分子生物学第5章蛋白质翻译
分子生物学名词解释最全

第一章名词解释1.基因(gene)是贮存遗传信息的核酸(DNA或RNA)片段,包括编码RNA和蛋白质的结构基因以及转录调控序列两部分。
2. 结构基因(structural gene)指基因中编码RNA和蛋白质的核苷酸序列。
它们在原核生物中连续排列,在真核生物中则间断排列。
3. 断裂基因(split gene 真核生物的结构基因中,编码区与非编码区间隔排列。
4. 外显子(exon)指在真核生物的断裂基因及其成熟RNA中都存在的核酸序列。
5. 内含子(intron)指在真核生物的断裂基因及其初级转录产物中出现,但在成熟RNA 中被剪接除去的核酸序列。
6. 多顺反子RNA(polycistronic/multicistronic RNA)一个RNA分子上包含几个结构基因的转录产物。
原核生物的绝大多数基因和真核生物的个别基因可转录生成多顺反子RNA。
7. 单顺反子RNA(monocistronic RNA)一个RNA分子上只包含一个结构基因的转录产物。
真核生物的绝大多数基因和原核生物的个别基因可转录生成单顺反子RNA。
8. 核不均一RNA(heterogeneous nuclear RNA, hnRNA)是真核生物细胞核内的转录初始产物,含有外显子和内含子转录的序列,分子量大小不均一,经一系列转录后加工变为成熟mRNA。
9. 开放阅读框(open reading frame, ORF)mRNA分子上从起始密码子到终止密码子之间的核苷酸(碱基)序列,编码一个特定的多肽链。
10. 密码子(codon)mRNA分子的开放读框内从5' 到3' 方向每3个相邻的核苷酸(碱基)为一组,编码多肽链中的20种氨基酸残基,或者代表翻译起始以及翻译终止信息。
11. 反密码子(anticodon)指tRNA分子反密码环中间3个相邻的核苷酸(碱基),它们与mRNA上的三联体密码子互补配对,确保蛋白质合成时氨基酸按照密码子对号入座。
分子生物学-第5章-分子生物研究法(上)精选全文完整版

限制性核酸内切酶
限制性核酸内切酶(restriction endonuclease, RE)
一类能识别和切割双链DNA分子中特定碱基顺序的核酸 水解酶
Bam HⅠ
GGATCC CCTAGG
GCCTAG+
GATCC G
分类: Ⅰ、Ⅱ、Ⅲ (基因工程技术中常用Ⅱ型)
命名
Hin dⅢ
Haemophilus influenzae 自主 复制能力的 DNA分子( vector),如 病毒、噬菌体 和质粒等小分 子量复制子都 可以作为基因 导入的载体。
1970年Mandel和Higa发现,大肠杆菌细胞经适量氯化钙处 理后,能有效地吸收λ噬菌体DNA。
1972年,Cohen等人又报道,经氯化钙处理的大肠杆菌细 胞同样能够摄取质粒DNA。
把磷酸基团加到多聚核苷酸链的5'-OH末端(进行末端标记 实验或用来进行DNA的连接 在双链核酸的3'末端加上多聚单核苷酸
从DNA链的3'末端逐个切除单核苷酸
从DNA链的5'末端逐个切除单核苷酸 切除位于DNA链5'或3'末端的磷酸基团
1972 - Paul Berg,
Produced first recombinant DNA using
5.1 重组DNA技术回顾 5.2 DNA基本操作技术 5.3 RNA基本操作技术 5.4 SNP的理论与应用 5.5 基因克隆技术 5.6 蛋白质组与蛋白质组学技术
5.1 重组DNA技术回顾
三大成就 :
1. 40年代确定了遗传信息的携带者,即基因的分子载体 是DNA而不是蛋白质,解决了遗传的物质基础问题;
• 基因工程是指在体外将核酸分子插入病毒、质粒 或其它载体分子,构成遗传物质的新组合,使之 进入原先没有这类分子的寄主细胞内并进行持续 稳定的繁殖和表达。
分子生物学---蛋白组学整理英文

proteomicsProteome: 细胞或组织或机体在特定时间和空间上表达的所有蛋白质。
Proteomics: 分析细胞内动态变化的蛋白质组成成分,表达水平于修饰状态,了解蛋白质之间的相互作用于联系,在整体水平上研究蛋白的组成与调控的活动规律。
研究蛋白组学希望达到的目标:By studying global patterns of protein content and activity and how these change during development or in response to disease, proteomics research is poised to boost our understanding of systems-level cellular behaviors. Clinical research also hopes to benefit from proteomics by both the identification of new drug targets and the development of new diagnostic markers.蛋白质组学研究内容:蛋白鉴定,蛋白定量,蛋白相互作用,蛋白修饰。
Why proteomics(为什么研究蛋白组学)•Proteins distinguish various types of cells, since all cells have essentially the same “Genome” their differences are dictated by which genes are active and the corresponding proteins that are made.•Similarly, diseased cells may produce dissimilar proteins to healthy cells.•Post-translational modifications can dramatically alter protein function - the task of studying proteins is often more difficult than genes.What’s MS(mass spectrometry),即质谱的工作原理1.The basic principle of MS is to generate ions from either inorganic or organiccompounds by suitable method, to separate these ions by their mass-to-charge ratio (m/z) and to detect them qualitatively and quantitatively by their respective m/z and abundance.即质谱能够实现不同质量离子的分离和相对定量,m/z(谱图中的x轴)值可以区分出不同的离子,intensity(谱图中的y轴)表示离子的相对丰度。
分子生物学--蛋白质的翻译课件

Initiation requires factors and free subunits
(2)细菌中有三种起始因子 IF-3:稳定30S亚基;辅助 30S亚基与mRNA上起始点特 异性结合; IF-1:与30S亚基结合在A位, 阻止氨酰-tRNA进入;阻止 30S与50S亚基结合。 IF-2:结合特定起始因子 tRNA,控制它进入核糖体; 有核糖体依赖GTP酶活性;
5.1.2 氨酰-tRNA合成酶引入的两种错误
◆蛋白质合成真实性主要决定于:
tRNA能否把正确的氨基酸放到新生多肽链的 正确位置。 ◆氨酰-tRNA合成酶会引入两种错误:
一种是将错误的氨基酸加在正确的tRNA上; 另一种是将正确的氨基酸加在错误的tRNA上。 前者现错的可能性更大。
Error rates differ at each stage of gene expression
Eplroontegiantibo5yn.2tRr.ai3bno蛋ssfoe白rmfer质omm合ovpee成spta的idloyn三l-gtR个mNRA阶NtAo段,aem简xitne介onadcinygl-tRNA
Termination Polypeptide chain is released from tRNA, and ribosome dissociates from mRNA
I: 次黄嘌呤
1.4.4 读码的连续性
生物合成过程中,mRNA的编码方向是 5`→3`,从N端向C端延伸肽链。一条肽链 合成起始后,密码子按3个一框读下去不重 叠也不跳格,直到终止。
2 蛋白质合成中使用的RNA
概述 2.1 mRNA 2.2 tRNA 2.3 rRNA
分子生物学第五章分子生物学研究法(上)

分子生物学第五章分子生物学研究法(上)——DNA、RNA及蛋白质操作技术第三节RNA操作技术第四节SNP的理论与应用第五节基因克隆技术第六节蛋白质组与蛋白质组学技术夏玉琼2013-10-10目录RNA操作技术cDNA文库的构建基因文库的筛选SNP的理论与应用基因克隆技术蛋白质与蛋白质组学技术分子生物学 夏玉琼 西安电子科技大学cDNA文库的构建切割位点用四碱基特异性的限制性内切酶部分消化DNA 片段,有的仍有切割位点质粒DNA将DNA 克隆进质粒DNA细菌克隆每个细菌都带有不同片段的DNA细菌转化分子生物学 夏玉琼 西安电子科技大学cDNA文库的构建cDNA的长度0.5-8 kb载体:质粒载体和噬菌体类载体完整的cDNA文库包含大于5*105的独立克隆分子生物学 夏玉琼 西安电子科技大学目录RNA操作技术cDNA文库的构建基因文库的筛选SNP的理论与应用基因克隆技术蛋白质与蛋白质组学技术分子生物学 夏玉琼 西安电子科技大学基因文库的筛选含义通过某种特殊方法从基因文库中鉴定出含有所需重组DNA分子的特定克隆的过程筛选方法核酸杂交法PCR筛选法免疫筛选法分子生物学 夏玉琼 西安电子科技大学核酸杂交法培养基上的菌落盖上硝酸纤维素膜移去硝酸纤维素膜裂解、中和去除细菌蛋白DNA 印迹32P 标记探针杂交放射自显影图像挑出阳性克隆保存母板分子生物学 夏玉琼 西安电子科技大学PCR筛选法需获得基因特异性引物将整个基因文库保存在多孔培养板上用设计好的基因探针对每个孔PCR筛选,挑出阳性的孔对阳性的孔再稀释到次级多孔板中PCR筛选重复稀释重复筛选直到与目的基因对应的单个克隆分子生物学 夏玉琼 西安电子科技大学免疫筛选法文库铺于E.coli 形成噬菌斑转移到硝酸纤维素膜吸收λ噬菌体中表达的外源蛋白保存原板,加入一抗筛选膜上的噬菌斑印迹洗去未结合的抗体加入酶偶联的二抗加底物显色从保存板上挑出阳性噬菌斑一抗:第一抗体,识别目标蛋白二抗:抗体的抗体,能增强信号,增加该方法的灵活性分子生物学 夏玉琼 西安电子科技大学目录RNA操作技术SNP的理论与应用SNP概述SNP的检测技术SNP的应用基因克隆技术蛋白质与蛋白质组学技术分子生物学 夏玉琼 西安电子科技大学SNP概述single nucleotide polymorphism,pronounced “snips”单核苷酸多态性基因组DNA序列中由于单个核苷酸的突变而引起的多态性,发生频率1%或更高例如:某些人的染色体上的某个位置为A,而另外一些人的同样位置是T,染色体DNA同一位置上的每个碱基类型叫做一个等位位点继RFLP和SSR之后的第三代遗传标记遗传标记:在遗传分析上用作标记的基因分子生物学 夏玉琼 西安电子科技大学第一代遗传标记:RFLPRFLP标记是发展最早的DNA标记技术。
分子生物学习题集答案 第五章

