枯萎菌毒素粗提液对4种陆地棉外植体愈伤组织诱导率的影响
不同浓度激素处理对棉花愈伤组织诱导的影响(1)

筛选,得出适宜培养基组成:MS无机盐+B5有机物+
0.1 mg/L KT+0.05 mg/L IBA+0.1 mg/L
图I
20天不同激素配比对愈伤组织诱导的影响
2,4-D(pH
6.4—6.5)。
3讨论
愈伤组织的形成过程是一个受培养基成分、外植
光照条件是影响愈伤组织诱导和生长发育的重要
成功的关键。现虽已从多种外植体获得愈伤组织,但
大量的试验结果表明,棉花下胚轴是获得愈伤组织的
较好的外植体,而且幼嫩组织比老化组织易诱导,一般 选择幼嫩下胚轴切段,长度在0.5—0.8 cm较佳。影
响愈伤组织诱导的主要因素是外植体种类、不同激素
天津科密欧化学品有限公司,其他常规试剂均购自天
津化学厂。 1.2实验方法 选取成熟饱满的百棉l号种子,脱绒剥取种皮,用 75%的酒精浸泡l min,用0.1%的HgCl:浸泡5—8 rain,进行表面消毒,在无菌环境下用无菌水漂洗4—6 次,每次5 rain。在超净工作台上将浸泡好的棉花种胚
SHEN Qiu.xiul,HU Gen.hai2,ZHAO
Cotton Callus Inducement
Wei.wei2,RAN Jing-chen93
摘要:选用百拂l号的下胚轴为外植体,在MSB培养基中分别 添加不同浓度组合的2,4.D、KT、IBA作为诱导培养基,以MSB 为继代培养基,研究不同激素处理对棉花体细胞发生的影响。 结果表明,适合百棉1号出愈2,4一D的浓度在0.05—0.1,ns/L 之间;KT的浓度在O.05—0.1 ms/L之间;IBA的浓度在O一
对照。诱导培养基的设计是按照k(34)正交试验设 计的,详见表l。
表l诱导愈伤组织培养基激素组成
稻瘟病菌粗毒素对水稻成熟胚愈伤组织诱导的影响及抗性筛选

经 毒 素筛 选 获得 的再生绿 苗 收获的 R 种子播 于秧 盘或 小钵 中,在 三叶一 心 期 ,进 行人 工 接种 鉴 定 ,采 1
用 各种 菌株 的分生 孢子悬 浮液 ( ×l5 子I 1 2 0孢 m )进 行 喷雾 接菌 ,每 10株 苗 喷 | ,保 湿 7一l 后 观察植 0 l 1 l 0天
维普资讯
第2 6卷第 5期
V 12 N . 0.6 o5
长春师 范学 院学 报 ( 自然 科学版 )
J ra o l cu o a n e i ( a rl c ne on l f mc hnN ma Ui rt N t a Si c ) u C l v sy u e
2 0 年 1 月 0r 7 O
Oc . 0O7 t2 r
稻 瘟 病 菌 粗 毒 素 对 水 稻 成 熟 胚 愈 伤 组 织 诱 导
的影 响及 抗 性 筛 选
高立宏 ,汉 丽 萍 ,辛树 权
( 春师范学 院生命科 学学 院 ,吉林长 春 长
[ 摘
10 2 ) 30 1
要]利 用稻瘟病不同生理小种菌株的液体培养 提取 的粗毒素对水稻成熟胚进行 处理 ,粗毒素液
・ 7 - 2
,
维普资讯
13 2 诱 导培 养基 中加入加 入稻 瘟病 菌毒素对 成熟胚 诱导及 分化 的影响 ..
取饱 满籽粒 灭菌 ( 方法 同上 )然后用 解剖 刀切下 种胚 ,分别接 种于含 有不 同浓度 的混合 的稻瘟 病 菌粗毒
素 的诱 导培养 基 上 ,每 瓶接种 l 2个 ,共 接种 4瓶 ,在 2 ℃条 Байду номын сангаас 下 暗培养 3 d 6 0 ,统计 其 诱 导 率 ;再 把愈 伤 组 织转 入分化 培养基 中培 养 2 d 5 ,统计 分化成 苗率 。 14 再生植 株 的抗病 性鉴定 : .
防治草莓枯萎病菌4种药剂的筛选试验

[] 6 张常隆 , 李扬苹 , 冯玉龙 , 表型可塑性和局域适应在紫茎泽 兰 等. 入侵不同海拔生境 中的作用[] 生态学报 , 0 , ( )14 J. 2 92 4 : 0— 0 9 9
14 9 6.
泽兰可较容易形成单优群落 。之后 , 以密集丛生的方式使各植 株相互遮阳 , 阔地蚕食 , 向开 从而打破 光照对其 入侵 的限制。 在开阔地中生长的紫茎泽兰的冠层高度一致性非常高 , 上部较
制的指示意义 [ ] J .植物生态学报 ,0 5 3 3 3—39 2 0 ( ):7 7.
于红 梅等 : 防治草莓枯萎病菌 4 种药剂 的筛 选试 验
制效果 。用体 积 比为 5: 5的丙酮无菌水溶 解表 1 所列 杀菌 剂各 处 理 质 量浓 度 1 L, 入 灭 菌 冷却 至 4 0m 加 5℃ 左 右 的 3 LP A 0m D 培养基中 , 充分 混匀后 均 匀到入 3个 灭 菌的培养 皿中, 制备平板 , 以等量 的无菌水作空 白对 照 。将制备 的 菌饼接种 于 4种杀菌剂稀释不同浓度处理 的 P A平板上 , D 每
表2 4种杀菌剂对草莓枯萎病菌的抑制效果
[] 3 邱 强 , 淼, 胡 王志 田, 等.原色西瓜甜瓜草莓病虫 与营养 诊断 图谱 [ . M] 北京 : 中国科学技术出版社 , 9. 1 6 9 [] 4 元菊丹 , 凯 , 山, 1 种杀菌剂对生姜腐霉茎基腐病菌 冯 齐军 等. 3 的毒力 测定 E] J.农药 , 1, ( )67 6963 2 15 8 :1 — 1,2. 0 0 [] 5涂 勇, 姚 昕, 余前嫒 , 等.不同杀菌剂对青枣炭疽病 菌的室 内
表 l 供试杀菌剂通用名 、 剂型及生产厂 家
p rm f p faai ) ou . r r e 从根部侵染维管束引致 的系统性土传 .s g a
稻瘟病菌粗毒素对水稻成熟胚愈伤组织生长和分化的影响及抗性筛选

1 材 料与 方法
1 1 供 试 品种 :丰 优 3 1 . 0 、丰 优 3 7 0 、优 9 —8 3 、农 大 3 、农 大 1 。 由吉林 农业 大学 水稻 研室 提供 。 9 12 稻 瘟病 菌 的培 养 . 12 1 进 行 预培 养 的菌种及 来 源 .. 3 —1 7 7 —1 ,90 0 —2 9 2 1 ,9 10 3 ,A ,17 —2 93 3 ,9 00 —2 9 63—2 9 4 , 13—1 ,由 吉林 省 农 业 科 学 ,w ,z8 6 —4 院植 保所 提供 。 12 2 利用 MS培 养液 培养 稻瘟 病菌 ..
.
2 分 化 培养 基 中加入 毒 素 。把 愈 伤组 织 转 入 含 有 不 同 浓度 的 混合 的稻 瘟 病 菌粗 毒 素 的 分 化 培 养 基 中 , . 每瓶 接种 1 愈伤 组织 ,共 接 种 2瓶 ,在 2  ̄ O块 6C、20hx 照条件 培 养 2d后 ,统 计 分化成 苗 率 。 00 光 5
强。水稻成熟胚组织培养 中进行抗稻瘟病 菌突变体筛选 的毒 素浓 度在 2 % ~3 %较好 ;愈伤组织 浸泡 5 o 1 ~2 h 以得 到分化出苗。在愈伤组织上直接加上毒素并 长时间处理 对愈伤组织 的生 长有 很大 的破 2 4可 坏作 用 ,每个 愈伤组织滴加 的毒素应小于 20 l .m 。不同的水稻 品种产生 的耐性不 同 ,可 以筛 选 出抗稻
瘟 病的突变体 。
[ 关键词 ]稻瘟病 菌粗毒素 ;成熟胚 愈伤组织 ;抗性筛选 [ 中图分类号 ]Q 4 93 [ 文献标识码 ]A [ 文章编号]10 —18 (0 8o —07 o 0 8 7x 2o )3 00一 4
棉花农杆菌介导转化体系中影响抗性愈伤组织诱导率的因素(1)

