丁玉强-形态学-2006
昆虫几丁质合成酶B基因的研究进展

黑龙江农业科学2023(11):144-150H e i l o n g j i a n g A gr i c u l t u r a l S c i e n c e s h t t p ://h l j n y k x .h a a s e p.c n D O I :10.11942/j.i s s n 1002-2767.2023.11.0144王晓曦,刘兴龙,王宇,等.昆虫几丁质合成酶B 基因的研究进展[J ].黑龙江农业科学,2023(11):144-150.昆虫几丁质合成酶B 基因的研究进展王晓曦1,刘兴龙1,王 宇1,王克勤1,樊 东2(1.黑龙江省农业科学院植物保护研究所/农业农村部哈尔滨作物有害生物科学观测实验站,黑龙江哈尔滨150086;2.东北农业大学植物保护学院,黑龙江哈尔滨150030)摘要:几丁质是昆虫的主要组成成分,由于几丁质不存在于植物和无脊椎动物中,被认为是环境友好型农药的设计靶标㊂几丁质合成酶B 是几丁质合成路径中的最后一个关键酶,近些年已成为国内外研究的热点内容㊂根据国内外对昆虫几丁质合成酶B 基因功能的研究,综述了几丁质合成酶B 对几丁质合成的调控研究进展,包括时空表达特性㊁基因功能㊁基因在害虫防治中的研究㊁并对R N A 干扰技术应用存在的问题及解决途径进行了展望㊂关键词:昆虫;几丁质;几丁质合成酶B ;环境友好型农药收稿日期:2023-05-18基金项目:黑龙江省重点研发计划项目(G A 23B 010);植物保护研究所青年基金项目(z b s qn 2023-3)㊂第一作者:王晓曦(1992-),女,硕士,研究实习员,从事作物虫害的防治研究㊂E -m a i l :2323458775@q q .c o m ㊂通信作者:樊东(1969-),男,博士,教授,博导,从事农作物昆虫及有害生物的防控研究㊂E -m a i l :d n f d @163.c o m ㊂几丁质又称甲壳素(C h i t i n ),为β-(1,4)-N -乙酰基-D -葡萄糖胺(G l c N A c )的线性高分子,含量仅次于膳食纤维素,为自然界中第二重要大分子化合物,占全部昆虫干质量的40%左右,广泛分布在昆虫体表,中肠和肠道围食膜内㊂它是进入昆虫体内的第一道屏障,有保护中肠上皮细胞免受食物颗粒机械损伤㊁阻止病原体入侵及为昆虫解除体内有毒物质等效果,对昆虫的生长和变态发育至关重要[1-2]㊂几丁质广泛存在于节肢动物㊁腕足类㊁软体动物㊁昆虫㊁真菌㊁线虫㊁藻类㊁原生生物㊁轮虫㊁海绵和墨鱼中㊂围食膜(P e r i t r o ph i c M a t r i x ,P M )是昆虫中肠细胞分泌的,隔开肠内容物与肠上皮细胞的一种半透膜[3]㊂几丁质是围食膜的主要成分之一,含量为3%~13%,构成围食膜的几丁质微纤丝具有高弹性,保护了围食膜的韧性和通透性[3-4]㊂昆虫几丁质的合成共8种酶参与,可以实现复杂的生理和生化反应,从而合成昆虫几丁质㊂1962年,C a N d y 与K i l b y 首次发现了昆虫体内甲壳素的生物合成途径,起步于海藻糖,终止于甲壳素,先后包括了海藻糖酶(t r e h a l a s e )㊁己糖激酶(h e x o k i N a s e )㊁葡萄糖-6-磷酸异构酶(gl u c o s e -6-p h o s ph a t e i s o m e r a s e )㊁谷氨酸盐:果糖-6-磷酸转氨酶(g l u t a m i N e :f r u c t o s e -6-p h o s p h a t e a m i n o t r a n s f e r a s e )㊁葡糖胺-6-磷酸-N -乙酰转移酶(gl u c o s a m i n e -6-p h o s p h a t e n -a c e t y l t r a n s f e r a s e )㊁磷乙酰氨基葡萄糖刺激变位酶(p h o s p h o a c e t y l gl u c o s a m i n e m u t a s e )㊁U D P -N -乙酰葡糖胺焦磷酸化酶(U D P -N -a c e t yl -g l u c o s a m i n e p y r o p h o s p h o r yl a s e )和几丁质合成酶(c h i t i ns yn t h a s e ,C H S )[5]㊂几丁质合成的路径,已有研究中最深入广泛之一是C H S ,即几丁质合成的最后一步关键酶㊂化学药剂型已使高危险性害虫形成了一定耐药性[6],且对人㊁作物㊁环境和食品的安全产生威胁,亟需新的绿色防控手段[7]㊂几丁质合成路径的异常会导致昆虫畸形㊁无法正常蜕皮及死亡率升高㊂由于在植物和脊椎动物体内不合成几丁质[8],控制几丁质合成的相关酶,就可以为新型环境友好型长效杀虫剂的设计与研制提出了潜在的优质靶标,进行害虫预防㊂近些年基于C H S 的生物化学抑制剂,以及基于R N A 干扰法(R N Ai n t e r f e r e n c e ,R N A i)的有害生物防治新途径的探讨和进展,也成为农业害虫防治方面的学术前沿和热点[9]㊂如对苯甲酰基脲类,作为我国最早㊁最常用的一类害虫甲壳素合成抑制剂,目前已经产生了许多商业化产品[10]㊂近年在害虫防治中随着R N A 干扰技术的开发与应用[11],凭借其专一化高㊁潜伏的靶点性强㊁对生态稳定等特点,以及运用基因表达的下调方式,在害虫防控应用研究上取得了新突破㊂目前鳞翅目㊁鞘翅目㊁双翅目和44111期 王晓曦等:昆虫几丁质合成酶B 基因的研究进展直翅目等昆虫的几丁质合成酶B 基因c D N A 序列已经克隆出来,部分进行了时空表达㊁基因功能㊁抑制剂筛选及基因干扰等的研究㊂本文主要综述了对几丁质合成途径中几丁质合成酶B 的构成㊁几丁质合成酶B 基因的空间表达特征㊁基因功能㊁基因结构㊁在害虫防控中的研究进展,并对存在问题和可能的破解路径等做出了比较系统的归纳总结,以期为几丁质合成的控制机理的进一步深入研究奠定基础,同时也为利用R N A i 技术进行害虫防控研究提出有价值的理论指导㊂1 昆虫几丁质合成酶B目前昆虫的几丁质合成酶C H S 包括了几丁质合成酶A (c h i t i n s y n t h a s eA ,C H S A )和几丁质合成酶B (c h i t i n s yn t h a s eB ,C H S B )两种,或称为C H S 2(E C2.4.1.16),功能为结合中肠上皮细胞的围食膜中的甲壳素,并在其中特异性表达[12]㊂几丁质决定了昆虫的生长发育,调控C H S B 基因的表达来防治害虫会有一定成效[13-14],如抑制C H S B 的活性会导致昆虫饥饿致死[15]㊂2 昆虫C H S B 的结构特性C H S B 为160~180kD 分子量的大型跨膜蛋白体[16],昆虫的C H S B 含14~17个跨膜螺旋及3个相同结构域A ㊁B 和C ,结构域A 在C H S B 的N 端,物种之间氨基酸序列相似性最低;结构域B 在C H S B 的中段,是高度保守的蛋白质催化中心,物种之间氨基酸序列相似性较高,含4个标签序列C A T MWH X T ,D X D ,E D R 和Q R R RW ;结构域C 在C H S B 的C 端,物种之间氨基酸序列差异性较高[12,17]㊂目前未有报道C H S B 有可变剪接[18]㊂第一个C H S B 基因是在1998年时由T h i r e o s 所鉴定的黑腹果蝇基因D m C H S B [19],现已从多个昆虫物种中克隆出C H S B 基因,目前已知黑腹果蝇(D r o s o p h i l am e l a n o g a s t e r )[20]㊁赤拟谷盗(T r i b o l i u m c a s t a n e u m )[21]㊁烟草天蛾(M a n d u c as e x t a )[22]㊁甜菜夜蛾(S p o d o pt e r a e x i g u a )[23]㊁家蚕(B o m b yxm o r i )[24]㊁亚洲玉米螟(O s t r i n i a f u r n a c a l i s )[17]㊁冈比亚按蚊(A n o ph e l e s ga mb i a e )[25]㊁东方黑腹果蝇(B ac t r o c e r ad o r s a l i s )[26]㊁飞蝗(L o c u s t a m i g r a t o r i a )[27]㊁稻纵卷叶螟(C n a p h a l o c r o c i sm e d i n a l i s )[28]㊁棉铃虫(H e l i c o v e r pa a r m i g e r a )[29]㊁致倦库蚊(C u l e x q u i n q u e f a s c i a t u s )[30]㊁马铃薯甲虫(L e p t i n o t a r s ad e c e m l i n e a t a )[31]㊁梨小食心虫(G r a ph o l i t h a m o l e s t a )[32]和草地贪夜蛾(S p o d o p t e r a f r u g i pe r d a )[33]等50多种害虫的C H S B 基因都已经克隆出来,并在G e n B a n k 上登录㊂3 昆虫C H S B 基因的时空表达特性对于昆虫C H S B 不同发育阶段和不同组织的表达模式研究上,整理了已发表文献的相关研究进展(表1)㊂昆虫不同发育阶段的研究表明,C H S B 基因在昆虫发育的各个阶段均有表达㊂可见,C H S B 基因在昆虫各个发育阶段进行调控,但相对表达量存在差异㊂飞蝗L m C H S 2基因和冈比亚按蚊A gC H S B 基因在受精卵发育的前期和中期基本不能表达[25,27,34],但在卵发育后期的表现数量却迅速增加㊂在卵发育时期高表达有何特定的功能尚待深入研究㊂大部分C H S B 在昆虫老熟幼虫和成虫中表达量相对较高,有部分在预蛹期表达量较高,如黑腹果蝇D m e C H S 2基因和梨小食心虫G m C H S 2基因[32,35],在低龄幼虫和蛹期表达量很低,可能与取食特性有关,它们取食水果,不需要过多的消化,所以不必形成大量围食膜㊂可见C H S B 基因在昆虫取食量高的时期高表达,以形成更大面积的围食膜㊂不同类型昆虫在不同阶段表达量的差异,可能和虫体中不同的激素代谢阶段密切相关㊂不同昆虫的发育历期不一致㊂昆虫不同组织的表达模式研究表明,除梨小食心虫G m C H S 2在前肠表达量最高外[32],其余昆虫在中肠中表达量最高,其次部分昆虫在前肠㊁后肠和马氏管表达量较高,C H S B 在昆虫取食过程中的肠道表达,参与围食膜的形成㊂飞蝗L m C H S 2也在胃盲囊中表达,推测可能是因为昆虫分属不同的目,与体内消化结构存在差异有关[27,34],锈赤扁谷盗C f C H S 2在腹部表达量最高,可能与中肠组织残留腹部有关[36]㊂茶尺蠖C S 2在形成围食膜期间表达[37]㊂说明C H S B 是昆虫中肠的特异性蛋白,合成中肠围食膜上的几丁质,C H S B 合成围食膜的β-几丁质和γ-几丁质,对昆虫的生长和发育起到关键作用㊂541黑 龙 江 农 业 科 学11期表1 昆虫不同发育阶段和不同组织C H S B 基因的表达目昆虫及其C H S B 基因登录号发育阶段组织参考文献鳞翅目(L e p i d o pt e r a )甜菜夜蛾S e C H S BD Q 9129294龄和5龄第1天表达量较高;4龄和5龄第2天及预蛹期表达量较低在中肠有最高表达[23][38]稻纵卷叶螟C m C H S BA J G 44539.1成虫期的表达量高于幼虫期4龄在中肠有最高表达,是表皮表达量的5倍;脂肪体中也有表达[28][39]梨小食心虫G m C H S 2K Y 242360预蛹期和成虫期表达量最高预蛹第1天在前肠的表达量最高,其次是后肠,其余组织表达量很少或不表达[32]家蚕B mC H S BJ Q 0741753~5龄中,3龄的第1天至第3天表达量最高,蜕皮后表达量下降,周期性变化5龄第3天在中肠特异性表达,头部和表皮有少量表达,丝腺㊁脂肪体㊁睾丸㊁卵巢和血细胞中几乎没有表达[24][40]亚洲玉米螟O fC H S B E U 3760264龄至成虫期中,预蛹期和5龄有最高表达,4~5龄蜕皮之间基因表达量降低,5龄第6天停止进食时表达量瞬降[17]粘虫M s C H S BK Y 3487761龄最后1天的基因表达量最低,3龄第1天和蛹期第1天的基因表达量最高5龄第2天在中肠特异性表达,其次为前肠后肠,体壁和脂肪体几乎没有可检测的表达[41]茶尺蠖(部分C S 2序列)E U 2901473~5龄幼虫取食阶段表达量高,蛹期不表达在中肠围食膜中表达[37]草地贪夜蛾S fC H S B M Z 364352主要在中肠中表达[33]小地老虎A i C H S 2O L 8267546龄幼虫表达量最高主要在前肠和中肠中表达[42]鞘翅目(C o l e o pt e r a )赤拟谷盗T c C H S 2A Y 291477老熟幼虫和成虫阶段表达量高[13]马铃薯甲虫L d C H S BK T 9647423龄第1天在中肠有最高表达,其次是马氏管[31]锈赤扁谷盗C fC H S 2MH 234580幼虫阶段1龄的相对表达量最多,随着龄期的增加表达量逐渐减少,在蛹期基因的表达量最低,成虫期表达量最高成虫在腹部表达量最高,其次中肠㊂在头部㊁胸部㊁脂肪体中有少量的表达,表皮几乎没有表达[36]玉米根萤叶甲D v v C H S 2MH 2568563龄幼虫期和成虫期表达量最高,其次是卵㊁1龄幼虫期和2龄幼虫期,在蛹期的表达量最低成虫在中肠有最高表达,在前肠㊁后肠和组织中表达极低[43]双翅目(D i pt e r a )致倦库蚊C qC H S 2A G A P 001205幼虫期表达量低,卵和蛹期表达量较低但差异小,成虫期表达量最高[30]东方果蝇(桔小实蝇)B d C H S BK C 354694卵到3龄幼虫表达量上升,成虫期表达量最高3龄在中肠有最高表达,其次是马氏管,脂肪体中表达最低[44]冈比亚按蚊A gC H S B X P _321951.2在成虫的表达量最高,在卵㊁4龄幼虫和蛹表达量均很低,尤其是幼虫阶段在前肠的表达水平最高,显著高于中肠[25]黑腹果蝇D m e C H S 2NM _079485幼虫期表达量较少,预蛹期显著升高[35]三叶草斑潜蝇L t C H S 2O N 453844蛹期表达量最高在成虫的头㊁胸㊁腹和肠中无显著差异[45]直翅目(O r t h o pt e r a )飞蝗L m C H S 2G U 067731卵期的2~11d 基本无表达,12~14d 表达量猛增;1~3龄表达量逐渐增加,3龄最高,之后保持在中肠和胃盲囊中特异性表达[27][34]64111期 王晓曦等:昆虫几丁质合成酶B 基因的研究进展4 昆虫C H S B 基因功能的研究进展4.1 进食、饥饿对虫体生长发育和C H S B 基因表达的影响各种环境因素中,食物对昆虫的繁殖与变态的影响显著㊂相关研究表明多数农业害虫幼虫期的取食对于生殖具有重要影响[46]㊂为了探究进食是否对C H S B 基因表达有影响,进而影响中肠的取食和消化,对试验昆虫进行饥饿处理㊂有研究选取5龄第2天飞蝗若虫,饥饿处理12,24和48h 之后,再复食0.5,1.0和4.0h ,解剖中肠观察,结果发现围食膜遭到了严重破坏,说明围食膜的完整性对害虫生长与发育至关重要,而饥饿也可能影响基因在中肠的表达;但由于饥饿持续时间拉长,L m C H S 2基因表达量逐步减少,在重新取食后表达量逐渐恢复[27]㊂对4龄桔小实蝇进行饥饿处理,饥饿组B d C H S B 基因表达量低于先饲喂组,且差别不明显;先饲喂-饥饿后,表达量变化并不明显;而先饥饿-饲喂后,基因的表达量明显增加至5倍以上,并对桔小实蝇几丁质含量的影响明显[44]㊂同样,对埃及伊蚊成虫饥饿-复食处理,发现饥饿时中肠A a C H S B 基因的表达量降低,恢复吸食血液后基因的表达量升高[47]㊂4龄第2天粘虫,喂食-饥饿,M s C H S B 基因表达量明显比持续喂食和持续饥饿低;饥饿-喂食,表达量明显比持续喂食和持续饥饿高㊂饥饿处理后M s C H S B 基因的表达量变化与中肠几丁质含量的变化具有相同的趋势[41]㊂研究表明,饥饿处理对C H S B 基因的表达有显著的抑制作用,验证了饥饿对昆虫的取食和中肠的消化与吸收具有一定影响,也进一步验证该基因在中肠大量表达㊂4.2 蜕皮激素对C H S B 基因表达的影响蜕皮激素(20E ,h y d r o x y e c d ys o n e )20E 诱导昆虫蜕皮和变态,保幼激素(J u v e n i l eH o r m o n e ,J H )为了维持平衡20E 带来的形态改变,使幼虫在蜕皮后始终处于雏体状态,它对控制昆虫变态至关重要㊂昆虫的生长发育,是由20E 和J H 共同配合完成的[48]㊂昆虫内源保幼激素较多时,就维持它幼虫或若虫的各个发育阶段形态,不会变态成下一个发育阶段的蛹或成虫;昆虫体内J H 含量较少且20E 较多时,全变态发育的幼虫才能变态成蛹[49]㊂目前20E 对C H S B 基因表达影响的研究还很少,已有的研究都表明,20E 可以诱导C H S B 基因的表达㊂对3龄幼虫和刚化蛹的果蝇C H S B 基因表达分析中,C H S B 基因的转录会响应20E 的刺激,数小时后表达水平上调,这与形成蛹前表皮的时间相吻合[20]㊂有学者向4龄蜕皮后2h 内的家蚕采用1μg㊃μL -1的20E 处理6h ,结果发现B m C H S B 基因表现明显优于对照组,诱导作用更明显;并在短时间控制了B mC H S B 基因的表达,后又得到了恢复[24,40]㊂对4龄第2天的粘虫注入20E ,20E 蜕皮激素对M s C H S B 基因的表达有明显诱导效果,对注入了10μg㊃μL -1的20E6h 的诱导效果尤为突出,相比对照,表达量升高了32.0%[41]㊂给2龄初和3龄第2天的马铃薯甲虫幼虫喂食20E 处理的马铃薯叶片12h 后,L d C H S B 基因表达水平均显著升高[50]㊂说明20E 影响C H S B 对几丁质的合成,从而影响昆虫生长发育㊂基因的表现不仅仅是受外源蜕皮激素单一控制,还和内源激素关系密切㊂外源20E 在与J H 协同作用下怎样控制C H S B 基因表达,也值得进一步深入研究㊂5 C H S B 基因在害虫防治中的研究进展5.1 几丁质合成酶抑制剂对靶标C H S B 的影响几丁质合成抑制剂同时还具有植物生长调节剂的功效,对害虫中的几丁质合成关键酶活性影响较大,通过其底物竞争性和C H S 结合,进而影响几丁质的正常合成,害虫畸形或死亡㊂有效霉素(V a l i d a m y c i n )又称井冈霉素(J i n g a n m y c i n ),是一类几丁质合成酶抑制剂,具有杀虫效果[38]㊂对4龄第2天的粘虫幼虫注射有效霉素,幼虫生长发育缓慢,M s C H S B 基因的表达量下降95%㊂有效霉素处理后粘虫M s C H S B 基因的表达量变化与中肠几丁质含量的变化具有相同的趋势㊂这也证实了在粘虫的形成与发育过程中,M s C H S B 基因的水平与中肠几丁质浓度之间的正相关关系[41]㊂定虫隆是一种昆虫生长调节剂,有研究用定虫隆饲喂锈赤扁谷盗,龄期延长㊁体重下降㊁几丁质含量下降,C fC H S B 基因表达量提高约27%㊂与其他研究得到相反的结果,推测C H S B受到了抑制,锈赤扁谷盗为维持自身稳定发育,提高C H S B 基因的表达,而触发了补偿性反馈机制,由此推测定虫隆可能作用于C H S B [51]㊂氟苯脲㊁除虫脲和虱螨脲是高效的几丁质合成抑制剂,是目前应用广泛的苯甲酰基脲类杀虫剂㊂4龄马铃薯甲虫经氟苯脲处理36h 后,L d C H S B 基因表达下调[52]㊂除虫脲处理对1龄桔小实蝇幼虫B d C H S B 基因没有显著影响[44]㊂虱螨脲处理对草地贪夜蛾S fC H S B 基因的表达无显著变化[33],而使小地老虎A i C H S 2基因表达显著提高[42]㊂几丁质合成抑制剂对主要天敌和非靶标生物的危害都很小,对昆虫也难以形成耐药性,因此可与低毒量和低残留危害性都很小的长效化学农药混合,以实现增效目的㊂同时也是几丁质合成抑741黑 龙 江 农 业 科 学11期制剂防治农作物害虫,在田间大范围使用的基础㊂随着未来对作用机理的深入研究,几丁质合成抑制剂类杀虫剂将在害虫化学防治中发挥重要作用㊂5.2 作为R N A i 的靶标基因R N A 干扰(R N Ai n t e r f e r e n c e ,R N A i)使靶基因沉默的技术特异性且高效,技术简便,目前已被广泛用于农业上主要的害虫绿色防控新技术开发中㊂现有许多研究利用R N A i 技术来抑制昆虫C H S B 基因的表达,取得了一定成效㊂有研究利用注射d s R N A 的方法干扰赤拟谷盗幼虫T c C H S B 基因的表达,幼虫身体皱缩,围食膜基本不再生成[13]㊂R N A 干扰了赤拟谷盗膜上的海藻糖酶基因(T r e -2),干扰中肠C H S B ,使中肠甲壳素水平降低了20%以上[53]㊂沉默飞蝗的L m C H S 2基因,引起飞蝗中肠围食膜残缺不全,中肠和胃盲囊生长减少,主食无法消化和吸收,最后由于饥饿而死去,蛛网膜下腔出血可达到75%以上[27];干扰3龄家蚕幼虫B