第五章基因表达自测题(一)选择题A型题1. 基因表达包括①复制②转录③逆转录④翻译A. ①+②B. ①+③C. ②+④D. ③+④E. ①+②+④2. 真核生物的转录不.需要A. 依赖DNA的RNA聚合酶B.三磷酸核苷C.DNAD.RNAE.蛋白质3. 关于转录和复制的区别,说法正确的是A. DNA双链均复制和转录B.复制的原料是一磷酸脱氧核苷,转录的原料是一磷酸核苷C.复制需要引物,转录不需要引物D.复制酶是依赖DNA的DNA聚合酶,转录酶是依赖RNA的DNA聚合酶E.均遵守碱基配对规律,模板中A对应的产物是T4. 关于转录和复制的相同点,说法错误..的是A. 核苷酸以3', 5'-磷酸二酯键连接B.以DNA为模板C.原料为dNTP或NTP,组成核酸链的是dNMP或NMPD.产物可与变性的模板杂交E.新生核酸链的延伸方向是3' → 5'5. 已知某基因转录产物的部分序列是5'-AUCCUGGAU-3',那么该基因中反意义链的相应序列为A.5'-ATCCAGGAT-3'B.5'-TAGGTCCTA -3'C.5'-TAGGACCTA-3'D.5'-UAGGACCUA-3'E.5'-ATCCUGGAT-3'6. E.coli的RNA聚合酶中,辨认转录起始点的组分是A. 核心酶B. σC. αD. βE. β'7. 真核生物中,RNA聚合酶II的转录产物是A. 45S rRNAB. 5S rRNAC. tRNAD. U6 snRNAE. hnRNA8. 关于真核生物的RNA聚合酶,说法正确的是A.RNA聚合酶II对鹅膏蕈碱极敏感B.RNA聚合酶I的转录产物往往是小分子RNAC.RNA聚合酶的氨基端重复序列可发生磷酸化D.不一定需要蛋白质因子辅助E.RNA聚合酶III的转录产物是45S rRNA9. 关于E.coli RNA聚合酶的σ因子的说法错误..的是A.不参与转录延长过程B.辨认–35区和–10区起始转录C.σ因子若不脱落,转录不能进入延长阶段D.σ因子的量少于核心酶,可反复利用E.转录开始后,补加σ因子可提高RNA的生成量10. 在某个体外转录体系中,若模板链含有连续的A,加入何种物质可对产物进行放射性标记A. α-32P-UMPB.γ-32P-dTTPC. β-32P-UDPD. α-32P-UTPE. γ-32P-UTP11. 下列哪项不.是ρ因子依赖的转录终止的特点A.转录终止信号存在于RNA而非DNA模板B.ρ因子有ATP酶活性和解螺旋酶活性C.ρ因子结合RNA的能力弱于对DNA的结合力D.生成的RNA链的3' 端往往富含CE.ρ因子通过与转录产物结合而发挥转录终止作用12. 真核生物II类基因的启动子核心序列通常位于A. –25区B. –10区C. –35区D. +1区E. +10区13. 真核生物中,与原核生物的Pribnow box功能相当的是A. GC boxB. CAAT boxC. CACA boxD. 八聚体(8 bp)元件E. Hogness box14. 下列物质中,能够辅助真核生物的RNA聚合酶结合启动子的是A. 起始因子B. 增强子C. 转录因子D. 延长因子E. σ因子15. 真核生物的RNA聚合酶II在进入转录延长阶段之前,其最大亚基羧基末端结构域发生磷酸化是必需的,催化这一步骤的转录因子是A. TF II BB. TF II DC. TF II HD. TF II EE. TF II F16. 关于转录因子的说法,错误..的是A. 原核细胞中转录起始复合物的形成不需要转录因子B.转录因子的化学本质是蛋白质C.转录因子通过DNA-蛋白质、蛋白质-蛋白质相互作用控制转录起始D.转录因子的转录激活域包括锌指、螺旋-转角-螺旋等结构域E.少量转录因子的不同排列组合即可精确调控众多基因的转录17. 关于真核生物的转录延长,说法正确的是A. 组蛋白发生磷酸化利于核小体解聚B.组蛋白发生乙酰化利于核小体解聚C.转录延长的同时出现多聚核蛋白体现象D.延长因子辅助RNA聚合酶催化核苷酸加入E.转录前后,核小体解聚并在原位重新聚合18. 下列哪种物质不.需要进行转录后加工即可发挥功能A. 大肠杆菌 mRNAB. 大肠杆菌tRNAC.大肠杆菌rRNAD.酵母mRNAE.酵母tRNA19. 关于加帽修饰,说法正确的是A. snRNA不能被加帽B.由加帽酶催化5' 端加入7甲基鸟苷酸C.加帽酶可以与RNA聚合酶I、II、III相结合D.hnRNA转录终止后才开始加帽E.RNA聚合酶发生磷酸化后,使加帽酶与之脱离20. 关于加尾修饰,说法错误..的是A. 组蛋白的成熟mRNA无需加poly A尾B.加尾信号包括AAUAAA和富含GU的序列C.剪切过程需要多种蛋白质因子的辅助D.加尾不需模板E.多聚A形成的过程是缓慢、匀速的反应21. 关于剪接的说法,正确的是A. 初级转录物中不存在内含子B.内含子一定是非编码序列C.内含子两端的结构通常是5'-GU-----AG-3'D. snRNP催化套索RNA发生转酯反应,使内含子和剪接接口一起被去除E.剪接后的mRNA不能与模板链DNA杂交22. RNA编辑发生在A. 成熟的mRNAB. tRNA和rRNA的前体C. hnRNAD.成熟的tRNA和rRNAE. snRNA23. 下列哪种情况下,DNA双链贮存的遗传信息没有..改变A. RNA编辑B.转换C.错义突变D.倒位E.移码突变24. 关于真核rRNA的转录以及加工,说法正确的是A. rDNA位于核仁,呈串联重复排列B.转录出的5S rRNA组成多顺反子RNA的一部分C.rDNA之间被可转录的基因间隔所分隔D.基因间隔等同于内含子E.45S rRNA前身可剪接成为16S、5.8S和28S rRNA25. 生命活动涉及核酸和蛋白质两类生物大分子之间的相互作用,下列物质中不.属于核酸-蛋白质复合物的是A. telomereB.splicesomeC.ribozymeD.SRPE.mRNP26. 蛋白质的生物合成不直接...需要A. RNAB. RNA剪切因子C.分子伴侣D.帽子结合蛋白E.GTP27. mRNA在蛋白质合成中的功能是A. 运输遗传密码所对应的氨基酸B.与蛋白质结合,提供合成场所C.与帽子结合蛋白结合启动翻译D.由三联体密码指引氨基酸的排列顺序E.通过剪切因子切除poly A尾调控翻译效率28. 某种情况下,基因的点突变可能不会影响它所编码的蛋白质的一级结构,遗传密码的哪种特点对此发挥了重要作用A. 方向性B. 连续性C. 摆动性D. 通用性E. 简并性29. 关于遗传密码说法错误..的是A. 遗传密码的阅读方向是5'→3'B. 细胞核和线粒体的遗传密码完全通用C. 61组有意义密码分别编码20种氨基酸D. 密码子的第三位的作用没有前两位重要E. 不同生物对密码子有不同的偏爱30. 密码子与反密码子之间存在摆动配对现象,如果密码子为CGU,则识别它的反密码子可能是A. ICGB. GCAC. CCGD. UCGE. GCI31. 原核生物的核糖体大亚基是A. 30SB. 40SC. 50SD. 60SE. 70S32. 真核生物的核糖体不.含有A. E位B. P位C. A位D. 5S rRNAE. 转肽酶33. 氨基酰-tRNA合成酶的特点是A. 能水解磷酸二酯键,发挥校正功能B. 对氨基酸有绝对专一性C. 对tRNA有相对专一性D. 对氨基酸和tRNA都有高度特异性E. 能水解肽键,发挥校正功能34. 活化后的氨基酸才能与tRNA结合,每个氨基酸的活化过程需要消耗几个高能磷酸键A. 1B. 2C. 3D. 4E. 535. 关于起始tRNA说法错误..的是A. 在肽链延长阶段,起始tRNA不能继续运输蛋氨酸B. 原核与真核起始tRNA所携带的氨基酸不同C. 真核生物中识别AUG的tRNA至少有两种D. 原核生物的起始tRNA可以辨认AUG、GUG和UUGE. 原核生物的起始tRNA可以运输N-甲酰甲硫氨酸、缬氨酸和亮氨酸36. 原核生物的70S 翻译起始复合物不.包括A. 50S亚基B. IF-2C. fMet-tRNAD. mRNAE. 30S 亚基37. 真核生物参与蛋白质合成的起始因子有几种A. 1B. 2C. 3D. 4E. > 538. 原核生物的翻译起始阶段,帮助fMet-tRNA结合AUG的是A. IF-2B. IF-1C. eIF-2D. eIF-3E. eIF-439. SD序列与下列哪种rRNA相互作用A. 5SB. 23SC. 16SD. 5.8SE. 18S40. 原核生物的翻译过程中,能进入P位的是A.fMet-tRNAB.Met-tRNAC.Val-tRNAD. Leu-tRNAE. Ile-tRNA41. 关于真核生物的翻译起始,不.正确的说法是A. 翻译起始的过程较原核生物复杂B. eIF-2是调节真核翻译起始的关键因子C. 起始氨基酰-tRNA进入核糖体的P位D. mRNA的SD序列与其在核糖体上就位有关Met组成翻译起始复合物E. 由80S核糖体、mRNA和Met-tRNAi42. 真核生物肽链合成的延长阶段,促使氨基酰-tRNA进入A位的蛋白质因子是A. EF-1B. EF-2C. EFTD. EFGE. 转肽酶43. 关于转肽酶的说法正确的是A. 是rRNA的一部分B. 由数种核糖体蛋白组成C. 成肽反应不需要延长因子D. 催化氨基酸的α-羧基与核苷酸的3'-OH形成酯键E. 催化新生的肽酰-tRNA转移到P位44. 狭义的真核生物核蛋白体循环所涉及的蛋白质因子是① IF ②转肽酶③ EF ④ RFA. ①+②+③+④B. ①+③+④C. ②+③+④D. ①+③E. ②+③45. 真核生物中参与翻译终止的释放因子的数目是A. 1B. 2C. 3D.4E. > 546. 蛋白质合成时消耗能量的步骤发生在①氨基酸活化②进位③成肽④转位⑤新生肽链的释放A. ①+③+⑤B. ①+②+③C. ①+④+⑤D. ①+②+④E. ①+②+④+⑤47. 蛋白质合成时,每增加1个肽键至少需要消耗几个高能磷酸键A. 1B. 2C. 3D. 4E. 548. 具有酯酶活性并能释放新生肽链的蛋白质因子是A. RF-3B. RRC. 转肽酶D. 转位酶E. IF49. 翻译后加工不.包括A. 由分子伴侣辅助折叠成空间构象B. 新生多肽链从tRNA上水解释放C. 亚基聚合D. 氨基酸残基的共价修饰E. 辅基的结合50. 关于分子伴侣的叙述,错误..的是A. 识别并结合非天然构象的蛋白质B. 自身是一类较为保守的蛋白质C. 热休克蛋白是常见的分子伴侣D. 广泛存在于原核和真核细胞E. 其作用不依赖于蛋白质一级结构的信息51. 新生多肽链中,不.发生共价修饰的氨基酸残基是A. CysB. ThrC. MetD. ProE. Lys52. 下列哪项不.是信号肽的特点A. N端一小段富含亲水氨基酸的序列B. 与细胞质中的信号识别颗粒结合C. SRP受体是蛋白进入内质网所必须的D. N端一小段富含疏水氨基酸的序列E. 翻译的同时引导新生多肽链穿过内质网膜53. 引导新生多肽链靶向输送到细胞核的核酸序列称为A. signal peptideB. NLSC. KDELD. MLSE. SD54. 采用翻译后转运机制的蛋白质是A. 免疫球蛋白B. 肽类激素C. 核DNA编码的线粒体蛋白D. 凝血因子E. 水解酶55. 哪一项不.属于基因表达的范畴A. mRNA模板指导的蛋白质合成B. DNA模板指导的hnRNA合成C. DNA模板指导的DNA合成D. DNA模板指导的rRNA合成E. DNA模板指导的snRNA合成56. 下列哪一项不.是转录的原料A. TTPB. ATPC. CTPD. UTPE. GTP57. 转录生成的RNA链中存在A. dAMPC. UDPD. dTTPE. UMP58. 初级转录产物是A. 以转录调控序列为模板转录的序列B. 以结构基因为模板转录的全部序列C. 以外显子为模板转录的序列D. 以内含子为模板转录的序列E. 以上都不对59. 断裂基因的转录过程是A. 不对称、不连续B. 对称、不连续C. 对称、连续D. 不对称、连续E. 以上都不对60. 在复制和转录中均起作用的是A. RNA引物B. DNA聚合酶D. dNTPE. 蛋白质因子61. 比较复制和转录的聚合酶,错误..的说法是A. 底物不同B. 催化聚合反应的方向不同C. 产物不同D. 碱基配对略有不同E. 模板链不同62. 以DNA为模板合成RNA的过程称为A. DNA replicationB. transcriptionC. reverse transcriptionD. translationE. RNA replication63. 转录时模板与产物之间不.存在的碱基对应关系是A. A→TB. U→AC. A→UD. C→GE. G→C64. RNA转录合成需要的酶是A. DNA-dependent DNA polymeraseB. DNA-dependent RNA polymeraseC. RNA-dependent RNA polymeraseD. RNA-dependent DNA polymeraseE. Taq DNA polymerase65. 真核生物的RNA聚合酶有几种A. 1B. 2C. 3D. 4E. 566. 原核生物的RNA聚合酶有几种A. 1B. 2C. 3D. 4E. 567. E.coli RNA聚合酶的核心酶含有的亚基是A. α、β、β'、σB. α、β、σ 、ωC. α、β'、σ 、ωD. α、β、β' 、ωE. β、β'、σ 、ω68. 大肠杆菌的转录全过程始终需要A. αβ2β'ωB. α2ββ'ωC. αββ'2ωD. α2β2β'ωE. αββ'ω69. E.coli RNA聚合酶的全酶与核心酶的差别在于A. α亚基B. β亚基C. β'亚基D. σ亚基E. 以上都包括70. 真核生物中催化合成tRNA的酶是ββ'A. α2ββ'σB. α2C. RNA polymerase ID. RNA polymerase IIE. RNA polymerase III71. 遗传密码的简并性是指A. 一种三联体密码编码一种氨基酸B. 一种三联体密码编码几种氨基酸C. 几种三联体密码编码同一种氨基酸D. 不同的三联体密码编码不同的氨基酸E. 三联体密码中第一位碱基的差异不影响编码氨基酸的特异性72. 关于各种原核生物RNA聚合酶的σ亚基,描述错误..的是A. 种类很多B. 分子量不同C. 参与转录延长过程D. 与转录起始点相结合E. 决定转录的特异性73. 原核生物RNA聚合酶的σ亚基辨认转录起始点的实质是A. DNA-蛋白质相互作用B. RNA-蛋白质相互作用C. 蛋白质-蛋白质相互作用D. DNA-DNA相互作用E. DNA-RNA相互作用74. 下列哪一项不.属于核酸A. 转录因子B. 操纵子C. 转录子D. 复制子E. 顺反子75. 下列位置关系叙述正确的是A. 启动子从不会位于转录起始点下游B. 增强子全部位于结构基因内部C. 大部分TATA盒位于转录起始点下游D. 转录调控序列全部位于基因之外E. 翻译起始密码子位于转录起始点下游76. 不.需要引物的酶是A. Klenow片段B. RNA聚合酶C. 反转录酶D. Taq DNA聚合酶E. DNA-pol δ77. 真核生物的RNA聚合酶发生哪种修饰后,进入转录延长阶段A. 磷酸化B. 去磷酸化C. 甲基化D. 去甲基化E. 乙酰化78. 大肠杆菌中催化mRNA链延长的酶是A. Klenow fragmentB. αββ'σ ω2C. DNA-pol IIIββ'ωD. α2E. RNA-pol II79. 转录起始生成RNA的第一位核苷酸最常见的是A. ATPB. AMPC. GTPD. GMPE. CMP80. 真核生物合成mRNA的起始阶段不.涉及A. Hogness boxB. release factorC. transcriptional factorD. RNA-pol IIE. cis-acting element81. 狭义的核糖体循环是指A. 翻译起始、延长和终止的全过程B. 翻译的起始过程C. 翻译延长的进位、成肽和转位过程D. 翻译的终止过程E. 翻译与翻译后加工的全过程82. 关于选择性剪接,正确的说法是A. 选择性转录内含子B. 选择性转录外显子C. 初级转录产物相同,后加工过程不同D. 成熟mRNA相同,翻译过程不同E. 初级转录产物的序列发生改变83. RNA编辑发生在哪一个层次A. 复制水平B. 转录水平C. 转录后水平D. 翻译水平E. 翻译后水平84. 真核生物中编码哪种蛋白质的mRNA不.具有poly (A) 尾A. 细胞色素CB. 胰岛素C. 角蛋白D. 血红蛋白E. 组蛋白85. 下列哪一项不.是蛋白质的分拣信号A. SD sequenceB. signal peptideC. MLSD. KDELE. NLS86. 在转录后加工过程中可以观察到A. 套索RNAB. 羽毛状结构C. 锤头状RNAD. 叉状结构E. 转录空泡87. 下列核酸-蛋白质复合物中,哪一项与真核生物的RNA合成无关..A. ribosomeB. hnRNPC. snRNPD. splicesomeE. mRNP88. 以mRNA为模板合成蛋白质的过程称为A. DNA replicationB. RNA replicationC. transcriptionD. reverse transcriptionE. translation89. 关于多聚核蛋白体的描述,正确的是A. 结合了核蛋白体的几条mRNA链聚在一起B. 一个核蛋白体连续翻译多肽链C. 一条mRNA链上结合多个核蛋白体D. mRNA、核蛋白体和多肽链的复合体E. DNA、mRNA、核蛋白体和多肽链的复合体90. 电子显微镜下观察到原核生物的基因表达呈现羽毛状结构,其根本原因在于A. 多聚核蛋白体形成B. 多个转录过程同时进行C. 多个翻译过程同时进行D. 转录与翻译偶联进行E. 新生的多肽链不脱落91. 原核生物mRNA中的哪种结构与核糖体相结合A. AUGB. SD序列C. ORFD. 5' 端非翻译区E. 3' 端非翻译区92. 原核生物的翻译起始阶段,mRNA中的SD序列与哪段序列互补结合A. 23S rRNA的5' 端序列B. 16S rRNA的5' 端序列C. 5S rRNA的5' 端序列D. 23S rRNA的3' 端序列E. 16S rRNA的3' 端序列93. 参与蛋白质合成的蛋白质因子不.包括A. 起始因子B. 延长因子C. 释放因子D. 转肽酶E. 反式作用因子94. 密码子与反密码子之间有时不严格遵守碱基配对规律,这是由遗传密码的哪种性质决定的A. 方向性B. 连续性C. 通用性D. 简并性E. 摆动性95. 遗传密码具有通用性,但下列哪一项例外..A. 大肠杆菌的基因组DNAB. 人的线粒体基因组C. 鲸鱼的核DNAD. 线虫的染色体基因组E. 小麦的染色体DNA96. 不.属于直接进入蛋白质合成的氨基酸是A. 羟赖氨酸B. 精氨酸C. 谷氨酰胺D. 瓜氨酸E. 胱氨酸97. 在翻译起始阶段发挥作用的蛋白质因子是A. IFB. EFC. RFD. RRE. 转肽酶98. 翻译的实质是A. 将4种脱氧核糖核苷酸的排列顺序转换为4种核糖核苷酸的排列顺序B. 将4种核糖核苷酸的排列顺序转换为4种脱氧核糖核苷酸的排列顺序C. 将4种脱氧核糖核苷酸的排列顺序转换为20种氨基酸的排列顺序D. 将4种核糖核苷酸的排列顺序转换为20种氨基酸的排列顺序E. 将蛋白质一级结构的信息转换为空间结构的信息99. 关于多顺反子mRNA,正确的说法是A. 几个mRNA分子有不同的开放阅读框B. 几个结构基因由不同的启动子调控转录C. 一个mRNA分子有几个开放阅读框D. 多个结构基因编码一类蛋白质E. 一个结构基因编码多种蛋白质100. 抑制真核生物蛋白质合成的抗生素是A. 放线菌酮B. 氯霉素C. 四环素D. 新霉素E. 链霉素B型题A. σB. αC. βD. β'E. CTD101. 原核生物的RNA聚合酶中,辨认转录起始点的组分是102. 原核生物的RNA聚合酶中,决定转录速度的亚基是103. 真核生物的RNA聚合酶的结构组成部分A.IFB.σ因子C.snRNPD.ρ因子E.TFII104. 与原核生物转录终止相关的是105.真核生物mRNA转录起始需要106. 真核生物mRNA剪接加工需要107. 原核生物的翻译起始需要A. 胱氨酸B. 谷氨酸C. 鸟氨酸D. 脯氨酸E. 甲硫氨酸108. 不属于...构成蛋白质的氨基酸是109. 能组成蛋白质但没有遗传密码的氨基酸是110. 能作为起始氨基酸的是111. 属于酸性氨基酸的是112. 属于亚氨基酸的是A. GTPB. UMPC. dTTPD. dAMPE. CDP113. 复制的原料包括114. 转录的原料包括115. 复制产物中包含116. 转录产物中包含117. 为翻译过程提供能量的核苷酸是A. U6 snRNAB. mRNAC. tRNAD. 5S rRNAE. 45S rRNA118. RNA聚合酶I催化合成119. RNA聚合酶II催化合成A. Taq DNA polymeraseB. Klenow fragmentC. RNA polymerase IIIββ'ωD. α2E. αββ'σ2120. 真核生物合成tRNA的过程依赖121. 原核生物合成mRNA的转录延长阶段需要122. 原核生物合成rRNA的转录起始阶段需要123. 体外催化DNA合成并能耐受高温的酶是A. 核糖体小亚基B. 核糖体大亚基C. 起始因子D. 起始氨基酰-tRNAE. mRNA124. 30S翻译起始复合物不.包括125. 70S翻译起始复合物不.包括X型题126. 基因表达的最终产物包括A.蛋白质B.mRNAC.tRNAD.rRNAE. 其他小分子RNA127. 转录的特点是A. 细胞内并非所有的基因都转录B. 不对称转录C. 特异性转录因子决定转录的时序性D. 模板可以分布在DNA的两条链上E. 不同单链上的模板转录方向一致128. 与模板链同义的是A. 正链B. 负链C. 有意义链D. 反意义链E. 编码链129. 原核生物的RNA聚合酶在转录延长过程中不.脱落的组分是A. σB. αC. βD. β'E.核心酶130. 可被RNA聚合酶保护的核酸区域是A. 5' 端帽子B. 顺反子C. Pribnow boxD. 羧基末端结构域E. –35区131. 转录生成的RNA链中,5′端第一个核苷酸通常是A. ATPB. CTPC. GTPD. CMPE. GMP132. 原核生物和真核生物的RNA聚合酶的区别在于A. 能否辨认Pribnow box或者TATA boxB.能否催化生成5' 端的四磷酸二核苷酸结构C.与模板结合需否辅助因子D.能否生成转录起始复合物E.是否对利福平敏感133. 真核生物细胞核内进行的mRNA转录后加工包括A. 5' 端加帽B.3' 端加尾C.修饰生成稀有碱基D.选择性剪接E.RNA编辑134. 基因组序列测定的研究结果表明,真核生物的基因数量远远少于预期值,你认为机体如何满足编码多种蛋白质的生理需要A. 自发突变B.多蛋白的水解修饰C.染色体易位D. RNA编辑E.选择性剪接135. 选择性剪接的作用机制包括A. 使用不同的剪接位点B.选择性使用外显子C.反式剪接D.使用不同的启动子E.使用不同的多腺苷酸化位点136. 不.属于有意义密码的是A. UGAB. UAAC. AUGD. UAGE. GUA137. 当mRNA中密码子的第三位是碱基C时,tRNA中反密码子的第一位碱基可能为A. IB. GC. UD. AE. C138. 30S翻译起始复合物和70S翻译起始复合物含有的相同组分是Met E. IF A. 50S亚基 B. 30S亚基 C. mRNA D. fMet-tRNAf139. 在真核生物的翻译起始阶段,参与大小亚基拆分的蛋白质因子是A. eIF-2B. eIF-3C. eIF-4ED. eIF-6E. eIF-4F 140. 在肽链合成的延长阶段,真核生物与原核生物的区别在于A. 反应体系B. 延长因子C. 卸载tRNA的脱落方式D. 肽链延伸的方向E. 消耗GTP141. 能占据核糖体A位的是A. fMet-tRNAB. IF-1C. eRFD. RF-3E. Val-tRNA 142. 蛋白质的生物合成过程具有高保真性,可能的机制包括A. 密码子与反密码子的碱基配对B. 氨基酰-tRNA合成酶的专一性和校正功能C. EF-GTP的存在时间极短,防止错误进位D. 耗能水解清除任何步骤出现的不正确连接E. mRNA和氨基酰-tRNA在核糖体中的正确就位(二)名词解释1. 基因表达(gene expression)2. 不对称转录(asymmetric transcription)3. 翻译(translation)4. TATA盒(TATA box)5. 套索RNA(lariat RNA)6. 选择性剪接(alternative splicing)7. RNA编辑(RNA editing)8. 密码子的简并性(degeneracy)9. 密码子的摆动性(wobble)10. SD序列(Shine-Dalgarno sequence)11. 多聚核糖体(polyribosome)12. 核糖体循环(ribosome cycle)13. 分子伴侣(molecular chaperon)14. 信号肽(signal peptide)(三)简答题1. 试比较DNA复制和转录的异同。
生物化学中文名词解释

