园艺植物丛枝菌根研究与应用(吴强盛著)思维导图
丛枝菌根真菌在园艺作物上的应用

丛枝菌根真菌在园艺作物上的应用1邹英宁,吴强盛*长江大学园艺园林学院,湖北荆州(434025)E-mail:wuqiangsh@摘要:丛枝菌根是土壤中的丛枝菌根真菌与植物根系结合的互惠共生体,能帮助植物吸收矿质营养和水分、促进植物生长、提高抗逆性、改善果实品质等。
提出了丛枝菌根真菌生产的技术流程,综述了丛枝菌根真菌在果树、蔬菜、花卉植物上的应用与效应。
关键词:丛枝菌根真菌;丛枝菌根;园艺作物;菌剂生产中图分类号:Q939.961. 引言菌根(Mycorrhizas)是一类与植物根系紧密结合互惠互利的联合体,其互惠互利表现在菌根通过其根系外的菌丝、根系内的丛枝及根内特殊的水分运输通道给寄主植物运送矿质营养和水分,而寄主植物将光合作用产生的碳水化合物通过物质流转运给菌根以维持其生长发育[1]。
菌根按照形态学分为三类:外生菌根(Ectomycorrhizas)、丛枝菌根(Endomycorrhizas)和内外生菌根(Ectoendomycorrhizas)[2]。
外生菌根指菌根真菌侵入到植物根系的皮层,在间隙里形成哈蒂氏网,大量的菌丝在根系外面形成一个菌套,主要与森林植物共生;丛枝菌根指菌根菌丝不仅侵入到根系皮层,而且还进入到细胞内部,形成丛枝(Arbuscules)结构,有的还在细胞间或者内部形成泡囊(Vesicles),在许多园艺作物如柑桔、桃、苹果、梨、番茄、西瓜、非洲菊、月季等都可以发现和找到这种结构;内外生菌根则同时具备外生菌根和丛枝菌根的特性,菌根菌丝在细胞间隙形成哈蒂氏网,根系表面形成菌套,菌丝在细胞内部也形成各种菌丝团,主要在一些松科和杜鹃花科植物存在。
目前的研究表明,在园艺作物上接种丛枝菌根真菌能够促进园艺作物的生长,增强园艺作物对矿质营养的吸收,提高抗逆性,改善水分代谢,提高果树和蔬菜的品质等[3]。
因此,在园艺作物根系上没有丛枝菌根的存在反而不正常[4],从而显示丛枝菌根在园艺作物上的重要性2. 丛枝菌根真菌菌剂的生产丛枝菌根真菌菌剂的生产是其应用于园艺作物的关键。
丛枝菌根真菌对限根栽培枳生长和根系构型的影响

丛枝菌根真菌对限根栽培枳生长和根系构型的影响作者:张艺灿赵思邹英宁吴强盛来源:《湖北农业科学》2014年第11期摘要:在限根栽培条件下,研究丛枝菌根真菌摩西球囊霉(Glomus mosseae)和隐类球囊霉(Paraglomus occultum)对枳(Poncirus trifoliata)生长和根系构型的影响。结果表明,2种菌根真菌在限根栽培下均显著提高了枳株高、茎粗、叶片数,以及一级、二级和三级侧根数,也增加了根系投影面积、表面积、体积和总长(主要是0~3 cm的根长),且G.mosseae的促进效果优于P.occultum。关键词:枳(Poncirus trifoliata);限根栽培;侧根;根系;丛枝菌根真菌中图分类号:S666;S718.81文献标识码:A文章编号:0439-8114(2014)11-2588-03Effects of Mycorrhizal Fungi on Growth and Root System Architecture of Trifoliate Orange under Root Restriction ConditionsZHANG Yi-can,ZHAO Si,ZOU Ying-ning,WU Qiang-sheng(College of Horticulture and Gardening, Yangtze University, Jingzhou 434025, Hubei, China)Abstract: The effects of mycorrhizal fungi including Glomus mosseae and Paraglomus occultum on growth and root system architecture of trifoliate orange under root restriction conditions were studied. Results showed that the two arbuscular mycorrhizal fungi significantly increased plant height, stem diameter, leaf number, the number of 1st, 2nd, and 3rd lateral roots, root surface area, root projected area, root volume, and root total length (mainly 0~3 cm size length). G. mosseae exhibited better effects than P. occultum did.Key words: citrus; root restriction; lateral root; root system; arbuscular mycorrhizal fungi基金项目:湖北省自然科学基金杰出青年人才基金项目(2012FFA001);长江大学优秀青年教师科研支持计划项目(cyq201324);长江大学大学生创新创业训练计划项目(2012350)植物根系通常具有非常多的生理功能,例如吸收、合成、贮藏、分泌和感知等,各种环境因子和栽培措施都会影响根系的功能。近年来在果树栽培中,限根成为一种重要栽培措施且被大量地推广[1]。限根指的是植株根系受到一定的空间限制,包括自然限根和人为限根。适当的采取限根措施可使果树地上部分的生长更容易向生殖生长方向转变,从而调节树体生长和改善树体营养的分配,可改善果实品质,目前已经在葡萄、苹果、无花果、杨桃等果树上取得了很好的效果[2]。柑橘(Citrus L.)多为浅根性植物,在田间条件下通常需要与土壤中的丛枝菌根真菌(Arbuscular mycorrhizal fungi)共生建立丛枝菌根,根系协同丛枝菌根共同从土壤中吸收养分和水分[3]。许多研究已经显示,丛枝菌根真菌接种能够显著改善柑橘的树体生长、增强抵抗性、促进养分吸收等[3]。根系构型描述的是同一植物的不同级别根系在生长介质中的空间分布状况,受到各种生物和非生物因素的影响[4]。在限根栽培的条件下,根系生长发育受到限制,必然影响果树根系的空间分布,从而改变根系构型[5]。研究表明,丛枝菌根真菌侵染能够改善枳和红橘的根系构型[6,7]。目前,还不清楚丛枝菌根真菌在限根栽培下是否还具有改变根系构型的功能。为此,试验在限根条件下,研究2种丛枝菌根真菌对柑橘砧木枳(Poncirus trifoliata)根系构型的效应,以期为今后丛枝菌根真菌的应用和限根栽培下菌根的管理提供理论依据。1材料与方法1.1试验材料材料选择我国柑橘产区广泛使用的砧木枳(Poncirus trifoliata L. Raf.),丛枝菌根真菌选择摩西球囊霉(Glomus mosseae)和隐类球囊霉(Paraglomus occultum)。采集成熟的枳果实,取出种子,用1 mol/L NaOH搓洗10~20 min,70%乙醇表面消毒,然后用去离子水冲洗干净,用纱布裹好进行催芽处理。2012年2月采集长江大学柑橘园土壤,高压蒸汽(121 ℃)灭菌40 min,目的是杀死土壤中原有的丛枝菌根真菌。将已萌芽的枳苗移栽到塑料盆(上口径18 cm,下口径13 cm,高16.5 cm)中。在塑料盆中套一个直径6 cm、高22 cm、孔径37 μm的尼龙网袋。枳幼苗种植在尼龙网袋中,根系无法穿过。1.2试验设计试验采用单因素完全随机区组设计。设接种G.mosseae、P.occultum两个处理,以不接种丛枝菌根真菌的为对照(CK),每处理4次重复,1盆为1小区,每盆2株枳实生苗。播种的时候进行接种处理,接种时将50 g接种体(包含有孢子、丛枝菌根真菌感染的白三叶根系)放置在根袋内,以等量灭菌的接种体为对照。接种时根袋放置在塑料盆中央区域,根袋内建立的根系区包括根系和菌丝,根系只能在根袋内生长,从而形成了对植株的限根栽培。1.3测定方法接种140 d后,于2012年8月3日结束试验,测定植株株高、叶片数及茎粗并收获植株地上部分及根系。将植株从尼龙网袋中小心取出,尽量保证根系的完整。将根系用自来水清洗干净,清洗时动作要小心,尽量不伤害根系。用EPSON平板扫描仪扫描根系,获得根系图片用WinRHIZO根系分析仪进行根系构型参数的分析,包括根系直径、长度、投影面积、表面积和体积。然后将根系平铺在托盘中,快速统计各级侧根数。根系菌根侵染率采用曲莉苯蓝染色法[3]进行。1.4数据统计分析运用SAS(8.1)软件进行处理间差异显著性分析,采用Duncan新复极差法进行多重比较。2结果与分析2.1限根栽培下接种丛枝菌根真菌对枳植株生长的影响从表1可以看出,在限根栽培下,接种丛枝菌根真菌后枳根系菌根侵染率在30.3%~45.6%,且G.mosseae处理效果显著高于P.occultum的接种效果。接种丛枝菌根真菌G.mosseae和P.occultum后的植株株高、茎粗、叶片数均显著高于对照,而接种处理间差异不显著。2.2限根栽培下接种丛枝菌根真菌对枳根系构型的影响从表2可以看出,与对照相比,接种丛枝菌根真菌后根系总长度显著提高(P从表3可以看出,在限根栽培下接种处理明显改善了枳根系构型参数。接种处理除了对根系直径没有显著影响外,对其他的参数如根系投影面积、表面积和体积都有显著影响(P3小结与讨论丛枝菌根真菌可以与植株形成共生体,两者为互惠的关系,丛枝菌根通过扩大根系的吸收范围,从而帮助植物更好地吸收土壤中的水分和养分[3]。试验结果表明,在限根栽培下,接种丛枝菌根真菌提高了枳株高、茎粗和叶片数,且菌种间差异不显著。说明在限根栽培下,菌根真菌仍旧能够起到促进养分吸收的功能,从而促进植物的生长。根系构型的优劣能够明显决定植物对土壤养分吸收的多少。在正常栽培下,已经显示接种菌根真菌处理能够改善柑橘(包括枳在内)的根系构型[6,7]。当前的研究显示,在限根栽培下,G.mosseae和P.occultum接种都显著增加了枳根系表面积、投影面积、体积和总长,这与之前在非限根栽培下的研究结果[6,7]相符。试验结果进一步显示,接种处理对于根总长的改善主要体现在0~1、1~2和2~3 cm长度的根系总长上,对3~4 cm和4 cm以上长度的根总长没有显著效应,说明接种菌根真菌主要对细小的短根产生影响。此外,接种菌根真菌处理也显著改变了侧根数,表现为提高了一级、二级和三级侧根数量。这与Berta等[8]在韭菜和Schellenbaum等[9]在葡萄上得到的结果相符。然而,Isobe等[10]发现,在菜豆上接种丛枝菌根真菌显著减少了一级和二级侧根数,说明丛枝菌根真菌接种对侧根的效应可能依赖丛枝菌根真菌—寄主植物的双方选择,其机制还有待进一步的研究。参考文献:[1] 樊怀福,杜长霞,朱祝军,等.限根栽培对大果型番茄浙杂204植物生长、果实品质和产量的影响[J].浙江农林大学学报,2011,28(3):343-348.[2] 胡艳丽,毛志泉,沈向,等.限根栽培中根系垂直深度和土壤质地对苹果树体发育的影响[J].山东农业科学,2009(8):37-40.[3] 吴强盛.园艺植物丛枝菌根研究与应用[M].北京:科学出版社,2010.[4] 管建慧,刘克礼,郭新宇.玉米根系构型的研究进展[J].玉米科学,2006,14(6):162-166.[5] 岳龙,董凤祥,徐迎春.控根容器苗根系构型研究进展[J].世界林业研究,2008,21(6):31-35.[6] WU Q S,HE X H,ZOU Y N,et al. Arbuscular mycorrhizas alter root system architecture of Citrus tangerine through regulating metabolism of endogenous polyamines[J]. Plant Growth Regul,2012,68(1):27-35.[7] WU Q S, ZOU Y N, HE X H, et al. Arbuscular mycorrhizal fungi can alter some root characters and physiological status in trifoliate orange (Poncirus trifoliata L. Raf.) seedlings[J]. Plant Growth Regul,2011,65(2):273-278.[8] BERTA G, FUSCONI A, TROTTA A,et al. Morphogenetic modifications induced by the mycorrhizal fungus Glomus strain E3 in the root system of Allium porrum L.[J]. New Phytol, 1990,114(2):207-215.[9] SCHELLENBAUM L, BERTA G, RAVOLANIRINA F, et al. Influence of endomycorrhizal infection on root morphology in a micropropagated woody plant species(Vitis vinifera L.)[J]. Ann Bot,1991,68(2):135-141.[10] ISOBE K, MURAKAMI S,TATEISHI A,et al. Effects of arbuscular mycorrhizal fungi on the morphology of kidney bean root[J]. Jpn J Crop Sci,2002,71(1):91-95.。
八年级生物知识思维导图整理