m C H S B 基因的表达致使家蚕大部分难以正常蜕皮[4]㊂喂食2龄和4龄马铃薯甲虫幼虫细菌表达的d s C H S B ,幼虫进食量减少㊁生长缓慢,且中肠几丁质含量降低[31]㊂喂食方式干扰3龄甜菜夜蛾4h 后,发现对试验组幼虫阶段基本无影响,使预蛹和成虫发育受阻,20%和25%无法化蛹和羽化[38]㊂注射方式干扰稻纵卷叶螟幼虫C m C H S 2的表达,4~5d 后表达量下降37%~50%,取食量减少,试验组部分幼虫蜕皮受阻,虫体显著变轻变小变黑,死亡率约41%[54-56]㊂玉米根萤叶甲成虫取食d s C H S 2,干扰对D v v C H S 2基因最高沉默效率达90%,取食1d 后,成虫D v v C H S 2基因表达量显著降低[43]㊂沉默锈赤扁谷盗C f C H S 2基因,会使幼虫蜷缩干瘪,无法正常爬行,导致死亡,7d 后,死亡率达63%且围食膜缺失,肠道上皮细胞严重破损㊂与上述实验结果不同,说明C H S 2在锈赤扁谷盗的中肠围食膜的生成途径中起到关键性的作用㊂持续干扰发现C fC H S 2基因有过表达现象,基因表达量先下降后上升再下降,这说明在C f C H S 2基因被d s R N A 沉默后,昆虫为了维持机体的稳定,使C fC H S 2基因的表达量升高,而形成的一种补偿反馈机制[36]㊂对4龄粘虫幼虫注射d s M s C H S B ,M s C H S B 基因的表达量下降,表明R N A 干扰成功抑制了M s C H S B 基因的表达㊂处理24和48h 后,该基因的表达水平分别下降了82.1%和86.0%,处理96h 后粘虫的死亡率最高,达46.7%㊂基因沉默影响了粘虫中肠几丁质的合成,可能破坏了中肠围食膜的通透性,进而影响进食导致虫体生长发育缓慢,无法正常蜕皮,产生异形,甚至死亡[41]㊂对小地老虎注射d s A i C H S 2干扰,化蛹率降低30%以上[42]㊂干扰三叶草斑潜蝇L t C H S 2在蛹中的表达,羽化率显著低于对照㊂综上研究,C H S B 在昆虫中肠围食膜的形成过程中发挥重要作用,影响昆虫生长发育[45]㊂干扰C H S B 基因的表达,会导致昆虫中肠围食膜受到破坏㊁降低中肠围食膜几丁质含量㊁进食量减少㊁身体皱缩变轻㊁生长缓慢㊁无法正常蜕皮化蛹和羽化㊁变黑㊁死亡率升高㊂6 展望R N A i 方法中因对C H S B 靶基因沉默方式的高度特异性和高效的优点,以及研究过程的简便性,目前已被广泛用于重大农业有害生物的防控研发技术中㊂C H S B 是重大农业有害生物形成与传播中的非常重要的基因,是潜在的杀虫有效靶标基因,也是将R N A i 方法应用于重大农业有害生物防治中的关键㊂如昆虫C H S B 是重大农业有害生物几丁质形成过程中的关键基因之一,但因为高等动㊁植物自身还没有形成这些蛋白质,所以被认为是比较好的对环境友好的杀虫剂靶标㊂体外喷洒d s R N A (核酸杀虫剂)对开展农作物害虫防治工作中有以下优点:第一,d s R N A 可高效专一地防治目标害虫,对天敌昆虫㊁作物和人畜健康没有影响;第二,d s R N A 可有效减少害虫;第三,可明显降低化学农药的施用量,从而有效保护了食品生产安全;第四,因为d s R N A 属于核酸化合物,对人类健康没有什么影响,是一种对人类真正友好的生物杀虫剂,它对降低农药残留和环境污染都具有重要的意义㊂目前,应用于d s R N A 的C H S B 基因作为靶标防控危险害虫的瓶颈在于:第一,实验室中通常制备d s R N A 是通过测定试剂盒中微量制备的,如果作为核酸外施或喷洒药物,就必须将大量的d s R N A 使用在田间㊂其二,d s R N A 合成成本过高㊂为此,利用害虫体内的高效靶标C H S B 基因的R N A i ,如何工厂化批量合成d s R N A ,且降低成本,还有待未来研究解决㊂农业害虫对药剂抗性也与靶标C H S 基因有关㊂比如,经虱螨脲诱导处理后,S f C H S A 基因的表达量显著提高,表明S fC H S A 基因在草地贪夜蛾对虱螨脲的抗性中发挥的作用大于S fC H S B 基因[33]㊂了解害虫对杀虫剂的抗药性水平㊁杀虫剂对害虫的主要作用机制以及害虫对杀虫剂的抗性机制,将有助于保护作物,延长该杀虫剂的使用寿命㊂另外,在与生物纳米材料偶联包被㊁脂质体修复技术㊁与B T 类杀虫药的结合,以及多靶标联用技术等方面迫切需要系统深入地开展研究[18]㊂并使R N A i 技术在防治有害生物领域广泛获得应用㊂相信随着对甲壳素合成途径84111期 王晓曦等:昆虫几丁质合成酶B 基因的研究进展的挖掘与研究,以及该途径的相关酶基因对几丁质生成的控制效果的深入研究,基于靶标基因的R N A i 研究将有助于今后害虫的绿色防治研究,为农业害虫的有效防控提供理论依据㊂为以几丁质生成途径的酶基因为主要工作靶点的绿色杀虫剂研究,及发现更有效的几丁质合成酶抑制剂等绿色杀虫剂,奠定了必要的科学基础㊂参考文献:[1] L Y O N E TP .T r a i t e a n a t o m i q u e d e l a c h e n i l l e [M ].L aH a ye :G r o s s eP i n e t ,1762:25-36.[2] B A L B I A N IE G.E t u d e sa n a t o m i q u e s e th i s t o l o g i qu e s s u r l e t u b e d i g e s t i f d e s C r y p t o p s [J ].A r c h i v e s d e Z o o l o g i e E x p e r i m e n t a l e e tG e n e r a l e ,1890(8):1-82.[3] T A B A S H N I KBE ,Z HA N G M ,F A B R I C KJA ,e t a l .D u a lm o d eo fa c t i o n o f B t p r o t e i n s :p r o t o x i n e f f i c a c y a g a i n s t r e s i s t a n t i n s e c t s [J ].S c i e n t i f i cR e po r t s ,2015,5(1):15107.[4] K U U S K S ,S ØR L I E M ,V ÄL J AMÄEP .T h e p r e d o m i n a n tm o l e c u l a r s t a t eo fb o u n de n z y m ed e t e r m i n e st h es t r e n gt h a n dt y p eo f p r o d u c t i n h i b i t i o n i nt h eh y d r o l ys i so f r e c a l c i t r a n t p o l y s a c c h a r i d e s b yp r o c e s s i v e e n z y m e s [J ].J o u r n a l o fB i o l o gi c a l C h e m i s t r y,2015,290(18):11678-11691.[5] M E R Z E N D O R F E R H ,Z I MO C H L.C h i t i n m e t a b o l i s mi ni n s e c t s :s t r u c t u r e ,f u n c t i o na n dr e g u l a t i o no fc h i t i ns y n t h a s e s a n dc h i t i n a s e s [J ].J o u r n a lo fE x p e r i m e n t a lB i o l o g y,2003,206(24):4393-4412.[6] 朱江,邱星辉.昆虫抗药性相关细胞色素P 450基因的表达调控机制[J ].昆虫学报,2021,64(1):109-120.[7] 谢丹洁,刘云朋,汪海鹏,等.美国白蛾核型多角体病毒与四种常用化学杀虫剂的复配增效作用研究[J ].应用昆虫学报,2023,60(1):49-57.[8] Y U HZ ,L IN Y ,X I E Y X ,e t a l .I d e n t i f i c a t i o na n d f u n c t i o n a l a n a l y s i s o f t w o c h i t i n s yn t h a s e g e n e s i n t h e c o m m o n c u t w o r m ,S p o d o pt e r a l i t u r a [J ].I n s e c t s ,2020,11(4):253.[9] 汪芳,党聪,金虹霞,等.R N A 干扰技术在害虫防治中的应用及其安全性[J ].浙江大学学报(农业与生命科学版),2022,48(6):683-691.[10] 杨霞.几丁质合成酶抑制剂的研究进展[J ].福建农业,2015(2):90.[11] 陈静,张道伟,钱正敏.白背飞虱几丁质合成酶1基因的结构及特性研究[J ].生物技术通报,2018,34(1):195-201.[12] M E R Z E N D O R F E R H.I n s e c t c h i t i ns yn t h a s e s :ar e v i e w [J ].J o u r n a l o fC o m p a r a t i v eP h y s i o l o g y B ,2006,176(1):1-5.[13] A R A K A N E Y ,MU T HU K R I S H N A NS ,K R AM E R KJ,e t a l .T h e T r i b o l i u m c h i t i ns y n t h a s e g e n e s T c C H S 1a n d T c C H S 2a r e s p e c i a l i z e df o r s y n t h e s i so f e pi d e r m a l c u t i c l e a n dm i d g u t p e r i t r o p h i cm a t r i x [J ].I n s e c t M o l e c u l a rB i o l o g y,2005,14(5):453-463.[14] T I A N HG ,P E N G H ,Y A OQ ,e t a l .D e v e l o pm e n t a l c o n t r o l o f a l e p i d o p t e r a n p e s t S p o d o p t e r a e x i gu a b y i n g e s t i o n o f b a c t e r i a e x p r e s s i n g d s R N Ao fan o n -m i d g u t g e n e [J ].P L o S O n e ,2009,4(7):e 6225.[15] C HA U D HA R IS S ,N O H M Y ,MO U S S I A N B ,e ta l .K n i c k k o p f a n d r e t r o a c t i v e p r o t e i n s a r e r e qu i r e d f o r f o r m a t i o n o f l a m i n a r s e r o s a l p r o c u t i c l e d u r i n g e m b r y o n i c d e v e l o pm e n t o f T r i b o l i u mc a s t a n e u m [J ].I n s e c tB i o c h e m i s r y a n d M o l e c u l a r B i o l o g y,2015,60(3):1-6.[16] T R UMA NJW ,R I D D I F O R D L M.I n s e c td e v e l o pm e n t a l h o r m o n e sa n dt h e i r m e c h a n i s m o fa c t i o n [J ].H o r m o n e s B r a i na n dB e h a v i o r ,2002(2):841-873.[17] Q U MB ,L I U T ,Y A N GJ ,e t a l .T h e g e n e ,e x p r e s s i o n p a t t e r n a n d s u b c e l l u l a r l o c a l i z a t i o no fc h i t i ns yn t h a s eBf r o mt h e i n s e c t O s t r i n i a f u r n a c a l i s [J ].B i o c h e m i c a l a n dB i o p h ys i c a l R e s e a r c hC o m m u n i c a t i o n s ,2011,404(1):302-307.[18] 许静静,李思琪,任梦圆,等.昆虫几丁质合成关键酶功能及其R N A i 技术在害虫防治中的研究进展[J ].陕西农业科学,2022,68(8):10.[19] TH I R E O SG ,K A F E T Z O P O U L O SD.D N Ae n c o d i n g an a r t h r o p o d c h i t i n s yn t h a s [J ].P a t e n t ,2000,W O 9853053-A .[20] G A G O U M E ,K A P S E T A K I M ,T U R B E R G A ,e ta l .S t a g e -s p e c i f i c e x p r e s s i o no f t h e c h i t i ns yn t h a s e D m e C h S A a n dD m e C h S B g e n e s d u r i n g t h e o n s e t o f D r o s o ph i l am e t a -m o r ph o s i s [J ].I n s e c tB i o c h e m i s r y a n d M o l e c u l a rB i o l o g y ,2002,32(2):141-146.[21] A R A K A N E Y ,H O G E N K AM P D G ,Z HU Y C ,e t a l .C h a r a c t e r i z a t i o no f t w oc h i t i ns yn t h a s e g e n e so ft h er e d f l o u r b e e t l e ,T r i b o l i u mc a s t a n e u m a n da l t e r n a t ee x o n u s a g e i n o n e o f t h e g e n e sd u r i n g d e v e l o p m e n t [J ].I n s e c tB i o c h e m i s r ya n d M o l e c u l a rB i o l o g y,2004,34(3):291-304.[22] H O G E N K AM P D G ,A R A K A N E Y ,Z I M O C H L ,e ta l .C h i t i ns y n t h a s e g e n e s i n M a n d u c a s e x t a :c h a r a c t e r i z a t i o n o f a g u t -s p e c i f i c t r a n s c r i p t a n dd i f f e r e n t i a l t i s s u e e x pr e s s i o n o f a l t e r n a t e l y s p l i c e d m R N A sd u r i n g d e v e l o pm e n t [J ].I n s e c t B i o c h e m i s r y a n d M o l e c u l a rB i o l o g y,2005,35(6):529-540.[23] K UM A RNS ,T A N G B ,C H E NXF ,e t a l .M o l e c u l a r c l o n i n g ,e x p r e s s i o n p a t t e r n a n d c o m p a r a t i v e a n a l y s i s o f c h i t i n s yn t h a s e g e n eB i n S p o d o p t e r a e x i g u a [J ].C o m p a r a t i v eB i o c h e m i s -t r y a n dP h y s i o l o g y P a r tB ,2008,149(3):447-453.[24] 孔令斐.家蚕几丁质酶与几丁质合成酶的基因表达与功能研究[D ].苏州:苏州大学,2010.[25] Z HA N G X ,Z H A N GJ ,P A R K Y ,e t a l .I d e n t i f i c a t i o na n dc h a r a c t e r i z a t i o no f t w oc h i t i ns y n t h a s e g e n e s i n A fr i c a n m a l a r i a m o s q u i t o ,A n o p h e l e s ga mb i a e [J ].I n s ec t B i o c h e m i s t r y a nd M o le c u l a rB i o l o g y,2012,42(9):674-682.[26] L IC ,Y A N G WJ ,L I N C ,e t a l .M o l e c u l a r c l o n i n g,c h a r a c -t e r i z a t i o na n d m R N A e x p r e s s i o n o fac h i t i ns yn t h a s e2g e n e f r o m t h e o r i e n t a l f r u i t f l y ,B a c t r o c e r a d o r s a l i s (D i p t e r a :T e ph r i t i d a e )[J ].I n t e r n a t i o n a l J o u r n a l o f M o l e c u l a r S c i e n c e s ,2013,14(8):17055-17072.[27] 刘晓健,崔淼,李大琪,等.飞蝗几丁质合成酶2基因的表达特性㊁功能及调控[J ].中国农业科学,2014,47(7):1330-1340.[28] 余海中,黄克慧,汪婉玲,等.稻纵卷叶螟几丁质合成酶及合成通路相关酶基因的鉴定及表达分析[J ].应用昆虫学报,2015,52(5):1181-1194.[29] 韩国英.棉铃虫H a C H S 和C D A 基因的克隆㊁表达及F e e d i n gR N A i 分析[D ].保定:河北大学,2015.[30] 赵文静,张春林,翟素珍,等.致倦库蚊几丁质合成酶基因C q C H S 1和C q C H S 2的表达与分析[J ].基因组学与应用生物学,2016,35(9):2317-2323.[31] S H I J F ,M ULL ,C H E NX ,e t a l .R N A i n t e r f e r e n c e o f c h i t i ns y n t h a s e g e n e si n h i b i t s c h i t i n b i o s yn t h e s i s a n d a f f e c t s l a r v a l p e r f o r m a n c e i n L e pt i n o t a r s a d e c e m l i n e a t a (S a y )[J ].I n t e r n a t i o n a l J o u r n a l o fB i o l o g i c a l S c i e n c e s ,2016,12(11):1319-1331.941。
STAT3入核的核定位序列研究

