~$西藏紫花针茅叶功能性状沿降水梯度的变化_胡梦瑶
浅析3种紫色植物不同叶龄花青素含量的变化
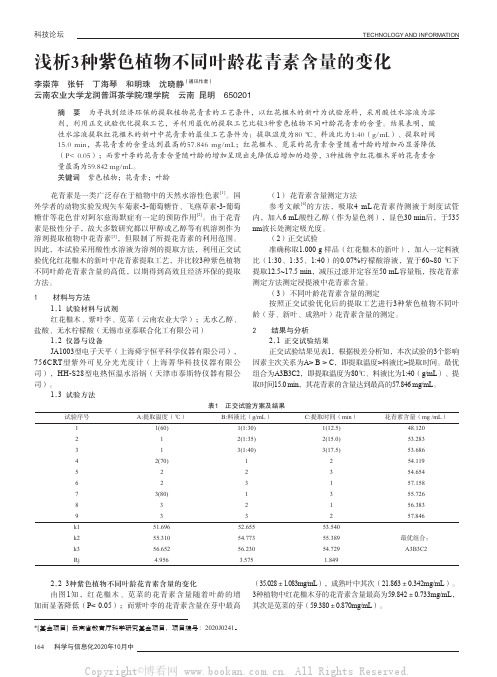
注:不相同字母表示差异显著(P <0.05)。
图1 3种紫色植物不用叶龄花青素的含量
结束语
采用正交试验对水提红花檵木的新叶中花青素工艺进行了
(上接第163页)
分类整理和传输各类数据,存储了数据信息之后,在分析数字天然气管道故障和优化调度等工作中,可以随时调取分析数据信息,提升天然气输送工作的安全性。
3.2 GIS系统
计算机技术不断高速发展,也带动了GIS系统发展,在很多领域广泛利用GIS系统。
结合GIS系统和其他技术,可以发挥着显著的作用。
结合数据库技术和网络通信技术,可以建立地物,在数字天然气管道管理中,GIS地区可以提供详细的天然气管道的地理信息,通过数据库技术管理天然气管道的信息,更加科学地管理数字天然气管道。
技术人员在GIS地图中。
藏药紫花黄华的研究进展

《西藏科技》2021年3期(总第336期)动植物研究藏药紫花黄华的研究进展*达瓦潘多(西藏藏医药大学,西藏拉萨850000)摘要:目的为藏药紫花黄华所含有的化学成分进行分类,从而筛选出该味药材的活性成分和最佳提取溶剂,为下一步制定质量标准奠定理论基础。
方法方法文章通过参考大量有关紫花黄华的研究文献和报道,首先对藏药紫花黄华在植物解剖学、显微鉴定、生物学以及生态学等方面的研究成果进行了较全面、系统地总结,其次利用天然药物溶剂提取法,用水、甲醇、乙醇三种溶剂制备藏药紫花黄华粉的提取液,再采用化学检识法之显色试验、荧光试验、沉淀试验以及纸色谱层析显色试验等方法鉴别该药材所含有的化学成分。
结果结果根据试验结果,分析各类显色现象和沉淀现象,得出藏药紫花黄华含有黄酮类、酚类、有机酸类、生物碱类以及植物甾醇、三萜类等天然药物化学成分。
结论将上述结果与黄华木属和沙冬青属化学成分作对比,进而提出紫花黄华活性成分筛选的方向。
关键词:紫花黄华研究进展*基金项目:2018年藏医药区域协同创新中心项目“藏药紫花黄华化学成分研究”(XTCX 2018026)藏药紫花黄华是藏药方剂中常用的药材之一。
它是豆科黄华属植物紫花黄华(Thermopsis barbata Benth.)的干燥根和根茎。
秋季采挖,切段并阴干[1]。
其性寒味苦,有小毒。
具有杀虫、止痛、消炎等功效。
用于虫病、高血压、中风、炭疽、水肿、肺热、咳嗽等症状。
1紫花黄华的药材鉴定学研究成果1.1植物地理学研究黄华属植物主要在亚洲和北美洲间断分布[4],但是两个地域没有发现同种植物。
根据袁永明等通过植物系统解剖研究,认为黄华属是介于沙冬青属和黄华木属之间的类群[2]。
其分布较广,生态环境的跨越幅度也比较大,有分布于低海拔地区的种类,也有分布于林下、干旱、半干旱的广大高原地区的。
紫花黄华就是分布于高寒干旱地区,它是黄华属常绿木本类群为适应环境而进化而成的极端例子[4]。
高原2种草本植物的光合作用和叶绿素荧光参数日动态

高原2种草本植物的光合作用和叶绿素荧光参数日动态高原2种草本植物的光合作用和叶绿素荧光参数日动态高丽楠1,张宏2,陈舒慧1,贺琼1,孙佳媛1【摘要】摘要:以来源于川西北高寒沼泽草甸2种优势草本植物木里苔草(Carex muliiensis Hand.-Mazz)和四川嵩草(Kobresia setchwanensis Hand.-Mazz)为试材,利用LI-6400便携式光合作用测量系统及PAM-2100调制叶绿素荧光仪,在夏季7月对其光合速率及叶绿素荧光特性的日动态进行测定.结果表明:木里苔草光合速率(Pn)和蒸腾速率(Tr)日动态均呈双峰曲线,而四川嵩草的光合速率和蒸腾速率日动态表现为单峰曲线,并且都出现光合“午休”现象.根据Pn、胞间CO2浓度(ci)、气孔限制值(Ls)的变化方向,推测木里苔草的光合“午休”主要受气孔因素限制,但四川嵩草主要受非气孔因素限制.清晨7:00,木里苔草和四川嵩草的Fv/Fm都低于0.8,发生了长期的光抑制.19:00,2种草本Fv/Fm均未恢复到7:00的水平,表明光系统Ⅱ(PSII)反应中心可能发生了不可逆失活,或光合机构受到了破坏或损伤.影响2种草本叶片Pn日变化的主要决定因子是Tr,主要限制因子是叶温(Tleaf).研究表明,木里苔草较低的Tr 和较高的水分利用效率(WUE)显示出在受水分胁迫的高温低湿环境下的适应性;但高寒草甸四川嵩草实际光化学反应效率(ΦPSII)和半饱和光强(Ek)都显著大于木里苔草,表明对强光耐受能力强.【期刊名称】四川师范大学学报(自然科学版)【年(卷),期】2015(038)004【总页数】11【关键词】关键词:高寒沼泽草甸;光合作用;叶绿素荧光【文献来源】https:///doc/3614503249.html,/academic-journal-cn_journal-sichuan-normal-。
高寒紫花针茅群落对不同形态氮素的利用策略与生态适应机制

高寒紫花针茅(Stipa purpurea)是生长在高寒地区的一种禾草植物,其群落对于不同形态的氮素的利用策略与生态适应机制是生态学研究领域的一个重要课题。
了解植物群落对氮素的响应和适应机制有助于更好地理解高寒生态系统中的养分循环、植物生长及相互作用等生态学过程。
### 1. **氮素在高寒紫花针茅群落中的重要性:**氮素是植物生长发育中的重要养分之一,对于植物体内蛋白质合成、光合作用等关键生理过程至关重要。
在高寒地区,氮素的获取和利用对于植物的生存和繁殖具有特殊的意义。
紫花针茅作为高寒地区的主要植被之一,其氮素利用策略对于整个高寒生态系统的稳定和健康具有重要影响。
### 2. **对不同形态氮素的利用策略:**高寒紫花针茅群落对不同形态的氮素可能展现出不同的利用策略。
一方面,硝态氮(NO3-)通常为植物较为容易吸收的形态,因此在土壤中硝态氮的浓度可能影响紫花针茅的生长。
群落可能通过增加根系对硝态氮的吸收能力来提高氮素利用效率。
另一方面,群落也可能对铵态氮(NH4+)有一定的适应性,通过调节植物根际微生物群落来增加对铵态氮的转化和吸收效率。
### 3. **生态适应机制:**在高寒紫花针茅群落中,可能存在多种生态适应机制,以适应不同的氮素环境。
首先,可能存在植物根系形态和生理特性的适应性变化,使其更好地获取土壤中的氮素。
例如,根系的生长深度和根系毛的密度可能受到氮素水平的调控,以更好地适应氮素的分布情况。
其次,可能存在一些共生微生物,如根际固氮细菌等,对提高土壤中氮素的有效性和可利用性有积极作用。
此外,植物本身的生长阶段和生命周期可能对其对氮素的利用策略产生影响,不同的生长阶段可能表现出不同的氮素利用特征。
### 4. **植物与土壤相互作用:**高寒紫花针茅群落中的氮素利用策略与土壤的相互作用密切相关。
土壤的氮素含量、pH值、质地等因素可能影响着植物对氮素的吸收和利用。
植物通过根系分泌物质调节土壤的氮素形态,与土壤中的微生物互动,构建一种良性的氮素循环系统。
青藏高原紫花针茅草原研究进展