八年级生物知识思维导图整理被子植物的有性生殖:精子受精卵种子的胚新一代植株有性生殖概念:有两性生殖细胞结合形成受精卵,再又受精卵发育成新个体的生殖方式特点:经过两性生殖细胞的结合,有性生殖的后代具有双亲的遗传特性概念:不经过两性生殖细胞的结合,由母体直接产生新个体的方式无性生殖特点:没有两性生殖细胞的结合,无性生殖产优点:加快繁殖速度,保持母体优良性状概念:剪去植物的枝条,将下部插入土无性生殖扦插举例:甘薯葡萄菊月季的应用概念:把一个植物同的芽或枝接在另一个植物体上,使之结合在一起。
嫁接方式:枝接和芽接关键:接穗和砧木的形成层紧密结合家蚕的生殖和发育发育过程:卵、幼虫、蛹、成虫变态发育:在由受精卵发育成新个体的过程中,幼体与成体的形态结构和生活习性差异很大的育过程发育过程完全变态发育过程:经过卵、幼虫、蛹、成虫四个时期其他昆虫的举例:菜粉蝶蝇生殖和发育不完全变态发育过程:经过卵、若虫、成虫三个时期举例:蝗虫蟋蟀蝼蛄螳螂等青蛙的生殖和发育过程:经过受精卵、蝌蚪、幼蛙、成蛙特点:有性生殖、特外受精,受精和发育过程离不开水环境,变态发育两栖动物的生殖和发育受环境条件的影响和限制卵壳和卵壳膜:保护作用胚盘:胚胎鸟卵的结构卵白和卵黄:为胚胎的发育提供营养物质和水分气室:为胚胎发育提供系带:固定卵黄鸟的生殖和发育过程:一般经过求偶、交配、筑巢、产卵、孵卵和育雏等几个阶段遗传:亲子间的相似性变异:亲子间及子代间的差异生物的性状:概念:生物体形态结构、生理和行为等特征的统相对性状:同种生物同一性状的不同表现基因控制生物的性状转基因生物性状表现是基因和基因:有遗传效应的DNA 片段基因DNA 和染色体 DNA :位于染色体上,主要的遗传物质,呈染色体:由蛋白质和DNA 组成,每种生物基因、DNA 、染色体和性状之间的关系基因经精子或卵现象:具有相对性状的纯种豌豆杂交,如高茎豌豆矮茎豌豆杂交,子一代都是高茎,子代杂种高茎豌豆自交,后代有高茎也有矮茎孟德尔豌豆杂交实验解释:控制相对性状的基因有显性D 和隐性d 之分,显性性状基因组成为DD和Dd ,隐性性状基因组成为dd.禁止近亲结婚:直系血亲和三代以内旁系血亲之间禁止结婚,生物的遗传与变异减少后代出现遗传病的机会常染色体(男女相同)22对。
丛枝菌根真菌对盐胁迫下园林植物再力花生长发育和光合特性的影响

