转录和转录组学优秀课件
合集下载
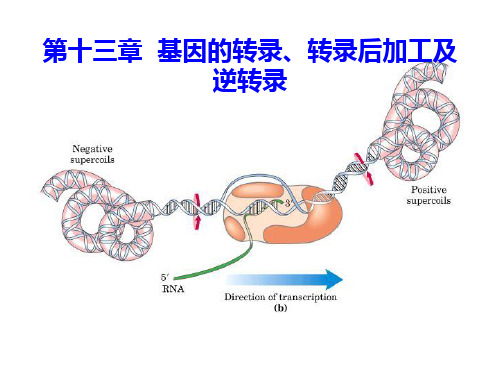
第十三章基因的转录、转录后加工及逆转录ppt课件
◆真核生物启动子 (1)DNA序列在转录起始点的5’端区(上游 区)(2)-25bp :TATA盒(Hogness box) (3)-90bp :GC盒 (4)-70bp :CAAT盒
-90
-70
GC
CAAT
RNA聚合酶Ⅱ催化的转录起始
RNA聚合酶Ⅱ催化各种前体mRNA的合 成
需要多种TF参与:TFⅡA-J
第一节 参与转录的酶
RNA聚合酶——依赖DNA的RNA聚合酶 (DNA-dependent RNA polymerase,DDRP)
以DNA为模板,催化2个游离的NTP 形成3’,5’-磷酸二酯键
一、原核生物RNA聚合酶
1、大肠埃希菌RNA聚合酶的组成 (1)全酶(holoenzyme)
由4种(5个)亚基α2ββ’σ组成 (2)核心酶(core enzyme)
作用位置
步骤1 步骤2
200~250
(3)真核生物mRNA转录后加工—剪接
内含子
外显子
DNA
hnRNA
●剪接所需条件
snRNA (U1-U6) + 蛋白质 (核内小分子核酸)
多种snRNP (核内小核蛋白颗粒)
多种snRNPs装配成
剪接体 (参与剪接过程)
4、RNA编辑(RNA editing)
二.真核生物RNA转录后的加工 1、rRNA转录后的加工
真核生物rRNA 的基因
(rDNA)
转录产物
成簇纵列串联排列
高度重复序列DNA
核质:(Ⅲ)--不需加 工
5s rRNA
核仁:(Ⅰ)--加工
5.8s rRNA 28s rRNA 18s rRNA
rDNA 内含子
基因间隔
转录和转录组学transcriptome PPT课件

DDRP )(1种)
1.2.1.组成
• 全酶: 2´ (核心酶 + ) • 核心酶 : 2´
1.2.2.作用
• α亚基: 决定那些基因被转录。 • β亚基: 催化与模板配对的相邻NTP
以3´, 5´-磷酸二酯键相连。
• β´亚基:促进酶与模板链结合,并使
DNA双链打开。 ( 核心酶: 催化RNA链的延长,参与整个 过程。)
• 1.单链小分子; • 2.含有稀有碱基或修饰碱基; • 3. 5′端总是磷酸化, 5′末端往往是pG; • 4. 3′端是CpCpAoH序列; • 5.三叶草结构; • 6.三级结构是倒L型。
三级结构呈倒L形
2.3 rRNA:
• 原核生物:70S--由50S和30S 组成 • 真核生物:80S--由60S和40S 组成
个茎。1~3个环,含13b保守序 列CAAA,AC,AGUC,GUG
核苷酸链断裂点
槌头状结构,最简单的核酶
核酶的意义
• 动摇了酶是蛋白质的传统概念。 • 为地球上生命起源早期可能是先出
现RNA提供证据。
• 为人工合成核酶以破坏某些病原微
生物,消除体内有害基因提供理论 基础。
2.5 核内不均一RNA(hnRNA)
2.7 反义RNA:
• 可与mRNA形成双链,抑制翻译。
2.8 microRNA 调节mRNA的水平
二、RNA的合成----转录(transcription)
指在RNA聚合酶催化下,以DNA为模板, NTP为原料,合成RNA的过程。
转录概述
• DNA为模板合成RNA的过程 • RNA聚合酶 • 原料:ATP,UTP,CTP,GTP (NTP) • Mg2+,Mn2+ • 合成方向:5´→3´ • 连接方式:3´,5´-磷酸二酯键
1.2.1.组成
• 全酶: 2´ (核心酶 + ) • 核心酶 : 2´
1.2.2.作用
• α亚基: 决定那些基因被转录。 • β亚基: 催化与模板配对的相邻NTP
以3´, 5´-磷酸二酯键相连。
• β´亚基:促进酶与模板链结合,并使