青藏高原紫花针茅草原研究进展
岳鹏鹏;彭敏
【期刊名称】《榆林学院学报》
【年(卷),期】2014(024)004
【摘要】紫花针茅草原是青藏高原高寒草原中分布面积最大、最重要的群落类型.综述了紫花针茅草原研究的发展过程,分析讨论了一些热点领域和有价值的研究结果,总结出紫花针茅草原研究中存在的三个主要问题.(1)局部分散研究多,系统研究少;(2)特征研究多,机理研究少;(3)静态研究多,动态研究少.在此基础上,笔者认为紫花针茅草原退化及植被-环境互作机理是未来的研究重点,演替监测和群落演替趋势还需深入研究,进而增强对青藏高原紫花针茅草原群落特点和规律的理解,更好的为青藏高原的生态保护和生态系统科学管理提供基础资料.
【总页数】6页(P1-6)
【作者】岳鹏鹏;彭敏
【作者单位】榆林学院生命科学学院,陕西榆林719000;中国科学院西北高原生物研究所,青海西宁810000
【正文语种】中文
【中图分类】S812
【相关文献】
1.天山高寒草原不同退化梯度紫花针茅群落结构特征研究 [J], 杨娇;安沙舟;张爱宁;阿依敏·波拉提;张晶晶
2.放牧和围封对藏北高寒草原紫花针茅群落生物量分配及碳、氮、磷储量的影响[J], 洪江涛;吴建波;王小丹
3.青藏高原高寒草地优势禾草-紫花针茅内生真菌分离和鉴定 [J], 鲍根生;李春杰
4.天山高寒草原紫花针茅群落特征的时间变化 [J], 蔺莹;欧其尔;冯丽;满良;马成仓
5.藏北高寒草原紫花针茅根系碳氮磷生态化学计量学特征 [J], 洪江涛;吴建波;王小丹
因版权原因,仅展示原文概要,查看原文内容请购买。
放牧对藏北紫花针茅高寒草原植物群落特征的影响

放牧对藏北紫花针茅高寒草原植物群落特征的影响段敏杰;高清竹;万运帆;李玉娥;郭亚奇;旦久罗布;洛桑加措【摘要】基于放牧试验,研究了不同放牧强度下藏北地区紫花针茅(Stipa purpurea)高寒草原植物群落特征的变化规律.结果表明,随着放牧强度的增强,植物群落盖度、地上生物量均呈现显著降低的趋势;紫花针茅等禾草类植物的重要值逐渐降低,莎草类中青藏苔草(Carex moorcroftii)、牲畜不喜食的杂类草及有毒有害植物均有增加的趋势;如果持续过度放牧,植物群落表现出由紫花针茅等禾草为建群种的草地型向青藏苔草(Carex moorcroftii)、杂类草(Herbarum variarum)草地型过度的趋势;在中度放牧强度下,紫花针茅高寒草原α物种多样性达到最高水平,而继续增强放牧强度,则造成各项指标的迅速降低.【期刊名称】《生态学报》【年(卷),期】2010(030)014【总页数】9页(P3892-3900)【关键词】放牧强度;群落特征;紫花针茅草原;α多样性;中度干扰假说【作者】段敏杰;高清竹;万运帆;李玉娥;郭亚奇;旦久罗布;洛桑加措【作者单位】中国农业科学院农业环境与可持续发展研究所,北京,100081;农业部农业环境与气候变化重点实验室,北京,100081;中国农业科学院农业环境与可持续发展研究所,北京,100081;农业部农业环境与气候变化重点实验室,北京,100081;中国农业科学院农业环境与可持续发展研究所,北京,100081;农业部农业环境与气候变化重点实验室,北京,100081;中国农业科学院农业环境与可持续发展研究所,北京,100081;农业部农业环境与气候变化重点实验室,北京,100081;中国农业科学院农业环境与可持续发展研究所,北京,100081;农业部农业环境与气候变化重点实验室,北京,100081;西藏自治区那曲地区草原站,那曲,852100;西藏自治区那曲地区草原站,那曲,852100【正文语种】中文高寒草原类草地在藏北高原广泛分布,其面积占全国草地总面积的10.6%[1],它不仅是亚洲中部高寒环境中典型的生态系统之一,而且在世界高寒地区也极具代表性[2]。
《2024年降水量变化对大针茅群落水分利用的影响》范文

《降水量变化对大针茅群落水分利用的影响》篇一一、引言大针茅群落是我国北方草原生态系统的重要组成部分,其生长和分布受到气候条件,特别是降水量的影响。
近年来,随着气候变化日益明显,降水量在不同地域和时间段的分布出现明显的变化,对生态环境产生重大影响。
本篇论文主要探讨了降水量变化对大针茅群落水分利用的影响。
二、研究背景与目的降水量是大自然中重要的水循环因素之一,直接影响植物生长的规律。
而大针茅作为一种多年生草本植物,在维护草地生态系统的稳定性和可持续性中发挥着重要作用。
因此,研究降水量变化对大针茅群落的影响,有助于我们更好地理解气候变化对草地生态系统的影响机制,为生态保护和恢复提供科学依据。
三、研究方法本研究采用文献分析、实地观察和数据分析等方法,综合研究降水量变化对大针茅群落水分利用的影响。
在实验设计中,我们选择了多个具有不同降水量变化的区域,并对大针茅群落进行定期的监测和记录。
四、降水量变化对大针茅群落的影响1. 生长状况随着降水量的变化,大针茅的生长状况也发生了明显的变化。
在降水量增加的地区,大针茅的生长更加旺盛,叶绿素含量更高,植物更加繁茂。
相反,在降水量减少的地区,大针茅的生长受到了明显的抑制。
2. 水分利用效率降水量变化对大针茅的水分利用效率也有显著影响。
在降水量增加的条件下,大针茅能够更有效地利用水分,提高其生物量和产量。
然而,在降水量减少的情况下,大针茅的水分利用效率会降低,植物可能会进入休眠状态以保存水分。
3. 群落结构随着降水量的变化,大针茅群落的结构也发生了变化。
在降水量增加的地区,大针茅群落的丰富度和多样性增加,而小植物和草本动物的数量也相应增加。
相反,在降水量减少的地区,大针茅群落的丰富度和多样性可能会降低。
五、结论与建议本研究表明,降水量变化对大针茅群落的水分利用有显著影响。
在降水量增加的条件下,大针茅的生长更加旺盛,水分利用效率更高,群落结构更加丰富和多样。
然而,在降水量减少的情况下,大针茅的生长和水分利用效率会受到抑制。
《2024年降水量变化对大针茅群落水分利用的影响》范文

《降水量变化对大针茅群落水分利用的影响》篇一一、引言降水量是决定植物群落分布和生存的重要因素之一。
大针茅群落作为我国北方草原的重要植被类型,其生长和繁衍与降水量密切相关。
随着全球气候变化,降水量变化成为影响大针茅群落生存和发展的重要因素之一。
本文旨在探讨降水量变化对大针茅群落水分利用的影响,以期为保护和合理利用大针茅群落提供科学依据。
二、研究区域与方法本研究选取了我国北方具有代表性的大针茅群落作为研究对象,运用实验方法和气象数据分析,综合探讨了降水量变化对大针茅群落水分利用的影响。
研究方法包括:文献综述、实地调查、实验设计和数据分析等。
三、大针茅群落的水分利用特点大针茅群落是一种典型的旱生植物群落,其生长和繁衍主要依赖于地下水和地表水的供应。
在干旱和半干旱的北方地区,大针茅群落的水分利用策略主要是通过根系深扎和叶面积减小来减少水分蒸发,从而维持其生存和生长。
四、降水量变化对大针茅群落的影响(一)降水量的变化对大针茅群落的生理影响随着降水量的减少,大针茅植物为了维持其生存和生长,必须采取更多的策略来应对水分的减少。
首先,植物的叶面积会减小,以减少水分的蒸发。
其次,植物的根系会向更深的地方延伸,以寻找更多的水分资源。
这些生理适应性调整使得大针茅群落在干旱条件下具有一定的抗逆性。
(二)降水量变化对大针茅群落结构的影响随着降水量的变化,大针茅群落的结构也会发生变化。
当降水量减少时,群落的密度和物种丰富度会降低,群落的生物量也会随之减少。
同时,不同物种在群落中的分布也会发生变化,一些适应干旱环境的物种可能会逐渐取代其他物种成为优势种。
(三)降水量变化对大针茅群落生态功能的影响大针茅群落作为生态系统的重要组成部分,其生态功能包括土壤保持、水源涵养、碳汇等。
随着降水量的变化,这些生态功能也会受到影响。
例如,当降水量减少时,土壤的侵蚀可能会加剧,水源涵养能力可能会降低,从而影响生态系统的稳定性。
五、结论与建议本研究表明,降水量变化对大针茅群落的水分利用、群落结构和生态功能具有显著影响。
《2024年降水量变化对大针茅群落水分利用的影响》范文

