三羧酸循环@生物化学精品讲义
中国海洋大学生物化学16 三羧酸循环-课件
海洋生命学院 32
苹果酸 三羧酸循环
2009-11-27
只有一些植物和微生物兼具有这样的 途径;
异柠檬酸裂解酶
柠檬酸合成酶
2009-11-27
海洋生命学院
33
乙醛酸循环的总反应:
2乙酰-CoA+NAD++2H2O→琥珀酸+2CoA+NADH+H+ 或2乙酰-CoA+2NAD++FAD→草酰乙酸+2CoA+2NADH+FADH2+2H+
2009-11-27 海洋生命学院 14
5、琥珀酰- CoA转化成琥珀酸
琥珀酰CoA合成酶
2009-11-27
海洋生命学院
15
催四 化聚 反体 应 可 逆 • 该反应特点 • 琥珀酸CoA合成酶(或琥珀酸硫激酶)催化 • 唯一底物磷酸化反应,产生1摩尔ATP
2009-11-27 海洋生命学院 16
2009-11-27
海洋生命学院
18
7、延胡索酸水合形成L-苹果酸
延胡索酸酶
四个亚基 活性中心-SH 具有立体专一性,产生L-苹果酸
2009-11-27 海洋生命学院 19
8、L-苹果酸脱氢形成草酰乙酸
苹果酸脱氢酶
2009-11-27
海洋生命学院
20
第苹 四果 次酸 氧脱 化氢 还酶 原 反 应
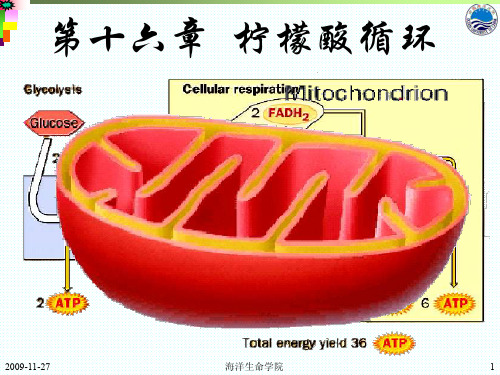
第十六章 柠檬酸循环
2009-11-27
海洋生命学院
1
• 三羧酸循环是由德国科学家 Hans Krebs于1937年提出, 生物化学领域的重大成就(当 时还没有同位素示踪法)。 Krebs于1953年获得诺贝 尔奖。 • 三羧酸循环 • 柠檬酸循环 • Tricarboxylic acid cycle(TCA cycle) • Krebs循环
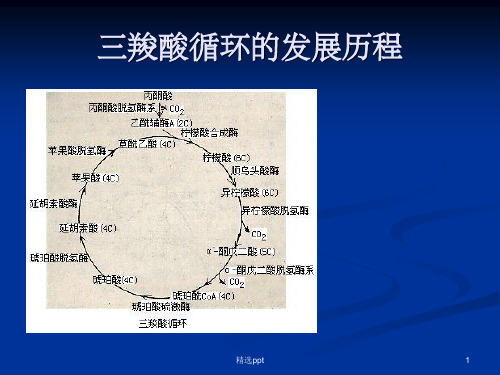
三羧酸循环的发展历程ppt课件
精选ppt
14
1948年,立体化学家奥格斯坦通过精辟的理 论分析认为,在不对称酶的“攻击”下,对称 的柠檬酸也可能表现出不对称性,从而又为 柠檬酸循环“恢复了名誉”。这就是柠檬 酸循环和三羧酸循环两个名称的来历。
休。
1958年受封为骑士,1979年获剑桥大学荣 誉博士学位。1981年逝世于牛津。
精选ppt
4
总论
克雷布斯教授的研究主要涉及的中间代谢 的各个方面。他研究的对象之一是在肝脏 合成尿素,尿酸和鸟嘌呤的合成,食品的 氧化的中间阶段,对电解质的主动运输机 制和细胞呼吸和磷酸腺苷的生成之间的关 系。
他的许多著作中是生物能量转换的显著的 调查。另外,他与H. L.科恩伯格合作讨论 了复杂的化学过程,提供生物体与高能磷 酸通过所谓的克雷布斯或柠檬酸循环。
精选ppt
6
三羧酸循环的发现历程
人们早在十八世纪就已注意到食物在生物 体内要经过一个缓慢“燃烧”的过程—氧化。
但直到二十世纪三十年代,生物氧化还是一 个 “剪不断、理还乱”的谜团。1932年后,经过众 多科学家的努力,特别是德国科学家迈耶霍夫 (OrorMeyerhof,1884一1951)等人的杰出贡献, 搞清了生物发酵—无氧氧化的具体步骤,称为 糖酵解途径(EMP)
精选ppt
13
但是在草酰乙酸和丙酮酸合成产物上却和
克雷布斯有根本分歧,根据他们的分析,柠檬 酸是对称性分子,由它所得到的两种。α一酮 戊二酸中所标记的碳原子也应是对称分布 的,但实验结果相反,被标记的分子只在一种 α一酮戊二酸中分布。因此他们认为草酰乙 酸和丙酮酸的合成产物不是对称性的柠檬 酸,而是非对称的顺乌头酸或异柠檬酸。
生物化学:第十章 三羧酸循环

The Citric Acid Cycle Oxidizes Two-Carbon Units