微小RNA(miRNAs)miRNAs是一类含19~25单核苷酸的单链RNA,在3’-端有1~2个碱基长度变化,广泛存在于真核生物中不编码任何蛋白,本身不具有开放阅读框架,具有保守性、时序性和组织特异性。成熟的miRNA可以和上游或下游序列不完全配对而形成基环结构。
▲
核酸的变性与复性核酸分子具有一定的空间结构,维持这种空间结构的作用力主要是氢键和碱基堆积力。有些理化因素会破坏氢键和碱基堆积力,使核酸分子的空间结构改变,从而引起核酸理化性质和生物学功能改变,这种现象称为核酸的变性。变性核酸在适当条件下,可使两条彼此分开的链重新由氢键连接而形成双螺旋结构,这一过程称为复性。
▲
糖脂糖脂是糖和脂类以共价键结合形成的复合物,组成和总体性质以脂为主体。根据脂质部分的不同,糖脂又可分为分子中含鞘氨醇的鞘糖脂,分子中含甘油酯的甘油糖脂,由磷酸多萜醇衍生的糖脂和由类固醇衍生的糖脂。
▲
脂多糖脂多糖是糖与脂类结合形成的复合物,与糖脂不同的是在脂多糖中以糖为主体成分。常见的脂多糖有胎盘脂多糖,细菌脂多糖等。
▲
糖蛋白糖蛋白是糖与蛋白质以共价键结合的复合分子,其中糖的含量一般小于蛋白质。常见的糖蛋白包括人红细胞膜糖蛋白、血浆糖蛋白、粘液糖蛋白等。
▲
蛋白聚糖蛋白聚糖是一类由糖与蛋白质结合形成的非常复杂的大分子糖复合物,其中蛋白质含量一般少于多糖。蛋白聚糖是构成动物结缔组织大分子的基本物质,也存在于细胞表面,参与细胞与细胞,或者细胞与基质之间的相互作用等。
酪氨酸蛋白激酶
第八章 生物氧化
生物氧化
黄素蛋白
铁硫蛋白
双加氧酶
单加氧酶
过氧化氢酶
过氧化物酶
底物水平磷酸化
氧化磷酸化
解偶联剂
呼吸链
第五章 基因表达2:蛋白质翻译 分子生物学习题