《降水量变化对大针茅群落水分利用的影响》篇一一、引言在全球气候变化的大背景下,降水量变化成为影响陆地生态系统结构和功能的关键因素之一。
大针茅作为典型的草原群落之一,在生态系统中扮演着重要的角色。
本文旨在探讨降水量变化对大针茅群落水分利用的影响,以揭示气候变化背景下大针茅群落的响应机制及适应性策略。
二、大针茅群落及其水分利用特点大针茅群落是一种以大针茅为主要种群的草原生态系统,具有独特的水分利用策略。
在生长季节,大针茅通过深根系吸收土壤中的水分,并在体内进行有效的水分利用和调节,以维持其正常的生长和繁殖。
大针茅的叶片具有较高的保水性,能够在干旱条件下保持较高的水分含量,以应对环境中的水分波动。
三、降水量变化对大针茅群落的影响1. 降水量减少的影响当降水量减少时,大针茅群落面临更为严峻的干旱条件。
由于土壤水分的减少,大针茅的根系需要更深入地寻找水源,这可能导致其生长受到抑制。
同时,大针茅的叶片保水性虽然能够在一定程度上缓解干旱的影响,但长期的水分不足仍可能导致其叶片出现萎缩、黄化等现象,甚至导致植株死亡。
2. 降水量增加的影响相反,当降水量增加时,虽然大针茅群落能够获得更多的水分供应,但过量的降水也可能带来负面影响。
过多的水分可能导致土壤中氧气含量降低,影响大针茅根系的呼吸和生长。
此外,过量的降水还可能引发洪涝等灾害,对大针茅群落的生长和繁殖造成不利影响。
四、大针茅群落的适应性策略面对降水量的变化,大针茅群落展现出了一定的适应性策略。
一方面,大针茅通过调整其根系分布和深度,以适应土壤中水分的分布和变化。
另一方面,大针茅的叶片结构和保水性也具有一定的可塑性,能够在一定程度上缓解水分波动对其造成的影响。
此外,大针茅群落还可能通过种间竞争和种间协作等方式,与其他植物种群共同应对环境的变化。
五、结论综上所述,降水量变化对大针茅群落的水分利用具有重要影响。
在降水量减少的情况下,大针茅需要更深入地寻找水源,以维持其生长和繁殖。
《2024年降水量变化对大针茅群落水分利用的影响》范文

《降水量变化对大针茅群落水分利用的影响》篇一一、引言降水量是决定植物群落分布、生长和生存的关键因素之一。
大针茅作为我国北方地区典型的植物群落之一,其生长和繁衍也受到降水量的直接影响。
随着全球气候变化,降水量变化成为了一个不可忽视的生态问题。
因此,研究降水量变化对大针茅群落水分利用的影响,对于理解大针茅群落的生态适应性及应对未来气候变化具有重要意义。
二、大针茅群落概述大针茅是一种适应力强、耐旱性强的草本植物,是我国北方地区主要的草原植被之一。
大针茅群落由大针茅以及其他相关的草本植物、昆虫和微生物等组成,是生态系统中的重要组成部分。
大针茅群落的水分利用主要依赖于自然降水,其生态适应能力在应对环境变化中起到了关键作用。
三、降水量变化对大针茅群落水分利用的影响(一)影响水分循环降水量的变化直接影响到大针茅群落的水分循环。
在干旱和半干旱地区,大针茅群落主要依赖自然降水进行水分补充。
当降水量减少时,大针茅群落的水分来源受到限制,植物生长受阻,进而影响到整个群落的生态平衡。
相反,当降水量增加时,虽然能够为植物提供更多的水分来源,但也可能导致土壤水分过多,影响植物的正常生长。
(二)影响植物生长降水量变化对大针茅群落的另一个重要影响是植物生长。
当降水量减少时,大针茅的生长速度减缓,生物量减少。
而当降水量增加时,虽然短期内能够促进植物生长,但长期来看,过多的水分可能导致土壤养分流失,反而对植物生长产生不利影响。
此外,不同季节的降水量变化也会对大针茅的生长产生不同的影响。
(三)影响群落结构降水量变化还会影响大针茅群落的结构。
当降水量减少时,大针茅群落的物种多样性可能会降低,优势种更加明显。
而当降水量增加时,可能会吸引更多喜湿性的植物种类进入群落,改变群落的结构。
此外,不同地区的降水量变化也可能导致大针茅群落的地理分布发生变化。
四、应对策略与建议为了应对降水量变化对大针茅群落的不利影响,我们可以采取以下策略和建议:1. 加强生态保护和恢复:通过植树造林、草原恢复等措施,提高大针茅群落的生态稳定性,增强其应对环境变化的适应能力。
干旱及复水对‘紫叶红花芋’干物质及氮、磷、钾积累和分配特征的影响

山地农业生物学报39(5):38~44,2020JournalofMountainAgricultureandBiology干旱及复水对‘紫叶红花芋’干物质及氮、磷、钾积累和分配特征的影响周竣宇1,李 慧1,欧珍贵2,张文娥1(1.贵州大学 农学院,贵州 贵阳 550025;2.贵州省亚热带作物研究所,贵州 兴义 562400)摘 要:以贵州主栽品种‘紫叶红花芋’为材料,通过盆栽试验,研究了干旱及复水条件下,干物质以及N、P、K积累及分配的变化规律,以期为‘紫叶红花芋’干旱条件下的高效栽培提供理论依据。
结果表明,干旱胁迫显著降低了‘紫叶红花芋’根茎、茎和叶中干物质积累及植株对N、P、K的吸收和积累,增加了根中干物质积累及分配。
干旱条件下,产量器官根茎的矿质元素和干物质积累受影响最大;3种矿质营养中,N的降幅最大,K次之,P受影响最小。
干旱胁迫条件下,N和K在叶片和根系中分配比例增加,P在根、茎和叶中的分配比例均有增加,而三者在根茎中的分配却持续降低。
‘紫叶红花芋’叶片的复水恢复效果最好,但复水7d对根茎几乎无正向补偿效应。
关键词:紫叶红花芋;干旱;复水;吸收与分配中图分类号:S539 文献标识码:A 文章编号:1008-0457(2020)05-0038-07 国际DOI编码:10.15958/j.cnki.sdnyswxb.2020.05.006TheEffectsofDroughtStressandRehydrationonCharacteristicsofAccumulationandTranslo cationofDryMatterandMineralElementsof‘Ziyehonghuayu’ZHOUJunyu1,LIHui1,OUZhengui2,ZHANGWene1(1.AgricultureCollege,GuizhouUniversity,Guiyang,Guizhou550025,China;2.GuizhouInstituteofSubtrop icalCrops,Xingyi,Guizhou562400,China)Abstract:Inthispaper,the‘Ziyehonghuayu’(Cannaeduliscv‘Ziyehonghuayu’),atraditionalcultivarofediblecanna,wasstudiedtoexploreitstheaccumulationandtranslocationcharacteristicsofdrymatterandmineralelements(N,PandK)underdroughtstressandrehydration,inordertoprovidetheoreticalrefer enceforitsefficientwater savingcultivation.TheresultsshowedthatdroughtstresssignificantlysuppressedC.edulisgrowth,decreasedbothdrymatteraccumulationofrhizomes,stemsandleaves,andtheuptakeandaccumulationofN,PandK,whiledrymatteraccumulationintherootwasincreasedbydrought.Fordiffer entorgans,theeffectwasthegreatestimpactontherhizomes.Amongthreemineralelements,Nhasthelar gestdrop,followedbyK,andPhasthesmallestimpactunderdroughtstress.Atthesametime,thedistribu tionratioofNandKinleavesandrootswasincreasedbydrought,andthatofPinroots,leavesandstemswasallraisedaswell,butthedistributionratioofthethreemneralelementsinrhizomeswasdeclineddramat ically.Insummary,therehydrationeffectontheleafofC.eduliswasthebest,butrehydrationfor7dhadal mostnopositivecompensationeffectonrhizome.Keywords:Ziyehonghuayu;drought;rehydration;uptakeanddistribution收稿日期:2020 09 08;修回日期:2020 09 20基金项目:国家自然科学基金(31460371);芭蕉芋种质资源创新与利用(黔科合支撑[2018]2291);贵州省生物学一流学科建设项目(GNYL[2017]009);滇桂黔石漠化地区特色作物产业发展关键技术集成示范项目(SMH2019 2021)通讯作者:张文娥(1976—),女,博士,教授,主要从事园艺植物生理生态与资源评价研究,E mail:wezhang@gzu.edu.cn. 芭蕉芋(CannaedulisKer)是一种集粮食、能源、饲料于一体的多用途作物,在我国云、贵、桂等省广泛栽培[1]。
基于MaxEnt模型和GIS的青藏高原紫花针茅分布格局模拟_胡忠俊

