氨酸处理、甘氨酸与谷氨酸协同处理能够有效阻断动物iGluRs拮抗剂DNQX 诱导的拟南芥幼苗下胚轴延伸, 之后转基因拟南芥表达水母发光蛋白表明: 甘氨酸处理、甘氨酸与谷氨酸协同处理可分别诱导胞质Ca2+浓度瞬时增加, 而这2个诱导都能被DNQX抑制. 推测AtGLRs可能是一种类似动物NMDAr (NMDA Receptor)的受体, 可以接受谷氨酸和甘氨酸的协同作用[11].进一步的研究发现, 除谷氨酸外, 甘氨酸、丙氨酸、天冬酰胺、半胱氨酸、丝氨酸和谷胱甘肽的处理都能诱导野生型拟南芥幼苗根尖细胞膜瞬间去极化和细胞外Ca2+大量流入, 而这一效应在突变体glr3.3中大幅度降低, 因此推测这6种氨基酸都能作为AtGLR3.3的激动剂[12]. 随后对野生型拟南芥幼苗的下胚轴使用上述 6种氨基酸中的任意2种进行先后处理(共36对处理), 发现这些氨基酸作为激动剂的作用并不是等效的, 例如谷氨酸引起的细胞膜去极化后的失敏现象不能被其他氨基酸信号解除, 而谷氨酸能够解除其他氨基酸信号引起的失敏现象, 由此推测GLRs离子通道可以分为A、B、C 3类: A类离子通道仅受谷氨酸激活和失敏; B类离子通道能够被丙氨酸、半胱氨酸、谷氨酸、甘氨酸激活和失敏; C类离子通道能被所有6种氨基酸激活和失敏. 通过对突变体glr3.3和突变体glr3.4的实验, 推测AtGLR3.3是3类离子通道的必要组成部分, 而AtGLR3.4是后2类通道的组成部分[24].4.6 其他高等植物中的GLRs在水稻OsGLR3.1基因的T-DNA插入短根突变体中, 因根尖分生组织的分裂能力受到阻碍, 细胞凋亡的增强导致短根, 说明OsGLR3.1基因与根尖的分生组织分裂和细胞发育密切相关, 参与调控根系的生长和发育[25]. 对小萝卜RsGluR基因的研究发现, 小萝卜GLRs是位于细胞膜上的谷氨酸门控的钙离子通道, 参与了受茉莉酸生物合成调控的病菌防御过程[18].因为异源表达系统构建不成功等原因, 拟南芥AtGLRs的功能研究至今没有突破性进展, 它们的生理功能、进化上意义也都没有确定, 然而通过电生理学、药理学、AtGLRs基因过度表达株系和反义AtGLR株系的研究, 越来越多证据表明, 植物的GLRs可能是一类特殊的GLRs受体, 对于它们的研究有可能开启探索“植物神经学”的大门.【相关文献】[1] Dingledine R, Borges K, Bowie D, et al. The glutamate receptor ion channels[J]. Pharmacological Reviews, 1999, 50:7-61.[2] Lam H M, Chiu J, Hsieh M H, et al. Glutamate receptor genes in plants[J]. Nature, 1998, 396:125-126.[3] Lacombe B, Becker D, Hedrich R, et al. The identity of plant glutamate receptors[J]. Science, 2001, 292:1486-1487. [4] Chiu J, Brenner E D, DeSalle R, et al. Phylogenetic and expression analysis of the glutamate receptor-like gene family in Arabidopsis thaliana[J]. Molecular Biology and Evolution, 2002, 19 (7):1066-1082.[5] Roy S J, Gilliham M, Berger B, et al. Investigating glutamate receptor-like gene co-expression in Arabidopsis thaliana[J]. Plant, Cell and Environment, 2008, 31(6):861-871. [6] Chiu J, DeSalle R, Lam H M, et al. Molecular evolution of glutamate receptors: Aprimitive signaling mechanism that existed before plants and animals diverged[J]. Molecular Biology and Evolution, 1999, 16(6):826-838.[7] Davenport R. Glutamate receptors in plants[J]. Annals of Botany, 2002, 90:549-557.[8] Oswald R E. Ionotropic glutamate receptor recognition and activation[J]. Advances in Protein Chemistry, 2004, 68:313-349.[9] Turano F J, Panta G R, Allard M W, et al. The putative glutamate receptors from plants are related to two superfamilies of animal neurotransmitter receptors via distinct evolutionary mechanisms[J]. Molecular Biology andEvolution, 2001, 18 (7):1417-1420. [10] Dennison K L, Spalding E P. Glutamate-gated calcium fluxes in Arabidopsis[J]. Plant Physiology, 2000, 124(4): 1511-1514.[11] Dubos C, Huggins D, Grant G H, et al. A role for glycine in the gating of plant NMDA-like receptors[J]. The Plant Journal, 2003, 35(6):800-810.[12] Qi Z, Stephens N R, Spalding E P. Calcium entry mediated by GLR3.3, an Arabidopsis glutamate receptor with a broad agonist profile[J]. Plant Physiology, 2006, 142(3):963-971.[13] Tapken D, Hollmann M. Arabidopsis thaliana glutamate receptor ion channel function demonstrated by ion pore transplantation[J]. Mol Biol, 2008, 383 (1):36-48.[14] Cho D, Kim S A, Murata Y, et al. De-regulated expression of the plant glutamate receptor homolog AtGLR3.1 impairs long-term Ca2+-programmed stomatal closure[J]. The Plant Journal, 2009, 58(3):437-439.[15] Turano F J, Muhitch M J, Felker F C, et al. The putative glutamate receptor 3.2 from Arabidopsis thaliana (AtGLR3.2) is an integral membrane peptide that accumulates in rapidly growing tissues and persists in vascular-associated tissues[J]. Plant Science, 2002, 163: 43-51.[16] Kim S A, Kwak J M, Jae S K, et al. Overexpression of the AtGluR2 gene encoding an Arabidopsis homolog of mammalian glutamate receptors impairs calcium utilization and sensitivity to ionic stress in transgenic plants[J]. Plant and Cell Physiology, 2001, 42(1):74-84.[17] 陈庆, 叶放, 宁永强, 等. 拟南芥 AtGLR1.3 和AtGLR3.3 启动子的 GUS 基因融合表达[J]. 宁波大学学报: 理工版, 2010, 23(3):1-5.[18] Kang S, Kim H, Lee H, et al. Overexpression in Arabidopsis of a plasma membrane targeting glutamate receptor from small radish increases glutamate-mediated Ca2+influx and delays fungal infection[J]. Molecules and Cells, 2006, 21(3):418-427.[19] Brenner E D, Martinez N, Clark A P, et al. Arabidopsis mutants resistant to S(+)-b-methyl-a,b-diaminopropionic acid, a cycadderived glutamate receptor agonist[J]. Plant Physiology, 2000, 124:1615-1624.[20] Kang J M, Turano F J. The putative glutamate receptor 1.1 (AtGLR1.1) functions as a regulator of carbon and nitrogen metabolism in Arabidopsis thaliana[J]. Proceedings ofthe National Academy of Sciences, 2003, 100(11):6872-6877.