DNA双链打开。 ( 核心酶: 催化RNA链的延长,参与整个 过程。)
• 1.单链小分子; • 2.含有稀有碱基或修饰碱基; • 3. 5′端总是磷酸化, 5′末端往往是pG; • 4. 3′端是CpCpAoH序列; • 5.三叶草结构; • 6.三级结构是倒L型。
三级结构呈倒L形
2.3 rRNA:
• 原核生物:70S--由50S和30S 组成 • 真核生物:80S--由60S和40S 组成
个茎。1~3个环,含13b保守序 列CAAA,AC,AGUC,GUG
核苷酸链断裂点
槌头状结构,最简单的核酶
核酶的意义
• 动摇了酶是蛋白质的传统概念。 • 为地球上生命起源早期可能是先出
现RNA提供证据。
• 为人工合成核酶以破坏某些病原微
生物,消除体内有害基因提供理论 基础。
2.5 核内不均一RNA(hnRNA)
2.7 反义RNA:
• 可与mRNA形成双链,抑制翻译。
2.8 microRNA 调节mRNA的水平
二、RNA的合成----转录(transcription)
指在RNA聚合酶催化下,以DNA为模板, NTP为原料,合成RNA的过程。
转录概述
• DNA为模板合成RNA的过程 • RNA聚合酶 • 原料:ATP,UTP,CTP,GTP (NTP) • Mg2+,Mn2+ • 合成方向:5´→3´ • 连接方式:3´,5´-磷酸二酯键
转录组课件

个体化医疗
通过对个体转录组的分析,有助 于实现个体化医疗和精准治疗, 提高治疗效果和降低副作用。
02
CATALOGUE
转录组测序技术
转录组测序技术的原理
转录组测序技术基于下一代测序技术,通过对基因转录本的检测和分析,揭示基因的表达水平和调控 机制。
该技术通过将生物体的基因组DNA切割成小片段,然后对这些小片段进行测序,再通过计算机技术将这 些测序数据进行比对和分析,从而确定每个基因的转录本。
转录组测序技术的应用
转录组测序技术在生命科学、 医学和农业等领域具有广泛的
应用价值。
在生命科学领域,转录组测序 技术可用于研究基因的表达模 式和调控机制,揭示生命活动
的奥秘。
在医学领域,转录组测序技术 可用于诊断疾病、预测疾病进 展和评估治疗效果等。
在农业领域,转录组测序技术 可用于研究作物生长发育、抗 逆性和品质等性状,为育种提 供有力支持。
转录组学还能够研究药物对机体其他系统的影响,有助于全面了解药物的 副作用和相互作用,提高药物使用的安全性和有效性。
在个性化医疗中的作用
转录组学在个性化医疗中具有重要应用 价值,通过对个体基因表达的差异分析 ,有助于实现精准医疗和个性化治疗方 案。
通过比较不同个体之间的转录组数据,可以 发现个体间的基因表达差异和疾病易感性, 为个体化诊断和治疗提供依据。
差异表达分析
比较不同样本间基因表达量的变化,筛选显著差异表 达的基因。
基因功能注释与分类
利用基因本体论(GO)和KEGG等数据库,对基因进 行功能注释和分类。
转录组数据可视化的方法
散点图和箱线图
用于展示基因表达量和差异表达情况。
聚类分析热图
用于展示基因在不同样本间的聚类关系。
分子生物学--转录与转录后加工课件

原核 DNA
ABC
P 转录
真核
DNA P A P B P C
转录
mRNA A B C
mRNA
A
B
C
• 真核细胞转录和翻译在时间上和空间上都是分 开的,转录在细胞核,翻译在细胞质。
• 真核细胞RNApol高度分工
• 启动子更复杂和更多样性,不同的RNA聚合酶 有不同的启动子
• 真核生物转录需要许多转录因子(transcription factor,TF)的参与。
不依赖ρ因子的终止子: E.coli有两类
依赖于ρ因子的终止子
不依赖ρ因子的终止子:
• 简单终止子,又称内在终止子 (Intrinsic terminator)