The cycle starts with the condensation of oxaloacetate (C4) and acetyl CoA (C2) to give citrate (C6), which is isomerized to isocitrate (C6). Oxidative decarboxylation of this intermediate gives –ketoglutarate (C5). The second molecule of carbon dioxide comes off in the next reaction, in which -ketoglutarate is oxidatively decarboxylated to succinyl CoA (C4). The thioester bond of succinyl CoA is cleaved by inorthophosphate to yield succinate, and a high phosphoryl transfer potential compound in the form of GTP is concomitantly generated. Succinate is oxidized to fumarate (C4), which is then hydrated to form malate (C4). Finally, malate is oxidized to regenerate oxaloacetate (C4). Thus, two carbon atoms from acetyl CoA enter the cycle, and two carbon atoms leave the cycle as CO2 in the successive decarboxylations catalyzed by isocitrate dehydrogenase and ketoglutarate dehydrogenase. In the four oxidation–reduction reactions in the cycle, three pairs of electrons are transferred to NAD and one pair to FAD. These reduced electron carriers are subsequently oxidized by the electrontransport chain to generate approximately 9 molecules of ATP. In addition, 1 molecule of a compound having a high phosphoryl transfer potential is directly formed in the citric acid cycle. Hence, a total of 10 molecules of compounds having high phosphoryl transfer potential are generated for each two-carbon fragment that is completely oxidized to H2O and CO 2. 44 浙江大学医学院徐立红
第4章第2节三羧酸循环

NAD+
NADH+H+ CO2
-酮戊二酸 脱氢酶
李新梅 生命科学与技术研究院
李新梅 生命科学与技术研究院
4C阶段
琥珀酸硫激酶
– (1)在好朋友琥珀酸硫激酶的帮助下变成琥珀 酸,同时释放一分子的GTP
乙酰CoA
李新梅 生命科学与技术研究院 李新梅 生命科学与技术研究院
7
–琥珀酰辅酶A生成琥珀酸,并生成GTP
2、什么是泛酸?
泛酸又称B5,什么是泛酸?
– α,γ-二羟基ββ-二甲基丁酸+β丙氨 酸 – 构成了重要的代谢中间产物乙酰CoA
蜂王浆中泛酸含量丰
富
– 青年工蜂咽头腺的分泌物, 是供给将要变成蜂王的幼 虫的食物 – 蛋白质约占蜂王浆干物质 的50% – 蜂王浆含有较多的维生素, 尤其是B族维生素特别丰 富
生物素 丙酮酸羧化酶
天冬氨酸 谷氨酸 丙酮酸 丙氨酸 COOH CH2 CO COOH 草酰乙酸
李新梅 生命科学与技术研究院
李新梅 生命科学与技术研究院
7、TCA在代谢中的地位:是糖、脂、蛋 白质三大物质代谢的共同通路
– 柠檬酸循环是有氧分解代谢的枢纽