[21] Kang J M, Mehta S, Turano F J. The putative glutamate receptor 1.1 (AtGLR1.1) in Arabidopsis thaliana regulates abscisic acid biosynthesis and signaling to control development and water loss[J]. Plant and Cell Physiology, 2004, 45(10):1380-1389. [22] Meyerhoff O, Müller K, Roelfsema M R, et al. AtGLR3.4, a glutamate receptor channel-like gene is sensitive to touch and cold[J]. Planta, 2005, 222(3):418-427.[23] Lewis B D, Karlin-Neumann G, Davis R W, et al. Ca2+-activated anion channels and membrane depolarizations induced by blue light and cold in Arabidopsis seedlings[J]. Plant Physiol, 1997, 114(4):1327-1334.[24] Stephens N R, Qi Z, Spalding E P. Glutamate receptor subtypes evidenced by differences in desensitization and dependence on the GLR3.3 and GLR3.4 Genes[J]. Plant Physiol, 2008, 146(2):529-538.[25] Li J, Zhu S, Song X, et al. A rice glutamate receptor-like gene is critical for the division and survival of individual cells in the root apical meristem[J]. The Plant Cell, 2006,18(2):340-349.。
gpcr家族分类

gpcr家族分类GPCR(G protein-coupled receptors)又称七膜通道受体家族,是人类体内最大的受体家族,也是目前最有效的药物靶点之一,研究和应用广泛。
GPCR家族包含了信号通路复杂多样的受体,如视觉、嗅觉、味觉、心血管、神经、内分泌等等,而这些受体的重要性和复杂性也促使了人们对该家族的深入研究。
GPCR家族中的受体根据其序列上的同源性分为不同的类别,目前已知的GPCR分类体系主要包括以下几种:一、Rhodopsin-like受体家族。
此类受体具有7个膜通道跨膜区域,是最大的GPCR亚型,包括了与视觉相关的视杆细胞色素(rodopsin)、视锥细胞色素(cones)、视蓝细胞色素(melanopsin),以及多个结合神经递质(neurotransmitter)的GPCR受体(如α和β肾上腺素受体、ATP受体等)。
二、Secretin-like受体家族。
此类受体亚型包括了用于体内的激素信号传递的类胰岛素肽受体(insulin-like peptide receptors)、钙调神经肽受体(calcitonin receptor-like receptors),以及调节胰高血糖激素(GLP-1)等生长因子的受体。
此外,他们还包括了Parathyroid hormone receptor(PTH)家族的受体;三、Metabotropic glutamate受体家族。
此类受体家族是指通过谷氨酸作为神经递质的信号路径进行通讯的主解剖受体群,是视神经系统中的必要元素。
结构上,他们与Rhodopsin-like受体有一定的相似性,但其激活和信号传递机制略有不同,这也是其与Rhodopsin-like受体区分开来的重要因素。
四、Adhesion受体家族。
此类受体家族的重要功能涉及细胞间的结构和黏附性,同时也参与支配细胞的信号转导。
作为细胞膜的结构部件,Adhesion受体对于细胞—细胞之间的黏附非常重要,其在色素痣出现、组织发展、疾病转移中都有很重要的作用。
禾本科植物根系发育的分子机制研究进展

禾本科植物根系发育的分子机制研究进展任永哲;徐艳花;张庆琛;梁峰【摘要】植物根系的主要功能是从土壤中吸收养分和水分,并起到固定植株的功能.因此根系对于植物完成生命周期是至关重要的.以往的研究多集中在对模式植物拟南芥的根系上,近年来,人们开始对禾本科植物根系发育的分子机制开展研究并取得了一定的进展,一些科学家开始利用QTL定位的方法对调控植物根系性状的基因进行定位,对根系表型鉴定的方法也进行了很多改良.现有的研究结果表明,禾本科植物根系发育的分子机制与拟南芥既有相似之处,又存在一定的差异.对近几年来禾本科植物根系发育的分子机制方面取得的研究结果进行了综述,并对根系育种的重要性及其困难进行了探讨.%Plant roots are required for the acquisition of water and nutrients, and fixition of plant. The proper establishment of root system architecture is of vital importance to fulfill its functional requirements, particularly in agronomically important crops such as cereals, which account for 70% of food production worldwide. In recent years, studies on the molecular mechanisms regulating root development have also been initiated in monocot cereals which have been well studied in Arabidopsis in the past ten years. The achievements of root development in cereals were reviewed in this paper. The application of quantitative trait locus mapping in studying root development and the application of root trait information in breeding were also discussed.【期刊名称】《湖北农业科学》【年(卷),期】2011(050)012【总页数】5页(P2380-2384)【关键词】植物根系;禾本科植物;QTL;育种【作者】任永哲;徐艳花;张庆琛;梁峰【作者单位】商丘师范学院生命科学系,河南商丘476000;商丘师范学院生命科学系,河南商丘476000;商丘师范学院生命科学系,河南商丘476000;商丘师范学院生命科学系,河南商丘476000【正文语种】中文【中图分类】S184根系的主要功能是从土壤中获取矿物元素和水分,并起到固定植株的作用;根系还可以与多种微生物形成共生结构,以提高植物的养分效率和抗逆性;此外根系还有一些其他功能比如光合产物的储存、植物激素的合成、无性繁殖等。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Glutamate Receptor-Like Channel3.3Is Involved in Mediating Glutathione-Triggered Cytosolic Calcium Transients,Transcriptional Changes,and Innate Immunity Responses in Arabidopsis1[W][OA]Feng Li,Jing Wang,Chunli Ma,Yongxiu Zhao,Yingchun Wang,Agula Hasi,and Zhi Qi*College of Life Sciences,Inner Mongolia University,Hohhot010021,ChinaORCID ID:0000-0002-0224-5701(Z.Q.).The tripeptide reduced glutathione(GSH;g-glutamate[Glu]-cysteine[Cys]-glycine)is a major endogenous antioxidant in both animal and plant cells.It also functions as a neurotransmitter mediating communication among neurons in the central nervous system of animals through modulating specific ionotropic Glu receptors(GLRs)in the membrane.Little is known about such signaling roles in plant cells.Here,we report that transient rises in cytosolic calcium triggered by exogenous GSH in Arabidopsis (Arabidopsis thaliana)leaves were sensitive to GLR antagonists and abolished in loss-of-function atglr3.3mutants.Like the GSH biosynthesis-defective mutant PHYTOALEXIN DEFICIENT2,atglr3.3showed enhanced susceptibility to the bacterial pathogen Pseudomonas syringae pv tomato DC3000.Pathogen-induced defense marker gene expression was also decreased in atglr3.3mutants. Twenty-seven percent of genes that were rapidly responsive to GSH treatment of seedlings were defense genes,most of which were dependent on functional AtGLR3.3,while GSH suppressed pathogen propagation through the AtGLR3.3-dependent pathway.Eight