第十三章 数量性状的遗传
遗传学数量性状的遗传分析

遗传学数量性状的遗传分析
目录
• 引言 • 数量性状遗传基础 • 数量性状遗传分析方法 • 数量性状基因定位 • 数量性状基因组关联分析 • 数量性状基因组编辑与优化
01
引言
研究背景
01
遗传学数量性状是生物体表型特 征中受多个基因和环境因素共同 影响的性状,如身高、体重等。
02
随着分子生物学和基因组学的发 展,遗传学数量性状的遗传分析 已成为遗传学研究的重要领域。
关联分析的软件工具
01
Plink
一款常用的关联分析软件,提供 多种统计分析和可视化工具,用 于处理和分析大规模遗传数据。
02
03
GAPIT
Tassel
基于R语言的关联分析工具包, 提供了丰富的统计方法和可视化 功能,适用于复杂数据分析。
主要用于基因组关联分析的软件, 支持多种数据格式和多种统计模 型,可进行大规模数据分析。
QTL定位的软件工具
QTL Cartographer
基于区间作图法的QTL定位软件,适用于大样本数据 集。
Tassel
综合关联分析和区间作图法的QTL定位软件,具有强 大的数据处理和分析能力。
R/qtl
基于R语言的QTL定位软件,提供了多种统计模型和 可视化工具。
05
数量性状基因组关联分析
关联分析的基本原理
广义遗传力
广义遗传力用于描述数量性状在遗传和环境变异中的贡献,计算公式为加性方差和显性方差占表型方差的比值。
狭义遗传力
狭义遗传力仅考虑基因型对表型变异的贡献,计算公式为加性方差占表型方差的比值。
遗传相关分析
遗传相关系数
用于描述两个数量性状之间的遗传关系,计算公式为两个数量性状的加性方差和显性方差之间的比值 。
遗传学 课后练习 复习题 总结 第十三章 数量性状的遗传

第十三章数量性状的遗传本章习题1.解释下列名词:广义遗传率、狭义遗传率、近交系数、共祖系数、数量性状基因位点、主效基因、微效基因、修饰基因、表现型值、基因型与环境互作广义遗传率:通常定义为总的遗传方差占表现型方差的比率。
狭义遗传率:通常定义为加性遗传方差占表现型方差的比率。
近交系数:是指个体的某个基因位点上两个等位基因来源于共同祖先某个基因的概率。
共祖系数:个体的近交系数等于双亲的共祖系数。
数量性状基因位点:即QTL,指控制数量性状表现的数量基因在连锁群中的位置。
主效基因:对某一性状的表现起主要作用、效应较大的基因。
微效基因:指一性状受制于多个基因,每个基因对表现型的影响较小、效应累加、无显隐性关系、对环境敏感,这些基因称为微效基因。
修饰基因:对性状的表现的效应微小,主要是起增强或减弱主基因对表现型的作用。
表现型值:是指基因型值与非遗传随机误差的总和即性状测定值。
基因型与环境互作:数量基因对环境比较敏感,其表达容易受到环境条件的影响。
因此,基因型与环境互作是基因型在不同环境条件下表现出的不同反应和对遗传主效应的离差。
2.质量性状和数量性状的区别在哪里?这两类性状的分析方法有何异同?答:质量性状和数量性状的区别主要有:①. 质量性状的变异是呈间断性,杂交后代可明确分组;数量性状的变异则呈连续性,杂交后的分离世代不能明确分组。
②. 质量性状不易受环境条件的影响;数量性状一般容易受环境条件的影响而发生变异,而这种变异一般是不能遗传的。
③. 质量性状在不同环境条件下的表现较为稳定;而控制数量性状的基因则在特定时空条件下表达,不同环境条件下基因表达的程度可能不同,因此数量性状普遍存在着基因型与环境互作。
对于质量性状一般采用系谱和概率分析的方法,并进行卡方检验;而数量性状的研究则需要遗传学方法和生物统计方法的结合,一般要采用适当的遗传交配设计、合理的环境设计、适当的度量手段和有效的统计分析方法,估算出遗传群体的均值、方差、协方差和相关系数等遗传参数等加以研究。
数量性状的遗传