• 转录终止不依赖任何辅助因子。 而依靠转录产物形成特殊的茎 环二级结构
具有茎环结构
•有茎环结构,富含GC •茎环结构后有寡聚尿苷
依赖ρ因子的终止子 (rho-dependent terminators )
• different genes may use different strands as template
(二)原核生物RNA聚合酶
DNA指导的RNA聚合酶 DNA dependent RNA polymerase(DDRP)
E.coli RNA聚合酶
全酶 (holoenzyme):
β’
α2β’ σ
模板的识别
启动子(promoter):RNA聚合酶识别,结合并 开始转录的一段DNA序列。
-35序列
TTGACA
-10序列
TATAAT
转录起 始位点
五种不同启动子的共同顺序
箭头表示突变,向上表示增加转录水平,向下表示降低转录水平
转录 ppt课件

启动子的一致序列是综合统计了多种基因的启动子序
列以后得出的结果,迄今为止,在E.coli中还没有发
现哪一个基因的启动子序列与一致序列完全一致。一
个基因的启动子序列与一致序列越相近,则该启动子
的启动效率就越高。不同基因在启动子序列上的差异,
是基因表达调控的一种重要途径。
ppt课件
18
3.2.3 转录的起始
ppt课件
13
因子与其相应的启动子
因子 基因 功能
-35区
间隔
-10区
70 rpoD 广泛
TTGACA
15-17bp TATAAT
32 rpoH 热休克 TCTCNCCCTTGAA 13-15bp CCCCATNTA
54 rpoN 氮代谢
CTGGNA
6bp
TTGCA
ppt课件
14
3.2.2 启动子的识别序列
ppt课件
12
不同的σ因子可以识别不同的启动子。如E.coli的
一般基因由σ70(Mr为70×103)识别, 枯草芽孢杆菌σ因 子的主要类型为σ43。识别热休克应激蛋白基因启动
子的为σ32,温度升高时,基因rpoH的产物σ32浓度升
高,温度回降时,σ32浓度降低。可以假设,σ70与σ32 通过竞争已有的核心酶,调控蛋白质合成的起始。诱 导σ32产生的基本信号是因温度升高引起的未折叠蛋 白的聚集。在固氮菌中识别固氮酶相关基因启动子的
为σ54,当培养基中缺乏氮时,E.coli中会有少量σ54存
在,在这种情况下,基因会转向利用其它可替代的N 源。
ω亚基由基因rpoZ编码,Mr为1.10×104,曾长期
被忽略,甚至许多人不把它作为聚合酶的组分。然而, 现在已经肯定,ω亚基是嗜热水生菌RNA pol必不可 少的组分,也是体外变性的RNA pol成功复性所必需 的,它与β亚基一起构成催化中心,稳定其与β' 亚基 的结合
列以后得出的结果,迄今为止,在E.coli中还没有发
现哪一个基因的启动子序列与一致序列完全一致。一
个基因的启动子序列与一致序列越相近,则该启动子
的启动效率就越高。不同基因在启动子序列上的差异,
是基因表达调控的一种重要途径。
ppt课件
18
3.2.3 转录的起始
ppt课件
13
因子与其相应的启动子
因子 基因 功能
-35区
间隔
-10区
70 rpoD 广泛
TTGACA
15-17bp TATAAT
32 rpoH 热休克 TCTCNCCCTTGAA 13-15bp CCCCATNTA
54 rpoN 氮代谢
CTGGNA
6bp
TTGCA
ppt课件
14
3.2.2 启动子的识别序列
ppt课件
12
不同的σ因子可以识别不同的启动子。如E.coli的
一般基因由σ70(Mr为70×103)识别, 枯草芽孢杆菌σ因 子的主要类型为σ43。识别热休克应激蛋白基因启动
子的为σ32,温度升高时,基因rpoH的产物σ32浓度升
