一类分数阶的病毒动力学模型
一类时滞分数阶计算机病毒模型的Hopf分岔研究

陕西理工大学学报!自然科学版)Journal of Shaanxi University of Technology ( Natural Science Edition)2021年6月第37卷第3期June. 2021Vol. 37 No. 3引用格式:高燕鑫,石剑平.一类时滞分数阶计算机病毒模型的Hgf 分岔研究[J ].陕西理工大学学报(自然科学版),2021,37(3) &52-60.一类时滞分数阶计算机病毒模型的Hopf 分岔研究高燕鑫,石剑平**收稿日期:2020-11甲8修回日期:2021-02-03基金项目:国家自然科学基金资助项目(11561034 );云南省教育厅科学研究基金资助项目(2017ZZX133)*通信作者:石剑平(1975—),女,侗族,贵州省黎平县人,博士,副教授,主要研究方向为非线性动力系统。
(昆明理工大学理学院,云南昆明650500)摘 要:研究了 一类时滞分数阶计算机病毒SIR 模型的Hopf 分岔问题。
首先通过再生矩阵方法分析系统基本再生数与正平衡f 之间的关系;其次以时滞作为分岔参数,利用线性化方法和Laplace 变换法分析系统在正平衡f 附近发生Hopf 分岔的条件,并推导了时滞临界值公式; 最后通过选择恰当的系统参数进行数值模拟,验证了理论分析结果的正确性,并显示了分数阶 变化对系统稳定域的影响。
关键词:分数阶;时滞;正平衡f ; Hopf 分岔;计算机病毒模型中图分类号:0193 文献标识码:A 文章编号:2096-3998(2021 )03-0052-09现实问题里很多系统的变化过程不仅依赖当前时刻的状态,还依赖于过去某个时刻或某段时刻的状态,这种特性称为时滞。
由于物质和能量在变化的时候往往不能瞬间传递,时滞发生在几乎所有类型 的自然和社会系统中口甲]。
计算机病毒模型$3甲%是一种以时间为状态变量的微分方程系统,引入时滞来描述和分析其动力学特征早就成为研究者的共识$5甲],并取得了很多有价值的成果。
几类分数阶传染病动力学模型研究

几类分数阶传染病动力学模型研究分数阶传染病动力学模型是对传染病传播过程进行建模和研究的数学方法。
与传统的整数阶模型不同,分数阶模型在描述传染病传播过程时引入了分数阶微分和分数阶积分的概念,能够更精确地描述传染病的动力学特性。
在研究中,分数阶传染病动力学模型主要可以分为以下几类。
首先,基础的分数阶SIR模型。
这类模型由分数阶微分方程组成,通常包括感染者数量、易感者数量和移动者数量等变量。
这类模型是传染病基本的传播模型,能够描述传染病的传播过程和基本动力学特性,如传播速率、传播范围等。
其次,分数阶SEIR模型。
这类模型在基础的SIR模型基础上引入了潜伏期概念,即将可感染的个体区分为潜伏期个体和易感个体。
潜伏期个体是指已经感染病毒但尚未出现症状的个体,通过分析潜伏期个体数量和易感个体数量的变化趋势,可以更准确地描述疫情的传播和爆发过程。
再次,分数阶SI模型。
这类模型通常用于描述传染病的最早期传播过程,不考虑恢复和治愈过程,即所有感染的个体都是永久性的感染者。
通过分析易感个体数量的变化趋势,可以预测传染病的传播速度和传播范围,为疫情的控制和预防提供科学依据。
最后,分数阶传染病模型的参数优化与控制。
在实际应用中,传染病的传播受到多种因素的影响,如人群流动、医疗资源分配等。
利用分数阶传染病模型可以推导出传播参数的数学表达式,进而进行参数优化和控制策略的设计。
通过优化模型参数,可以最大限度地减少疫情的传播速度和传播范围,为疫情防控提供有力支持。
综上所述,分数阶传染病动力学模型是对传染病传播过程进行建模和研究的一种重要方法。
在分析疫情特征、预测疫情走势以及指导疫情防控方面具有重要意义。
随着分数阶微积分的理论和方法的不断发展,分数阶传染病动力学模型的应用将会更加广泛和深入。
一类具反馈控制的病毒感染模型的稳定性与周期解

一类具反馈控制的病毒感染模型的稳定性与周期解程荣福【摘要】研究了一类具有反馈控制的病毒感染动力学模型.利用常微分方程定性与稳定性方法,通过分析特征方程,讨论了该模型各个平衡点的局部稳定性;通过构造适当的Lyapunov泛函,证明了未感染平衡点和反馈控制病毒感染平衡点的全局稳定性;最后,利用重合度理论中的延拓定理,给出了保证其周期系统存在正周期解的充分条件.【期刊名称】《北华大学学报(自然科学版)》【年(卷),期】2011(012)001【总页数】8页(P10-17)【关键词】病毒感染模型;反馈控制;重合度理论;稳定性;周期解【作者】程荣福【作者单位】北华大学数学学院,吉林,吉林,132033【正文语种】中文【中图分类】O175.11 引言数学生态学模型具有非常深刻的实际背景,历来受到学术界的重视,现已有了大量工作(参见文献[1-6]及其参考文献).基于人与自然的协调发展和生态资源的可持续发展,带有反馈控制的数学生态学模型更具有深远意义,反馈控制已被越来越多的学者所关注.在具体的生态问题中,为了某种需要,需人为地改变种群规模的正平衡态,一种有效的办法是在所考虑的生态系统中引入反馈控制变量,借以改变生态系统正平衡点的大小.文献[7-8]提出了如下反馈控制系统(1.1)其中:n(t)表示t时刻的种群数目;u(t)表示t时刻的反馈控制变量.给出了该模型正平衡点全局渐近稳定的充分条件.文献[9]研究了如下一类具有非线性感染率的病毒动力学模型(1.2)其中:x(t)表示t时刻的易感细胞的数目;y(t)表示t时刻的感染细胞的数目.讨论了非线性感染率对病毒感染动力学性态的影响,给出了病毒消失平衡点和病毒幸存平衡点全局渐近稳定的充分条件.近年来,对带有反馈控制的数学生态学模型的研究,大多集中于无病毒感染发生的自治系统或非自治系统来讨论其动力学性质[10-12],考虑带有反馈控制的病毒感染模型却不多见.基于以上工作,本文考虑如下具反馈控制的病毒感染模型(1.3)其中:x(t),y(t)分别表示t时刻易感者种群的密度和已染病者种群的密度;u(t)表示t 时刻的控制变量的密度;β(y)表示非线性感染发生率;参数a,b,c,d,p,r,α,β均为正常数.模型(1.3)的建立基于以下假设:1)易感者种群:以常数速率r产生,以速率dx死亡并以速率β(y)x变为感染者;2)感染者种群:以速率β(y)x产生,以速率ay死亡并以速率αyu被控制变量所捕获;3)控制变量:以速率by产生并以速率cu死亡.3:x(t)≥0,y(t)≥0,u(t)≥0}.在本文中,我们首先利用常微分方程定性分析方法,对模型(1.3)在内的非负平衡点性态进行分析;然后通过构造Lyapunov泛函,利用LaSalle不变性原理,讨论模型(1.3)非负平衡点全局渐近稳定性的充分条件;最后考虑模型(1.3)为周期系统时,研究周期系统的正周期解存在的充分性判据.模型(1.3)具有生态背景和一般性.2 局部稳定性易知,模型(1.3)存在一个未染病平衡点E0(x0,0,0);当rβ>ad时,模型(1.3)存在一个带有反馈控制的病毒感染平衡点Ω(x*,y*,u*).其中:;;; .式中:A=bα(dp+β);B=ac(dp+β)+bαd;C=c(rβ-ad).据此,我们有如下结论: 引理2.1 模型(1.3)存在一个未染病平衡点E0(x0,0,0);当rβ>ad时,模型(1.3)存在一个带有反馈控制的病毒感染平衡点Ω(x*,y*,u*).下面我们在引理2.1的基础上,运用常微分方程定性分析方法,讨论模型(1.3)的两个非负平衡点的局部稳定性.1)模型(1.3)在未染病平衡点E0(x0,0,0)处的线性化矩阵为特征方程为(λ+b)(λ+d)(λ-βx0+a)=0.显然,特征方程有3个实根λ1=-b<0,λ2=-d<0,当rβ>ad时,λ3>0,于是E0(x0,0,0)是不稳定的;当rβ<ad时,λ3<0,于是E0(x0,0,0)是局部稳定的.2)模型(1.3)在带有反馈控制的病毒感染平衡点Ω(x*,y*,u*)处(rβ>ad)的线性化特征方程为λ3+n2λ2+n1λ+n0=0,其中:易知n2n1-n0>0,由Hurwitz判据[13]知特征方程的所有根均具有负实部,因此,Ω(x*,y*,z*)是局部稳定的.综上,我们得到如下结论:定理2.1 模型(1.3)的两个非负平衡点E0(x0,0,0),Ω(x*,y*,z*)性态如下:1)当rβ<ad时,E0(x0,0,0)是局部稳定的;当rβ>ad时,E0(x0,0,0)是不稳定的.2)当rβ>ad时,Ω(x*,y*,z*)总是局部稳定的.3 全局稳定性下面我们在定理2.1的基础上,通过构造适当的Lyapunov泛函,利用LaSalle不变性原理[13],讨论模型(1.3)两个非负平衡点全局渐近稳定的充分条件.定理3.1 如果rβ<ad,则模型(1.3)的未感染平衡点E0(x0,0,0)是全局渐近稳定的.证明将模型(1.3)改写为如下等价系统(3.1)设(x(t),y(t),u(t))T是系统(3.1)的任意正解,构造Lyapunov泛函其中:待定常数ai>0(i=1,2,3).计算V1(t)沿着系统(3.1)的解的导数得D+V1(t)= a1(x(t)-x1)x′(t)+a2y′(t)+a3u′(t)=-a1(x(t)-x0)2-(a1x0-a2)βx(t)+a2a-a1β+a2apy(t)-a3b-a3bpy(t)-(a2py(t)+a3c)u(t),当rβ<ad时,选取注意到则易见模型(1.3)在M1中的最大不变集只有点E0(x0,0,0),因此E0(x0,0,0)是全局吸引的.由定理2.1知,当rβ<ad时E0(x0,0,0)是局部稳定性的,进而由LaSalle不变性原理知,模型(1.3)的未感染平衡点E0(x0,0,0)是全局渐近稳定的. 定理3.2 如果rβ>ad,则模型(1.3)带有反馈控制的病毒感染平衡点Ω(x*,y*,z*)是全局渐近稳定的.证明将模型(1.3)改写为如下等价系统(3.2)设(x(t),y(t),u(t))T是系统(3.2)的任意正解,构造Lyapunov泛函其中:待定常数ci>0(i=1,2,3).计算V2(t)沿着系统(3.2)的解的导数得D+V2(t)= c1(x(t)-x1)x′(t)+c2(y(t)-y1)+c3(u(t)-u*)u′(t)=当rβ>ad时,选取则易见模型(1.3)在M2中的最大不变集只有点Ω(x*,y*,z*),因此Ω(x*,y*,z*)是全局吸引的.由定理2.1知,当rβ>ad时Ω(x*,y*,z*)是局部稳定性的,进而由LaSalle不变性原理知,模型(1.3)带有反馈控制的病毒感染平衡点Ω(x*,y*,z*)是全局渐近稳定的.4 周期解的存在性真正符合实际的生态系统其系数都是时变的,特别地,如果考虑到季节环境的周期性变化、动物繁殖的周期性变化以及流行病的周期性变化等,均可以假设生态系统系数是有公共周期的连续周期函数.因此,我们假设模型(1.3)中参数a,b,c,d,r,α,β都是关于t的ω-周期连续函数,状态常数p>0,这样模型(1.3)就成为一个ω-周期系统(4.1)系统(4.1)在t时刻的生态意义与模型(1.3)相同.下面研究系统(4.1)存在正周期解的充分性判据.为此,我们引入重合度理论中的延拓定理[14].设X,Z是赋范向量空间,L:DomL⊂X→Z为线性映射,N:X→Z为连续映射.如果dim KerL=co dim ImL<+∞且ImL为Z中闭子集,则称映射L为指标为零的Fredholm映射.如果L是指标为零的Fredholm映射且存在连续投影P:X→X以及Q:Z→Z使ImP=KerL,ImL=KerQ=Im(I-Q),则LDomL∩KerP:(I-P)X→ImL可逆,设其逆映射为KP.设Ω为X中的有界开集,如果有界且是紧的,则称N在上是L-紧的.由于ImQ与KerL同构,因而存在同构映射J:ImQ→KerL.引理4.1 设X、Z是Banach空间,同构映射J:ImQ→KerL,L是指标为零的Fredholm算子,在上是L-紧的,其中Ω是X中的有界开集,且满足:ⅰ) Lx≠λNx,∀x∈∂Ω∩DomL,λ∈(0,1);ⅱ) QNx≠0,∀x∈∂Ω∩K erL;ⅲ) deg{JQN,Ω∩KerL,0}≠0.则方程Lx=Nx在内至少存在一个解.为着以下证明,对连续的ω-周期函数g(t),本文采用记号:定理4.1 如果并且则系统(4.1)至少有一个ω-周期正解.证明作变换x(t)=ex1(t),y(t)=ex2(t),u(t)=ex3(t),则系统(4.1)化为如下等价系统(4.2)易知,如果系统(4.2)有ω-周期解那么就是系统(4.1)的ω-周期正解.为此,我们只需证明系统(4.2)的ω-周期解的存在性.取X=Z={x=(x1(t),x2(t),x3(t))T∈C(,3):x(t+ω)=x(t)},x=xi(t),则X、Z在如上范数·下为Banach空间.令L:DomL∩X,DomL={x∈C1(,3)},N:X→X,定义两个投影P及Q为显然,是X的闭子空间,而且dim KerL=3=dim(Z\ImL),因此L是指标为零的Fredholm算子,容易证明P、Q是连续投影且使得ImP=KerL,KerQ=ImL=Im(I-Q).因此LP的逆映射KP具有形式KP:ImL→DomL∩KerP, KP(x)=x(s)ds-x(s)dsdt.显然,利用Lebesgue收敛定理可以证明QN及KP(I-Q)N是连续的.利用Arzela-Ascoli定理,能够证明对X中的任何有界开集和是相对紧的.因此,对X中的任何有界开集Ω,N在上是L-紧的.对应算子方程Lx=λNx,λ∈(0,1),有(4.3)设x∈X是系统(4.3)对应某个λ∈(0,1)的解,将系统(4.3)中各式两端同时从0到ω积分得dt=0, (4.4)dt=0, (4.5)b(t)ex2(t)-x3(t)-c(t)dt=0. (4.6)因为x∈X,所以存在ηi,ξi∈[0,ω]使得(t) (i=1,2,3).(4.7)于是由式(4.4)~(4.7)有(4.8)-a(η2)-α(η2)ex3(η2)=0,(4.9)b(η3)ex2(η3)-x3(η3)-c(η3)=0,(4.10)以及(4.13)由式(4.8)有则得M1.(4.14)由式(4.10)有b(η3)ex2(η3)=c(η3)ex3(η3),即(4.15)由式(4.9)有=a(η2)+α(η2)ex3(η2)>a(η2),根据定理条件和式(4.14)可得M2.(4.16)将式(4.16)代入式(4.15)得到M3.(4.17)另一方面,由式(4.11)有即m1.(4.18)由式(4.13)有(4.19)利用式(4.17)~(4.18),对式(4.12)有于是,根据定理条件可得m2>0.(4.20)将式(4.20)代入式(4.19)得到m3.(4.21)综上,由式(4.14)、(4.16)~(4.18)、(4.20)~(4.21)得xi(t)<max{lnMi,lnmi}Bi(i=1,2,3).显然,Bi(i=1,2,3)与λ的选取无关.由于代数方程组在定理所给的条件下有唯一正解因此,令这里B0取充分大,使得(lnx*,lny*,lnu*)T=lnx*+lny*+lnu*<B0.令Ω={x∈X:x<B}.则Ω满足引理4.1的条件ⅰ).当x∈∂Ω∩KerL=∂Ω∩3时,x是3的一个常向量且满足x=B,于是这样Ω满足了引理4.1的条件ⅱ).直接计算得deg{JQN,Ω∩KerL,0}=sgn=-1≠0.这里同构J映射可取恒同映射,因为ImQ=KerL.这就证明了Ω也满足引理4.1的条件ⅲ).至此,Ω满足引理4.1的所有条件,由引理4.1知,方程Lx=Nx在中至少有一个解,即系统(4.3)在Ω内均至少存在一个ω-周期解.从而,由变换x(t)=ex1(t),y(t)=ex2(t),u(t)=ex3(t)知,周期系统(4.2)至少存在一个ω-周期正解.【相关文献】[1] Zeng G Z,Chen L S,Chen J F.Persistence and Periodic Orbits for Two-species Nonautonomous diffusion Lotka-Volterra Models [J].Math Comput Modeling,1994,20(12):69-80.[2] 程荣福,常亮.具无限时滞和非单调功能性反应捕食系统的多周期解[J].吉林大学学报:理学版,2010,48(5):761-765.[3] Gao P,Cheng R F.Existence of Multiple Periodic Solutions of a Predator-Prey System with Infinite Delay[J].Word Academic Union,2010,16:77-80.[4] 赵明,程荣福.一类具生物控制和比率型功能反应的食物链系统周期解的存在性[J].吉林大学学报:理学版,2009,47(4):730-736.[5] Gao P,Cheng R F.Periodic of Predator-Prey Diffusion System with Depositings and Bedding-Type[J].Word Academic Press,2010,4(6):201-205.[6] Cheng R F.Global Existence of Positive Periodic Solution of a Predator-Prey Diffusive Model with Double-Density Restrict and Functional Response[J].Word Academic Union,2010,7:52-55.[7] Gopalsamy K.Stability and Oscillations in Delay Differential Equation of Population Dynamics[M].Boston:Kluwer Academic Publishers,1992.[8] Gopalsamy K,Weng P X.Feedback Regulation of Logistic Growth[J].Internal Math and Math Sci,1993,16:177-192.[9] 杨茂,王开发.一类非线性感染率病毒动力学模型分析[J].四川师范大学学报:自然科学版,2009,32(3):304-306.[10] 程荣福,孙吉荣.具生物控制的时滞阶段结构种群模型的稳定性[J].北华大学学报:自然科学版,2008,9(2):97-103.[11] 丁孝全,程述汉.具反馈控制的时滞阶段结构种群模型的稳定性[J].生物数学学报,2006,21(2):225-232.[12] 程荣福.一类具生物控制的多滞量捕食模型正周期解的存在性[J].北华大学学报:自然科学版,2010,11(1):1-6.[13] 马知恩,周义仓.常微分方程定性与稳定性方法[M].北京:科学出版社,2001:49-70.[14] Gaines R E,Mawhin J L.Coincidence Degree and Nonlinear DifferentialEquations[M].Berlin:Springer-Verlag,1977:40-45.。
一类具有细胞感染年龄和一般饱和感染率的病毒感染动力学模型的稳定性分析