㊀山东农业科学㊀2023ꎬ55(12):79~88ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2023.12.011收稿日期:2023-07-12基金项目:福建省科技厅引导性项目(2021N0002)ꎻ国家自然科学基金项目(31572088)ꎻ国家林业局森林公园工程中心开放课题(PTJH1500210)作者简介:刘振凡(1989 )ꎬ男ꎬ福建莆田人ꎬ硕士研究生ꎬ研究方向为植物生理㊁碳排放和园林工程ꎮE-mail:159****8176@163.com通信作者:刘文鑫(1983 )ꎬ男ꎬ福建厦门人ꎬ博士ꎬ副教授ꎬ研究方向为园艺园林植物生理ꎮE-mail:liuwxin_1983@126.com丛枝菌根真菌对盐胁迫下园林植物再力花生长发育和光合特性的影响刘振凡1ꎬ崔广强2ꎬ吴成华2ꎬ刘文鑫3(1.福建农林大学金山学院ꎬ福建莆田㊀351262ꎻ2.福建江夏学院ꎬ福建福州㊀350108ꎻ3.福建农林大学林学院ꎬ福建福州㊀350002)㊀㊀摘要:为提高园林湿地水生植物的耐盐性ꎬ本试验以再力花为材料ꎬ研究接种丛枝菌根真菌(arbuscularmycorrhizalfungiꎬAMF)对NaCl胁迫下植株生长发育㊁根系活力㊁渗透调节物质㊁抗氧化酶活性及叶片光合特性的影响ꎮ结果表明ꎬ接菌+盐胁迫组(T1)再力花的盐害指数显著低于不接菌+盐胁迫组(N-CK)ꎬ且T1组根系活力㊁株高㊁茎粗㊁地上地下干物质量均显著高于N-CKꎻ与N-CK相比ꎬ施加AMF(T1)显著提高可溶性蛋白含量ꎬ降低可溶性糖和脯氨酸含量ꎬ同时还显著降低丙二醛含量ꎬ显著提高抗氧化酶(SOD㊁CAT㊁POD)活性ꎻ随盐胁迫时间延长ꎬAMF处理(T1)叶片净光合速率㊁气孔导度㊁蒸腾速率及胞间CO2浓度均呈现上升趋势ꎬN-CK组则均呈下降趋势ꎮ综之ꎬ施加AMF可以有效缓解NaCl对再力花的渗透胁迫和氧化损伤作用ꎬ增强其对盐胁迫的抗性ꎬ从而促进植株生长和提高光合作用效率ꎮ关键词:丛枝菌根真菌ꎻ再力花ꎻ盐胁迫ꎻ生长发育ꎻ光合特性中图分类号:S682.1+61㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2023)12-0079-10EffectsofArbuscularMycorrhizalFungionGrowthandPhotosyntheticCharacteristicsofOrnamentalPlantThaliadealbataunderSaltStressLiuZhenfan1ꎬCuiGuangqiang2ꎬWuChenghua2ꎬLiuWenxin3(1.JinshanCollegeofFujianAgricultureandForestryUniversityꎬPutian351262ꎬChinaꎻ2.FujianJiangxiaUniversityꎬFuzhou350108ꎬChinaꎻ3.CollegeofForestryꎬFujianAgricultureandForestryUniversityꎬFuzhou350002ꎬChina)Abstract㊀ToimprovethesalttoleranceofaquaticplantingardenwetlandsꎬtheexperimentwascarriedoutwithThaliadealbataasmaterialtostudytheeffectsofinoculationofarbuscularmycorrhizalfungi(AMF)onitsplantgrowthanddevelopmentꎬrootactivityꎬosmoticregulationsubstancesꎬantioxidantenzymeactivityandleafphotosyntheticcharacteristicsunderNaClstress.TheresultsshowedthatthesaltdamageindexofT1group(inoculatingAMFundersaltstress)wassignificantlylowercomparedwiththeN ̄CKgroup(non ̄inocu ̄lationofAMFundersaltstress)ꎬwhiletherootactivityꎬplantheightꎬstemdiameterꎬabovegroundandunder ̄grounddrymatteramountweresignificantlyhigherthanthoseoftheN ̄CKgroup.ComparedwithN ̄CKgroupꎬtheinoculationofAMFsignificantlyincreasedthecontentofsolubleproteinꎬreducedthecontentofsolublesugarandprolineꎬandalsosignificantlyreducedthecontentofmalondialdehydeꎬsignificantlyincreasedtheactivitiesofantioxidantenzymes(SODꎬCATꎬPOD).Asthetimeincreaseofsaltstressꎬthenetphotosynthet ̄icrate(Pn)ꎬstomatalconductance(Gs)ꎬtranspirationrate(Tr)andintercellularCO2concentration(Ci)oftheleavestreatedwithAMFshowedupwardtrendsꎬwhilethoseofN ̄CKgroupshoweddownwardtrends.ThereforeꎬinoculationofAMFcouldeffectivelyalleviatetheosmoticstressandoxidativedamageofNaCltoThaliadealbataꎬenhanceitsresistancetosaltstressꎬsoastopromotetheplantgrowthandincreasethephoto ̄syntheticefficiency.Keywords㊀ArbuscularmycorrhizalfungiꎻThaliadealbataꎻSaltstressꎻGrowthanddevelopmentꎻPho ̄tosyntheticcharacteristics㊀㊀湿地与森林㊁海洋并称为全球三大生态系统ꎮ湿地包含城市人工湿地和园林湿地ꎬ是生产力最高的生态系统之一ꎬ被誉为 地球之肾 ꎬ具有涵养水源㊁净化水质㊁调节气候㊁改善环境㊁维护生物多样性等重要生态功能[1]ꎮ福建省有湿地园林面积18.86万hm2ꎬ此外ꎬ还有河流水面15.86万hm2㊁水库水面7.36万hm2㊁坑塘水面11.26万hm2[2]ꎮ近年来ꎬ沿海湿地长时间受到盐分侵蚀ꎬ再加上滨海养殖业㊁深层采矿业的迅速发展和盐碱地的大规模开垦ꎬ含盐废水排放量不断增加ꎬ园林湿地水生植物面临的盐胁迫亦越来越严重[3]ꎮ水生植物作为园林湿地中的主要生产者ꎬ是湿地生态系统稳定的基础ꎬ湿地盐碱化的加重严重威胁着湿地水生植物生长ꎮ丛枝菌根真菌(arbuscularmycorrhizalfungiꎬAMF)是一类广泛定殖于植物根系且对植物生长发育起着重要作用的共生内生真菌[4]ꎮAMF不仅能与绝大多数陆生植物共生ꎬ还能与大多数水生植物共生ꎬAMF与植物形成互惠共生体后可以提高宿主植物对养分的吸收ꎬ对植物生长起着重要的积极作用ꎬ同时还能提高植物在逆境胁迫中的生存能力[5]ꎮZong等[6]研究发现AMF通过提高文冠果的光合作用㊁抗氧化酶活性和渗透调节作用来缓解盐胁迫ꎻDong等[7]研究表明AMF通过提高杨树叶绿素荧光和光合作用参数ꎬ增强其对水分㊁K+和Ca2+的吸收ꎬ促进杨树的生长发育和提高植物耐盐碱能力ꎻWang等[8]研究发现AMF可以通过增加青冈栎生物量和液泡中K+含量并区隔化Na+㊁Cl-来提高青冈栎的耐盐性ꎮ研究表明ꎬAMF可以与植物形成共生关系ꎬ可以提高植物从外界环境中吸收营养元素的能力ꎬ提高渗透调节作用ꎬ改善胞间离子平衡ꎬ还可以提高细胞的抗氧化酶活性来缓解盐胁迫造成的氧化损伤ꎬ此外还可以通过提高叶片光合色素含量来增强叶片的光合作用ꎬ进而提高植物的抗盐胁迫能力[9]ꎮ因此ꎬ在盐碱地上通过接种AMF提高植物的耐盐能力是一种有效方法ꎮ再力花(ThaliadealbataFraser)ꎬ别称水莲蕉ꎬ为竹芋科多年生挺水草本植物ꎬ主要生长于公园湿地㊁河流㊁湖泊㊁沼泽以及滨海滩涂等水湿洼地ꎬ具有很强的水肥吸收能力且繁殖能力强ꎬ植株相对高大ꎬ外观美丽ꎬ其紫色花朵和叶子具有很高的观赏价值ꎬ而且对水体具有较强的去污和净化能力ꎬ常用于园林人工湿地污水净化修复ꎬ然而其对高盐水的耐受性较弱ꎬ盐分过高会抑制植株生长和繁殖[10]ꎮ目前ꎬ有关再力花对重金属胁迫的修复作用研究较多[11-12]ꎬ但关于再力花对盐胁迫的研究报道较少ꎬ尤其是在盐胁迫下接种AMF对再力花光合生理特性影响的研究更少ꎮ因此ꎬ本试验以园林水生植物再力花为材料ꎬ研究NaCl胁迫下接种AMF对再力花幼苗生长发育和光合特性的影响ꎬ以阐明AMF提高再力花植株的耐盐作用机制ꎬ为其在园林湿地高盐水中的应用提供技术参考ꎮ1㊀材料与方法1.1㊀试验材料试验于2021年5月至2022年3月在福建农林大学国家林业和草原局森林公园工程技术研究中心开展ꎮAMF菌剂(硅藻土为基质ꎬ包含真菌08山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀孢子㊁菌丝ꎬ孢子量为20~25个/g)㊁地表多样孢囊霉(Diversisporaepigaea):北京农林科学院提供ꎮ再力花苗:青州市诚韵花卉苗木种植专业合作社提供ꎬ株高25~30cmꎮ育苗栽植桶50L(主体直径400mmꎬ口径400mmꎬ高530mm):常州多成塑料包装材料有限公司产品ꎮHoagland营养液:福建万果农资有限公司提供ꎮ栽培土:采自福州市闽江河口国家湿地公园(119.62ʎEꎬ26.03ʎN)ꎬ有机质含量为18.77g/kg㊁有效氮55.84mg/kg㊁有效磷11.88mg/kg㊁速效钾62.74mg/kgꎬpH值6.3ꎬ用前高压灭菌3hꎮ1.2㊀试验设计与管理前期筛选试验得到再力花盐胁迫的最适浓度为150g/kgꎬ此浓度下再力花能维持生长且生长抑制作用较明显ꎬ因此本研究调制桶中土壤盐浓度为150g/kgꎮ选取长势㊁根系生长较一致的再力花苗100株ꎬ每桶栽1株ꎬ桶中装入等量的栽培土ꎬ每桶浇灌等量的无菌水和Hoagland营养液50mLꎮ试验设置4个处理:不接菌+不胁迫(CK组)㊁不接菌+盐胁迫(N-CK组)㊁接菌+不胁迫(T0组)㊁接菌+盐胁迫(T1组)ꎬ每个组各栽培25桶ꎬ置于大棚内进行试验ꎮ1周后待再力花苗长出新叶时ꎬT0㊁T1组每桶接种50g菌剂ꎬCK㊁N-CK均不接菌ꎻ继续培养1周ꎬT1组㊁N-CK组分别加入盐水30L(水面淹没土壤约10cm)ꎬ使得桶中盐浓度为150g/kgꎬT0组㊁CK组加入无菌水30Lꎬ记下刻度线ꎮ后面管理中隔2周均向桶中加入Hoagland营养液50mL并用无菌水补充至水面刻度ꎮ1.3㊀测定指标及方法1.3.1㊀盐害指数分析㊀盐害分级参考赵华等[13]的方法并适当修改ꎬ于盐胁迫开始后的第7天观察植株生长情况ꎮ盐害分级:0级ꎬ植株正常生长ꎬ未出现盐害症状ꎻ1级ꎬ植株生长较正常ꎬ少数植株叶片出现症状ꎬ单株出现0~1片叶发黄ꎻ2级ꎬ植株生长相对缓慢ꎬ单株出现1~3片叶发黄ꎻ3级ꎬ植株生长明显缓慢ꎬ大部分植株出现盐害症状ꎬ单株出现4~6片叶发黄ꎻ4级ꎬ植株生长非常缓慢ꎬ单株绿叶低于2片ꎻ5级ꎬ植株基本停滞生长ꎬ叶片全部变黄ꎬ甚至枯萎ꎮ按照下式计算盐害指数:y=ðaˑb5ˑcˑ100ꎮ式中ꎬy为盐害指数ꎬ单位%ꎻa为N级ꎻb为N级株数ꎻc为总株数ꎮ1.3.2㊀植株农艺性状和生物量测定㊀盐胁迫处理第14天时ꎬ各组分别选取5株ꎬ用卷尺㊁游标卡尺测量株高和茎粗ꎮ同时ꎬ采集整株并分为地上部分和地下部分ꎬ清洗干净后烘箱中105ħ杀青30minꎬ再65ħ烘干至恒重ꎬ用分析天平分别称取地上部和地下部干物质量ꎮ1.3.3㊀根系活力和可溶性糖㊁可溶性蛋白㊁脯氨酸含量测定㊀分别在盐胁迫第7㊁14㊁28天时ꎬ各组随机选取5株拔出ꎬ清洗干净根系ꎬ用纸巾吸干表面水分ꎬ放入液氮中带回实验室ꎮ采用氯化三苯基四氮唑(TTC)法测定根系活力[14]ꎬ采用蒽酮比色法[15]测定根系可溶性糖含量ꎬ采用考马斯亮蓝法[16]测定可溶性蛋白含量ꎬ采用茚三酮显色法[17]测定游离脯氨酸含量ꎮ1.3.4㊀根系抗氧化酶活性和丙二醛含量测定㊀分别于盐胁迫第7㊁14㊁28天时测定根系超氧化物歧化酶(SOD)㊁过氧化氢酶(CAT)㊁过氧化物酶(POD)活性和丙二醛(MDA)含量ꎬ测定方法参考南京建成生物工程研究所试剂盒ꎮ1.3.5㊀叶片光合特性测定㊀采用美国Li-cor6400XT便携式光合作用测定系统ꎬ设置光照强度为1500μmol/(m2 s)ꎬ分别在盐胁迫第7㊁14㊁28天晴天上午9ʒ30 11ʒ30之间ꎬ选择植株顶部下的第2~3片功能性叶片ꎬ分别测定净光合速率(Pn)㊁气孔导度(Gs)㊁蒸腾速率(Tr)和胞间CO2浓度(Ci)ꎮ1.4㊀数据处理与分析所有数据均测量3次ꎬ采用平均值ʃ标准差表示ꎮ数据处理和单因素方差分析采用SPSS22.0软件ꎬ用LSD法进行差异显著性检验(P<0.05)ꎮ采用Origin2021软件作图ꎮ2㊀结果与分析2.1㊀AMF对盐胁迫下再力花植株盐害指数的影响18㊀第12期㊀㊀㊀刘振凡ꎬ等:丛枝菌根真菌对盐胁迫下园林植物再力花生长发育和光合特性的影响由图1可知ꎬCK和T0组植株未出现叶片变化和萎蔫症状ꎻN-CK组叶片变黄和萎蔫症状较严重ꎬ盐害指数为67.54%ꎻT1组叶片出现少量变黄和萎蔫症状ꎬ盐害指数为21.76%ꎮ与N-CK相比ꎬT1组盐害指数降低67.78%ꎮ表明盐胁迫使得再力花植株出现盐害症状ꎬAMF灌根处理显著缓解再力花的盐害程度ꎬ提高抗盐能力ꎮ柱上不同小写字母表示处理间差异显著(P<0.05)ꎮ图1㊀再力花植株盐害指数情况2.2㊀AMF对盐胁迫下再力花根系生长和根系活力的影响图2A显示ꎬ与CK相比ꎬN-CK根系生长明显受到抑制ꎬ根系稀少且短ꎬT0组根系显著增多且生长密集ꎬT1组根系较密集ꎬ但差异不大ꎻ与N-CK相比ꎬT1组根系明显增多增长ꎮ可见ꎬ盐胁迫抑制再力花的根系生长ꎬAMF具有促进根系生长的作用ꎬ一定程度上缓解再力花的盐胁迫效应ꎮ图2B显示ꎬ盐胁迫第7㊁14㊁28天时ꎬT0组根系活力均最高ꎬN-CK根系活力均最低ꎬ与CK相比N-CK㊁T1根系活力均显著降低ꎬ与N-CK相比T1根系活力均显著提高ꎬ分别达14.22%㊁55.47%㊁149.02%ꎮ另外ꎬ从第7天到第28天期间ꎬCK㊁T0组根系活力变化差异均不显著ꎬN-CK呈逐渐显著降低趋势ꎬT1出现先显著升高后缓慢升高趋势ꎮ综上表明ꎬ盐胁迫降低再力花的根系活力ꎬ且随着胁迫时间延长根系活力出现降低趋势ꎬAMF具有提高再力花根系活力的作用ꎬ同时还具有缓解盐胁迫效应ꎮB图中不同小写字母表示不同处理不同天数间差异显著(P<0.05)ꎬ下同ꎮ图2㊀再力花根系生长情况(A)和根系活力值(B)2.3㊀AMF对盐胁迫下再力花植株生长的影响由表1可知ꎬ与CK相比ꎬ单独盐胁迫下(N-CK)ꎬ再力花株高㊁茎粗㊁地上干物质量和地下干物质量均显著降低ꎬ说明盐胁迫显著抑制再力花生长ꎻ而施用AMF(T0组)则显著提高植株的各指标值ꎬ说明AMF具有促进植株生长的作用ꎮ与N-CK相比ꎬ盐胁迫下施用AMF(T1组)显著提高株高㊁茎粗㊁地上干物质量和地下干物质量ꎬ分别提高14.73%㊁10.58%㊁19.24%和49.77%ꎮ说明ꎬ接种AMF不仅促进再力花植株生长ꎬ还有利于提高再力花植株对盐胁迫环境的适应性ꎮ㊀㊀表1㊀㊀AMF对盐胁迫下再力花植株生长的影响处理株高/cm茎粗/mm地上干物质量/(g/株)地下干物质量/(g/株)CK42.78ʃ0.45b6.52ʃ0.27b24.56ʃ0.28b6.85ʃ0.21bN-CK36.32ʃ0.36c5.86ʃ0.31c19.85ʃ0.22c4.36ʃ0.16cT045.82ʃ0.84a6.87ʃ0.44a27.33ʃ0.31a7.38ʃ0.24aT141.67ʃ0.62b6.48ʃ0.39b23.67ʃ0.19b6.53ʃ0.18b㊀㊀注:同列数据后不同小写字母表示处理间差异显著(P<0.05)ꎮ2.4㊀AMF对盐胁迫下再力花根系渗透调节物质含量的影响由图3可知ꎬ非盐胁迫下ꎬ同时期CK和T028山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀组根系渗透调节物质(可溶性蛋白㊁可溶性糖及脯氨酸)含量均不存在显著差异(胁迫7天时脯氨酸含量除外)ꎬ说明施用AMF对根系渗透调节物质的影响不显著ꎻ盐胁迫下ꎬN-CK和T1组根系渗透调节物质含量均大幅升高ꎬ说明盐胁迫对再力花根系产生渗透胁迫ꎬ使得细胞内积累大量渗透物质以缓解外界环境的胁迫作用ꎮ胁迫第7㊁14㊁28天时ꎬ与N-CK相比ꎬT1组可溶性蛋白含量分别升高7.83%㊁39.79%㊁75.88%且第14㊁28天时达到显著水平ꎬ而可溶性糖含量则分别降低28.14%㊁31.56%和34.34%ꎬ脯氨酸含量分别降低16.41%㊁24.55%和24.38%ꎬ且均达显著水平ꎮ这说明AMF具有提高根系可溶性蛋白含量和降低可溶性糖㊁脯氨酸含量的作用ꎮ图3㊀AMF对盐胁迫下再力花根系㊀㊀㊀渗透调节物质含量的影响2.5㊀AMF对盐胁迫下再力花根系MDA含量和抗氧化酶活性的影响2.5.1㊀对根系MDA含量的影响㊀图4显示ꎬ在非盐胁迫下ꎬCK和T0组根系MDA含量均较低ꎬT0组均显著低于CK组ꎬ随着生长天数的增加ꎬ两者MDA含量变化均不显著ꎻ盐胁迫下ꎬN-CK和T1组根系MDA含量均显著增加ꎮ表明ꎬAMF具有降低非盐胁迫下根系MDA含量的作用ꎬ而盐胁迫可使根系产生大量的MDAꎮ与N-CK相比ꎬ盐胁迫第7㊁14㊁28天时ꎬT1组MDA含量分别显著降低达29.68%㊁39.42%和55.20%ꎬ另外ꎬ随着胁迫天数增加ꎬN-CK组MDA含量出现显著增加趋势ꎬ而施用AMF处理可以缓解根系中MDA的净生成速率ꎮ由此可见ꎬ盐胁迫下施用AMF可以降低根系MDA含量ꎬ缓解细胞的脂质过氧化损伤ꎮ图4㊀AMF对盐胁迫下再力花根系MDA含量的影响2.5.2㊀对根系SOD㊁CAT㊁POD活性的影响㊀图5显示ꎬ非盐胁迫下ꎬCK和T0组根系抗氧化酶(SOD㊁CAT㊁POD)活性均维持在一个较低水平ꎬ但施用AMF的T0组抗氧化酶活性均不同程度地高于CK组ꎮ这主要是因为再力花根系在正常生理状态下细胞中氧化和抗氧化水平维持在动态平衡中ꎬ其抗氧化酶活性较低ꎬ在AMF与根系形成共生作用时ꎬAMF可能通过某种途径相对地提高根系抗氧化酶活性ꎮ盐胁迫下ꎬN-CK和T1组根系抗氧化酶活性均出现大幅升高ꎬ这说明150g/kgNaCl水溶液给再力花根系造成胁迫作用ꎬ体内产生大量自由基ꎬ为维持氧化和抗氧化平衡ꎬ从而激活抗氧化酶的活性ꎮ随着盐胁迫天数增加ꎬN-CK组SOD活性出现显著降低趋势ꎬCAT和38㊀第12期㊀㊀㊀刘振凡ꎬ等:丛枝菌根真菌对盐胁迫下园林植物再力花生长发育和光合特性的影响POD活性则出现先显著升高后显著降低趋势ꎻT1组抗氧化酶活性呈现显著升高趋势ꎮ与N-CK相比ꎬ盐胁迫第7㊁14㊁28天时ꎬT1组SOD活性分别显著升高29.50%㊁95.11%和190.21%ꎬCAT活性分别显著升高29.68%㊁51.52%和140.14%ꎬPOD活性分别显著升高37.11%㊁22.31%和101.78%ꎮ综上ꎬ盐胁迫使再力花根系抗氧化酶活性不同程度升高ꎬ施用AMF后ꎬ抗氧化酶活性得到进一步提高ꎬ表明AMF具有提高抗氧化酶活性的作用ꎮ图5㊀AMF对盐胁迫下再力花根系SOD㊁㊀㊀CAT㊁POD活性的影响2.6㊀AMF对盐胁迫下再力花叶片光合特性的影响2.6.1㊀对Pn的影响㊀叶片的净光合速率(Pn)反映光合作用强弱和有机物积累程度ꎬ它直接决定了植株生物量的积累ꎬ其值越大单位时间内生成的有机产物就越多[18]ꎮ由图6可知ꎬ非盐胁迫组(CK和T0)叶片Pn均大于盐胁迫组(N-CK和T1)ꎬ且随着栽培周期增加ꎬCK㊁T0㊁T1的Pn均呈现上升趋势ꎬ而N-CK呈现下降趋势ꎮ盐胁迫第7㊁14㊁28天时ꎬ与CK相比ꎬT0的Pn均显著升高ꎬ分别升高12.10%㊁20.30%和28.96%ꎬN-CK的Pn分别显著降低15.85%㊁33.60%和44.52%ꎬ说明非盐胁迫下施用AMF可以提高再力花叶片光合效率ꎬ盐胁迫则大大降低叶片的光合效率ꎻ与N-CK相比ꎬT1的Pn分别显著升高5.98%㊁36.24%和57.77%ꎮ综上表明ꎬ盐胁迫降低叶片的光合作用ꎬ施用AMF可以缓解盐害对叶片光合效率的抑制ꎬ提高净光合速率ꎬ促进体内有机物的合成ꎮ图6㊀AMF对盐胁迫下再力花净光合速率的影响2.6.2㊀对Gs的影响㊀气孔导度表示气孔张开的程度ꎬ是与外界环境因子(如二氧化碳㊁水蒸气分子)交互的门户ꎬ可以调控植物光合作用效率ꎬ其值越大光合效率就越高[19]ꎮ由图7可知ꎬ非盐胁迫组(CK和T0)的Gs均大于盐胁迫组(N-CK和T1)ꎬ再力花生长过程中ꎬCK㊁T0㊁T1的Gs均呈现上升趋势ꎬ而N-CK呈现下降趋势ꎮ盐胁迫第7㊁14㊁28天时ꎬ与CK相比ꎬN-CK的Gs分别显著降低35.59%㊁70.89%和92.85%ꎬT0组分别显著升高23.53%㊁26.67%和17.79%ꎬ说明盐胁迫降低叶片的气孔导度ꎬ而在非盐胁迫下施用AMF可以提高叶片的气孔导度ꎻ与N-CK相比ꎬT1的Gs分别显著升高41.55%㊁174.81%和942.65%ꎮ上述结果表明ꎬ盐胁迫限制叶片气孔的张开程度ꎬ施用AMF可以缓解再力花的盐胁迫效应ꎬ气孔导度增大ꎬ从而增强叶片的光合作用ꎮ2.6.3㊀对Tr的影响㊀由图8可知ꎬ非盐胁迫组(CK和T0)的Tr均大于盐胁迫组(N-CK和48山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀T1)ꎬ再力花生长过程中ꎬCK㊁T0㊁T1的Tr均呈现上升趋势ꎬ而N-CK呈现下降趋势ꎬ这间接说明CK㊁T0㊁T1为正常生长ꎬN-CK在盐胁迫下生长受到抑制ꎮ盐胁迫第7㊁14㊁28天时ꎬ与CK相比ꎬN-CK的Tr分别显著降低34.16%㊁57.49%和91.00%ꎬT0组分别显著升高12.87%㊁7.29%和16.26%ꎬTr的变化趋势与Gs相似ꎻ与N-CK相比ꎬT1的Tr分别显著升高33.83%㊁83.81%和765.38%ꎮ上述结果表明ꎬ盐胁迫下叶片蒸腾速率明显降低ꎬ施用AMF提高再力花的蒸腾速率ꎬ其光合作用随之增强ꎮ图7㊀AMF对盐胁迫下再力花气孔导度的影响图8㊀AMF对盐胁迫下再力花蒸腾速率的影响2.6.4㊀对Ci的影响㊀由图9可知ꎬ再力花植株从盐胁迫第7天到第28天ꎬCK㊁T0㊁T1的胞间CO2浓度(Ci)呈逐渐升高趋势ꎬN-CK呈现下降趋势ꎮ第7㊁14㊁28天时ꎬ与CK相比ꎬN-CK的Ci分别显著降低43.25%㊁56.29%和68.28%ꎬT0组分别显著升高10.83%㊁16.55%和16.76%ꎻ与N-CK相比ꎬT1的Ci分别显著升高22.23%㊁65.44%和158.78%ꎮ上述结果表明ꎬCi的变化趋势与Pn㊁Gs㊁Tr趋势相似ꎬ说明Ci与Pn呈现正相关ꎬ叶片光合速率的变化主要是由气孔因素引起ꎬ盐胁迫下叶片胞间CO2浓度明显降低ꎬ施用AMF提高了胞间CO2浓度ꎮ图9㊀AMF对盐胁迫下再力花胞间CO2浓度的影响3㊀讨论3.1㊀AMF对盐胁迫下再力花植株生长的促生作用根系活力对植物生长起着重要影响ꎬ活力越高其对土壤水分和营养元素的吸收作用越强ꎬ植物生长才更加茂盛[20]ꎮAMF在植物根系成功定殖后ꎬ形成庞大的菌丝网络ꎬ可以提高根系对水分㊁营养物质的吸收和利用效率ꎬ而且还能加强植物体内营养元素的运输ꎬ从而促进植物的生长发育ꎮ安晓霞等[21]研究发现AMF可以提高紫花苜蓿根系对磷的吸收和利用ꎬ具有较好的促生作用ꎻ王紫瑄等[22]研究发现AMF可以提高蒙古沙冬青幼苗根系活力和根面积ꎬ同时提高土壤酶活性ꎬ促进其根系阳离子交换ꎬ从而促进植株生长和生物量积累ꎻFall等[23]研究发现AMF增加土壤中有效氮磷的浓度ꎬ提高土壤肥力ꎬ促进根系对养分的吸收ꎬ从而提高玉米产量ꎮ本研究结果显示ꎬ非盐胁迫下接种AMF的再力花根系活力显著高于对照组ꎬ且根系生长形态优于对照组ꎬ显著提高株高㊁茎粗以及地上地下干物质量ꎮ本研究结果与上述学者的研究结果均表明ꎬAMF在宿主植物根系定殖后ꎬ可以促进植株生长ꎮ盐胁迫下ꎬ植株最明显的表现就是矮化和生物量下降ꎬ盐害不仅会影响根系Na+大量积累和K+流失ꎬ还会影响根系对其他营养元素的吸收ꎬ使植株处于营养亏缺状态ꎬ导致生长不良ꎮ紫花苜蓿[24]㊁粗枝木麻黄[25]㊁皂荚[26]在受到盐胁迫时ꎬ其植株会变矮ꎬ58㊀第12期㊀㊀㊀刘振凡ꎬ等:丛枝菌根真菌对盐胁迫下园林植物再力花生长发育和光合特性的影响茎秆变得纤细ꎬ叶片出现黄化以及生物量大幅下降等现象ꎬ接种AMF后植株的盐害现象得到显著改善ꎬ干物质积累量显著提高ꎮ本研究中ꎬ盐胁迫下未接种AMF时ꎬ再力花植株性状受到明显抑制ꎬ而接种AMF可以明显缓解盐害效应ꎬ其株高㊁茎粗㊁干物质量均显著高于未接种的盐害组ꎮ这表明AMF提高了再力花对盐胁迫环境的适应性ꎮ3.2㊀AMF对盐胁迫下再力花根系渗透调节和氧化应激的影响可溶性蛋白㊁可溶性糖㊁脯氨酸作为植物体内重要的渗透调节物质ꎬ植物受到盐害时体内可以合成并大量积累这些调节物质来提高细胞的渗透势ꎬ从而促进根系对水分的吸收和运输[27-28]ꎮLiu[29]㊁赵艳兰[30]㊁陈晓楠[31]等分别在研究盐胁迫对紫花地丁㊁白车轴草㊁疏叶骆驼刺的影响时发现ꎬ与对照组相比ꎬ盐胁迫下植株体内可溶性蛋白㊁可溶性糖㊁脯氨酸含量均显著升高ꎬ且接种AMF后体内这些渗透调节物质含量均出现一定程度的降低ꎬ植株的盐害症状明显好转ꎮ本研究中ꎬ盐胁迫组再力花根系可溶性蛋白㊁可溶性糖㊁脯氨酸含量均显著高于正常对照组ꎬ接种AMF后可溶性蛋白含量显著高于非接种组ꎬ而可溶性糖和脯氨酸含量则显著低于非接种组ꎮ可溶性蛋白含量的升高可能是由于AMF改善根系的盐胁迫效应ꎬ参与细胞内物质运输蛋白以及产生大量抗氧化酶ꎬ而可溶性糖含量和脯氨酸含量的下降则可能是由AMF提高根系抗盐性而使细胞活性逐渐恢复到正常情况㊁盐害效果降低所致[32]ꎮ上述这些研究均表明ꎬAMF可以通过调节渗透物质含量来缓解盐胁迫对植株的盐害作用ꎮ植物体内有一套较复杂的氧化应激系统ꎬSOD㊁CAT㊁POD是植物体内重要的抗氧化酶ꎬ组成了酶促防御系统的第一道防线[33]ꎮ植物处在盐胁迫环境中时ꎬ根系细胞内会产生大量的自由基ꎬ引起细胞的脂质过氧化而产生大量MDAꎬ进而导致细胞膜表面结构被损坏ꎬ体内抗氧化酶活性会升高以清除体内过多的自由基ꎬ维持细胞内氧化还原平衡[34]ꎮ薛红飞[35]㊁曹岩坡[36]㊁Bencherif[37]等分别研究发现ꎬ草莓㊁芦笋㊁苜蓿在盐胁迫下ꎬ体内MDA含量会急剧升高ꎬ抗氧化酶活性会显著升高ꎬ然而随着盐胁迫时间延长ꎬMDA含量会进一步提高ꎬ抗氧化酶活性则出现下降趋势ꎻ施加外源AMF植物体内抗氧化酶活性会显著高于未接种组ꎬMDA含量则低于未接种组ꎮ本研究结果表明ꎬ盐胁迫下再力花根系MDA含量和抗氧化酶活性均显著升高ꎬ施加AMF后ꎬ根系MDA含量低于未接种组ꎬ抗氧化酶活性显著高于未接种组ꎬ与盐胁迫组(N-CK)相比ꎬ第7㊁14㊁28天时ꎬ施加AMF组的MDA含量分别显著降低29.68%㊁39.42%和55.20%ꎬSOD活性分别显著升高29.50%㊁95.11%和190.21%ꎬCAT活性分别显著升高29.68%㊁51.52%和140.14%ꎬPOD活性分别显著升高37.11%㊁22.31%和101.78%ꎮ本研究结果与上述学者的研究结果相似ꎬ表明AMF可以缓解盐害下再力花植株根系细胞内的氧化应激水平ꎮ3.3㊀AMF对盐胁迫下再力花叶片光合特性的影响植物生物量的积累主要由叶片光合作用强弱决定ꎬ光合作用是植物初级代谢的关键过程ꎬ该过程中光能转换㊁电子运输对盐胁迫作用很敏感ꎬ盐胁迫下叶肉细胞的光合活性会被抑制ꎬ另外ꎬ叶绿素的合成同样对盐胁迫敏感[38]ꎮ研究表明盐胁迫下植物叶片的光合特性会发生变化ꎬ以此适应逆境胁迫[39]ꎮ龚远博[40]㊁马仕林[41]㊁赵华[13]等分别研究发现ꎬ杜梨㊁榉树㊁番茄幼苗在盐胁迫下其叶片净光合速率下降ꎬ部分气孔关闭ꎬ气孔导度㊁蒸腾速率㊁胞间CO2浓度降低ꎬ整片叶的光合性能降低ꎬ而施加AMF后叶片的光合作用逐渐增强ꎬ说明AMF可以激活叶片的光合性能ꎬ缓解盐胁迫对幼苗叶片的光损伤ꎮ本研究中ꎬ再力花植株在盐胁迫下叶片的Pn㊁Gs㊁Tr㊁Ci均随着胁迫时间的延长而逐渐降低ꎬ施加AMF后ꎬ叶片的Pn㊁Gs㊁Tr㊁Ci均升高ꎬ且随着时间延长而逐渐升高ꎮ这说明AMF具有提高叶片光合作用的能力ꎮ研究认为ꎬ植物在盐胁迫下接种AMF能够通过提高叶片气孔交换能力㊁光合色素含量以及光合系统中光合活性ꎬ从而使叶片的光合作用增强[42]ꎬ本研究中AMF提高再力花的光合作用可能与此相68山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀关ꎬ崔令军[43]㊁张永志[44]等同样研究发现盐胁迫下施加AMF可提高植物叶片光合色素含量和光合作用ꎮ这些研究结果均表明ꎬAMF可以通过改善气体交换参数和叶绿素光合参数来提高植物对光能的利用能力ꎬ从而缓解盐分对植物造成的胁迫作用ꎬ增强植物的耐盐性ꎮ4㊀结论盐胁迫下再力花盐害指数显著高于正常对照组ꎬ且植株生长明显受到抑制ꎮ施加AMF处理(T1)对植株生长的促进作用明显ꎬ与盐胁迫未接菌组(N-CK)相比ꎬ根系活力得到显著提高ꎬ且显著提高株高㊁茎粗㊁干物质量ꎮAMF具有通过调控再力花根系渗透物质积累来缓解盐害胁迫的作用ꎬ同时还具有降低根系MDA含量和提高抗氧化酶活性的作用ꎬ从而缓解盐胁迫对根系造成的氧化损伤ꎮAMF还对盐胁迫下再力花叶片光合特性具有一定的改善作用ꎬ提高叶片的光合效率ꎮ综之ꎬAMF不仅对再力花植株生长具有促进效应ꎬ还具有缓解盐胁迫对植株造成损伤的作用ꎮ其作用机制可能是ꎬAMF通过渗透调节作用ꎬ激活抗氧化酶活性并降低MDA含量以及保护叶肉细胞的光合色素ꎬ提高光合特性ꎮ参㊀考㊀文㊀献:[1]㊀徐长林ꎬ任君临ꎬ贺强.黄河三角洲盐沼湿地蟹类对芦苇种群动态的下行控制作用及空间变异[J].复旦学报(自然科学版)ꎬ2022ꎬ61(4):417-423.[2]㊀郭舜ꎬ陈峰ꎬ吕国梁.河流湿地生态修复规划:以福建龙岩东山湿地公园为例[J].湿地科学与管理ꎬ2022ꎬ18(3):51-54.[3]㊀黄珂.NaCl胁迫下湿地植物的形态变化及其对生理应激的影响[J].山东农业大学学报(自然科学版)ꎬ2023ꎬ54(3):373-377.[4]㊀MuzafarHDꎬSyedMRꎬNarenderSꎬetal.Arbuscularmycor ̄rhizalfungiforsalinitystress:anti ̄stressroleandmechanisms[J].Pedosphereꎬ2023ꎬ33(1):212-224.[5]㊀LiangSMꎬLiQSꎬLiuMYꎬetal.Mycorrhizaleffectsongrowthandexpressionsofstress ̄responsivegenes(aquaporinsandSOSs)oftomatoundersaltstress[J].J.Fungiꎬ2022ꎬ8(12):1305.[6]㊀ZongJWꎬZhangZLꎬHuangPLꎬetal.ArbuscularmycorrhizalfungialleviatessaltstressinXanthocerassorbifoliumthroughimprovedosmotictoleranceꎬantioxidantactivityꎬandphotosyn ̄thesis[J].FrontiersinMicrobiologyꎬ2023ꎬ14:1138771. [7]㊀DongFXꎬWangYHꎬTaoJꎬetal.Arbuscularmycorrhizalfun ̄giaffecttheexpressionofPxNHXgenefamilyꎬimprovephoto ̄synthesisandpromotePopulussimoniiˑP.nigragrowthundersaline ̄alkalistress[J].FrontiersinPlantScienceꎬ2023ꎬ14:1104095.[8]㊀WangYHꎬDongFXꎬChenHꎬetal.Effectsofarbuscularmy ̄corrhizalfungusonsodiumandchlorideionchannelsofCasua ̄rinaglaucaundersaltstress[J].InternationalJournalofMo ̄lecularSciencesꎬ2023ꎬ24(4):3680.[9]㊀郭娜ꎬ张玥ꎬ刘贤雍ꎬ等.丛枝菌根真菌提高植物耐盐性生理机制研究进展[J].江苏农业科学ꎬ2023ꎬ51(4):16-23. [10]陈满军ꎬ方燕ꎬ戢茜ꎬ等.再力花强化人工湿地除汞效能的试验研究[J].环境生态学ꎬ2021ꎬ3(8):59-65.[11]迟韵阳ꎬ付宇新ꎬ何素琳ꎬ等.10种湿地植物对不同程度模拟污染水体的净化效果研究[J].南方林业科学ꎬ2021ꎬ49(3):37-41ꎬ69.[12]张洲ꎬ张单ꎬ祖艳群ꎬ等.5种植物的矿区生长及对底泥重金属富集特征[J].环境科学与技术ꎬ2021ꎬ44(7):115-122. [13]赵华ꎬ任晴雯ꎬ王熙予ꎬ等.丛枝菌根真菌对盐胁迫下番茄抗氧化酶活性和光合特性的影响[J].浙江农业学报ꎬ2021ꎬ33(11):2075-2084.[14]赵世杰ꎬ史国安ꎬ董新纯.植物生理学实验指导[M].北京:中国农业科学技术出版社ꎬ2002:47ꎬ120.[15]王学奎.植物生理生化实验原理和技术[M].第2版.北京:高等教育出版社ꎬ2006.[16]刘萍ꎬ李明军.植物生理学实验技术[M].北京:科学出版社ꎬ2007.[17]ShabnamNꎬTripathiIꎬSharmilaPꎬetal.Arapidꎬidealꎬandeco ̄friendlierprotocolforquantifyingproline[J].Protoplasmaꎬ2016ꎬ253(6):1577-1582.[18]高海哲ꎬ贾志国ꎬ张丽.盐胁迫对平欧杂种榛幼苗生长及光合生理的影响[J].江西农业学报ꎬ2023ꎬ35(4):74-77ꎬ97. [19]李果ꎬ韩聪颖ꎬ张宝忠.夏玉米气孔导度模型适用性分析[J].节水灌溉ꎬ2023(4):32-38ꎬ51.[20]李晴ꎬ段文艳ꎬ李鑫ꎬ等.丛枝菌根真菌对元宝枫生长及其根系形态的影响[J/OL].西北农林科技大学学报(自然科学版)ꎬ2024:1-8.http://doi.org/10.13207/j.cnki.jnwafu.2024.01.009.[21]安晓霞ꎬ李想ꎬ曹冠华ꎬ等.菌磷互作对紫花苜蓿地上生物量及营养品质的影响[J].中国草地学报ꎬ2023ꎬ45(4):90-99.[22]王紫瑄ꎬ解甜甜ꎬ王雅茹ꎬ等.丛枝菌根真菌(AMF)对蒙古沙冬青幼苗的促生特性及作用机制[J].干旱区研究ꎬ78㊀第12期㊀㊀㊀刘振凡ꎬ等:丛枝菌根真菌对盐胁迫下园林植物再力花生长发育和光合特性的影响。
丛枝菌根(AM)对植物矿质营养影响-PPT文档资料