previously identified putative AtGLR3.3ligands,GSH,oxidized glutathione,alanine,asparagine,Cys,Glu,glycine,and serine,all elicited the AtGLR3.3-dependent cytosolic calcium transients,but only GSH and Cys induced the defense response,with the Glu-induced AtGLR3.3-dependent transcription response being much less apparent than that triggered by GSH.Together,these observations suggest that AtGLR3.3is required for several signaling effects mediated by extracellular GSH,even though these effects may not be causally related.Reduced glutathione(GSH;g-Glu-Cys-Gly)is the most abundant short-chain peptide in cells and is enzymati-cally synthesized from Glu,Cys,and Gly.It is present in both intra-and extracellular compartments,although at least90%is found inside the cell.Depending on the cell type,glutathione levels have been estimated to range from1to10m M,90%of which is in reduced form,thus representing the major pool of endogenous nonprotein thiols within organisms(Noctor et al.,2011).Its Cys sulfur group can perform reversible redox reactions to form disulfides,notably with another glutathione Cys residue to produce oxidized glutathione(GSSG),which is continuously recycled at high rates to GSH by glutathi-one reductase existing in the cytosol and other cellular organelles.Therefore,glutathione serves a central role in free radical scavenging,maintaining the thiol status of proteins,and functions as a general antioxidant protect-ing cell membranes against oxidative stress and DNA against radiation(for review,see Franco and Cidlowski, 2009;Schmidt and Dringen,2012).In plants,GSH typically accumulates in cells to milli-molar concentration.Intracellular GSH is essential for development and reactive oxygen species scavenging as well as oxidative signaling during stress responses(Foyer and Noctor,2011;Noctor et al.,2012).Arabidopsis(Arab-idopsis thaliana)knockout mutants completely deficit in GSH biosynthesis show either embryo-or seedling-lethal phenotypes(Cairns et al.,2006;Pasternak et al.,2008), demonstrating its indispensable role in plant develop-ment.Other Arabidopsis mutants that have various de-grees of reduction in endogenous GSH content show altered expression of stress-related genes(Ball et al.,2004; Mhamdi et al.,2010;Han et al.,2013a,2013b),enhancing sensitivity to excessive zinc(Shanmugam et al.,2012)and cadmium(Jozefczak et al.,2012)as well as pathogens (Parisy et al.,2007;Dubreuil-Maurizi and Poinssot,2012). In animal cells,GSH not only functions as an essential antioxidant as in plant cells,but GSH in the extracellular space of neurons in the central nervous system can act like a neurotransmitter or neuromodulator through specific interactions with Glu receptors(Levy et al.,1991; Juurlink,1999;Dringen,2000;Oja et al.,2000;Shaw et al.,2001;Aoyama et al.,2008).Glu receptors are either ionotropic(iGLRs)or metabotropic.iGLRs form ionic1This work was supported by the National Natural Science Foun-dation of China(grant no.31171364),the Program for New Century Excellent Talents in University from the Ministry of Education,the Major Basic Science Research Open Program from the Inner Mongolia Science and Technology Department,and a Startup Grant from Inner Mongolia University(to Z.Q.).*Corresponding author;e-mail qizhi@.The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors()is: Zhi Qi(qizhi@).[W]The online version of this article contains Web-only data.[OA]Open Access articles can be viewed online without a subscrip-tion./cgi/doi/10.1104/pp.113.217208cation channels permeable to Ca2+,Na+,and K+and are named after their agonists N-methyl-D-Asp(NMDA), 2-amino-3-hydroxy-5-methyl-4-isoxazolepropionate,and kainite(Flores-Soto et al.,2012).Metabotropic Glu re-ceptors are coupled to G proteins and functionally linked either to the formation of inositol phosphates and diacylglycerol or to the metabolism of cyclic nucleotides (Loane et al.,2012).The molecular structure of GSH tends to make it neuroactive,because all of its three individual amino acid residues can either function as neurotransmitters or interfere with glutamatergic neuro-transmission,with Glu and Gly being excitatory and in-hibitory neurotransmitters,respectively(Hnasko and Edwards,2012).Furthermore,Cys is neurotoxic at high concentrations(Slivka and Cohen,1993).GSH can stimulate NMDA receptor-dependent membrane depo-larization and Ca2+entry into rat neurons(Leslie et al., 1992;Janáky et al.,1993;Ogita et al.,1995;Varga et al., 1997),cultured hippocampal neurons(Chin et al.,2006), and pig cerebral cortical synaptic membranes(Janáky et al.,2000).To the best of our knowledge,thefirst report of a po-tentially similar signaling role