一类具有细胞感染年龄和一般饱和感染率的病毒感染动力学模型的稳定性分析李梁晨;徐瑞【摘要】为了了解病毒在人体内的感染、受制、清除等动力学过程,研究一类具有细胞感染年龄和一般饱和感染率的病毒感染动力学模型,证明当病毒的基本再生率大于1时,模型存在唯一的病毒感染稳态解.通过分析相应特征方程讨论了可行稳态解的局部稳定性,在构造Lyapunov泛函和应用LaSalle不变集原理的基础上,证明了当基本再生率小于1时,病毒未感染稳态解是全局渐近稳定的;当基本再生率大于1时,病毒感染稳态解是全局渐近稳定的.【期刊名称】《河北科技大学学报》【年(卷),期】2016(037)004【总页数】8页(P349-356)【关键词】稳定性理论;细胞感染年龄;饱和感染率;Lyapunov泛函;LaSalle不变集原理【作者】李梁晨;徐瑞【作者单位】军械工程学院基础部,河北石家庄 050003;军械工程学院基础部,河北石家庄 050003【正文语种】中文【中图分类】O175传染病是威胁人类生存、发展的重要因素之一。
近年来,很多学者开始从微观角度对传染病的流行条件进行研究[1],为此他们建立了一系列描述人体内感染过程的病毒感染动力学模型,这些模型关注于染病个体体内的疾病动力学过程。
通过研究病毒感染动力学模型,可以了解病毒在人体内的感染、复制(繁衍)、清除等动力学过程,为临床制定合理的治疗方案提供理论依据。
1996年NOWAK等[2]提出了病毒感染动力学的基本模型。
受其启发,在文献[2]中模型的基础上,许多学者进行了扩展性的研究,对基于不同病毒的具体感染特征提出了大量的数学模型[3-7]。
在这些模型中,被感染细胞的死亡率和产生病毒的速率都被设为了常数。
然而,文献[8—9]通过实验证明,被感染细胞的死亡率和产生病毒的速率是随着细胞的感染年龄(健康细胞被感染后的时长)而改变的。
因此,文献[10]提出并研究了如下具有细胞感染年龄的HIV感染模型:其中:x(t), v(t)分别表示t时刻健康T细胞的密度和细胞外具有感染性的病毒的密度;a为细胞的感染年龄;y(a,t)表示在t时刻感染年龄为a的染病T细胞的密度;s,d,u,β均为正常数,s为健康T细胞的产生率,d为健康T细胞的死亡率,u为病毒的死亡率;βx(t)v(t)为双线性感染率;δ(a),k(a)分别表示感染年龄为a的染病T细胞的死亡率和产生病毒的速率。
一类具有常数免疫输入的病毒灢免疫应答动力学模型分析