2.2丛枝菌根与植物的氮素营养
氮是植物体 中蛋白质、酶、叶绿素、维生素等的 重要组成元素,缺氮将限制植物生长、减少产量 和降低品质。大量试验研究表明,接种AM真菌可 促进植物对氮的吸收利用,改善植物的氮素营养, 最终促进植物生长发育,提高产量,改善品质。 有研究表明,接种AM菌显著提高了生姜叶片和根 中氮素含量。陈洁敏等在试验中得出,AMF处理 的玉米氮素吸收 比对照增加24.14%~56.65%。 Gupta注意到,在田间条件下AM极显著地提高了 薄荷茎叶中氮的含量,接种AM菌的植株根际土壤 中氮素明显呈现亏缺。
光合速率、蒸腾速率与碳水化合物含量 研究表明,与对照相比,接菌植株的光合速率、 蒸腾速率有一定程度或显著的提高,其中气孔阻 力减小,碳水化合物含量明显增加。 与非接菌株相比,接菌黄瓜 的光合速率与蒸腾速 率提高了24.8%与11.7%;水分胁迫条件下沙田柚 实验组和对照组幼苗叶片的光合速率比移栽时分 别增加了3.9和3.5倍,表明沙田柚接种AM真菌后 有利于提高其水分胁迫阶段叶片的光合速率。
改变根际土壤pH,活化难溶性磷酸盐 研究表明,pH是影响AM菌丝际能否有效吸收利 用土壤磷的重要因素。石灰性土壤中根际pH值的 下降有利于植物对磷的吸收,在缺磷条件下,AM 真菌通过分泌质子和有机酸,改变菌丝际的pH以 活化土壤中难溶性磷酸盐,并促使磷从土壤有机 磷酸盐的金属螯合物中释放出来,从而增加土壤 磷浓度,提高植株的吸磷量。
根据其形态和解剖学特征,菌根可分为外生菌根、内生菌 根和内外生菌根3种类型。
1.1 外生菌根
菌根具有菌丝套。 部分菌丝侵入根的外皮层细胞间隙,形成哈氏网。 菌丝不进入皮层细胞之中。 加强植物对矿质营养元素的吸收。 向植物提供生长素、维生素、细胞分裂素、抗生 素和脂肪酸等代谢产物,促进植物生长。 提高植物对病原菌侵染和对温度、干旱和过酸或 过碱等极端环境的抗性。
高中生物知识结构网络图(完整版)
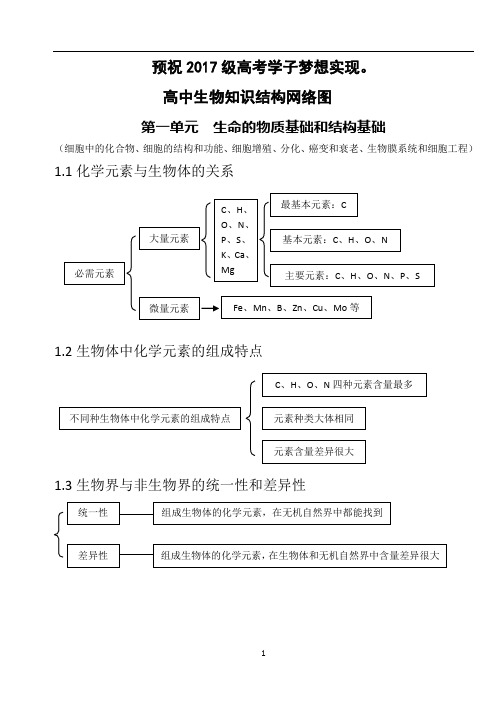
预祝2017级高考学子梦想实现。
高中生物知识结构网络图第一单元 生命的物质基础和结构基础(细胞中的化合物、细胞的结构和功能、细胞增殖、分化、癌变和衰老、生物膜系统和细胞工程)1.1化学元素与生物体的关系1.2生物体中化学元素的组成特点1.3生物界与非生物界的统一性和差异性1.4细胞中的化合物一览表1.5蛋白质的相关计算设 构成蛋白质的氨基酸个数m ,构成蛋白质的肽链条数为n ,构成蛋白质的氨基酸的平均相对分子质量为a , 蛋白质中的肽键个数为x , 蛋白质的相对分子质量为y ,控制蛋白质的基因的最少碱基对数为r ,则 肽键数=脱去的水分子数,为 n m x -= …………………………………①蛋白质的相对分子质量 x ma y 18-= ………………………………………②或者 x a ry 183-=………………………………………③ 1.6蛋白质的组成层次1.7核酸的基本组成单位1.8生物大分子的组成特点及多样性的原因1.9生物组织中还原性糖、脂肪、蛋白质和DNA的鉴定1.10选择透过性膜的特点1.11水被选择的离子和小分子其它离子、小分子和大分子亲脂小分子高浓度——→低浓度不消耗细胞能量(ATP)离子、不亲脂小分子低浓度——→高浓度需载体蛋白运载消耗细胞能量(ATP)1.12线粒体和叶绿体共同点1、具有双层膜结构2、进行能量转换3、含遗传物质——DNA4、能独立地控制性状5、内含核糖体6、有相对独立的转录翻译系统7、能自我分裂增殖1.13真核生物细胞器的比较1.14细胞有丝分裂中核内DNA、染色体和染色单体变化规律1.15理化因素对细胞周期的影响注:+ 表示有影响1.16细胞分裂异常(或特殊形式分裂)的类型及结果1.18已分化细胞的特点 1.19分化后形成的不同种类细胞的特点G 21.20分化与细胞全能性的关系1.211.22癌细胞的特点分化程度越低全能性越高,分化程度越高全能性越低分化程度高,全能性也高分化程度最低(尚未分化),全能性最高扁平梭形 球形成纤维细胞癌变如癌细胞膜糖蛋白减少,细胞黏着性降低,易转移扩散。
丛枝菌根真菌提高植物耐盐性的机理研究进展