第四章数量性状的遗传目的要求掌握数量性状与质量性状的区分、特征,多基因假说的要点,数量性状表现值的分解,遗传力的概念;了解通径系数概念与意义,基因的非加性效应与加性效应的意义,遗传力公式的推导及计算方法;掌握遗传力的应用。
第一节数量性状的遗传基础生物的性状基本上可分为两大类:质量性状(qualitative trait):变异可以截然区分为几种明显不同的类型,一般用语言来描述;数量性状(quantitative trait):个体间性状表现的差异只能用数量来区别,变异是连续的。
阈性状(threshold trait):表现型呈非连续变异,与质量性状类似,但不是由单基因决定,性状具有一个潜在的连续型变量分布,遗传基础是多基因控制的,与数量性状类似。
一、数量性状的一般特征数量性状的特点:①数量性状是可以度量的;②数量性状呈连续性变异;③数量性状的表现容易受到环境的影响;④控制数量性状的遗传基础是多基因系统。
学习数量性状的方法①统计学思想贯穿数量性状遗传的全部内容;②确定性与不确定性的矛盾时时体现;③研究对象在个体与群体间的相互转换;④遗传与变异的矛盾。
二、数量性状的遗传基础1.多基因假说瑞典遗传学家尼尔迩·埃尔(Nilsson-Ehle)通过对小麦籽粒颜色的遗传研究,提出了数量性状遗传的多基因假说。
多基因假说的要点(1)数量性状是由许多微效基因决定的,每个基因的作用的微效的;(2)基因的作用是相等的,且可以累加、呈现剂量效应,等位基因间通常无显隐关系;(3)基因在世代相传中服从孟德尔定律,即分离规律和自由组合规律,以及连锁交换规律2.基因的非加性效应基因的非加性效应包括显性效应和上位效应。
(1)显性效应由等位基因间相互作用产生的效应。
例1:有两对基因,A1、A2的效应各为20cm,a1、a2的效应名为10cm,基因型A1A1a2a2按加性效应计算其总效应为60cm 。
而在杂合状态下,即A 1a 1A 2a 2同样为两个A 和两个a ,其总效应可能是75cm(2)上位效应或互作效应 由非等位基因之间相互作用产生的效应。
遗传学_ 数量性状遗传_
个体的基因型
✓ 个体性状的表现型数值,称为表现型值,以P表示。 ✓ 表现型值有两部分组成:
一个是基因型所决定的数值,称为基因型值,以G表示; 一个是环境条件引起的变异,用E表示。 ✓ 表现型值、基因型值,和环境变异值三者之间的数量关 系可用以下公式表示:P=G+E
环境条件的影响
✓ 表型变异用表型方差(即总方差)VP表示; ✓ 遗传变异用遗传方差(即基因型方差)VG表示; ✓ 环境变异用环境方差VE表示。 ✓ 三者的数量关系可用下式表示:Vp=VG +VE
三、纯系学说
(三)纯系学说的发展
“ 纯系的纯是相对的、暂时的,绝对的纯系
是不存在的,纯系内继续选择可能是有效的。 纯系繁育过程中,由于突变、天然杂交和机械 混杂等因素必然会导致纯系不纯,产生新的遗 传变异,可能出现更优个体。
“
遗传率及估算方法
一、数量性状变异的表示方法
生物性状 表现的 决定因素
超矮秆表型是由于D18的突变导致。 该种突变体除株高显著降低后,其他 农艺性状与野生型无显著性差异。
小麦粒色简单划分,表现质量性状,单细致 观察,籽粒颜色红到白,表现连续变异,数量性 状的特点。
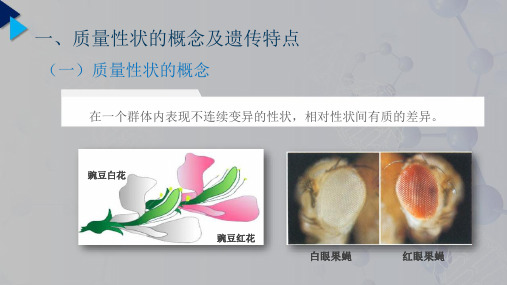
二 、数量性状的概念及遗传特点
(三)数量性状和质量性状的相对性
生物还有一些性状为阈性性状: 表型呈非连续变异,而其基本物质 的数量呈潜在的连续变异的性状, 即只有超越某一遗传阈值时才出现 的性状,如动植物甚至包括人类的 抗病力、死亡率以及单胎动物的产 仔数等性状。
3 数量性状对环境条件的变化反应敏感。
4 研究方法上,依靠群体,必须用统计方法,对在杂种和后代进行分析。
二 、数量性状的概念及遗传特点
(三)数量性状和质量性状的相对性
数量性状的遗传名词解释

数量性状的遗传名词解释数量性状,是指在自然界或人工条件下产生的各种特征以数量的方式表现出来的遗传性状。
它指的是通过对种群中大量个体进行测量或计量,将结果以数量化的形式呈现出来的遗传特征。
数量性状通常具有连续变异的特征,即在一个种群中存在着一系列不同的表现形式,而不是像离散性状那样只有几个确定的表型。
在数量性状的研究中,有一些重要的遗传名词需要加以解释。
其中包括基因型、表型、遗传方差、环境方差、遗传相关等。
基因型是指个体在基因水平上的遗传组成。
它决定了个体对特定数量性状的表现。
每个数量性状通常由多个基因共同决定,因此基因型的组合将决定这些基因在个体上的表现形式。
表型是指个体在外部表现上的特征。
它受到基因型和环境的共同影响。
数量性状的表型通常呈现连续性变化,这是因为数量性状通常受到多种基因的共同作用,以及环境因素的影响。
例如,人体身高就是一种典型的数量性状,它受到多基因的影响,同时还受到营养、运动等环境因素的调节。
遗传方差是指数量性状中由基因所引起的表型变异程度。
它可以通过研究个体间的表型差异以及表型与基因型之间的关系来估计。
遗传方差的大小反映了数量性状中遗传因素的重要程度。
如果遗传方差较大,说明遗传因素在数量性状的表达中起到了重要作用,反之则说明环境因素的贡献较大。
环境方差是指数量性状中由环境因素所引起的表型变异程度。
环境方差通常通过比较同一种群中不同个体之间的差异来估计。
环境方差的大小表示了环境对数量性状的影响程度。
如果环境方差较大,说明环境因素在数量性状的表达中起到了重要作用,反之则说明遗传因素的贡献较大。
遗传相关是指在同一种群中不同数量性状之间的遗传联系。
它反映了一种或多种数量性状随着基因型的变化而变化的程度。
通过研究数量性状之间的遗传相关,可以了解不同数量性状之间的遗传关系及其对进化和适应的影响。
例如,身高和体重之间的遗传相关可以帮助我们理解这两个数量性状在人类进化中是如何相互影响的。
以上介绍了数量性状的遗传名词解释,包括基因型、表型、遗传方差、环境方差和遗传相关等概念。
数量性状的遗传分析