# ## 通讯联系人; # (60:$!$?@@A*!*"&,R?W/,0:+X6.J9Y WZ; 56,; -87; 5. # 收稿日期:"$$*?$&?!A,接受日期:"$$*?$@?*$
-//);(!(,) 8 8 8 8 8 8 8 生物化学与生物物理进展8 8 IH>/J K.>=G:8J K.>9G@3J
·&&·
!"# "!# ""# !!# !!# #$! !"! !"%&’,引物 &: (’%$"$ $"# #"$ #"# $!# "!! """ !#$ !!# !#$ "!#%&’,引物 ):(’%""" $$#"!!( !"#* !) "$! ### ##$ ##! $#" #"$ "!%&’+ 引物 , 和引 物 ) 用于扩增野生全长 $%&%& 基因,引物 , 和引物 - 扩增 $%&%& 基因的 , . , -/0 12 得产物 $,引物 和 ) 扩增 $%&%& 基因的 , -34 . - &,/ 12 得产物 5, 将产物 $ 和产物 5 纯化,利用产物 $ 和产物 5 的 部分重叠,加入引物 , 和引物 ) 进行 6"7 对 $%&%& 基因的 , -/0 . , -34 12 之间的碱基缺失+ !" #$ %&’&( 及其缺失突变体的真核表达载体的构建 8 8 将纯化后的 6"7 产物经 ’()"和 !"#*!酶切 过 夜+ 经 凝 胶 分 离 纯 化 后, 克 隆 在 载 体 259:;<=>?2@%AB 的 ’() " 和 !"#* ! 酶 切 位 点 之 间+ 经酶切鉴定和上海生物工程公司测序证实正确后, 亚克隆于表达载体 2C#D6%E, 的 ’()"和 !"#*!酶 切位点 之 间+ 分 别 命 名 为 <!$!&%#D6 ( F?9* @G2; <!$!&%2C#D6%E, ) 和 H<!$!&%#D6 ( *;9;@;* <!$!&%2C#D6%E,)+ !" )$ 转染及激光共聚焦显微镜观察 8 8 分 别 将 - #I 的 空 载 体 2C#D6%E,, <!$!&% #D6,H<!$!&%#D6 和 ( #9 J?2KL;=@MN?O; -/// 分别 稀释于 -(/ #9 的无双抗和无血清的 HPCP 中,将 HE$ 滴入脂质体中轻轻混匀,室温 静 止 -/ N?O+ 转染激光共聚焦专用培养皿培养 -) Q 的 ERS&!& 和 -0&! 细 胞,转 染 -) Q 后 经 ,// #I T J RJ%U 刺 激 -/ N?O 或直接用激光共聚焦显微镜观察+
花器官发育的ABC模型研究进展

! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! 形成萼片状、心皮状或花瓣状的器官代替心皮。 ! 类功能缺失则 "#$ 延伸表达,心皮代替萼片,雄 蕊代替花瓣。 根据在矮牵牛中的研究结果,!%& 模型被扩展
a类功能基因突变后ap2突变体ag就会在第类功能基因的表达不受影响这样在突变体中轮花器官只有c类功能基因作用发育为心皮花器官中的基因表达模式不变仍然发育成雄蕊和pi突变体轮花器官中只有ac类功能基因表达而分别发育成花萼和心皮同时第14轮花器官的发育不受影响分别发育形成花萼和心皮由此形成的花只有花萼和心皮结构
["]
面一轮是心皮,是雌性生殖器官,常常是几个心皮 结合,胚珠和种子在里面形成。尽管发育完全后萼 片、花瓣、雄蕊和心皮的结构和功能可能差异很 大,每个花器官都是由花分生组织的一些凸起(一 小团未分化细胞)开始发育的。在花原基发育过程 中每个细胞都必须获悉在花中的位置,相应地分化
[$] 成适当的细胞类型 。
[L] 的工作证实了这一点:在 6J>63 双突变体 +.K/64 中,第 *、< 轮花器官均转化成叶片(绿色,有托
究花发育的模式植物。两者有不同的优点,前者的 优点是有优良性状的移动因子短,植株体积小 。已经发现了它们已有大量的突
农业生物技术科学 中国农学通报
第 *L 卷
第I期
>&&; 年 *& 月
-11J: O O P41=Q !-246R.S7468Q 401Q !4
Graves病动物模型构建的研究进展

的造模 果,分 建
和病
模
型,质粒造模组予pcDNA3. 1-TSHR 55陆皮内注
射,造模组予肌肉注射Ad-TSHR289,重组腺病毒
造模组中所有小鼠的TRAb和T4水平均升高,
9
中 6 出 现 状 大、 状
织增生等甲亢症状,而质粒造模组仅2只小鼠出
现较 TSHR
反应且总 T4 平 升高,
状腺组织未发生增生性改变。另一研究U4]发现,
第25卷
模型,研究者以携带人TSHR基因的真核表达载 体转染RT4.14HP后的细胞为载体,免疫AKRN (H-2K)小鼠,每2周免疫1次,共计6次,最终约 25%的模型小鼠出现总甲状腺素(T4)水平增高、 TSAb阳性及甲状腺弥漫性肿大,提示造模成功。 此后,有学者[5]采用表达人TSHR和小鼠TSHR 的M12(hM12、mM12)细胞免疫BALB/c小鼠,最 终成功诱导TSAb、T4水平升高并出现典型甲亢 症状;而后应用表达TSHR的重组腺病毒转染树 突状细胞并注射免疫BALB/c小鼠,亦成功诱导 约25%的小鼠出现甲亢[1]0但以上造模方式繁 琐,均需使用其他载体转染细胞后再免疫小鼠,且 成模率较低、稳定性差等,逐渐淡出研究者视线。 1.2 表达TSHR的质粒DNA
表达TSHR的质粒DNA也可有效诱导GD 发生。既往研究⑶发现,将表达TSHR的质粒
DNA(pc DNA 3.1-2SHR)在小鼠双侧股四头肌注 射,结果小鼠体内血清TRAb浓度增高,T4、TSAb 水平均增高,而TBAb水平差异无统计学意义; 甲状腺组织呈现甲状腺滤泡增生、胶质浓缩等 GD的病理表现,提示该质粒可有效建立GD小鼠 模型。同样,XIAN等⑷采用pcDNA3.1质粒载体 构建pcDNA3. 1-T229重组质粒DNA,使90%小鼠 出现明显的甲状腺肿大和甲亢表现,89%实验小 鼠血清T4、TRAb水平显著升高合并甲状腺滤泡肥 大和增生,同时MRI显像能明显观察到小鼠眼眶 肌肉成纤维细胞增大,提示应用质粒DNA可成功 构建GD小鼠模型。此外,pcDNA3. 1-TSHR229 重组质粒也可显著提高小鼠模型的TSHR水 平[7],但重组质粒免疫的小鼠常会出现甲减而非 甲亢⑻。质粒DNA重组方式多样,一部分如 pBccMam-2或pTriCx-2. 1构建的质粒DNA,具有 更高的稳定性,但代价较高⑼。部分质粒DNA在 短期实验中无法有效诱导小鼠发生GD[6] 0上述不 足限制了质粒DNA的应用,仍有待进一步完善。 1.3 表达TSHR的重组腺病毒
晶体结构及与牛血清白蛋白(BSA)的相互作用

Solvothermal Synthesis, Crystal Structure of a 1D Coordination Polymer [Er(CCA)3(H2O)2]n•2nH2O (HCCA=Coumarin-3-Carboxylic Acid) and the Interaction between the Polymer and Bovine Serum
杨树平*,a 韩立军 b 王大奇 c 潘 燕 d 余志群 a 叶海燕 a
(a 淮海工学院化学工程学院 连云港 222005) (b 淮海工学院数理学院 连云港 222005) (c 聊城大学化学工程学院 聊城 252059)
( d 北京大学医学部基础医学院药理系及国家天然仿生药物重点实验室
北京 100191)
血清白蛋白是血浆中含量最丰富的重要载体蛋白 质, 能与许多内源及外源性物质结合, 具有储存和运输 内源性代谢产物和外源性药物小分子及离子等重要生
理功能[27,28], 同时, 血清白蛋白也是一种重要的模型蛋 白用于研究药物分子与蛋白质作用机理[29]. 牛血清白蛋 白(BSA)由于具有实用性、成本低、稳定性和独特的键 合性能, 以及其结构与人血清白蛋白的相似性, 它已作 为模型蛋白被人们广泛的研究与应用[30,31]. 研究药物与 蛋白的相互作用将有助于解释药物的代谢和运输过程, 将有助于解释蛋白质结构和功能之间的关系, 对于了解 生命过程、研究药物作用机制及药物分子设计等, 具有 重要的理论意义[32]. 目前, 血清白蛋白与香豆素小分 子 、 稀 土 离 子 及 稀 土 配 合 物 相 素-3-羧酸稀土配合物与牛血清白蛋白 (BSA)间相互作用的研究未见报道.
Roh 等[20]采用 ErCl3 与香豆素-3-羧酸盐反应获得了 具 有 荧 光 性 能 的 香 豆 素 -3- 羧 酸 铒 配 合 物 [Er(CCA)3(H2O)3], 并利用 FT-IR 和 UV 对其结构进行了 表征. 本课题组利用溶剂热法合成并得到了分子式与文 献 [20] 报 道 不 同 的 香 豆 素 -3- 羧 酸 铒 (III) 配 位 聚 合 物 [Er(CCA)3(H2O)2]n•2nH2O, 并通过元素分析和 IR 及 X 射 线单晶衍射等技术对配合物进行了结构表征及晶体结 构分析. 同时, 用荧光光谱法研究了配合物与 BSA 的相 互作用, 计算了 BSA 与配合物之间的猝灭常数(KSV)、结 合常数(Ka)、结合位点数(n)、BSA 与配合物间的距离(r), 热力学参数焓变(∆H)、熵值的改变(∆S)和自由能改变 (∆G), 据此判断了 BSA 与配合物之间相互作用类型及 强弱, 为今后进一步开发、利用香豆素稀土配合物, 并 为该类配合物在生物无机化学、药物化学、分子生物学 等领域的研究提供了一些具有理论和应用价值的信息.
【北京市自然科学基金】_提取方法_基金支持热词逐年推荐_【万方软件创新助手】_20140730
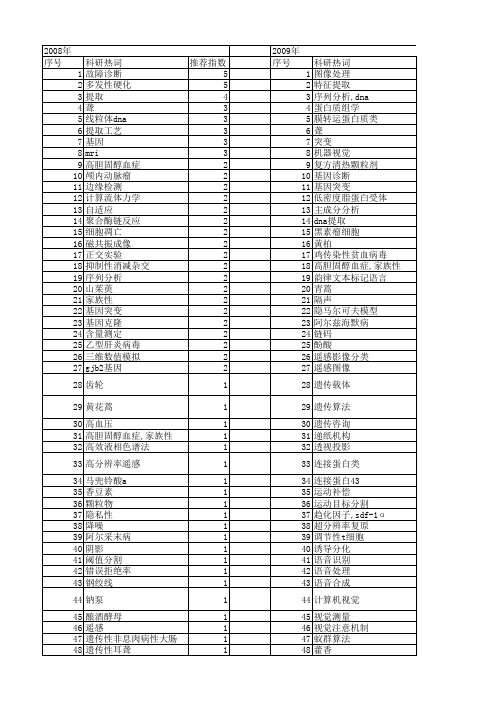
科研热词 推荐指数 故障诊断 5 多发性硬化 5 提取 4 聋 3 线粒体dna 3 提取工艺 3 基因 3 mri 3 高胆固醇血症 2 颅内动脉瘤 2 边缘检测 2 计算流体力学 2 自适应 2 聚合酶链反应 2 细胞凋亡 2 磁共振成像 2 正交实验 2 抑制性消减杂交 2 序列分析 2 山茱萸 2 家族性 2 基因突变 2 基因克隆 2 含量测定 2 乙型肝炎病毒 2 三维数值模拟 2 gjb2基因 2 齿轮 1 黄花蒿 1 高血压 1 高胆固醇血症,家族性 1 高效液相色谱法 1 高分辨率遥感 1 马兜铃酸a 1 香豆素 1 颗粒物 1 隐私性 1 降噪 1 阿尔采末病 1 阴影 1 阈值分割 1 错误拒绝率 1 钢绞线 1 钠泵 1 酿酒酵母 1 遥感 1 遗传性非息肉病性大肠癌 1 遗传性耳聋 1 连锁分析 1 连续小波分析 1 过氧化氢 1 辛弗林 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106
肿瘤坏死因子-α 肾肿瘤 肝炎病毒,乙型 肝炎,乙型,慢性 聚类分析 聚合酶链反应-单链构象多态性 聚合酶链反应 耐药 耐甲氧西林金黄色葡萄球菌 网格图像 绘制 结构光焊缝图像 细菌含量 细胞色素p450 细胞增殖 细胞培养 细胞分离 线粒体 线段分析 线性特征 线性判别分析 突触的可塑性 突触体 稀疏成分分析 禹白附 示例学习 硫氧还原蛋白还原酶 石膏 矩形元 真菌病,蕈样 疲劳 生物学标记 环介导等温扩增技术(lamp) 玄参 猪肉新鲜度 灰度共生矩阵 漂亮 溯源 游程长矩阵 混合像元分解 淋巴瘤,t细胞,皮肤 活动轮廓 注意焦点 水蒸气蒸馏 水平集 气相色谱-质谱 毫米波光纤 模式识别 植物多酚 根系 校准 柿叶 板料零件 束波变换
公示内容应包括如下方面

公示内容(应包括如下方面)一、中华医学科技奖医学科学技术奖推荐项目:1.推荐奖种:中华医学科技奖医学科学技术奖一等奖2.项目名称:新分子锌指蛋白ZBTB20的生理功能及其致病机制3.推荐单位或推荐科学家:中国人民解放军第二军医大学4.推荐意见:该项目组围绕自主发现的新型转录因子ZBTB20,利用国际上首创的ZBTB20全身性和组织特异性敲除小鼠模型,开展了该转录因子在个体发育、学习记忆、痛觉感受、糖脂代谢、垂体发育和免疫应答等过程中的重要生理和病理作用的前瞻性研究,取得了系列独创性研究成果。
在国际核心刊物PNAS、Gastroenterology、Nature Communications等发表的13篇代表性论文,多次被包括Science、Cell、Nature Genetics等国际权威学术刊物引用。
这些成果对于认识Primrose综合征、认知障碍和糖尿病等重大疾病的分子机制具有重要理论创新意义,并为相关疾病的诊治提供了新的理论基础。
此外,该项目培养了一批优秀的硕博士研究生,搭建了与国内外多家知名实验室间深层次合作研究的平台,也使该实验室的承载学科---病理生理学获得了长足的发展。
该项目提供的材料真实可信,特此推荐申报2018年中华医学科技奖医学科学技术奖一等奖5.项目简介:锌指蛋白是一类含有锌指结构域的功能蛋白,在细胞增殖、分化、凋亡和功能等细胞生物学过程中发挥重要的调节作用。
不同种类和成员众多的锌指蛋白构成了庞大的锌指蛋白超家族,约占人类基因的3%。
迄今为止,该超家族的大多数成员的生物学功能尚不清楚。
因此,锌指蛋白被认为是解析生命活动规律的金钥匙。
近十年来,本研究团队围绕自主发现、并在国际上率先报道的新基因锌指蛋白ZBTB20,利用在国际上首次建立的全身性和组织特异性基因敲除小鼠模型,首次系统揭示了该分子在多种组织细胞中的生理功能及其作用机制,取得了如下系列创新性成果,产生了重要国际影响:(1)在代谢方面,ZBTB20是维持机体血糖稳态的关键分子。
会议通知|2022年亚太肝病学会肝纤维化专题会