丛枝菌根真菌提高植物耐盐性的机理研究进展叶贤锋;吴强盛;孙润生;赵伦杰【摘要】土壤盐渍化是影响植物正常生长的主要因素之一,丛枝菌根真菌能提高植物的耐盐性.分析了土壤盐渍化对丛枝菌根真菌生长、发育的影响,重点从营养吸收、光合作用、根系、抗氧化防御系统和脯氨酸等5个方面阐述了丛枝菌根真菌提高植物耐盐性的机理.%Soil salinization was a factor affecting normal growth of plants. Arbuscular mycorrhizal fungi could improve the salt tolerance of plants. The effects of soil salinization on the growth and development of arbuscular mycorrhizal fungi was analyzed,and the possible mechanisms that arbuscular mycorrhizas enhance the salt tolerance of plants from aspects of nutrient uptake,photosynthesis, antioxidative defence system, root and proline were elaborated.【期刊名称】《湖北农业科学》【年(卷),期】2011(050)001【总页数】3页(P9-11)【关键词】丛枝菌根真菌;土壤盐渍化;植物耐盐性机理【作者】叶贤锋;吴强盛;孙润生;赵伦杰【作者单位】长江大学园艺园林学院,湖北,荆州,434025;长江大学园艺园林学院,湖北,荆州,434025;华南农业大学农业部生态农业重点开放实验室,广州,510640;长江大学园艺园林学院,湖北,荆州,434025;长江大学园艺园林学院,湖北,荆州,434025【正文语种】中文【中图分类】S154.36近年来,土壤盐渍化现象已越来越严重,在干旱、半干旱地区更为严重,世界陆地面积的7%已经被盐渍化;在可持续发展农业中提高植物在盐碱土壤中的正常生长显得尤为重要。
丛枝菌根真菌根外菌丝形成时间及对牧草的促生长效应