第36卷 第6期 陕西科技大学学报 V o l.36N o.6 2018年12月 J o u r n a l o f S h a a n x iU n i v e r s i t y o f S c i e n c e&T e c h n o l o g y D e c.2018* 文章编号:2096-398X(2018)06-0176-07一类具有常数免疫输入的病毒-免疫应答动力学模型分析李亚男,王晓琴*,李建全(陕西科技大学文理学院,陕西西安 710021)摘 要:在假设病毒以饱和形式刺激免疫应答的基础上,将免疫细胞的常数输入引入病毒-免疫应答模型.此模型平衡点的存在性和稳定性得到了完整的分析.其全局稳定性是通过构造恰当的L y a p u n o v函数证得.并发现免疫存在时的病毒内禀增长率决定模型的动力学性态.也就各免疫因素对病毒持续的影响进行了讨论,同时用数值模拟验证了所得的理论结果.关键词:病毒感染;免疫应答;平衡点;稳定性中图分类号:O175.12 文献标志码:AA n a l y s i s o f a c l a s s o f v i r u s-i m m u n e r e s p o n s e d y n a m i cm o d e lw i t h c o n s t a n t i m m u n e i n p u tL IY a-n a n,WA N G X i a o-q i n*,L I J i a n-q u a n(S c h o o l o fA r t s a n dS c i e n c e s,S h a a n x iU n i v e r s i t y o f S c i e n c e&T e c h n o l o g y,X i'a n710021,C h i n a)A b s t r a c t:B a s e do n t h e a s s u m p t i o n t h a t t h ev i r u s s t i m u l a t e s t h e i mm u n e r e s p o n s e i na s a t u-r a t e d f o r m,t h e c o n s t a n t i n p u t o f t h e i mm u n e c e l l i s i n c o r p o r a t e d i n t o t h ev i r u s-i mm u n e r e-s p o n s em o d e l.T h e e x i s t e n c e a n d s t a b i l i t y o f e q u i l i b r i a o f t h em o d e l a r e c o m p l e t e l y a n a l y z e d.T h e i r g l o b a l s t a b i l i t i e s a r e p r o v e db y c o n s t r u c t i n g t h ea p p r o p r i a t eL y a p u n o v f u n c t i o n s.O u r r e s u l t s s h o wt h a t t h e d y n a m i c s o f t h em o d e l i s d e t e r m i n e db y t h e i n t r i n s i c g r o w t h r a t e o f t h e v i r u s i n t h e p r e s e n c e o f i mm u n i t y.T h e e f f e c t o f v a r i o u s i mm u n e f a c t o r s o n t h e p e r s i s t e n c e o f v i r u s i s a l s o d i s c u s s e d.T h e t h e o r e t i c r e s u l t s o b t a i n e d a r e v e r i f i e db y m e a n s o f n u m e r i c a l s i m-u l a t i o n.K e y w o r d s:v i r a l i n f e c t i o n;i mm u n e r e s p o n s e;e q u i l i b r i u m;s t a b i l i t y0 引言众所周知,乙肝病毒(H B V)㊁丙肝病毒(H C V)和艾滋病病毒(H I V)等在人体内可以建立长久的慢性感染,对人们的健康甚至生命造成了严重的威胁.病毒在人体内的慢性感染程度,通常与个体的免疫应答强度密切相关.目前,在临床上已基本了解人体免疫系统对大多数病毒的作用机理,但具体的作用方式仍未完全理解.于是,借助数学模型来刻画免疫系统对病毒感染作用的具体形式,对认识和了解以及控制病毒对人体的感染有着重要的现实意义,并且已取得一些显著的成果[1,2].人体的免疫形式分为体液免疫和细胞免疫两类.体液免疫通常是以抗体对病毒的抑制作用为*收稿日期:2018-06-06基金项目:国家自然科学基金项目(11371369)作者简介:李亚男(1992-),女,山西长治人,在读硕士研究生,研究方向:生物数学通讯作者:王晓琴(1973-),山西忻州人,副教授,研究方向:非线性动力系统,w a n g x i a o q i n@s u s t.e d u.c n Copyright©博看网 . All Rights Reserved.第6期李亚男等:一类具有常数免疫输入的病毒-免疫应答动力学模型分析主,而细胞免疫则主要以免疫T 细胞对作为抗原的被感染细胞进行溶解等作用来控制病毒的产生[3].对此,用数学模型描述病毒的感染过程时,至少需要引入未感染细胞㊁被感染细胞和病毒三种变量[2,4,5].若再考虑免疫应答的作用,就需要增加数学模型中的变量个数,这样模型的结构变得比较复杂,同时也增加了模型动力学性态的分析难度.进一步,若是以H o l l i n g I I 或I I I 等非线性函数来刻画相互作用形式时,模型的动力性质往往难以得到较全面的理论分析结果[6,7].为了能清晰地研究病毒和免疫应答的直接作用方式,一些不考虑宿主细胞,而仅考虑病毒和免疫应答相互作用的简单数学模型已经提出,这就为后续的数学分析提供了方便[8-12].在文献[10,11,12]中,对于免疫细胞的输入,均假设仅因个体体内存在的病毒刺激免疫系统而产生,这就意味着病毒不存在时,免疫系统的免疫力消失.同时以具有饱和形式H o l l i n g I I 函数描述病毒对免疫细胞的刺激产生率.但临床检测已经发现,当病毒不存在时,由免疫系统(如淋巴系统)分泌的免疫细胞会处于一种动态平衡状态.因此,本文将在已有文献所述模型的基础上,再将不依赖于病毒的免疫细胞输入引入模型之中,进而分析其动力学性态.1 模型及其有界性为了用简单模型描述免疫应答与个体体内病毒的相互作用,假设当免疫应答不存在时,病毒的变化符合l o gi s t i c 模型,即v '=r v 1-v æèçöø÷k ,其中v =v (t )表示t 时刻个体体内的病毒载量,r 表示病毒的內禀增长率,k 表示个体体内病毒的最大量.假设没有病毒存在时,免疫细胞数量的变化满足模型z '=λ-d z ,其中z =z (t )表示t 时刻个体体内的免疫细胞数量,λ表示免疫细胞的输入率,d 表示免疫细胞的死亡率系数.在现实中,当个体被病毒感染时,个体体内的病毒与免疫细胞会相互作用.免疫细胞对病毒具有杀伤作用,在此假设这种杀伤作用符合质量作用定律,即杀伤率与病毒载量和免疫细胞数量的乘积成正比,设比例系数为p ,则t 时刻免疫细胞对病毒的杀伤率为p v z .另一方面,病毒的出现会刺激个体的免疫系统分泌产生相应的免疫细胞.这里假设被刺激的免疫系统以饱和形式c v z v +ε产生免疫细胞,其中c 为最大的免疫细胞刺激产生率,ε为半饱和常数.在以上假设下,可建立如下病毒与免疫细胞相互作用的数学模型v '=r v 1-v æèçöø÷k -p v zz '=λ+c v z v +ε-ìîíïïïïd z (1) 对于模型(1)有定理1 对t ≥0,模型(1)在非负初始条件下的解(v (t ),z (t))非负且最终一致有界.证明:由模型(1)的第一个方程可得,当v (0)=0时,对所有的t ≥0,都有v (t)≡0.由于v (t )≡0是模型(1)的解,所以根据自治系统解轨线的唯一性知,当v (0)>0时,对所有的t ≥0,都有v (t)>0.对于z (0)≥0,由模型(1)第二个方程可知,对于t >0,z =z (t )总是正的.事实上,如果z (0)>0,假设存在t 0(t 0>0)使得z (t )在t 0处首次为零,即z (t 0)=0,且当t ∈[0,t 0)时,z (t )>0,所以d z (t )d t t =t 0≤0.但由模型(1)的第二个方程有d z (t )d t t =t 0=λ>0,与实际相矛盾,因此假设不成立.故对所有的t ≥0,z (t )>0.如果z (0)=0,则d z (t )d t t =0=λ>0.根据z =z (t )的连续性,存在⎺t 使得当t ∈(0,⎺t )时z (t )>0.同情形z (0)>0类似可知,对于t >0有z (t)>0.综上所述,当t ≥0时,模型(1)在非负初始条件下的解(v (t ),z (t))是非负的.取L 1=v +p cz ,则L 1沿着模型(1)的解的全导数为d L 1d t =r v 1-v æèçöø÷k -p v z +p λc +pv z v +ε-p d cz ≤r v 1-v æèçöø÷k +p λc -p d cz =v r 1-v æèçöø÷k +éëêùûúd +p λc -d v +p c æèçöø÷z ≤ k (r +d )24r +p λc-d v +p c æèçöø÷z (2)所以l i m t →∞s u p v (t )+p c z (t éëêùûú)≤k (r +d )24r d +p λc d .因此模型(1)在非负初始条件下的解是最终有界的.定理1证毕.同时,对于v ,z ≥0,由模型(1)的第一个方程有v '=r v 1-v æèçöø÷k -p v z ≤r v 1-v æèçöø÷k ,所以l i m t →∞s u p v (t )≤k .由模型(1)的第二个式子得到z '=λ+c v z v +ε-d z ≥λ-d z ,故l i m t →∞i n f z (t )≥λd .于是结合定理1的证明知,模型(1)有正不变集㊃771㊃Copyright©博看网 . All Rights Reserved.陕西科技大学学报第36卷Γ=(v ,z ):v ≥0,z >0,v ≤k ,z ≥λd ,v +p c z ≤k (r +d )24r d +p λìîíïïïïüþýïïïïc d.因此,只需在Γ上分析模型(1)的动力学性态.2 平衡点的存在性和局部渐近稳定性模型(1)的平衡点P (v ,z )坐标满足方程组:r v 1-v æèçöø÷k -p v z =0λ+c v z v +ε-d z =ìîíïïïï0(3)显然,方程组(3)总存在解0,λæèçöø÷d ,因此模型(1)总存在边界平衡点P 00,λæèçöø÷d .下面讨论模型(1)正平衡点的存在性.当v ≠0时,由方程组(3)中的第一个方程有 z =r p1-v æèçöø÷k (4) 由于在正不变集Γ上z ≥λd,所以由式(4)知,对于模型(1)的平衡点P (v ,z )有r p 1-v æèçöø÷k ≥λd,即v ≤k 1-p λæèçöø÷r d =:⎺v .这意味着对于正平衡点P (v ,z )有v <k .为了保证v 的正性,r d >p λ就是模型(1)正平衡点存在的必要条件.进一步,将式(4)代入方程组(3)中的第二个方程,并整理可得k λk -v +r p (c -d )v -d εv +éëêêùûúúε=0(5)记f (v )=k λk -v +r p (c -d )v -d εv +éëêêùûúúε易得f (⎺v )=r c ⎺v p (⎺v +ε)>0,f '(v )=k λ(k -v )2+r c εp (v +ε)2>0,且当r d >p λ时,f (0)=λ-r d p <0.因此,函数f (v )在区间(0,⎺v )上有唯一正零点,记为v *.相应地z *=r p 1-v *æèçöø÷k .于是模型(1)当且仅当r d >p λ时有唯一正平衡点p *(v *,z *).综上所述,关于模型(1)在正不变集Γ上的平衡点存在性有如下定理.定理2 模型(1)总有边界平衡点P 00,λæèçöø÷d .当且仅当r d >p λ时模型(1)还有唯一正平衡点p *(v *,z *),其中v *是方程f (v )=0的正根,z *=r b 1-v *æèçöø÷k .下面讨论平衡点P 0和P *的局部渐近稳定性.模型(1)在P 0点的J a c o b i 矩阵为:J (P 0)=r -p λd0c λd ε-æèçççöø÷÷÷d 矩阵J (P 0)的两个特征根分别为 λ1=r 1-p λæèçöø÷r d ,λ2=-d 所以当r d <p λ时模型(1)的边界平衡点P 0是局部渐近稳定的;当r d >p λ时P 0是不稳定的.当r d =p λ时,矩阵J (P 0)的两个特征根为λ1=0和λ2<0.因此P 0是模型(1)的高阶平衡点,同时在P 0附近满足局部中心流形存在的条件.下面用中心流形来判断P 0在正不变集Γ上的稳定性.为了便于计算,首先做变换,令u =v ,y =z -λd,则模型(1)变为u '=-p u y +λk d æèçöø÷u y '=c λd εu -d y +c u u +εy -λu d æèçöø÷ìîíïïïïε(6)将系统(6)第二式中1u +ε在原点处T a yl o r 展开得 æèçöø÷u y '=A æèçöø÷u y +f 1(u ,y )g 1(u ,y æèçöø÷)(7)其中:A =00c λd ε-æèççöø÷÷d ,f 1(u ,y )=-p u y -p λk d u 2,g 1(u ,y )=c εu y -λc d ε2u 2-c ε2u 2y +λc d ε3u 3+ o (|u ,y |3). 直接计算知,ξ1=d ,c λd æèçöø÷εT ,ξ2=0,c λdæèçöø÷εT分别是矩阵A 的两个特征值λ1=0,λ2=-d 对应的特征向量.利用线性变换 æèçöø÷u y =d 0c λd εc λd æèççöø÷÷εæèçöø÷w x (8)系统(7)变为 æèçöø÷w x '=000-æèçöø÷d æèçöø÷w x +f2(w ,x )g 2(w ,x æèçöø÷)(9)其中:f2(w ,x )=-p c λd ε+λæèçöø÷k w 2-p c λd εw x ,g2(w ,x )=a 1w 2+a 2w x +a 3w 3+a 4w 2x + o (|w ,x |3)其中a 1㊁a 2㊁a 3㊁a 4分别为:a 1=p λc d ε+1æèçöø÷k +d (c -d )ε,a 2=p c λd ε+d c ε,㊃871㊃Copyright©博看网 . All Rights Reserved.第6期李亚男等:一类具有常数免疫输入的病毒-免疫应答动力学模型分析a 3=-d 2(c -d )ε2,a 4=-d 2c ε2. 根据中心流形定理[13],假设系统(9)在P 0的局部中心流形为x =h (w )=bw 2+o (w 2)(10) 其中:b 为待定常数.对式(10)两边沿着系统(9)的解关于t 求导得-2b p c λd ε+λæèçöø÷k w 3-2b p c λd εw 2x +o (|w ,x |3)=-d x +a 1w 2+a 2w x +a 3w 3+a 4w 2x +o (|w ,x |3)(11)再将x =h (w )=bw 2+o (w 2)代入式(11)得-2b pc λd ε+λæèçöø÷k w 3-2b 2p c λd εw 4+o (w 4)=(a 1-b d )w 2+(a 2b +a 3)w 3+a 4b w 4+o (w 4)(12)比较式(12)两端w 的同次幂系数得b =a 1d,即b =p λd c d ε+1æèçöø÷k +c -d ε.于是系统(9)在P 0的局部中心流形为x =h (w )=p λd c d ε+1æèçöø÷k +c -d éëêêùûúúεw 2+o (w 2)(13)进一步将式(13)代入系统(9)中的第一方程,得中心流形上系统(9)的退化方程为w '=-p λc d ε+1æèçöø÷k w 2- p c λd εp λd c d ε+1æèçöø÷k +c -d éëêêùûúúεw 3+o (w 3)(14) 注意到在式(14)右端w 的最低次数是2,且w 2的系数小于零,并结合式(8)以及平移变换易知当r d =pλ时,边界平衡点P 0在正不变集Γ上是局部渐近稳定的.模型(1)在正平衡点P *的J a c o b i 矩阵为J (P *)=r -2r v *k -pz *-p v *c εz *(v *+ε)2c v *v *+ε-æèççççöø÷÷÷÷d 借助v *和z *满足的方程组(3)可将矩阵J (P *)化简为J (P *)=-r k v*-p v *c w z *(v *+ε)2-λz æèçççöø÷÷÷* 显然,d e t J (P *)>0,t r J (P *)<0.因此正平衡点P *只要存在就是局部渐近稳定的.于是关于模型(1)平衡点的局部稳定性有如下定理.定理3 当r d ≤p λ时,模型(1)的边界平衡点P 0在正不变集Γ上是局部渐近稳定的,当r d >p λ时P 0是不稳定的;模型(1)的正平衡点P *只要存在,在正不变集Γ内就是局部渐近稳定的.3 平衡点的全局渐近稳定性关于模型(1)平衡点的全局稳定性有如下定理.定理4 当r d ≤p λ时,模型(1)的边界平衡点P 0在正不变集Γ上是全局渐近稳定;正平衡点P *只要存在,在正不变集Γ内就是全局渐近稳定的.证明:为了讨论模型(1)边界平衡点P 00,λæèçöø÷d 的全局稳定性,可将模型(1)化为v '=v r 1-v æèçöø÷k -p λd -p z -λæèçöø÷éëêêùûúúd z '=z λ1z -d æèçöø÷λ+c v v +éëêêùûúúìîíïïïïε(15) 定义L y a pu n o v 函数L 2=c p l n v +εæèçöø÷ε+z -λd -λd l n d z æèçöø÷λ则L 2沿着系统(15)解的全导数为L '2|(15)=c p 1v +εv '+1-λæèçöø÷d z z '=c p v v +εr -p λd -r k æèçöø÷v +λ2-λd z -d z æèçöø÷λ 显然,当r d ≤p λ时L '2|(15)在正不变集Γ上关于点P 0负定,因此由L y a pu n o v 稳定性定理[14]知,当r d ≤p λ时,模型(1)的边界平衡点P 0在正不变集Γ上是全局渐近稳定的.对于模型(1)的正平衡点P *(v *,z *),由方程组(3)有 r =r k v *+p z *,d =λz *+c v *v *+ε于是模型(1)可表示为v '=-v r k (v -v *)+p (z -z *éëêêùûúú)z '=z λ1z -1z æèçöø÷*+c v v +ε-v *v *+æèçöø÷éëêêùûúúìîíïïïïε(16) 为了证明平衡点P *在正不变集Γ内的全局渐近稳定性,构造函数L 3=F (v )+p z *(v *+ε)cG (z ),其中F (v )=(v *+ε)l n (v +ε)-v *ln v G (z )=z z *-ln zz * 易知,v =v *是函数F (v )在(0,∞)上的唯一极值点,并且F (v )在v =v *处取到最小值.同样,z =z *是函数G (z )在(0,∞)上的唯一极值点,并且G (z )在z =z *处取到最小值.直接计算可得,函数L 3沿着系统(16)解的全导数为L '3|(16)=F '(v )d v d t +p z *(v *+ε)cG '(z )d z d t =㊃971㊃Copyright©博看网 . All Rights Reserved.陕西科技大学学报第36卷v *+εv +ε-v *æèçöø÷v v '+p z *(v *+ε)c 1z *-1æèçöø÷z z '=-εr k (v -v *)2v +ε-p λc z *(v *+ε)z z*2(z -z *)2因此L '3|(16)关于正平衡点P*是负定的.所以由L y a pu n o v 稳定性定理[14]知,正平衡点P *(v *,z *)在正不变集Γ内是全局渐近稳定的.定理4证毕.4 数值分析与讨论前述定理2㊁3㊁4,从平衡点的存在性㊁局部稳定性和全局稳定性在理论上说明了模型(1)的全局动力学性态.为进一步从数值上进行验证,下面将对r d <p λ,r d =pλ,r d >p λ三种情形予以数值模拟以显示理论结果的正确性.由图1(a )㊁(b )㊁(c )可以看出,对于情形r d <pλ,此时模型的可行平衡点仅为单重无感染平衡点P 0(0,4),是全局渐近稳定的;对于情形r d =pλ,模型的可行平衡点仅为重无感染平衡点P 0(0,4),是全局渐近稳定的;对于情形r d >p λ,模型的可行平衡点有单重无感染平衡点P 0(0,4)和感染平衡点P *(0.1,5.7).这里P 0是不稳定的,P *在可行域内是全局渐近稳定的.(a )情形r d <p λ时的轨线图(k =2㊁p =1㊁λ=2㊁c =2.5㊁ε=1.5㊁d =0.5㊁r =3)(b )情形r d =p λ时的轨线图(k =2㊁p =1㊁λ=2㊁c =2.5㊁ε=1.5㊁d =0.5㊁r =4)(c )情形r d >p λ时的轨线图(k =2㊁p =1㊁λ=2㊁c =2.5㊁ε=1.5㊁d =0.5㊁r =6)图1 r d <p λ㊁r d =p λ㊁r d >p λ三种情形的轨线图根据本文的模型假设,当没有病毒存在z '=λ-d z 时,免疫细胞数量的变化服从于方程,则其最终收敛于z =λd.因此当免疫细胞以处于无病毒时的平衡值杀伤病毒时,对病毒的杀伤率系数就为pλd .此时病毒的内禀增长率即为r -p λd.本文对模型(1)的分析结果显示,r d pλ=1即r -p λd =0是决定模型(1)动力学性态的阈值条件.因此,当免疫系统发生作用时,病毒的内禀增长率决定模型(1)的动力学性态.同时,模型的最终状态是要么收敛于无感染平衡点,即病毒被清除,要么感染持续,病毒载量收敛于v *.病毒被清除,即边界平衡点P 0全局渐近稳定,因此平衡点P 0可称为无感染平衡点,相应的条件为r ≤p λd.而正平衡点P *的全局渐近稳定意味着感染虽然受到免疫应答的抑制,但仍持续存在,因此平衡点P *可称为免疫控制感染平衡点,相应的条件为r >p λd.从这里的两个条件可以看出免疫系统的作用如何决定病毒感染的最终结局.下面进一步考虑在免疫作用下,感染持续并处于平衡状态时病毒载量对各免疫因素p ㊁λ和c 的依赖性.当免疫系统起作用时,若模型(1)的正平衡点存在,则感染平衡点P *(v *,z *)中病毒的分量v *满足方程f (v )=0.此方程也可写为 f 1(v ):=p λr+c v v +æèçöø÷ε1-v æèçöø÷k =0(17) 根据v *的存在范围有c v *v *+ε<d 和v *<k .所以直接计算可显示f '1(v *)>0.进一步,对于由㊃081㊃Copyright©博看网 . All Rights Reserved.第6期李亚男等:一类具有常数免疫输入的病毒-免疫应答动力学模型分析式(17)所确定的隐函数v=v*(p),v=v*(λ)和v =v*(c),有∂v*∂p<0,∂v*∂λ<0以及∂v*∂c<0.由于模型(1)中的参数p㊁λ和c分别表示免疫细胞对病毒的杀伤率系数㊁免疫细胞的输入率和免疫细胞受病毒刺激的最大产生率,所以由∂v*∂p< 0,∂v*∂λ<0以及∂v*∂c<0可知,任何能使得参数p㊁λ和c中任意一个增大的措施都能降低个体体内的病毒载量.图2(a)㊁(b)㊁(c)分别显示了病毒载量平衡值对参数p,λ和c的依赖性.(a)v*对p的依赖(r=6,k=2,λ=2,c=2.5,ε=1.5,d=0.5)(b)v*对λ的依赖(r=6,k=2,p=1,c=2.5,ε=1.5,d=0.5)(c)v*对c的依赖(r=6,k=2,p=1,λ=2,ε=1.5,d=0.5)图2 病毒载量平衡值对参数p、λ和c的依赖性应当注意,根据定理2㊁3㊁4,决定感染能否灭绝的阈值条件不依赖于参数c,这就意味着增大免疫细胞的刺激产生率只会降低病毒载量,不会使得感染灭绝.相反,增大免疫细胞对病毒的杀伤率系数或免疫细胞的输入率都可清除感染.5 结论本文考虑了一类具有常数免疫输入的病毒-免疫应答动力学模型,通过对模型进行分析和研究得到模型的边界平衡点和正平衡点,并对模型的边界平衡点和正平衡点的局部稳定性和全局稳定性进行了详细的分析和研究,发现免疫存在时的病毒内禀增长率决定模型的动力学性态.最后针对各免疫因素对病毒持续的影响进行了讨论.参考文献[1]A n d e r s o nR M,M a y R M.T h e m a t h e m a t i c so f i n f e c t i o n[J].S c i e n c e,1991,254(2):591-592.[2]M A N o w a k,R M M a y.V i r u sd y n a m i c s:M a t h e m a t i c a lp r i n c i p l e s o f i mm u n o l o g y[J].V i r u s R e s e a r c h,2001,80(1):85-86.[3]刘燕明.免疫学与病原生物学[M].北京:中国中医药出版社,2008.[4]P e r e l s o nAS.M o d e l l i n g v i r a l a n d i mm u n e s y s t e md y n a m-i c s[J].N a t u r eR e v i e w s I mm u n o l o g y,2002,2(1):28-36.[5]L i J,W a n g K,Y a n g Y.D y n a m i c a lb e h a v i o r so fa n H B Vi n f e c t i o nm o d e l w i t h l o g i s t i c h e p a t o c y t e g r o w t h[J].M a t h-e m a t i c a la n d C o m p u t e r M o d e l l i n g,2011,54(1-2):704-711.[6]A M E l a i w,N H A l s h a m r a n i.D y n a m i c so f v i r a l i n f e c t i o nm o d e l s w i t h a n t i b o d i e sa n d g e n e r a ln o n l i n e a ri n c i d e n c ea n dn e u t r a l i z e r a t e s[J].I n t e r n a t i o n a l J o u r n a l o fD y n a m i c sa n dC o n t r o l,2016,4(3):303-317.[7]R e g o e sRR,W o d a r zD,N o w a kM A.V i r u s d y n a m i c s:T h ee f f e c t o f t a r g e tc e l l l i m i t a t i o na n di mm u n er e s p o n s e so nv i r u s e v o l u t i o n[J].J o u r n a l o fT h e o r e t i c a lB i o l o g y,1998,191(4):451-462.[8]S h uH,W a n g L,W a t m o u g h J.S u s t a i n e d a n d t r a n s i e n t o s-c i l l a t i o n s a n dc h a o s i nd u ce db y d e l a y e da n t i v i r a l i mm u n er e s p o n s e i na ni mm u n o s u p p r e s s i v ei n f e c t i o n m o d e l[J].J o u r n a l o fM a t h e m a t i c a l B i o l o g y,2014,68(1-2):477-503.[9]B o e rRJD,B o e r l i j s tM C.D i v e r s i t y a n dv i r u l e n c e t h r e s h-o l d s i nA I D S[J].P r o c e e d i n g s o f t h eN a t i o n a lA c a d e m y o fS c i e n c e s,1994,91(2):544-548.[10]K o m a r o v aN L,B a r n e sE,K l e n e r m a nP,e ta l.B o o s t i n gi mm u n i t y b y a n t i v i r a ld r u g t h e r a p y:As i m p l er e l a t i o n-s h i p a m o n g t i m i n g,e f f i c a c y,a n ds u c c e s s[J].P r o c e e d i n g so f t h e N a t i o n a l A c a d e m y o fS c i e n c e s,2003,100(4):㊃181㊃Copyright©博看网 . All Rights Reserved.陕西科技大学学报第36卷1855-1860.[11]李 佳,李建全,李益群.一类带有特异性免疫细胞钟形增殖率的慢性病毒感染模型的全局动力学性态[J ].工程数学学报,2017,34(4):424-436.[12]W o d a r zD ,H a l l SE ,U s u k uK ,e t a l .C y t o t o x i cT -c e l l a -b u n d a nc e a n dv i r u s l o ad i nh u m a n i mm u n o def i c i e n c y v i -r u s t y p e 1a n dh u m a nT -c e l l l e u k a e m i av i r u s t y p e 1[J ].P r o c e e d i n g s B i o l o g i c a l S c i e n c e s ,2001,268(1473):1215.[13]K a z a r i n o f f N D.N o n l i n e a ro s c i l l a t i o n s ,d y n a m i c a ls y s -t e m s ,a n db i f u r c a t i o n so f v e c t o r f i e l d s .b y J o h n g u c k e n -h e i m e r ;p h i l i p ho l m e s [J ].S i a m R e v i e w ,2006,26(4):600-601.[14]S m i t h M C .T h e g e n e r a l p r o b l e mo f t h e s t a b i l i t y o fm o -t i o n [J ].A u t o m a t i c a ,1999,31(2):14-15.【责任编辑:蒋亚儒 】(上接第153页)[6]R u g g i u M ,K o n g X.M o b i l i t y a n dk i n e m a t i ca n a l ys i so f a p a r a l l e lm e c h a n i s m w i t hb o t hP P Ra n d p l a n a ro p e r a t i o n m o d e s [J ].M e c h a n i s m &M a c h i n eT h e o r y,2012,55(9):77-90.[7]王 波,刘向东,韩 强,等.三自由度平面并联机构位姿分析[J ].机械制造与自动化,2012,41(1):170-171.[8]G o s s e l i nC M ,M e r l e t JP .T h e d i r e c t k i n e m a t i c s o f p l a n a rp a r a l l e lm a n i p u l a t o r s :S pe c i a l a r c h i t e c t u r e s a n dn u m b e r of s o l u t i o n s [J ].M e c h a n i s m &M a c h i n e T h e o r y ,1994,29(8):1083-1097.[9]岑豫皖,余晓流,储刘火.三自由度并联姿态测量机构及其运动学参数最优化设计[J ].中国机械工程,2005,16(3):189-191.[10]刘延斌,韩秀英,郭志佳,等.一种新型3-P P R 并联机构及其运动学分析[J ].中国机械工程,2011,22(19):307-311.[11]高晓雪.3-P P R 平面并联机构的运动学和动力学性能研究[D ].太原:中北大学,2014.[12]A l e j a n d r oC .O l i v i e r i ,S a n t i a g oB o r t o l a t o ,F r a n c oA l l e g r i -n i .E r r o rm o d e l i n g a n de x p e r i m e n t a l v a l i d a t i o no f a p l a -n a r 3-P P R p a r a l l e lm a n i p u l a t o rw i t h j o i n t c l e a r a n c e s [J ].J o u r n a l o fM e c h a n i s m s&R o b o t i c s ,2013,4(4):4-8.[13]李增刚.A D AM S 入门详解与实例[M ].2版.北京:国防工业出版社,2006.[14]赵建国,李 兵.一种新型三自由度平面并联机构的运动学研究[J ].机械工程师,2008,13(1):76-78.[15]C h a b l a tD ,W e n g e rP .T h ek i n e m a t i ca n a l y s i so f as y m -m e t r i c a l t h r e e -d e g r e e -o f -f r e e d o m p l a n a r p a r a l l e lm a n i p u -l a t o r [J ].C o m pu t e r S c i e n c e ,2007,46(1):1-7.[16]J i a n M ,Z h a n g D ,L i Z .A c c u r a c y a n a l y s i so f a3-d o f p l a -n a r p a r a l l e l r o b o t [J ].J o u r n a lo fJ i a n g s u U n i v e r s i t y of S c i e n c e&T e c h n o l og y,2009,131(1):889-894.【责任编辑:陈 佳 】(上接第175页)[12]H e r n a n d e zC ,V o g i a t z i sG ,B r o s t o w GJ ,e t a l .N o n -r i g i d p h o t o m e t r i c s t e r e ow i t h c o l o r e d l i g h t s [C ]//I E E EI n t e r -n a t i o n a l C o n f e r e n c e o nC o m pu t e rV i s i o n .R i o d e J a n e i r o :I E E E ,2007:1-8.[13]Y u H ,W i l b u r nB ,M a t s u s h i t aY ,e ta l .E d g e -p r e s e r v i n g p h o t o m e t r i c s t e r e ov i ad e p t hf u s i o n [C ]//I E E E C o n f e r -e n c e o nC o m p u t e rV i s i o n a n dP a t t e r nR e c o g n i t i o n .P r o v -i d e n c e :I E E E ,2012:2472-2479.[14]I k e h a t aS ,W i p fD ,M a t s u s h i t aY ,e t a l .P h o t o m e t r i c s t e r -e ou s i n g s p a r s eb a y e s i a nr e gr e s s i o nf o r g e n e r a ld i f f u s e s u r f a c e s [J ].I E E E T r a n s a c t i o n so n P a t t e r n A n a l y s i s &M a c h i n e I n t e l l i ge n c e ,2014,36(9):1816-1831.[15]P o w e l lM W ,S a r k a rS ,G o l d g o fD.As i m p l e s t r a t e g y fo r c a l i b r a t i n g t h e g e o m e t r y o fl i g h t s o u r c e s [J ].I E E E T r a n s a c t i o n so n P a t t e r n A n a l y s i s &M a c h i n eI n t e l l i -ge n c e ,2001,23(9):1022-1027.[16]S p e n c e A ,C h a n t l e r M.O p t i m a l i l l u m i n a t i o nf o rt h r e e -i m a g e p h o t o m e t r i c s t e r e o a c q u i s i t i o no f t e x t u r e [J ].P r o -g r e s s o fT h e o r e t i c a l P h ys i c s ,2003,153(2):149-159.【责任编辑:陈 佳】㊃281㊃Copyright©博看网 . All Rights Reserved.。
一类分数阶金融风险模型的混沌动力学行为分析及控制