高,温度回降时,σ32浓度降低。可以假设,σ70与σ32 通过竞争已有的核心酶,调控蛋白质合成的起始。诱 导σ32产生的基本信号是因温度升高引起的未折叠蛋 白的聚集。在固氮菌中识别固氮酶相关基因启动子的
为σ54,当培养基中缺乏氮时,E.coli中会有少量σ54存
在,在这种情况下,基因会转向利用其它可替代的N 源。
ω亚基由基因rpoZ编码,Mr为1.10×104,曾长期
被忽略,甚至许多人不把它作为聚合酶的组分。然而, 现在已经肯定,ω亚基是嗜热水生菌RNA pol必不可 少的组分,也是体外变性的RNA pol成功复性所必需 的,它与β亚基一起构成催化中心,稳定其与β' 亚基 的结合
转录与转录组学PPT讲稿

Severo Ochoa
1/2 of the prize USA New York University, College of Medicine New York, NY, USA b. 1905 (in Luarca, Spain) d. 1993
Arthur Kornberg
1/2 of the prize USA
个茎。1~3个环,含13b保守序 列CAAA,AC,Biblioteka GUC,GUG核苷酸链断裂点
槌头状结构,最简单的核酶
核酶的意义
2.4 核酶:
分三类:异体催化的剪切型;自 体催化的剪切型;内含子的自我剪 接型。
二级结构:锤头结构;13(或11) 个保守核苷酸。
• 核酶的发现
1982年Cech在研究低等真核生物四膜 虫的rRNA加工成熟过程中发现其rRNA 前体有自我剪接作用,即酶的作用,故 把有催化活性的RNA称核酶。
• 底物部分含GU,催化部分包括3
• 1.单链小分子; • 2.含有稀有碱基或修饰碱基; • 3. 5′端总是磷酸化, 5′末端往往是pG; • 4. 3′端是CpCpAoH序列; • 5.三叶草结构; • 6.三级结构是倒L型。
三级结构呈倒L形
2.3 rRNA:
• 原核生物:70S--由50S和30S 组成 • 真核生物:80S--由60S和40S 组成
The Nobel Prize in Physiology or Medicine 1959
"for their discovery of the mechanisms in the biological synthesis of ribonucleic acid and deoxyribonucleic acid"
系统生物学-第三讲-转录组学PPT课件

.
18
• 第一个被确认的miRNA——在线虫中首次发现的lin-4 和 let-7 ,可以通过部分互补结合到目的mRNA靶的3’非编 码区(3’UTRs),以一种未知方式诱发蛋白质翻译抑制, 进而抑制蛋白质合成,通过调控一组关键mRNAs的翻译 从而调控线虫发育进程。
继线虫之后,随后多个研究小组在包括人类、果蝇、植物等多 种生物物种中鉴别出数百拼接和注释表达与分类功能分析作用机理分析qpcr验证est软件平台est序列库序列的质量检查测序量监控聚类和拼接检查借助于基因组信息全长orf寻找发现全长基因研究表达基因概况的主要实验手段dnachipproteomics的先驱功能分类表达量分析交替剪接检测est特有信息microarray和genechip大规模表达谱或全景式表达谱globalexpressionprofile
.
40
表达序列标签(EST)测定及分析
1、什么是EST? 2、EST的应用 3、EST序列测定及分析过程
.
41
1、表达序列与表达序列标签概念
(1) 什么是表达序列?
基因组表达为RNA的序列: mRNA和功能RNA
(2) 什么是表达序列标签?
(expressed sequence tag, EST)
• 转录组研究是基因功能及结构研究的基础和 出发点,是解读基因组功能原件和揭示细胞 及组织分子组成所必需的。
.