表10-2 玉米穗长度的遗传
图10-2 玉米穗长度的遗传
短
长
4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
F1
穗长
8 9 10 11 12 13 14 15 16
穗长
F2
6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
阈性状
有一类特殊的生物性状,不完全等同于数量性状或质量性状,其 表现呈非连续变异,与质量性状类似,但是又不服从孟德尔遗传 规律。一般认为这类性状具有一个潜在的连续型变量分布,其遗 传基础是多基因控制的,与数量性状类似。即由微效多基因控制 的,呈现不连续变异的性状。通常称这类性状为阈性状 (threshold character)。
单击此处添加副标题
10 数量性状的遗传分析
Genitics of Quantitative Character
单林娜 制作
1
上次课中所讲的性状差异,大多是明显的不连续差异。例如
豌豆种子的圆与皱,子叶的黄与绿
水稻的粳与糯
鸡羽的芦花斑纹和非芦花斑纹
这类性状在表面上都显示质的差别,所以叫做质量性状 (qualitative character)。质量性状的遗传可以比较容易地由分 离定律和连锁定律来分析。
10.3.1 广义遗传力(heritability in the broad sense) 的估算方法
因为方差可用来测量变异的程度,所以各种变异都可 用方差来表示,这样,
P = G + E 就可表示为:
VP = VG + VE 遗传方差;VE:环境方差)
(VP:表型方差;VG:
我们把遗传方差占总方差的比值称为广义遗传力
数量性状遗传教案

数量性状遗传教案数量性状是指一种性状可以通过计数或测量来度量的性状。
这些性状在遗传中的传递方式可以通过数量性状遗传教案来进行教学和学习。
教案:一、教学目标:1.理解数量性状的概念和特点。
2.掌握数量性状的遗传规律和方法。
3.能够应用数量性状遗传的知识来解决问题和进行实验。
二、教学内容:1.数量性状的定义和特点。
2.数量性状的遗传规律。
3.数量性状的测量方法。
4.应用数量性状遗传的实例分析。
三、教学过程:1.导入(5分钟):通过展示一组根据身高排序的学生图片,引发学生对数量性状的认知和兴趣。
2.知识讲解(15分钟):a.介绍数量性状的定义和特点:数量性状是指可以通过计数或测量来度量的性状,如身高、体重、产量等。
这些性状受多个基因和环境因素的影响。
b.数量性状的遗传规律:数量性状由多对基因控制,遵循连续变异规律。
常见的遗传模式包括寡基因控制和多基因控制。
c.数量性状的测量方法:根据具体性状的特点选择相应的测量方法,如身高可以通过直接测量或问卷调查等方式获得。
3.案例分析(30分钟):通过实例分析来加深学生对数量性状遗传的理解。
a.案例一:一个家族中的兄弟姐妹身高差异很大,其中一个兄弟非常高大而且父母也身高较高。
请分析此现象的遗传原因,并给出可能的遗传模式。
b.案例二:在一个小麦品种改良项目中,科研人员通过测量不同品种小麦的产量来筛选高产的品种。
他们发现产量呈正态分布,即大部分品种产量中等,极少数有很高或很低的产量。
请问这个产量数量性状的遗传基础是什么?4.实验设计(30分钟):让学生设计一个实验来验证数量性状的遗传规律。
a.主题:探究小麦高产性状的遗传方式。
b.实验材料:不同产量的小麦品种种子。
c.实验步骤:-步骤一:选择5个不同产量的小麦品种,分别种植在相同的土壤和环境条件下。
-步骤二:测量和记录每个品种的产量。
-步骤三:通过基因分析或后代观察,确定每个品种高产性状的遗传模式。
-步骤四:总结实验结果,并得出结论。
数量性状的遗传—数量性状遗传的特征(遗传学课件)

所以数量性状在农业中显得特别重要。 (三)人类
人的身高、体重、胖瘦、寿命……
三、认识数量性状
特点:变异不容易分为截然不同的组别,其间有 一系列的过渡类型,只有数量的不同,没有质的 差别。
10
0
5
6
7
8
9 10 11 12 13 14 15 16 17 18 19 20 21
《遗传学》
知识目标
学习目标
一、 二、 三、
知道 清楚 数量 数量
熟悉 数量 性状
性状 性状 与质
的概 的遗 量性
念 传特 状的
点
区别
能力目标
能用分析 数量性状 的方法分 析育种与 生产中的 实际问题
Gregor Mendel 1822-1884
(一)数量性状与质量性状的区别
五、数量性状与质量性状的关系 (二)数量性状与质量性状的相对性 1、数量性状与质量性状的区别不是绝对的; 2、生物的性状都有其质和量两个方面,只是在一 定条件下质和量表现出主次关系。 3、在不易区分一个性状是质量性状或数量性状时, 就必须根据F1或F2遗传动态特征来作出判断。
30
亲 本 25
20
玉米穗长遗传的柱形图
15
10
5
0
5
6
7
8
9 10 11 12 13 14 15 16 17 18 19 20 21
18
16
F 14 1 12
10
8 6 4
2 0
5
6
7
8
9 10 11 12 13 14 15 16 17 18 19 20 21
遗传学-数量性状的遗传分析