of extracellular GSH in plants to that found in the animal central nervous system was a study with tobacco(Nicotiana tabacum)expressing the Ca2+reporter luminescence protein aequorin in the cytosol(Gomez et al.,2004).The authors reported that exogenous application of5m M GSH induces cytosolic Ca2+([Ca2+]cyt)transient elevation in the leaf(Gomez et al.,2004).In Arabidopsis roots,our previous study also demonstrated that1m M GSH can induce strong[Ca2+]cyt elevation(Qi et al.,2006).Furthermore,GSH can induce AtGLR3.3-dependent membrane depolarization(Qi et al., 2006).AtGLR3.3is one of20Glu receptor genes in Arab-idopsis(Lacombe et al.,2001).The predicted protein structure of AtGLRs includes conserved transmembrane and extracellular ligand binding domains of iGLRs and G protein-coupled receptors(GPCRs;Chiu et al.,1999; Turano et al.,2001).Different plant GLR members are suggested to play roles in various physiological pro-cesses,including mineral nutrient homeostasis(Kim et al., 2001;Dubos et al.,2003;Kang et al.,2006;Aouini et al., 2012),carbon/nitrogen balance(Kang and Turano,2003), root development(Li et al.,2006),stomata movement (Cho et al.,2009),abscisic acid sensing(Kang et al.,2004), gravitropism sensing(Miller et al.,2010),and pollen tube elongation(Michard et al.,2011).Recently,pharmaco-logical approaches produced evidence that AtGLRs could be involved in innate immunity response in Arabidopsis seedlings(Kwaaitaal et al.,2011,2012).In light of the established roles of GSH in plant defense responses (Ferrari et al.,2003;Parisy et al.,2007;Schlaeppi et al., 2008)and the requirement of AtGLR3.3for GSH-induced signaling events in the roots(Qi et al.,2006),we hy-pothesized that GSH could mediate innate immunity responses in the leaf through AtGLR3.3-dependent pathways.This study reports evidence that strongly supports this notion and gives rise to several intriguing new questions about this novel signaling cascade in the defense response.RESULTSExogenous GSH Induces AtGLR3.3-Dependent Heterogeneous[Ca2+]cyt RiseTo investigate the signaling role of extracellular GSH in the leaf,we monitored how exogenous application of GSH([GSH]ext)affects[Ca2+]cyt in detached Arabidopsis leaves,using cytosolic expression of the Ca2+reporter protein aequorin.As observed previously(Qi et al.,2006), the background[Ca2+]cyt concentration was around0.1 m M(Fig.1).Immediately after the delivery of a10m M GSH solution into the detached leaf,a rise in[Ca2+]cyt was observed(Fig.1).Increasing the[GSH]ext to100and 1,000m M further promoted the response,which showed a strong concentration-dependent pattern(Fig.1).In our previous study with Arabidopsis roots,we demonstrated that1m M[GSH]ext induced membrane depolarization and that this effect largely depends on AtGLR3.3(Qi et al.,2006),one of20Glu receptor-like genes in Arabidopsis.We hypothesized that the ob-served[GSH]ext-induced[Ca2+]cyt rise in this study was also mediated by AtGLR3.3.As an initial test of this hypothesis,we pretreated the leaf with the iGLRs antagonists(2R)-amino-5-phosphonopentanoate(AP5) and6,7-dinitroquinoxaline-2,3-dione.Both ofthem,Figure1.[GSH]ext-induced concentration-dependent[Ca2+]cyt rise in the leaf.A,Representative recording curve of10,100,and1,000m M GSH-induced[Ca2+]cyt transient rise in a detached leaf.B,Averaged peak values with SE of the responses(n=5),which are significantly different to each other at P,0.01.The dash line indicates the back-ground[Ca2+]cyt.Li et al.especially AP5,signi ficantly suppressed the [GSH]ext -induced responses (Fig.2).We next introduced aequo-rin gene into the atglr3.3-1and atglr3.3-2knockout mutant backgrounds through cross pollination and monitored its response to [GSH]ext .The GSH-induced [Ca 2+]cyt rise observed in the wild type was completely absent in the mutants (Fig.3).We next used pharmacological approaches to investi-gate which Ca 2+mobility pathways contribute to the [GSH]ext -induced and AtGLR3.3-dependent [Ca 2+]cyt rise.Removal of extracellular Ca 2+and including 1m M Ca 2+chelator EGTA in the recording buffer signi ficantly,but not completely,suppressed 100m M GSH-induced [Ca 2+]cyt rise (Fig.2).This indicates that extracellular Ca 2+in flux through certain plasma membrane Ca 2+-permeable chan-nels partially,but not fully,contributes to the observed response.cAMP-gated calcium channels have been iden-ti fied in the plasma membrane of Arabidopsis leaf protoplasts (Ali et al.,2007).Alloxan can interrupt the cAMP-dependent Ca 2+in flux pathway by inhibiting adenylate cyclase activity and so blocking cAMP produc-tion.Pretreatment for 1h with 1m M alloxan signi ficantly inhibited the rise in [Ca 