2009,21(4):271~274
棉花农杆菌介导转化体系中影响抗性愈伤组织诱导率的因素
赵常燕,王省芬。韩改英,迟吉娜,张桂寅,马峙英’
(河北省作物种质资源重点实验室,河北农业大学,河北保定071001) 摘要:以冀无2031为材料,采用正交设计方法,研究了农杆菌介导的棉花下胚轴遗传转化过程 中菌液浓度、侵染时间、共培养时间和共培养温度对抗性愈伤转化效率的影响。结果表明,4
1
材料和方法 供试棉花品种为陆地棉冀无2031、珂312、中
1.1试验材料
521和农大94—7,由河北农业大学棉花遗传育种 研究室提供,所有品种均经多年自交。其中,冀无 2031用于优化转化体系的筛选,其它3个品种用 于验证筛选出的最佳组合。大肠杆菌菌株为
E.coli
1.2.4农耔菌侵染和共培养。用制备好的菌液 侵染下胚轴。弃去菌液,用无菌滤纸将表面菌液吸 干,接种于共培养培养基中培养。 1.2.5愈伤绍织诱导及筛选。将共培养后的下 胚轴接种于含抗生素的愈伤组织诱导培养基上, 培养1~2个月。统计抗性愈伤组织的诱导率。 1.2.6数据处理。抗性愈伤组织诱导率(出愈 率)一(诱导出愈伤组织外植体数/接种的外植体
93.85%、86.72%和85.83%。
表3不同因素卡那抗性愈伤组织诱导率的方差分析
Table 3 Variance analysis of the induction ratio of
另外,本研究还发现初始诱导的愈伤组织状态很
差,呈淡黄色、质地湿润或呈绿色有硬块。调节培
kanamycin-resistant calli for different factors
个因素均对抗性愈伤的诱导率有极显著影响,其诱导效应依次是共培养温度>菌液OD值>
棉花自毒物质对幼苗生理作用和棉花枯、黄萎病菌丝生长的影响

(n tt t fEc n mi Cr p , n in a e fAg iut r l ce c s Ur mq 3 0 1 Chn ) I siu eo o o c o s Xij gAc d my o rc lu a in e , u i 0 9 , ia a S 8
c n r ton u e he e f c xta t d s u i n. e t a i nd r t fe tofe r c e ol to The a tv t u v fS0D, c i iy c r e o CAT nd POD is o e t n a fr t r s he do r ppe t o h c v fVe tc li d;he gr wt ur e o r i ilum it w l myc lu fr t r e t n d o e I s a l z d t att e e i m is os he r pp d; twa na y e h h n r a o h a d a o pto fc t o e d i s r ots t m r m p d d b h o t xt a t d s l o m lgr wt n bs r i n o o t n s e lng o ys e we e i e e y t e c ton e r c e o u— to .The a tv t fSOD , in c i iy o CA T nd POD fc ton s e ln a t ha e t e gr wt oton s e ln a o o t e d i g pl n s c ng d, h o h ofc t e d i g wa nh bie n oto d a t t x ct fe t s i i t d a d c t n ha u o o i iy e f c . Ke r s: c t n pl n ;a t t xiiy;phy i l ia ;Fu a i m l n r i ilu wit y wo d oto a t u o o c t so og c l s r u wit a d Ve tc l i m l my e i m c lu
陆地棉愈伤组织诱导影响因素初探
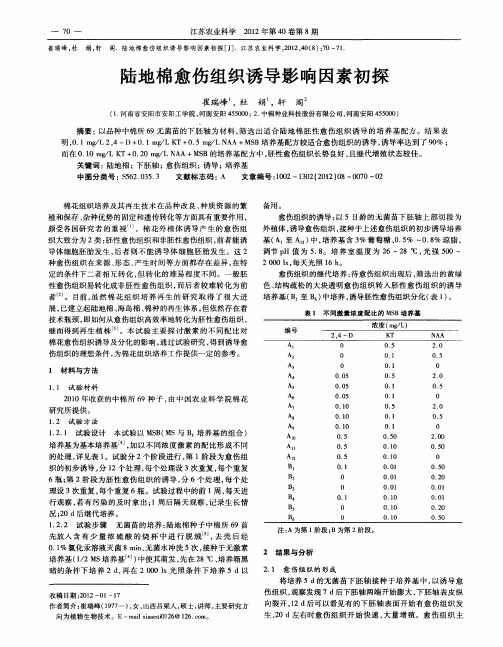
愈伤组织的继代培养 : 待愈伤组织 出现后 , 筛选出的黄绿 色、 结构疏松 的大块透明愈伤组织 转入胚性 愈伤组织 的诱 导 培养基 ( 至 B ) B 中培养 , 诱导胚性愈伤组织分化 ( 1 。 表 )
一 7 l一
要有淡黄色 、 淡绿色 、 深绿色 和 白色等 , 质地有疏 松 、 紧密 、 颗
粒状等 。 由表 2可以看出 , 同处理 的愈 伤组织诱 导率及 其愈伤 不 组织状态明显不同。诱 导率 较高的为附加 0 10 5mgL24 . 、. / ,
将培养 5d的无菌苗 下胚轴 接种于培养 基 中, 以诱 导愈 伤组织 , 观察发现 7d 后下胚轴两端开始膨大 , 下胚 轴表皮纵 向裂开 ,2d后可以看见有 的下胚轴表 面开始有愈伤组 织发 1 生 ,0d左右时愈伤组织 开始 快速 、 2 大量 增殖 。愈 伤组织 主
崔瑞峰等 : 陆地棉愈伤组织诱导影 响因素初探
收稿 日期 :0 2一 l—l 21 O 7 作者简介 : 崔瑞峰 (9 7 ) 女 , 17 一 , 山西 吕梁人 , 硕士 , 讲师 , 主要研究方 向为植物生物技术 。E—m ixaci16 2 .o al i uO 2 @16 tm。 o
2 结 果与 分 析 2 1 愈 伤 组 织 的形 成 .
表 1 不 同激 素 浓度 配 比 的 MS B培 养 基
棉花愈伤组织诱导及分化 的影 响, 通过试验研究 , 得到诱导愈
伤 组 织 的理 想 条 件 , 棉 花 组 织 培 养工 作 提 供 一 定 的 参 考 。 为
1 材 料 与 方 法
1 1 试 验 材 料 .
旱芹粗提物对棉花枯萎病菌丙二醛含量、电导率及保护酶活性的影响

t e m e r n i i sp r x d z d s ro sy a t rb i g t e t d wih t x r c i n, l n t r a e i e p o h mb a e l d wa e o i ie e i u l , fe e n r a e t hee t a t p o a o g wi t e t d t r - h m
t ie f rtci n y si ua im xsou fs .vs fcu w r tde . h eut idctd ta i t s oe t ee zme F sr vi o p v n u oyp rm .p ai etm ees i T ersl iae h t n u d sn
F s ru x s ou f s .a ifcu u a im o y p r m .p v sne tm
W e q a g, Xi i i iLi i n e Hu q n, Ya g De o g n s n , Li i n i S n Le uJ a we , u i
棉花黄萎病菌毒素诱导植株产生抗病性的机理

12 6 毒素诱导后棉苗叶绿素分 析 ..
12 6 1 叶绿素含量测定 ... 取新 鲜 的叶片 0 5 g 剪碎研磨 . ,
后 加 入 5m L乙醇 研 磨 至 组 织 发 白 , 5m n 过 滤 , 乙 醇 3— i 后 用 定 容 至 10m , 别 于 6 5 6 94 0n 测 吸 光度 。3种 叶 绿 0 L分 6 、4 、7 m
加 入 2 5m . o L双 氧水 , 于 3 . L0 1m l / 置 0℃ 水 浴 1 i。然 后 0r n a
的茎 和叶片扩展 , 终侵染 整个植株 J 最 。依据 棉花植株 依
靠毒素侵染产生抗病性 的生理特性进 行抗性筛选是培育棉花 抗性植株 的一种重要方法 。植物抗 病性诱导分 3个阶段 : 诱 导处理 , 号传递 , 信 抗性 表达 。其 中任何一个 阶段都与基础理 论有重要关系 , 因而研究新疆 棉花黄萎病菌 毒素诱导棉 花产 生抗 性 的机 理 尤 为 重 要 。
一
12一 0
江苏农业科学
21 0 2年第 4 0卷第 8期
李
凤, 耿雅文 , 孙健 富, 等.棉花黄萎病菌毒素诱导植株 产生抗病性 的机理[ ] J .江苏农业科学,0 2,0 8 :0 2 1 4 ( ) 12—15 0
棉花黄萎病菌毒素诱导植株产生抗病性 的机理
李 凤, 耿雅文 , 孙健 富, 焦天奇 , 欧秀玲, 王爱英
1 材 料 与 方 法 1 1 材 料 .
1 号至 3号瓶分别加 1 %硫酸 25m , 0 1m lm 0 . L 用 . o L高锰酸 /
钾滴定 至粉红色 ( 酸碱滴定法) 记 录结果 。 ,
12 52 脯氨酸含量测 定 ... 分别取 不 同处理 的叶片 0 2g . , 加 2 0mL的 8 % 乙醇 和少 量 石 英研 钵 中冰 浴 研 磨 , ℃、 0 4 1 0 mn离心 2 i , 上清 液。每种 上清液 加入 冰醋 000r i / 0m n 取 酸 2m , 三酮 2m , L茚 L 水浴 1 i 5rn后置于 55n a 1 m下 比色 , 并 无菌 操作条件下 将新疆棉
萱草叶枯病菌粗毒素液对分化芽生理生化代谢的影响