基于MaxEnt 模型和GIS 的青藏高原紫花针茅分布格局模拟*胡忠俊1,2,3张镱锂1,2**于海彬1(1中国科学院地理科学与资源研究所陆地表层格局与模拟院重点实验室,北京100101;2中国科学院青藏高原地球科学卓越创新中心,北京100101;3中国科学院大学,北京100049)摘要气候变化对物种分布的影响是生物地理学研究的热点问题.本研究以广泛分布在青藏高原高寒草甸区域的建群种紫花针茅为研究对象,通过植物标本库查询和实地调查了解紫花针茅在青藏高原的分布情况,利用MaxEnt 模型模拟紫花针茅在青藏高原历史、当前及未来的分布趋势,分析紫花针茅各历史时期的分布格局,探讨了物种分布变化的产生原因.结果表明:青藏高寒草地的针茅属物种具有较高的物种多样性,代表性植物紫花针茅的主要分布区域为青藏高原腹地及沿喜马拉雅地区;紫花针茅的分布强烈地受最暖季降水量、最湿季降水量和年均降水量的控制.依据紫花针茅在末次冰期的分布格局以及青藏高原的地理地质特点,研究认为:冰期来临时,羌塘以南藏北核心区和喜马拉雅西部阿里地区是紫花针茅潜在分布的核心区,此处提供了比其他地区更适合紫花针茅生存的栖息地,这些区域成为紫花针茅的避难所,当前的紫花针茅都是从这些避难所迁移并扩展开来.生物避难所的存在有助于理解青藏高原高寒植被起源和分化的相关问题.关键词紫花针茅;MaxEnt 模型;生态位;分布格局;冰期避难所文章编号1001-9332(2015)02-0505-07中图分类号X171文献标识码ASimulation of Stipa purpurea distribution pattern on Tibetan Plateau based on MaxEnt modeland GIS.HU Zhong-jun 1,2,3,ZHANG Yi-li 1,2,Yu Hai-bin 1(1Key Laboratory of Land Surface Pat-tern and Simulation ,Institute of Geographical Sciences and Natural Resources Research ,ChineseAcademy of Sciences ,Bejing 100101,China ;2Center of Chinese Academy of Sciences Excellence in Tibetan Plateau Earth Sciences ,Beijing 100101,China ;3University of Chinese Academy of Sciences ,Beijing 100049,China ).-Chin.J.Appl.Ecol .,2015,26(2):505-511.Abstract :The impact of climate change on species distribution is a hot issue in biogeography re-search.This study utilized the constructive species Stipa purpurea as the research object ,which was widely distributed in alpine meadow of the Tibetan Plateau ,investigated its distribution in the Tibe-tan Plateau through the field survey and herbarium search ,and used MaxEnt model to simulate its historical ,current and future distribution trends to analyze its distribution pattern in each historical period and explore the cause of species distribution changes.Research results showed that diversity of Stipa species in alpine grassland of the Tibetan Plateau was high ,its main distribution area was the hinterland of the Tibetan Plateau and areas along the Himalaya ,and its distribution was strongly affected by precipitation of warmest quarter ,precipitation of wettest quarter and annual precipitati-on.According to the distribution pattern of S.purpurea in the Last Glacial Maximum ,and geograph-ical and geological features of the Tibetan Plateau ,this paper proposed that :North Tibet core area of South Qiangtang and Ali region of west Himalaya mountainous area were the core area of the potential distribution for S.purpurea ,since these regions could provide more suitable habitats for S.purpurea than other regions and be the refugia where the current S.purpurea was migrated and differentiated from.The presence of refugia may contribute to the understanding of related issues of the alpine plants ’origin and differentiation in the Tibetan Plateau.Key words :Stipa purpurea ;MaxEnt model ;niche ;distribution pattern ;glacial refugia.*中国科学院战略性先导科技专项(XDB03030500)和国家自然科学基金项目(41271068,41371120)资助.**通讯作者.E-mail :Zhangyl@igsnrr.ac.cn 2014-04-29收稿,2014-09-12接受.应用生态学报2015年2月第26卷第2期Chinese Journal of Applied Ecology ,Feb.2015,26(2):505-511DOI:10.13287/j.1001-9332.2015.0004紫花针茅(Stipa purpurea)是青藏高原及其毗邻区域的寒旱生、多年生丛生禾草,主要分布于中国西藏、青海、甘肃、四川、新疆等省区,其主要分布中心为阿里中部、羌塘高原、雅鲁藏布江中上游高山区域及藏南高原湖盆区[1].以该物种为建群种组成的紫花针茅群落是高原上高寒植被的最主要代表群落之一,为高寒草原中分布面积最大及最重要的植物类型[2-3].其分布区域的平均气温约0 3ħ,年降水量150 300mm;它对于寒冷、干旱及强风条件具有非常强的抵御能力[4].严酷的环境条件使得紫花针茅草原不仅稀疏低矮,而且生长发育节律较晚.紫花针茅粗蛋白质含量高,粗纤维少,营养价值较高,茎叶柔软,耐牧性强,是青藏高寒草原重要的牧草资源[5-6].紫花针茅分布区域辽阔,垂直幅度较高,除了形成大面积的单优势群落外,还常常与其他优势植物如青藏苔草(Carex moorcroftii)、小嵩草(Kobresia humilis)、羊茅(Festuca ovina)、变色锦鸡儿(Caraga-na versicolor)和金露梅(Potentilla fruticosa)等分别组成不同的群落[1,7].气候环境因素是影响生物分布的决定性因素,也是主导生物体内各种生理代谢和生物繁衍生息的关键因素[8].生物受气候变化驱动会产生生理活性和分布范围的变化.随着获取数据的丰富以及研究手段和运算能力的提高,生物分布研究朝着两个不同的方向发展:一方面,利用气候数据驱动植物群落、植被及整个生物圈的格局和过程成为宏观生态学的一个重要方面,同时人们也更加关注未来情景下的全球变化;另一方面,利用物种分布模型(spe-cies distribution models,SDM)研究物种隔离、物种形成及自然选择与非生物因子之间的关联具有很大潜力,利用该模型寻找物种的分布区、限制因子及生境条件等也成为保护区划定、入侵生物防治和生物多样性保护等方面的重要依据[8-9].有些学者分析美国西部植被分布与环境因子的关系后没有发现它们之间有明显的关联性[10-11],表明物种分布模型虽然是很实用的研究工具,但存在一定的争议性,需要不断改进,使其能够更准确地模拟复杂气候环境变化对物种及群落分布的影响.青藏地区高寒植物能够最先感受到地表气候的异常,因而它们的进化往往同地表气候变迁、生命历史及人类起源和进化相关联[12-13];另外,由于青藏高原的地质、地理特殊性及相对封闭的自然状态,其一直是进行生态学、地质学和进化生物学等研究的理想实验室.研究青藏高原生物的分布、多样性及其相互关系有助于理解生物在复杂的气候和环境条件下适应和扩散的过程和机制,并为该区域农牧业生产和管理提供理论依据和指导.为此,本文采用MaxEnt模型和ArcGIS10.1中的SDM工具箱对紫花针茅的分布和不同时期面积变化进行模拟和分析,得出其不同时期种群分布及群落面积变化趋势,并进一步推测其在青藏高原的可能避难所,为紫花针茅的合理保护和管理措施的科学制定提供依据.1研究数据与研究方法1.1物种分布模型的选择物种分布模型主要是利用物种的分布数据与环境数据,依据特定算法估计物种的生态位,并投射到景观中,结果可以解释为物种出现的概率、生境适宜度或物种丰富度等[14-15].物种及其多样性和丰富度的地理分布取决于人们是否真正理解和认识它们的生态位,对量化物种分布和丰富度的环境因子的测量是加强对物种生态位认识的重要一步[16-18].生态位是物种在群落内部位置的一个抽样特征,取决于促成物种进化的资源梯度在时空上的分布变化.基于生态位的大多数模型是靠一般植物、动物和生物多样性的研究发展而来[19-21].最大熵模型(MaxEnt)作为机器学习算法模型的一种,是目前应用最广泛、预测效果较好的物种分布模型之一[22-23].最大熵模型可对物种实际存在点位和相关环境数据的整合进行分析,通过每个栅格表达不同环境变量作用下的物种适宜性,即该栅格具有最适合该物种生存的气候及环境条件[22-24].与其他模型相比,只要依据存在数据,MaxEnt模型就能推测环境变量下的最大熵的可能分布,具有操作简单、运算速度快、预测结果好等优点[25-29].1.2模拟数据源通过野外调查和植物标本馆查询一共获取紫花针茅在青藏高原的285个点位分布信息:2013年8—9月中国科学院植物研究所对青藏高原高寒草原及高寒草甸区域的植被进行样方调查(图1),获取84个紫花针茅的点位数据;通过中国数字植物标本馆(CVH)查询获取201个紫花针茅点位数据,其中,来自中国科学院植物研究所标本馆的点位数据74个,来自中国科学院西北高原生物研究所标本馆的点位数据87个,来自中国科学院昆明植物研究所标本馆点位数据40个,物种在青藏高原的详细分布情况如图2所示,模拟所需要的末次间冰期(LIG,605应用生态学报26卷图1青藏高原野外调查路线图Fig.1Vegetation survey roadmap on Tibetan Plateau.Ⅰ:县County ;Ⅱ:样地Plot ;Ⅲ:调查路线Surveyline.图2紫花针茅物种在青藏高原分布点位及来源Fig.2Distribution points and source of Stipa purpurea in Tibetan Plateau.Ⅰ:实地调查Field survey ;Ⅱ:中国科学院植物研究所标本馆He-barium of Botony Institute ,CAS ;Ⅲ:昆明植物所标本馆Hebarium ofKunming Institute of Botony ,CAS ;Ⅳ:西北高原所标本馆Hebarium of Northwest Institute of Plateau Biology ,CAS.120 140ka )、末次冰期(LGM ,21ka )、当前(2000年)以及未来(2080年)4个时期的气候变量数据从WorldClim 数据库(www.worldclim.org )下载.将气候数据和紫花针茅分布点位数据输入MaxEnt 3.3模型中进行分布模拟及验证[30].1.3模型验证物种的现实分布状态有存在和不存在两种情况,一般用数字“0”表示不存在,“1”表示存在.这时模拟结果就变成了0 1之间连续变化的值,数值越高代表物种适应度越高,该区域越有可能存在此物种,数值越低,则正好相反.本研究把一套紫花针茅分布数据按照80/20的比例划分成两部分,分别用于模拟和验证.接受曲线(receive operating character-istic ,ROC )是目前物种分布模拟结果准确性验证最为广泛使用的指标,通过计算接受曲线下方的面积得到.ROC 的数值范围为0.5 1,ROC 越大表示预测越精确,ROC >0.9表示模拟结果拥有很高的精确度,一般来说,ROC >0.85的模拟验证结果就可以采纳.1.4物种分布面积变化的计算利用ArcGIS 10.1打开一个新的ArcMap 文件,双击“SDM ”工具→MaxEnt 工具→二进制SDM 图层的分布范围变化→“Distribution Changes Between Binary SDMs ”工具,进入工具界面进行操作.SDM 工具箱操作步骤如下:输入图层1:紫花针茅于LIG /LGM /Current 时期的SDM 图层;输入图层2:紫花针茅于LGM /Current /2080年的SDM 图层,建立栅格文件之间的关联,选择输出图层的位置,选择一个合适的等面积投影,最后以Tiff (.tif )格式的文件类型输出.2结果与分析2.1影响紫花针茅分布的主要环境变量本文利用19个生物气候指标以及高程数据信息(DEM )作为紫花针茅物种分布模拟的环境变量(表1).在20个环境因子中,年均降水量、最湿季度降水量和最暖季度降水量所占比重最大(约33% 35%),表明它们是限制紫花针茅物种在青藏高原分布的主要因子;其他变量如最湿月降水量、湿度季节性变化标准差、最冷月最低温度、年均温变化范围及DEM 虽然对于植物生长的贡献率相对较小,但在高寒恶劣多变的高原生态环境下,其对于预测紫花针茅的分布仍然具有重要意义,其中DEM 比较特殊,在不同模型中的贡献率差别很大,比如在GBM 中可以达到42.1%,但在MaxEnt 中却只有24%,表明各个模型对于环境变量的处理是不一样的,同时由于本研究的地理跨度较大,DEM 对各区域紫花针茅分布所起的作用也各不相同.紫花针茅区域分布与生长季降雨、年均降水量和最湿季度降水量有很强的相关性,在19个气候变量及海拔的贡献值中,它们排在前3位,贡献值都超过了30%,意味着紫花针茅的分布区主要处于青藏高原年降水量特别是生长季降水量较多的区域;而最干季度降水量及最冷季度降水量的贡献率很小(<10%),表明青藏高原腹地区域虽然冬季温度较低、降水量稀少,但这些区域的紫花针茅却广泛分布;模型预测的紫花针茅DEM 的分布区间在3500 5200m.表明紫花针茅对于水分的依赖性较强,对温度和海拔的容忍度较大.2.2紫花针茅在青藏高原分布格局的模拟对模型进行验证的结果显示,利用现有标本点位信息模拟的精度较低(ROC =0.778,图3),表明模型模拟的分布与验证的一致性较差;紫花针茅分布7052期胡忠俊等:基于MaxEnt 模型和GIS 的青藏高原紫花针茅分布格局模拟表1物种分布模拟的20个环境变量Table 120environmental variables in SDMs的最高适宜度只有0.7,低于利用采集样品和标本信息相结合进行分析得到的最高适宜度[MaxEnt 模型的最高适宜度为0.89,广义线性模型(GLM )、广义相加模型(GBM )和任意树模型(RF )的最高适宜度也大于0.85],表明利用这些标本获取的紫花针茅模拟分布准确性低于利用采集样品与标本数据相结合的模拟结果;从分布范围来看,标本样品的模拟分布区集中于包括藏北高原腹地、藏西南和青海湖周围的广阔区域(图3).这与紫花针茅在青藏高原分布的实地调查及植被分布图并不相符(图4),模拟的分布范围比紫花针茅的实际分布范围大,特别是图3基于标本馆分布数据利用MaxEnt 模拟紫花针茅的潜在分布及ROC 值Fig.3Potential distribution and ROC value of Stipa purpureamodeled by MaxEnt based on Herbarium distribution data.A :训练值Training data ;B :检测值Test data ;C :随机预测Randon prediction.下同The same below.Ⅰ:紫花针茅点位Stipa purpurea plot.青海湖周围区域.实际调研发现:紫花针茅在该区域属于零星分布,或者作为其他物种的伴生种存在,而不是作为建群种及优势种存在,因为该区域的易进入性,早期的植物学者在此区域调研的频率高于青藏高原其他地区,所以会让人误认为紫花针茅等物种是作为优势物种或者建群种在青海湖周围区域广泛存在.实地调查及利用植被图提取植物分布范围有助于人们纠正这样的认识误区.结合植物标本馆标本点位数据及野外植物采集的点位数据,对分布密集区进行疏化,剔除其中的无效及重复数据,基于模型MaxEnt 模拟紫花针茅4个历史时期的潜在分布,训练集和验证集的ROC 值均大于0.85,表明模型预测结果的准确性较好,再利用ArcGIS 10.1中“SDM ”工具对不同时期的紫花针茅物种潜在分布面积变化进行计算,结果(图4)表明,紫花针茅主要分布区域为羌塘无人区以南藏北高原图4紫花针茅群落在青藏高原的现实分布及本文推测的可能冰期避难所Fig.4Real distribution of Stipa purpurea communities in Tibetan Plateau and potential glacial refugia speculated by the research.Ⅰ:冰期避难所Glacial refugia ;Ⅱ:等花针茅群落分布区Distribution area of Stipa purpurea community.805应用生态学报26卷图5基于MaxEnt及GIS的末次间冰期(LIG)、末次冰期(LGM)、当前时期(Current)及2080年紫花针茅的潜在分布图、不同时期分布变化比较和当前时期的ROC值和各环境因子贡献率Fig.5Stipa purpurea potential distribution map in LIG,LGM,current period and2080,comparison of the distribution changes be-tween different periods,ROC value and contribution rate of each environment factors of current period based on MaxEnt model and GIS.图例中数字“-1”代表面积扩张的区域,0代表不存在物种分布的区域,1代表分布没有发生变化的区域,2代表面积收缩的区域-1stood for ar-ea expansion region,0stood for“no-species”region,1stood for region distribution was not changed,2stood for area contraction region.Ⅰ:无变量No variable;Ⅱ:单变量Only variable;Ⅲ:全部变量All variables.腹地(包括班戈县、双湖县、申扎县和尼玛县等地)和以阿里地区为中心的西喜马拉雅地区.从4个历史时期物种分布变化来看,从末次间冰期到当前时期,紫花针茅适宜性栖息地分布范围总体上呈现退缩的趋势,但在局部区域的物种适宜性栖息地既有扩张也有退缩.比较末次间冰期和末次冰期,羌塘以南藏北高原腹地紫花针茅种群适宜栖息地面积发生退缩,而喜马拉雅地区紫花针茅种群适宜性面积向东西两方向同时扩展,藏南湖盆区及雅鲁藏布江沿江区域出现紫花针茅的可能性增加;比较末次冰期和当前时期物种的分布范围,紫花针茅在青藏高原的适宜栖息地发生了不小变化,羌塘以南藏北高原腹地紫花针茅适宜性栖息地有扩大现象,喜马拉雅地区适宜性栖息地东西方向发生退缩,南北宽度也有明显缩减;比较当前时期和2080年,喜马拉雅区域的紫花针茅适宜栖息地面积有所扩展,羌塘以南藏北高原腹地种群适宜性区域面积有所缩减,但变化均不大,表明在80年较短的时间间隔内,紫花针茅在青藏高原的分布没有发生显著变化,呈现相对稳定的状态.有研究指出,青藏高原未来会呈现干旱化趋势[31],意味着在更远的将来,适宜紫花针茅生长的适宜栖息地面积会因为降水量的减少而减少,紫花针茅在青藏高原的分布有可能发生进一步萎缩.3讨论3.1物种分布模拟准确性的影响因素以紫花针茅标本经纬度存在点位进行物种分布模拟的精度比利用采集的样品点位和标本存在点位相结合进行模拟的精度低,因为这些标本点年代不统一、分布较分散、经纬度信息不够明确,很多需要通过Google Earth进行点位确认,因而存在一定的经纬度误差,而有些采样点的原有物种在采样后几十年几乎消失不见,该物种的环境变量也产生了较大变异,并可能会掩盖存在于标本采集地的分布信息.利用这些环境变量和物种信息进行模拟,会削弱模型对于该物种生态位的界定,并直接导致物种分布模型模拟能力和精度的下降.标本数据的精度对于模拟结果的影响非常大.如果样品数据不正确,很可能给整个模型传递错误的信息,从而产生错误的9052期胡忠俊等:基于MaxEnt模型和GIS的青藏高原紫花针茅分布格局模拟模拟结果.对于模型验证来说,该信息会导致模型精度的降低.由于模型的调参和验证是同时进行的,样品点位数据会交替用于调参和验证,因而必然会对整个模型及其精度产生不利影响.3.2物种分布与群落分布的比较影响物种分布的因素很多,除了受气候因素影响外,种间作用、微地形及其引起的局地小气候也会对物种分布产生影响.由于不同物种在群落中的地位和作用不同,其所采取的生存策略也会有所不同.对于紫花针茅而言,由于该物种在群落中的分布不能完全由群落所代表,所以取样通常不能完全覆盖到该群落的所有分布区,图4显示的是利用1ʒ1000000青藏高原植被图提取的紫花针茅群落在青藏高原的现实分布,可以明显看出紫花针茅群落与物种点位分布范围的差异.模拟精度主要取决于物种在群落的未分布区域能否通过其他预测变量检测出来.因为紫花针茅分布的限制因子往往不都是其群落的限制因子,例如由于竞争排斥作用产生的生物因素以及该物种及其敏感的环境因素,这些限制因子都需要在预测变量中体现;通过以往对群落构建和演替机制的分析可知,群落基本上都是受气候因素的影响[32].因此,利用气候变量可以较精确地实现对群落的模拟,这一点在BIOME模拟平台和其他生物地理模型中已经有不少相关研究[33-34].而物种分布模型的统计特征表明,大部分模型同样也可以用于生态群落或植被的分布模拟[35-36],模拟结果的可靠性主要取决于数据来源的代表性和准确性.3.3紫花针茅在青藏高原冰期避难所的推测生物避难所作为生物体或生物群落不利气候条件下退守的栖息地,具有时间和空间的持续性和地理范围的最大收缩性.在不利的气候变化影响下,可有限度地减轻其范围内生物种的生存困难[37-38].有研究对高山嵩草(Kobresia pygmaea)在青藏高原的冰期避难所推测认为:它们可能位于藏东南川藏交接区域和横断山区[39].利用最大熵模型及ArcGIS得到的紫花针茅4个时期适宜性栖息地的位置及其面积变化可以推测得知:羌塘以南藏北核心区域及喜马拉雅山西部阿里地区是紫花针茅的两个可能避难所(图4).羌塘以南藏北高原腹地处于唐古拉山脉和冈底斯山脉之间,在冰期来临的时候,由于山脉阻挡和众多湖泊对气候的调节,局部区域形成相对独立的小气候,这些区域提供了比高原其他地区更适合紫花针茅生存的栖息地,紫花针茅具有耐寒耐旱的特点,会比其他物种更不容易达到生存极限,其他物种都因为冰期来临而灭绝后,少数如紫花针茅等物种得以存活;而阿里地区由于喜马拉雅造山运动,使当地层次分明地存在着从寒带到热带的各种生物,在多次地质变动后,这里实际上成为物种数量庞大的生物避难所,多种生物在这种过渡环境中保存下来,紫花针茅也不例外,在漫长的末次冰期中维持在该地的持续存在,并在气候条件变好的时候扩展开来.致谢感谢中国科学院植物研究所郭柯研究员、内蒙古大学赵利清教授和中国科学院西北高原生物研究所陈世龙馆长在青藏高原野外植物调查和紫花针茅点位数据获取过程中提供的支持和帮助.参考文献[1]Wu Z-Y(吴征镒).Flora of China.Beijing:Science Press,1987(in Chinese)[2]Chen G-X(陈桂香),Shao Y-L(邵业立),Wang Z-L (王周龙),et al.Analysis of relationship between Stipapurpurea and site factor based on number level.Grass-land of China(中国草地),1988,55(1):53-59(inChinese)[3]Guo K(郭柯).Vegetation of Qinghai Hol Xil region.Journal of Plant Ecology and Geobotany(植物生态学与地植物学学报),1993,17(2):120-152(in Chi-nese)[4]Yue PP,Lu XF,YeRR,et al.Distribution of Stipa purpurea steppe in the Northeastern Qinghai-XizangPlateau(China).Russian Journal of Ecology,2011,42:50-56[5]China Encyclopedia(中国百科网).Grass Encyclopedia.[EB/OL](2008-01-09)[2014-04-28].http://www.chinabaike.com/article/396/cao/2008/200801091127533.html(in Chinese)[6]Zhou X-M(周兴民),Wang Z-B(王质彬),Du Q (杜庆).Vegetation of Qinghai.Xi’ning:QinghaiPeople Press,1987(in Chinese)[7]Sun H-L(孙鸿烈).The Formation and Evolution of Ti-betan Plateau.Shanghai:Shanghai Science and Techno-logy Press,1996(in Chinese)[8]Kozak KH,Graham CH,Wiens JJ.Integrating GIS-based environmental data into evolutionary biology.Trends in Ecology&Evolution,2008,23:141-148[9]Elith J,Leathwick JR.Species distribution models:Ecological explanation and prediction across space andtime.AnnualReview of Ecology,Evolution and Systemat-ics,2009,40:677-697[10]LentzRD,Simonson GH.Correspondence of soil proper-ties and classification units with sagebrush communitiesin southeastern Oregon.Soil Science Society of AmericaJournal,1987,51:1263-1271[11]Jensen ME,Simonson GH,Dosskey M.Correlation be-tween soils and sagebrush dominated plant communitiesof northeastern Nevada.Soil Science Society of America015应用生态学报26卷Journal,1990,54:902-910[12]Brochmann C,Brysting AK,Alsos IG,et al.Polyploidy in arctic plants.Biological Journal of the Linnean Socie-ty,2004,82:521-536[13]Koch MA,Kiefer C,Ehrich D,et al.Three times out of Asia Minor:The phylogeography of Arabis alpina L.(Brassicaceae).Molecular Ecology,2006,15:825-839[14]Peterson AT,Soberon J,PearsonRG,et al.Ecological Niches and Geographic Distributions.Princeton,NJ:Princeton University Press,2011[15]Yu H-B(于海彬),Zhang Y-L(张镱锂),Li S-C(李士成),et al.Predicting the dispersal routes of alpineplant Pedicularis longiflora(Orobanchaceae)based onGIS and species distribution models.Chinese Journal ofApplied Ecology(应用生态学报),2014,25(6):1669-1673(in Chinese)[16]Woodward FI.Climate and Plant Distribution.New York:Cambridge University Press,1987[17]Peterson AT,Soberon J,Sanchez-Cordero V.Conserva-tism of ecological niches in evolutionary time.Science,1999,285:1265-1267[18]WhittakerRJ,Willis KJ,FieldR.Scale and species richness:Towards a general,hierarchical theory of spe-cies diversity.Journal of Biogeography,2001,28:453-470[19]Rouget M,Richardson DM,Nel JL,et al.Mapping the potential ranges of major plant invaders in South Africa,Lesotho and Swaziland using climatic suitability.Diversi-ty and Distributions,2004,10:475-484[20]Ward DF.Modelling the potential geographic distribution of invasive ant species in New Zealand.Biological Inva-sions,2007,9:723-735[21]Poutsma J,Loomans AM,Aukema B,et al.Predicting the potential geographical distribution of the harlequinladybird,Harmonia axyridis,using the CLIMEX model.BioControl,2008,53:103-125[22]Phillips SJ,AndersonRP,SchapireRE.Maximum en-tropy modeling of species geographic distributions.Eco-logical Modelling,2006,190:231-259[23]Phillips SJ,Dudik M.Modeling of species distributions with Maxent:New extensions and a comprehensive eval-uation.Ecography,2008,31:161-175[24]Suarez-Seoane S,Morena ELG,Prieto MBM,et al.Maximum entropy niche-based modelling of seasonalchanges in little bustard(Tetrax tetrax)distribution.Ecological Modelling,2008,219:17-29[25]Graham CH,HijmansRJ.A comparison of methods for mapping species ranges and species richness.GlobalEcology and Biogeography,2006,15:578-587[26]Fitzpatrick MC,Gove AD,Sanders NJ,et al.Climate change,plant migration,and range collapse in a globalbiodiversity hotspot:The Banksia(Proteacea e)of West-ern Australia.Global Change Biology,2008,14:1337-1352[27]Elith J,Phillips SJ,Hastie T,et al.A statistical expla-nation of MaxEnt for ecologists.Diversity and Distribu-tions,2011,17:43-57[28]Zhang JP,Zhang YL,Liu LS,et al.Predicting potential distribution of Tibetan spruce(Picea smithiana)in Qo-molangma(Mount Everest)National Nature Preserve:Using maximum entropy niche-based model.ChineseGeographical Science,2011,21:417-426[29]CregoRD,Didier KA,Nielsen CK.Modeling meadow distribution for conservation action in arid and semi-aridPatagonia,Argentina.Journal of Arid Environments,2014,102:68-75[30]Elith J,Philips SJ,Hastie T.Maxent Software for Spe-cies Habitat Modeling.Version3.2.19.Princeton,NJ:Princeton University,2011[31]Wu S-H(吴绍洪),Yin Y-H(尹云鹤),Zheng D(郑度).Climate changes in the Tibetan Plateau during thelast three decades.Acta Geographica Sinica(地理学报),2005,60(1):3-11(in Chinese)[32]Rousseau DD,Duzer D,Etienne JL,et al.Pollen re-cord of rapidly changing air trajectories to the NorthPole.Journal of GeophysicalResearch:Atmosphere,2004,109:116-123[33]Prentice IC,Jolly D,Afanas’eva NB,et al.Mid-Holocene and glacial-maximum vegetation geography ofthe northern continents and Africa.Journal of Biogeog-raphy,2000,27:507-519[34]Thuiller W,Lafourcade B,EnglerR,et al.BIOMOD:A platform for ensemble forecasting of species distribu-tions.Ecography,2009,32:369-373[35]Ferrier S,Guisan A.Spatial modelling of biodiversity at the community level.Journal of Applied Ecology,2006,43:393-404[36]Olden JD,Joy MK,DeathRG.Rediscovering the spe-cies in community-wide predictive modeling.EcologicalApplications,2006,16:1449-1460[37]Hampe A,PetitRJ.Conserving biodiversity under cli-mate change:The rear edge matters.Ecology Letters,2005,8:461-467[38]Hu Z-J(胡忠俊),Zhang Y-L(张镱锂),Liu L-S(刘林山),et al.Refugia and their identification methods:A review.Chinese Journal of Ecology(生态学杂志),2013,32(12):3397-3406(in Chinese)[39]Wu L(吴良).Eco-geographical Distribution and Phylogenetic of Kobresia pygmaea Willd.on the Qinghai-Tibetan Plateau.PhD Thesis.Beijing:Institute of Geo-graphical Sciences and NaturalResourcesResearch,Chinese Academy of Sciences,2011(in Chinese)作者简介胡忠俊,男,1979年生,博士研究生.主要从事生物地理学、青藏高原建群种分布研究.E-mail:Huzj.11b@ igsnrr.ac.cn责任编辑杨弘1152期胡忠俊等:基于MaxEnt模型和GIS的青藏高原紫花针茅分布格局模拟。
青海湖地区紫花针茅型高寒草原群落特征变化