• 糖、脂肪和氨基酸的有氧分解代谢都汇集在柠檬 酸循环的反应
乙酰辅酶A
李新梅 生命科学与技术研究院 李新梅 生命科学与技术研究院
5、泛酸缺乏症
爱荷华州立医院所做的人体泛酸缺乏实验
– 年轻的受试者有倦怠、头痛、眩晕、虚弱、心 跳加速、抽筋、持续感冒、上呼吸道感染疾病 等症状;并且变得沮丧不安、消沉、怨恨、暴 躁易怒、挑衅 – 那些饮食营养均衡,只缺乏泛酸的受试者,在6 个星期之后,健康状况极度恶化,即使每日补 充4000毫克泛酸及可的松,仍然恢复得很慢 – 身体的每一个细胞都需要泛酸,否则无法将糖 或脂肪转化为能量
三羧酸循环(TCA)(课堂PPT)

医学应用
进一步探索三羧酸循环在疾 病诊断和治疗中的潜力,如代 谢性疾病和肿瘤等。
三羧酸循环相关的实验技术
色谱技术
利用液相色谱和气相色谱检 测三羧酸循环中的中间体和 相关代谢产物。可定量分析 各种酶促反应的变化。
光谱分析
采用紫外-可见分光光度法和 核磁共振波谱法测定三羧酸 代谢物的浓度和结构。能更 精确地监测循环中各步反应 。
三羧酸循环的研究发展历程
1937年
汉斯·克雷布斯发现并描述了三羧酸循环的化学过程,为生物化学领域带来 了重大突破。
1970年代
电子传递链的发现推动了三羧酸循环与细胞呼吸的联系,为能量代谢的理解 奠定了基础。
1
2
3
1940年代
研究人员通过同位素示踪实验进一步证实了三羧酸循环的反应机理,并揭示 了其在代谢过程中的中心地位。
图示分析
通过生动形象的图示,帮助学生 直观地理解三羧酸循环的复杂 过程。
互动讨论
鼓励学生积极参与讨论,分享见 解,加深对三羧酸循环的理解。
实际应用
解释三羧酸循环在生物医学、 工业生产等领域的广泛应用,增 强学生的兴趣。
结语及问答环节
通过对三羧酸循环的深入探讨,我们对这一重要代谢过程有了更全面的认知。 让我们总结一下关键要点,并开放现场提问,以加深对这一主题的理解。
三羧酸循环中的关键中间体
柠檬酸
异柠檬酸
作为三羧酸循环的第一个中间体,它为 它在三羧酸循环中起到了关键的催化
后续反应提供了重要的碳骨架。
作用,调节了整个循环的速率。
α-酮戊二酸
这一中间体在三羧酸循环中起核心作 用,是其他氨基酸合成的前体。
琥珀酰-CoA
这一重要的中间体连接了三羧酸循环 与电子传递链,产生ATP。
生物化学ii(苏维恒)20130329-三羧酸循环

编辑版ppt
15
乙酰二氢硫辛酰胺
乙酰CoA 二氢硫辛酰胺
二氢硫辛酰胺转乙酰酶还催化乙酰硫辛酰胺上的乙酰基转 移给辅酶A生成乙酰辅酶A后,离开酶复合体,同时氧化 过程中的2个电子使硫辛酰胺上的二硫键还原为2个巯基。
编辑版ppt
16
硫辛酸
编辑版ppt
17
编辑版ppt
18
砷化物对硫辛酰胺的抑制作用
编辑版ppt
19
甘油醛-3-磷酸氧化成1, 3-二磷酸甘油酸
甘油醛-3-磷酸
甘油醛-3-磷酸 脱氢酶
糖酵解中唯一的 脱氢反应
1,3-二磷酸甘油酸
砷酸盐是磷酸的类似物,可以代替磷酸结合到甘油酸 的1位,并很快水解,使得不能形成1,3-二磷酸甘油酸, 不能产生ATP,导致解偶联。
编辑版ppt
20
还原型E2被氧化反应(三)
O
葡萄糖
C==O
CH3-C-SCoA
CH3 CoASH
乙酰CoA
CO2
丙酮酸
NAD+ NADH+H+
三羧酸 循环
葡萄糖的有氧分解
编辑版ppt
3
➢概念
有氧氧化是指体内组织在有氧条件下,葡萄糖彻底氧化分 解生成CO2和H2O的过程。
C6H12O6 + 6O2
6 CO2 + 6 H2O + 30/32 ATP
丙酮酸脱 氢酶系
三种酶
E1-丙酮酸脱羧酶(也叫丙酮酸脱氢酶) E2-二氢硫辛酸乙酰基转移酶 E3-二氢硫辛酰胺脱氢酶。
五种辅助因子 焦磷酸硫胺素(TPP)、硫辛酸、 COASH、FAD、NAD+
编辑版ppt
10
丙酮酸脱氢酶复合体的组成
三羧酸循环讲课文档

(3). 异柠檬酸的转变有两条途径:一是当需要能量时,进行氧化
脱羧形成-酮戊二酸;二是在能量充足时,经异柠檬酸裂解
酶作用,生成琥珀酸和乙醛酸。
第二十七页,共48页。
4. α-酮戊二酸氧化脱羧成为琥珀酰-CoA
4.1 反应方程式
第二十八页,共48页。