三、微效基因表型值的推算
累加作用(每个显性基因的作用以一定的数值与纯隐性亲本 的表型值相加) 纯显性亲本表型值=每个显性基因表型值X纯显性亲本基因数+ 纯隐性亲本表型值 如短穗玉米x=6.6,长穗玉米x=16.8,F2中长、短穗各占群体 的1/16 4n=16,n=2 控制长穗玉米穗长的显性基因为2对(4个). 每个显性基因表型值=纯显亲本表型值-纯隐亲本表型值/纯显 亲本基因数=16.8-6.6/4=2.55 所以,含一个显性基因的玉米穗长:6.6+2.55=9.15cm 含2个显性基因的玉米穗长:6.6+(2×2.55)=11.7cm 依此类推。
狭义遗传率
计算基因的相加效应的方差VA在总的表型方差中所占的百分率。
Aa同AA回交的子代个体为B1,同aa回交的子代个体为B2。 B1的遗传方差的计算 f x fx fx2 AA 1/2 a 1/2a 1/2a2 Aa 1/2 d 1/2d 1/2d2 合计 1 1/2(a+d) 1/2(a2+d2) B1的遗传方差:VB1=1/2(a2+d2) -1/4(a+d)2=1/4(a-d)2 B2的遗传方差的计算 f x fx fx2 Aa 1/2 d 1/2d 1/2d2 aa 1/2 -a -1/2a 1/2a2 合计 1 1/2(d-a) 1/2(a2+d2) B2的遗传方差:VB2=1/2(a2+d2)- 1/4(d-a)2=1/4(a+d)2
例如小麦籽粒颜色两对基因控制的遗传动态 P 红R1R1R2R2 白r1r1r2r2 R1r1R2r2 红 1 4 6 4
F1
F2
1
4R
深红
3R
中深红
13 数量性状的遗传(6h)PPT共63页

F2基因型 及其比率:
1a1a1a2a2 增效基因数:0 表型: 白粒 表型比: 1/16
A1A1A2A2 × a1a1a2a2
红粒
白粒
A1a1A2a2(粉红粒)
2a1a1A2a2 2A1a1a2a2
1a1a1A2A2 4A1a1A2a2 1A1A1a2a2
2A1A1A2a2 2A1a1A2A2
1
2
3
淡红粒 中红粒 红粒
4/16
6/16
4/16
1A1A1A2A2
4 深红粒 1/16
13.2.3 数量性状基因座 (QTL)
❖ 概念:
控制数量性状的基因是微效多基因,这些基因往 往相对集中存在于一个染色体或多个染色体的某一 区段——数量性状基因座(quantitative trait locus,QTL)。
↓
粉红粒 ↓
粉红粒 ↓
F2 3/4红粒 1/4白粒
15/16红粒 1/16白粒
63/64红粒 1/64白粒
❖ 试验结果分析:
分析结果表明,小麦和燕麦中存在3对与种皮颜色有关、种类不 同但作用相同的基因,这3对基因中的任何一对在单独分 离时都可以产生3∶1的比率,当3对基因同时分离时,则 产生63/64∶1/64的比率;上述杂交在F2的红粒中又呈现各 种程度的差异,按红色的程度分为:
➢微效基因之间往往缺乏显性,用大写字母表示增 效,用小写字母表示减效。
13.2.1 多基因假说(polygene hypothesis)
➢微效基因对环境敏感,因而数量性状的表现易受环 境因素的影响而发生变化。微效基因的作用常常被 整个基因型和环境的影响所遮盖,难以识别个别基 因的作用。
➢多基因往往有多效性。多基因一方面对某一性状起 微效基因的作用,同时对其它性状起修饰基因的作 用,使之成为其它基因表现的遗传背景。
数量性状和遗传

数量性状和遗传
根据一般实验经验,例如说水稻穗选,穗重而谷粒多的,其后代平均讲来,穗的重量也要大些。
这是我们在选种中最基本最普通的常识。
可是有时穗选的效果很显著,产量也有很大的提高;有时穗选的效果不显著,产量未能有所提高,这是什么道理呢?在本世纪初,约翰逊把这个问题作了深入分析,得到了发人深思的结果。
他用市场上买来的菜豆做实验,有些菜豆有轻有重,参差不齐,轻的只有15厘克,中的可达90厘克。
他从轻重不一的19粒菜豆出发,建立了19个纯系。
因为菜豆是高度自花授粉植物。
每粒种子的后代应该都是纯合的,所以约翰逊称单粒种子的自交后代为纯系。
不同纯系间的平均粒重可有明显差异,而在一个纯系内,豆粒也有轻有重,且呈连续分布,但其平均粒重与亲代几乎没有差异。
所以约翰逊认为,一个纯系内的粒重变异是不遗传的,而不同纯系间的变异至少一部分是遗传的。
下图就是他的一个纯系数据。
在连续六年内,选出纯系内最大的种子和最小的种子分别种下,后代种子的平均重量始终都一样,没有什么区别。
数量性状的遗传