34
• 转录组的特点:受到内外多种因素的调节,因而是 动态可变的。能够揭示不同物种、不同个体、不同 细胞、不同发育阶段及不同生理病理状态下的基因 差异表达信息。
.
35
• 转录组学(Transcriptomics):研究细胞在某 一功能状态下所含mRNA的类型与拷贝数;比较不
第三章转录-课件

RNA pol 执行多功能
(1) 识别DNA双链上的启动子; (2) 使DNA变性在启动子处解旋成单链; (3) 通过阅读启动子序列,RNA pol确定它
自己的转录方向和模板链。 (4)最后当它达到终止子时,通过识别停止转录。
原核生物的RNA聚合酶
大肠杆菌RNA聚合酶由2个α亚基、一个β亚基、 一个β′亚基和一个ω亚基组成,称为核心酶。加 上一个σ亚基后则成为聚合酶全酶
细菌中是RNA聚合酶识别启动 子。
二、转录机器的主要成分
(一)RNA聚合酶 1960年Weiss,S,B等发现RNA聚合酶(RNA Pol) 其特点是: (1)以核糖核苷三磷酸(rNTR)为底物; (2)主要以双链DNA为模板; (3)按5′-3′方向合成; (4)无需引物的存在能单独起始链的合成; (5)第一个引入的rNTP是以三磷酸形式存在; (6)在体内DNA双链中仅一条链作为模板; (7)RNA的序列和模板是互补的。
RNA聚合酶II:核质中,合成mRNA 前体
RNA聚合酶III:核质中,合成tRNA、 5srRNA以及一些小分子RNA前体
H
A
13-15
CCCCATNTA
σ54 rpo 氮代谢 N
CTGGNA
6
TTGCA
转录的起始从化学过程来看是单个 核苷酸与开链启动子-酶复合物相 结合构成新生RNA的5‘端,再以磷 酸二酯键的形式与第二个核苷酸相 结合,起始的终止反映在σ因子的 释放
真核生物RNA聚合酶
RNA聚合酶I:核仁中,合成rRNA前 体
研究发现, 由β和β′亚基组成了聚合酶的催化 中心,它们在序列上与真核生物RNA聚合酶的两 个大亚基有同源性。
Β亚基能与模板DNA、新生RNA链及核苷酸底物相 结合。
第五章转录ppt课件

一、E.coli RNA聚合酶
E.coli RNA聚合酶由5个亚基组成(5个 多肽链),即α2ββ′σ
αββ′σ四个亚基的分子量分别为36.5KDa、 150 KDa、160KDa和82 KDa,整个酶 分子的分子量为465KDa
分别是基因rpoA、rpoB、rpoC和rpoD的
产物
RNA聚合酶Ⅲ也存在于核质中,其功 能是合成tRNA和5S rRNA以及转录 Alu序列
在细胞质中也能发现一些RNA聚合酶 Ⅲ,它是从细胞核中渗漏出来的。
三种主要的RNA聚合酶的分子量都在 500 KDa左右(14S-15S),每种酶分子 含有两个大亚基和4~8个小亚基,每 个小亚基的分子量为10 KDa-90KDa
这些名称最早是依据它们从DEAE-纤维素柱上洗脱的 先后顺序而定出来的。后来发现不同生物的三种RNA聚 合酶的洗脱顺序并不相同,因而改用三种不同的RNA聚 合酶对于α-鹅膏蕈碱(α-amanitine)的敏感性不同来 进行区别。RNA聚合酶I基本不受α-鹅膏蕈碱的抑制, 在大于10-3 mol/L时才表现出轻微的抑制作用;RNA聚 合酶Ⅱ对于α-鹅膏蕈碱最为敏感,在10-9-10-8mol/L 浓度下就会被抑制;RNA聚合酶Ⅲ的敏感性介于RNA聚 合酶Ⅰ和Ⅱ之间,在10-5-10-4 mol/L时表现抑制作用。
与RNA聚合酶相结合的一个很小的蛋白质(MW= 10KDa),叫做ω亚基,其功能尚不清楚,有人认 为ω亚基对于RNA聚合酶的结构和功能没有太大的 影响。
原核生物的 RNA聚合酶
亚基 分子量
36512
150618
155613
70263
功能
决定哪些基因被转录 催化功能
转录ppt(共66张PPT)

RNA 聚合酶
Ⅰ
Ⅱ
Ⅲ
转录
产物