第五章基因表达2:蛋白质翻译名词解释:SD序列、无义突变和错义突变、EF-Tu、同义密码填空:1.可使每个氨基酸和它相对应的tRNA分子相偶联形成一个2.核糖体包括两个tRNA分子的结合位点:即P位点,紧密结合与多肽链延伸尾端相连接的tRNA分子;即A位点,结合带有一个氨基酸的tRNA 分子。
3.蛋白质合成的起始过程很复杂,包括一系列被催化的步骤4.tRNA的三叶草型结构中,其中氨基酸臂的功能是_________,反密码环的功能是___________。
5.核糖体沿着mRNA前进时,它需要另一个延伸因子,这一步需要的水解。
当核糖体遇到终止密码、、的时候,延伸作用结束,核糖体和新合成的多肽被释放出来。
翻译的最后一步被称为,并且需要一套因子。
6.tRNA的反密码子为GGC,它可识别的密码子为和7.遗传密码中第个碱基常很少或不带有遗传信息8.真核细胞多肽合成的起始氨基酸均为,而原核细胞的起始氨基酸应为。
9.蛋白质生物合成是从_____端到______端10.原核生物中的释放因子有三种,其中RF-1识别终止密码子_____________、____________;RF-2识别__________、____________;真核中的释放因子只有___________一种。
判断:1、因为AUG是蛋白质合成的起始密码子,所以甲硫氨酸只存在于蛋白质的N末端()2、原核生物蛋白质合成的起始氨基酸为甲硫氨酸,真核生物蛋白质合成起始氨基酸为甲酰甲硫氨酸()3、核糖体小亚基最基本功能是连接mRNA和tRNA大亚基则催化肽键的形成。
()选择题:1、反密码子中哪个碱基参与了密码子的简并性()A、第一个B、第二个C、第三个D、第一个与第二个E、第二个与第三个2、“同工tRNA”是指()A、识别同义mRNA密码子(具有第三个碱基简并性)的多个tRNAB、识别相同密码子的多个tRNAC、代表相同氨基酸的多个tRNAD、由相同的氨酰tRNA合成酶识别的多个tRNA3、核糖体的E位点是()A、真核mRNA加工位点B、tRNA离开原核生物核糖体的位点C、核糖体中受EcoR限制的位点D、电化学势驱动转运的位点4、蛋白质合成所需的能量来自()A、ATPB、GTPC、ATP和GTPD、CTP5、mRNA的5’-ACG-3’密码子相应的反密码子是()A、5′-UGC-3′B、5′-TGC-3′C、5′-CGU-3 ′D、5 ′ -CGT-3 ′6、在蛋白质合成过程中,下列哪些说法是正确的?()A、氨基酸随机地连接到tRNA上去B、新生肽链从C一端开始合成C、通过核糖核蛋白体的收缩,mRNA不断移动D、合成的肽链通过一个tRNA与核糖核蛋白相连问答题:1、简述遗传密码的性质2、原核生物蛋白质合成的过程3、蛋白质前体加工包括哪些?有一个被认为是mRNA的核苷酸序列,长300个碱基,你怎样才能:1.证明此RNA是mRNA而不是tRNA或rRNA。
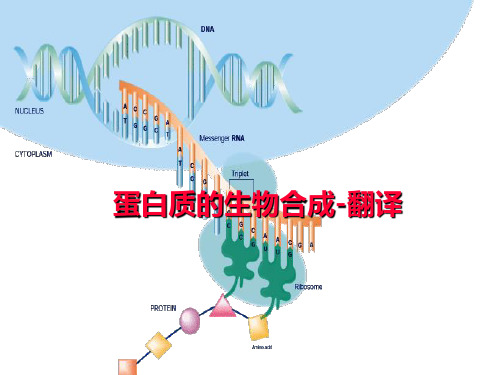
蛋白质的翻译过程
起始密码
➢肽链合成的起始
❖30s起始复合物形成
1.核糖体亚基的拆离
2.mRNA在小亚基上就位 3.fmet-tRNAfmet的结合
起始序列(SD 序列)
30S小亚基与mRNA识别、结 合
IF1、IF3协助 fmet-tRNAfmet -IF2-GTP 通 过
其反密码与mRNA上的起始密
码
AUG相配对
蛋白质的生物合成-翻译
分子生物学的中心法则(central dogma)
复制 DNA
RNA复制
转录
RNA
翻译
蛋白质
逆转录
2
翻译(蛋白质的生物合成)
蛋白质生物合成体系
➢以氨基酸为原料
➢以mRNA为模板 ➢以tRNA为运载工具 ➢以核糖体为合成场所
➢起始、延长、终止各阶段蛋白因子 参与合成后加工成为有活性蛋白质
❖氨基酰tRNA合成 酶
❖催化反应
❖氨基酰tRNA
氨基酰tRNA合成酶
A.A+特异tRNA
氨基酰tRNA
ATP AMP+PPi
氨基酸 பைடு நூலகம் ATP-E 氨基酰-AMP-E + PPi
氨基酰-AMP-E+tRNA 氨基酰tRNA+AMP+E (-COOH) (3’-CCA-OH)
16
2 、氨基酰tRNA合成酶 的高度专一性
➢核糖体与特异蛋白质、mRNA、tRNA的反应 部位
➢新技术 低温电子显微镜技术 中子散射技术
14
第二节 蛋白质合成的过程
原核生物 氨基酸的活化与转运 肽链合成的起始 肽链的延长 “核糖体循环” 肽链合成的终止 蛋白质的加工、修饰
分子生物学-蛋白质的翻译课件