[J].BestPractResClinGastroenterol,2011,25(2):195-206.DOI:10.1016/j.bpg.2011.02.005.[2]MEDERACKEI,HSUCC,TROEGERJS,etal.Fatetracingrevealshepaticstellatecellsasdominantcontributorstoliverfi brosisindependentofitsaetiology[J].NatCommun,2013,4:2823.DOI:10.1038/ncomms3823.[3]RAYK.Liver:hepaticstellatecellsholdthekeytoliverfibrosis[J].NatRevGastroenterolHepatol,2014,11(2):74.DOI:10.1038/nrgastro.2013.244.[4]PIERA-VELAZQUEZS,MENDOZAFA,JIMENEZSA.Endo thelialtomesenchymaltransition(EndoMT)inthepathogene sisofhumanfibroticdiseases[J].JClinMed,2016,5(4):45.DOI:10.3390/jcm5040045.[5]SUNX,NKENNORB,MASTIKHINAO,etal.Endothelium-mediatedcontributionstofibrosis[J].SeminCellDevBiol,2020,101:78-86.DOI:10.1016/j.semcdb.2019.10.015.[6]RUANB,DUANJL,XUH,etal.Capillarizedliversinusoidalendothelialcellsundergopartialendothelial-mesenchymaltransitiontoactivelydepositsinusoidalECMinliverfibrosis[J].FrontCellDevBiol,2021,9:671081.DOI:10.3389/fcell.2021.671081.[7]THOMSONJG,RUCKEREB3rd,PIEDRAHITAJA.MutationalanalysisofloxPsitesforefficientCre-mediatedinsertionintogenomicDNA[J].Genesis,2003,36(3):162-167.DOI:10.1002/gene.10211.[8]KOSCH.Cre/loxPsystemforgeneratingtissue-specificknockoutmousemodels[J].NutrRev,2004,62(6Pt1):243-246.DOI:10.1301/nr2004.jun243-246.[9]JIANGZJ,YUEZS,YANGYC,etal.ImprovementmethodofisolationofmouseliverSinusoidalendothelialcell[J].ProgModBiomed,2018,18(6):1034-1039.DOI:10.13241/j.cnki.pmb.2018.06.007.蒋子剑,岳振生,杨毅聪,等.小鼠肝血窦内皮细胞分离与鉴定新方法[J].现代生物医学进展,2018,18(6):1034-1039.DOI:10.13241/j.cnki.pmb.2018.06.007.[10]WANGS,FRIEDMANSL.Hepaticfibrosis:Aconvergentre sponsetoliverinjurythatisreversible[J].JHepatol,2020,73(1):210-211.DOI:10.1016/j.jhep.2020.03.011.[11]TERKELSENMK,BENDIXENSM,HANSEND,etal.Tran scriptionaldynamicsofhepaticsinusoid-associatedcellsafterliverinjury[J].Hepatology,2020,72(6):2119-2133.DOI:10.1002/hep.31215.[12]TRAUTWEINC,FRIEDMANSL,SCHUPPAND,etal.Hepaticfibrosis:Concepttotreatment[J].JHepatol,2015,62(1Suppl):S15-S24.DOI:10.1016/j.jhep.2015.02.039.[13]ZEISBERGEM,TARNAVSKIO,ZEISBERGM,etal.Endothe lial-to-mesenchymaltransitioncontributestocardiacfibrosis[J].NatMed,2007,13(8):952-961.DOI:10.1038/nm1613.[14]ZEISBERGEM,POTENTASE,SUGIMOTOH,etal.Fibro blastsinkidneyfibrosisemergeviaendothelial-to-mesenchymaltransition[J].JAmSocNephrol,2008,19(12):2282-2287.DOI:10.1681/ASN.2008050513.[15]HASHIMOTON,PHANSH,IMAIZUMIK,etal.Endothelial-mesenchymaltransitioninbleomycin-inducedpulmonaryfibrosis[J].AmJRespirCellMolBiol,2010,43(2):161-172.DOI:10.1165/rcmb.2009-0031OC.引证本文:XUH,RUANB,LIZW,etal.Establishmentofamousemodelofvascularendothelial-mesenchymaltransdifferentiationgenetictracinganditsroleinliverfibrosisstudies[J].JClinHepatol,2022,38(4):832-836.许皓,阮柏,历志文,等.血管内皮间质样转分化遗传示踪小鼠模型的构建及其在肝纤维化研究中的应用[J].临床肝胆病杂志,2022,38(4):832-836.(本文编辑:刘晓红)·国内外会议动态·会议通知|2022年亚太肝病学会肝纤维化专题会APASLSTC2022:Liverfibrosisintheeraoftranslationalmedicine 会议时间:2022年6月3-5日会议地点:中国北京会议网站:http://www.apaslstc.com会议主席:尤红(首都医科大学附属北京友谊医院),徐小元(北京大学第一医院),贾继东(首都医科大学附属北京友谊医院)本次会议主题为“肝纤维化转化研究”,将邀请相关领域国际学术权威和专家从肝纤维化发病机制、最新诊疗技术以及慢性肝病病因治疗、抗纤维化新药临床试验进展等方面开展探讨,分享国内外学者关于肝纤维化和肝硬化的新药研发以及最新诊治管理策略的专业理念和创新技术。
考博

生命科学学院北京市玉泉路19号(甲)生命科学学院071009 细胞生物学01 环境与健康、糖尿病丁文军①1001英语一②2464分子生物学③3477细胞生物学02 骨细胞生物学杨建虹同上03 自由基生物学/分子医学陆忠兵①1001英语一②2573自由基生物学③3477细胞生物学071010 生物化学与分子生物学01 医用酶钟良玮①1001英语一②2464分子生物学③3687生物化学02 植物抗逆的分子生物学柴团耀①1001英语一②2493植物生理学③3493植物分子生物学03 环境与健康、糖尿病丁文军①1001英语一②2464分子生物学③3477细胞生物学04 植物次生代谢的分子调控王红①1001英语一②2493植物生理学③3493植物分子生物学05 生物分析丁永胜①1001英语一②2464分子生物学③3477细胞生物学06 哺乳动物的遗传与发育袁莉同上07 环境与健康糖尿病丁文军①1001英语一②2181分子生物学(A)③3250细胞生物学(A)为中丹学院代招,挂靠生命科学学院招生生物物理研究所北京朝阳区大屯路15号071009 细胞生物学01 氧化应激与蛋白质巯基修饰,衰老相关疾病与调控;一氧化氮生物功能陈畅①1001英语一②2753生物化学与分子生物学③3896细胞生物学只招硕博连读生02 病毒复制的分子机理及病毒与宿主的相互作用邓红雨同上只招硕博连读生03 天然免疫信号调控免疫应答的分子和细胞机制侯百东①1001英语一②2702分子免疫学A③3896细胞生物学04 慢性炎症恶性转化的分子机制;抗肿瘤免疫学机制和免疫治疗新策略王盛典同上只招硕博连读生05 肿瘤免疫学:1. HBV和肿瘤微环境;2. 肝癌转移机制;3. 血管稳态调节。
杨鹏远同上06 树突状细胞发育和功能的分子机制张立国同上07 淋巴器官的发育、稳态、重塑和功能朱明昭同上只招硕博连读生08 基因组稳定性和蛋白质人工进化杭海英①1001英语一②2753生物化学与分子生物学③3896细胞生物学09 内质网的形态和功能胡俊杰同上只招硕博连读生10 细胞多信号体系紊乱与疾病发生和干预机制;胚胎干细胞生物学姬广聚同上11 细胞生长与代谢、增殖与细胞分化的机理焦仁杰同上12 生物大分子药物输送系统及免疫应答机制梁伟同上只招硕博连读生13 基于干细胞的治疗和药物筛选体系;人类衰老和长寿的分子遗传学基础刘光慧同上只招硕博连读生14 脂质储存与代谢,脂滴生物学;代谢疾病机理;微生物生物能源刘平生同上只招硕博连读生15 生物大分子间相互作用的分子机马跃同上只招硕制和动力学性质博连读生16 细胞极性建立的分子机制、细胞运动机理研究、精子活化的调控机理苗龙同上17 炎症因子在肿瘤免疫中的作用及机制研究秦志海①1001英语一②2702分子免疫学A③3896细胞生物学18 人多能干细胞曲静①1001英语一②2753生物化学与分子生物学③3896细胞生物学19 多细胞生物自噬作用的机理和调控机制;神经退行性疾病的发生机制张宏同上只招硕博连读生20 以秀丽线虫为模式研究凋亡细胞的清除机制及溶酶体动态变化在发育和胁迫应答中的作用王晓晨同上21 感染免疫唐宏①1001英语一②2702分子免疫学A③3896细胞生物学22 肿瘤免疫学刘阳同上23 发现肿瘤新靶点,发展肿瘤靶向抗体药物,发展肿瘤诊断新方法阎锡蕴①1001英语一②2753生物化学与分子生物学③3896细胞生物学只招硕博连读生071010 生物化学与分子生物学01 黏膜免疫与肠炎刘志华①1001英语一②2753生物化学与分子生物学③3896细胞生物学02 结核病系统生物学研究毕利军同上03 细胞极性调控及分子马达运输相关蛋白质复合物的结构与功能研究冯巍同上只招硕博连读生04 细胞重编程、肿瘤干细胞与肿瘤免疫治疗范祖森同上只招硕博连读生05 感染免疫、肿瘤免疫的机理及重大病毒性疾病和肿瘤免疫治疗的研究傅阳心①1001英语一②2702分子免疫学A③3896细胞生物学06 天然免疫抗HIV机理,重点是抗病毒因子的鉴定和分子机理分析高光侠同上只招硕博连读生07 蛋白质晶体学手段研究疾病相关蛋白质复合体和膜蛋白结构江涛①1001英语一②2753生物化学与分子生物学③3896细胞生物学只招硕博连读生08 蛋白质的折叠、修饰与质量控制柯莎同上只招硕博连读生09 染色质结构与表观遗传调控李国红同上只招硕博连读生10 重大疾病相关的蛋白质结构生物学研究刘迎芳同上只招硕博连读生11 蛋白质翻译过程与肿瘤发生秦燕同上12 线粒体动态的分子机制; 生物超大分子复合体结构功能;生物成像方法学研究孙飞同上13 肿瘤发生和转移的机制研究屠亚平同上14 病原结构生物学:研究疾病发生与防御的蛋白质结构基础与分子机理王大成同上只招硕博连读生15 合成生物学;酶的进化与设计;活细胞中生物大分子的标记王江云同上16 RNA干扰相关蛋白与RNA复合物的结构与功能研究王艳丽同上只招硕博连读生17 内质网蛋白质氧化折叠系统及其调控与二型糖尿病等疾病以及衰老间的联系王志珍同上只招硕博连读生18 基因转录的表观遗传调控和RNA 转录后加工许瑞明同上只招硕博连读生19 线粒体-溶酶体动态变化及稳态杨福愉同上只招硕维持的分子机理博连读生20 结构生物学叶克穷同上21 重要生命活动相关膜蛋白的结构与功能研究张凯同上只招硕博连读生22 分析病原微生物学张先恩同上23 膜蛋白的结构与功能赵永芳同上24 组蛋白变体的结构和功能及其关联的表观遗传调控机制周政同上只招硕博连读生25 表观遗传学朱冰同上26 染色质高级结构与表观遗传调控;病毒感染与复制机制朱平同上只招硕博连读生27 应用生物质谱等高级分析技术研究糖及蛋白质组学的基础和临床问题李岩同上28 结构生物学、蛋白质核酸相互作用龚为民同上29 天然免疫信号通路相关蛋白的结构功能研究,基于结构的药物设计刘志杰同上只招硕博连读生30 重要传染病以及与人类疾病相关关键蛋白质的结构与功能的研究饶子和同上只招硕博连读生31 RNA结合蛋白和非编码RNA在细胞重编程、干细胞分化及癌症发生中的功能与机制薛愿超同上32 病原菌感染和宿主免疫防御机制邵峰同上只招硕博连读生微生物研究所北京市朝阳区北辰西路1号院中科院微生物研究所071005 微生物学5901 工业酵母菌的进化基因组学白逢彦①1001英语一②2434生物化学A③3479分子生物学02 植物病原真菌学蔡磊①1001英语一②2466生物化学B③3493真菌学03 甲烷古菌冷适应机理东秀珠①1001英语一②2434生物化学A③3479分子生物学04 微流控生化分析芯片及微生物学应用杜文斌同上05 单细胞单分子生物化学傅钰同上06 菌根菌的碳氮循环调控机制郭良栋①1001英语一②2434生物化学A③3493真菌学07 医学真菌功能基因组学黄广华①1001英语一②2466生物化学B③3479分子生物学08 放线菌系统学与生态功能黄英①1001英语一②2434生物化学A③3479分子生物学09 真菌遗传李少杰同上10 合成生物学与代谢工程李寅同上11 基于珍稀药用真菌生物活性物质的新药与新功能食品研发刘宏伟①1001英语一②2466生物化学B③3479分子生物学12 芳烃类化合物对微生物降解途径和降解机理的研究刘双江①1001英语一②2434生物化学A③3479分子生物学13 病毒与宿主相互作用刘文军同上14 真菌群体基因组学研究刘杏忠①1001英语一②2466生物化学B③3493真菌学15 环境微生物学刘志培①1001英语一②2434生物化学A③3479分子生物学16 多糖合成蛋白的结构与功能马旅雁同上17 极端微生物适应机制与合成生物学马延和同上18 重要病毒分子生物学和免疫学研究田波同上19 通过表型与基因型相结合的综合分析方法进行地衣型真菌的系统分类学研究魏江春①1001英语一②2495地衣生物学③3479分子生物学20 食药用菌深层次开发利用姚一建①1001英语一②2434生物化学A③3493真菌学21 海洋微生物天然产物的高通量药物筛选张立新①1001英语一②2434生物化学A③3505天然产物化学22 菌生真菌与宿主真菌之间相互作用庄文颖①1001英语一②2466生物化学B③3479分子生物学071007 遗传学01 微生物次级代谢产物的生物合成与基因组挖掘陈义华①1001英语一②2434生物化学A③3479分子生物学02 植物基因功能及其调控方荣祥同上03 植物RNA沉默和植物-病原的相互作用郭惠珊同上04 植物-微生物互作的细胞及分子机理孔照胜同上05 丝状真菌次级代谢产物的生物合成与调控刘钢同上06 植物抗病蛋白组学刘俊①1001英语一②2466生物化学B③3479分子生物学07 细菌识别寄主的分子机制钱韦①1001英语一②2434生物化学A③3479分子生物学08 基因组编辑技术及植物抗病免疫蛋白磷酸化网络邱金龙同上09 微生物次级代谢产物的生物合成及其分子调控谭华荣同上10 微生物的代谢工程与系统生物学温廷益同上11 古菌CRISPR-Cas系统分子机制向华同上12 乳酸菌分子遗传学钟瑾同上13 基因组学朱宝利①1001英语一②2520免疫学③3479分子生物学071010 生物化学与分子生物学01 病毒感染与肿瘤发生机理陈吉龙①1001英语一②2434生物化学A③3479分子生物学02 酶分子生物学董志扬同上03 NK细胞在流感病毒感染后的免疫应答反应特征及控制感染的作用机制研究方敏①1001英语一②2520免疫学③3479分子生物学04 细胞免疫识别的结构基础高福同上05 极端环境微生物遗传机制及其环境适应性黄力①1001英语一②2434生物化学A③3479分子生物学06 真菌来源化合物抗肿瘤增殖机制姜学军同上07 蛋白质翻译后修饰与微生物生长发育机制金城同上08 合成生物学及生物调控系统的设计理论和组装方法娄春波同上09 乙肝病毒与肿瘤免疫孟颂东同上10 高价值化合物的生物合成及酶工程唐双焱同上11 全细胞催化、代谢工程、合成生物学陶勇同上12 Ⅰ类抗原提成分子β2m在特异性抗体增高中的分子机制和作为免疫佐剂的应用前景王北难①1001英语一②2520免疫学③3479分子生物学13 疱疹等重要病毒与宿主相互作用及致病机制研究严景华同上14 微生物代谢调控杨克迁①1001英语一②2434生物化学A③3479分子生物学15 病毒感染及细胞周期调控的叶昕同上分子机理16 真菌次级代谢产物基因调控与合成生物学尹文兵①1001英语一②2466生物化学B③3479分子生物学17 细胞免疫在抗感染和自身免疫病中机制的研究张福萍①1001英语一②2520免疫学③3479分子生物学18 调节性T细胞的免疫负调控周旭宇同上遗传与发育生物学研究所北京市朝阳区北辰西路1号院2号071007 遗传学01 植物分子遗传学唐定中①1001英语一②2026生物化学③3023遗传学谢旗同上胡赞民同上凌宏清同上童依平同上张爱民同上左建儒同上张劲松同上傅向东同上程祝宽同上李家洋同上周奕华同上李霞同上周俭民同上沈前华同上限招直博生韩方普同上刘翠敏 同上 王永红 同上 02 植物功能基因组学李传友同上 薛勇彪同上限招直博生曹晓风 同上 储成才 同上 王国栋 同上 田志喜 同上 03 植物分子遗传育种 王道文 同上 张相岐 同上 朱保葛 同上 翟文学同上 李俊明 同上 姚善国 同上 高彩霞 同上 陈化榜 同上 林少扬 同上 04 人类与动物分子遗传学马润林同上 钱文峰同上限招硕转博071008 发育生物学01 植物发育生物学杨维才 ①1001英语一②2026生物化学③3023遗传学陈凡 同上 李云海 同上 焦雨铃同上02 人类与动物发育生物学孙方臻同上王朝晖同上黄勋同上鲍时来同上张建同上限招直博生John R.Speakman同上071009 细胞生物学01 人类与动物细胞生物学王秀杰①1001英语一②2026生物化学③3023遗传学代屠强招生杨崇林同上李巍同上限招直博生孟文翔同上税光厚同上北京基因组研究所北京市朝阳区北辰西路1号院104号,北京基因组研究所071007 遗传学2501 遗传与进化基因组学吴仲义①1001英语一②2045基因组学③3040生物信息学或3075遗传学02 复杂性状或疾病的进化基因组学吕雪梅同上03 疾病和演化相关基因组研究曾长青同上该研究方向仅招收硕转博研究生04 肿瘤遗传学赵永良同上该研究方向仅转博研究生05 癌症与干细胞的表观遗传调控孙英丽同上该研究方向仅招收硕转博研究生071010 生物化学与分子生物学01 重大疾病相关非编码RNA研究陈非①1001英语一②2092生物化学与分子生物学③3075遗传学或3133细胞生物学02 基因组不稳定性与疾病相关性研究郭彩霞同上该研究方向仅招收硕转博研究生03 RNA甲基化表观调控杨运桂同上该研究方向仅招收硕转博研究生04 癌症中非编码RNA的作用研究米双利同上该研究方向仅招收硕转博研究生05 蛋白质组学吴琳同上该研究方向仅招收硕转博研究生06 蛋白质组学吴琳①1001英语一②2265生物化学与分子生物学③3236细胞生物学为中丹学院代招,挂靠北京基因组研究所招斯奇导师合招07 蛋白质组学及疾病标志物吴琳同上为中丹学院代招,挂靠北京基因组研究所招生。
巴西橡胶树14—3—3蛋白基因的克隆及表达分析