山西农业科学2019,47(10):1725-1729萱草叶枯病菌粗毒素液对分化芽生理生化代谢的影响杨丽莉,杨睿,常建忠,蒋丹(山西省农业科学院旱地农业研究中心,山西太原030031)摘要:研究了萱草叶枯病菌毒素液对大花萱草愈伤组织分化芽不同生长时间内生长量、成活率和保护酶(SOD 、POD 、CAT )活性的影响。
结果表明,随着病菌毒素液剂量的增加毒害作用增强;添加40%叶枯病菌毒素液为抗病筛选的半致死剂量,培养24d 后,成活率达50%左右;叶枯病菌毒素液对SOD 、POD 和CAT 这3种酶活性的影响均表现为先升高后降低,活性升高的程度与病菌毒素液的浓度呈正相关,其中,POD 酶活性峰值出现在第7天,SOD 和CAT 酶活性峰值出现在第2天。
关键词:大花萱草;萱草叶枯病菌;病菌毒素液;生长状态;保护酶活性中图分类号:S682.1+9文献标识码:A文章编号:1002-2481(2019)10-1725-05Effect of Toxin Liquid of Extracted fromon Physiology,Biochemistry and Metabolism of Differentiated BudsYANG Lili ,YANG Rui ,CHANG Jianzhong ,JIANG Dan(Research Center of Dryland Farming ,Shanxi Academy of Agricultural Sciences ,Taiyuan 030031,China )Abstract :In this paper,the impact of bacterial toxin liquid of Kabatiella microsticta extracted from Hemerocailis hybridai on the amount of growth,survival rate and protective enzymes (SOD,POD,CAT )activity of Hemerocailis hybridai callus differentiation buds was studied at different growth times.The result showed that the toxic liquid effect enhanced with the increase of toxin dose.40%of Kabatiella microsticta toxin liquid was added to be half lethal dose for disease resistance screening.24d after culture,the survival rate reached approximately 50%.The impact of Kabatiella microsticta toxin liquid on the enzymatic activity of SOD,POD and CAT showed increase first and then decrease in different levels.The degree of increased activity was positively correlated with concentration of Kabatiella microsticta toxin liquid.The peak value of POD enzymatic activity was found in the 7th day,while SOD and CAT appeared on the 2nd day.Key words :Hemerocailis hybridai ;Kabatiella microsticta ;bacterial toxin liquid;growth state;protective enzyme activity收稿日期:2019-05-22基金项目:山西省重点研发计划项目(201603D221017-1);山西省农业科学院特色农业攻关项目(YGG17037)作者简介:杨丽莉(1964-),女,河北唐山人,研究员,主要从事生物技术种质资源创制研究工作。
拟轮枝镰孢菌毒素粗提液对玉米种子发芽及幼苗生长的影响

拟轮枝镰孢菌毒素粗提液对玉米种子发芽及幼苗生长的影响作者:刘春元邢小萍韩卫丽来源:《安徽农业科学》2019年第19期摘要为建立玉米杂交种和自交系利用拟轮枝镰孢菌毒素抗性筛选的试验方法,制备拟轮枝镰孢菌毒素粗提液,测定不同稀释倍数毒素粗提液对2个玉米杂交种和4个玉米自交系处理4 和6 d后的种子发芽抑制率、胚根生长抑制率以及幼苗致萎效果,分析不同抗性玉米种质、毒素稀释倍数和处理时间对玉米种子发芽、胚根和幼苗生长的影响。
结果表明,易感杂交种豫玉22与抗性杂交种农大108的种子发芽抑制率和胚根生长抑制率均存在极显著差异(P<0.01);在处理后4、6 d时,前者的种子发芽抑制率、胚根生长抑制率分别比后者高7.2%、11.3%、12.7%、11.8%。
在对玉米杂交种幼苗致萎蔫试验中,用稀释2倍和5倍的毒素粗提液处理,豫玉22与农大108杂交种幼苗间的萎蔫程度也存在显著差异(P<0.05)。
拟轮枝镰孢菌毒素粗提液对不同抗性玉米种质种子发芽、幼苗生长的影响存在显著差异,为利用拟轮枝镰孢菌毒素粗提液对玉米品种进行苗枯病抗性鉴定和筛选提供理论依据。
关键词玉米苗枯病;拟轮枝镰孢菌;毒素粗提液;发芽率;胚根生长中图分类号 S432.1文献标识码 A文章编号 0517-6611(2019)19-0158-04doi:10.3969/j.issn.0517-6611.2019.19.046Abstract In order to establish a simple method for evaluating the resistance of maize hybrids and inbred lines, the crude toxin extract from the culture medium of Fusarium verticillioides was prepared , and the seed germination inhibition rate, radicle growth inhibition rate, as well as the wilting degree of 2 maize hybrids and 4 inbred lines under the treatment of the crude toxin at various dilutions and different treatment duration (4 or 6 d) were determined. The results showed that the susceptible hybrid Yuyu22 and resistant hybrid Nongda108 significantly differed in the inhibition rates of seed germination and radicle growth (P<0.01), and the inhibition rates of seed germination and radicle growth of Yuyu22 were 7.2%, 11.3% and 12.7%, 11.8% higher than those of Nongda 108. An obvious difference between Yuyu22 and Nongda108 was also found under the treatment of the crude toxin extract at 2 and 5 diluted solution (P<0.05).The effects of crude extracts of fusarium rotatum toxin on seed germination and seedling growth of different resistant maize germplasms were obviously different, which provided certain basis for identification and screening of resistance to bacterial blight of maize varieties by using crude extracts of fusarium rotatum toxin.Key words Maize seedling wilt disease;Fusarium verticillioides;Crude toxin extract;Germination rate;Radicle growth玉米苗枯病是玉米生產中发生广泛、为害严重的病害之一。
棉花抗枯萎病抗性转化机理的初步研究

棉花抗枯萎病抗性转化机理的初步研究新陆早7号和新陆早16号棉花品种对棉花枯萎病菌( Fusarium oxysporum f.sp.vasinfectum)7号生理小种表现高度感病。
本文以这两个棉花品种和其经过病圃定向筛选3年、2年对枯萎病抗性有明显提高的材料为研究对象,运用液体培养棉苗孢子悬浮液浸根接种的方法,针对接种后抗、感材料的病理学变化和差异表达的基因进行了研究,旨在从病理学和分子生物学角度揭示感病棉花品种经过病圃加压定向筛选后抗性发生转化的机理。
1、通过对2种不伤根的棉花枯萎病接种方法,即全生育期的田间病圃鉴定法和液体培养棉苗孢子悬浮液浸根法进行对比,结果表明,液体培养棉苗孢子悬浮液浸根法(接种浓度为107个分生孢子·mL-1)全周期只需要30天,通过这种方法,可以在较短时间内鉴定出棉花品种对枯萎病菌的抗、感病性。
对比不同浸根接种时间(10、20、30、40、50、60 min)对结果的影响,发现浸根接种40 min可加快发病,鉴定结果和田间病圃鉴定的结果基本一致。
2、通过电子显微镜和光学显微镜观察接种后抗、感材料根部组织的病理学变化,发现二者在宏观和微观病理学方面均存在一定差异。
在宏观病理学方面的差异表现为:感病品种接种7天开始显症,25天严重发病,整个植株维管束变色;但抗病材料生长健康或只有轻微症状,维管束变色部位只局限在子叶节以下。
在细胞和组织病理学方面的差异为:侵入后前期,抗病材料被侵入的细胞中有细胞壁加厚现象;侵入中期,病菌可通过表皮层侵入感病品种的薄壁组织,却只能到达抗病材料的皮层组织中;侵入后期,抗病材料中出现了大量寄主细胞的降解物质或分泌物将菌丝包围,阻止了病菌进一步扩展。
抗病材料无论抗侵入还是抗扩展的能力都比感病品种大大增强。
3、利用差异显示PCR技术对接种枯萎病菌前后的抗、感材料中差异表达的基因进行了初步研究。
结果显示,反转录获得cDNA第一链,用3个锚定引物和4个随机引物组合扩增的DDRT-PCR产物,经测序胶电泳分离回收,二次扩增共得到约62个差异条带。
枯萎病菌毒素粗提液对美洲南瓜幼苗生长及生理的影响