数量性状的遗传数量性状指的是一个生物体的某种性状具有连续性质,在一个种群中表现出一定的变异程度,且受多种基因和环境因素的影响。
例如人体身高、体重等就是数量性状。
数量性状由多个基因的作用所决定,被称为多基因性状。
与单基因性状不同的是,多基因性状不符合孟德尔遗传定律。
数量性状的遗传规律经过长时间的探究,现已初步得出。
从基因层面探究数量性状的遗传数量性状的基因型及其表现形式比较复杂,同一基因型的个体之间也会存在表现形式的差异。
基因由两条相同或不同的基因座构成,分别来自父母亲。
在数量性状的遗传中,每个基因座所对应的基因影响数量性状的大小和表现型。
同时,多个基因座共同作用于数量性状,这种作用关系被称为加性效应(additive effect)。
数量性状的遗传规律主要有:性状值=基因值+环境值,基因型对数量性状的影响呈现正态分布,且受到染色体上多个基因的影响。
数量性状的遗传模式数量性状的遗传规律有三种模式:常染色体显性遗传、常染色体隐形遗传以及性联遗传。
常染色体显性遗传的表现形式是当一个自由基因突变,双等位基因后者扰动的时候,显性基因造成的表现现象。
例如,人体的眼睛颜色就是常染色体显性遗传的一种表现。
常染色体隐性遗传与常染色体显性遗传类似,不同的是表现基因是一种隐性基因。
这种遗传模式表现突变基因表现在两条染色体上都具有相同的表现现象。
例如,某些人患有系统性红狼疮就是常染色体隐性遗传的一种表现。
性联遗传指由X和Y染色体来遗传。
X染色体上的基因对于女性来说是双等位基因,由于女性有两个X染色体,所以会出现多种表现型。
而男性由于只有一个X 染色体,所以表型变化更加显著和恒定。
例如,红绿色盲就是一种典型的性连锁遗传疾病。
数量性状的计算分析数量性状的遗传变异分析可以通过基因型频度分析、亲权分析和遗传连锁分析来进行。
(1)基因型频度分析:由于每个基因座共有两个等位基因,因此可将一个种群中某一基因座的等位基因频率进行 PA+Pa=1,其中PA为某一基因座等位基因A 的频率,Pa为某一基因座等位基因a的频率。
数量性状的遗传

47 42.8
轻粒种子
35.8 40.2 31.4 38.3 37.9 37.4 36.8
重粒种子
34.8 41.0 32.6 39.2 39.9 37.0 37.4
轻粒种子
60 55 50 43 46 46 51.7
重粒种子 轻粒种子
70
63.2
80
75.2
87
54.6
73
63.6
84
74.4
81 79.2
2、双亲之间相差的基因对数不同,某些性状有时表 现质量性状,有时表现为数量性状
3、因观察的层次不同,性状的遗传方式不同 宏观层次上不连续分布,则表现质量性状,但决
定性状的基本物质是连续分布的,则表现为数量性状。
11
五、阈性状
概念 阈性状(threshold traits) :数量性状在表现型上
数量性状
连续性
连续性(中亲值或有偏向) 连续性(正态分布) 易受环境条件影响 产生变异 微效多基因控制 作用相等,累加效应 群体大, 世代数多 采用统计方法
10
1、区分性状的方法不同,有些性状既有数量性状的 特点又有质量性状的特点 例如:小麦粒色 两对基因 F2 15:1 为质量性状 F2 1 4 6 4 1 为数量性状
某一性状在群体中的表现型可以分为许多变异 型,各变异型之间的区别不明显,总体上呈 连续变异(continuous variation)。
如人的身高、肤色的深浅、体重;棉花的纤 维长度、奶牛的泌乳量等等。
又可分为计数和量测两种情况。
2
例. 1913年,R.Emerson 和E.East公布了他们关于玉米 果穗长度的遗传实验。用玉米长果穗和短果穗两个品系进行杂 交,结果见表及图。
数量性状的遗传(下)

• 状态相似 :
• 群体中可能还有另外一类具有BB基因型的个体 ,其中的两个等位基因B不是血统相同的,而 仅是状态相似的(alike in state)。 • 如果所有纯合基因型个体都不具有血统相同的 等位基因,这些等位基因都可以认为是状态相 似的
近交程度的度量
近交系数(F)—— 动物
指某一个体(二倍体生物)任何基因座位上两 个基因来自共同祖先的同一基因的概率。 0≤F≤1 F=0, 表明某一个体的两个亲本没有亲缘关系 F=1, 表明某一个体的两个亲本是同基因型的纯合体
F1
A1B1C1 D1E1 A2B2C2 D2E2
2+2+2+2+2=10
根据这一假说,等位基因间没有显隐性关系。 杂合等位基因间的相互作用显然大于纯合等位 基因间的相互作用
(三)、两种假说比较
• 两者的相似之处: – 都立论于杂种优势来源于双亲基因间的相互关系,也就是说双亲 间基因型的差异对杂种优势起着决定性作用。 – 都没有考虑到非等位基因间的相互作用(上位性作用)。
ACEIHB和ACEIKHB。
因此,Z的近交系数为:
7 6 7
C A Z
D B
1 1 1 1 1 1 1 2 2 2 128 64 128 32
若共同祖先F≠0 则
F=∑(1+F i)
i
1 2
ni
F i =(1/2)3=0.125
一、杂种优势的表现:定义与表示方法
• 杂种优势(heterosis/hybrid vigor)是生物界的普遍现
象,指两个遗传组成不同的亲本的杂种第一代,在生 长势、繁殖力、抗逆性、产量和品质上比其双亲优越 的现象。 • 杂种优势所涉及的性状大都为数量性状,故以性状数
《数量性状遗传》课件