主要 以 同源或 异 源二 聚体 形式 存 在 .可 以同 时与 两个 靶 蛋 白或者 与 一 个 靶蛋 白 的两 个 结构 域 相 互 作 用 .
通过 与靶 蛋 白上 一小 段 共有 序列 的磷酸 化 丝氨 酸/ 苏氨 酸残 基结 合 来发 挥 其调 控 功能【 1 — — 4 4 3 3蛋 白在 调 J 控植 物 基础代 谢 中处于 中 心地位 .通 过与 硝 酸还 原酶 、蔗糖磷 酸 合 成酶 、叶绿 体前 体 蛋 白等 的相 互作 用 来调 控糖 、氨基 酸 、核酸和 蛋 白质等物 质 的生物合 成 ;1 — — 4 3 3蛋 白通 过 直接蛋 白质一 白质 相互作 用 对植 蛋 物代谢关 键酶 、质膜 H 一 T 十A P酶 等发挥 广泛调 节作用[o 5] -。相关研 究结果 表 明 。1 — — 1 4 3 3蛋 白通过 与转 录因子
l— — 4 3 3蛋 白家 族虽 在 序列 上 高度保 守 ,但 不 同 物种 、同一 物 种 中不 同 1 — — 4 3 3蛋 白成员 的 N端 和 C
端均 存在 差异 。由于 N端 和 C端 的差 异 .1— — 4 3 3蛋 白家 族 中的不 同成 员行 使不 同的功 能I 1 — — 4 3 3蛋 自 3 】 。
群结腐霉细胞壁降解酶的活性检测及基因表达分析

櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄[18]郑 ,王昊宁,刘靖文,等.牡丹花发育相关基因PsPI的克隆与表达分析[J].分子植物育种,2021,19(7):2185-2192.[19]EstruchJJ,KadwellS,MerlinE,etal.Cloningandcharacterizationofamaizepollen-specificcalcium-dependentcalmodulin-independentproteinkinase[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1994,91(19):8837-8841.[20]YoonGM,DowdPE,GilroyS,etal.Calcium-dependentproteinkinaseisoformsinpetuniahavedistinctfunctionsinpollentubegrowth,includingregulatingpolarity[J].ThePlantCell,2006,18(4):867-878.[21]麻 浩,王 爽,周亚丽.植物中钙依赖蛋白激酶的研究进展[J].南京农业大学学报,2017,40(4):565-572.[22]丁艳芬.玉米ZmCPK11在ABA诱导的抗氧化防护中的功能分析[D].南京:南京农业大学,2012.[23]FedosejevsET,GerdisSA,YingS,etal.Thecalcium-dependentproteinkinaseRcCDPK2phosphorylatessucrosesynthaseatSer11indevelopingcastoroilseeds[J].TheBiochemicalJournal,2016,473(20):3667-3682.[24]王娇娇,韩胜芳,李小娟,等.钙依赖蛋白激酶(CDPKs)介导植物信号转导的分子基础[J].草业学报,2009,18(3):241-250. 陈一帆,薛太强,陈思桥,等.群结腐霉细胞壁降解酶的活性检测及基因表达分析[J].江苏农业科学,2023,51(4):46-51.doi:10.15889/j.issn.1002-1302.2023.04.007群结腐霉细胞壁降解酶的活性检测及基因表达分析陈一帆1,2,薛太强2,陈思桥2,张金凤2,周冬梅2,魏利辉1,2(1.江苏大学环境与安全工程学院,江苏镇江212013;2.江苏省农业科学院植物保护研究所,江苏南京210014) 摘要:群结腐霉(Pythiummyriotylum)是一种死体营养型病原菌,可侵染生姜引起茎基腐病,造成生姜产量损失,严重危害该产业发展。
生物学通报(月刊)2006年第41卷总目次

生物学通报(月刊)2006年第41卷总目次2006年第41卷第12期生物学通报生约也搬(月刊)SHENGWUXUETONGBAO总目次题目作者期页科学家论坛节肢动物的分类和演化………………………宋大祥0301马的演化遗传多样性研究……宁眺张亚平080l鼠瞧…………………………………方喜业王光明0901秉志院士,lk绩永存——纪念束志先生诞辰120周年…………………………………………………………………翟启慧l001综述与进展野牛鸟类对禽流感爆发与传播的影响…霄富民赵德龙0101超氧化物歧化酶研究进展……………………于平0l04蛋白质组学及其在植物科学研究中的应用……………………………………………………范海延陈捷曲波等0106定量蛋白质组学研究的技术体系...叶雯洪华珠0l09有毒污染物对生物体的毒性效应与自由基反应……………………………………………………………安鑫龙间启星020l蝇源抗菌肽的研究进展…土婷婷金小宝朱家勇0304分于伴俏结构与功能的研究进展………………………………………………………………潘云娣杨文鸽侯温甫等0307细胞静息膜电位的产乍和维持………………左明雪040l含硫氨基酸的抗氧化作用…霍湘王安利杨建梅0403绿色生命和科学发展舰……苗以农朱长甫石连旋等05叭洞穴型蝙蜗的栖息环境选择,生态作及保护……………………………………………………………张成菊吴毅0504植物细胞的伞能性……………………………肖尊安060l水稻基因图…………………………刘髓王勇0604昆虫蛹滞育的神经内分泌调控...…王忠婵王方海0606饥饿对鱼类的影响………钱周兴徐永清方一峰0609青藏铁路格唐段植被特征及其保护与恢复对策………………………………………………陈桂琛』孟延山卢学峰等0701棉花转基技术和转基冈棉花……………………………………………………………段春燕侯小改张亚冰等07动物适应的生理反应…………………………柳劲松07抗生素菌种选育的研究进展……...周梅先蓖钧锋08海南岛热带花卉资源研究与利用……………欧阳欢09中闷亚洲象现状及研究进展…………………张立1l性染色体与精于功能研究进腱………………………………………………………………张红国刘睿智段晓刚l1前列腺癌生物标记与早期诊断……………………………………………………………吕赫哲黄珊郭振光等12植物遗传资源的评价,保持和利用……………t林生12植物蛋白质组学的研究进展………万华方粱颖12基础知识动物的试一错学习行为…………………………尚玉昌01苔藓植物提取液的经济价值…………………车未艾01人类最忠实的伴侣动物——家犬……………居尼和01钠通道及其相关疾病综合征………徐妍肖玉成01系统生物学——后基因组时代的生物学……王JI101内质网和高尔基体的功能浅析………………金春钰0l双生子印性相似诊断法…十建平吴中华郑坚01低能离束介导转基囚技术尧俊英李景原李清禄01线粒体的研究……………刘红梅蒋继志廖祥儒02线粒体融合…………………………郭晓强郭振清02流感病毒与人流感和禽流感…………………徐志毅02动物的顿悟学习行为…………………………尚玉吕02黑猩猩与人类基冈相似度达96%以上………奇云02植酸的生物学特性与脚用……………………陈红霞02分子标记技术的发展及应川闷华超高岚李桂兰02发光鱼类……………………………张世义伍玉明O2鱼类的嗅觉…………………………李信书郑伟02寄牛植物慨述…………盛晋华张雄杰刘宏义等03秦岭金丝猴的觅食生物学王晓卫吕九全郭松涛等03动物的求偶喂食行为…………………………尚玉昌0320O6年第41卷硒的生物活性与相关疾病………………………黄峙0317植物次生物的代谢途径……季志平苏印泉张存莉o319苏尼特左旗发菜资源的利『{】及保护…………………………………………………………………叶长鹏王长荣高坤I¨0321生物多倍体及其存农业上的应用………………杨万立0323转基因食品的安全性探讨……………丁存宝刘海燕o325蚂蚁的社会生活…………………………………尚玉吕0405褐藻多糖…………………………………………张海容0407我国红树林的分布现状,保护及生态价值…………………………………………………………张忠华胡刚梁士楚0409还水鸟一片洁净的水域………………周可新霍培书04l】我国的桃花水母……………符方尧王丹丽徐善良o413寄生虫的起源与进化…………………李亮张路平04l5生物活性肽……………………………黄德娟黄德超04l7细胞静息膜电位的测量…………………………左明雪0507几种高通量定向筛选技术及其比较…………………………………………………………………王绘砖陈喜文陈德富O5O8线粒体微卫星DNA不稳定性与肿瘤关系的研究进展……………………………………………郭晓华昌恩梓冯典兴05l0氮,磷对水华发生的影响………………………陈琼05l2寄生虫与宿主的相互作用李亮李晓娜徐真等05l5农杆菌Ti质粒的改造及其衍生的质粒载体…………………………………………………………………李淑萍康浩05l9动作电位形成的机制……………………………左明雪06l2蚂蚁的化学通讯…………………………………尚玉昌0614杜鹃花赏析…………………朱春艳包志毅唐宇力06l6湖南2个民族5种运动类型的研究……………………………………………………………皮建辉雷鸣枝吴亿中等06l8丝裂原活化蛋白激酶信号通路相关研究………………………………………吴小萍陈晓萍腔肠动物的毒索……...李兆英吕倩免疫组织化学中的抗原修复技术……………………………………………魏仲梅俞诗源核酸探针的原理及应用…张明琴窦远药朋植物的多倍体育种…朱智青梁运章胚胎干细胞技术…………李松苗增民斯舁亮辛蕾严峰等张建华等李玉峰等高佩安等蜱螨慨述…………………………………………陈军蚂蚁的视觉,听觉和触觉通讯…………………尚昌人参果简介………………………………………范甲朋神经系统与内分泌系统的相互影响与协同作用………?- ……………………………张凌燕温庆城张志文小议生物扩散…………………………肖治术张知彬试论人类头目化石在研究人类起源和演化中的作用……-. ……………………………………………………周必文人类的性别与性别畸形……张淑玲于海涛张春晶加压素和催产素在rrl枢神经系统巾的作用……………………………………………何凤琴邰发道张育辉等哺乳动物的冬眠及其影响因素……………………………一………………………………刁颖祁冲袁丽丽淡水鱼类精子的冷冻与保存………………………………? ……………………………廖馨严维辉唐建清等动物行为的研究方法……………………………尚玉昌一种绿色燃料——生物柴油…………邓文辉李新生从酸雨到酸沉降——关于对酸雨的认识………………………………张昕孙腊梅周淑梅白蚁的社会组织和社会行为……………………尚玉昌高尔基体的结构与功能研究简介………………鲍时来细胞膜的双层磷脂结构与功能…………………徐国恒半胱天冬酶在艾滋病发病机制中的作用………詹建立纳米生物效应及安全性研究……………………王川I鲨鱼……………………………………张世义伍玉明同位素示踪法与生命科学发展………郭申生任衍钢06200621070707070707070911l315l72I2307240727080508070810081408160818O82O08O909o909O9090922071O1115172O22~∞∞叭~∞~m¨"∞¨"如∞总目次Ⅱ生物学通报2oo6年第41卷第12期腐生真菌的消长规律………………于淑玲赵静简述中心体的结构与功能……………………桑建利香蕉的生物学特性及其组织培养技术..…….吴坤林动物的行为节律………………………………尚玉昌从基因组3非编码区看黄病毒科各成员的进化关系……. ………………………………………张舒蕾肖明麝的通讯行为………姜海瑞王清薛文杰等我国的生物产业状况与前景沈光涛常灏黄耀江牛膝茎节膨大现象与原因的探究……………李新华基因工程在环境保护中的应用……陈怀侠赵永志高尔基体堆形成的分子机制………张照亮鲍时来北京颐和园昆明湖的鱼类物种多样性…………………….. …………………………邢迎春赵亚辉王玉风等线虫与果蝇先天免疫系统的比较…….……..卜兴江持久性有机污染物的植物修复………………李坤陶奇怪的血型——孟买型和非孟买型….……..季雄严重危害人类的8种病毒性传染病…………陈瑞我国常用的灭螺方法…毕研云图立红李枢强等海藻肥料的应用前景…………………………隋战鹰09241003l00510081010l013l0151017102ll104l10611l1l1121114l1151117l119植物维生素E代谢工程研究……………………………………¨一…………………………钱文成陶苏丹陈德富等1213昆虫温度感受器的结构与功能………………时磊1216心绞痛的痛觉产生机制……………王小蕾徐国恒1218二三峡库区的渔业资源及保护………幸奠权李建勇122O内耳淋巴液的来源,性质和成分……………王艳1223生物能源的研究和利用现状…………………戎茜12245种严重影响水质的化学物质…………………………………………………………………谢汇魏翠英魏凌燕1226糖生物学的产生和发展………………………傅雷1227教学参考资料《本草纲目》一些植物种名中蕴涵的动植物之间的相互关系………………………………………李新华尹晓明0125关于人类白细胞抗原HLA……………………张云佐0223物质的内吞,外排与质膜,内膜系统的关系…杨志明0224恶性杂草——豚草……………………………张葵0225关于植物的求救信号…………………………周新生O226利用SSR分子标记解析遗传学三大规律…邵元健o327电磁辐射对人类健康的影响……刘志华时丽冉o330关于"世代交替"的一些疑惑…………………………………………………………………俞佩芳张文华孙凯0331再议细胞周期中染色体,DNA的数量变化规律………………………………………………杨静利张莉姚世贤0419单基因遗传病的种类判断和概率计算……池君o420遗传学中基因频率和基因型频率的有关问题白绍静o421应激性与反射关系的辨析………………李保兴05目次Ⅱ从自由组合与连锁交换看基因排列的"效率原则"……………………………………………………赵阳顾蔚0517有机食品,绿色食品和无公害食品张万海原维芳0520谈谈植物的性别——由一道选择题引起的思考…………………………………………………………………王玲敏0726有关种群增长的教学思考……………………刘四清0729百合大孢子母细胞的减数分裂与雌配子体形成——植物学显微摄影彩色照片系列之七…周仪袁勤0824对钙离子几种生理学效应的归纳与探讨…杨明鄄0826多倍体植株为什么高大且营养丰富………周飞彪0828对一个概率计算问题的分析…………………李其安0843食物中核酸的消化与吸收……………………苏成清0926奇特的海马……………………………………李晓梅0927母性影响和细胞质遗传………………………游隆信o928性反转………………………………杜继煜董建新1018小议"细胞分化"………………………………颜景军1019DNA杂交与DNA指纹技术…………………郎红梅l121关于遗传密码于的3个问题…………………张云佐1122对血糖激素调节的商榷………………………张辉l123关于胰岛素和胰高血糖素参与血糖调节作用……………………………………..………………….肖洋徐国恒l124合理使用维生素C……………………………黄富生ll25记忆细胞与二次免疫…………………………常永红l126适用于本科实验教学的衣藻核遗传转化方法研究…………………………………耿云红奇Et迈励图鲁迪红等ll27浅析生态系统中的三大功能类群…………赵玉彩ll29植物学海藻野外实习的几点做法…王元军张福海l229课程?教材改革探究性学习实施中的几个问题………………朱正威0l27突出实践特色培养创新意识——《生物技术实践》(中图版)教材简介…………………石健冯永康0228高中生物学课程的目标是提高公民科学素养——苏教版高中《生物》新教材简介……………………汪忠0422高中生物学新课程标准教学研讨"细胞的分子组成"的教学构思………………………………………郑春和0829普通高中生物学课程标准教学研讨——"细胞的结构"的教学提示……………………………………朱正威0929高中生物学新课程标准教学研讨——"细胞的代谢"教学构思(1)……………………………………陈月艳1023高中生物学课程标准教学研讨——"细胞的代谢"教学构思(2)……………………………………陈月艳l131高中生物学新课程标准教学研讨一"细胞的增殖,分化,衰老和凋亡"的教学构思……………吴成军1230课改实验交流新课标理念指导下的"教学相长"典型案例——汗液和尿液是一样的吗?………………………吴光举赵广宇O129生物学教师如何"活用"新教材…………………席广涛o231关于"观察DNA和RNA在细胞中的分布"实验的改进建议……………………………………………………何彬0249让生物学课堂成为学生自主学习的新天地——"植物茎的输导功能"一节教学的点评……李其柱刘迎侠o333对新课程实施中问题的反思……………………文学艺o336 "采集和测算空气中的尘埃粒子"实验中的几处改进…………………………………………………………………田美容o337"鸟类"一节教学…………………………………赵广宇o425开展"探索生长素类似物促进插条生根的最适浓度"探究活动的方法和策略……………………………沈雁0523设计制作生态缸观察"生态系统的稳定性"…………………………………………………卢菁菁沈初见陈莺莺等O527关于"植物的营养生殖方式"一节课的几点思考戴春0623探索生长索类似物促进插条生根的最适浓度…卓铭阳0625探究生物体维持pH稳定的机制……陈龙珠沈初见0731"探究蚂蚁的通讯"实验改进……………………田春华O832建立血糖调节的模型…………………俞小燕沈初见0833"测量不同植被环境的空气温度和湿度"活动改进与实施建议……………………………………………尚德兰0835样方法调查草地中某种双子叶植物的种群密度……………………………………………………………徐建林沈初见0932"模拟自然选择"课堂活动的改进………………王予红0935土壤中小动物类群丰富度的研究……丁泾芳沈初见1027从探究"蚂蚁的通讯"谈对学生创新能力的培养……………………………………………………………田春华温淑香1029土壤微生物分解作用的研究…………陈栩沈初见l136课堂教学设计"基因的分离定律"的教学分析与设计…………苏明学0130关于"基因突变和基因重组"的教学……………苏明学o338 "细胞膜——系统的边界"一节教学设计………张斌o426"捕获光能的色素和结构"的教学设计…………操明权O524 "光合色素的提取与分离"的实验教学…………杨志明O532 "食物的消化"一节的教学设计………赵文瑾周然0631"全员参与,全程参与,有效参与"理念在生物学教学中的运用——以"体液调节"第1课时为例………薛静尧O633 "微生物的实验室培养"一节的教学设计………杨富芝0730 "真菌"一节教学…………………………………徐瑛0736课堂教学中问题意识的培养——"细胞中的无机物"教学例举…………………………………杨学志张静0836"从柳苗生长之谜说起"一课的教学设计及反思…王卿璜0936 "细胞的代谢"主题分析及教学建议…龚雷雨夏娉娉1031"细胞呼吸"教学方案……………………………陈廷华1032 "影响光合作用因素"教学设计…………………余厚军ll34 "光合作用的发现"一节的教学设计及教后反思…李艳华ll382006年第41巷第12期生物学通报"捕获光能的色素和结构"一课教学设计……黄复华生K素的发现——科学史在探究式教学设计中的应用………………………………………杨晓梅王燕"细胞呼吸"总复习课的教学设计……..….康红STS教育在中学生物学教学中的探索一例——"如何面对艾滋病"一节教学………………………马千里探究单履虫对刺激反应的实验…张良慧刘济滨教学专题研究l23412361238l240新课程初中牛业卜物学科学,考试的基本经验与若干思考…………………………………………………………施紫雄0133科学探究能力评价的作业设计……I张海和l刘恩山0234初一{j生物学素养多元评价的方案….._-■任小贝0240生物学教育与信息能力培养…………………余自强0340高中生物学教学中开展EEFS教育对学生情感态度与价值观的影响…………………………蔡微李韶山0343建构主义理论与生物学新课程的教学模式………………………………………………………………曹惠玲徐金良0346生物析冈实验没计的5种逻辑方法…………陆建身0428基于新课程背景下的高中生物学课堂教学评价体系的构建…………………………………………………马明0432高中生物学实验教学技能目标的理解与实施…黄敏0529对生物学课程在学校健康教育巾地位的反思…刘海涛0636试论"描述""概述"的区别…………………张涛0734高一fl新旧课程中模拟实验的初步研究张文胜£学丽0740如何在实验探究中作m假设与预期.………..苏明学0838在课堂教学中运用"单元问题"教学策略…………………………………………………………李秀菊张颖之刘恩山0939卜海市青少年生物科技活动历年获奖课题探析………………………………………………张文华俞佩芳刘怡等1035初中生物学发展性学生评价体系的研究……………………………………………………………………李明霞尼志杰1038生物学猜想与假设能力的评价………….…..钱军敏l04l小组合作探究模式的实验研究.……………..爱盈l140生物科学史与探究能力——利用"光合作用发现史"培养学生探究能力的研究……………………陈维汗忠1241影响课堂教学中分组讨论学习实效性的调查与思考…………………………………………………………………平亚茹1245新教师园地一堂竞赛课得到的启示………………………胡宇红O232感悟生物科学实验思想的好教材——谈"DNA是主要的遗传物质"一节的教学……………………刘青0628课堂教学中学生情感伤害的反思…金钧陈秉初O629基于学生实际进行有效教学…………………任小文O738红细胞的几个"例外"特征……………………金钧1033教学实践与交流从"遗传物质基础"的经典实验体味生命科学史蕴涌的教育价值………………………………………王俊霞O139一个"影响酶活性的条件"探究设计和一道原创题…………………………………………………………傅厚春沙丽萍ol41生物学备考中的概念复习(1)…………………郑春和O143生物学备考中的概念复习(2)…………………郑春和O237高考生物学实验专题复习探索………………袁细清O242浅谈牛物学教学中"问题点"的设计及"探究链"的筑建………………………………………………………………吴兴芳O245"种子"专题复习………………………………徐彤O246概念图及其在教与学中的应用………………吴举宏O348谈高考试题中的实验设计……………………温太强0349复习课要改变思路……………………………费世汲O351让学生在习题讲评课中"动"起来……………伊丽梅O353再谈如何指导撰写小论文……………………余力0430确定口标积极评价促进小组合作学习...庄荣婉0435中学开展分.生物学实验的探索与思考.…..赵晓刚0437关于有丝分裂的探究性实验没计……………陈维0439用"预习提纲"引导学生提…………………曾文俊0534学案导学在生物学网络课堂教学巾的应用…林美娟0537多媒体环境下植物教学的课堂控制策略……………………………………………………陈晔陶劲松方亮等0539浅谈学生课外专题实践活动的指导………………………………………………………………李德成赵广宁胡铃冬0639总目次Ⅲ关于遗传学实验教学中果蝇实验的改革探索………………………………………………………方茹杨少宗侄莉0641生命科学实验教学与信息技术整合的实践探索顾佩琴0643生物学课程改革中教研活动的推进作用………沈军0746在生物学教学中培养学生热爱家乡的情感……荐0747网络环境下的协作探究学习……………………刘世华0749"吸烟有害健康"主题式研究性学习与高实验复……………………………………………………………………许允文0840初巾新课程生物学科学业考试的几点尝试……金海尔0842培养学生元认知能力提高生物学复习课效率………………………………………………………………梁沽滑宝平0844夯实基础提升能力——高第1轮复习之找见………………………………………………………………………胡秀隽0846浅谈细胞生物学实验教学改革………张俭刘胜贵O941探究性学习在综合科学教学中的实践…………郑剑锋0943"探索pH埘酶活性的影响"敦学反思…………黄英颖0947对"渗透现象"演示实验的改进…………………陈维0948初叶1开展金鱼养殖研究性学习的实践探索……『E美华0949 "噬菌体侵染细菌"实验的问题式探究学习……周菏0951紧扣课本培养能力——从高考试题看探究性文验试题的取材…………………………………………周可富1042幽¨螺杆菌发现过秤对牛物学教学的启示……徐岳敏1045开展自我指导学爿培养终身学习能力………………………………………………………………………沈霞余胜光l144在大学,丰物学实验教学叶1矸设开放性实验的尝试…………………………………………………张金风刘洋胡冬侮1146设计问题导向指导研究性学习——由"大l=∈今现象"引发的研究课题…………………………………韩玉平l147高兰专题复习:生命系统的动态性……………吴举宏l149讲究讲评方法提高复习效率…………………土珍1248充分发挥校内外资源优势全面培养学生科学素质………………………………………………………………………周湘l250新课稃背景下作业与练习设汁的思考…………谢建rf11252命题与考试高考生物学命题技术拾零(--i.)…………………依兰0145一道遗传题的两种典型错解……………………姚宝骏02l9高考试题点评与口常教学反思………孙仲平刘锡志0254从一道高考试题看植物多倍化…………………鲁吕雄0259遗传学中基冈敛争E类型题解析…………………李生清0355基冈频率解题法…………………………………余顺舟o356以显微镜为背景的高考试题……………………顺红梅0357生物学中的汁赞:题归类…………………………秦改英0441对两个概率计算『口J题的分析……………………岳朝宽o442 同位索标记法专题复习…………………………十永0552提高解题能力从学会审题开始…………………李艳华0645试题难度预估方法的探索实践……李洪滔李振来0743例析高巾生物学试题严谨性的失误……………余波0751《普通高中生物课程标准》(实验)F的考试大纲…依兰0953生物学探究性试题的编制……………林静刘恩山1046研究性学习案例菖蒲对污水净化作用的探索与实践…………………………………………………………………王开诚朱健米刘洋0156低温推迟果蝇交配行为的研究………………………………………………………………………郝大翠姚明镜刘焰0253不同肥料对土壤动物的影响朱健来王开诚刘洋0359关于DNA长度的趣味探究……………………高显奎0361戈于小白鼠记忆力的一则探究活动………………………………………………………………康建国薛达刘宇初等054l探究动物的学习行为……………………………王若燕O543 同…宿舍女生月经周期趋于一致的调查…………………………………………………………王苗李海莉张云慧等0652宝兴蜂桶寨短雉玉IlJ竹营养分析……徐单彬张君0654城市水域,砂地,草坪,他岗岩温度测量分析思考孛晓0853实验与技术一种新的金属电极绝缘方法……槐瑞托于志豪01曰次Ⅱ转基因鱼品系的纯化…………………龙华黄燕0l49埘"植物体内的转氨基作用"实验的改进……………………………………………………………………榭金彩周悻0150平板菌落计数的改进方法………………………李华0151总目次Ⅳ生物学通报2006年第41卷第12期动物骨髓细胞染色体标本制备失败的原因分析………………………………………………黄燕赵小平余红等0152对"观察细胞质流动"实验的改进……………罗益群0248大苞萱草染色体的核型分析…………………李国泰0250果蝇的饲养……………………………………陈晓芸0251"光对鼠妇生活影响"实验的几种改进方法…向群0252巧用硬枝扦插快速繁育银杏……………姬生祥03目次Ⅱ一种洋葱快速生根的方法……………………龚慧明0306用大蒜做植物细胞有丝分裂实验的材料更好…赵培军o360不经染色直接从SDS—PAGE制备凝胶中准确切下所需蛋白条带的方法……………………陈鹏孙群0362改装显微镜………………………………路景泽04目次Ⅱ治疗糖尿病的短肽药物GIJP一1在毕赤酵母中的分泌表达…………………………………侯建华王翠艳方园园等0444番茄碱研究进展以及对鸡红细胞膜Caz+一Mg2*-ATPase活性的影响…………………孙晓甜赵丽段江燕等0447观赏花卉矮牵牛与野生牵牛花的染色体核型比较……………………………………………蔡华张传和王业精等0449太平洋牡蛎养殖技术…………………………李华琳0450"DNA的粗提与鉴定"实验改进………………张叶o452如何观察植物细胞的后含物和质体…………张秀玲0453"叶绿体中色素的提取和分离"实验的改进………………………………………………………郝琦蕾姜晋国刘艳芳0455用旧家具改制光照培养箱…………江廷磊王重力0456蛙肠系膜平铺片的制作……………刘瑜琦刘济滨0461对高中生物学实验材料选取的改进…朱坚毅钱克容0526研究神经发生的方法…………………………包春莹0536一种简易的家兔离体心脏灌流方法……………………………………………………………一林鹏吴萌魏香0544室内饲养蜥蜴的一种好方法......…董丙君王晶琦0545生物墨线绘图的方法与技巧…………………唐安科0546简易唾液收集器的制作……………张海智王雁0548"DNA的粗提取与鉴定"实验改进……………郑焕强0549光学立体最微…………………………………陈宝明0550四棱豆在向光性实验中的应用………………全妙华0551狼尾草在草履虫培养和保存中的应用…一….石玉珍0554蚯蚓观察器的制作……………………………张萍0562红叶椿腋芽增殖培养基的筛选…………………………………………………………………张华辛海波亢秀萍等0647紫花地丁的组织培养与快速繁殖…………………………………………………………………王峰样张福鑫郑泽荣0649磁珠法快速提取鉴定DNA的实验研究………………………………………………………王平康骆延王光志等0650对昆虫标本传统制作技术的一些改进………苏绍科0653观察细胞分裂和染色体变化的压片法制作方法…张志川0662用木耳培养黑根霉……………………………张葵0708投影仪的新用途………………………………何丹龙0739 "DNA粗提取与鉴定"实验的探究………………………………………………………………姜先荣周衍茂鲁红侠0753蝌蚪的呼吸……………………………………单迎春0754利用花卉废料简易制取香花………兰伟蔡健0755一种从淡水中分离变形虫的简易方法…孑L健孙群0809溶菌酶提取过程中出现的问题及改进措施…………………………………………………………………刘洋b梦磊0848手持实验技术在中学生物学教学中的应用…………………………………………………………………朱勋林王重力0850用饮料瓶制作鸟类自动饲喂器……………………………………………………………………白岩青朱勋林蔡金红0852培养温度和保存时间对草履虫密度的影响…于宝霞0856实验设计——证明叶绿素是光合作用的必要条件……………………………………………………………………孙鹏远086l家蚕一对相对性状的遗传实验………………林全顺0952荧光定量PCR应用常见问题………王磊任东明0954一种简易塑化动物标本的制作方法…………唐安科0956辣椒染色体常规镜检技术…张菊平刘珂珂巩振辉等1050翼手类动物染色体的制备方法…………………………………………………………………余燕马金友孙瑞1052"转录","翻译"活动图的制作与演示……………………………………………………………陈丽纯赖乃友谢卫英l053小鼠腹腔注射甲醛后骨髓细胞微核变化检测…王慧阳1055肝脏研磨液的制备和保存…郑俊启王惠琴刘文阁l058"比较过氧化氢在不同条件下的分解"实验的注意事项及改进措施………………………………………丁稳双草履虫分裂生殖过程排队装片制作方法…………………一………………………………王青赵惠玲赵惠卿改进的蛋白银法在伪尾枝虫类群上的应用……施心路XSD640型显微镜常见故障及排除……………罗益群玻璃盒镶蝴蝶标本的制作………………………张焕欣碱性染料用酸配制的原因分析…………………杨红辉桃花水母的采集培养与固定方法……徐善良沈勤指导学生认识蘑菇的孢子印…………李洪臣杨秀艳花粉萌发与花粉管生长实验的改进…王友保张莉国外参考资料美国教材中的小组活动解析…………韩宁王重力1049问题解答单倍体还是多倍体?……………………………余晓丽0638生物学奥林匹克竞赛第14届全国中学生生物学竞赛实验(二)试题分析……………………………………………………刘宁陈星01542004年第15届国际生物学奥林匹克竞赛实验试题F(4)………………………………赵进东许崇任蒋志刚等01582004年第15届国际生物学奥林匹克竞赛实验试题F(4)………………………………赵进东许崇任蒋志刚等02602005年第16届国际生物学奥林匹克竞赛理论考试(1)………………………………………………赵进东许崇任04572005年第16届国际生物学奥林匹克竞赛理论考试(2)………。
小分子化合物在干细胞定向分化诱导研究中的应用