中国瓜菜2023,36(4):81-88美洲南瓜(Cucurbita pepo)是葫芦科南瓜属一年生蔓性草本植物,因其营养成分种类多,药用和食用价值极高,在我国很多地区都有种植[1-2]。
甘肃省武威市是美洲南瓜重要的育种基地,美洲南瓜也是当地重要的经济作物之一,生产的南瓜籽远销海外[3]。
瓜类枯萎病(Fusarium wilt)是由镰孢菌引起的土传病害,也是瓜类生产中的毁灭性病害[4]。
其中,以西瓜发病最为严重,其次是南瓜,一般发病率为20%~30%,严重时达80%[5],有时在连作地甚至造成全田死亡。
因此,瓜类枯萎病是瓜类生产上的一大难题,也是限制美洲南瓜生产的重要因素之一[6]。
前人研究表明,引起甘肃省武威市美洲南瓜枯萎病的致病枯萎病菌毒素粗提液对美洲南瓜幼苗生长及生理的影响刘佳,柴改凤,郭风清,张树武,徐秉良(甘肃省农作物病虫害生物防治工程实验室·甘肃农业大学植物保护学院兰州730070)摘要:为了揭示美洲南瓜枯萎病菌产生的毒素粗提液对幼苗的影响,研究了病原菌产毒的条件以及不同浓度毒素粗提液对美洲南瓜幼苗生长及生理的影响。
结果表明,美洲南瓜枯萎病菌在pH值=7的Czapek培养液中24h连续光照、振荡培养15d时,产毒效果最明显,对美洲南瓜种子胚根生长抑制率最高,毒素粗提液亦具有较强的热稳定性。
80%的病原菌毒素粗提液,对美洲南瓜株高、茎粗和根长的生长抑制效果最好。
处理11d时,80%毒素粗提液处理下,美洲南瓜幼苗根系和叶片的细胞膜透性分别达到62.71%和62.31%;叶片MDA含量为对照的2.61倍,叶片PAL活性最高,为84.25U·g-1,是对照处理PAL酶活性的2.09倍。
研究结果可为揭示美洲南瓜枯萎病菌的致病机制提供一定的理论依据。
关键词:美洲南瓜;枯萎病菌;毒素;生长;细胞膜透性;生理指标中图分类号:S642.6文献标志码:A文章编号:1673-2871(2023)04-081-08Effects of crude toxin extract in the pathogen causing Cucurbita pepo Fu-sarium wilt on the growth and physiology of seedlingsLIU Jia,CHAI Gaifeng,GUO Fengqing,ZHANG Shuwu,XU Bingliang(Biocontrol Engineering Laboratory of Crop Diseases and Pests of Gansu Province/College of Plant Protection,Gansu Agricultural University,Lanzhou730070,Gansu,China)Abstract:In order to reveal the effect of crude toxin produced by pathogen causing Cucurbita pepo Fusarium wilt,the toxin-producing conditions of Fusarium oxysporum and the effects of different concentrations of toxin crude extract on the growth and physiology of Cucurbita pepo seedlings were studied.The results showed that:In the Czapek medium of pH=7,24hours of continuous light and shaking culture for15days,the toxigenic effect was the most obvious,and the growth inhibition rate of the radicle of Cucurbita pepo seeds was the highest.The crude toxin extract has strong thermal stability.80%of the toxin crude extract had the greatest growth inhibitory effect on plant height,stem diameter and root length.Under the treatment of80%crude toxin extract,the cell membrane permeability of Cucurbita pepo seedlings roots and leaves reached62.71%and62.31%on the11th day of treatment,respectively;The content of MDA in leaves was2.61times that of the control,and the activity of PAL in leaves was the highest,which was84.25U·g-1,which was2.09times that of the control.The research results provided the theoretical basis for revealing the pathogenic mechanismof Cucurbita pepo Fusarium wilt.Key words:Cucurbita pepo;Fusarium wilt;Toxin;Growth;Cell membrane permeability;Physiological indicators收稿日期:2022-09-08;修回时期:2022-12-01基金项目:2021年甘肃省教育厅青年博士基金项目(2021QB-025);甘肃农业大学科技创新基金(GAU-XKJS-2018-156);国家自然科学基金(31460518)作者简介:刘佳,女,讲师,研究方向为植物与病原菌互作。
棉花黄萎病菌毒素对棉花生化代谢的影响

棉花黄萎病菌毒素对棉花生化代谢的影响侯丽娟;李卫;刘燕霞;杨家荣【期刊名称】《西北农业学报》【年(卷),期】2010(019)012【摘要】为明确棉花黄萎病菌毒素对棉花生化代谢的影响,采用棉花黄萎病菌毒素处理棉花黄萎病抗病和感病品种的棉苗后,测定棉株体内超氧化物歧化酶(SOD)、过氧化物酶(POD)、苯丙氨酸解氨酶(PAL)的活性以及丙二醛(MDA)和木质素含量的动态变化.结果表明,抗病品种陕7191对病菌毒素的抗性显著高于感病品种中棉19;接种毒素后2品种棉苗叶片SOD活性都降低,感病品种降幅高于抗病品种;感病品种POD活性升高较快,到达峰值后迅速下降,而抗病品种POD活性持续上升;2品种PAL活性变化趋势相似,接种后抗病品种活性始终高于感病品种;2品种MDA和木质素含量均增多,感病品种MDA含量的增幅和增量高于抗病品种.【总页数】5页(P63-67)【作者】侯丽娟;李卫;刘燕霞;杨家荣【作者单位】西北农林科技大学,植物保护学院,陕西省农业分子生物学重点实验室,陕西杨凌,712100;西北农林科技大学,植物保护学院,陕西省农业分子生物学重点实验室,陕西杨凌,712100;西北农林科技大学,植物保护学院,陕西省农业分子生物学重点实验室,陕西杨凌,712100;西北农林科技大学,植物保护学院,陕西省农业分子生物学重点实验室,陕西杨凌,712100【正文语种】中文【中图分类】S435.621.24【相关文献】1.棉花黄萎病菌毒素诱导植株产生抗病性的机理 [J], 李凤;耿雅文;孙健富;焦天奇;欧秀玲;王爱英2.培养条件对棉花黄萎病菌毒素产生的影响 [J], 吴蔼民;夏正俊3.棉花黄萎病菌液体培养特性及其毒素的生物测定 [J], 侯丽娟;李卫;刘燕霞;杨家荣;毛岚4.棉花黄萎病菌毒素对棉花作用机制的初步探讨 [J], 章元寿;王建新5.用棉花黄萎病菌毒素快速检测棉花抗病性研究初报 [J], 章元寿;王建新;顾本康;李经仪;朱月英因版权原因,仅展示原文概要,查看原文内容请购买。
【棉花种植】滴二水前后枯黄萎病再不防就没机会了