05
总结与展望
数量性状遗传研究的重要意义
揭示生物多样性
数量性状遗传研究有助于揭示生物多样性的遗传基础,理解生物进 化的机制。
指导育种工作
通过数量性状遗传研究,可以更有效地进行动植物育种,提高农作 物的产量和品质,以及改善动物的生长性能和健康状况。
遗传方差与环境方差的比较
01
02
03
遗传方差
表示数量性状受基因控制 的变异程度,包括加性方 差和显性方差。
环境方差
表示数量性状受环境因素 影响的变异程度。
比较意义
了解遗传方差和环境方差 的相对大小,有助于理解 数量性状的变异来源和选 择潜力。
遗传进度与选择效率的关系
遗传进度
指选择过程中一个或多个世代的 遗传改变量。
应用
QTL定位在动植物育种、人类医学等领域具有广泛的应用价值,有 助于深入了解数量性状的遗传基础和进行相关研究。
03
数量性状遗传的应用
作物育种中的应用
提高产量和品质
通过研究数量性状遗传,育种家可以培育出产量更高、品质更优的作物品种。例如,通过 选择具有理想株高、穗粒数等数量性状的个体,可以获得抗逆性强、适应性广的作物品种 。
《数量性状遗传》ppt课件
Hale Waihona Puke 录• 数量性状遗传概述 • 数量性状遗传的遗传机制 • 数量性状遗传的应用 • 数量性状遗传的未来发展 • 总结与展望
01
数量性状遗传概述
数量性状遗传的定义
数量性状遗传是指多个基因位点 共同作用,对个体表现型产生影
响的遗传现象。
它与质量性状遗传不同,质量性 状遗传是由单一或少数基因位点 控制,表现为明显的孟德尔遗传
《数量性状遗传》课件