4.2 反应机制
α-酮戊二酸脱氢酶系催化的每步机制和丙酮酸脱 氢酶复合体相一致,需要TPP、硫辛酸、CoA、FAD、NAD+、Mg2 +6种辅助因子。
还原型乙酰 硫辛酰胺
乙酰辅酶A
还原型 硫辛酰胺
羟乙基-TPP
氧化型硫辛酰胺
第二步反应, 在二氢硫辛酰转乙酰基酶分子E2上结合着的乙酰基,由E2催化,将乙酰基转移 到CoA-SH分子上,形成游离的乙酰-CoA分子,从而使二氢硫辛酰转乙酰基酶E2由氧化型变成 还原型。
第十二页,共48页。
2.2.1 反应机制
氧化型硫辛酰胺
这一步反应是使氧化型硫辛酰胺再生的反应。
第十四页,共48页。
2.3.1反应机制
这一步反应使氧化型硫辛酰胺再生,在此反应中,催化此反应的酶为二 氢硫辛酸脱氢酶E3(其辅基为FAD),使二氢硫辛酰胺再氧化,从而使其完
成整个反应过程,重新形成氧化型二氢硫辛酰转乙酰基酶。
活泼
互換反应
氧化型二氢硫辛酸 还原型二氢硫辛酰
丙酮酸-TPP加成物脱羧,形成羟乙基-TPP,由于TPP环上带正电荷 的氮原子起电子“陷井”作用,使脱羧后形成的羟乙基上产生较稳定的
负碳离子。
起电子“陷井”作用
丙酮酸-TPP加成物‧E1
较稳定的负碳离子
羟乙基-TPP-E1 (暂时稳定的共振形式)
第十页,共48页。
大学精品课件:第10章 三羧酸循环

三羧酸循环
Tricarboxylic Acid Cycle
的发现
Discovery of the Citric Acid Cycle
2
一、三羧酸循环是三类营养物质氧化分解的 (共同)第二阶段
* 营养物在生物体内氧化的一般过程 糖原 三酯酰甘油 蛋白质
5
1937 年, Hans Krebs 利用鸽子胸肌(这块肌
肉在飞行中有相当高的呼吸频率,因此特别适合
于氧化过程的研究)的组织悬液,测定了在不同 的有机酸作用下,丙酮酸氧化过程中的耗氧率, 首次提出在动物组织中丙酮酸氧化途径的假说。
6
Albert Szent-Gyorgyi等已经发现动物肌肉组
葡萄糖 脂酸 + 甘油 氨基酸
乙酰CoA
TCA 循环
CO2 2H
3
ADP+Pi
ATP
H2O
呼吸链
在真核生物,TCA循环在线粒体中进行, 与呼吸链在功能和结构上相偶联。
4
二、Krebs发现三羧酸循环
三羧酸循环亦称柠檬酸循环
(citric acid cycle),这是因 为循环反应中的第一个中间产物是 一个含三个羧基的柠檬酸 。由于 Krebs 正式提出了三羧酸循环的 学说,故此循环又称为 Krebs 循 环。
GDP
ADP
ATP
(六)琥珀酸脱氢生成延胡索酸
• 由琥珀酸脱氢酶催化,其辅酶是FAD,是三羧酸 循环中唯一与内膜结合的酶。
20
(七)延胡索酸加水生成苹果酸
• 延胡索酸酶催化此步反应
21
(八)苹果酸脱氢生成草酰乙酸
苹果酸脱氢酶催化,辅酶是NAD+。
三羧酸循环 ppt课件

若从丙酮酸开始,加上纽带 生成的1个 NADH,则共产生10+2.5=12.5个ATP。 若从葡萄糖开始,共可产生 12.5×2+7=32个ATP。(二版及其他教材为38个ATP,
NADH3ATP,FADH2 2ATP)
可见由糖酵解和TCA循环相连构成的糖的 有氧氧化途径,是机体利用糖氧化获得能量 的最有效的方式,也是机体产生能量的主要 方式。
3、由琥珀酰CoA形成琥珀酸时,偶联有底物水平磷酸化生成1个GTP, 1GTP 1ATP。
4、循环中消耗两分子水。
5、3NADH
7.5 ATP ,
1FADH2
个GTP
6、单向进行
7、整个循环不需要氧,但离开氧无法进行。
1.5ATP,再加上1
1分子乙酰CoA通过TCA循环被氧化,可生成10分子
ATP。
乙酰CoA+
▪反C2应O+不N可A逆D,H+分H5+步进行,由丙酮酸脱氢酶复合体催化。
▪丙酮酸脱氢酶复合体是一个十分大的多酶复合体,包 括丙酮酸脱氢酶E1、二氢硫辛酸乙酰转移酶E2、二氢 硫辛酸脱氢酶E3三种不同的酶及焦磷酸硫胺素(TPP)、 硫辛酸,FAD, NAD+,CoA 及Mg2+六种辅助因子组 装而成。
• 你所经历的课堂,是讲座式还是讨论式? • 教师的教鞭