生物医学工程研究Journal of Biomedical Engineering Research基金项目:教育部博士点基金资助项目(20050422013)。
作者简介:赵阳(1984-),男,山东大学生命科学学院生物科学专业学生。
小分子化合物在干细胞定向分化诱导研究中的应用3赵阳,丁丽娜,苗俊英(山东大学生命科学学院发育生物学研究所,山东济南250100)摘要:干细胞的治疗潜能越来越受到重视,如何诱导干细胞定向分化是当前研究的热点化学生物学的发展为干细胞研究提供了多种多样的生物活性小分子。
本研究对小分子化合物在体外诱导干细胞定向分化及在疾病治疗中的应用作了综述。
关键词:干细胞;小分子化合物;诱导;定向分化;应用中图分类号:R318 文献标识码:A 文章编号:167226278(2006)0120058204Application of Sm all Molecule Compoundsin Directed Differentiation of Stem CellsZHAO Yang ,DING Li 2na ,MIAO Jun 2ying(Institute o f Developmental Biology ,School o f Life Science ,Shandong Univer sity ,Jinan 250100)Abstract :The therapeutic potential of stem cells is becoming m ore and m ore im portant.H ow to induce directed differentiation of stem cells is one of the m ost valuable biological subjects.The development of chemical genetics provides various functional small m olecules for stem cell biology.In this review ,how small m olecule com pounds regulate directe differentiation of stem cells in vitro and the application in clinical application are focused on.K ey w ords :S tem cells ;Small m olecule com pounds ;Induce ;Directed differentiation ;Application1引 言干细胞是个独特的细胞群体,既能自我更新,又能分化为具有各种不同功能的成熟细胞。
Wnt信号通路与后口动物体轴的进化发育

HEREDITAS (Beijing) 2011年7月, 33(7): 684―694 ISSN 0253-9772 综 述收稿日期: 2010−12−25; 修回日期: 2011−02−20基金项目:国家自然科学基金项目(编号: 30830023, 31071110)资助作者简介:钱光辉, 博士研究生, 专业方向: 动物发育遗传与分子进化。
E-mail: qianguanghui@通讯作者:王义权, 博士, 教授, 博士生导师, 研究方向: 动物分子遗传与进化。
E-mail: wangyq@网络出版时间: 2011-5-23 16:53:35URL: /kcms/detail/11.1913.R.20110523.1653.004.htmlDOI: 10.3724/SP.J.1005.2011.00684Wnt 信号通路与后口动物体轴的进化发育钱光辉, 王义权厦门大学生命科学学院, 厦门 361005摘要: 动物体轴极性的建立和最初胚轴的形成涉及到一系列信号通路的调控, Wnt 信号通路是其中一条十分保守的信号通路, 并且Wnt/β-catenin 信号通路中的关键成员早在海绵动物中就有发现, 暗示这一信号通路相对于其他信号路径来说可能是最早参与原始后生动物体轴发育的信号通路之一, 并且在体轴后端和腹部的发育及命运分化方面发挥着重要作用。
近年来, 随着体外功能实验体系的建立, 人们发现Wnt 信号通路中很多基因都不同程度地影响了早期胚轴的形成, 例如wnt 基因、母源性基因β-catenin 以及一系列转录因子等。
文章首先对参与后生动物体轴发育的wnt 基因家族的起源与进化关系做一简要分析, 并进一步就经典的Wnt/β-catenin 通路与后口动物的海胆、文昌鱼、斑马鱼、爪蟾和小鼠等类群体轴极性的建立乃至整个体轴形成方面的研究进展做一综述。
关键词: 动物体轴; Wnt 信号通路; β-catenin ; 进化; 发育Wnt signaling pathway and the Evo-Devo of deuterostome axisQIAN Guang-Hui, WANG Yi-QuanSchool of Life Sciences , Xiamen University , Xiamen 361005, ChinaAbstract: A series of signal transduction pathways have been found to regulate the polarity establishment and formation of animal primary body axis. Among them, Wnt signaling pathway is extremely conserved and several key components in the pathway have been identified in the demosponge lineage. This implies that it is one of the earliest pathways involved in the ancestral metazoan axis development and might play an important role in specification and development of posterior and ventral fate of animal axis. Recently, with the establishment of functional experiments in vitro , the body plan formation has been found to be affected, in varying degrees, by many genes in the Wnt signaling pathway, such as members of wnt gene family, maternal gene β-catenin and some transcription factor encoding genes. In this review, we analyzed the evolu-tionary origin of the wnt gene family involved in development of metazoan body plans, and then made a brief review on the roles of canonical Wnt/β-catenin signaling in the polarity establishment and formation of primary body axis in diverse deu-terostomes including sea urchin, amphioxus, zebrafish, frog, and mouse.Keywords: animal body axis; Wnt signaling pathway; β-catenin ; evolution; development动物是如何建立并形成最初的体轴(Body plan formation)是生命科学研究领域中一个重要的生物学基础问题。
高中生物选修1中的2个实验探索

高中生物选修1中的2个实验探索
沈浩宁;丁火华
【期刊名称】《中学生物学》
【年(卷),期】2006(22)8
【摘要】对新课程中的部分实验(芳香油的提取、乳腐的制作)进行了操作和分析,并探索出有效的实验操作过程及其注意事项,降低了实验成本.
【总页数】2页(P24,56)
【作者】沈浩宁;丁火华
【作者单位】南京师范大学生命科学学院,江苏,南京,210097;南京师范大学生命科学学院,江苏,南京,210097
【正文语种】中文
【中图分类】Q-331
【相关文献】
1.高中生物与通用技术选修模块资源共享之探索——基于通用技术现代农业选修模块正常开设的思考 [J], 姜松祥
2.高中生物与通用技术选修模块资源共享之探索——基于通用技术现代农业选修模块正常开设的思考 [J], 姜松祥;
3.高中生物"DNA粗提取与鉴定"实验探索 [J], 王跃光;赵冬阳;斯玲莉
4.学生核心素养的形成——从高中生物实验探索法谈起 [J], 王姗
5.微信公众平台在高中生物学选修课程中的应用 [J], 马凯阳;曹芝琳
因版权原因,仅展示原文概要,查看原文内容请购买。
大鼠内侧视前区支配Barrington核内骶髓投射神经元的光,电镜研究