青海湖地区紫花针茅型高寒草原群落特征变化
王婷;史倩倩;祁得胜;陈克龙;杜岩功
【期刊名称】《草地学报》
【年(卷),期】2024(32)4
【摘要】气候变暖促进青藏高原高寒草原分布面积逐渐增加,但有关紫花针茅(Stipa purpurea)型高寒草原生产力与物种丰富度长期变化特征及其驱动因素尚不明确。
本研究连续13年观测青海湖地区高寒草地地上生物量、物种丰富度和盖度等指标,阐明了植被群落特征的长期变化规律,采用结构方程模型,解析了紫花针茅型高寒草原生产力和物种丰富度的主要影响因素。
结果表明,高寒草原地上生物量和物种丰富度均呈现较强的波动上升趋势,平均鲜重和物种数分别为215.41 g·m^(-2)和15,高物种丰富度促进生产力增加;高寒草原群落总盖度和平均高度呈现显著增加趋势。
紫花针茅型高寒草原地上生物量和总盖度分别受到年均气温和生产力的显著影响。
未来增温气候情景将会显著增加青海湖地区紫花针茅型高寒草原生物量,但降雨量增加会降低物种丰富度。
【总页数】6页(P1204-1209)
【作者】王婷;史倩倩;祁得胜;陈克龙;杜岩功
【作者单位】青海师范大学生命科学学院;青海湖国家级自然保护区管理局;中国科学院西北高原生物研究所
【正文语种】中文
【中图分类】S821.43
【相关文献】
1.天山高寒草原不同退化梯度紫花针茅群落结构特征研究
2.放牧和围封对藏北高寒草原紫花针茅群落生物量分配及碳、氮、磷储量的影响
3.青海湖地区紫花针茅草原群落特征
4.天山高寒草原紫花针茅群落特征的时间变化
5.放牧对藏北紫花针茅高寒草原植物群落特征的影响
因版权原因,仅展示原文概要,查看原文内容请购买。
藏北高寒草原紫花针茅根系碳氮磷生态化学计量学特征