遗传模型构建方法
遗传力模型
通过构建遗传力模型,分 析数量性状的遗传变异程 度,并估计遗传力和相关 参数。
遗传相关模型
通过构建遗传相关模型, 分析不同数量性状之间的 遗传相关控制的群体遗传现象, 通过混合模型进行基因型 和环境交互作用的分析。
数量性状遗传在自然界中广泛存在,如人的身高、 体重、智力等都属于数量性状。
数量性状遗传的特点
数量性状遗传具有连续变异的 特点,即在一个群体中,个体 的表现型值可以连续变化。
数量性状遗传受多个基因位点 的影响,这些基因位点通常具 有微效作用,即每个基因位点 对表现型的影响较小。
数量性状遗传还受到环境因素 的影响,环境因素可以影响个 体表现型值的变异范围和分布 。
数量性状遗传在动物育种中的应用
生长速度
通过研究动物生长速度的数量性 状遗传,育种家可以培育出生长 快速的动物品种,提高养殖效益
。
繁殖性能
通过选育具有优良繁殖性能的数 量性状基因,可以提高动物的繁
殖效率,加速品种改良进程。
抗病性
通过研究动物抗病性的数量性状 遗传,育种家可以培育出具有较 强抗病能力的动物品种,降低养
利用新一代测序技术和遗传资源发掘,精细定位和克隆控制数量性状的基因或基因组区域 。
解析数量性状基因的互作网络
研究基因之间的相互作用关系,解析数量性状形成的复杂网络调控机制。
探索表观遗传修饰对数量性状的影响
研究DNA甲基化、组蛋白修饰等表观遗传修饰对数量性状表达的调控作用。
加强数量性状遗传与其他学科的交叉研究
03
数量性状遗传分析方法
统计分析方法
01
02
03
方差分析
通过比较不同群体或处理 组之间的变异程度,确定 数量性状是否受遗传控制 。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第十三章数量性状的遗传[关闭窗口]本章习题1.解释下列名词:广义遗传率、狭义遗传率、近交系数、共祖系数、数量性状基因位点、主效基因、微效基因、修饰基因、表现型值、基因型与环境互作广义遗传率:通常定义为总的遗传方差占表现型方差的比率。
狭义遗传率:通常定义为加性遗传方差占表现型方差的比率。
近交系数:是指个体的某个基因位点上两个等位基因来源于共同祖先某个基因的概率。
共祖系数:个体的近交系数等于双亲的共祖系数。
数量性状基因位点:即QTL,指控制数量性状表现的数量基因在连锁群中的位置。
主效基因:对某一性状的表现起主要作用、效应较大的基因。
微效基因:指一性状受制于多个基因,每个基因对表现型的影响较小、效应累加、无显隐性关系、对环境敏感,这些基因称为微效基因。
修饰基因:对性状的表现的效应微小,主要是起增强或减弱主基因对表现型的作用。
表现型值:是指基因型值与非遗传随机误差的总和即性状测定值。
基因型与环境互作:数量基因对环境比较敏感,其表达容易受到环境条件的影响。
因此,基因型与环境互作是基因型在不同环境条件下表现出的不同反应和对遗传主效应的离差。
2.质量性状和数量性状的区别在哪里?这两类性状的分析方法有何异同?答:质量性状和数量性状的区别主要有:①. 质量性状的变异是呈间断性,杂交后代可明确分组;数量性状的变异则呈连续性,杂交后的分离世代不能明确分组。
②. 质量性状不易受环境条件的影响;数量性状一般容易受环境条件的影响而发生变异,而这种变异一般是不能遗传的。
③. 质量性状在不同环境条件下的表现较为稳定;而控制数量性状的基因则在特定时空条件下表达,不同环境条件下基因表达的程度可能不同,因此数量性状普遍存在着基因型与环境互作。
对于质量性状一般采用系谱和概率分析的方法,并进行卡方检验;而数量性状的研究则需要遗传学方法和生物统计方法的结合,一般要采用适当的遗传交配设计、合理的环境设计、适当的度量手段和有效的统计分析方法,估算出遗传群体的均值、方差、协方差和相关系数等遗传参数等加以研究。
3.叙述表现型方差、基因型方差、基因型×环境互作方差的关系。
估计遗传协方差及其分量在遗传育种中有何意义?答:表现型方差由基因型方差(V G)、基因型×环境互作方差(V e)和环境机误方差()构成,即,其中基因型方差和基因型×环境互作方差是可以遗传的,而纯粹的环境方差是不能遗传的。
由于存在基因连锁或基因的一因多效,生物体的不同数量性状之间常存在不同程度的相互关连。
在统计分析方法中常用协方差来度量这种相互关联的变异程度。
由于遗传方差可以进一步区分为基因型方差和基因型×环境互作方差等不同的方差分量,故遗传协方差也可进一步区分为基因型协方差和基因型×环境互作协方差等分量。
在作物遗传改良过程中,对某一性状进行选择时常会引起另一相关性状的变化,为了取得更好地选择效果, 并使一些重要的性状能够得到同步改良, 有必要进行性状间的协方差即相关性研究。
如基因加性效应对选择是有效的, 细胞质效应亦可通过母本得以传递,因此当育种的目标性状不易测定或遗传率较低、进行直接选择较难取得预期效果时, 利用与其具有较高加性相关和细胞质相关的其它性状进行间接选择, 则较易取得育种效果。
显性相关则是控制性状的有关基因的显性效应相互作用而产生的相关性, 杂交一代中表现尤为强烈, 在杂种优势利用中可以加以利用。
但这种显性相关会随着世代的递增和基因的纯合而消失, 且会影响选择育种中早代间接选择的效果, 故对于显性相关为主的成对性状应以高代选择为主。
所以, 进行各种遗传协方差分析更能明确性状间相关性的遗传本质, 有利于排除环境因素对间接选择的影响,取得更好的选择效果,对于作物的选择育种具有重要的指导意义。
4.基于对数量性状遗传本质的理解,叙述数量性状的多基因假说的主要内容。
答:在遗传机制方面,数量性状受多基因控制,基因与基因间的关系错综复杂; 数量基因的表达对环境条件的变化比较敏感,基因的作用与环境条件的影响混杂在一起。
