基因转录课件
合集下载
第十三章基因的转录、转录后加工及逆转录ppt课件
◆真核生物启动子 (1)DNA序列在转录起始点的5’端区(上游 区)(2)-25bp :TATA盒(Hogness box) (3)-90bp :GC盒 (4)-70bp :CAAT盒
-90
-70
GC
CAAT
RNA聚合酶Ⅱ催化的转录起始
RNA聚合酶Ⅱ催化各种前体mRNA的合 成
需要多种TF参与:TFⅡA-J
第一节 参与转录的酶
RNA聚合酶——依赖DNA的RNA聚合酶 (DNA-dependent RNA polymerase,DDRP)
以DNA为模板,催化2个游离的NTP 形成3’,5’-磷酸二酯键
一、原核生物RNA聚合酶
1、大肠埃希菌RNA聚合酶的组成 (1)全酶(holoenzyme)
由4种(5个)亚基α2ββ’σ组成 (2)核心酶(core enzyme)
作用位置
步骤1 步骤2
200~250
(3)真核生物mRNA转录后加工—剪接
内含子
外显子
DNA
hnRNA
●剪接所需条件
snRNA (U1-U6) + 蛋白质 (核内小分子核酸)
多种snRNP (核内小核蛋白颗粒)
多种snRNPs装配成
剪接体 (参与剪接过程)
4、RNA编辑(RNA editing)
二.真核生物RNA转录后的加工 1、rRNA转录后的加工
真核生物rRNA 的基因
(rDNA)
转录产物
成簇纵列串联排列
高度重复序列DNA
核质:(Ⅲ)--不需加 工
5s rRNA
核仁:(Ⅰ)--加工
5.8s rRNA 28s rRNA 18s rRNA
rDNA 内含子
基因间隔
真核生物基因的转录

(4)上游元件的多样性
Octamer
CAAT
GC
TATA
Startpoint
SV40 early
胸苷激酶 Thymidine kinase
Histone H2B
-140 -120 -100 -80 -60 -40 -20
没有哪一种上游元件是所有启动子所共同必需的
TF II B
(3) TFIIB 覆盖靠近起始点的启动位置,C端与TFIID和DNA的复合物结合,N-端与TFⅡF协同作用募集RNA聚合酶II TFIIB与TFIID结合,并为RNA聚合酶结合起一个桥梁作用。
(4)与RNA聚合酶与TFIIF相连的复合体结合
TF II F
Pol II
TF II F 结合Pol II并带向启动子; 两个亚基: RAP74(ATP依赖性解旋酶),可能参与DNA 双链的溶解 RAP30(与细菌因子有同源性),与RNA 聚合酶Ⅱ紧密结合
01
03
02
3、tRNA基因转录的起始
Pol III
TF III C
boxB
boxA
TF III B
(二)5S rRNA 基因的转录
特点:串连排列,形成基因簇 (是唯一单独被转录的rRNA亚基)
5S rRNA 基因:
C框 ;A框
启动子:
转录因子:
TFIIIB: TBP + BRF + B//
TFIIIA :结合位点为C box 。
(5)TF II E 扩大DNA覆盖区至+30
TF II E
TF II H 和TF II J加入复合物
TF II H 有多种酶活性,包括ATP酶、解旋酶、和可使Pol II 的CTD 磷酸化的激酶活性。
第八章 真核基因转录调控(共84张PPT)

(一) 基因表达是基因经过转录、翻译、产生有生物活 性的蛋白质的整个过程。同原核生物一样,转录 依然是真核生物基因表达调控的主要环节。
但真核基因转录发生在细胞核(线粒体基因的转录 在线粒体内),翻译则多在胞浆,两个过程是分开 的,因此其调控增加了更多的环节和复杂性,转 录后的调控占有了更多的分量。