藏北高寒草原紫花针茅根系碳氮磷生态化学计量学特征洪江涛;吴建波;王小丹【期刊名称】《山地学报》【年(卷),期】2014(032)004【摘要】植物碳(C)、氮(N)和磷(P)元素平衡是当前生物地球化学循环和全球变化生态学研究的热点和焦点.以往关于植物营养元素的研究多集中于植物叶片水平,而对植物根系元素的研究较少.为此,沿藏北高寒草原安多-那曲-班戈-申扎-尼玛-改则-革吉-噶尔样带,在34个样点采集优势物种紫花针茅(Stipa purpurea)根系样品,分析其C、N和P元素含量及其相互比值与环境因子的关系.研究结果显示:紫花针茅根系C、N和P含量变化范围分别为357.17 ~458.99 mg/g,6.49 ~ 16.42mg/g,和0.22 ~ 0.77 mg/g;紫花针茅根系N含量与全球植物根系平均值基本一致,但是其根系P含量低于全球平均值;紫花针茅根系C∶N∶P生态化学计量学特征受纬度、经度、生长季降水量、土壤磷含量的影响较为显著,而海拔、生长季均温和土壤氮含量对其影响则不显著.紫花针茅通过对高寒环境因子的长期适应,其根系C∶N∶P生态化学计量学特征已经形成了一定的空间分布格局.研究结果有助于揭示极端环境下植物对营养物质的分配和利用策略,为应对未来气候变化对高寒生态系统的影响提供参考.【总页数】8页(P467-474)【作者】洪江涛;吴建波;王小丹【作者单位】中国科学院水利部成都山地灾害与环境研究所,山地表生过程与生态调控重点实验室,四川成都610041;中国科学院大学,北京100049;中国科学院水利部成都山地灾害与环境研究所,山地表生过程与生态调控重点实验室,四川成都610041;中国科学院水利部成都山地灾害与环境研究所,山地表生过程与生态调控重点实验室,四川成都610041【正文语种】中文【中图分类】Q142【相关文献】1.放牧和围封对藏北高寒草原紫花针茅群落生物量分配及碳、氮、磷储量的影响[J], 洪江涛;吴建波;王小丹2.藏北紫花针茅高寒草原适宜放牧率研究 [J], 干珠扎布;郭亚奇;高清竹;段敏杰;万运帆;李玉娥;旦久罗布3.人为干扰对藏北高寒草原群落生物量及其碳氮磷含量特征的影响 [J], 鄢燕;马星星;鲁旭阳4.放牧对藏北紫花针茅高寒草原植物群落特征的影响 [J], 段敏杰;高清竹;万运帆;李玉娥;郭亚奇;旦久罗布;洛桑加措5.氮素添加对贝加尔针茅草原土壤团聚体碳、氮和磷生态化学计量学特征的影响[J], 李明; 秦洁; 红雨; 杨殿林; 周广帆; 王宇; 王丽娟因版权原因,仅展示原文概要,查看原文内容请购买。
茂兰喀斯特森林木本植物叶功能性状与其相互关系