【棉花种植】滴二水前后枯黄萎病再不防就没机会了这几天,有些农户都准备进二水了,有些往年有病的棉田在滴完头水、二水后陆续有枯萎病的发生。
随着天气慢慢转暖,棉田进水后,肥料刺激棉花开始快速生长,随着蒸腾作用棉田中的枯萎病病菌也开始大量从土壤中通过根系向主茎传导,根据这几天农之道微信大讲堂里许多学员反应,有很多棉田都出现了枯萎病。
枯萎病到底是怎么发生的呢?今天农之道小编就和大家再次谈谈枯萎病的发生及如何防治。
棉花感染枯萎病后,轻者减产10%~20%,重者减产40%~50%,甚至绝收。
一旦发生将会给棉农带来巨大损失,因此在田管中需要重点关注。
棉花枯萎病幼苗至成株均可发病,现蕾前后发病最盛。
可归纳为5种类型:(1)黄色网纹型:病株叶脉变黄,叶肉保持绿色,叶片局部或大部分呈黄色网纹状,叶片逐渐萎缩枯干。
(2)黄化型:叶片边缘局部或大部变黄,萎缩枯干。
(3)紫红型:叶片局部或大部变紫红色,叶脉也呈紫红色,萎缩枯干。
(4)青枯型:叶片突然失水,叶色稍变深绿,叶片变软变薄,全株青干而死亡,但叶片一般不脱落,叶柄弯曲。
(5)皱缩型:5-7片真叶时,大部分病株顶部叶片皱缩,畸型,色深绿,节间缩短,比健株矮小,一般不死亡。
病株根茎剖面木质部变成黑褐色。
枯萎病从棉花苗期至吐絮期都可能发生,但以盛蕾期为发病高峰期,最为常见的为黄化型、网纹型和青枯型。
枯萎病病菌寄生的寄主、载体很多,但以老棉田发病的土壤中隐藏最深,存活时间长达6~8年。
棉花生育期间,若遇持续阴雨天气,病菌(尖孢镰刀菌)很快就可从棉花根部的根毛、表皮伤口处入侵,随即急速暴发,来势迅猛。
棉花枯萎病的发病与温湿度关系密切,一般土温在20℃左右开始显症,土温上升到25℃-28℃时,形成发病高峰,当土温上升到33℃以上时病症受抑,出现暂时性隐症,入秋后待土温下降到25℃左右时,又出现第二次发病高峰。
夏季暴雨或多雨年份,因土温下降,发病严重。
地势低洼、土质粘重、偏碱、排水不良、偏施氮肥、棉田连作、耕作粗放的棉田重发。
抗生素对新疆棉花愈伤组织诱导和生长的影响

新疆农业大学学报 1998,21(4):267~269J ou rna l of X inj iang A g ricu ltu ra l U n iversity抗生素对新疆棉花愈伤组织诱导和生长的影响王冬梅 郝秀英(新疆农科院核生所)摘 要 通过抗生素对新疆3个棉花品种愈伤组织形成和生长影响的试验,结果表明头孢霉素和羧苄青霉素对其都有影响,其中羧苄青霉素的抑制作用较强,头孢霉素作为抑制农杆菌的抗生素较为适宜。
关键词 棉花 抗生素 愈伤组织诱导中图分类号 S562.035.3目前,农杆菌介导法是棉花遗传转化中较为常用的一种方法。
因此,用于抑制农杆菌生长的抗生素选择是十分必要的,抗生素的选用和使用量的确定都会直接影响到棉花愈伤组织的诱导和生长。
故对其进行研究可以更好地指导棉花的遗传转化。
1 材料和方法1.1 材 料以陆地棉栽培品种新陆早6号、新陆早7号和新陆早8号的脱绒种子为材料。
首先培养成无菌苗,然后将生长一致的无菌苗下胚轴切成0.5c m长的切段,作为外植体,接种在诱导培养基上。
1.2 含抗生素诱导培养基的制备基本培养基为M S无机盐,B5维生素,100m g·L21肌醇,0.1m g·L212.42D,0.1 m g·L21KT,30g·L21葡萄糖和7.0g·L21琼脂粉,pH5.8~6.0。
待灭菌后的培养基冷却至50℃左右,加入定量的过滤灭菌的抗生素,混匀分装(头孢霉素由中科院遗传所提供分装产品,羧苄青霉素由上海四药股份有限公司提供)。
1.3 方 法将外植体接入含有不同浓度抗生素的诱导培养基中,培养条件为28~30℃,每天光照16h。
培养6d时,调查切段(外植体)启动情况,30d时将切段愈伤组织进行称重。
每个浓度处理重复3次,取其平均值,以不加抗生素处理为对照。
收稿日期:1998-09-22“九五”国家科技攻关计划资助项目862 新 疆 农 业 大 学 学 报 1998年2 结果分析2.1 头孢霉素对棉花下胚轴切段启动、愈伤组织生长的影响头孢霉素对新陆早7号棉花切段启动有一定的影响,浓度为300m g·L21时,6d时切段启动频率(R6)为100%;浓度为400m g·L21时,R6为100%;浓度为500m g·L21时,R6则为50%,对照为100%。
诱导提取的dsRNA粗提液对黄瓜绿斑驳花叶病毒的抑制作用_郑海刚