• “不怕太阳晒,也不怕那风雨狂,只怕先生骂我 笨,没有学问无颜见爹娘 ……”
• “太阳当空照,花儿对我笑,小鸟说早早早……”
一、由丙酮酸形成乙酰CoA
丙酮酸进入线粒体转变为乙酰CoA,这是连接糖酵解 和三羧酸循环的纽带:
丙酮酸+CoA+NAD+
❖ GTP+ADP
三羧酸循环——精选推荐

三羧酸循环第23章三羧酸循环(⽣物化学下册p92) 3学时学习重点:◆熟悉柠檬酸循环途径中的各步酶促反应,以及各步反应酶的作⽤特点。
◆会分析和计算酵解和柠檬酸循环中产⽣的能量,以及底物分⼦中标记碳的去向。
葡萄糖的有氧氧化包括四个阶段。
①糖酵解产⽣丙酮酸(2丙酮酸、2ATP、2NADH)②丙酮酸氧化脱羧⽣成⼄酰CoA③三羧酸循环(CO2、H2O、A TP、NADH)④呼吸链氧化磷酸化(NADH-----ATP)三羧酸循环:⼄酰CoA经⼀系列的氧化、脱羧,最终⽣成CO2、H2O、并释放能量的过程,⼜称柠檬酸循环、Krebs循环。
原核⽣物:①~④阶段在胞质中真核⽣物:①在胞质中,②~④在线粒体中⼀、丙酮酸脱羧⽣成⼄酰CoA1、反应式:2、丙酮酸脱氢酶系丙酮酸脱氢酶系是⼀个⼗分庞⼤的多酶体系,位于线粒体膜上,电镜下可见。
E.coli丙酮酸脱氢酶复合体:分⼦量:4.5×106,直径45nm,⽐核糖体稍⼤。
酶辅酶每个复合物亚基数丙酮酸脱羧酶(E1)TPP 24⼆氢硫⾟酸转⼄酰酶(E2)硫⾟酸24⼆氢硫⾟酸脱氢酶(E3)FAD、NAD+12此外,还需要CoA、Mg2+作为辅因⼦这些肽链以⾮共价键结合在⼀起,在碱性条件下,复合体可以解离成相应的亚单位,在中性时⼜可以重组为复合体。
所有丙酮酸氧化脱羧的中间物均紧密结合在复合体上,活性中间物可以从⼀个酶活性位置转到另⼀个酶活性位置,因此,多酶复合体有利于⾼效催化反应及调节酶在反应中的活性。
3、反应步骤反应过程(1)丙酮酸脱羧形成羟⼄基-TPP(2)⼆氢硫⾟酸⼄酰转移酶(E2)使羟⼄基氧化成⼄酰基(3)E2将⼄酰基转给CoA,⽣成⼄酰-CoA(4)E3氧化E2上的还原型⼆氢硫⾟酸(5)E3还原NAD+⽣成NADH4、丙酮酸脱氢酶系的活性调节从丙酮酸到⼄酰CoA是代谢途径的分⽀点,此反应体系受到严密的调节控制,此酶系受两种机制调节。
(1)可逆磷酸化的共价调节丙酮酸脱氢酶激酶(E A)(可被ATP激活)丙酮酸脱氢酶磷酸酶(E B)磷酸化的丙酮酸脱氢酶(⽆活性)去磷酸化的丙酮酸脱氢酶(有活性)(2)别构调节ATP、CoA、NADH是别构抑制剂ATP抑制E1CoA抑制E2NADH抑制E35、能量1分⼦丙酮酸⽣成1分⼦⼄酰CoA,产⽣1分⼦NADH(2.5A TP)。
三羧酸循环.PPT

和 FMN) 铁硫蛋白(Fe-S) 辅酶Q 细胞色素类
作用 递氢体
递氢体 单电子传递体 递氢体 单电子传递体
6
三、呼吸链中各组分的排列顺序
(一)依还原电位大小排列 (二)呼吸链复合物重组 (三)利用光谱变化观察线 粒体各传递体的氧化顺序
一、概念 二、呼吸链的组分 三、呼吸链中各组分
的排列顺序 四、线粒体中两条重
要呼吸链 五、胞液中NADH和
NADPH的氧化
一、概 念
催化生物氧化电 子传递的酶及其辅酶 的连锁反应体系称为 电子传递链。
由于电子传递链 与细胞摄取氧的呼吸 过程有关,通常又称 为氧化呼吸链。
5
二、呼吸链的组分
组分
尼克酰胺腺嘌呤二核苷 酸 (NAD+)
7
四、线粒体內的两条重要呼吸链 (一)NADH 氧化呼吸链 (二 )琥珀酸 氧化 呼吸链
8
9
五、胞液中NADH及NADPH的氧化 (一)-磷酸甘 油穿梭系统 (二)苹果酸穿 梭系统
10
11
(一)-磷酸甘油穿梭系统
胞液
线粒体 内膜
12
(二)苹果酸穿梭系统
胞液
内膜
线粒体
13
二、ATP的生成
(一)ATP生成的方式 (二)氧化磷酸化偶联部位 (三)氧化磷酸化偶联机理 (四)ATP合成的结构基础 (五)氧化磷酸化的调节 (六)氧化磷酸化抑制剂
14
(一)ATP生成的方式
1.底物水平磷酸化:底物的高能磷酸基团 直接转移给ADP生成ATP。
2.氧化磷酸化:是指在电子传递过程中,释放 的能量使ADP磷酸化成ATP的过程。又称 电子传递水平磷酸化