加工
45SrRNA
18SrRNA 28SrRNA
hnRNA tRNA 5SRNA
snRNA
mRNA
利福平
抑制原核生物的RNA聚合酶
鹅膏蕈碱
抑制真核生物的RNA聚合酶
利福平 主要用于治疗结核病、麻风病等
鹅膏蕈碱 鬼笔鹅膏
三、真核生物的转录产物为单顺
反子
与原核生物不同,真核生物一个转 录单位仅生成一个mRNA分子,经翻 译生成一条多肽链
(三)都形成3’, 5’-磷酸二酯键
二、复制与转录的不同点
模板
原料
酶
校读
复制 两条链 dNTP DNA聚合酶
有
转录 一条链
NTP RNA聚合酶
无
配对
产物
引物
复制
A=T GC
子代双
链DNA
需要
转录
A=U T=A
GC
mRNA tRNA rRNA
不需要
第一节 原核生物的转录
一、转录模板 二、RNA聚合酶 三、模板和酶的辨认结合
第 11 章
RNA的生物合成(转录)
DNA DNA DNA
RNA
蛋白质
中心法则
目录
前 言 复制与转录的异同 第1节 原核生物的转录
第2节 真核生物的转录特点
第3节 真核生物转录后加工
前言 复制与转录的异同
一、相同点 二、不同点
一、复制与转录的相同点
(一)服从碱基配对规则
(二)合成方向都是5’-3’
DNA-dependent RNA polymerase (二)合成方向都是5’-3’ (四)腺嘌呤脱氨成为次黄嘌呤 σ亚基从三元起始复合物上脱落后,核心酶的构象随之发生改变,并沿着模板链的3’→5’方向滑行,进入延长阶段 第1节 原核生物的转录 原核生物的RNA聚合酶仅1种,合成全部RNA 主要用于治疗结核病、麻风病等 真核生物rRNA转录后加工 α2 β β’ ω σ称为全酶
转录组学 ppt课件

Experil population A
Cell population B
AA B B
RNA extraction
Quantify pixel intensities.
AA BB
Reverse transcription
Sample A labelled with cy5 dye
组 (四)大规模平行信号测序系统
MPSS(massively parallel signature sequencing,MPSS)。
五、特点
• 转录组的特点:受到内外多种因素的 调节,因而是动态可变的。能够揭示 不同物种、不同个体、不同细胞、不 同发育阶段及不同生理病理状态下的 基因差异表达信息。
二、生产背景
• 首先提出转录组是指特定细胞在某一功能 状态下全部表达的基因总和,代表了 每一 个基因的身份和表达水平同一细胞在不同 的生长时期及生长环境下,基因表达情况 是不完全相同的,具有特定的空间。能够 提供全部 基因的表达调节系统和蛋白质的 功能相互作用的信息转录组学研究作为一 种整体的方法,改变了单个基因的研究模 式,将基因组学研究带入了高速发展的时 代
• 4、在土壤、海洋生态环境中的应 用目前宏转录组学研究大部分仍集 中在对海洋与土壤生态环境群落微 生物宏转录组的研究中,表1归纳 了部分海洋与土壤微生物宏转录组 的研究概况。
七、展望
• 转录组学可以直接反应实时环境表达 信息,这种研究不仅为微生物资源的 开发和利用提供宝贵的信息,而且也 为未培养微生物的研究提供了新的思 路,具有巨大的生物学意义。随着科 学技术的不断发展,转录组学将在微 生物群落中发挥越来越重要的作用。
Klenow label incorporation
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Roger D. Kornberg Stanford University, CA, USA
"for his studies of the molecular basis of eukaryotic transcription".