一类分数阶金融风险模型的混沌动力学行为分析及控制作者:张文娟张晓丹张英晗来源:《中国管理信息化》2017年第14期[摘要]本文提出了一类分数阶的金融风险模型。
首先,研究了风险模型对初始值的敏感依赖性;然后,研究了风险模型对风险传递效率及分数阶次的敏感性;最后,采用增加反馈增益矩阵的方法,构造了风险模型的受控系统,数值模拟结果表明该控制方法可以有效地控制混沌。
[关键词]一类分数阶;金融风险模型;混沌动力学行为doi:10.3969/j.issn.1673 - 0194.2017.14.069[中图分类号]F832.5;O415.5 [文献标识码]A [文章编号]1673-0194(2017)14-0-020 引言混沌是由确定性系统产生的随机现象,对初始条件的高度敏感性是其主要特征。
混沌经济学也叫非线性经济学,它是基于非线性动力学方法来解释真实的经济现象的。
因为金融风险总是存在于金融活动中,而且金融风险的存在会扰乱正常的金融秩序。
所以,金融风险管理成为了一个热门的研究课题。
近年来,已经发表了不少关于金融风险管理的文章。
然而,这些模型较少地考虑金融变量的长期记忆性,所以不能更好地反映风险管理过程的真实情况。
与整数阶微积分相比,分数阶微积分最大的优势在于具有记忆效应。
受此启发,本文提出了一类分数阶的金融风险传染模型。
1 分数阶金融风险模型的提出其中,状态变量R,U,V,W分别表示风险发生值,分析值,控制值和传染值。
而且R,U,V,W均是非负的。
参数a>0表示风险分析效率,b>0表示风险传递效率,c∈R表示风险控制的失真系数,d>0表示风险强度和k∈R表示风险传染系数。
2 风险模型对初值的敏感依赖性现在讨论模型(1)的初值敏感性。
取分数阶次为α1=α2=α3=α4=0.95及参数为(a,b,c,d,k)=(10,28,8/3,36,0.5),当初始值分别取(10,10,10,10)和(10,10,10,10.001)时,模型(1)的时域图(见图1)。
病毒传播动力学模型与预测