详细描述
核糖体通过识别mRNA上的起始密码子与mRNA结合,形成 翻译起始复合物。这个过程需要消耗能量,以确保核糖体正 确地定位在起始密码子上。
起始复合物的形成
总结词
起始复合物的形成是翻译过程的重要步骤,它涉及到多个蛋白质和RNA分子的相互作 用。
详细描述
起始复合物的形成涉及多个步骤。首先,核糖体与mRNA结合后,需要招募翻译起始 因子,如IF3和IF2。这些因子帮助核糖体正确地定位在起始密码子上,并确保翻译的准 确性。随后,氨酰-tRNA结合到核糖体的A位点上,准备开始多肽链的合成。至此,起
肽链的延长
01
02
03
肽键的形成
氨基酸在加入到肽链中后, 通过肽键的形成相互连接, 形成多肽链。
转肽酶的作用
转肽酶在肽键形成过程中 起催化作用,促进氨基酸 之间的连接。
核糖体的移动
随着肽链的延长,核糖体 沿着mRNA移动,确保下 一个密码子被正确识别和 翻译。
终止密码子的识别
终止密码子的种类
终止密码子有UAA、UAG和UGA三种,它们作为翻译终止的信号 被核糖体识别。
翻译的起始
02
起始密码子
总结词
起始密码子是mRNA上的一个特定 序列,用于标记蛋白质合成的起始位 置。
详细描述
起始密码子是mRNA上的三个连续的 核苷酸,通常为AUG。它不仅标记了 翻译开始的位点,还决定了从这里开 始合成多肽链的方向。
核糖体与mRNA的结合
总结词
核糖体是负责蛋白质合成的细胞器,它通过与mRNA的结合 开始翻译过程。
无意义校正是指当mRNA上的终止密码子提前出现时,核 糖体会提前终止多肽链的合成。这种机制有助于减少多肽 链的错误合成。
5第五章现代分子生物学研究方法——DNA、RNA及蛋白质操作技术

DNA的基本操作技术——核酸凝胶电泳 以琼脂糖凝胶电泳为例:
DNA的基本操作技术——核酸凝胶电泳
凝胶浓度的高低影响凝胶介质孔隙的大小,浓度越高,孔隙越小, 其分辨能力就越强。反之,浓度降低,孔隙就增大,其其分辨能力 就越弱。
DNA的基本操作技术——核酸凝胶电泳
溴化乙锭(ethidium bromide,EB)能插入到DNA或RNA分子的相 邻碱基之间,并在紫外灯光照射下发出荧光,所以常用EB来检测凝 胶介质中的核酸条带。
x 25 中的聚合酶可能很多正处于复制状态,
如果此时降到室温,将会影响最终产 率。所以再留出5分钟,以使正在复制 中的DNA能够复制完全,以合成更多 的目的分子。
DNA的基本操作技术——重组载体构建 PCR完成之后,需要有什么操作?
DNA的基本操作技术——重组载体构建
DNA的基本操作技术——重组载体构建
转化(transformation):是指重组质粒DNA分子通过与膜蛋白结合 进入受体细胞(一般指细菌),并在受体细胞内稳定维持与表达的 过程。
转染(transfection):是真核细胞主动或被动导入外源DNA片段而 获得新的表型的过程。(与转化类似,只是受体细胞不同)
转导(transduction):是指通过病毒(如λ噬菌体)颗粒感染宿主细 胞将外源DNA分子导入到受体细胞内并稳定遗传的过程。
DNA的基本操作技术——聚合酶链式反应技术
DNA的基本操作技术——聚合酶链式反应技术
常用的PCR反应体系:引物、DNA聚合酶、dNTP、模板、缓冲液。
常用的PCR反应程序:
预变性 95℃ 3 min
变性 95℃ 30 s
退火 55℃ 30 s
x 25
延伸 72℃ 1 min
蛋白质翻译总结

氨基酸的活化a.起始信号(AUG-甲硫氨酸密码子)和缬氨酸(GUG)极少出现i.真核生物起始氨基酸—甲硫氨酸,原核生物-甲酰甲硫氨酸ii.SD序列:存在于原核生物起始密码子AUG上游7~12个核苷酸处的一种4~7个核苷酸的保守片段,与16srRNA3’端反向互补。
功能将mRNA的AUG起始密码子置于核糖体的适当位置以便起始翻译作用。
1)原核生物的SD序列:原核mRNA起始密码子上一段可与核糖体结合的序列。
30s小亚基首先与翻译因子IF-1(与30s结合)和IF-3(稳定小亚基,帮助其与mRNA结合位点的识别)结合,通过SD序列与mRNA模板相结合。
iii.真核生物依赖于结合5'帽,核糖体小亚基沿mRNA5'端帽子结构扫描到RBSiv.在IF2起始因子和GTP的帮助下,fMet-tRNA进入小亚基的P位,tRNA上的反密码子与mRNA密码子配对。
v.小亚基复合物与50s大亚基结合,GTP水解,释放翻译起始因子vi.翻译的起始b.后续氨基酸与核糖体的集合:第二个氨酰-tRNA与EF-Tu.GTP形成复合物,进入核糖体的A位,水解产生GDP并在EF-Ts的作用下释放GDP并使EF-Tu结合另一分子GTP形成新的循环。
i.肽键的生成:AA-tRNA占据A位,fMet-tRNA占据P位,在肽基转移酶的催化下,A位上的AA-tRNA转移到P位,P位上的起始tRNA转移至E位,与fMet-tRNA上的氨基酸生产肽键。
起始RNA随后离开。
ii.移位:核糖体通过EF-G介导的GTP水解所提供的能量向mRNA模板3'末端移动一个密码子,二肽基-tRNA完全进入P位点iii.肽链的延申c.当终止密码子UAA,UAG,UGA出现在核糖体的A位时,没有相应的AA-tRNA能与其结合,而释放因子能识别密码子并与之结合,水解P位上的多肽链与tRNA之间的二酯键,然后新生的肽链释放,核糖体大小亚基解体i.肽链的终止d.N端fMet或Met的切除i.二硫键的形成ii.特定氨基酸的修饰iii.新生肽段非功能片段的切除iv.蛋白质前体的加工e.无义突变:DNA序列中任何导致编码氨基酸的三联密码子突变转变为终止密码子UAA,UGA,UAG中的突变,使得蛋白质合成提前终止,合成无功能或无意义的多肽。
第五章分子生物学基本研究法(上)DNA、RNA及蛋白质操作技术可编辑全文