• (二)真核基因的转录与染色质的结构变化相 关。
❖ ④增强子的作用机理虽然还不明确,但与其他顺 式调控元件一样,必须与特定的蛋白质结合后才 能发挥增强转录的作用。
❖ 增强子一般具有组织或细胞特异性,许多增强子只在 某些细胞或组织中表现活性,是由这些细胞或组织中 具有的特异性蛋白质因子所决定的。
• ①影响模板附近的DNA双螺旋结构,导 致DNA双螺旋弯折或在反式因子的参与 下,以蛋白质之间的相互作用为媒介形 成增强子与启动子之间“成环”连接,活 化基因转录;
• 实验表明这段序列甲基化可使其后的基因不能转录,甲基 化可能阻碍转录因子与DNA特定部位的结合从而影响转录
。
• 如果用基因打靶的方法除去主要的DNA甲基化酶,小鼠的 胚胎就不能正常发育而死亡,可见DNA的甲基化对基因表 达调控是重要的。
• 由此可见,染色质中的基因转录前先要有一个被激活的过程, 但目前对激活机制还缺乏认识。
• 真核启动子一般包括转录起始点及其上游约100- 200bp序列,包含有若干具有独立功能的DNA序列元 件,每个元件约长7-30bp。
• 最常见的哺乳类RNA聚合酶Ⅱ启动子中的元件
哺乳类RNA聚合酶Ⅱ启动子中的元件序列
元件名称 共同序列
名称
结合的蛋白因子 分子量 结合DNA长度
TATAbox TATAAAA
• 三、真核基因转录水平的调控
但真核基因转录发生在细胞核(线粒体基因的转录 在线粒体内),翻译则多在胞浆,两个过程是分开 的,因此其调控增加了更多的环节和复杂性,转 录后的调控占有了更多的分量。
• (二)真核基因的转录与染色质的结构变化相 关。
❖ ④增强子的作用机理虽然还不明确,但与其他顺 式调控元件一样,必须与特定的蛋白质结合后才 能发挥增强转录的作用。
❖ 增强子一般具有组织或细胞特异性,许多增强子只在 某些细胞或组织中表现活性,是由这些细胞或组织中 具有的特异性蛋白质因子所决定的。
• ①影响模板附近的DNA双螺旋结构,导 致DNA双螺旋弯折或在反式因子的参与 下,以蛋白质之间的相互作用为媒介形 成增强子与启动子之间“成环”连接,活 化基因转录;
• 实验表明这段序列甲基化可使其后的基因不能转录,甲基 化可能阻碍转录因子与DNA特定部位的结合从而影响转录
。
• 如果用基因打靶的方法除去主要的DNA甲基化酶,小鼠的 胚胎就不能正常发育而死亡,可见DNA的甲基化对基因表 达调控是重要的。
• 由此可见,染色质中的基因转录前先要有一个被激活的过程, 但目前对激活机制还缺乏认识。
• 真核启动子一般包括转录起始点及其上游约100- 200bp序列,包含有若干具有独立功能的DNA序列元 件,每个元件约长7-30bp。
• 最常见的哺乳类RNA聚合酶Ⅱ启动子中的元件
哺乳类RNA聚合酶Ⅱ启动子中的元件序列
元件名称 共同序列
名称
结合的蛋白因子 分子量 结合DNA长度
TATAbox TATAAAA
• 三、真核基因转录水平的调控
第五章转录ppt课件

一、E.coli RNA聚合酶
E.coli RNA聚合酶由5个亚基组成(5个 多肽链),即α2ββ′σ
αββ′σ四个亚基的分子量分别为36.5KDa、 150 KDa、160KDa和82 KDa,整个酶 分子的分子量为465KDa
分别是基因rpoA、rpoB、rpoC和rpoD的
产物
RNA聚合酶Ⅲ也存在于核质中,其功 能是合成tRNA和5S rRNA以及转录 Alu序列
在细胞质中也能发现一些RNA聚合酶 Ⅲ,它是从细胞核中渗漏出来的。