福建农林大学学报(自然科学版)第40卷第4期Journal of Fujian Agriculture and Forestry University(Natural Science Edition)2011年7月诱导提取的dsRNA粗提液对黄瓜绿斑驳花叶病毒的抑制作用郑海刚,陈启建(福建农林大学植物病毒研究所,福建福州350002)摘要:在已测序的黄瓜绿斑驳花叶病毒运动蛋白基因序列的基础上,设计特异引物,扩增全长基因,将其插入到原核表达载体L4440的2个T7启动子之间,构建能诱导形成双链RNA(dsRNA)的原核表达载体L4440-MP,并转化大肠杆菌HT115(DE3),经IPTG诱导,提取了高质量的dsRNA.将提取的dsRNA对黄瓜进行抗病性鉴定,ELISA检测结果表明,提取的dsR-NA能有效地抑制黄瓜绿斑驳花叶病毒的侵染,抗病率达到40%左右.关键词:黄瓜绿斑驳花叶病毒;RNA沉默;双链RNA中图分类号:S432.1文献标识码:A文章编号:1671-5470(2011)04-0346-05Inhibition of crude extracts of extracted dsRNA by inductionon the cucumber green mottle mosaic virusZHENG Hai-gang,CHEN Qi-jian(Institute of Plant Virology,Fujian Agriculture and Forestry University,Fuzhou,Fujian350002,China)Abstract:The movement protein(MP)gene of cucumber green mottle mosaic virus was cloned using specific primers designed ac-cording to the sequenced gene.Then the PCR products were inserted into the vector L4440,which contains T7promoter sites flanking each side of the multiple cloning site(MCS)and can thus produce dsRNAs.The resulted plasmids were transformed into the Esche-richia coli HT115(DE3).After IPTG induction,dsRNAs of high quality were obtained.The effects of the dsRNAs on the disease re-sistence of cucumber were then evaluated.ELISA experiments showed that about40%of plants applied with dsRNAs were virus-free.Key words:cucumber green mottle mosaic virus;RNA silencing;dsRNA植物病毒病有植物“癌症”之称,其每年对粮食作物、园艺作物、经济作物等造成的经济损失约数百亿元[1].黄瓜绿斑驳花叶病毒(cucumber green mottle mosaic virus,CGMMV)是严重威胁葫芦科作物生产的重要病毒之一.2006年12月中华人民共和国农业部将CGMMV确定为我国农业植物检疫性有害生物.因此对该病毒的防治显得尤为重要[2-3].目前,防治病毒病常用的方法有:经组织脱毒后获得无病毒的繁殖材料;使用农药驱除或杀死昆虫介体;通过基因工程培养获得转基因植株等[4].不过这些方法都有一定的局限性,如组织脱毒法成本高、工作量大,喷洒农药会造成严重的环境污染,获得转基因植物的几率小,转化体系不成熟等.RNA沉默机制的揭示为植物病毒的防治提供了新的思路[5].RNA干扰(RNA interference,RNAi)是由双链RNA(dsR-NA)介导的、在转录后特异性地降解与之同源的mRNA,从而抑制相应基因表达的过程,是生物体中存在的一种现象,在植物中称转录后基因沉默[6-7].RNAi是生物体对病毒基因等外源核酸侵入的一种保护性反应[8-10].目前,RNA沉默已成为研究植物基因功能的一个有力工具,在动物病毒病的防治和人类病毒病的治疗等方面被广泛应用[11-13].dsRNA是RNA沉默起始的关键分子,然而在大肠杆菌(Escherichia coli)细胞中存在RNaseⅢ,产生的dsRNA会被其他特异的酶切掉,因此在一般的大肠杆菌中较难获得大量的dsRNA.利用大肠杆菌RNaseⅢ缺陷型菌株HT115(DE3)可诱导表达靶基因dsRNA,并可生产高质量的大量的dsRNA.提取的高质量的dsRNA摩擦接种植株可启动RNA沉默的开始,从而抑制病毒的侵染[14-16].本试验研究诱导提取的dsRNA对CGMMV的抑制效果,以期为植物病害的防治提供参考.收稿日期:2010-11-05修回日期:2011-01-10基金项目:国家自然科学基金项目(30770089);福建省自然科学基金项目(2009J1069).作者简介:郑海刚(1983-),男,硕士研究生.研究方向:分子病毒.Email:zhenghaigang1010@163.com.通讯作者陈启建(1967-),副研究员,博士,硕士生导师.研究方向:植物病毒.1材料与方法1.1菌株质粒毒源CGMMV 由福建农林大学植物病毒研究所(以下简称病毒所)保存并繁殖于黄瓜上.E.coli DH5α、E.coli HT115(DE3)及L4440载体、CGMMV MP 的一抗血清也由病毒所保存.1.2酶和有关试剂各种限制性内切酶、Taq DNA 聚合酶、T4DNA 连接酶、M-MLV 反转录酶、DNA 酶、RNaseA 等购自TAKARA 公司;氨苄青霉素(AMP )、四环素(TET )、TrizoL 试剂、DNA 回收试剂盒、质粒小提试剂盒购自天根生物有限公司;以碱性磷酸酶标记的羊抗兔(IgG-AP )二抗、IPTG 购自SIGMA 公司;其他试剂均为国产分析纯.1.3引物根据病毒所已测序过的CGMMV MP 的基因序列,利用Primer Premier 5.0软件设计,并由TAKARA 公司合成,引入酶切位点(Pst Ⅰ、Sac Ⅰ).引物序列如下:F :AACTGCAGATGTCTCTAAGTAAGGTGTCGG Pst ⅠR :CGAGCTCCTAGGTGTGATCGGATTGTAAG Sac Ⅰ划线部分分别为酶切位点Pst Ⅰ、Sac Ⅰ.1.4目的基因CGMMV MP 的获得先用Trizol 法[17]提取病毒的总RNA.cDNA 合成:取2μL 提取的病毒总RNA ,加1μL 下游引物和2μL DEPC 水,放入PCR 仪中(70ħ)10min ,然后迅速置于冰上5min ;冷却后加混合物(0.5μL M-MLV 反转录酶、2μL 5ˑM-MLV buffer 、2.5μL dNTPs 、0.25μL Ribonuclease inhibitor ),于42ħ合成60min ;以此为模板进行PCR 反应.PCR :取0.2mL Eppendorf 管,依次加入1μL cDNA 模板、1μL 10pmol ·L -1上游引物、1μL 10pmol ·L -1下游引物、2μL 2.5mmol ·L -1dNTPs 、2μL 10ˑPCR 反应缓冲液、0.3μL Taq DNA 聚合酶,加灭菌ddH 20至终体积为25μL ;反应程序:95ħ预变性5min ,95ħ变性30s ,55ħ退火30s ,72ħ延伸60s ,循环35次后72ħ延伸10min.反应结束后,电泳检测扩增结果.1.5L4440-MP 原核表达载体的构建割胶回收MP 片段,并分别用Pst Ⅰ-Sac Ⅰ双酶切已经回收的MP 片段和L4440载体.电泳检测后再分别割胶回收,之后连接转化DH5α,用Pst Ⅰ-Sac Ⅰ双酶切鉴定重组质粒,经测序正确后命名为L4440-MP.1.6dsRNA 的诱导表达将构建好的质粒L4440-MP 转化E.coli HT115(DE3),筛选含有重组质粒的转化子.将筛选的转化子接种于LB 培养基(含AMP 、TET )中,在37ħ培养约16h.取适量菌液重新接种于新鲜的含抗生素的LB培养基中,37ħ、250r ·min -1振荡2h ,使D 600nm ≈0.5.加IPTG (终浓度0.8mmol ·L -1)进行诱导表达,37ħ、250r ·min -1振荡培养4h 后用于dsRNA 的提取.1.7dsRNA 的提取取2mL 菌液于5000r ·min -1离心1min ,瞬离,倒掉上清,加500μL TE 缓冲液(10mmol ·L -1Tris 、1mmol ·L -1EDTA ,pH 7.5)重新悬浮;加30μL 10%SDS 振荡混匀,37ħ恒温孵育30min ;加100μL NaCl (5mol ·L -1)混匀,再加入80μL CTAB /NaCl ,混匀,65ħ热激10min ;加等体积酚/氯仿/异戊醇(25ʒ24ʒ1),混匀,12000r ·min -1离心5min ;取上清液,用0.8倍体积的异丙醇沉淀,12000r ·min -1离心10min ;弃上清,用75%乙醇(500μL )洗涤,6000r ·min -1离心4min ;干燥后溶于TE 缓冲液,电泳检测;分别用DNase 、RNaseA 消化后电泳检测.1.8dsRNA 介导的病毒抗性鉴定将提取的来源于CGMMV MP 基因的dsRNA 粗提液以约2μg ·μL -1的浓度溶解于TE 缓冲液中.选取30棵4-5叶苗龄的黄瓜,用dsRNA 溶液和CGMMV 病汁液等体积混合后摩擦接种于黄瓜叶片,进行抗病性鉴定.阳性对照用CGMMV 病汁接种,以pH 7.010mmoL ·L -1磷酸钠缓冲液按1ʒ10的比例(W /V )研磨CGMMV 侵染的叶片作为接种液;阴性对照用TE 缓冲液(10mmol ·L -1Tris 、1mmol ·L -1EDTA ,pH 7.5)接·743·第4期郑海刚等:诱导提取的dsRNA 粗提液对黄瓜绿斑驳花叶病毒的抑制作用种.在接种后的30d 内每天观察并记录症状,定期采样,用间接ELISA 法[18]检测黄瓜中CGMMV 的含量.1:CGMMV MP ;M :Lambda DNA Eco R Ⅰ/Hin d Ⅲmarker.图1CGMMV MP PCR 产物Fig.1PCR amplification of CGMMV MP2结果与分析2.1重组质粒L4440-MP 载体的构建从含CGMMV 的黄瓜病叶中提取总RNA ,用特异引物(F 、R )RT-PCR 获得CGM-MV MP 全长,1%琼脂糖凝胶电泳检测,大小为795bp (图1),表明扩增成功,割胶回收MP 片段.然后用Pst Ⅰ-Sac Ⅰ双酶切已经回收的MP 片段和质粒L4440载体,电泳检测后再次回收(图2).将两者连接转化DH5α,通过筛选阳性克隆及双酶切鉴定重组质粒,可以切下目的片段,表明载体构建成功,命名为L4440-MP (图3).1:载体L4440;M :Lambda DNA Eco R Ⅰ/Hin d Ⅲmarker.图2载体L4440双酶切Fig.2Analysis of L4440digested by restriction enzyme 1:质粒L4440-MP ;M :Lambda DNA Eco R Ⅰ/Hin d Ⅲmarker.图3重组质粒L4440-MP 双酶切Fig.3Analysis of L4440-MP digested by restriction enzyme2.2dsRNA 的诱导表达和提取将含有质粒L4440-MP 的E.coli HT115(DE3)菌株,用0.8mmoL ·L -1IPTG 诱导表达,提取dsRNA.经电泳检测可见,在975bp 位置有清晰的条带,比MP 多180bp (图4).因为诱导成dsRNA 时含有L4440上多克隆位点180bp 的序列,所以大小为975bp ,表明提取成功.将提取的dsRNA 分别用DNase 和RNaseA 消化,大小正确,条带单一,证实得到的核酸是dsRNA (图5).M :150bp DNA marker ;1、2、3:dsRNA.图4未经DNase 和RNaseA 消化的dsRNAFig.4Extraction products of dsRNAM :Lambda DNA Eco R Ⅰ/Hin d Ⅲmarker ;1:用DNase消化的dsRNA ;2:用RNaseA 消化的dsRNA.图5经DNase 和RNaseA 消化的dsRNAFig.5Extraction products of dsRNA treated by DNase and RNaseA2.3dsRNA 介导的病毒抗性鉴定ELISA 检测结果如表1所示.结果表明,构建的来源于CGMMV MP 基因的原核表达载体表达的dsR-·843·福建农林大学学报(自然科学版)第40卷NA 可以有效地防治CGMMV.在接种病毒后的一个月,阳性对照组全部发病,阴性对照组全部健康.在试验组中,大约40%的植株表现健康,病毒的含量与阴性对照组接近;其余的植株表现典型的发病症状,病毒的含量与阳性对照组接近.表1ELISA 检测dsRNA 介导的病毒抗性效果1)Table 1ELISA detection of resistance mediated by dsRNA transcriptsdsRNA 溶液表型病毒的滴定度10dpi 20dpi 30dpi 阳性对照S10/100.524ʃ0.0060.612ʃ0.0040.586ʃ0.007阴性对照R10/100.048ʃ0.0050.042ʃ0.0030.039ʃ0.006L4440-MPR12/300.051ʃ0.0040.049ʃ0.0020.047ʃ0.003S18/300.484ʃ0.0050.503ʃ0.0030.516ʃ0.0041)病毒的滴定度为ELISA 检测的平均值ʃ标准偏差(n =3);R 和S 分别表示抗病(resistant )和感病(susceptible );dpi 表示接种后的天数(days post inoculation ).3讨论在植物抗病毒基因工程领域,利用来源于植物病毒本身的核苷酸序列转化植物来获得抗病毒的工程植株已成为增强作物对病毒病抗性的主要措施之一,而RNA 介导的抗性是近年来发现并引起广泛关注和深入研究的一种策略.研究表明,dsRNA 能高效诱导RNA 沉默,从而导致植物体内同源的内源基因表达受到抑制,瞬时表达或摩擦接种的dsRNA 能启动RNA 沉默的起始,从而抑制植物病毒的侵染.Timmons et al [19]利用HT115生产的dsRNA 诱导小杆线虫内源基因,从而发生了RNA 沉默.HT115菌株是RNase Ⅲ缺失体,细菌中绝大多数的dsRNA 能被RNase Ⅲ降解,因此,HT115菌株可以生产大量的dsRNA.于是,Tenllado et al [20]创立了利用HT115菌株大量表达病毒来源dsRNA 的简单、有效的方法.Ten-llado et al [21]还发现通过喷雾器将病毒的dsRNA 喷到植物叶片几天后,再在植物的同一叶片接种该病毒,该植物会对病毒产生免疫作用.本试验以L4440载体为基础,将CGMMV MP 全长基因插入其中,构建了能表达dsRNA 的原核表达载体,在E.coli HT115(DE3)中实现了dsRNA 的大量表达,并提取了大量的dsRNA ,且ELISA 检测结果表明,提取的dsRNA 能有效地抑制CGMMV 的侵染.提取高质量的dsRNA 是试验的关键,其中,对含转化子的E.coli HT115(DE3)的诱导效果又是提取高质量dsRNA 的关键.这里需注意2点:(1)接种的菌必须新鲜;(2)菌的浓度要合适,D 600nm ≈0.5最佳,诱导时间3-4h.本研究发现,在混合接种CGMMV 和HT115菌株生产的CGMMV MP 的dsRNA 粗提物的黄瓜中,表现为抗病的植株,在接种后整个生活周期内均不发病;表现为感病的植株,出现了CGMMV 典型症状;只接种CGMMV 的黄瓜,均表现为发病症状,说明HT115菌株生产CGMMV MP 的dsRNA 对CGMMV 病毒有明显的抑制作用.因此,推测dsRNA 分子进入植物细胞并形成小干扰RNA (small interfering RNA ,siRNA )的机制:dsRNA 分子能借助于摩擦产生的表皮细胞伤口与病毒粒子一起进入植物细胞中,然后这些dsRNA 分子被植物中的Dicer 切割成能指导同源病毒RNA 切割的siRNA 分子,随后siRNA 分子整合入RISC 复合物,并介导RISC 对其同源病毒的RNA 分子进行降解.本研究结果证实了RNA 沉默的原理,根据病毒的基因构建能表达dsRNA 原核表达载体,最终诱导植物对病原物产生抗性,形成了一种防治植物病害的新途径.参考文献[1]FRASER R S S.Plant resistance to viruses ∥GRANOFF A ,WEBSTER R G.Encyclopedia of Virology [M ].San Diego ,Cali-fornia :Academic Press ,1999:1300-1307.[2]秦碧霞,蔡健和,刘志明,等.侵染观赏南瓜的黄瓜绿斑驳花叶病毒的初步鉴定[J ].植物检疫,2005,19(4):198-200.[3]CHANG K S ,KI S H ,JUNG H L ,et al.Isolation and characterization of watermelon isolate of cucumber green mottle mosaicvirus (CGMMV-HY1)from watermelon plants with severe mottle mosaic symptoms [J ].Plant Pathology Journal ,2005,2(2):167-171.[4]程海鹏,朱睦元,金伟,等.植物转基因沉默研究进展[J ].生物工程进展,2001,21(6):42-47.[5]TENLLADO F ,CESAI L ,JOSERANMON D R.RNA interference as new biotechnological tool for the control of virus disease·943·第4期郑海刚等:诱导提取的dsRNA 粗提液对黄瓜绿斑驳花叶病毒的抑制作用in plants [J ].Virus Res ,2004,102:85-96.[6]SMITH N A ,SINGH S P ,WANG M B ,et al.Total silencing by intron-spliced hairpin RNAs [J ].Nature ,2000,407:319-320.[7]WATERHOUSE P M ,WANG M B ,LOUGH T.Gene silencing as an adaptive defence against viruses [J ].Nature ,2001,411:834-842.[8]ROVERE C V ,MARIANA D V ,ESTENBAN H.RNA-mediated virus resistance [J ].Current Opinion in Biotechnology ,2002,13:167-172.[9]KARPALA A J ,DORAN T J ,BEAN A G D.Immune responses to dsRNA :implications for gene silencing technologies [J ].Immun Cell Bio ,2005,83:211-216.[10]DYKXHOORN D M ,LIEBERMAN J.Silencing viral infection [J ].PLoS Med ,2006,3:1000-1004.[11]THOMAS C L ,LEH V ,LEDERER C ,et al.Turnip crincle virus coat protein mediates suppression of RNA silencing in Nic-otiana benthamiana [J ].Virology ,2003,306:33-41.[12]HAMMOND S M ,CANDY A A ,HANNON G J.Post-transcriptional gene silencing by double-stranded RNA [J ].Nat RevGenet ,2001,2:110-119.[13]LI F ,DING S W.Virus counterdefense :diverse strategies for evading the RNA-silencing immunity [J ].Annu Rev Microbi-ol ,2006,60:503-531.[14]ANASTASIA M ,KRITON K ,ALEXANDRA B ,et al.Generation of transgenic potato plants highly resistant to potato virus Y(PVY )through RNA silencing [J ].Mol Breeding ,2004,14:185-197.[15]张德咏,张战泓,刘勇,等.表达dsRNA 的细菌提取液可抑制黄瓜花叶病毒对烟草的侵染[J ].植物病理学报,2008,38(3):304-311.[16]GAN D F ,ZHANG J ,JIANG H B ,et al.Bacterially expressed dsRNA protects maize against SCMV infection [J ].Plant CellRep ,2010,29:1261-1268.[17]张梅娟,沙伟,都艳霞.几种提取苔藓植物总RNA 方法的比较[J ].齐齐哈尔大学学报:自然科学版,2009,25(1):51-54.[18]李泽,王洪超,吴玉梅,等.利用ELISA 和RT-PCR 法检测甜菜丛根病毒[J ].中国甜菜糖业,2008(2):52-55.[19]TIMMONS L ,COURT D L ,FIRE A.Ingestion of bacterially expressed dsRNA can produce specific and potent genetic inter-ference in Caenorhabditis elegans [J ].Gene ,2001,263:103-112.[20]TENLLADO F ,LIAVE C ,DIAZ-RUIZ J R.RNA interference as a new biotechnological tool for the control of virus diseasesin plants [J ].Virus Res ,2004,102:85-96.[21]TENLLADO F ,MARTINEZ-GARCIA B ,VARGAS M ,et al.Crude extracts of bacterially expressed dsRNA can be used toprotect plants against virus infections [J ].BMC Biotechnol ,2003,3:3.(责任编辑:杨郁霞)·053·福建农林大学学报(自然科学版)第40卷。
五倍子粗提液对棉花黄萎病的防治效果