《应用与环境生物学报》Chin J Appl Environ Biol Doi: 10.19675/ki.1006-687x.2020.09069茂兰喀斯特森林木本植物叶的功能性状及其相互关系熊玲龙翠玲**廖全兰薛飞贵州师范大学地理与环境科学学院贵阳550001摘要叶功能性状是连接植物与外界环境的重要桥梁,与植株资源获取与利用密切相关,能直观反映植物适应环境变化的生存对策。
为探讨原生性喀斯特森林植物叶片性状的适应性特征,以茂兰自然保护区不同地形(槽谷、漏斗、阴坡、阳坡)的木本植物为研究对象,通过测定叶片的叶鲜重(FW)、叶干重(DW)、叶厚度(LT)、叶面积(LA)、比叶面积(SLA)、叶干物质含量(LDMC)、叶绿素含量(Chl)和叶组织密度(LTD)8项叶性指标,研究叶功能性状之间的关系,并对不同生长型、不同地形木本植物叶功能性状进行比较。
结果显示:(1)茂兰喀斯特森林木本植物呈现出叶面积和比叶面积大、叶厚度及叶干物质含量低的性状组合,叶性状的变化范围不同,其中叶鲜重和叶干重的变异系数较大,叶绿素含量较稳定。
(2)叶功能性状之间的相关性好,比叶面积与叶鲜重、叶干重、叶厚度、叶绿素含量及叶干物质含量之间呈极显著负相关,叶鲜重对叶干重、叶面积、叶厚度及叶绿素含量表现为促进效应。
(3)不同生长型植物叶功能性状不同,常绿树种的叶厚度、叶干物质含量和叶绿素含量高于落叶树种,比叶面积则相反,落叶树种的叶干重数值较为集中,常绿树种的叶厚度和叶组织密度数值较为分散。
(4)不同地形木本植物叶功能性状存在显著差异,叶面积表现为阴坡>槽谷>阳坡>漏斗,比叶面积为阴坡>阳坡>漏斗,叶厚度则是阳坡>槽谷>阴坡。
乔木的叶干物质含量为阳坡>阴坡>槽谷>漏斗,灌木的差异性不显著。
本研究表明原生性喀斯特森林木本植物的LA、SLA较大而LDMC较小,叶功能性状间的相互作用强烈,植物采用不同的叶性策略来适应微生境。
紫花杧花期调控试验

紫花杧花期调控试验
唐晶;李现昌;杜德平;许建楷
【期刊名称】《果树学报》
【年(卷),期】1995(0)S1
【摘要】紫花杧成熟期集中在7月中旬至8月上旬。
为调节鲜果上市期,采用赤霉素(GA_3)和多效唑(PP_(333))进行花期调控。
赤霉素30mg·1^(-1)于冬季喷洒试
验株3次,抑制花芽分化。
次年3月土施多效唑5、10g·株^(-1)(以有效成分计),诱导花芽形成和开花。
紫花杧成熟期可推迟至10月中旬以后。
单株平均产量达10.9、17.5kg,品质良好。
试验株次年仍取得一定的产量。
【总页数】3页(P82-84)
【关键词】花期调控;花芽分化;果实品质;花芽萌发;花序数;有效成分;坐果率;抑制作用;多效唑;联合公司
【作者】唐晶;李现昌;杜德平;许建楷
【作者单位】广东广州市国营新塘农工商联合公司;华南农业大学园艺系
【正文语种】中文
【中图分类】S667.7
【相关文献】
1.紫花勒杜鹃花期调控技术初探 [J], 罗泽榕;罗少郁;陈劲晖
2.'桂热杧82号'花期短截花枝试验 [J], 陈千付;韦淑丹;钟勇;黄杰;王烨熔;黄慧俐;杨谨瑛;蒋强;曹元鑫
3.枣树花期调控试验分析 [J], 张黎;彭刚;梁刚;唐燕
4.杧果品种紫花芒砧木比较试验 [J], 黄桂香;徐炯志;李荣耀;黄寿山
5.应用植物生长调节剂调控杧果花期及保果初探 [J], 赵文振;沈雪玉
因版权原因,仅展示原文概要,查看原文内容请购买。
牦牛藏羊混合放牧对高寒草地紫花针茅功能性状的影响