三羧酸循环(TCA)ppt课件

以NAD+为电子受体,存在于线粒体 中,需Mg2+。
以NADP+为电子受体,存在于胞 液中,需Mn2+。
完整版课件
13
6、α—酮戊二酸氧化脱羧生成琥珀酰CoA
第二次脱氢脱羧 不可逆 消耗1NAD+,生成1NADH+H+,1CO2
完整版课件
14
生成一个高能键“~”,此步 类似于丙酮酸的氧化脱羧。
47
1——3步
完整版课件
48
完整版课件
49
(二)可逆的非氧化阶段 (4——8) 戊糖互变 4、5—P 核酮糖(5—P—Ru)异构化为
5—P核糖(5—P—R) 官能团异构
完整版课件
50
5、5—P 核酮糖(5—P—Ru)异构化为 5—P木酮糖(5—P—Xu)
差向异构
完整版课件
51
4——5步
完整版课件
HO—CHCOOH
+NADP+
CH2COOH
再由苹果酸脱氢酶催化:
HO—CHCOOH
+NAD+ O=CCOOH
CH2COOH
完整版课件
+ NADH+H+ 36
CH2COOH
5、α—酮戊二酸和Asp 经转氨作用 生成Glu和草酰乙酸
完整版课件
37
第五节 磷酸戊糖途径(HMP PPP)
磷酸戊糖途径的概念:是G分解的另一条途径: 在6—P—G上直接氧化,再分解产生5—P—核糖。
CO2+H2O,说明另有途径。
(2)用同位素14C标记C1和C6 ,如果是EMP、TCA,那么生 成的14C1O2和14C6O2 分子数应相等,但实验表明14C1 更容易 氧化为CO2,说明另有途径。
大连理工大学生物化学课件--三羧酸循环(共58张PPT)

HSCoA
丙酮酸脱氢酶复合体的组成
NAD+
酶
辅酶
E1 :丙酮酸脱氢酶
TPP CoA-SH
E2 :二氢硫辛酰转乙酰基酶 硫辛酸
E3 :二氢硫辛酸脱氢酶
FAD NAD+
8
9
10
乙酰COA系列具有很高的酰基转移势能
11
功能是转送乙酰基或 其他酰基或氢
结合与蛋白质上的硫辛酸像 “摆动壁”一样把电子和酰基 从复合体中的一个酶转送到 另一个酶
丙酮酸+HCO3-+ATP
丙酮酸羧化酶 草酰乙酸+ADP+Pi
PEP+ CO2+GDP
PEP羧激酶
草酰乙酸+GTP
PEP+ HCO3-
PEP羧激酶 草酰乙酸+Pi
苹果酸+ HCO3- +NAD(P)H 苹果酸酶 苹果酸+NAD(P)+
41
厌氧细菌中不完整的TCA
TCA中间产物是某 些物质的合成原料 (前体)
ADP与NAD 浓度 :使三羧酸循环 被氧化。
5、植物中乙醛酸循环是柠檬酸循环的支路
+
乙酰辅酶A进入TCA生成10个ATP
ATP与NADH浓度 :使三羧酸循环 TCA中间产物是某些物质的合成原料(前体)
TCA中间产物是某些物质的合成原料(前体)
44
45
三羧酸循环的调节
糖有氧氧化的调节是基于能量的需求
有能量释放的过程。
C6H12O6 + 6O2
6CO2 + 6H2O + 能量
过程分三阶段,第一阶段在胞液(同酵解),后两个阶 段在线粒体中进行。
三羧酸循环PPT课件

催化丙酮酸生成乙酰CoA。
04
三羧酸循环的调控 机制
酶的别构效应
01
别构效应的概念
别构效应是指小分子化合物与酶活性中心外的某个部位非共价可逆结合
,引起酶的构象改变,从而影响酶的活性的调节方式。
02 03
别构效应的机制
别构效应的机制包括正协同效应和负协同效应。正协同效应是指小分子 化合物与酶结合后,增加酶的活性;负协同效应则是指小分子化合物与 酶结合后,降低酶的活性。
产生ATP
三羧酸循环是细胞内ATP的主 要来源,通过氧化磷酸化过 程,将底物的高能磷酸键转
化为ATP的高能磷酸键,为机 体提供大量的能量。
提供能量供给
三羧酸循环产生的ATP能够供 给细胞内各种生命活动的能 量需求,如维持离子梯度、 合成代谢等。
维持ATP水平
三羧酸循环通过维持ATP水平 来保持细胞内能量平衡,保 证细胞的正常功能。
THANKS
感谢您的观看
在三羧酸循环中,一系列的酶促反应将脂肪、蛋白质和糖等营养物质转化为能量。
三羧酸循环与柠檬酸循环的关系:三羧酸循环是柠檬酸循环的继续,它们之间没有 本质的区别。
循环的重要性
为生物体提供能量
三羧酸循环是细胞呼吸的主要途径,为生物体的生命活动提供能量 。