病毒传播动力学模型与预测病毒传播是一种复杂而又重要的现象,已经给全球带来了巨大的挑战和影响。
了解和预测病毒传播的动力学模型是疫情防控和公共卫生应对的关键。
本文将介绍病毒传播动力学模型的基本原理以及它们在预测疫情发展方面的应用。
传播动力学模型是基于数学和统计学原理建立的一种工具,用于描述病毒在人群中的传播方式、速率和规模等基本特征。
其中最常用的模型是SIR模型,即易感者(Susceptible)、感染者(Infected)、恢复者(Recovered)三个互相转化的状态。
SIR模型基于一个重要的假设,即人群中的易感者不断暴露于感染者,并按照一定速率感染,进而成为感染者。
感染者在一段时间内保持传染性,然后转化为恢复者并具有免疫力。
这种模型通常基于人群之间的联系和传播方式,如近距离接触、空气传播等。
然而,简单的SIR模型并不能完全准确地描述病毒传播的复杂性。
因此,人们发展了各种改进型的模型,如SEIR模型、SIER模型等。
这些模型可以考虑更多因素,如潜伏期、暴露程度、治疗措施等,并且对不同人群之间的传播差异进行建模。
在预测疫情发展方面,传播动力学模型可用于估计病毒的基本传染数R0,即一个感染者平均会感染多少个人。
当R0大于1时,疫情将呈现指数增长;当R0小于1时,疫情将逐渐消退。
这个指标对于评估防控措施的有效性至关重要。
此外,传播动力学模型还可以预测疫情的时间和规模等重要参数。
通过结合实际的流行病数据,如接触网络、人群迁移流动等,可以对整个疫情的传播过程进行模拟和预测。
这些模型可以帮助决策者调整措施、规划医疗资源、制定疫苗接种策略等,从而更好地控制和阻断病毒传播。
然而,传播动力学模型也面临许多挑战和限制。
首先,模型的建立需要大量的数据支持,如感染人数、接触网络等,而这些数据往往很难获取。
其次,模型在不同病毒和人群之间的适用性也存在差异,因此需要根据具体情况进行调整。
最后,模型建立的假设和参数选择也可能引入一定的偏差,影响预测的准确性。
一类考虑抗体免疫反应具有标准传染率的病毒动力学模型