03
营养成分分析
测定牧草叶片光合速率、叶绿素 含量、根系活力等生理指标,以 揭示促生长机制。
分析牧草中蛋白质、脂肪、纤维 等营养成分含量,以评价牧草品 质改善情况。
促生长效应评估及机制探讨
1 2 3
促生长效应评估
综合比较不同处理组牧草生长指标和生理指标差 异,评估丛枝菌根真菌对牧草的促生长效应。
机制探讨
创新点与不足之处
创新点
本研究首次系统地研究了丛枝菌根真菌根外菌丝的形成时间及其对牧草的促生长效应,为丛枝菌根真 菌在牧草生产中的应用提供了理论依据。
不足之处
由于实验条件和时间的限制,本研究未能涵盖所有可能的菌种和宿主植物组合,也未能深入研究丛枝 菌根真菌与牧草之间的互作机制。
未来研究方向与应用前景
而影响根外菌丝的形成时间。
牧草种类
温度、湿度、光照等环境条件对 丛枝菌根真菌的生长和繁殖具有 重要影响,因此也会影响根外菌 丝的形成时间。
环境条件
土壤中的营养成分、pH值、透 气性等因素也会影响丛枝菌根真 菌的生长和根外菌丝的形成时间 。
04 丛枝菌根真菌对 牧草的促生长效 应
实验设计与实施
实验材料选择
选用代表性丛枝菌根真菌菌种和适宜牧草品种。
实验处理设置
设置不同接种量、接种时间和培养条件等处理组,以探究最佳促 生长条件。
实验重复与对照
确保实验具有可重复性和对照性,以准确评估促生长效应。
牧草生长指标测定
01
生长量测定
定期测定牧草株高、叶面积、生 物量等生长指标,以评估促生长 效果。
02
生理指标测定
THANKS
感谢观看
本研究采用了先进的分子生物学技术 和显微观察手段,对丛枝菌根真菌根 外菌丝的形成过程进行了深入细致的 研究,为揭示其促生长机制提供了有 力支持。
丛枝菌根