代谢途径的连接
三羧酸循环与其他代谢途径相互连接,如糖代谢、脂肪代谢和氨基 酸代谢等,确保了生物体内能量的有效利用。
调节代谢关键酶
三羧酸循环中的一些关键酶如柠檬酸合酶、异柠檬 酸脱氢酶等,能够调节其活性来控制代谢的速率和 方向。
参与信号转导
三羧酸循环中的一些产物如AMP、ADP等 可以作为信号分子参与细胞内的信号转导过 程,调节细胞的生理功能。
第十节:三羧酸循环 考研生物化学精编辅导讲义

第十节:三羧酸循环三羧酸循环(包括乙醛酸循环),又称柠檬酸循环,KREB循环。
是糖、脂肪、氨基酸等化合物生物氧化的共同通路,也是各代谢途径连接的枢纽。
生物氧化可分三个阶段:Stage1:乙酰CoA的生成。
Stage2:乙酰CoA的彻底氧化。
(TCA cycle)Stage3:电子传递和氧化磷酸化。
生物氧化特点:逐步缓和地释放能量,能量储存在ATP中。
丙酮酸氧化脱羧生成乙酰CoA 反应部位:线粒体(严密调节的不可逆步骤)该复合酶体的组成:丙酮酸脱氢酶E1,二氢硫辛酰转乙酰基酶E2,二氢硫辛酸脱氢酶E3;5种辅酶包括TPP(硫胺素焦磷酸),硫辛酰胺,CoA,FAD,NAD+三羧酸循环的反应部位:真核细胞的线粒体和原核细胞的胞浆2C的乙酰基以乙酰CoA的形式加入循环,首先和草酰乙酸反应,经过8步反应,释放2CO2和能量,最后再生一个草酰乙酸。
柠檬酸——顺乌头酸——异柠檬酸——×α酮戊二酸+CO2——×琥珀酰辅酶A+CO2——×琥珀酸——×延胡索酸——L苹果酸——×草酰乙酸通过氧化磷酸化,1分子NADH产生2.5ATP,1分子FADH2产生1.5ATP。
1分子葡萄糖彻底氧化产生30~32ATP。
P96-97关于TCA循环中能量的产生。
➢生理意义:TCA补充,才能维持TCA的正常进行。
一些厌氧细菌缺乏α-酮戊二酸脱氢酶,它们通过不完整的TCA获得生物合成的前体。
这种不完整的TCA可能是TCA早期进化的一个阶段➢①②共价修饰丙酮酸脱氢酶复合体中还有两个调节蛋白,一个是磷酸激酶,另一个是磷酸酶。
磷酸①三羧酸循环中三步不可逆的放能反应是其调节位点。
②催化这三步反应的酶受到产物反馈抑制(如柠檬酸、琥珀酰CoA等),也受到高能荷物质抑制(如ATP、NADH等)。
③催化这三步反应的酶受到低能荷物质(如ADP等)或能量需求信号(如Ca2+)激活。
④由于三羧酸循环的中间物可作为前体用于生物合成,因此底物的浓度对于调节也是比较重要的。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
是糖、氨基酸和脂肪酸最后共同的代谢途径
也称为柠檬酸循环和Krebs循环
糖酵解产生的丙酮酸(实际上是乙酸)被降解成CO2 产生一些ATP 产生更多的NADH NADH进入呼吸链,通过氧化磷酸化产生更多的ATP。
完整的三羧酸循环
乙酰CoA的形成
脂肪酸的β氧化 氨基酸的氧化分解 丙酮酸的氧化脱羧——由丙酮酸脱氢酶系催化
转乙酰酶
哺乳动物60
CoA
泛酸 辅酶 移到CoA
E3 二氢硫辛酸 大肠杆菌12、酵母12、 FAD
B2
辅基 氧化型硫辛
脱氢酶
哺乳动物6
NAD+
PP
辅酶 胺的再生
大肠杆菌内丙酮酸脱氢酶系的电镜照片
丙酮酸转变成乙酰-CoA的四步反应
丙酮酸脱氢酶的催化机理
砒霜的毒性机理
反应1:柠檬酸合酶
柠檬酸的合成
30或31或32
三羧酸循环中间物的去向
草酰乙酸的回补反应
乙醛酸循环
植物和微生物的三羧酸循环的变化形式
☻琥珀酸的类似物丙二酸是该酶的竞争性抑制剂
反应7:富马酸酶
双键的水合
☻水分子加成反式的双键 ☻反应机制还不清楚
反应8:苹果酸脱氢酶
依赖于NAD+-的氧化还原反应
☻这是三羧酸循环的最后一步反应,也是三羧酸循 环中的第四次氧化还原反应 ☻ΔGo‘ = +30 kJ/mol,意味着在热力学上极不利于正 反应的进行,但是,在体内反应产物草酰乙酸可 以迅速被下一步不可逆反应消耗,NADH则进入呼 吸链被彻底氧化,因此,整个反应被“强行拉向” 正反应。
3NADH,1FADH2
2H2O被使用作为底物 绝对需要O2
吵, 您顺意吵,(吵得)铜壶呼盐瓶!