无病平衡点 E 全局稳定 ; 。 如果R > 1则每一起始的病毒感染细胞产生平均大于一个新的感染细胞 , 。 , 病毒
在 最初 时间 内呈 指数 增 长 , 随着未感 染 细胞 数 量下 降 , 毒感 染新 的细胞 的机 会 下降 , 病 病毒 数 量增 长减 慢 ,
最后 系统 经 阻尼 震荡 趋 向于 正平衡 点 E 正平衡 点 稳定 . ,
1 2 平衡 点及 其稳 定性 .
我 们首 先研 究 系统 ( )的平 衡 点存 在情 况 . 1 系统 总存 在 一个 无病 平衡 点 : 一 ( o 0 o =( , , ) Eo T , , ) 0 o ,
此平 衡 点代 表宿 主 体 内不存 在病 毒 的状 态 , 本 再生 数 R。 , R。 1 则系统 存 在一 个正 平衡 点 基 一 若 > ,
率. 被感 染 细胞 以速率
生成, 以速 率 , 亡. 离 病 毒 由被感 染 细 胞 死 亡裂 解 而产 生 , 速 率 为 J死 游 其
*
收 稿 日期 :0 10 —1 2 1-22
作者简介 : 毕
滂 ( 93 )男 , ' 7 - , 山西 省 太 原 市 人 , 师 , 士 , 究 方 向 : 用 数 学 1 讲 硕 研 应
类 考 虑 抗 体 免 疫 反 应 具 有 标 准 传 染 率 的病 毒 动 力 学模 型
一
毕 溺 ,肖湘 萍
( 北 大 学 理 学 院 ,山 西 太 原 中 005) 30 1
摘 要 : 究 了一类考 虑抗体 免 疫反应 的病毒 动 力学模 型的 全局性 态 , 明 了 当基本 再生数 R 研 证 0
≤1时 , 病毒在 体 内清除 , 无病 平衡 点稳 定 ; R > 1时 , 而 。 病毒在体 内持 续 生存 , 正平衡点稳 定.
一类分数阶的病毒动力学模型

一类分数阶的病毒动力学模型庄科俊;温朝晖【摘要】提出了一类分数阶的具有Beddington-DeAngelis非线性发生率的病毒动力学模型,研究了模型解的非负性和有界性,得到了无病平衡点和病毒平衡点的稳定性条件,并给出了相应的数值例子.【期刊名称】《北华大学学报(自然科学版)》【年(卷),期】2013(014)005【总页数】4页(P508-511)【关键词】分数阶;病毒动力学模型;稳定性【作者】庄科俊;温朝晖【作者单位】安徽财经大学统计与应用数学学院应用数学研究所,安徽蚌埠233030;安徽财经大学统计与应用数学学院应用数学研究所,安徽蚌埠233030【正文语种】中文【中图分类】O175.14在研究慢性乙型肝炎病毒携带者的临床试验中,文献[1-2]为了描述宿主体内未感染细胞、感染细胞和游离病毒之间的关系,提出了如下的微分方程模型:其中:x(t),y(t),v(t)分别表示在t时刻未感染的宿主细胞(即肝细胞)、感染的宿主细胞和游离病毒的数量;未感染的肝细胞的产生速率为s,死亡率为dx,感染率为βxv;感染的肝细胞的产生速率为βxv,死亡率为py;游离病毒的产生率为ky,死亡率为uv.这里的参数s,d,β,p,k,u均为正的常数.由模型(1)可知,肝细胞的总数越大,即一个人的肝越大,则越容易被病毒感染,这显然是不合理的[3].因此,需要用饱和发生率去描述正常细胞的感染率[4].基于这些考虑,文献[5]提出了具有Beddington-DeAngelis功能响应的模型:文献[5]通过构造Lyapunov函数,讨论了(2)的非负平衡点的全局稳定性.另外,文献[6-7]还讨论了模型(2)的时滞情形的稳定性.最近,分数阶微积分被广泛应用于各个领域,在建立具有生物学背景的数学模型时,分数阶的微分方程比传统的整数阶微分方程更有优势[8],因此,将分数阶导数引入模型(2),得到下列的分数阶模型:本文采用Caputo定义的α阶微分,即这里n-1lt;α≤n,并记Dα为Caputo分数阶微分算子.关于分数阶微积分的更多性质参见文献[8].本文将讨论模型(3)解的非负性与有界性,以及非负平衡点的稳定性,并通过数值例子验证理论分析的结果.首先给出本文用到的几个重要结论.引理1[9-10] (分数阶Routh-Hurwitz准则) 若分数阶系统在平衡点处的特征方程λ3+a1λ2+a2λ+a3=0的所有根满足arg(λ)≥πα/2,则系统的平衡点是渐近稳定的.记(ⅰ) 若D(P)gt;0,则平衡点局部渐近稳定的充分必要条件是a1gt;0,a3gt;0,a1a2-a3gt;0.(ⅱ) 若D(P)lt;0,a1≥0,a2≥0,agt;0,则当αlt;2/3时平衡点是局部渐近稳定的;若D(P)lt;0,a1lt;0,a2lt;0,αgt;2/3,则平衡点不稳定.(ⅲ) 若D(P)lt;0,a1gt;0,a2gt;0,a1a2-a3=0,则平衡点局部渐近稳定.(ⅳ) 平衡点局部渐近稳定的必要条件是a3gt;0.引理2[11] 对α∈(0,1],假设f(x)∈C[a,b],Dαf(x)∈(a,b].若对∀x∈(a,b),有Dαf(x)≥0,则函数f(x)在[a,b]上是递增的;若对∀x∈(a,b),有Dαf(x)≤0,则函数f(x)在[a,b]上是递减的.定理1 若系统(3)的初值非负,则当tgt;0时,该系统的解唯一,且保持非负、有界.证明:根据文献[12]的定理3.1和注记3.2可知,系统(3)的解存在且唯一.由Dαxx=0=s≥0,Dαyy=0=βxv/(1+ax+bv)≥0,Dαvv=0=ky≥0,根据引理2,系统(3)在非负的初始条件下保持正不变.另外,令σ=min{d,p},由Dα(x+y)x+y=s/σ≤s-σ(x+y)x+y=s/σ=0可知,0≤x+y≤s/σ,且0≤y≤s/σ.另一方面,由Dαvv=ks/(uσ)≤ks/σ-uvv=ks/(uσ)=0可得0≤v≤ks(uσ).证毕.由于分数阶不改变系统的平衡点的个数和位置,因此,当R0=lt;1时,系统(3)仅有一个无病平衡点E0=(s/d,0,0);当R0=gt;1时,系统除了无病平衡点外,还存在正平衡点E1=(x*,y*,v*),其中:x*=,y*=1-,v*=·1-.下面,将利用分数阶的Routh-Hurwitz准则讨论平衡点的稳定性.根据引理1不难发现,当特征根均为负实数时,判别条件arg(λ) ≥πα/2始终成立,从而平衡点渐近稳定;当特征方程有正实根时,平衡点必不稳定;而特征方程有正实部的共轭复根时,只要阶数α较小,仍能保证平衡点的稳定性.定理2 当R0lt;1时,无病平衡点E0渐近稳定;当R0gt;1时,无病平衡点E0不稳定.证明:系统(3)在点E0处的特征方程为当R0lt;1时,三个特征根均为负实数,从而E0渐近稳定;当R0gt;1时,有一个正实根,从而E0不稳定.下面讨论R0gt;1时正平衡点E1的稳定性.通过计算系统(3)在E1处的Jacobian 矩阵,可以得到相应的特征方程为其中:b1=,b2=,b3=,a1=p+u-b1,a2=kb2+pu-ub1-pb1,a3=-dkb2-pub1.根据引理1,可以得到关于正平衡点E1稳定性的结论.证毕.定理3 当R0gt;1时,有如下结论成立:(ⅰ)若判别式D(P)gt;0,则E1渐近稳定的充要条件是a1gt;0,a3gt;0,a1a2gt;a3.(ⅱ)若D(P)lt;0,a1≥0,a2≥0,a3gt;0,0.5lt;αlt;2/3时,则E1渐近稳定. (ⅲ)若D(P)lt;0,a1gt;0,a2gt;0,a1a2=a3,0.5lt;αlt;1时,则E1渐近稳定. (ⅳ)若D(P)lt;0,a1lt;0,a2lt;0,αgt;2/3时,则E1不稳定.为获得分数阶系统(3)的数值解,采用Adams型预估-纠错方法[13].根据文献[1-3],选定参数s=5,d=0.2,a=b=1,p=0.25,k=7.5,u=2.5.取β=0.05时,R0=0.576 923lt;1,此时仅有渐近稳定的无病平衡点E0=(25,0,0),见图1.取β=1时,R0=11.538 5,有两个非负平衡点:不稳定的E0=(25,0,0),正平衡点E1=(4.552 24,16.358 2,49.074 6).借助Mathematica软件,容易算得a1=3.773 5,a2=3.376 09,a3=0.652 391,D(P)=6.270 64gt;0,a1a2-a3=12.087 3gt;0,根据定理3可知,此时E1渐近稳定,见图2.通过图1和图2不难发现,当参数满足一定的条件时,平衡点在整数阶和分数阶情形下具有相同的稳定性.当阶数α距离1越远时,解的曲线和整数阶情形相差也越大.至于α会如何影响病毒模型的动力学性质,还有待进一步研究.【相关文献】[1] Nowak M A,Bonhoeffer S,Hill A M,et al.Viral Dynamics in Hepatitis B Virus Infection[J].Proceedings of the National Academy of Sciences of the United States of America,1996,93:4398-4402.[2] Nowak M A,Bangham C R.Population Dynamics of Immune Responses to Persistent Viruses[J].Science,1996,272:74-79.[3] Min Lequan,Yumei Su,Kuang Yang.Mathematical Analysis of a Basic Virus Infection Model with Application to HBV Infection[J].Rocky Mountain Journal of Mathematics,2008,38:1573-1585.[4] Hews S,Eikenberry S,Nagy J D,et al.Rich Dynamics of a Hepatitis B Viral Infection Model with Logistic Hepatocyte Growth[J].Journal of Mathematical Biology,2009,60(4):573-590.[5] Huang Gang,Ma Wanbiao,Takeuchi Yasuhiro.Global Properties for Virus Dynamics Model with Beddington-Deangelis Functional Response[J].Applied Mathematics Letters,2009,22:1690-1693.[6] Huang Gang,Ma Wanbiao,Takeuchi Yasuhiro.Global Analysis for Delay Virus Dynamics Model with Beddington-Deangelis Functional Response[J].Applied Mathematics Letters,2011,24:1199-1203.[7] Wang Xia,Tao Youde,Song Xinyu.A Delayed HIV-1 Infection Model with Beddington-Deangelis Functional Response[J].Nonlinear Dynamics,2010,62:67-72.[8] Podlubny I.Fractional Differential Equations[M].San Diego:Academic Press,1999.[9] Ahmed E,El-Sayed A M A,El-Saka Hala A A.On Some Routh-Hurwitz Conditions for Fractional Order Differential Equations and Their Applications in Lorenz,Rossler,Chua and Chen Systems[J].Physics Letters A,2006,358:1-4.[10] WANG X Y,HE Y J,WANG M J.Chaos Control of a Fractional Order Modified Coupled Dynamos System[J].Nonlinear Analysis,2009,71:6126-6134.[11] Ding Yongsheng,Ye Haiping.A Fractional-Order Differential Equation Model of HIV Infection of C T-Cells[J].Mathematical and Computer Modelling,2009,50:386-392. [12] Lin Wei.Global Existence Theory and Chaos Control of Fractional Differential Equations[J].Journal of Mathematical Analysis and Applications,2007,332:709-726. [13] Diethelm K,Ford N J,Freed A D.A Predictor-Corrector Approach for the Numerical Solution of Fractional Differential Equations[J].Nonlinear Dynamis,2002,29:3-22.。
一类病毒动力学模型概周期解的存在性