5. 2. 3聚合酶链式反应(PCR)技术 聚合酶链式反应是快速扩增DNA序列最常用 的方法。 PCR反应的模板DNA若是基因组上的某个片 段,就称为genomic PCR,若是mRNA反转 录产生的cDNA,就称为RT-PCR。
基因工程是指在体外将核酸分子插入病毒、质粒或 其它载体分子,构成遗传物质的新组合,使之进入 原先没有这类分子的寄主细胞内并进行持续稳定的 繁殖和表达。
基因工程技术是核酸操作技术的一部分,只不过它 强调了外源核酸分子在另一种不同的寄主细胞中的 繁衍与性状表达。
事实上,这种跨越物种屏障、把来自其它生物的基 因置于新的寄主生物细胞之中的能力,是基因工程 技术区别于其它技术的根本特征。
50℃~60℃,退火
引物与模板DNA相 结合
DNA合成(第三步):
将反应混合物的温度上升到72℃左右保 温1-数分钟,在DNA聚合酶的作用下, 从引物的3'-端加入脱氧核苷三磷酸,并 沿着模板分子按5'→3'方向延伸,合成新 生DNA互补链。
PCR扩增,72℃左右, 按 每 分钟1000个碱基对设计
5. 2. 1 核酸的凝胶电泳
自 从 琼 脂 糖 ( agarose ) 和 聚 丙 烯 酰 胺 (polyacrylamide)凝胶被引入核酸研究以来, 按分子量大小分离DNA的凝胶电泳技术,已经 发展成为一种分析鉴定重组DNA分子及蛋白质 与核酸相互作用的重要实验手段。
一种分子被放置到电场中,它就会以一 定的速度移向适当的电极。我们把这种 电泳分子在电场作用下的迁移速度,叫 做电泳的迁移率,它与电场强度和电泳 分子本身所携带的净电荷数成正比,与 片段大小成反比。
山东大学分子生物学章节习题及参考答案05蛋白质的生物合成(翻译)

第五章蛋白质的生物合成(翻译)一、选择题1.仅有一个密码子的氨基酸是A.色氨酸、赖氨酸B.苏氨酸、甘氨酸C.甲硫氨酸、甘氨酸D.亮氨酸、丙氨酸E.色氨酸、甲硫氨酸2.密码与反密码配对时,不遵从碱基配对规律,称为A.密码的简并性B.密码的偏爱性C.密码的连续性D.密码的摆动性E.密码的通用性3.真核生物核蛋白体中没有的rRNA是A.18SB.23SC.5SD.28SE.5.8S4.反密码存在于A.DNAB.tRNAC.mRNAD.rRNAE.cDNA5.不符合密码的通用性的细胞器是A.细胞核B.微粒体C.线粒体D.内质网E.高尔基体6.氨基酰-tRNA合成酶的校正活性是A.水解酯键B.水解3’,5’磷酸二酯键C.水解磷酸酯键D.形成酸酐键E.形成磷酸酯键7.关于核蛋白体,错误的是A.由rRNA和多种蛋白质组成B.分为大小亚基C.是翻译的场所D.在细胞核内起作用E.一个mRNA上可附着多个核蛋白体8.能促使大小亚基解离的因子是A.IF1B.IF2C.IF3D.EF-TsE.IF1与IF39.EF-Tu的功能是A.协助氨基酰-tRNA进入A位B.促进核糖体亚基聚合C.促进核糖体解聚D.促进mRNA与核糖体分离E.促进肽酰-tRNA移位10.延长因子EFG具有哪种酶的活性A.转肽酶B.酯酶C.转位酶D.转甲酰酶E.转氨酶11.肽链延长过程的叙述,错误的是A.又称为核蛋白体循环B.每循环一次延长一个氨基酸C.分为进位,成肽和转位三步D.需要EFT、EFGE.需要ATP供能12. 翻译终止时激活转肽酶为酯酶活性的是A.RF-1B.RF-2C.RF-3D.RF-4E.RR13. 蛋白质合成中不消耗能量的阶段是A.氨基酸活化B.翻译起始C.进位D.成肽E.转位14. 关于多肽链一级结构的翻译后修饰,描述错误的是A.蛋白质合成过程中N端总是甲酰甲硫氨酸B.天然蛋白质N端多数不是甲酰甲硫氨酸C.脱甲酰基酶可除去N端甲酰基D.氨基肽酶可除去N端氨基酸E.翻译终止才能除去N端甲酰基15. 鸦片促黑皮质素原水解加工生成的是A.胰岛素B.糖蛋白C.脂蛋白D.ACTHE.TSH16. 可被信号肽酶裂解的部位是A.加工区B.疏水核心区C.碱性氨基末端区D.酸性羧基末端区E.亲水区17.关于信号肽识别粒子(SRP)的描述,错误的是A.由蛋白质与RNA组成的复合体B.能特异识别结合信号肽C.具有暂停蛋白质合成的作用D.可将正在合成蛋白质的核蛋白体带至膜外E.SRP需与对接蛋白结合18. 白喉毒素可共价修饰的因子是A.EF3B.eEF1C.EF1D.eEF2E.EF219. 干扰素通过何种方式使eIF2失活A.甲基化B.ADP核糖基化C.羧化D.磷酸化E.乙酰化20. 可辨认结合分泌蛋白新生肽链N端的是A.转肽酶B.信号肽识别颗粒C.GTP酶D.RNA酶E.对接蛋白二、名词解释1. 多聚核蛋白体(polyribosome)2. 信号肽(signal peptide)3.开放阅读框架(open reading frame, ORF)三、问答题1.三种RNA在蛋白质合成中各起何作用?2.原核与真核生物翻译起始阶段各有何异同?3.细胞核蛋白合成后如何靶向输送到细胞核?4.举例说明抗生素在翻译水平抑菌的作用机理。
生物化学和分子生物学英语术语