三种主要的RNA聚合酶的分子量都在 500 KDa左右(14S-15S),每种酶分子 含有两个大亚基和4~8个小亚基,每 个小亚基的分子量为10 KDa-90KDa
这些名称最早是依据它们从DEAE-纤维素柱上洗脱的 先后顺序而定出来的。后来发现不同生物的三种RNA聚 合酶的洗脱顺序并不相同,因而改用三种不同的RNA聚 合酶对于α-鹅膏蕈碱(α-amanitine)的敏感性不同来 进行区别。RNA聚合酶I基本不受α-鹅膏蕈碱的抑制, 在大于10-3 mol/L时才表现出轻微的抑制作用;RNA聚 合酶Ⅱ对于α-鹅膏蕈碱最为敏感,在10-9-10-8mol/L 浓度下就会被抑制;RNA聚合酶Ⅲ的敏感性介于RNA聚 合酶Ⅰ和Ⅱ之间,在10-5-10-4 mol/L时表现抑制作用。
与RNA聚合酶相结合的一个很小的蛋白质(MW= 10KDa),叫做ω亚基,其功能尚不清楚,有人认 为ω亚基对于RNA聚合酶的结构和功能没有太大的 影响。
原核生物的 RNA聚合酶
亚基 分子量
36512
150618
155613
70263
功能
决定哪些基因被转录 催化功能
转录因子PPT课件

(DNA-蛋白质) 转录抑制因子 共调节因子 (蛋白质-蛋白质)
特点:
至少含3个功能结构域:DNA结合功能域,转录活性功能域,其他转 录 因子结合功能域;
能识别并结合上游调控区中的顺式作用元件; 对基因表达有正性或负性调控作用,即激活或阻遏基因表达。
目前研究较广泛的有:识别TATA区的TFⅡD,识别CAAT 区的CTF,识别GGGCGG的SP1,识别热激蛋白启动区的 HSF。
通过从一侧逐段缺失 来确定启动子的边界. 当一段缺失不会阻碍 RNA合成, 而下一段缺 失使转录不再发生时, 那么我们可以确定启 动子的边界必然存在
于两者之间.
启动子克隆的几种方法
启动子的克隆对于构建基因工程载体,表达目的蛋白有着重要的意义。启动子 克隆的方法很多,从常用的利用启动子探针型载体筛选启动子到PCR方法的应 用,此后相继问世的一些基于PCR法的克隆启动子技术,像I-PCR、P-PCR、 SSP.PCR、YADE、TAIL-PCR等,为克隆启动子提供了更可靠,更合理的方 法。
并非任何细胞型特异的蛋白在任何情况下都起作用,而是取决于核心 启动子提供一个合适的环境来决定基因是被激活还是被抑制。
构建多个启动子连接多个基因的表达载体,虽然可以实现同时转入多 个基因的愿望,但这种载体一方面构建困难,另一方面如果引入的启 动子之间仅仅有90bp 的同源性顺序,导入生物体内,就会引起所谓的 基因表达”共抑制”现象,而使基因沉默。因此,将极性启动子人工改 造为高效双向表达的启动子,对于促进基因工程的进展具有重要意义 。
2.1.1 基本转录因子(general transcription factors)
是RNA聚合酶结合启动子所必需的一组蛋白因子, 决定三种RNA(mRNA、tRNA及rRNA)转录的类别,包 括 TF Ⅱ A、TF Ⅱ B、TF Ⅱ D、TF Ⅱ E、TF Ⅱ F、TF Ⅱ H、 TFⅡI等7种因子。
特点:
至少含3个功能结构域:DNA结合功能域,转录活性功能域,其他转 录 因子结合功能域;
能识别并结合上游调控区中的顺式作用元件; 对基因表达有正性或负性调控作用,即激活或阻遏基因表达。
目前研究较广泛的有:识别TATA区的TFⅡD,识别CAAT 区的CTF,识别GGGCGG的SP1,识别热激蛋白启动区的 HSF。
通过从一侧逐段缺失 来确定启动子的边界. 当一段缺失不会阻碍 RNA合成, 而下一段缺 失使转录不再发生时, 那么我们可以确定启 动子的边界必然存在
于两者之间.