目前宿 主体 内的群 体动力 学研 究 1益 受 重视 , 3 其研究 主要集 中于持 续 性病 毒感 染 n 3.文 献 [ ] -] 1 研究 了 如下一 类具 有周 期免疫 反应 的病毒 动力 学模 型
』 A ) —一, d d x
,
【t fy a 一 ( y o = x —y 6t r l { y )
作者简介 :陈晓英 (9 3一) 女 ,福建诏安人 , 18 , 福州大 学数 学与计算机科学学院助教. 张 娜( 94一) 女 ,山东济宁人 , 18 , 福州 大学数 学与计算机科学学院助教.
i c r e u h w t e f a i i t ft e man r s l . s a r d o tt s o e sbl y o i e u t i o h i h s Ke r s i n e t n mo e ;amo tp r d c s lt n;u i r y a y t t al tb h y wo d :v r i fc i d l l s ei i ou i l a o o o n f ml s mp o c y s i t o il a y
C N a — i g r.Z NG Na HE Xio y n HA
( .C lg 1 o eeo te ai n o p t c ne F zo n e i F zo ,Fj n 3 00 ,hn ; l fMahm t s dC m u r i c, uhuU i  ̄@, u u ua , 5 18 C i ca eSe v h i a 2 h hn o ee F zo n e i , u o , ua 50 2 C i ; .Z i egC lg , uh uU i rt F z u Fj n3 0 0 , n ) c l v sy h i h a
几类分数阶传染病动力学模型研究

几类分数阶传染病动力学模型研究几类分数阶传染病动力学模型研究近年来,随着传染病的不断爆发,传染病动力学模型的研究变得越来越重要。
而传统的传染病动力学模型往往假设传播速率和恢复速率为常数,忽略了真实世界中传染病传播速率和恢复速率的不确定性和复杂性。
为了更准确地描述传染病传播过程,分数阶微积分被引入到传染病动力学模型中。
分数阶微积分是传统微积分的推广,它将微分和积分的次数泛化为实数或复数,并且能够描述非线性和非局域性的动力学过程。
因此,分数阶微积分在传染病动力学模型中的应用具有重要的意义。
分数阶传染病动力学模型主要分为分数阶微分方程和分数阶微分方程组两种类型。
其中,分数阶微分方程主要用于描述单一传染病的传播,而分数阶微分方程组则用于描述多种传染病的联合传播。
在分数阶微分方程中,分数阶导数的引入使得传播速率和恢复速率能够具有记忆性,并且能够更好地反映传染病的非线性和非局域性特征。
例如,在SIR模型中,传染速率和恢复速率可以用分数阶导数来描述,使得传染病传播的过程更加准确和真实。
在分数阶微分方程组中,不同传染病之间的相互作用被引入到模型中,使得传染病之间的协同和竞争关系能够被更好地描述。
例如,在SIRS模型中,分数阶微分方程组可以描述多种传染病在人群中的传播和相互竞争的过程,从而更好地预测和控制传染病的传播。
此外,分数阶传染病动力学模型还可以用于研究传染病的扩散速度和控制措施的优化。
传统的传染病动力学模型往往假设传染病的传播速率和恢复速率为固定值,而实际上传染病的传播速率和恢复速率会受到很多因素的影响,例如人群行为、环境条件等。
分数阶传染病动力学模型可以用于描述传染病的传播速率和恢复速率的变化趋势,并且可以通过优化控制参数来减缓传染病的扩散速度。
总之,分数阶传染病动力学模型在描述传染病传播过程和预测传染病传播趋势方面具有重要的意义。
分数阶微积分的引入使得传染病动力学模型具备了更好地描述非线性和非局域性动力学过程的能力,并且能够更准确地反映传染病的传播特征。
一类具有饱和传染率的分数阶SIR传染病模型的动力学性质

一类具有饱和传染率的分数阶SIR传染病模型的动力学性质李明山;张渝曼;梁凤;黄静珊;李晓培
【期刊名称】《湛江师范学院学报》
【年(卷),期】2017(038)003
【摘要】研究了一类具有饱和发生率分数阶SIR传染病模型的动力学性质.首先介绍了分数阶微分方程的基本性质与相关引理.然后通过分析特征值与系数参数的关系得到了平衡点稳定性.接着证明了该模型一致稳态正解的存在唯一性,最后考虑了具有饱和治疗函数的分数阶SIR传染病模型的平衡点稳定性及该模型一致稳态正解的存在唯一性.
【总页数】10页(P32-41)
【作者】李明山;张渝曼;梁凤;黄静珊;李晓培
【作者单位】岭南师范学院数学与统计学院,广东湛江 524048;岭南师范学院数学与统计学院,广东湛江 524048;岭南师范学院数学与统计学院,广东湛江524048;岭南师范学院数学与统计学院,广东湛江 524048;岭南师范学院数学与统计学院,广东湛江 524048
【正文语种】中文
【中图分类】O175
【相关文献】
1.一类具有饱和传染率、免疫接种和垂直传染的SIR传染病模型的全局稳定性分析[J], 朱春娟;
2.一类具有垂直传染和连续治疗的SIRS传染病模型的动力学性质 [J], 宋志强;李明山;周效良
3.一类具有饱和传染率、免疫接种和垂直传染的SIR传染病模型的全局稳定性分析[J], 朱春娟
4.一类具有饱和发生率的SIRS传染病模型的全局性分析 [J], 崔倩倩;张强
5.一类具有饱和传染率的分数阶SIR传染病模型的动力学性质 [J], 李明山;张渝曼;梁凤;黄静珊;李晓培;
因版权原因,仅展示原文概要,查看原文内容请购买。
HIV病毒动力学模型

-
-d T T V 0
dT * TV T * dt dV N T * cV dt
TV T * 0
N T * cV 0
T
d V
T * TV
V N T * c
T
c
,T * (R0 1)
dc
,V
(R0 1)
d
N
N
完整ppt课件
模型推广
在上述中的λ表示个体内如胸腺产生新的 CD4+T细胞的速率,另外CD4+T细胞还可以通过自 身的分裂进行增值.现在我们在此基础上考虑CD4+ 细胞以Logistic形式的增长模式,建立模型:
模型有两个平衡点,E0(T ,0,0), E(T ,T *,V ),其中
T
(Nci),T*
cV
(Ni)
,
V
f (T )
T
完整ppt课件
模型推广
平衡点的求解
dT f (T ) TV
dt
dT* TV T *
dt
dV NT * cV iTV
dt
f (T ) T V 0
TV T * 0
HIV病毒动力学模型 及基本再生数R0计 算,MATLAB仿真
杜金霄 150610309
完整ppt课件
1
1 HIV病毒动力学模型 2 基本再生数R0的计算 3 模型推广 4 MATLAB仿真
完整ppt课件
(一)模型的背景
病毒体在细胞外是处于静止状态,本身不能 复制,但是一旦有适宜它们生长的环境,这 些病毒就会侵入细胞,而细胞的分裂就会产 生更多的病毒,这些细胞继而又会感染更多 的细胞。HIV就是一种逆转录病毒,CD4+T 淋巴细胞是HIV感染的主要靶细胞,而其本 身又是免疫反应的中心细胞,所以CD4+细 胞的数量和病毒载量的检测对艾滋病治疗效 果的判断和对患者免疫功能的判断有重要作 用。该模型就是分析CD4细胞和病毒数量在 感染者体内动态变化
几类新型传染病模型动力学分析及其研究

几类新型传染病模型动力学分析及其研究几类新型传染病模型动力学分析及其研究摘要:传染病是人类社会面临的重大威胁之一,为了更好地理解传染病的传播机理,科学家们发展了多种数学模型来进行动力学分析。
本文将介绍几类新型传染病模型的动力学分析及相关研究。
1. SI模型SI模型是一种最简单的传染病模型,它假设人群分为易感者(Susceptible)和感染者(Infectious)两类。
传染病的传播通过感染者直接接触易感者而发生,易感者一旦被感染则成为感染者,不具有自愈能力。
传播动力学分析表明,SI模型中传染病的传播速度取决于感染率和易感者的数量。
2. SIR模型SIR模型是建立在SI模型基础上的一种改进模型,它引入了恢复者(Recovered)的概念。
恢复者指的是被感染后经过一定时期的治疗或免疫后康复的人群。
SIR模型考虑到了恢复者对传染病传播的抑制作用。
动力学分析结果显示,恢复率对传染病传播的速度有重要影响,较大的恢复率可以降低传染病流行的程度和速度。
3. SEIR模型SEIR模型进一步引入了潜伏期(Exposed)的概念,潜伏期指的是人群被感染后,病毒在人体内潜伏的时间。
在这段时间内,患者可能没有明显的症状,但却具有传染性。
SEIR模型的动力学分析表明,潜伏期的长短对传染病传播的速度和规模有重要影响。
4. SEIRS模型SEIRS模型是在SEIR模型的基础上加入了免疫失效(Susceptible to Exposed to Infectious to Recovered to Susceptible)的过程。
免疫失效指的是人群在恢复一定时间后,再次成为易感者的过程。
动力学分析研究表明,免疫失效的存在使得传染病流行时传染者的数量会呈现周期性变化。
5. SIQR模型SIQR模型是一种考虑隔离的传染病传播模型。
在这个模型中,除了易感者、感染者和康复者,还引入了隔离者(Quarantined)的概念。
隔离者是被感染者或疑似感染者通过隔离措施被特别关注和控制的人群。
1传染病动力学模型简介

传染病动力学模型简介摘要:应用传染病动力学模型可描述疾病发展变化的过程和传播规律,预测疾病发生的状态,评估各种控制措施的效果,为预防控制疾病提供决策依据。
本文介绍传染病动力学的最基本模型――SIR模型,综述了各种传染病模型在医学领域的应用,探讨传染病动力学模型的发展进程和研究动向。
关键词:传染病;动力学模型;SIR模型A brief introduction to dynamics model of infectiousdiseasesAbstract:The dynamics models of infectious diseases can be used to describe the spread characters of infectious diseases, predict the status of the infection and evaluate the efficacy of control strategies, which are useful tools in diseases control decision making. A brief introduction to the basic dynamics model SIR was made, and we also reviewed the application of several dynamic models and discussed its future direction in the paper.Key words: epidemic; dynamic model; SIR model传染病和新出现的疫病严重危害人类健康与社会经济发展。
对传染病发病机理、传播规律和防治策略研究的重要性日益突出。
目前,对传染病的研究方法主要有描述性研究、分析性研究、实验性研究和理论性研究。
传染病动力学[1]是对进行理论性定量研究的一种重要方法,是根据种群生长的特性,疾病的发生及在种群内的传播、发展规律,以及与之有关的社会等因素,建立能反映传染病动力学特性的数学模型,通过对模型动力学性态的定性、定量分析和数值模拟,来分析疾病的发展过程,揭示流行规律,预测变化趋势,分析疾病流行的原因和关键因素,寻求预防和控制的最优策略,为防制决策提供理论依据。
抗病毒药物治疗人类免疫缺陷病毒1型(HIV-1)的分数阶模型研究