因此,数量性状的多基因假说的主要内容是:⑴.数量性状受制于多对微效基因或称多基因的联合效应;⑵.各对微效基因的效应相等而且是累加的,故又可称是累加基因;⑶.各对基因对某一性状的效应微小,多基因不能予以个别的辨认,只能按性状的表现作为一个多基因体系进行研究;⑷.微效基因之间无显隐性关系,一般用大写字母表示增效、小写字母表示减效作用;⑸.微效基因对环境敏感,因而数量性状的表现易受环境的影响而发生变化;⑹.微效基因具有多效性,除对数量性状起微效多基因的作用外,对其它性状有时也可能产生一定的修饰作用;⑺.微效基因和主效基因均处于细胞核的染色体上,具有分离、重组、连锁等性质。
5.叙述主效基因、微效基因、修饰基因对数量性状遗传作用的异同。
答:主效基因、微效基因、修饰基因在数量性状遗传中均可起一定的作用,其基因表达均可控制数量性状的表现。
但是它们对数量性状所起的作用又有所不同,主效基因的遗传效应较大,对某一数量性状的表现起着主要作用,一般由若干个基因共同控制该性状的遗传;修饰基因的遗传效应微小,主要是对主效基因起修饰作用,起增强或减弱主基因对表现型的作用;而微效基因是指控制数量性状表现的基因较多,而这些基因的遗传效应较小,它们的效应是累加的,无显隐性关系,对环境条件的变化较敏感,且具有一定的多效性,对其它性状有时也可能产生一定的修饰作用。
6.什么是普通遗传率和互作遗传率?他们在育种实践上有何指导意义?答:遗传率是指基因型方差(V G)占表型总方差(V p)的比值,它是衡量基因型变异和表型总变异相对程度的遗传统计量。
遗传率反映了通过表型值预测基因型值的可靠程度,表明了亲代变异传递到子代的能力。
同时也可以作为考查亲代与子代相似程度的指标。
由于导致群体表现型产生变异的遗传原因可以进一步区分为由遗传主效应产生的普通遗传变异和由基因型×环境互作效应产生的互作遗传变异,故遗传率可以分解为普通遗传率和互作遗传率两个分量。
其中普通遗传率是指由遗传主效应引起的那部分遗传率,一般指遗传方差占表现型方差的比率;互作遗传率是指由基因型×环境互作效应引起的那部分遗传率,一般指基因型×环境互作方差占表现型方差的比率。
育种实践表明,根据遗传率的大小可以决定不同性状的选择时期和选择方法,这对于改进育种方法,避免育种工作的盲目性和提高育种效果是很有效的。
一些遗传率较高的性状,可在杂种的早期世代进行选择,收效比较显著:而对于遗传率较低的性状,则需要在杂种后期世代进行选择才能收到更好的效果。
一般而言,当数量性状的基因型×环境互作效应越强,其互作遗传率就会越大,该性状的遗传表现就越易因环境而异,通过选择只能获得适应某一年份或某一特殊环境(如某一生态区域)的品种或组合;而基因型×环境互作效应小的性状则其普通遗传率就会越大,容易通过选择来改良育种材料的遗传组成,获得能够适应不同年份或不同环境的品种(组合)。
故普通遗传率适用于不同环境条件下的选择,而互作遗传率则只适用于某一特定条件下的选择。
某一年份或环境下的选择总效益,可以根据总的遗传率大小(普通遗传率 + 某一环境中的互作遗传率)进行预测和分析,以了解通过选择个体或个体群改良其基因型的准确性和选择效率。
7.什么是基因的加性效应、显性效应及上位性效应?它们对数量性状遗传改良有何作用?答:基因的加性效应(A):是指基因位点内等位基因的累加效应,是上下代遗传可以固定的分量,又称为"育种值"。
显性效应(D):是指基因位点内等位基因之间的互作效应,是可以遗传但不能固定的遗传因素,是产生杂种优势的主要部分。
上位性效应(I):是指不同基因位点的非等位基因之间相互作用所产生的效应。
上述遗传效应在数量性状遗传改良中的作用:由于加性效应部分可以在上下代得以传递,选择过程中可以累加,且具有较快的纯合速度,具有较高加性效应的数量性状在低世代选择时较易取得育种效果。
显性相关则与杂种优势的表现有着密切关系,杂交一代中表现尤为强烈,在杂交稻等作物的组合选配中可以加以利用。
但这种显性效应会随着世代的递增和基因的纯合而消失, 且会影响选择育种中早代选择的效果, 故对于显性效应为主的数量性状应以高代选择为主。
上位性效应是由非等位基因间互作产生的,也是控制数量性状表现的重要遗传分量。
其中加性×加性上位性效应部分也可在上下代遗传,并经选择而被固定;而加性×显性上位性效应和显性×显性上位性效应则与杂种优势的表现有关,在低世代时会在一定程度上影响数量性状的选择效果。
8.什么是基因的加性×环境互作效应、显性×环境互作效应及上位性×环境互作效应?它们对数量性状遗传改良作用与基因的遗传主效应有何异同?答:加性×环境互作效应(AE):是指基因加性效应与环境互作产生的遗传效应,是一部分可以在上下代传递、并加以固定的遗传效应,但会因环境条件的变化而产生较大差异。
显性×环境互作效应(DE):是指基因显性效应与环境互作产生的遗传效应,是一部分可以遗传、但不能固定的遗传效应,主要与杂种的优势表现有关,这部分效应也会因环境的变化而异。
上位性×环境互作效应(IE):是指基因上位性效应与环境互作产生的遗传效应,在不同环境中会有较大差异。
其中加性×加性上位性互作效应部分经选择可被固定;而加性×显性上位性互作效应和显性×显性上位性互作效应与杂种优势的表现有关,在低世代时会在一定程度上影响数量性状的选择效果。
不同的环境互作效应与遗传主效应的作用一样,对数量性状的遗传改良起着重要作用,可以影响数量性状的表现和选择效果。
但两者也有较大的差异,特别是一些受基因加性、显性或上位性等遗传主效应控制的数量性状在遗传改良中较为稳定,不同环境条件对这些数量性状的选择效果影响较小,通过选择容易获得适合不同年份或不同地点的育种材料或组合。
相反,某一数量性状的基因型与环境互作效应越强,该性状的遗传表现就越容易受到环境变化的影响,通过选择一般可以获得适合某一特定年份或某一特定环境的育种材料。
因此不同环境下的基因型稳定性对作物种子品质育种目标的制定非常重要。
9.以下是陆地棉4个亲本及其F1在8月9 日和9月3日的平均单株成铃数的分析资料(1981,1985)表1 方差和协方差计算结果:表2 基于群体均值的F1平均优势的预测结果(%)⑴. 估算8月9日和9月3日的普通广义遗传率、互作广义遗传率、普通狭义遗传率、互作狭义遗传率,说明这两个时期单株成铃数的遗传规律及其对选择育种的指导意义;⑵. 根据F1普通平均优势和互作平均优势的预测结果,评价不同杂交组合的杂种优势利用的潜力。