2+]cyt (Fig.2).Our previous study implicated Cyclic Nucleotide Gated Channel2(CNGC2)in mediating the [Ca 2+]cyt rise associated with innate im-munity (Ma et al.,2009).However,in this study,we found a normal response of the cngc2mutant expressing aequorin to the [GSH]ext (data not shown).Chelating extracellular Ca 2+did not abolish the GSH-induced [Ca 2+]cyt response (Fig.2),implicating Ca 2+re-lease from certain internal Ca 2+storage pools in the response.1,4,5-trisphosphate (IP3)-mediated Ca 2+release from internal Ca 2+pools has been demonstrated in Arab-idopsis guard cells (Tang et al.,2007).Neomycin can block the IP 3-mediated Ca 2+mobility pathway by inhibiting the activity of phospholipase C,which catalyzes pro-duction of IP 3from phosphatidylinositol 4,5bisphos-phate.One-hour 1m M neomycin pretreatment also signi ficantly suppressed the [GSH]ext -induced Ca 2+re-sponse (Fig.2).Taken together,these data demon-strated that the [GSH]ext -induced Ca 2+response fully depended on AtGLR3.3but was caused by increased Ca 2+mobility through various pathways.AtGLR3.3Mediates Part of the Early Transcriptional Response of the Leaf to [GSH]extLigand-induced [Ca 2+]cyt rise is often linked to down-stream transcriptional responses (Dodd et al.,2010).To focus on the early transcriptional responses to [GSH]ext and to establish the importance of AtGLR3.3for the GSH-mediated signaling cascades,we treated the wild-type and atglr3.3-2seedlings for 1h with either the control (CK)recording buffer or the buffer containing 100m M GSH.Two independent biological replicates were con-ducted for whole genome transcriptional pro filing anal-ysis with Agilent Arabidopsis microarrays.Under the CK condition,there were 35genes showing at least 3-fold differential expression between the mutant and the wild type (Supplemental Table S1).As expected,AtGLR3.3was the lowest expressed transcript in the atglr3.3mutant compared with the wild type,with 47-and 50-fold changes in the two biological replicates,respectively (Supplemental Table S1).GSH treatment of the wild type altered expression of 97genes,with at least 3-fold transcript abundance change (Fig.4A;Supplemental Tables S2and S3).ByFigure 2.Pharmacological study of the [GSH]ext -induced [Ca 2+]cyt transient response in the leaf.A,Representative recording curve of 100m M [GSH]ext -induced [Ca 2+]cyt rise in the CK and pretreated with 1m M various Ca 2+mobility pathway blockers as indicated.B,Averaged peak values of these responses with SE (n =4for EGTA and n =5for the rest).The asterisk stands for the SD to the CK at P ,0.01.The average background [Ca 2+]cyt was around 0.08m M.Figure 3.[GSH]ext -induced [Ca 2+]cyt response was absent in the atglr3.3mutant.Averaged recording curves with SE of 100m M [GSH]ext -induced [Ca 2+]cyt rise in the leaf cells of the ecotype Columbia (Col-0)and atglr3.3-1and atglr3.3-2mutants expressing aequorin (n =10for each).Glutathione and Glutamate Receptor in Innate Immunitycomparing the genes modulated by GSH in the atglr3.3mutant,we found that 70(Supplemental Table S2)of the 97genes showed no signi ficant change in the mutant.The remaining 27genes showed the same expression trends in response to GSH in both the wild type and the mutant (Fig.4A;Supplemental Tables S3and S4).Thus,this expression pro filing analysis indicated that 72%(70/97)of the genes rapidly modulated by GSH in the wild type were dependent on the AtGLR3.3pathway and 28%(27/97)independent of it.To validate the transcriptional pro filing data set,inde-pendent biological samples were produced,and eight genes selected from the list of genes regulated by GSH in the wild type were chosen for quantitative reverse transcript (qRT)-PCR analysis (Supplemental Tables S2and S3).All eight genes showed signi ficantly increased expression in response to GSH,an effect that was signi ficantly attenuated in the atglr3.3-2mutant back-ground (Supplemental Fig.S1).The trends of gene expression in Supplemental Figure S1were consistent with those revealed by the microarray (Supplemental Table S2).These observations provide further evi-dence that early transcriptional responses induced by GSH are at least partially acting through AtGLR3.3.GSH-Regulated AtGLR3.3-Dependent Defense and Signaling-Related GenesTo explore which cellular pathways were regulated by GSH in an AtGLR3.3-dependent manner,we performed categorization of these genes by Gene Ontology anno-tation analysis combined with manual checking of available literature associated with each of the 70genes in Supplemental Table S2through searching The Arabi-dopsis Information Resource ().This analysis showed that 27%,20%,20%,20%,and 16%of these