TCA循环的功能
产生更多的ATP
提供生物合成的原料
是糖、氨基酸和脂肪酸最后的共同分解途径 某些代谢中间我作为其他代谢途径的别构效应物 产生CO2
一分子葡萄糖彻底氧化过程中的ATP 收支情况
与ATP合成相关的反应Fra bibliotekCA循环中C的命运
☻乙酰CoA的羰基C只有在第2轮循环转变成CO2 ☻乙酰CoA的甲基C能完全留在两轮循环中,但是以后每一轮循
环有一半离开。
TCA 循环总结
总反应: 乙酰-CoA+3NAD+
+FAD+GDP+Pi+2H2O→2+C+OC2o+A3NADH+FADH2+GTP+2H
1个乙酰-CoA通过三羧酸循环产生2CO2, 1 ATP,
反应3:异柠檬酸脱氢酶(IDH)
异柠檬酸氧化脱羧产生α-酮戊二酸
☻先是脱氢,然后是β-脱羧 ☻有两种形式的异柠檬酸脱氢酶,分别使用辅酶I和辅酶II作为
氢的受体
反应4: α-酮戊二酸脱氢酶系
第二次氧化脱羧反应
☻酶几乎等同于丙酮酸脱氢酶系——结构上或者机制上 ☻5种辅酶——TPP、CoASH、硫辛酸 NAD+、FAD ☻也是亚砷酸的作用对象
合成ATP的方式
合成ATP的量
糖酵解(包括氧化磷酸化) 己糖激酶 PFK-1 磷酸甘油酸激酶 丙酮酸激酶 甘油醛-3-磷酸脱氢酶 (NADH)
丙酮酸脱氢酶系
三羧酸循环 异柠檬酸脱氢酶(NADH) α-酮戊二酸脱氢酶系(NADH) 琥珀酰-CoA合成酶 琥珀酸脱氢酶(FADH2) 苹果酸脱氢酶(NADH)
☻柠檬酸合酶由两个相同的亚基组成,它被视为酶“诱导契 合”学说又一代表性的例子
☻在没有底物结合的情况下,酶的两个亚基的构象是开放型 的;当结合底物以后,则被诱导为紧密型。反应动力学为 序列有序型,在反应中,OAA首先与酶活性中心结合,这 种结合迅速诱导活性中心的构象发生变化,从而创造出乙 酰-CoA的结合位点。随后,乙酰-CoA结合到酶的活性中心, 并与OAA形成中间产物—柠檬酰-CoA,这时,酶的构象再 次发生变化,远离活性中心的一个关键的Asp残基被拉入到 柠檬酰-CoA上的硫酯键,很快硫酯键被切开,终产物辅酶A 和柠檬酸被依次释放。
反应5:琥珀酰-CoA合成酶
TCA循环唯一的一步底物水平磷酸化反应
☻ATP或GTP被合成 ☻它的催化过程牵涉到一系列高能分子的形成,因此能量的损失
微乎其微 ☻反应机制涉及一个磷酸组氨酸
琥珀酰-CoA合成酶的反应机理
反应6:琥珀酸脱氢酶
产生FADH2
☻此酶实际上是呼吸链复合体II的主要 成分
柠檬酸异构化成异柠檬酸
☻柠檬酸不是氧化的好底物
☻S异柠檬酸却不一样,经过异构化,三级羟基变成了 易氧化的二级羟基
☻在形成的异柠檬酸分子中,羟基只会与来源于草酰乙 酸的β-碳原子而绝对不会与来源于乙酰-CoA的β-碳 原子相连!
☻顺乌头酸酶使用铁硫蛋白
铁硫蛋白在顺乌头酸酶反应中的作用
顺乌头酸酶催化反应中产物的立体专一性
丙酮酸跨线粒体内膜的转运
丙酮酸脱氢酶系的结构和组成
缩写 酶活性
亚基数目 (个数)
辅助因子 维生素 辅助因 催化的反应 前体 子类型
E1 丙酮酸脱氢 大肠杆菌24、酵母60、 TPP
酶
哺乳动物20或30
B1
辅基
丙酮酸
氧化脱羧
E2 二氢硫辛酸 大肠杆菌24、酵母60、 硫辛酰胺 硫辛酸 辅基 将乙酰基转
丙酮酸的氧化脱羧
丙酮酸如何进入线粒体? 丙酮酸脱氢酶系的结构与组成——丙酮酸脱氢
酶系由丙酮酸脱氢酶(E1)、二氢硫辛酸转乙酰 酶(E2)和二氢硫辛酸脱氢酶(E3)通过非共价 键结合在一起的稳定复合物 催化机制 亚砷酸和有机砷的作用对象——氧化型硫辛酰 胺的再生对于丙酮酸脱氢酶系的持续运转十分重 要,砒霜的主要成分亚砷酸能够与还原型的硫辛 酰胺形成共价的复合物而阻止它的再生 。
☻柠檬酸合酶在催化过程中所发生的由底物OAA和中间产物 柠檬酰-CoA分别诱导的两次构象变化既防止了乙酰-CoA的 提前释放,也大大降低了乙酰-CoA在活性中心被Asp残基水 解成乙酸的可能性。
柠檬酸合酶催化的反应
氟代乙酸在细胞内的代谢转变及其对TCA循环的影响
柠檬酸合酶的两种构象
反应2:顺乌头酸酶
总ATP量
消耗ATP 消耗ATP 底物水平磷酸化 底物水平磷酸化 氧化磷酸化
氧化磷氧化磷酸化酸化 氧化磷酸化 氧化磷酸化 底物水平磷酸化 氧化磷酸化 氧化磷酸化
5或6或7 -1 -1 +2 +2 +3或+4或+5(取决于 NADH通过何种途径进入 呼吸链)
2×2.5=5
19 2.5×2=5 2.5×2=5 1×2=2 1.5×2=3 2.5×2=5