大鼠内侧视前区支配Barrington核内骶髓投射神经元的光,
电镜研究
王丹;丁玉强
【期刊名称】《中国神经科学杂志》
【年(卷),期】1999(015)001
【摘要】采用光镜及电镜双标记技术,对大鼠内侧视前区(MPO)神经元发出
的投射到脑桥Barrington核的轴突与其内投射到骶髓的神经元的胞体和树突之间的突触关系进行了探讨。
将顺行追踪剂结合生物素的葡聚糖胺(BDA)注射于MPO用来标记它发出的轴突终末;将逆行追踪剂辣根过氧化物酶(HRP)注射于脊髓腰骶段,以此标记Barrington核内投射至腰骶髓的神经元。
光镜观察发现Barrington核内含有大量BD
【总页数】4页(P50-53)
【作者】王丹;丁玉强
【作者单位】第四军医大学解剖学教研室梁Qiu琚脑研究中心;第四军医大学解剖
学教研室梁Qiu琚脑研究中心
【正文语种】中文
【中图分类】R329.481
【相关文献】
1.大鼠内侧视前区支配Barrington氏核内骶髓投射神经元的光电镜研究 [J], 焦西英;林英华;王丹;丁玉强;鞠躬
2.电针足三里穴对溃疡性结肠炎大鼠骶髓后联合核内神经元和胶质细胞形态学变化的影响 [J], 秦明;饶志仁;杨琦;王景杰;段丽;曹荣;黄裕新
3.大鼠腰骶髓后连合核内5—HT样阳性纤维和终末的来源 [J], 徐俊卿;丁玉强
4.中脑导水管周围灰质支配桥脑Barington's核内骶髓投射神经元的电镜双标记研究 [J], 张淼丽;王丹;丁玉强
5.大鼠Barrington核内脊髓投射神经元直接接受腰骶髓传入的电镜研究 [J], 丁玉强;郎兵;宿长军;王红;鞠躬
因版权原因,仅展示原文概要,查看原文内容请购买。
蛋白质溶液NMR结构测定的一些新进展(1)

invasion.Acta U niversitatis Ouluensis M edica,1996,D382(4): 17~363 Reponen P.The Primary structure and developmental expression ofmouse type collagenases.Acta Universitatis Ouluensis,A S cirntiae Rerum Naturalium,1994,A262(1):1~334 Hulboy D L,Rudolph L A,M atrisian L M.M atri x metalloprote-inases as mediators of reproductive function.M olecular Human Re-production,1997,3(1):27~455 Giannelli G,Falk-M arzillier J,Schirald O,et al.Induction of cellmigration by matrix metalloproteinase-2cleavage of lami n i n-5.S cience,1997,277(11):225~2286 Woessner J F.M atrix metalloproteinases and th eir inhibitors inconnective tis sue remodeling.FAS EB J,1991,5(8):2145~ 21547 M urphy G,Docherty J P.T he matrix metalloproteinases and theirinhibitors.Am J Resp Cell M ol Biol,1992,7(2):120~1258 Baragi V M,Fli szar C J,Conroy M C,et al.Contribution of theC-terminal domain of metalloproteinases to binding by tissue inhibitor of metall oproteinases.C-term i nal truncated stromelysin and matrilysin exh i bit equally compromised binding affi n i ties as compared to ful-l length stromelysin.J Biol Chem,1994,269(17):12692~126979 Puente X S,Pendas A M,Liano E,et al.M olecular cloning of anovel membrane-type matri x m etalloprotei nase from a human breast carcinoma.Cancer Res,1996,56(5):944~94910Zhu C,Woessner J F JR.A tissue inhibitor of metalloproteinases and -macroglobulins in th e ovulating rat ovary:Possible regulators of collagen matrix breakdow n.Biology of Reprod,1991,45(2): 334~34211Bjorn S F,Hastrup N,Lund L R,et al.Co-ordi nated expressi on of M M P-2and its putative activator,M T1-M M P,in human placentati on.M ol Hum Reprod,1997,3(8):713~723Extra Cellular Matrix and Matrix Metalloprote-i nases.ZH AO Yun-Ge,Orbit1)ANWAR,ZH U Cheng(The State K ey L aboratory of Rep r oductive Biology,I nstitute of Zoology,The Chinese A cademy o f Sciences,Beij ing100080,China;1)Dep ar tment of Biology,X inj iang Norm al University, r m qi 830053,China).Abstract Extra cellular matrix(ECM)is a dy namic reticular structure,which distributes among cell spac-es.The structural proteins of ECM are varied and in-clude collagens,proteoglycans,multidomain glycop-roteins and other biomacromolecules.These proteins can combine w ith the specific receptors w hich exist on the surface of cells.This w ill result in different gene ex pression by connecting w ith intracellular skeleton frame directly or initiating signal transduction cas-cades,and lead to cells proliferation and differentia-tion.M atrix Metalloproteinases(M MPs)and their tissue inhibitors(T IM Ps)play an important role in the process of ECM destruction and remodeling. MM Ps is also a Zn2+dependant proteolytic enzyme. MM Ps expression is regulated strictly by grow th fac-tors,cytokines,and hormones at transcription level, by their natural activators and inhibitors at protein level.MM P participate various cells physiological and pathological process by deg rading different ECM com-ponents.Key words ex tra cellular matrix,base membrane, matrix metalloproteinases,tissue inhibitors of m atrix metalloproteinases蛋白质溶液NMR结构测定的一些新进展高广华 王大成(中国科学院生物物理研究所,北京100101)摘要 新的标记技术的进展和采用稀释的液晶作为溶剂以提供额外的结构信息,提高了核磁共振技术测定蛋白质溶液三维结构的精度,扩大了分子质量测定范围.目前已经利用多维15N,13C,2H标记NM R测定了许多分子质量为30ku左右的蛋白质溶液结构,这一上限可能还会被进一步提高.关键词 核磁共振,蛋白质三维结构,液晶,偶极偶合,同位素标记学科分类号 Q51,Q6-33多维核磁共振作为测定生物大分子,特别是蛋白质溶液三维结构最重要的手段,近年来取得了飞速的发展.1997年10月Nature结构生物学分册的增刊全面综述了NM R用于生物大分子研究的最新进展[1].在这一领域一直存在着两个备受关注的问题,即NMR测定蛋白质溶液结构的分子质量有无上限[2]和蛋白质溶液NM R结构能否达到与X射线晶体结构相比拟的分辨率水平[3].一般认为NMR 最适于中等分子质量的蛋白质的溶液结构测收稿日期:1998-02-24,修回日期:1998-06-04表1 1995年7月PDB库中蛋白质及多肽NMR结构统计残基数目NM R结构所占百分比小于408320 2%40~8018645 4%81~1208119 8%121~1604711 5%大于160133 2%总 计410100%数目为40至120的蛋白质结构占很大的比例,而160残基以上的蛋白质结构较少.当时分子质量最大的粒细胞集落刺激因子有178个残基,分子质量为20ku左右.目前这一上限已被大大提高,完整测定NMR 结构的蛋白质(单体)分子质量已经突破35ku.表2是已测定的高分子质量蛋白质的情况.本文将就最近在提高NM R结构分析的分辨率和增大被解析结构的分子质量方面的主要进展作一简要介绍.表2 已测定的高分子质量蛋白质NMR结构蛋白质名称PDB文件分子质量 u(残基数) 1色氨酸阻遏物-操纵子DNA复合物370002大肠杆菌磷酸烯醇丙酮酸:糖磷酸转移酶系统中酶 的N端结构域(EIN)1EZA1EZB1EZC30000(259aa)3rRNA转甲基酶(ErmAm)300004人碳酸脱氢酶(HCA )290005大肠杆菌色氨酸全、去阻遏物25000(216aa) 6Bc-l x L-Bak复合物1BXL196aa+16aa 7Bc-l x L蛋白质1LXL196aa8Shc磷酸化酪氨酸结合结构域190aa+12aa 9粒细胞集落刺激因子1GNC178aa1 液晶与高分辨率溶液NMR结构核欧佛豪斯效应(nuclear Overhauser enhance-ment,NOE)是核间距的定性指标,从NOE得到的距离约束的定性本质限制了所得到的生物大分子时间平均构象的精度.原子核间的偶极偶合作用包含许多结构信息,然而在各向同性的溶液中,由于蛋白质分子在各个方向上的翻滚,这种作用平均为零而无法观测.如果能够从偶极偶合作用中直接测定成键原子对相对于分子其他部分的取向,就将极大地提高结构测定的精度.当射频脉冲作用于一个磁性核时,附近另一个磁性核的存在将会由于偶极偶合作用而在两个共振频率之间产生裂分信号,信号的强度对该化学键相对于外磁场的取向十分敏感.人们曾试图用各种方法使蛋白质在溶液中部分取向以观察偶极偶合作用.当蛋白质分子置于强磁场时会因非零的磁感应各向异性而产生微小角度的部分取向.这时可以观测相隔一个化学键的15N-1H 和13C-1H的偶极偶合.由于这些偶极偶合的核间距(即键长)是固定的,因此这些数据就提供了相应的化学键相对于蛋白质磁感应张量的取向的直接信息.然而对于单个的蛋白质分子来说这种磁相互作用的能量非常微弱而难以准确测量.液晶可以使溶剂分子产生定向,但是这种作用对蛋白质分子又太强,所有的分子都被牢牢束缚并完全定向.这时偶极偶合作用就不仅仅出现在相邻原子间而是在整个分子内部,所以将产生大量信号.即使对很小的分子,这些数据也根本无法解析.1997年,T jandra和Bax[3]用简单通用的方法巧妙地解决了这个问题.他们使用的是一种二己酰卵磷脂(DH PC)和二豆蔻酰卵磷脂(DMPC)混合物(1 2 9),室温或略高于室温时这种混合物在水溶液中可以在较低的重量体积比(3%)下从凝胶转化为液晶,这些磷脂分子形成直径为数十纳米厚约4nm的盘状微粒,而且由于这些微粒具有抗磁性,它们沿磁场方向取向.它们之间的空间比通常的蛋白质分子的尺寸大得多,所以蛋白质分子可以在溶液中自由地扩散,只是偶尔与这些微粒碰撞.由于蛋白质分子通常呈椭球状,这种持续的碰撞会使蛋白质分子在液晶排列的方向上发生部分的取向.蛋白质取向的程度与蛋白质分子的形状有关,如图1所示.图1 蛋白质分子在液晶中部分取向示意图通过改变液晶的浓度可以方便地调节蛋白质分子取向的程度,这种作用比现在最强的磁场所产生的取向作用强两个数量级.液晶分子与蛋白质之间没有明显的相互作用,不需要重新再作谱峰的识别.测定这种偶极偶合作用所需的同位素标记的蛋白质样品比测定弱NOE 峰要少,一般只需0 1~0 5mmol L.选择适当的液晶浓度可以只使邻近的核给出可观测的偶极偶合作用.这时蛋白质分子所产生的取向程度已经足以精确地测定多种化学键(13C13C ,13C1H ,15N1H ,13C15N )连接的两个核之间的偶极相互作用,同时又不会使谱图变得极为复杂,这些准确的结构信息可以大大提高NMR 测定的蛋白质结构的质量.Tjandra 和Bax [3]对人泛激素(76aa,8600u)进行的测定结果与0 16nm 高分辨率X 射线晶体结构非常吻合.此外这种方法还有可能提高NM R 测定蛋白质结构的分子质量上限,用高度氘代蛋白质在稀释的液晶溶液中可以快速直接地获得偶极偶合信息,作为与长程NOE 相互补充的结构参数,从而使高分子质量蛋白质溶液结构达到较好的分辨率.虽然目前这种方法还没有用于测定完整的蛋白质结构,但是在原理上已无太大的问题.一旦用这种方法测定的蛋白质NM R 结构的精度能一般地达到高分辨率晶体结构的水平,NMR 技术将进入一个新的阶段.2 氘代与高分子质量蛋白质主链识别随着蛋白质分子质量增大,分子中氢原子的数目急剧增加,谱峰的线宽变宽,导致谱峰严重重叠,谱图的解析几乎不可能进行.如果将蛋白质分子中的1H 用2H 取代将大大简化谱图.目前,三共振(triple resonance)实验结合氘代技术是解决溶液NM R 分子质量限制的最好办法.1993年氘代技术用于三共振实验,并出现了将序列相邻酰胺基团相关的4D H N (COCA )NH 实验.Yamazaki 等[4]设计了一系列用于测定主链NH,15N,13C 和侧链13C 化学位移的实验.在高度氘代的蛋白质中,得到13C 和13C 的化学位移有助于识别二级结构,因为这时NOE 峰的缺失要求尽可能多地测定其他的结构信息.在此之后大量用于主链识别的实验相继出现.最近Yamazaki 等[5]报道了一种获得仅在13C 位置质子化而其他部分高度氘代的蛋白质的方法.这种标记方法特别有助于因酰胺质子交换速率过快而无法进行的,基于NH 的主链识别方法.Yamazaki 等[4]还设计了一套用于主链识别的实验来利用通过上述方法制备的的蛋白质样品.Farmer 和Venters [6]设计了一系列用于高度氘代的蛋白质侧链识别的实验.在质子化的15N,13C 标记蛋白质中,侧链化学位移的获得通常是将产生于烷基质子的信号传递到相邻的碳原子然后通过接力传递到主链的15N 和NH 自旋.虽然这种办法易于分析,但这类实验的灵敏度在蛋白质的分子质量超过15ku 时会大大降低.对这类实验的许多改进已经应用于高度氘代的人碳酸脱氢酶(HCA ,259个残基,29ku)的主链识别,如使磁化强度从烷基碳开始传递等.与全质子化样品中磁化强度产生于质子的实验相比,经过改进的实验的灵敏度有明显提高.Farmer 和Venters [6]还设计了许多实验来进行高度氘代样品中侧链15N NH 共振峰的识别.在高度氘代样品中可用于结构测定的NOE 数目减少,所以分子中保留下来的所有质子都必须尽可能的识别出来以增加结构信息,这些质子包括Glu 和Asp 的酰胺质子和Arg 的 和 (如果能够得到)质子.在对H CA 的研究中,仅对高度氘代样品才能进行主链的全识别,而质子化的15N,13C 标记样品仅能达到85%识别.与此类似,Shan 等[7]进行了分子质量达64ku 的色氨酸阻遏物,色氨酸和有22个碱基对的DNA 片段的三聚体的NH ,15N,13C ,和13C 化学位移的接近完全的识别,其中蛋白质部分高度氘代(>90%),而氘代率为70%时所进行的识别未能成功.Constantine 等[8]用高度氘代的样品进行了无底物结合和NADP +结合的UDP -N -乙炔醇丙酮酸葡萄糖胺还原酶(M urB)的接近完全的主链识别(包括13C ).Garrett 等[9]利用全氘代15N,13C 标记的大肠杆菌磷酸烯醇丙酮酸:糖磷酸转移酶系统中酶 的N端结构域(EIN),长度为259个残基的片段的蛋白质样品来识别在全质子化样品中无法识别的8个残基.这些结果表明对于分子质量达30ku或更高的蛋白质进行完全或接近完全的识别必须使氘代达到很高的程度.目前许多三共振实验用于烷基位置高度氘代的蛋白质样品时灵敏度都相当好,但是存在的问题是氘代使能够用于结构计算的NOE数目减少.因此Laue[10]建议使用约50%氘代的样品作为折衷方案,一方面通过与主链15N和NH相关进行的侧链1H 和13C共振峰识别的实验可以获得合适的灵敏度,另一方面又可以得到较高灵敏度的NOE谱.3 反标记与蛋白质的溶液三维结构NM R进行结构测定主要依赖于建立起质子间的距离约束,这些约束将空间上靠近(约0 5nm 范围之内)却通常在一级序列上远离的质子联系起来.由于高度氘代使这些质子的数目大大减少,从而使高分子质量的蛋白质的结构测定面临极大的困难.虽然位于可交换位置的质子(如主链和侧链的NH)能够在样品制备之后被重新引入,在高度氘代样品中仍然可以建立起NH-NH间的距离约束,且仅仅依靠NH-NH间的距离约束也可以得到一定分辨率的蛋白质结构,但是实验表明,测定精确的蛋白质结构还需要额外的距离信息作为NH-NH间距离约束的补充.为此目的,许多研究小组致力于研究某些关键基团质子化而其他位置均高度氘代的15N,13C标记的样品,这种方法现在被称为 反标记(reverse labeling procedures) .很明显最合适的选择是Val,Leu和Ile的甲基.因为一方面它们是蛋白质疏水核心中最常见的残基,另一方面它们的堆积是稳定整个分子折叠的重要因素.此外即使在高分子质量的蛋白质分子中围绕甲基三重轴的旋转也会使13C和1H的线宽变窄,有利于谱图解析.最后,相对于谱图的其他部分,13C-1H相关谱中的甲基区域较易分析.现在已有多种不同的方法来制备甲基质子化的高度氘代的蛋白质样品.Metzler等[11]和Laue 等[12]的方法是将15N,13C,1H-Ile,Leu,Val引入待研究的蛋白质中以增加可以获得结构信息的位置的数目.培养基中含有氘代的葡萄糖,95%D2O, 15N,13C,1H-Ile,Leu,Val及一种15N源如15NH4Cl.他们都报道在生物合成引入Leu,Val,Ile的过程中C 位置可以获得高度的氘代,使用于主链和侧链识别的许多实验的灵敏度大大提高. Rosen等[13]报道了另外一种得到甲基质子化高度氘代蛋白质样品的方法.培养基含有13C,1H-丙酮酸酯(CH3COCOOR)作为碳源,95%D2O以及NH4Cl.质谱和NMR的结果表明这个方法得到的蛋白质中大多数侧链位置是氘代的,而Ala,Val, Leu,和Ile(仅 2位置)的甲基是质子化的,它们都来自丙酮酸酯的质子化的甲基.甲基化的程度从Leu的约80%,Ile(仅 2位置)和Val的60%到Ala的40%.另外一些位置,包括Asx的C 和Glx,Arg,Pro的C 也有一定程度的质子化.但重要的是位于Ala,Val,Leu和Ile的质子化的甲基和主链酰胺基团间所有的碳原子均是高度氘代的,这将有利于用磁化强度沿着侧链碳原子传递到主链酰胺的脉冲序列进行侧链的识别.Kay等[14]建立了制备Val,Leu,Ile(仅 1位置)甲基质子化的高度氘代的蛋白质样品的方法.蛋白质由大肠杆菌高效表达,15N,13C,2H培养基中含有氘代葡萄糖作为碳源并辅以[2,3-2H2]-15N,13C-Val和[3,3-2H2]-13C-2-氧代丁酸酯(CH3C H2COCOOR).现在已经建立了综合利用CH3-CH3,CH3-NH 和NH-NH间NOE信息来测定蛋白质结构的方法,与只利用NH-NH间NOE的方法相比,结构的正确性和精度都明显提高.PLC SH2结构域[15], Grb2NSH3 SOSE蛋白质-多肽复合物[11]和ras p21蛋白质GDP[12]的结构测定证实了其正确性.如果要求更高的分辨率,就要在蛋白质中引入更多的质子.Farmer等[16]和Laue等[12]制备了芳香残基质子化的高度氘代的M urB-NADP+复合物样品.此外,还可以利用二面角的构象数据库或13C 和13C 的化学位移来精化由有限的NOE约束获得的NMR结构.尽管在高度氘代的样品中选择性质子化对于高分子质量蛋白质的结构测定是必需的,但值得注意的是可以利用多个氘代程度不同的样品进行实验. Fesik等[17]采用这种方法测定了分子质量为29ku 的rRNA转甲基酶的结构,主链和侧链的识别由部分氘代(约75%)的15N,13C标记的样品获得. NH对间的距离约束来自高度氘代(>90%)的样品,而另外的NOE则来自全质子化的15N,13C标记的蛋白质样品.Fesik领导的研究小组已经利用这种方法测定了Shc磷酸化酪氨酸结合结构域(190aa)[18]、Bc-l x L蛋白质(209aa)[19]及Bc-l x L 与Bak多肽复合物的结构[20].Garrett等[9]采用类似的方法研究了E IN的结构.高度氘代的15N,13C 标记样品用来证实由全质子化样品所建立起来的对NH,15N和13C的识别,并得到了在未氘代时不可能进行的识别.全氘代样品则用来以高的灵敏度记录NH-NH间的NOE.对于质子化样品的谱图质量较好的高分子质量蛋白质如EIN和Bc-l x L来说,结构测定的最好方法就是首先利用甲基质子化高度氘代的样品获得的CH3-CH3,CH3-NH,NH-NH 间的NOE约束建立初始结构模型,然后再利用质子化样品获得的大量NOE进行结构的精化.目前NMR测定的蛋白质完整溶液三维结构的最大分子质量为37ku,主链完全识别的最大分子质量已达64ku,精度最高可达0 15nm分辨率.与X射线衍射方法相比,NMR还是一种尚未完全成熟的技术,其各个方面都在经历着不断的变革.从已经显露的技术发展趋势看,似乎可以明确回答本文开篇所提到的问题,即NMR结构解析的分子质量还有很大提高的余地,不久将可能扩展到50至60ku,结构解析的分辨率可以达到与X射线单晶结构分析相当的水平.随着理论知识的日益完善,实验技术的新的进展以及硬件性能的不断提高, NMR技术将在结构生物学领域发挥更大的作用.参 考 文 献1 W agner G.An account of NM R in structural biology.NatureStructural Biology,1997,4(supplement):841~8442 Kay L E,Gardn er K H.Solution NM R spectroscopy beyond25kDa.Current opinion in structural biology,1997,7(5):722 ~7313 T jandra N,Bax Ad.Direct measurement of di stances and angles inbiomolecules by NM R in a dilute li quid crystalline medium.S cience,1997,278(5340):1111~11144 Yamazaki T,Lee W,Revington M,et al.An HNCA pulsescheme for the backbone assignment of15N,13C,2H labeled proteins:application to a37kDa trp repressor-DNA complex.J Am Ch em S oc,1994,116(14):6464~64655 Yamazaki T,Tochio H,Furui J,et al.Assignment of backboneres onances for larger proteins usi ng the13C-1H coherence of1H , 2H,13C and15N labeled sample.J Am Chem Soc,1997,119(5):872~8806 Farmer B T,Venters R A.Assignm ent of aliphatic side-chain1HN 15N resonances in perdeuterated proteins.J Biomol NM R, 1996,7(1):59~717 Shan X,Gardner K H,M uhandi ram D R,et al.Assignment of15N,13C ,13C and HN resonances in an15N,13C,2H labeled 64kDa trp repressor-operator complex using triple resonance NM R spectroscopy and2H-decoupling.J Am Ch em Soc,1996,118(28):6570~65798 Constantine K L,M ueller L,Goldfarb V,e t al.Characterizationof NADP+bindi ng to perdeuterated M urB:backbond atom NM R assignments and chemica-l shift changes.J M ol Bi ol,1997,267(5):1223~12469 Garrett D S,S eok Y J,Liao D I,e t al.Solution structure of30kDa N-term i nal domain of enzyme I of the Escherichia coli phosphoenolpyruvate:sugar phosphotransferase system by multidimensional NM R.Bi ochemis try,1997,36(9):2517~ 253010Nietlispach D,Clow es R T,Broadhurst R W,et al.An approach to the s tructure determ i nation of larger proteins usi ng tri ple resonance NM R experiments i n conjunction w ith random fractional deuteration.J Am Chem Soc,1996,118(2):407~41511M etzler W J,W ittekind M,Goldfarb V,et al.Incorporation of 1H 13C 15N-(Ile,Leu,Val)into a perdeuterated,15N labeled protein:potential in structure determination of large proteins by NM R.J Am C hem S oc,1996,118(28):6800~680112Smith B O,Ito Y,Raine A,et al.An approach for global fold determination usi ng li m i ted NM R data from larger protei ns selec-tively protonated at specific residues.J Biomol NM R,1996,8(3):360~36813Rosen M K,Gardn er K H,W i llis R C,et al.Selective methyl group protonati on of perdeuterated proteins.J M ol Biol,1996, 263(5):627~63614Gardner K H,Kay L E.Producti on and incorporation of15N, 13C,2H(1H- 1methyl)isoleucine into proteins for multidimen-si onal NM R s tudies.J Am Ch em S oc,1997,119(32):7599~ 756015Gardner K H,Rosen M K,Kay L E.Global folds of highly deu-terated,methyl protonated protei ns by multidimensional NM R.Bi ochemis try,1997,36(6):1389~140116Farmer B T,Constantine K L,Goldfarb V,et al.Localizing the NADP+binding site on the M urB enzyme by NM R.Nat Struct Bi ol,1996,3(12):995~99717Yu L,Petros A M,Schnuchel A,et al.Solution structure of an rRNA methyltransferase(ErmAm)that con fers M LS antibiotic resistance.Nat Struct Biol,1997,4(6):483~48918Zhou M M,Ravichandran K S,Olejniczak E F,et al.Structure and ligand recognition of the phosphotyrosine binding domai n of Shc.Nature,1995,378(6577):584~59219M uchmore S W,Sattler M,Liang H,et al.X-ray and NM R structure of human Bc-l x L,an inhibitor of programmed cell death.Nature,1996,381(6580):335~34120Sattler M,Liang H,Nettesheim D,et al.Structure of Bc-l xL-Bak peptide complex:recognition betw een regulators of apoptosis.Scien ce,1997,275(5302):983~986Progress in the Determination of Protein Solution Structure by NMR.GAO Guang-Hua,WANG Da-Cheng(I nstitute of Biop hysics,The Chinese Academy o f Sciences,Beijing100101,China). Abstract Advances in new label strategies and application of liquid crystalline medium for g iving additional structural information have improved theaccuracy of solution protein structures determined by NM R,and extended the molecular w eight lim it. Several structures of proteins of30ku have been solved using15N,13C,2H labeling multidimensional NM R techniques,and this lim it w ill probably be surpassed in the near future.Key words nuclear mag netic resonance,protein three-dimensional structure,liquid crystal,dipolar coupling,isotope labeling纤维素酶分子结构和功能研究进展*阎伯旭 齐 飞 张颖舒 高培基1)(山东大学微生物技术国家重点实验室,济南250100)摘要 概述了近10年来利用结构生物学和蛋白质工程技术在纤维素酶分子结构和功能方面研究的进展,包括:酶分子结构域的拆分、催化域和纤维素结合结构域的结构和功能的研究,纤维素酶的分子折叠.并展望了该领域的研究前景.关键词 纤维素酶,催化结构域,纤维素结合域结构域,分子折叠,活性位点,双置换机制学科分类号 Q936纤维素这一巨大可再生资源的有效利用对解决环境污染、食品短缺、能源危机具有重大现实意义,利用微生物产生的纤维素酶来分解和转化纤维素则是纤维素利用的有效途径,故对纤维素酶的研究越来越引起人们的重视.纤维素酶的研究,自从1904年在蜗牛消化液中被发现,至今已经历三个发展阶段.第一阶段是80年代以前,主要工作是利用生物化学的方法对纤维素酶进行分离纯化.但由于纤维素酶来源广泛、组分复杂、纯化甚为困难,故进展缓慢.第二阶段是1980年至1988年,主要工作是利用基因工程的方法对纤维素酶的基因进行克隆和一级结构的测定.Tr ichoder ma reesei 的内切酶(EG 、EG )和外切酶(CBH 、CBH )、Cellulomonas f imi的内切酶(CenA、CenB、CenC、CenD)和外切酶(CbhA、CbhB、Cex)、Clostridium ther mocellum的内切酶(CelA、CelB、CelC、CelD)的基因已被克隆和测序,并在大肠杆菌,酵母菌等中得到表达.第三阶段是1988年至今,主要工作是利用结构生物学及蛋白质工程的方法对纤维素酶分子的结构和功能进行研究,包括纤维素酶结构域的拆分、解析、功能性氨基酸的确定、水解的双置换机制的确立,分子折叠和催化机制关系的探讨.1 纤维素酶分子结构域的拆分1986年T ilbeurgh[1]用木瓜蛋白酶有限酶切Tr ichoder ma reesei的CBH 分子得到具有独立活性的两个结构域:一个是具有催化功能的催化域(catalytic domain,CD),另一个是具有结合纤维素功能的纤维素结合(吸附)区(cellulose binding domain,CBD),之后用类似的方法在多种细菌和真菌的纤维素酶中发现类似的结构.CBD在纤维素酶中位于氨基端或羧基端,它通过一段高度糖基化的linker与催化区相连.细菌纤维素酶的linker富含Pho、T hr,而真菌纤维素酶的linker富含Gly、Ser、Thr;细菌纤维素酶的CBD与CD夹角为135 ,而真菌为180 ;细菌纤维素酶有两个酶切位点可将CBD与linker 分别切去,而真菌纤维素酶一般只有一个酶切位点可将CBD与linker一同切去.由于纤维素全酶分子呈蝌蚪状、其linker高度糖基化且具有较强柔韧性,故很难得到结晶.而结构域的拆分使呈球形催化域的结晶成为可能,为纤维素酶的结构和功能研究开辟了道路.2 催化域结构和功能的研究采用X光衍射的方法1990年Rouvinen[2]对T.reesei的CBH 的催化域、1992年Juy[3]对C. therm ocellum的CelD的催化域、1993年Spezio[4]*国家教委博士点基金资助项目.1)通讯联系人.收稿日期:1998-02-19,修回日期:1998-05-29。
GDNF激活星形胶质细胞保护PD模型大鼠黑质多巴胺能神经元