genes were functionally categorized as Defense,Signaling,Unknown,Transcription Factor,and Others,respectively (Fig.4B;Supplemental Table S2).There were eight genes that were common to thecategories Defense and Transcription Factor.All of the genes in the Defense category have been demonstrated using corresponding mutants to play roles during de-fense responses or found to be induced by various pathogens at transcriptional level (Table I;Supplemental Table S2).These genes notably include the essential sal-icylic acid (SA)biosynthesis gene ISOCHORISMATE SYNTHASE1(ICS1;Wildermuth et al.,2001)and its key regulator SYSTEMIC ACQUIRED RESISTANCE DEFICIENT1(SARD1;Zhang et al.,2010)as well as the transcription factors CALMODULIN BINDING PROTEIN60-LIKEG (CBP60G )(Wang et al.,2009),WRKY33,and WRKY46(Birkenbihl et al.,2012;Hu et al.,2012;Moreau et a.,2012),all of which are essential genes for medi-ating defense response of Arabidopsis.Genes in the Signaling category include a kinase (At1g67000),receptor-like proteins RLP7and RLP20,cal-cium homeostasis-related proteins such as Glu receptors AtGLR1.2,AtGLR2.8,and AtGLR2.9,and CALMODULIN LIKE37.These proteins could be involved in relaying the GSH-induced AtGLR3.3-dependent rise in [Ca 2+]cyt tran-sient to downstream signaling cascades (Fig.3).In addi-tion,this category includes At3g28580,an AAA-ATPase,which was up-regulated 6-fold by GSH in an AtGLR3.3-dependent manner (Supplemental Table S2).This com-ponent has recently been reported to play a central role in mediating plant responses to reactive oxygen species in conditions of environmental stress (Simkováet al.,2012).Taken together,this analysis demonstrated that the AtGLR3.3-dependent early transcriptional responses of Arabidopsis to [GSH]ext primarily involved cellular de-fense and signaling,functional categories that account for 47%of the modulated genes (Supplemental Table S2).AtGLR3.3Is Required for GSH-Mediated Innate Immunity Response in Arabidopsis LeafArabidopsis mutants defective in GSH biosynthesis show altered defense responses to variouspathogensFigure 4.Overview of the GSH-and Glu-mediated AtGLR3.3-dependent gene expression.A,Number of overlapping genes modulated by GSH and Glu in the ecotype Columbia (Col-0)and atglr3.3mutant.B,Functional category of the genes mod-ulated by GSH with AtGLR3.3dependence.C,Functional category of the genes modulated by Glu with AtGLR3.3dependence.The numbers in and the percentage symbol outside the pie chart indicate the number and percentage of the genes in the category,respectively.U,Unknown;TF,Transcription Factor;S,Signaling;O,Others;D,Defense.Li et al.(Glazebrook and Ausubel,1994;Roetschi et al.,2001; van Wees et al.,2003;Bohman et al.,2004;Parisy et al., 2007;Dubreuil-Maurizi and Poinssot,2012).Innate immunity responses involve complicated signaling networks.We hypothesized that the atglr3.3mutants may have overlapping defense phenotypes with Arab-idopsis mutants,such PHYTOALEXIN DEFICIENT2-1 (pad2-1),defective in GSH biosynthesis.To test its pathogen phenotype,we inoculated mature leaves of the wild type and two allelic knockout mutants(atglr3.3-1 and atglr3.3-2)with the hemibiotrophic model bacterial pathogen Pseudomonas syringae pv tomato DC3000(Pst DC3000).Pathogen growth was monitored at0,1,2,and 3d post inoculation(DPI).Figure5showed that the two atglr3.3mutants had significantly higher susceptibility to the pathogen infection than the wild type,indicating that AtGLR3.3has a role in innate immunity of Arabi-dopsis.To validate our experimental system,we reex-amined the pathogen phenotype of pad2-1along with the wild type.Supplemental Figure S3showed that the pathogen population per unit in the mutant at2DPI was significantly higher than that of the wild type,which was consistent with the previous report(Glazebrook and Ausubel,1994).To investigate the role of AtGLR3.3in the response, we monitored expression of two defense marker genes, PATHOGENESIS-RELATED GENE1(PR1)and WRKY33,in response to pathogen infection in both the wild type and the mutants at0,12,24,and48h post inoculation. The pathogen-induced expression of the defense marker genes was significantly attenuated in both allelic atglr3.3 loss-of-function mutants(Fig.6).To investigate the physiological significance of the GSH-mediated AtGLR3.3-dependent defense gene expression(Fig.4;Table I),we next examined the effect of[GSH]ext on pathogen propa-gation inside the leaves of the wild type and the mutants. One day prior to the pathogen infiltration,the leaves were infiltrated with the CK buffer or the CK buffer con-taining100m M GSH.On the day after,all leaves were infiltrated with the pathogen