五倍子粗提液对棉花黄萎病的防治效果袁媛;冯鸿杰;师勇强;赵丽红;魏锋;冯自力;朱荷琴【期刊名称】《中国棉花》【年(卷),期】2018(045)004【摘要】以五倍子为对象,研究其粗提液对大丽轮枝菌的抑菌效果及对棉花黄萎病的田间防治效果,为研制用于棉花黄萎病防治的新型植物源制剂提供技术支持.结果表明,当五倍子粗提液体积分数达到0.6%时,其对分离自不同棉区的大丽轮枝菌强致病力菌株Vd080、Vd147、Vd021的菌落生长抑制率和产孢量抑制率均达到100%.田间叶面喷施五倍子粗提液对棉花黄萎病的防治效果最高可达到58.8%,40、80倍液处理能显著提高棉花纤维的断裂伸长率,且前者能显著提高籽棉产量.推荐生产上在棉花黄萎病发生前或发生初期叶面喷施40~80倍的五倍子粗提液进行防治.【总页数】5页(P8-11,40)【作者】袁媛;冯鸿杰;师勇强;赵丽红;魏锋;冯自力;朱荷琴【作者单位】棉花生物学国家重点实验室/中国农业科学院棉花研究所,河南安阳455000;棉花生物学国家重点实验室/中国农业科学院棉花研究所,河南安阳455000;棉花生物学国家重点实验室/中国农业科学院棉花研究所,河南安阳455000;棉花生物学国家重点实验室/中国农业科学院棉花研究所,河南安阳455000;棉花生物学国家重点实验室/中国农业科学院棉花研究所,河南安阳455000;棉花生物学国家重点实验室/中国农业科学院棉花研究所,河南安阳455000;棉花生物学国家重点实验室/中国农业科学院棉花研究所,河南安阳455000【正文语种】中文【中图分类】S435.621【相关文献】1.黄萎病菌毒素粗提液对棉花抗性酶的诱导 [J], 马春红;范尉尉;董文琦;李运朝;崔四平;贾银锁2."入田"拌种对棉花枯黄萎病的防治效果 [J], 张德政; 朱荷琴; 冯自力; 师勇强; 冯鸿杰; 赵丽红; 魏锋; 彭军3.2种微生物菌剂对棉花农艺性状和黄萎病防治效果的影响 [J], 王凡;邵青龙;张栋海;姜明军4.微生物菌肥对棉花黄萎病的防治效果研究 [J], 吕博;孟庆忠;张成;韩光明;陈全求;房健;易先达5.杠柳根皮乙醇粗提液对菜青虫的拒食作用及其防治效果 [J], 朱九生;乔雄梧;王静;秦曙因版权原因,仅展示原文概要,查看原文内容请购买。
香蕉枯萎病菌粗毒素的毒性及其模型