菌根分类AM:丛枝菌根(苔藓、蕨类、裸子、被子)ECM:外生菌根(蕨类、裸子、被子)EM:内生菌根EEM:内外兼生菌根(裸子、被子)ARM:浆果鹃类菌根MM:水晶兰类菌根ERM:欧石楠类菌根OM:兰科菌根结构AM真菌包括;菌丝、丛枝、泡囊、辅助细胞、孢子和孢子果等结构1)菌丝(Hyphae)任何一种菌根都由植物根系、两个相关的菌丝系统三部分组成,其中菌丝系统一个是分布于土壤中的,另一个是分布于根系内的。
AM真菌中,分布于土壤中的菌丝称为外生菌丝或根外菌丝,通常呈网状结构,有时形成二分叉吸收结构。
根外菌丝从形态上又可分为两种:厚壁菌丝和薄壁菌丝。
根外菌丝对损伤的愈合能力较强。
在较粗的根外菌丝上可以产生大量的休眠孢子。
分布于根系内的菌丝称为内生菌丝或根内菌丝。
内生菌丝又可分为胞间菌丝和胞内菌丝。
胞间菌丝是在皮层薄壁管胞中间由圈状菌丝或由侵入菌丝分叉直接形成。
(2)丛枝(Arbuscule)AM真菌侵入宿主植物根系皮层细胞内,经过连续二叉分枝生长形成树枝状或花椰菜状结构,即丛枝。
丛枝是AM真菌最重要的结构,它是AM真菌侵染宿主植物根细胞组织内部进一步延伸的端点,被认为是宿主植物与AM真菌进行物质和能量交换的优势位点或主要场所。
(3)泡囊(Vesicle)泡囊是由根内菌丝顶端膨大而形成的球形、棒形、圆柱形、椭圆形或不规则形结构,可在根系皮层细胞内或细胞间生长发育。
并非所有的AM真菌都产生泡囊,如巨孢囊霉属和盾巨孢囊霉属的真菌则不再根内产生泡囊。
关于泡囊的功能有两种观点:一种认为它是繁殖器官;另一种则认为它是储藏器官。
(4)辅助细胞(Auxiliary cell)辅助细胞是巨孢囊霉真菌所特有的结构,这个科的真菌不在根系皮层细胞内或间隙产生泡囊。
巨孢囊霉科菌根真菌的繁殖体萌发而尚未侵染寄主根系的过程中,及侵入根系后,菌丝在根外分叉,末段隆起、膨大形成辅助细胞(根外泡囊)。
巨孢囊霉科的根外辅助细胞与球囊霉科和无梗囊霉科的根内泡囊一样,被认为是储存营养的器官。
丛枝菌根真菌对福鼎大白茶生长、侧根数和根系内源激素的影响

丛枝菌根真菌对福鼎大白茶生长、侧根数和根系内源激素的影响作者:夏庭君吴强盛邵雅东来源:《广西植物》2018年第12期摘要:; 该研究以盆栽福鼎大白茶(Camellia sinensis ‘Fuding Dabaicha’)为材料,通过对其接种丛枝菌根真菌(AM真菌)幼套球囊霉(Clariodeoglomus etunicatum)、地表球囊霉(Diversispora versiformis)、粘屑多孢囊霉(D. spurca)以及上述三菌种的混合菌剂,研究AM真菌对茶生长、侧根数及根系內源激素的影响。
结果表明:接种12周后福鼎大白茶根系能被AM真菌侵染,为18.85%~40.23%。
接种AM真菌处理促进了福鼎大白茶株高、叶面积、主根长以及一级侧根和三级侧根数量,但抑制了二级侧根数(除混合菌种)。
单一的AM 真菌接种显著提高了福鼎大白茶根系脱落酸、玉米素核苷、赤霉素和油菜素内酯的含量,但降低了根系茉莉酸甲酯含量(除Clariodeoglomous etunicatum)。
相关性分析揭示菌根诱导的福鼎大白茶根系激素变化与菌根促进福鼎大白茶侧根数有关。
此外,幼套球囊霉的促生效果最显著,而混合菌种对根系形态和侧根数影响最显著。
今后茶树栽培中应加强菌根管理。
-关键词:白茶,土壤微生物,脱落酸,共生真菌,侧根-中图分类号:; Q945文献标识码:; A文章编号:; 1000-3142(2018)12-1635-06在植物生长过程中,根系在养分和水分的吸收上扮演着十分重要的角色,但也经常受到植物激素、土壤酸碱性和土壤微生物的影响(Osmont et al, 2007)。
研究表明,接种丛枝菌根真菌(arbuscular mycorrhizal fungi, AM真菌)可改善宿主植物根系形态、促进生长和营养元素的吸收(Wu et al, 2011, 2017)。
在杨树上接种2种不同AM真菌均可显著增加二、三级侧根的长度和数量,而对一级侧根没有影响(Hooker et al, 1992)。
植物系统发育 丛枝菌根真菌群落

植物系统发育丛枝菌根真菌群落篇一:《神奇的植物世界:丛枝菌根真菌群落与植物系统发育》嘿!同学们,你们知道植物的世界有多奇妙吗?今天我就来给大家讲讲植物系统发育和丛枝菌根真菌群落那些超级有趣的事儿!想象一下,植物就像是一个个小小的王国,它们有着自己独特的成长方式和秘密。
而丛枝菌根真菌群落呢,就像是植物王国里的神秘助手。
咱们先来说说植物系统发育。
这就好比是植物们的家族族谱,记录着它们一代一代的变化和发展。
有的植物从很久很久以前就存在了,像那些古老的大树;而有的植物则是新出现的“小朋友”。
这难道不神奇吗?再看看丛枝菌根真菌群落,它们和植物的关系那叫一个密切!就好像是植物的好朋友,总是在默默地帮助它们。
比如说,植物要吸收营养,丛枝菌根真菌群落就会像勤劳的小蜜蜂一样,帮植物找到更多的养分,让植物能长得壮壮的。
有一次,我和小伙伴一起去植物园玩,看到一棵大树长得特别茂盛。
我就好奇地问老师:“老师,为啥这棵树能长得这么好呀?”老师笑着说:“也许是因为它有丛枝菌根真菌群落这个好帮手呢!”我当时就想,哇,这些小小的真菌居然有这么大的作用!你们说,丛枝菌根真菌群落是不是很像植物的超级英雄?它们虽然小小的,我们用肉眼都看不到,但是却能为植物做这么多事情。
而且呀,不同的植物和丛枝菌根真菌群落的关系还不一样呢!有的植物和它们是“铁哥们”,离不开;有的植物则只是偶尔需要它们的帮忙。
这就像我们在学校里,有的同学总是一起玩,有的同学只是偶尔一起做个活动。
植物系统发育和丛枝菌根真菌群落的关系,也像是一场精彩的舞蹈。
植物在不断地发展变化,而丛枝菌根真菌群落也在配合着它们的步伐,一起跳出美丽的旋律。
所以说,植物的世界真是充满了惊喜和奥秘!我们一定要好好保护它们,让它们能继续在这个地球上快乐地生长。
我的观点就是:植物系统发育和丛枝菌根真菌群落的相互作用太重要啦,我们得多多了解它们,才能更好地保护我们美丽的大自然!篇二:《神奇的植物与菌根真菌的奇妙世界》嘿!同学们,你们知道植物的世界有多神奇吗?今天我就来给大家讲讲植物系统发育里特别有趣的一部分——丛枝菌根真菌群落!先来说说植物吧,就像我们在学校里有不同的班级一样,植物也有各种各样的种类。
菌根化苹果苗木及其生产技术

栽培技术菌根化苹果苗木及其生产技术顾雨非,隋秀奇,郑建强(烟台现代果业发展有限公司山东烟台264003)随着科学技术的进步和人们对菌根认识的不断深入,菌根学理论研究日益引起世界各国学者的广泛关 注,菌根化在果树领域的应用技术研究也日趋深入。
且随着我国近年来苹果栽植面积的增加和老园改造、 矮化密植栽培技术的发展,培养健壮根系、提高苗木质量己经成为果树生产的基本要求。
菌根是土壤有益真菌与植物吸收根系共生形成的复合吸收器官,既具有植物根系的作用又具有某些专 化真菌的功能,在长期的进化过程中菌根已成为植物存活的决定因素及其生活的自然供养体系。
在现阶段 的研究中,丛枝菌根的研究可以占到整个研究体系的 60%以上,这与其在自然界的广泛分布与重要地位是分不开的。
对于苹果苗木来说,丛枝菌根也是最适宜在 生产中应用的,其对于提高苗木的营养吸收能力和抗 逆性至关重要。
1菌根在苹果苗木生产中的主要作用1.1提高苹果苗木对水肥的吸收能力丛枝菌根能够提高植物对各类矿质元素的吸收能 力,特别是移动性较差的P 、Zn 、Cu 等,能够加速有机质 的分解,提高对N 的捕获能力,从而提高肥料利用率。
1.2分泌苗木所需的植物生长调节剂,促进植物生长菌根真菌能够分泌多种酶和植物生长调节剂,且其 庞大的菌丝系统能够扩大植物的养分吸收范围。
菌根真菌的根外菌丝扩展到土壤中将土壤的矿质营养提供 给植物,也把宿主植物的碳水化合物提供给土壤和土 壤生物体,将植物和土壤有机地联系起来,形成良性循环系统。
1.3提高苗木的抗逆性,特别是对干旱、盐碱和重金属的抵抗能力菌根真菌的菌丝能直接吸收水分,随着根外菌丝的延伸和扩展,菌根真菌的侵染能增加宿主植物根长和 根密度或者改变根系形态,增强植物根系的吸收能力,扩大根的吸收范围,提高植物对干旱的抵抗能力;刘春 艳等研究表明菌根真菌通过改变植物体内碳水化合物、氨基酸组分和含量来改变根组织中的渗透平衡,提 高植物耐盐能力;菌根及其庞大的菌丝体网可在土壤中分泌大量的生物化学物质,改变植物根际环境及重金属的存在状态,以降低重金属的毒性。
《300种园林植物栽培与应用》读书笔记思维导图PPT模板下载