GDNF激活星形胶质细胞保护PD模型大鼠黑质多巴胺能神经元刘洪梅;余景考;丁艳霞;王红军;王炎强;高殿帅【期刊名称】《神经解剖学杂志》【年(卷),期】2006(22)4【摘要】探索在胶质细胞源性神经营养因子(GDNF)保护受损多巴胺(DA)能神经元过程中,星形胶质细胞的可能作用。
成年大鼠右侧纹状体内注射6-羟多巴胺(6-OHDA)制备Parkinson病(PD)模型。
动物模型右侧黑质内注射GDNF,于注射后第7、14和21d进行动物行为学分析,行黑质节段连续冠状石蜡切片,以抗酪氨酸羟化酶(TH)、胶质原纤维酸性蛋白(GFAP)抗体免疫组化染色,分别显示DA能神经元和激活的星形胶质细胞,光镜观察并进行细胞计数。
用免疫印迹法分析注射后第21d 黑质内TH、GFAP蛋白表达量,蛋白条带用图像处理仪扫描,LabWorks软件分析处理。
结果显示:动物在注射GDNF后第21d,行为学功能出现显著性恢复;TH阳性神经元数量显著增加;在检测的各时间点均可观察到大量的GFAP阳性细胞;TH、GFAP的蛋白表达量显著高于对照组。
以上结果提示,黑质内注射GDNF可能通过激活了的星形胶质细胞保护PD模型大鼠黑质DA能神经元。
【总页数】6页(P367-372)【关键词】Parkinson病模型;星形胶质细胞;多巴胺能神经元【作者】刘洪梅;余景考;丁艳霞;王红军;王炎强;高殿帅【作者单位】徐州医学院神经生物学教研室【正文语种】中文【中图分类】Q421;Q813.11【相关文献】1.表皮生长因子加强胶质细胞系源性神经营养因子对PD模型大鼠黑质多巴胺能神经元的保护作用 [J], 刘洪梅;丁艳霞;王炎强;高殿帅2.PD模型中GDNF与星形胶质细胞对黑质DA能神经元的影响 [J], 金梅芳;李洁3.GDNF对PD模型大鼠黑质神经祖细胞和神经元前体细胞增殖及分化的影响 [J], 刘洪梅;丁艳霞;王红军;王炎强;高殿帅4.塞来昔布对脂多糖所致PD大鼠黑质多巴胺能神经元损伤的保护作用 [J], 牛平;陈鑫;王聪杰;刘宝茹;辛志强5.6-OHDA所致PD大鼠模型黑质多巴胺能神经元与胶质细胞的变化 [J], 张思源;齐建国;章为;何毅因版权原因,仅展示原文概要,查看原文内容请购买。
2018年全国优秀博士学位论文

复杂两相流动中颗粒碰撞的DEM-LES/DNS耦合 模拟研究
桂南
樊建人
浙江大学
2013048
远程量子通信的实验研究
金贤敏
潘建伟
中国科学技术大学
2013049
量子点光学性质的经验赝势计算
龚明
郭光灿
中国科学技术大学
2013050
纳米纤维宏观组装体的制备及功能化研究
梁海伟
俞书宏
中国科学技术大学
韩波
陈应春
四川大学
2013082
基于纹波的开关功率变换器控制技术及其动力
学行为研究
周国华
许建平
西南交通大学
2013083
低相关序列设计及其相关编码研究
周正春
唐小虎
西南交通大学
2013084
基于四维天线理论和强互耦效应的阵列天线技 术研究
陈益凯
杨仕文
电子科技大学
2013085
在胚珠表皮中时空调控生长素的生物合成提高 转基因棉花的纤维产量和品质
振成像研究
朱雪玲
姚树桥
中南大学
2013075
质料先天与人格生成一一对舍勒现象学的质料 价值伦理学的系统研究
张伟
倪梁康
中山大学
2013076
Alexandrov空间上的Ricci曲率
张会春
朱熹平
中山大学
2013077
农林生物质半纤维素分离纯化、结构表征及化学 改性的研究
彭锋
孙润仓
华南理工大学
2013078
基于网络引证关系的知识交流规律研究
杨思洛
邱均平
武汉大学
2013063
流域梯级电站群多目标联合优化调度与多属性 风险决策
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
4. Ultra-thin sectioning with
a diamond knife.
5. Observation after staining
with uranyl acetate and lead
Rough ER
Smooth ER
Golgi apparatus
Secretary granule
Endosome Multivesicular body Primaryiary lysosome
Myelinated fiber
Ranvier node
Immunoelectron Microscopy
For subcellular localization of antigen in a particular subcellular organelle or compartment.
Two major approaches:
Pre-embedding immunoelectron microscopy Post-embedding immunoelectron microscopy
(2x10-5 m) Resolution: > 0.2 m vs nm Images: film and digital
Categories in Biology
Transmission EM 透射电镜
Scanning EM 扫描电镜
Transmission EM
光源:电子枪,超高
工作原理: 用一束极细的电子 束扫描样品,在样品表面激发 出次级电子,次级电子的多少 与样品的表面结构有关 。次级 电子由探测体收集,转化为扫 描图像。图像为立体形象,反 映了标本的表面结构。
为了使标本表面发射出次级电 子,标本在固定、脱水后,要 喷涂上一层重金属微粒,重金 属在电子束的轰击下发出次级 电子信号。
Advantages:
Good for preservation of antigens
Disadvantages:
1. The antigen could not be precisely localized, because of diffused DABbased reaction products. 2. Ultrastructure: so so. (Triton X-100)
For examination of ultra-structures at subcellular level (Membrane, Lysosome,
Golgi apparatus, Synapse)
The same approach as light microscopy
Section: m vs 60 nm (ultrathin section) Light wave (0.4-0.7 m) vs electron “wave”
扫描电镜的最大有效放大倍率 为20k。
Part II
In situ hybridization
Brief introduction
To detect cellular distribution
of mRNA
1. Differ from Southern Blot 2. Cellular distribution of
Electron microscope &
In situ hybridization
Lab of Neural Development, ION
丁玉强
E-mail: dingyq@
Part I Electron Microscopy
Brief introduction of EM
mRNA may differ from
that of protein.
Advantages compared with IHC
Any interested genes, and time-saving
Synapse
Routine processing for EM
1. Fixation: Glutaraldehyde is
required
2. Post-fixation: 1% OsO4 to
preserve lipid and lipoprotein structures.
3. Embedding: Dehydration
压,电子束
注:在使用时,电镜镜体 内保持真空状态。抽真空 需要若干小时
成像系统:电磁场,
会聚成像
观察系统:荧光屏,
胶片,数码设备
Just show you several examples and then give you a brief
introduction of how to do it
1. Precise localization of antigen.
2. Multiple labeling 3. Good for GABA and Glu.
Disadvantage
Tricky in some cases.
Scanning electron microscopy
扫描电子显微镜:观察标本的 表面结构
Pre-embedding for EM
Tissue preparation:
Fixation: 4% PFA+0.05% Glu Sectioning:
Vibratome not cryostat or others
Advantages and disadvantages of pre-embedding IHC for EM
Bao, et al. PNAS 1997
New method is available now to overcome the limitation
Nanogold intensification
Post-embedding for EM
immunogold labeling
Advantages