suspension at53105 colony-forming units(cfu)mL–1.The GSH pretreatment significantly suppressed pathogen propagation in the wild-type leaf.However,suppression of propagation by[GSH]ext was not evident in the atglr3.3mutants(Fig.5).Thus,[GSH]ext promotes innate immunity responses in the leaf through AtGLR3.3-dependent pathways.Ligand Specificity of AtGLR3.3in the InnateImmune ResponseOur previous study with Arabidopsis root and hy-pocotyl cells suggests that there are up to eight puta-tive ligands of AtGLR3.3:GSH,GSSG,Ala,Asn,Cys, Gly,Glu,and Ser(Qi et al.,2006;Stephens et al.,2008). Among these,Glu plays a dominant role(Stephens et al.,2008).Given the results shown in Figure5,we hypothesized that if any of the eight putative ligands are true biological ligands of AtGLR3.3in vivo,they should be able to evoke AtGLR3.3-dependent innate immunity response in the leaf by exogenous applica-tion,in a similar fashion to GSH(Figs.3–5).To test this hypothesis,we infiltrated the leaf of the wild type and two alleles of mutants with CK or CK solution con-taining100m M GSSG,Ala,Asn,Cys,Gly,and GluTable I.Defense genes induced by GSH in ecotype Columbia but not glr3.3-2Gene ID FC01a FC02a Average Annotation Reference Having confirmative role in defenseAT5G1332017.9 6.112GRETCHEN HAGEN3.12;SA biosynthesis Okrent et al.,2009AT2G4640015.7 4.910.3WRKY46transcription factor Hu et al.,2012AT1G3396012.537.75AVIRULENCE-INDUCED GENE1Reuber and Ausubel,1996 AT1G74710 5.8 4.8 5.3Isochorismate Synthase1;SA biosynthesis Wildermuth et al.,2001 AT1G72520 3.3 6.6 4.95Lipoxygenase4;jasmonic acid biosynthesis Acosta and Farmer,2010 AT2G35980 5.1 3.9 4.5Yellow-leaf-specific gene9Po-Wen et al.,2013AT5G26920 4.4 3.4 3.9CBP60G Wang et al.,2009AT1G73805 3.6 3.5 3.55SARD1;SA biosynthesis Zhang et al.,2010AT2G38470 3.4 3.4 3.4WRKY33transcription factor Birkenbihl et al.,2012 Having putative role in defenseAT5G3697014.823.118.95HAIRPIN-INDUCED1LIKE25;induced by pathogens Varet et al.,2002AT5G47850713.210.1CRINKLY44related4;protein kinase;response to insect Little et al.,2007AT1G212407.5 3.3 5.4Wall associated kinase3;induced by SA He et al.,1999AT2G18060 3.5 6.3 4.9NAC(NAM,ATAF1/2and CUC2)domain containing protein37;induced by chitinLibault et al.,2007AT3G50930 3.7 5.7 4.7ATPases associated with diverse cellular activities type;inducedby bacterial pathogenNavarro et al.,2004AT3G232303 6.3 4.65Ethylene response factor98;induced by pathogen and jasmonic acid McGrath et al.,2005AT2G43000 3.7 5.1 4.4NAC domain containing protein42;camalexin biosynthesis Saga et al.,2012AT1G145404 3.8 3.9Peroxidase4;induced by oxidative stress and fungus infection Rasul et al.,2012AT1G18570 3.2 4.1 3.65MYB51transcription factor;induced by pathogen Maekawa et al.,2012 AT4G330504 3.1 3.55Embryo sac development arrest39;response to chitin and virus Libault et al.,2007a Fold changes of two replicates.Glutathione and Glutamate Receptor in Innate Immunity1d before the pathogen inoculation.We also included Asp,a nonexciting amino acid,as a control.At 2DPI,consistent with the previous results (Fig.5),the atglr3.3mutants showed enhanced bacterial growth (Fig.7).Among the compounds tested,only Cys and Asp were able to act similarly to GSH to signi ficantly suppress pathogen propagation in the wild-type leaf (Fig.7).The suppressing effects of Cys,but not Asp,were an-nulled in the atglr3.3loss-of-function mutants (Fig.7).Thus,of the two amino acids,only the suppressive effect of Cys acts through an AtGLR3.3-dependent pathway,similar to GSH.The Eight Putative Ligands,but Not Asp,Induce an AtGLR3.3-Dependent [Ca 2+]cyt RiseIt was unexpected that the eight ligands and Asp would have distinct effects on the plant ’s innate im-munity responses (Figs.5and 7).We hypothesized that the difference could be related to (1)varying ability to initiate the transient rise in [Ca 2+]cyt and (2)different dependence on AtGLR3.3in the leaves.We therefore examined effects of seven ligands and Asp on the [Ca 2+]cyt in the leaf cells of the wild type and atglr3.3mutants.All the tested ligands,except Asp but including Cys,evoked signi ficant [Ca 2+]cyt re-sponses (Fig.8).These were of varying amplitudes,but all were dependent on AtGLR3.3(Fig.8).This result demonstrated that innate immunity responses triggered by the ligands GSH and Cys are not sim-ply correlated with their ability to induce [Ca 2+]cyt transients.The Glu-Mediated Early Transcriptional Response Is Distinct to That Induced by GSHGlu is the principle exciting amino acid that acti-vates GLRs in the central nervous system (Flores-Soto et al.,2012).It also evokes strong [Ca 2+]cyt responses in the leaf (Fig.8A),which are at least