指标(植株高度、单株重、叶厚、比叶面积)进行测定,旨在明确放牧方式对紫 花 针 茅 的 影 响。 结 果 显 示:不 同 放 牧 方
式对紫花针茅碳含量、氮含量、碳氮比、叶厚无显著影响,对 植 株 高 度、单 株 重、比 叶 面 积 影 响 显 著 (高度、单株重、比叶 面 积 受 放 牧 影 响 最 显 著(P<0.05),与 禁 牧 相 比,分 别 降 低
SUN Cai-cai,FENGBin,DONG Quan-min,LIU Wen-ting,SHIGuang,LIU Yu-zhen, ZHANG Xiao-fang,YU Yang,ZHANG Chun-ping,YANG Xiao-xia*
(AcademyofAnimalScienceandVeterinary Medicine,QinghaiUniversity,QinghaiProvincialKeyLaboratoryofAdaptiveManagement onAlpineGrassland,Xining,QinghaiProvince810016,China)
(青海省畜牧兽医科学院,青海大学,青海省高寒草地适应性管理重点实验室,青海 西宁 810016)
摘要:植物功能性状是植物响应草地变化的主要指标,对于了解草地发展进程 具 有 重 要 意 义。 本 试 验 对 牦 牛、藏 羊
混合放牧(不同放牧方式)下对紫花针茅(Stipapurpurea)叶片元素含量(碳含量、氮含量、碳氮比)及 种 群 表 型 性 状
为显著。
关 键 词 :高 寒 草 地 ;牦 牛 藏 羊 混 合 放 牧 ;紫 花 针 茅 ;功 能 性 状
中 图 分 类 号 :S812
文 献 标 识 码 :A
文 章 编 号 :1007-0435(2021)07-1477-07
青藏苔草和紫花针茅形态结构及其生态适应性的开题报告

青藏苔草和紫花针茅形态结构及其生态适应性的开题报告摘要:青藏高原是全球高原环境的代表地区,其独特的生态系统为研究高原植物形态结构与生态适应性提供了重要条件。
本文以青藏苔草和紫花针茅为研究对象,分析其形态结构及生态适应性,探究其适应高原环境的生存策略,为揭示高原植物的适应机制提供理论依据。
关键词:青藏苔草;紫花针茅;形态结构;生态适应性;高原植物一、研究背景及意义随着全球气候变化和环境污染的加剧,对高原生态系统的保护和恢复已经成为全球关注的热点问题。
高原植物是高原生态系统的重要组成部分,其形态结构和生态适应性对于理解高原植被的生态与进化过程具有重要意义。
青藏高原是世界上最大的高原区域之一,其独特的自然环境和植被多样性吸引了众多学者的关注。
青藏苔草和紫花针茅作为高原植物中的代表性物种,在高原植物的形态结构和生态适应性方面具有重要的研究价值。
二、研究内容和方法本研究的主要内容是对青藏苔草和紫花针茅的形态结构和生态适应性进行分析和比较,并探究其适应高原环境的生存策略。
研究方法主要包括野外调查和实验室分析。
野外调查主要采用样带法,对青藏高原不同海拔高度和不同生境类型中的青藏苔草和紫花针茅进行调查,记录其群落特征、生长状况和分布情况等信息;实验室分析主要包括形态结构、光合作用特性和生理生化指标等方面的分析,采用常规的组织学、生化分析和光谱技术等实验手段进行。
三、研究预期成果本研究预期获得以下成果:(1)对青藏苔草和紫花针茅的形态结构和生态适应性进行深入分析,揭示高原植物的适应机制;(2)比较两种植物的差异和相似之处,探究高原植物的多样性及其影响因素;(3)为青藏高原植物保护和恢复提供理论依据。
四、研究进展目前,本研究已经完成了野外调查和样品采集工作,进入实验室分析阶段。
初步结果显示,青藏苔草和紫花针茅在形态结构、生长状况和分布情况等方面存在明显的差异,但在适应高原环境的生存策略方面具有一定的相似性。
进一步的实验分析将为对高原植物的形态结构与生态适应策略的理解提供更为详细的信息。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
植物生态学报 2012, 36 (2): 136–143 doi: 10.3724/SP.J.1258.2012.00136 Chinese Journal of Plant Ecology 西藏紫花针茅叶功能性状沿降水梯度的变化胡梦瑶1,2张林1*罗天祥1沈维1,21青藏高原环境变化与地表过程重点实验室, 中国科学院青藏高原研究所, 北京 100085; 2中国科学院研究生院, 北京 100049摘要植物叶功能性状与环境因子的关系是近10年来植物生态学的研究热点。
该文以广泛分布于青藏高原干旱、半干旱草地的优势植物种紫花针茅(Stipa purpurea)为研究对象, 沿降水梯度(69–479 mm)系统测定了日土、改则、珠峰、当雄和纳木错5个调查地点紫花针茅比叶面积(SLA)、单位重量和单位面积叶氮含量(N mass, N area)、叶密度和厚度等叶功能性状以及土壤全氮含量等因子, 试图验证干旱胁迫地区同一物种内SLA-N mass关系沿降水梯度的策略位移现象是否具有普遍性, 并对是否出现策略位移现象提出可能的解释。
研究结果表明: 1) SLA和N mass与生长季温度和降水以及土壤全氮含量均没有显著关系, SLA与N mass的关系在干旱半干旱区(年降水/蒸发比< 0.11)与半湿润区(年降水/蒸发比> 0.11)之间并没有出现典型的位移现象; 2)叶密度是决定半湿润区SLA变化的主导因子, 而叶厚度则是干旱半干旱区SLA变化的控制因子, 两者与SLA均呈负相关, 随着温度增加或降水减少, 叶厚度增加而叶密度降低, 导致SLA随温度和降水变化不明显; 3)半湿润区的叶密度增加引起N area增加, 而干旱半干旱区的叶厚度增加并没有造成N area的显著变化, 导致N area沿降水梯度没有显著变化; 4)紫花针茅地上生物量与N area具有显著正相关关系, 表明N area的增加有助于提高植被生产力。
结果表明, 在干旱胁迫下, 植物通过增加叶厚度来维持不变的N area可能有助于保持与较湿润地区相似的光合生产和水分利用效率。
叶厚度和叶密度对比叶面积的相对影响在干旱半干旱区与半湿润区之间发生转变, 这为进一步检测高寒草地植被的水分限制阈值提供了新思路。
关键词叶密度, 叶氮含量, 叶厚度, 比叶面积, 紫花针茅, 策略位移Variations in leaf functional traits of Stipa purpurea along a rainfall gradient in Xizang, China HU Meng-Yao1,2, ZHANG Lin1*, LUO Tian-Xiang1, and SHEN Wei1,21Key Laboratory of Tibetan Environment Changes and Land Surface Processes, Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100085, China; and 2Graduate University of Chinese Academy of Sciences, Beijing 100049, ChinaAbstractAims Stipa purpurea is the dominant species in alpine arid and semi-arid grasslands on the Tibetan Plateau. Our objectives are to determine if this species exhibits a strategy shift in its specific leaf area (SLA) to nitrogen (N) concentration relationship along a rainfall gradient and to detect possible effects of environmental factors on re-lated leaf traits.Methods We investigated variations in leaf traits of S. purpurea associated with climatic and soil factors along an east-west transect with a rainfall gradient (69–479 mm) but similar altitudes (4 300–4 700 m). Five locations from east to west are Damxung, Namco, Gêrzê, Mount Qomolangma and Rutog. We measured SLA, mass- and area-based leaf N concentration (N mass, N area), leaf density and thickness and soil total N along the transect. Important findings In pooled data, SLA and N mass varied little with the growing season mean temperature and precipitation and the soil total N concentration. The SLA-N mass relationship in S. purpurea did not shift between the semi-humid areas (ratio of rainfall to evaporation > 0.11) and the arid and semi-arid areas (ratio < 0.11), al-though there was a positive correlation between SLA and N mass across the five locations. Variation in SLA was mainly determined by leaf density in the semi-humid areas and by leaf thickness in the arid and semi-arid areas; both were negatively correlated with SLA. With increasing temperature or declining precipitation, leaf density de-creased and leaf thickness increased, leading to non-significant relationships between SLA and climatic factors. The increase of leaf density in the semi-humid areas was correlated with the increase of N area, but the increase of leaf thickness in the arid and semi-arid areas did not lead to change of N area, resulting in unchanged N area along the rainfall gradient. A positive correlation was detected between aboveground biomass and N area in S. purpurea, indi-cating that increased N area may increase plant productivity. Our findings suggest that alpine plants in arid and semi-arid areas may maintain a constant N area by increased leaf thickness in order to achieve a similar photosyn-——————————————————收稿日期Received: 2011-07-20 接受日期Accepted: 2011-12-16* 通讯作者Author for correspondence (E-mail: zhanglin@)胡梦瑶等: 西藏紫花针茅叶功能性状沿降水梯度的变化 137doi: 10.3724/SP.J.1258.2012.00136thetic productivity and water use efficiency compared to the relatively humid areas. The relative impacts of leaf density and leaf thickness on SLA shifted between the semi-humid areas and the arid and semi-arid areas, which may provide insight in detecting the threshold of water limitation in alpine grasslands.Key words leaf density, leaf nitrogen concentration, leaf thickness, specific leaf area, Stipa purpurea, strategy shift叶功能性状(如最大光合/呼吸速率、比叶面积和叶氮含量等)是植物为适应环境而表现出来的叶片水平功能特征。
研究这些叶性状的相互关系及其随环境梯度的变化规律有助于深刻理解植物为获得最大光合生产所形成的适应策略(Reich et al ., 1997; Wright & Westoby, 2002), 相关研究已成为当前植物生态学研究的热点(Wright et al ., 2005; He et al ., 2009)。