启动子克隆的几种方法
启动子的克隆对于构建基因工程载体,表达目的蛋白有着重要的意义。启动子 克隆的方法很多,从常用的利用启动子探针型载体筛选启动子到PCR方法的应 用,此后相继问世的一些基于PCR法的克隆启动子技术,像I-PCR、P-PCR、 SSP.PCR、YADE、TAIL-PCR等,为克隆启动子提供了更可靠,更合理的方 法。
并非任何细胞型特异的蛋白在任何情况下都起作用,而是取决于核心 启动子提供一个合适的环境来决定基因是被激活还是被抑制。
构建多个启动子连接多个基因的表达载体,虽然可以实现同时转入多 个基因的愿望,但这种载体一方面构建困难,另一方面如果引入的启 动子之间仅仅有90bp 的同源性顺序,导入生物体内,就会引起所谓的 基因表达”共抑制”现象,而使基因沉默。因此,将极性启动子人工改 造为高效双向表达的启动子,对于促进基因工程的进展具有重要意义 。
2.1.1 基本转录因子(general transcription factors)
是RNA聚合酶结合启动子所必需的一组蛋白因子, 决定三种RNA(mRNA、tRNA及rRNA)转录的类别,包 括 TF Ⅱ A、TF Ⅱ B、TF Ⅱ D、TF Ⅱ E、TF Ⅱ F、TF Ⅱ H、 TFⅡI等7种因子。
基因转录调控表达PPT课件

原核基因转录系统 真核基因转录系统
三、基因概念的延伸 1、移动基因(Movable genes ) 1)插入序列(insertion sequence)
2)转座子(transponson)
侧翼IS 因子取向不同的两种复合转座子 Tn3系转座子的形体图
3)转位作用
保留型转座
复制型转座
4)逆转录转座子
个人观点供参考,欢迎讨论!
3、基因的概念
A gene is a physical and functional unit of genetic information with the potential to be expressed,i.e.to be used as a template to generate one or more gene products of RAN or protein.
第三章 基因转录、调控、表达
基因的概念
染色体上的DNA与基因的关系: 摩尔根的果蝇遗传试验告诉我们基因定位在染色体上:(遗传单位外段DNA): 这段DNA怎样控制性状的表达——中心法则。
一、基因与顺反子(cistron)、基因与酶的关系 1、基因与顺反子(cistron)的关系 2、基因与酶的关系
A cistron is a unit of genetic function defined by a complementation test. The bacterial gene is the same as a cistron--a unit of genetic function which corresponds to an open reading frame .
Using PCR to detect gene targeting events without the use of a selectable marker
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
真核生物RNA的转录终止
AAAAAAA· · · · · · 3
mRNA
3加尾
核酸酶 5 3
3 5
AATAAA
GTGTGTG
RNA-pol
转录终止的修饰点
真核生物转录后加工
1.加帽(adding cap): • 即在mRNA的5'-端加上m7GTP的结构。此 过程发生在细胞核内,即HnRNA即可进行 加帽。 • 加工过程首先是在磷酸酶的作用下,将5'端的磷酸基水解,然后再加上鸟苷三磷酸, 形成GpppN的结构,再对G进行甲基化。
• 真核生物转录延
长:与原核生物 类似,但由于存 在核小体的高级 结构,故在转录 延长过程中可观 察到核小体移位
和解聚现象。
• 真核生物转录终止:
在poly A修饰位点的下游存在一组共同序 列AATAAA和GTGTGT……,为转录终 止的识别修饰位点。 在转录越过修饰点后,RNA链在修饰点处
被切断,随即进行加帽和加尾修饰。
可变剪接(Alternative splicing)
可变剪接是指从一个特定的基因转录物中通过利用不同 5 ’、3’剪接位点产生出不同的成熟的mRNA的过程。主 要有四种方式:
(1)利用不同的启动子
(2)利用不同的Poly(A)位点
(3)保留某些内含子
(4)保留或去除某些外显子
4. RNA的编辑、再编码和修饰
• RNA的编辑(RNA editing)
RNA的一种加工方式,它导致了RNA所编码 的遗传信息的改变(如 单碱基突变),特别 是mRNA编辑,是在mRNA水平上信息发生 改变(碱基取代、插入或缺失)的过程
单碱基突变 尿苷酸的缺失和添加
DNA序列
mRNA序列 蛋白质序列
GA
4. 化学修饰
tRNA前体的转录合成
DNA
TGGCNNAGTGC
GGTTCGANNCC
RNA pol Ⅲ
tRNA前体
tRNA前体的切断和剪接加工
tRNA3'-末端-CCA序列的添加
tRNA的碱基修饰 (1)甲基化 如:A Am (2)还原反应 如:U DHU (3)核苷内的转位反应 如:U ψ
• (抗)终止因子
协助RNA聚合酶识别终止信号的蛋白质称为终止因子
(termination factor)。
原核生物中的终止因子蛋白是一种六聚体的蛋白质,亚
基的分子量为50kd。蛋白能识别转录终止信号,并与 RNA紧密结合,导致RNA的释放。
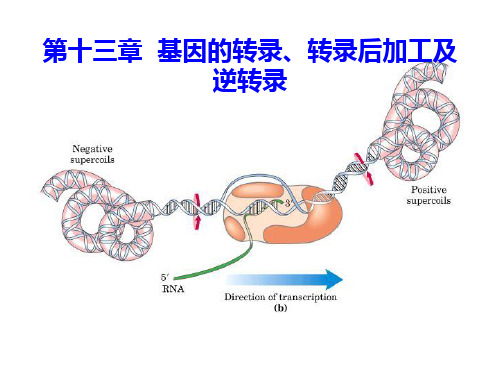
转录过程
起始 双链DNA 局部解开
启动子 (promoter)
• 有特定的起始和终止位点
RNA转录合成时,只能以DNA分子中的某一段作 为模板,故存在特定的起始位点和特定的终止位点。
特定起始点和特定终止点之间的DNA链构成一个转
录单位,通常由转录区和有关的调节顺序构成。
转录过程
• 参与转录合成的酶和蛋白因子
原料:NTP (ATP, UTP, GTP, CTP)
pGpA
pGpA
GpU 第二次转酯反应 G-OH
UpU
核酶与传统酶的区别:
一般的酶是纯的蛋白质,而核酶是 RNA
或带蛋白的RNA
有的核酶既是催化剂又是底物。而酶仅
催化反应
核酶发现的重大意义:
突破了酶的概念
揭示了内含子自我剪接的奥秘,促进了RNA的研究
为生命的起源和分子进化提供了ห้องสมุดไป่ตู้的依据
由终止因子(因子)识别特异的终止信号,并促
使RNA的释放。
富含 G-C
系列U
A. 不依赖于Rho() 的终止子 A. 依赖于Rho() 的终止子
大肠杆菌两类终止子的回文结构
1.非依赖Rho的转录终止:
模板DNA链在接近转
录终止点处存在相连的
富含GC和AT的区域,
使RNA转录产物形成
寡聚U及发夹形的二级 结构,引起RNA聚合 酶变构及移动停止,导 致RNA转录的终止。
Cap 0
100%
G
A
Guanine-7-methyl-transferase 2`-O-methyl-transferase
A
10~15%
2.加尾(adding tail):
• 这一过程也是细胞核内完成,首先由核 酸外切酶切去3'-端一些过剩的核苷酸, 然后再加入polyA。
• polyA结构与mRNA的半寿期有关。
特点:依赖于特定序列(GU…AG,AU…AC);SnSNP 形成剪接体;转酯反应 • Ⅱ型内含子(核酶)(mRNA,rRNA)(自我剪接) 特点:依赖于特定序列(U…U、U…G等);特殊结 构,自身催化;转酯反应
tRNA的转录后加工
• 主要有以下几种加工方式:
1. 切断
2. 剪接 3. 3’-末端-CCA序列添加
• 真核生物转录起始:
首先由TFⅡD的TBP
亚基识别并结合TATA
盒,然后在其他转录
因子的配合下,与
RNA聚合酶Ⅱ组装形
成转录起始前复合物
(pre-initiation
complex, PIC)。
• RNA聚合酶Ⅱ催化第一个磷酸二酯键形成。