香蕉枯萎病菌粗毒素的毒性及其模型许文耀;兀旭辉;林成辉【期刊名称】《热带作物学报》【年(卷),期】2004(025)004【摘要】以香蕉组织培养苗为受试植物,蕉苗受毒素作用后的病情指数为指标,测定香蕉枯萎病菌(Fusarium oxysporum f.sp.cubense)粗毒素对蕉苗的毒性.结果表明,毒素处理72,96,120和144 h引致香蕉苗受害程度(以病情指数表示)达90的剂量(TD90)的质量浓度分别为2 057.20,1 245.49,549.54,380.19 μg/mL,蕉苗的受害程度存在着随处理时间延长和处理剂量的增加而加重的趋势.根据毒素对蕉苗的毒性测定结果,建立了蕉苗受毒素作用的"时间-剂量-受害程度"模型.根据模型的分析结果认为,粗毒素引致蕉苗受害的有效时间为96~120 h,有效剂量质量浓度为260.0~130.0 μg/mL.【总页数】5页(P25-29)【作者】许文耀;兀旭辉;林成辉【作者单位】福建农林大学植物保护学院,福州,350003;福建农林大学植物保护学院,福州,350003;福建农林大学植物保护学院,福州,350003【正文语种】中文【中图分类】S436.68【相关文献】1.香蕉枯萎病菌及其粗毒素对香蕉的致病性比较 [J], 李梅婷;严琰;张绍升2.稻瘟病菌粗毒素对水稻品种的毒性与产毒菌株致病力的关系 [J], 许文耀;王金陵3.福建水华微囊藻粗毒素对原代培养大鼠肝细胞毒性作用的初步研究 [J], 庞春艳;林炜;陈华;林锦峰4.离体培养中利用病菌粗毒素检测香蕉枯萎病抗性研究 [J], 罗燕羽;刘绍钦;黄绍力;刘伟光;张木清;黄炽辉5.百菌清与多菌灵对香蕉枯萎病菌抑制条件的研究 [J], 罗芳;谯顺彬因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
西 北 植 物 学 报 2 0 ,2 6 : 7 — 1 8 02 2 () 1 7 3 3 3
Ae a Bot Bo e . 0c i nt t . r a1 一 c de .Si n.
文 章 编 号 :0 04 2 ( 0 2 0 — 3 70 1 0 —0 5 2 0 ) 61 7 — 7
枯 萎 菌 毒 素 粗 提 液 对 4种 陆 地 棉 外 植 体 愈 伤 组 织 诱 导 率 的 影 响
肖 娅萍, 浩, 齐 王莆之, 刘全宏
( 西师范 大学生 命科 学学 院 , 安 706 ) 陕 西 1 0 2
摘
要 : 培 养 5d 的 4种 陆 地 棉 栽 培 品 种 军 棉 1号 、 棉 2号 、 2 4及 中棉 1 将 泗 23 2号 无 菌 苗 , 取
c ptbiiy e i lt of dif r t xpl nt t t c f e en e a s o he onc n r to of us i e T oxi ha di tn- e t a in F arc m n s s i—
guihed. s
Ke r s u l n o t n; x l n  ̄ al s Fu a ie o y p r m . p. a if cu y wo d : p a d c to e pa t c lu ; s rcm x s o a f s v sn e t m
西
北
植
物
学
报
棉 花 枯 萎 病 害 是 我 国 棉 区 的 重 要 病 害之 一 , 使 皮 棉 产 量 降 低 3 ~ 7 , 重 影 可 0 0 严
响 了 棉 花 的产 量 。 近 年 来 , 国 在 陆 地 棉 抗 枯 萎 病 领 域 的工 作 获 得 较 大 突 破 , 我 已成 功 培 育 出 一 些 高 抗 品 种 。 过 建 立 陆 地 棉组 织 细 胞 培 养 系 统 , 陆 地 棉 的细 胞 工 程 和 基 因 工 程 的 通 为 遗 传 操 作 及 抗 性 细 胞 系 的筛 选 奠 定 了基 础 [ 。本 研 究 利 用 4种 陆 地 棉 栽 培 品 种 的 外 植 】 叫] 体 , 不 同 枯 萎 菌毒 素 梯 度 的 培 养 基 中进 行 培 养 观 察 , 在 旨在 为 陆地 棉 愈 伤 组 织 的组 织 形 态 学研究 、 枯萎 品种的形态学鉴定及筛选提供依据 。 抗
其 根 、 胚 轴 及 子 叶 接 种 在 含 不 同 浓 度 枯 萎 菌 毒 素 粗 提 液 的 培 养 基 上 , 隔 3d观 察 统 计 其 下 每 成 活 率 , 计 至 3 。 结 果 表 明 : 地 棉 外 植 体 愈 伤 组 织 的 诱 导 率 与 枯 萎 菌 毒 素 浓 度 呈 负 相 统 3d 陆 关 , 不 同的外 植体 对 枯萎 菌毒 素浓 度 的敏感 程度 有所 差异 。 而 关 键 词 : 地 棉 ; 植 体 ; 伤 组 织 ; 萎 菌 陆 外 愈 枯 中 图 分 类 号 : 4 . Q9 8 1 5 文献标 识 码 : A
223 a 4 nd Zho ngm i n ha b n c t e or a 12, d ee ulur d f 5 day n he c dii kil s i t on ton l ba t rum . cei
The hei o n t r r ot,owe l u a xi nd c y e l r p m l r a s a ot l don we e noc l t d on t ulur e — r i u a e he c t e m di
I l nc f Fus i e To i n c l u nduc i i y nf ue e o ar c m x n o a l si tv t o x a n f ur upl nd c t ns f e pl nt i o a ot o
XI AO a p n Q IH a W AN G e z i LI Q u n h n Y - i g, o, Zh — h , U a — o g
ur s ont ni g dif r nt c n c ai n fe e onc nt ato us ie T oxi e r i n of F ar c m n.T he ur va r t w e e s vi l a e r ob — s ve a ounr d e er hid ay .T he e u t s ow e t t t c lu i er d nd c e v y t r d s r s l s h d ha he a l s ndu tviy of c i t
e p a t r e a i e y i t r e a e t h o c n r to f Fu a i e To i . x l n s a e n g tv l n e r l t d wih t e c n e t a i n o s r c m x n The S S U —
收 稿 日期 :0 2 0 — o 修 改 稿 收 到 日期 :0 20 —6 20—32 ; 2 0 —4 0 作者 简介 ; 肖娅 萍 ( 9 6 ) 女 ( 族 ) 副 教 授 , 事 植 物 学 及 植 物 生 物 技 术 教 学 科 研 工 作 。 15一 , 汉 , 从
维普资讯
( le e ofLi c e e, a n r a n v r iy, 'n 71 0 2, Co l g f S i nc Sh a xiNo m l U i e s t Xi 0 6 Chi a) e a n
Ab t a t Th o ru ln o t n , ih we ec lia so u m in 1 Si i n 2, in a s r c : e f u p a d c to s wh c r u tv r f n a , m a M a hu J