抗病毒药物治疗人类免疫缺陷病毒1型(HIV-1)的分数阶模型研究崔周进;陈桂东【期刊名称】《杭州师范大学学报(自然科学版)》【年(卷),期】2014(13)4【摘要】讨论了一类分数阶的抗病毒药物治疗人类免疫缺陷病毒1型模型,对分数阶模型的基本动力学行为,如平衡点、稳定性进行研究,并进行数值模拟验证理论分析的结果。
%A fractional model of anti-viral drug treatment for HIV-1 was proposed in this study .The basic dynamical behaviors of this fractional model were investigated ,such as the equilibrium andstability .Furthermore ,the numerical simulations were used to illustrate and verify the results .【总页数】4页(P406-409)【作者】崔周进;陈桂东【作者单位】江苏海事职业技术学院基础部,江苏南京 211170;解放军理工大学理学院数学教研中心,江苏南京 211101【正文语种】中文【中图分类】O175.1【相关文献】1.慢性乙肝核苷类抗病毒药物治疗价值评估模型研究 [J], 胡善联2.一类时滞分数阶计算机病毒模型的Hopf分岔研究 [J], 高燕鑫;石剑平3.分数阶可学习CPG模型的研究 [J], 刘傲;董诗康;董有为;陆强4.一类时滞分数阶计算机病毒模型的Hopf分岔研究 [J], 高燕鑫;石剑平5.混凝土中变扩散系数的时间分数阶氯离子扩散模型研究 [J], 王一晗;白羽;张艳;巩璇因版权原因,仅展示原文概要,查看原文内容请购买。
分数阶动力学模型

分数阶动力学模型一、引言分数阶动力学模型是一种描述非线性系统行为的数学模型,它考虑了系统动力学的非整数阶特性。
相比传统的整数阶动力学模型,分数阶动力学模型能更准确地描述一些复杂系统的行为。
二、分数阶微积分基础1. 分数阶导数和积分在整数阶微积分中,导数和积分是通过整数次幂的幂函数来定义的。
而在分数阶微积分中,导数和积分是通过分式幂函数来定义的。
分数阶导数和积分可以用分数阶微分方程来描述系统的行为。
2. 分数阶微分方程分数阶微分方程是一种包含了分数阶导数的微分方程。
它可以描述一些非线性系统的演化规律。
分数阶微分方程的求解通常需要使用迭代方法或数值计算方法。
三、分数阶动力学模型的应用1. 生物学领域分数阶动力学模型在生物学领域的应用非常广泛。
例如,分数阶微分方程可以用来描述肿瘤生长的动力学过程,从而帮助医生更好地了解肿瘤的发展规律。
2. 经济学领域分数阶动力学模型在经济学领域的应用也得到了广泛关注。
例如,分数阶微分方程可以用来描述股票市场的波动性,从而帮助投资者制定更科学的投资策略。
3. 物理学领域分数阶动力学模型在物理学领域的应用也是非常重要的。
例如,分数阶微分方程可以用来描述粒子在非整数阶力场中的运动规律,从而帮助科学家更深入地理解粒子的行为。
四、分数阶动力学模型的优势和挑战1. 优势分数阶动力学模型能更准确地描述一些复杂系统的行为,对于一些非线性系统的建模和分析具有重要的意义。
它可以帮助科学家们更深入地理解系统的动力学特性。
2. 挑战分数阶动力学模型的求解相对复杂,需要使用迭代方法或数值计算方法。
此外,分数阶微分方程的数学理论和方法相对较新,仍然存在很多待解决的问题和挑战。
五、总结分数阶动力学模型是一种描述非线性系统行为的重要数学模型。
它在生物学、经济学和物理学等领域的应用具有重要的意义。
虽然分数阶动力学模型的求解相对复杂,但它能更准确地描述系统的行为,对于科学研究具有重要的价值。
注意:本文档旨在提供关于分数阶动力学模型的基本概念和应用领域的介绍,不涉及具体的数学推导和求解方法。
一类分数阶HIV模型的稳定性分析
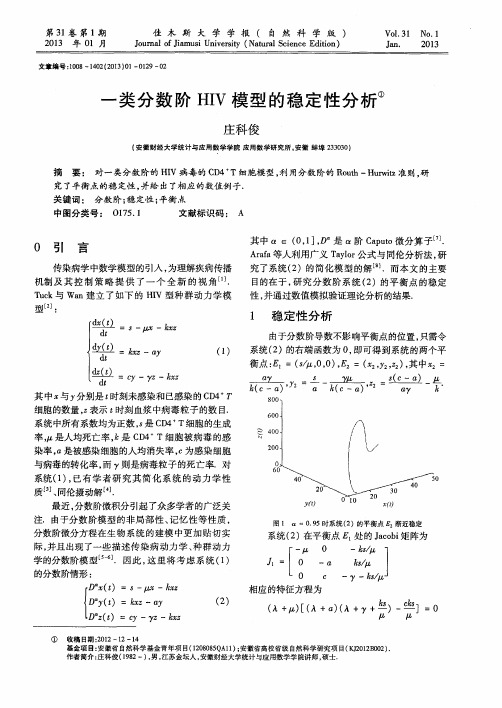
( 2 )
( A+ ) [ ( A+0 ) ( A+ +堕)一 坐 ]=0
收稿 日期 : 2 0 1 2—1 2—1 4 基金项 目: 安徽省 自然科 学基金青年项 目( 1 2 0 8 0 8 5 Q A l 1 ) ; 安徽省高校省级 自然科学研究项 目( K J 2 0 1 2 B 0 0 2 ) 作者简介 : 庄 科俊 ( 1 9 8 2一) , 男, 江苏金坛人 , 安徽 财经大学统计与应用数学学院讲师 , 硕士.
V o 1 . 3 1 No . 1
J a n .
2 0 1 3
文章编号 : 1 0 0 8—1 4 0 2 ( 2 0 1 3 ) 0 1-0 1 2 9— 0 2
一
类分数 阶 H I V模 型 的稳定 性 分 析①
庄科俊
( 安徽财经大学统计与应用数学学院 应用数学研究所 。 安徽 蚌埠 2 3 3 0 3 0)
文 献标 识码 : A
0 引 言
传染病学中数学模型的引入 , 为理解疾病传播 机制及 其 控制 策 略 提供 了一 个 全 新 的 视角….
T u c k 与 Wa n建立了如下 的 H I V型种群动力学模
型 :
= s 一 一
其中 ∈ ( 0 , 1 ] , D 是 阶 C a p u t o 微分 算子 . A r a f a 等人利用广义 T a y l o r 公式与同伦分析法 , 研 究了系统( 2 )的简化模型 的解㈨.而本文的主要
=
音一
=
一
其 中 与 Y分别是 t 时刻未 感 染和 已感 染 的 C D 4 T 细胞 的数 量 , 表 示 t 时刻 血浆 中病毒 粒 子 的数 目. 系 统 中所 有 系数均 为正 数 , s 是C D 4 T细胞 的生成
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Vi r u s Dy n a mi c s Mo d e l wi t h a Cl a s s o f Fr a c t i o n a l — o r d e r
Z HU A N G K e - j u n , WE N Z h a o — h u i
0c t . 2 01 3
文章 编 号 : 1 0 0 9 48 2 2 ( 2 0 1 3 ) 0 5 - 0 5 0 8 0 - 4
D OI : 1 0 . 1 1 7 1 3 / j . i s s n . 1 0 0 9 48 2 2 . 2 0 1 3 . 0 5 . 0 0 3
Ab s t r a c t:A c l a s s o f f r a c t i o n a l — o r d e r v i r u s d y n a mi c s mo d e l wi t h Be d d i ng t o n — De Ang e l i s no n l i n e a r i n c i d e n c e r a t e i s p u t f o r wa r d. Th e p o s i t i v i t y a n d b o u n d e d n e s s o f s o l u t i o ns a r e i n v e s t i g a t e d. T he s t a b i l i t y c o n d i t i o n s o f u n i n f e c t e d
d£
d v i Fra bibliotek0f y —U V,
其 中: ( t ) , Y ( t ) , ( t )分 别表示 在 t 时刻 未感染 的 宿主细 胞 ( 即肝 细胞 ) 、 感染 的宿 主细胞 和 游离病 毒 的数 量; 未 感染 的肝 细胞 的产 生速 率为 s , 死 亡率 为 , 感 染率 为 t G x v ; 感染 的肝 细胞 的产 生速 率 为 为p Y ; 游离 病毒 的产 生率 为 , 死 亡率 为 U V . 这里 的参数 s , d , , P , , “均 为正 的常 数.
由模 型 ( 1 )可 知 , 肝 细胞 的总 数越 大 , 即一个 人 的 肝越 大 , 则越 容易被病毒感 染 , 这显然是 不合理 的 .
, 死 率
因此 , 需 要 用 饱 和 发 生率 去描 述 正 常 细 胞 的 感 染 率 . 基 于这 些 考 虑 , 文献 [ 5 ]提 出 了具 有 B e d d i n g t o n 一
一
类 分 数 阶 的 病 毒 动 力 学 模 型
庄科俊 , 温朝 晖
( 安 徽 财 经 大 学 统 计 与 应 用 数 学 学 院应 用 数 学 研 究 所 , 安徽 蚌埠
2 3 3 0 3 0 )
摘要 : 提出了一 类分 数阶的具有 B e d d i n g t o n — D e A n g e l i s 非线性发生 率的病毒 动力学模 型, 研究 _ r 模 型解的非 负性 和有界性 , 得到了无病平衡点和病 毒平衡 点的稳 定性 条件 , 并给 出了相应 的数值例子. 关键词 : 分数阶 ; 病毒动力学模型 ; 稳定性
1 引
言
在 研究 慢性 乙型肝 炎病 毒携 带者 的临 床试验 中 , 文献 [ 1 . 2 ] 为 了描述 宿主 体 内未 感 染 细胞 、 感 染 细 胞 和游离 病毒 之 间的关 系 , 提 出 了如 下 的微分 方程模 型 :
d x
一 一
n
,
:
一
( 1 )
第 1 4卷 第 5期
北华大学学报 ( 自然科学版 )
J O U R N A L O F B E I H U A U N I V E R S I T Y( N a t u r a l S c i e n c e )
Vo 1 . 1 4 No . 5
2 0 1 3年 l O月
a n d i n f e c t e d e q ui l i b r i a a r e o b t a i n e d. S o me n ume r i c a l e x a mp l e s a r e g i v e n.
Ke y wo r ds:f r a c t i o na l — o r d e r; v i us r d y n a mi c s mo de l ; s t a b i l i t y
收稿 日期 : 2 0 1 3 4 ) 4 . 2 5 基金项 目: 安徽省 自然科学基金青年项 月( 1 2 0 8 0 8 5 Q A1 1 ) ; 安徽省高校省级 自然科学研究项 目( K J 2 0 1 2 A 0 0 1 , K J 2 0 1 3 B 0 0 3 ) 作者简介 : 庄科俊 ( 1 9 8 2一) , 男, 讲帅 , 主 要 从 事 生 物 数 学研 究 .
( I n s t i t u t e o f A p p l i e d Ma t h e m a t i c s , S c h o o l o f S t a t i s t i c s a n d A p p l i e d Ma t h e m a t i c s A n h u i U n i v e r s i t y f o F i n a n c e a n d E c o n o m i c s , B e n g b u 2 3 3 0 3 0, C h i n a )