生物化学和分子生物学英语术语*第一章蛋白质的结构与功能protein 蛋白质amino acid 氨基酸peptide bond 肽键isoelectric point 等电点peptide 肽oligopeptide 寡肽polypeptide 多肽residue 氨基酸残基amino terminal 氨基末端(N-端)carboxyl terminal 羧基末端(C-端)peptide unit 肽单元α-helix α-螺旋β-pleated sheet β-折叠β-turn β-转角random coil 无规卷曲motif 模体primary structure 一级结构secondary structure 二级结构tertiary structure 三级结构quaternary structure 四级结构domain 结构域molecular chaperone 分子伴侣protein family 蛋白质家族homologous protein 同源蛋白质cooperativity 协同效应positive cooperativity 正协同效应negative cooperativity 负协同效应allosteric effect 别构效应prion protein, PrP 朊病毒蛋白denaturation 变性renaturation 复性dialysis 透析*第二章核酸的结构与功能nucleic acid 核酸deoxyribonucleic acid, DNA 脱氧核糖核酸ribonucleic acid, RNA 核糖核酸nucleotide(,nt) 核苷酸deoxyribonucleotide 脱氧核糖核苷酸ribonucleotide 核糖核苷酸purine 嘌呤pyrimidine 嘧啶Adenine, A 腺嘌呤Guanine, G 鸟嘌呤Uracil, U 尿嘧啶Thymine, T 胸腺嘧啶Cytosine, C 胞嘧啶β-N-glycosidic bond β-N糖苷键phosphodiester bond 磷酸二酯键base sequence 碱基序列base pair, bp 碱基对kilobase pair, kbp 千碱基对anti-parallel 反向平行right-handed helix 右手螺旋major groove 大沟minor groove 小沟complementary base pair 互补碱基对complementary strand 互补链base stacking interaction 碱基堆积力telomere 端粒superhelix/supercoil 超螺旋结构chromatin 染色质chromosome 染色体nucleosome 核小体histone 组蛋白messenger RNA, mRNA 信使RNA open reading frame, ORF 开放(阅)读框codon 密码子transfer RNA, tRNA 转运RNArare base 稀有碱基stem-loop structures 茎环结构anticodon 反密码子ribosomal RNA, rRNA 核糖体RNA ribosome 核糖体non-coding RNA, ncRNA 非编码RNA DNA denaturation DNA变性hyperchromic effect DNA的增色效应renaturation 复性annealing 退火heteroduplex 杂化双链nucleic acid hybridization 核酸杂交deoxyribonuclease, DNase DNA酶ribonuclease, RNase RNA酶*第三章酶enzyme 酶monomeric enzyme 单体酶oligomeric enzyme 寡聚酶multienzyme complex 多酶复合物multienzyme system 多酶体系multifunctional enzyme 多功能酶tandem enzyme 串联酶simple enzyme 单纯酶conjugated enzyme 缀合酶(结合酶) coenzyme 辅酶prosthetic group 辅基metalloenzyme 金属酶metal activated enzyme 金属激活酶active center 活性中心essential group 必需基团binding group 结合基团catalytic group 催化基团isoenzyme/isozyme 同工酶activation energy 活化能turnover number 转换数specificity 特异性/专一性absolute specificity 绝对专一性relative specificity 相对专一性proximity effect 邻近效应orientation arrangement 定向排列desolvation 脱溶剂化surface effect 表面效应general acid-base catalysis 酸-碱催化作用nucleophilic catalysis 亲核催化covalent catalysis 共价催化electrophilic catalysis亲电催化maximum velocity 最大反应速率Michaelis constant, K m米氏常数turnover number 转换数optimum temperature 最适温度optimum pH 最适pHinhibitor 抑制剂irreversible inhibition 不可逆性抑制reversible inhibition 可逆性抑制competitive inhibition 竞争性抑制non-competitive inhibition 非竞争性抑制uncompetitive inhibition 反竞争性抑制apparent K m表观K mactivator 激活剂allosteric regulation 别(变)构调节allosteric enzyme 别构酶allosteric site 别构部位allosteric effector 别构效应剂covalent modification 共价调节zymogen 酶原*第五章维生素与无机盐vitamin 维生素lipid-soluble vitamin 脂溶性维生素water-soluble vitamin 水溶性维生素trace element 微量元素macroelement 常量元素*第六章糖代谢carbohydrates 糖(碳水化合物)Na+-dependent glucose transporter, SGLT Na+依赖型葡萄糖转运体glucose transporter, GLUT 依赖葡糖转运蛋白glycolysis 糖酵解lactic acid fermentation 乳酸发酵ethanol fermentation 乙醇发酵aerobic oxidation 有氧氧化anaerobic oxidation 无氧氧化hexokinase 己糖激酶glucokinase 葡糖激酶6-phosphofructokinase-1, PFK-1 磷酸果糖激酶-1 substrate-level phosphorylation 底物水平磷酸化phosphoenolpyruvate, PEP磷酸烯醇式丙酮酸pyruvate kinase 丙酮酸激酶fructose-2,6-biphosphate, F-2,6-BP 果糖-2,6-BP citric acid cycle 柠檬酸循环tricarboxylic acid cycle 三羧酸循环oxaloacetate 草酰乙酸citric acid 柠檬酸succinic acid 琥珀酸Pasteur effect 巴斯德效应pentose phosphate pathway 磷酸戊糖途径glycogen 糖原glycogenesis 糖原合成primer 糖原引物glycogenin 糖原蛋白branching enzyme 分支酶glycogenosis 糖原分解gluconeogenesis 糖异生blood sugar 血糖insulin 胰岛素glucagon 胰高血糖素glucose tolerance 葡萄糖耐量第七章脂质代谢lipids 脂质lipoid 类脂triglyceride, TG 甘油三酯fatty acid 脂肪酸phospholipid 磷脂ceramide 神经酰胺cholesterol 胆固醇glycerol 甘油acyl CoA 脂酰CoAchylomicron, CM 乳糜颗粒acetyl CoA 乙酰CoApalmitic acid 软脂酸citrate pyruvate cycle 柠檬酸-丙酮酸循环acetyl CoA carboxylase 乙酰CoA羧化酶fat mobilization 脂肪动员β-oxidation β-氧化carnitine acyl transferaseⅠ肉碱脂酰转移酶ketone bodies 酮体acetoacetate 乙酰乙酸β-hydroxy-butyrate β-羟丁酸acetone 丙酮3-hydroxy-3-methyl glutaryl CoA 羟甲基戊二酸单酰CoAphosphatidyl choline, PC 磷脂酰胆碱phosphatidyl ethanolamine, PE 磷脂酰乙醇胺HMG-CoA reductase HMG-CoA还原酶squalene 鲨烯bile acid 胆汁酸lipoprotein 脂蛋白very low density lipoprotein, VLDL 极低密度脂蛋白low density lipoprotein, LDL 低密度脂蛋白high density lipoprotein 高密度脂蛋白apolipoprotein, Apo 载脂蛋白第八章生物氧化biological oxidation 生物氧化oxidative respiratory chain 氧化呼吸链electron transfer chain 电子传递链nicotinamide adenine dinucleotide, NAD+烟酰胺腺嘌呤二核苷酸flavoprotein 黄素蛋白flavin mononucleotide, FMN 黄素单核苷酸coenzyme Q, CoQ, Q 泛醌;辅酶Q cytochrome, Cyt 细胞色素substrate level phosphorylation 底物水平磷酸化oxidative phosphorylation 氧化磷酸化ATP synthase ATP合酶adenylate kinase 腺苷酸激酶creatine phosphate, CP 磷酸肌酸第九章氨基酸代谢nitrogen balance 氮平衡nutritionally essential amino acid 营养必需氨基酸non-essential amino acid 非必需氨基酸nutrition value 营养价值pepsin 胃蛋白酶pepsinogen 胃蛋白酶原autocatalysis 自身激活作用endopeptidase 内肽酶exopeptidase 外肽酶γ-glutamyl cycle γ-谷氨酰基循环putrefaction 腐败作用half time, t1/2半寿期ubiquitination 泛素化aminoacid metabolic pool 氨基酸代谢库transamination 转氨基作用transaminase 转氨酶alanine transaminase, ALT 丙氨酸转氨酶aspartate transaminase, AST 天冬氨酸转氨酶transdeamination 转氨脱氨作用α-keto acid α-酮酸glucogenic amino acid 生糖氨基酸ketogenic amino acid 生酮氨基酸glucogenic and ketogenic amino acid 生糖兼生酮氨基酸alanine-glucose cycle 丙氨酸-葡萄糖循环ornithine cycle 鸟氨酸循环urea cycle 尿素循环N-acetyl glutamic acid, AGA N-乙酰谷氨酸one carbon unit 一碳单位tetrahydrofolic acid, FH4 四氢叶酸S-adenosyl methionine, SAM S-腺苷甲硫氨酸methionine cycle 甲硫氨酸循环creatine 肌酸creatine phosphate 磷酸肌酸creatinine 肌酐第十章核苷酸代谢de novo synthesis 从头合成途径salvage pathway 补救合成途径phosphoribosyl pyrophosphate, PRPP 磷酸核糖焦磷酸adenine phosphoribosyl transferase, APRT 腺苷酸磷酸核糖转移酶hypoxanthine-guanine phosphoribosyl transferase, HGPRT 次黄嘌呤-鸟嘌呤磷酸核糖转移酶uridine monophosphate, UMP 尿嘧啶核苷酸第十一章非营养物质代谢biotransformation 生物转化作用oxidation 氧化reduction 还原hydrolysis 水解conjugation 结合反应ethanol 乙醇bile 胆汁bile acid 胆汁酸heme 血红素hemoglobin 血红蛋白oxidative stress 氧化应激jaundice 黄疸第十二章物质代谢的整合与调节oxaloacetate, OAA 草酰乙酸adenylate deaminase 腺苷酸脱氨酶substrate cycle 底物循环key enzyme 关键酶multi-enzyme system 多酶体系feedback inhibition 反馈抑制covalent modification 共价修饰ubiquitin 泛素第十九章细胞信号转导的分子机制cell communication 细胞通讯signal transduction 信号转导ligand 配体receptor 受体cross-talking 交叉调控signal transduction network 信号转导网络signal transducer 信号转导分子second messenger 第二信使adenylate cyclase, AC 腺苷酸环化酶protein kinase G, PKG 蛋白激酶Gprotein phosphatase, PP 蛋白磷酸酶guanine nucleotide-binding protein, G protein 鸟苷酸结合蛋白,G 蛋白**被生物体直接用于合成蛋白质的20种氨基酸1.非极性脂肪族氨基酸甘氨酸Glycine, Gly丙氨酸Alanine, Ala缬氨酸Valine, Val亮氨酸Leucine, Leu异亮氨酸Isoleucine, Ile脯氨酸Proline, Pro2.极性中性氨基酸丝氨酸Serine, Ser半胱氨酸Cysteine, Cys甲硫氨酸Methionine, Met天冬酰胺Asparagine, Asn谷氨酰胺Glutamine, Gln苏氨酸Threonine, Thr3.含芳香环的氨基酸苯丙氨酸Phenylalanine, Phe酪氨酸Tyrosine, Tyr色氨酸Tryptophan, Trp4.酸性氨基酸天冬氨酸Aspartic acid, Asp谷氨酸Glutamic acid, Glu5.碱性氨基酸精氨酸Arginine, Arg赖氨酸Lysine. Lys组氨酸Histidine, His。
分子生物学 第五章

成多肽的氨基酸顺序,再进行比较分析来推测各氨基酸的
密码子。
三、Khorana的实验
Khorana就是用这种方法将所有的遗传密码都破译了。 这项实验还同时证实了三联密码子的准确性以及简并性的存在。
Robter Holley发现并得到了 tRNA的精确结构,应用上述各种方 法仅用了4年时间,于1965年完全查清了20种基本氨基酸所对应 的全部61个密码子,编出了遗传密码字典(教材表 5-4)。AUG 为起始密码子(initiation codon),其余61个密码子对应20种氨 基酸。另外的3个密码子(UAA、UAG和UGA)是终止密码子 (termination codons or stop codons),它们不编码任何氨基酸, 只结束蛋白质的合成。
除了色氨酸和甲硫氨酸仅有一个密码子外,其它氨基酸 都有一个以上的密码子,如:亮氨酸、精氨酸、丝氨酸 都有6个密码子,大多数氨基酸有2个、3个或4个密码子 (教材表5-5)。
二、密码的简并性(degeneracy) 密码子的简并性可以减少有害的突变,具有非常 重要的生物学意义。
三、密码的摆动性
密码的简并性往往表现在密码子的第三位碱基上,如甘 氨酸的密码子是GGU、GGC、GGA、GGG,丙氨酸的 密码子是GCU、GCC、GCA、GCG,它们的前两位碱基 是相同的,只是第三位碱基不同。有些氨基酸只有两个密 码子,通常第三位碱基或者都是嘌呤,或者都是嘧啶。 显然,密码子的专一性基本上取决于前两位碱基,第三位 碱基起的作用有限。即第三位碱基有较大的灵活性。
蛋白质分子的20种氨基酸编码,不可能是一对一的关系,两个碱基 决定一个氨基酸也只能编码16种氨基酸,如果用三个碱基决定一个 氨基酸,43=64,就足以编码20种氨基酸。这是编码氨基酸所需碱 基的最低数目,故密码子应是三联体(triplet)。
分子生物学名词解释