• RNA聚合酶Ⅱ的羧基末端结构域(CTD) 被磷酸化修饰,大部分转录因子脱离,聚合 酶向下游移动延伸RNA链。
并起始转录有关的一些DNA调控序列被称
为启动子(promoter)。
原核生物启动子的保守序列
RNA聚合酶保护区 结构基因
5 3 5 3 -35 区 TTGACA AA C T G T RNA-pol辨认位点 (recognition site) -10 区 开始转录
-50 -40 -30 -20 -10 1
3、RNA的剪接(RNA splicing)
• 真核生物结构基因,由若干个编码区和非编码区互相
间隔开但又连续镶嵌而成,去除非编码区再连接后,
可翻译出由连续氨基酸组成的完整蛋白质,这些基因
称为断裂基因
A
B
C
非编码区
D
编码区 A、B、C、D
• tRNA
• rRNA
• Ⅰ型内含子(mRNA,rRNA)
(3) (4)
(2)
(1) (1)
(4)脱氨反应 如:A I
rRNA的转录后加工
在高等真核生物中,前体以沉降速率来 命名,命名为45SRNA,真核生物中前 体包括了18S rRNA、5.8S rRNA和28S rRNA序列
rRNA前体的转录和剪接加工
Ⅰ型内含子:hnRNA和 snRNA: 核内带有外显子和内含子编码序列的初级 RNA转录产物称为杂化核RNA (heteronuclear RNA, hnRNA) hnRNA经剪接加工除去内含子编码序列
3 5
10
3 5
T A T A A T Pu A T A T T A Py (Pribnow box)
• 流产转录 • 通过启动子(起始)快:强启动子 慢: 弱启动子
转录起始复合物
RNApol (2) - DNA - pppGpN- OH 3
• 延长
因子从全酶上脱离,余下的核心酶继续沿DNA链移 动,按照碱基互补原则,不断聚合RNA。
剪接体
U1 GU
U1
OH
UACUAAC U2 AG
U1
OH
U2 U4/6 U5
U1 U G UACUAAC AG
U5 U2 U4/6
OH
套索的形成及剪接 1
GU A AG
2
GU
A
AG
套索结构
U G A
AG
1
2
Ⅱ型内含子
• 线粒体与叶绿体的rRNA 中 • 1981年T.Cech和S.Atman 四膜虫 rRNA
模板:单链DNA
酶:RNA聚合酶(RNA-pol)
其他蛋白质因子:如转录因子、终止因子等
• RNA聚合酶
依赖DNA的RNA聚合酶,该酶在单链DNA
模板以及四种核糖核苷酸存在的条件下,
不需要引物,即可从5‘→3’聚合形成RNA。
原核生物中的RNA聚合酶全酶由五个亚基构成,即2'。 亚基与转录起始点的识别有关,在转录合成开始后被释 放;余下的部分(2')被称为核心酶,与RNA链的聚 合有关。
• 核酶(ribozyme):是指一类具有催化功
能的RNA分子,通过催化靶位点RNA链中
磷酸二酯键的断裂,特异性地剪切底物
RNA分子,从而阻断基因的表达
锤头型核酶的二级结构和空间立体结构示意图
剪接机制
外显子1 内含子 外显子2 GpU 第一次转酯反应 U-OH
UpA
pG-OH (ppG-OH, pppG-OH)
The 3` ends of mRNAs are generated by cleavage and polyadenylation;The sequence AAUAAA is necessary for cleavage to generate a 3` end for polyadenylation.
2,依赖 Rho因子的转录终止
DNA 序列缺乏共性,不能形成强的发卡结构
ATP
6聚体, 具有 NTP酶 和解螺旋酶
真核生物的转录过程
真核生物启动子保守序列
参与转录位点确定起始
控制转录起始频率
• 在远离受控基因处存在的,能够增强基因转录活性的调控 序列称为增强子(enhancer)。增强子特点: 远距离效应;无方向性;顺势调控;广泛性,相位性
转录
DNA
RNA
基本特点
• 不对称性
以双链DNA中的一条链作为模板进行转录,从而 将遗传信息由DNA传递给RNA