comparable with,if not stronger than,those induced by GSH (Figs.2and 3).However,Glu and GSH had essentially distinct effects on the plant defense responses.Most notably,Glu had no suppressive effect on the pathogen prop-agation (Figs.5and 7).To further explore the in fluence of the two com-pounds,we compared Glu-elicited transcriptomic re-sponses with those induced by GSH (Fig.4B;Table I;Supplemental Table S2).In total,treatment of wild-type leaves with 100m M Glu modulated expression of 70genes,69of which showed no signi ficant changes in response to Glu in the atglr3.3mutant and can therefore be categorized as Glu-modulated AtGLR3.3-dependent genes (Fig.4A;Supplemental Tables S5,S6,and S7).Among the 69genes,only six genes were common to the GSH-modulated AtGLR3.3-dependent group (Fig.4A;Supplemental Tables S2and S5).Functional catego-rization of the 69Glu-modulated AtGLR3.3-dependent genes showed that 37%,16%,23%,3%,and 6%of these genes were classi fied as Unknown,Others,Signaling,Transcription Factor,and Defense,respectively (Fig.4C).The Defense section only included four genes:PR1,FLAVIN-DEPENDENT MONOOXYGENASE1(Mishina and Zeier,2006),CELL WALL-ASSOCIATED KINASE1,and CELL WALL-ASSOCIATED KINASE3(Brutus et al.,2010;De Lorenzo et al.,2011).Hence,it wasstrikinglyFigure 5.Role of AtGLR3.3in the innate immunity and GSH-triggered defense response in the leaf.A,Leaves of ecotype Columbia (Col-0)and atglr3.3-1and atglr3.3-2mutants were infiltrated with either CK solution or the solution containing 100m M GSH 1d before the inoculation of Pst DC3000at 53105cfu mL –1.The pathogen proliferation was monitored at 0,1,and 2DPI.Values presented are average with SE from at least four leaves from different plants.At 1,2,and 3DPI,there is SD between the Col-0and two mutants at P ,0.01.Two asterisks indicated significance at P ,0.01.Five independent experiments were conducted and showed comparable results.B,Leaf symptoms of the Col-0,atglr3.3-1,and atglr3.3-2plants preinfiltrated with CK solution or the solution containing 100m M GSH 3DPI with Pst DC3000.C,Same leaves in B were kept 1d longer,e.g.4DPI,in a sealed agar plate.Li et al.different from the transcriptomic patterns elicited by GSH in an AtGLR3.3-dependent manner (Fig.4B).Taken together,these results demonstrate that al-though GSH and Glu induce comparable AtGLR3.3-dependent [Ca 2+]cyt responses,they induce distinct downstream early transcription responses.Crucially,only GSH was able to induce expression of a signi fi-cant proportion of defense-related genes,which could explain the speci fic capacity of GSH to suppress path-ogen propagation in the leaves (Fig.5).DISCUSSIONSince the discovery of AtGLRs in Arabidopsis ge-nome (Lam et al.,1998),there has been considerable interest in deciphering their physiological functions (Kudla et al.,2010).Previously,in root cells,we dem-onstrated that AtGLR3.3is required for the membranedepolarization induced by eight putative ligands,in-cluding GSH,GSSG,Ala,Asn,Cys,Glu,Gly,and Ser (Qi et al.,2006).It is possible that different AtGLRs expressed in different tissue and under various con-ditions show different ligand speci ficity.The first evi-dence to support this notion was the finding that D -Ser,but not Glu and Gly,activates AtGLR1.2-dependent Ca 2+conductance in the plasma membrane of the pollen tube (Michard et al.,2011).Among theeightFigure 6.Pst DC3000-induced defense market gene expression in ecotype Columbia (Col-0)and atglr3.3mutants.Pst DC3000at 53105cfu mL –1was infiltrated into the leaves of Col-0,atglr3.3-1,and atglr3.3-2.The leaves were harvested at 0,12,24,and 48h post in-fection (HPI)for PR1(A)and WRKY33(B)gene expression analysis with qRT-PCR.The transcript abundance was normalized with refer-ence gene Tubulin2in Col-0at 0h post infection.Values presented are average with SE from three independent experiments.The asterisk in-dicated the SD between the mutants and Col-0at same time point at P ,0.05.Figure 7.Pst DC3000infection assay of ecotype Columbia (Col-0)and atglr3.3with putative ligands and Asp.The experiment procedure and analysis methods are same as those in Figure 5.Pst DC3000prolifer-ation in these leaves was monitored at 0(A)and 2DPI (B)preinfiltrated with either CK solution or the solution containing 100m M GSSG,Glu,Asp,Ala,Cys,and Asn.C,The same assay was conducted separately for Ser and Gly.At 2DPI in B and C,except for Asp,the values of atglr3.3-1and atglr3.3-2were significantly higher than that of Col-0at P ,0.01.The asterisk in B indicated there was SD between the treat-ment and the CK for the corresponding genotype at P ,0.01.Glutathione and Glutamate Receptor in Innate Immunity。