01
七、剪股 颖(见彩 图264)
02
八、狗牙 根(见彩 图265)
03
九、结缕 草(见彩 图266)
04
十、假俭 草(见彩 图267)
06
十二、大 花马齿苋 (见彩图 269)
05
十一、紫 茉莉(见 彩图268)
01
十三、德 国鸢尾 (见彩图 270)
02
十四、玉 簪(见彩 图271)
04
十六、荆 条(见彩 图226)
06
十八、九 里香(见 彩图228)
05
十七、紫 珠(见彩 图227)
第六章攀缘植物
01
一、台尔 曼忍冬 (见彩图 237)
02
二、南蛇 藤(见彩 图238)
03
三、凌霄 (见彩图 239)
04
四、紫藤 (见彩图 240)
06
六、茑萝 (见彩图 242)
05
五、络石 (见彩图 241)
七、枸骨 (见彩图 217)
02
八、阔叶 十大功劳 (见彩图 218)
03
九、狭叶 十大功劳 (见彩图 219)
04
十、石楠 (见彩图 220)
05
十一、海 桐(见彩 图221)
06
十二、紫 叶矮樱 (见彩图 222)
01
十三、金 叶莸(见 彩图223)
02
十四、珍 珠梅(见 彩图224)
03
十五、牛 奶子(见 彩图225)
02
八、南洋 杉(见彩 图105)
03
九、雪松 (见彩图 106)
04
十、日本 金松(见 彩图107)
05
十一、日 本五针松 (见彩图 108)
根-真菌(思维导图)
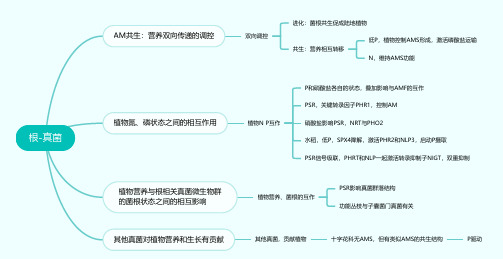
根-真菌AM共生:营养双向传递的调控双向调控
进化:菌根共生促成陆地植物
共生:营养相互转移
低P,植物控制AMS形成,激活磷酸盐运输
N,维持AMS功能
植物氮、磷状态之间的相互作用植物N P互作
P和硝酸盐各自的状态,叠加影响与AMF的互作
PSR,关键转录因子PHR1,控制AM
硝酸盐影响PSR,NRT与PHO2
水稻,低P,SPX4降解,激活PHR2和NLP3,启动P摄取
PSR信号级联,PHRT和NLP一起激活转录抑制子NIGT,双重抑制
植物营养与根相关真菌微生物群
的菌根状态之间的相互影响植物营养、菌根的互作
PSR影响真菌群落结构
功能丛枝与子囊菌门真菌有关
其他真菌对植物营养和生长有贡献其他真菌,贡献植物十字花科无AMS,但有类似AMS的共生结构P驱动。
丛枝菌根侵染率研究资料讲解

丛枝菌根研究方法一、检测孢子含量的方法A 湿筛倾注法1. 称取一定重量的土壤样品(最好是取自15cm 表层植物根系附近的土壤),放在容器内用水浸泡20-30min ,使土壤松散。
如果土壤粘性很大,也可加入各种土壤分散剂。
2. 选用一套洁净的具有孔径为0.5-0.034mm 的土壤筛,依次重叠起来。
最底层用一物体垫着(如培养皿、木块等物),使筛面稍微倾斜。
3. 用玻璃棒搅动浸泡的水溶液,停置几秒钟后,使大的石砾和杂物沉淀下去,即将悬浮的土壤溶液慢慢地倒在最上一层孔径最大的土壤筛上。
倾倒时,最好集中倒在筛面的一个点上,不要使整个筛面都沾有土壤溶液。
4. 用清水依次轻轻冲洗停留在筛面上的筛出物,以免在上层粗筛面的剩留物中夹藏有VA 菌根真菌孢子。
5. 用洗瓶将停留在筛面上的筛出物轻轻冲洗到一个清洁的培养皿里面,再将滤液通过细筛并用水冲洗。
在冲下来的筛出物中,除有许多细的沙砾和杂质外,就含有VA 菌根真菌的不同直径的孢子。
6. 将含有筛出物的培养皿放在双目实体解剖显微镜下观察。
B 蔗糖离心法1. 称取10 g 菌剂,置入大烧杯中加500 ml 水,搅拌,静置10 s。
2. 先后过80目分样筛、400目分样筛,将400目筛子上的残余物用药匙转入50ml 离心管中,后用清水冲洗筛子,将残余物全部转入离心管中,配平,3000转/min离心10 min。
(注分样筛最好直径为12 cm 左右,便于下面放置烧杯过筛)3. 去掉上清液,在离心管中加入预先配制好的质量分数为50%的蔗糖溶液,玻璃棒搅匀,配平,3000转/min离心10 min (注意离心前离心管壁上不能有残余物)。
4. 将400 目筛子呈一斜面放置,离心后的蔗糖溶液过筛子的下侧,用水将筛子上的残留物轻轻洗入划线培养皿中。
(注意水不能加太多以防影响检测,培养皿划线便于统计)。
5. 解剖镜镜检统计培养皿中的孢子数目,计算出菌剂中的孢子含量。
注:溶于蔗糖溶液中的孢子仍可进行接种。
丛枝菌根真菌

丛枝菌根真菌对香蕉试管苗的驯化07生物技术1班于澎0701024123日期:1999年5月24日摘要:丛枝菌根真菌的影响(AMF)的香蕉试管苗上进行了评估在驯化期。
植物接种无梗scrobiculata,绣球clarum和Glomus etunicatum。
在种植后温室3个月,株高,叶面积,鲜重和干物质的根,芽,AMF的殖民化的水平营养水平,光合作用和蒸腾率,水势和气孔导进行了测定。
丛枝菌根真菌孢子的生产数量在每个治疗也决心。
苗接种与丛枝菌根真菌具有更大的株高,叶面积和新鲜地上部和根系的重量,以及较高的光合作用和蒸腾比对照组。
植物与血管球接种均优于在最评估参数。
关键词:穆萨菌,内生菌根,菌根菌,气候适应引言:水果的营养快繁,观赏和森林物种,是一个良好的生产条件,转基因植物检疫植物和均匀大量的主要工具。
到温室栽培植物体外转移是在结构和生理适应的最重要的准备过程中试管苗的步骤之一。
这一阶段,由于水土不服,是一种对植物自养的存在开始,以期为生存所必需的生理过程的开始。
在这段时间内,必须增加水的试管苗和矿物质,光合速率的吸收。
试管苗,病免费的,但他们还缺乏丛枝(AMF)的菌根真菌。
AMF的是众所周知的增加,增加水和矿物营养素的吸收,尤其是磷(P)植物的活力。
此外,AMF的病原体可以保护寄主植物的根和减轻极端温度变化,pH值和水分胁迫(迪克森和马克思1987年的影响; Siqueira 1994年)。
接种AMF的成功在驯化期间(格兰杰等人的开始。
1983年; Brazanti等。
1992年;罗杰古勒明等。
1995年),甚至在体外培养已被证实。
三是与从组织培养植物的根系形成共生互利的效果表现在蓬勃植物的光合作用和蒸腾速率高,养分和水分,提高抗逆性。
接种丛枝菌根真菌在植物组培苗生长初期当然可以对体外培养,通过积极对rootmeristem活动菌根共生效应,高殖利率。
支持这个假说是由伯塔等人的结果。
(1995年),谁表明,AMF的协会改变了红叶李根的分枝格局。
丛枝菌根真菌生态学功能

从枝菌根真菌在陆地生态系统中的作用姓名:蒋胜竞专业:兰州大学植物学从枝菌根真菌在陆地生态系统中的作用生态系统主要强调生物界与非生物界的物质循环和能量流动,而AMF是在生态系统是介于生物界和非生物界的枢纽位置,它可以促进植物的营养吸收,因此,AMF在生态系统中有着很重要的作用。
AMF可与陆地上大部分植物形成共生体,通过菌丝与植物发生相互作用。
目前,对AMF的生态学研究大多集中在生物个体水平上,比如植物的生理,生长和繁殖。
然而,在生态系统水平上AMF的重要性确没有太多的报道。
AMF在生态系统研究中的一个难点就是很难去量化其物质的转移和能量的流动,也没有一种可行的方法去检测AMF 在生态系统中的生物量。
在生态系统模式甚至是土壤C循环的模式中都没有AMF的存在,我觉得这是不够完善的,因为每公顷土壤中AMF菌丝的C大约会有50-900Kg。
AMF的存在会通过影响植物的群落结构、土壤微生物的群落结构、植物的生理特性以及土壤有机质来影响生态系统,这四个过程是彼此独立且又相互联系的,比如AMF所引起的植物群落结构的变化也会引起地下微生物群落的变化。
现在我就分别具体地介绍下着四个途径。
1 AMF可以通过影响植物群落的组成从而间接的影响生态系统一些报道已经证实了AMF的存在和群落结构会对植物的群落结构产生强烈的影响,AMF 对植物群落的影响有积极的也有消极的,其对植物多样性的影响主要在于其菌根依赖性的植物在植物群落中有什么样的生态位。
比如,在植物群落中如果优势物种是高菌根依赖性的,那么当菌根真菌消失时,就会增加植物群落的多样性。
此外,AMF的存在也会改变植物间的相互竞争作用。
在植物生态中,关于植物群落的特性对生态系统所产生的影响有很多的数据和理论框架,比如植物的群落结构可以影响生态系统中的营养结构及对资源的利用。
然而对于AMF所介导的植物群落的变化所引起生态过程的变化这方面一直缺乏实验性数据。
AMF 和植物的群落结构是相互独立的两个个体,但在自然界中却是有着共同变化趋势的,因此,很难区分生态过程中的一些变化是由于AMF造成的还是由于植物群落结构造成的。