二1.Single-strand binding protein:Single-strand bindingprotein的中文名称是单链结合蛋白(SSB蛋白),又称DNA结合蛋白,是一种在远低于解链温度时使双链DNA分开,并牢牢地结合在单链DNA上的蛋白。
SSB蛋白的作用是保证被解链酶解开的单链在复制完成前能保持单链结构,它以四聚体形式存在于复制叉处,待单链复制完成后才离开,重新进入循环。
SSB蛋白可以保持单链的存在,并没有解链的作用。
2.SNP:SNP 全称Single NucleotidePolymorphism,中文名称为单核苷酸的多态性,是指在基因组DNA 序列中由于单个核苷酸(A、T、C和G)的突变而引起的多态性。
单核苷酸多态性是基因组最简单最常见的多态性形式,具有很高的遗传稳定性,其核苷酸变化类型主要有两种:转换,即嘌呤与嘌呤、嘧啶与嘧啶之间的转化;颠换,即嘌呤与嘧啶之间的转化。
3.半保留复制:DNA的半保留复制是指在DNA复制过程中,双螺旋的DNA分子解螺旋后,分别作为模板,按照碱基互补配对原则,在DNA聚合酶的作用下合成新的互补链,形成了两个DNA分子,与原来的DNA分子比较,每个子代分子的一条链来自亲代DNA,另一条链是新合成的一种DNA复制方式。
4.冈崎片段:冈崎片段是指在DNA半不连续复制中沿着后随链的模板链合成长度为1000~2000个碱基的短的DNA片段,能被连接形成一条完整的DNA链。
冈崎片段的长度在真核与原核生物中存在差别,真核生物的冈崎片段长度约为100~200核苷酸残基,而原核生物的冈崎片段长度约为1000~2000核苷酸残基。
5.拓扑异构酶(topoisomerase):拓扑异构酶是指通过切断DNA链中的磷酸二酯键然后重新缠绕和封口来改变两条链的环绕次数的酶在复制过程中,随着DNA的解旋,双螺旋的盘绕数T减少,而超螺旋数W增加,使正超螺旋增加,未解链部分的缠绕更加紧密,形成的压力使解链不能继续进行。
分子生物学 第五章 蛋白质的生物合成

密码利用率(codon usage)
从一般规律看,当密码子的第1、2碱基 为嘧啶碱基,生物在进化过程中往往选择 第3碱基为嘌呤碱基的密码子来编码结构基 因的氨基酸信息。
了解不同生物的密码子使用率对实施遗 传转化,基因表达,基因工程以及蛋白质 工程研究具有重要的理论指导意义。
(三)rRNA和核蛋白体
蛋白质生物合成的 场所。
核糖体的主要作用
一是提供tRNA,mRNA和相关蛋白质因子的 结合位置,使它们在核糖体上保持正确的相对位 置;
二是包括rRNA在内的组分具有催化功能,能 执行翻译中许多关键的化学反应。
核糖体在胞内除了以多聚核糖体形式参 与蛋白质合成外,还有一部分以游离状态 存在,游离状态的核糖体占总核糖体20% 左右。
当第2661个核苷酸由G突变为C后,氨 基酰tRNA-EF-Tu-GTP复合体进入A位的速 度明显减慢,当第2252、第2253位的G双 突变为C,将对转肽酶的活性产生抑制。
核 糖 体 的 组 装
• 大肠杆菌核糖体的
空间结构为一椭圆 球体,其30S亚基 呈哑铃状,50S亚 基带有三角,中间
凹陷形成空穴,将 30S小亚基抱住, 两亚基的结合面为
遗传密码的连续性
遗传密码的摆动配对
密码的简并性具有的生物学意义
它允许生物体的DNA碱基有较大变异 的余地,使基因突变可能造成的为害降至 最低程度,而不影响物种形状的表达,对 环境的适应和物种遗传的稳定。
例如细菌DNA中G+C含量变动很大, 但不同G+C含量的细菌却可以编码出相同 的多肽链。
这归因于同义密码子的分布规则。
除了少数例外(如脊椎动物线粒体基 因组编码的tRNA),所有tRNA分子都有三 叶草式的二级结构。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
---aa 接受臂 ;
loading aa at 3‟ end ---TΨ C loop; contact with 5s rRNA
---DHU loop;
contact with AARS ---anti-codon loop;
34
---extra loop;
分类标准
tRNACys表示转运半胱氨酸的tRNA Cys-tRNACys 表 示 携 带 了 半 胱 氨 酸 的 tRNACys
●
Paracodon ---由若干Nt组成,存在于tRNA不定位置上 ---与AARS侧链基团的分子发生特异的“契合” ---成为tRNA准确负载氨基酸的机制之一
●
tRNA的“L”三维结构与功能 “L”构型的结构力 ---二级结构中双链区的碱基堆积力和氢键 ---二级结构中非双链区在“L”结构中,形成氢键结合
成为核糖体识别第一个 AUG的信号
细菌 16S
AMEAMECCUGCGGUUGGAUGACCUCCUU
哺乳动物 18S rRNA AMEAMECCUGCGGAAGGAUGAUUA
高度相似
5.2.3. mRNA
mRNA分子5‟端的序列对于起始密码子选择 有重要作用
● In Prokaryote
poly-cistron
a) 简并现象的概念; ---一种氨基酸受2个或2个以上codon编码的遗传现象 ---编码一种aa的4个codon间, 仅3rd Nt 不同, 称为codon family 例;Ser (6 codons) 1 codon family & 2 extra codons
密码子的简并性
b) 简并现象的机理;
5.3. Genetic Code
5.3.1. Universal triplet codon
codon的特征
●
codon是mRNA 上连续排列的三个核苷酸序列, 并编码一个氨基酸信息的遗传单位 codon具有通用性与保守性(除mt)
●
●
在一个基因序列中 codon具有5‟→3‟方向编码、不
重叠性和无标点性
--- 34th摇摆位点被修饰的频率高
导致配对原则的改变 尤以A34 I I=A/ I= C / I=U
--- 34th几乎无 A
--- 线粒体中 U34 = N
when U U* = A/G only
5.3.3. tRNA abundance & codon usage (codon bias)
● tRNA as a adapter for codon & amino acid ● tRNA loading aa by aa-tRNAaa 合成酶 (AARS) & paracodon ● tRNA recognition codon by anti-codon
● codon;
universal triplex codon two of three reading codon codon in codon
密码子的破译 (1968. nobel prize)
Marshall Nirenberg (1961) In vitro Poly(U) Poly(C) poly(Phe) peptide poly(Pro) peptide
Poly(A)
Poly(G)
poly(Lys) peptide
poly(Gly) peptide poly(Ser-Leu-Ser-Leu…)
5.2. 基 本 元 件
5.1.1. tRNA
● mini RNA, 4S, (74-93 Nt)
● tRNA phe, 77Nt cloverleaf form (1964 Holly R.) ● Nt more modified by methylation
● 4 arms & 4 loops
生物GC%不等 各种codon的频率不等 进化过程
中度重复基因tRNA的拷贝数与codon使用频率的对应
--- 识别同一氨基酸的不同tRNA(isoacceptor)量不等
--- 不同生物间同一isoacceptor的量不等
tRNA abundance ; codon usage (codon bias) 是进化中形成的基因表达调控机制之一
Codon usage Observed for E.coli Ribosome Protein
1209 codons
Codon usage in the genes of Animals
2244 codons
a) tRNA abundance ~ 正相关 ~ codon usage
有关codon usage 高
Codons Comparing between in usual and in mt
密码子 AGA AUA
UGA
通常含义 Arg Ile
终止密码子
线粒体 终止密码子 Met
Trp
● Codon-reading
“三中读二”方式(密码子家族中)
codon anti-codon UCU-----UCA-----UCG-----UCC AGU
tRNAaa Ribosome Nitrocellulose filter
Ser-C14….
Leu-C14 ….
Lys-C14 ….
Gly-Cห้องสมุดไป่ตู้4 ….
61个codons被破译,仅剩UAA,UAG,UGA (Stop codon).
Genetic codon字典
5.3.2. Degeneracy of codon (密码子的简并现象)
真核生物核糖体80S
●
5s rRNA 与TΨ C loop of tRNA部分同源,并可配对
● In Prok.
3‟-end of 16S rRNA富含CCU保守序列 互补
mRNA 5‟ 端的前导序列 Shine-Dilgarno seq, 富含(AGG)
● 23s rRNA
--- 6 domains
But
poly(UCUCUC…)
UCU/CUC
Ser/Leu ?
M. Nirenberg & P. Leder (1964. Science 145;1399) In vitro UCU (tri-nucleotide) Ser-C14, Leu, Lys, Arg,… Ser, Leu-C14, Lys, Arg,… Ser, Leu, Lys-C14, Arg,… ……
● codon degeneracy;
wobble hypothesis
isoacceptor ● codon usage (codon bias)
● mechanism of exactly translation initiation, loading, elongation, proofreading
5‟-end;
300±Nt leading seq. (A/G-------------AUG) Shine-Dalgarno seq. (S.D): 富含嘌呤
S.D seq---------------------AUG 9Nt better rich A,U, → G mut. translation go down
aa’-tRNAaa
--- TΨ C loop & DHU loop 位于“L”两臂的交界处, 利于“L”结构的稳定 ---“L”结构中碱基堆积力大 使其拓扑结构趋于稳定
--- wobble base
位于“L”结构末端 堆积力小 自由度大 使碱基配对摇摆
5.2.2. rRNA
Ribosomal genes (rDNA) are different
---有的与对抗生素的抗性有关 ---2660±Nt region binding with complex of aa-tRNAaa~(EF)-Tu~GTP
In Euk.
3‟-end of 18s rRNA 与原核生物高度相似,
但无与 S.D.seq.互补的保守序列
在 mRNA的AUG上游存在CCACC核糖体扫描序列
•but other RNA synthesized in nucleoplasm
●
Prokaryote 23s, 16s, 5s / Eukaryote 28s-5.8s, 18s, 5s
●
Rich methylation (m2U, m3A, m3U, m26A(二甲基)…)
细菌核糖体70S
有关codon usage 低
相应tRNA量多
相应tRNA量少
b) codon 与anti-codon间的作用强度
codon usage
G … U 弱氢键配对
aa-tRNAaa 以求结合稳定
需较长时间
into A site of ribosome
G C 强氢键配对
aa-tRNAaa
需较长时间
融解温度高
● In Eukaryote
leading seq.
mono-cistron
5‟ m7Gppp--- -----CCA/GCC-----A/G-3---A1U2G3G4—
核糖体小亚基扫描AUG 的信号序列
至关准确翻译起始
But mRNA of chloroplast shows similarities to prokaryote.
from other nuclear gene
• have GC content of 60% • unusually repetitive, each cell contains from several hundred to over 20,000 copies of rDNA gene • rRNA synthesized in nucleolus