Roger D. Kornberg, born 1947 (59) in St Louis, MO, USA (US citizen). PhD from Stanford University, CA, USA. Mrs. George A. Winzer Professor in Medicine at Stanford University School of Medicine, CA, USA.
一、核糖核酸的类型、结构和功能
1. 类型 • 编码蛋白质:编码RNA和非编码RNA • 功能:rRNA, tRNA, mRNA • 小RNA (snRNA, snoRNA, scRNA)
2.核糖核酸的结构和功能
4种核苷糖以磷酸二酯键连接的长链, A、U、 C、G;戊糖是核糖。
2.1 mRNA
2.1.1.原核生物mRNA结构的特点:
Bushnell, D.A., Westover, K.D., Davis, R.E. and Kornberg, R.D. (2004) Structural basis of transcription: An RNA polymerase II – TFIIB cocrystal at 4.5 angstroms. Science 303, 983988.
2.4 核酶:
分三类:异体催化的剪切型;自 体催化的剪切型;内含子的自我剪 接型。
二级结构:锤头结构;13(或11) 个保守核苷酸。
• 核酶的发现
1982年Cech在研究低等真核生物四膜 虫的rRNA加工成熟过程中发现其rRNA 前体有自我剪接作用,即酶的作用,故 把有催化活性的RNA称核酶。
• 底物部分含GU,催化部分包括3
Gnatt, A.L., Cramer, P., Fu, J., Bushnell, D.A. and Kornberg, R.D. (2001) Structural basis of transcription: An RNA polymerase II elongation complex at 3.3 Å resolution. Science 292, 1876-1882.
• (1)多顺反子; • (2) mRNA 5′端无帽子结构,3′端无多聚
A尾;
• (3) mRNA一般没有修饰碱基。
2.2.2.真核生物mRNA结构的特点:
(1) 5′端有帽子结构; (2) 大多3′端有多聚A尾; (3)分子中可能有修饰碱基:主要有甲化; (4)分子中有编码区与非编码区。
2.2 tRNA
转录和转录组学优秀课件
Replication
DNA
Transcription
Replication
RNA
Reverse Transcription
Translation
中心法则
(the central dogma) P r o t e i n
The Royal Swedish Academy of Sciences has decided to award the Nobel Prize in Chemistry for 2006 to
Severo Ochoa
1/2 of the prize USA New York University, College of Medicine New York, NY, USA b. 1905 (in Luarca, Spain) d. 1993
Arthur Kornberg
1/2 of the prize USA
• 1.单链小分子; • 2.含有稀有碱基或修饰碱基; • 3. 5′端总是磷酸化, 5′末端往往是pG; • 4. 3′端是CpCpAoH序列; • 5.三叶草结构; • 6.三级结构是倒L型。
三级结构呈倒L形
2.3 rRNA:
• 原核生物:70S--由50S和30S 组成 • 真核生物:80S--由60S和40S 组成
The Nobel Prize in Physiology or Medicine 1959
"for their discovery of the mechanisms in the biological synthesis of ribonucleic acid and deoxyribonucleic acid"
Original scientific articles
Cramer, P., Bushnell, D.A. and Kornberg, R.D. (2001) Structural basis of transcription: RNA polymerase II at 2.8 ångstrom resolution. Science 292, 1863-1876.
Stanford University bS. t1a9n1fo8rd, CA, USA
2006年诺贝氏生理医学奖得主 Dr. Fire A and Dr Me功能 二、RNA的合成 三、真核细胞转录后修饰 四、转录水平的基因表达调控 五、RNA分析 六、转录组学
个茎。1~3个环,含13b保守序 列CAAA,AC,AGUC,GUG
核苷酸链断裂点
槌头状结构,最简单的核酶
Review article
Boeger, H., Bushnell, D.A., Davis, R., Griesenbeck, J., Lorch, Y., Strattan, J.S., Westover, K.D. and Kornberg, R.D. (2005). Structural basis of eukaryotic gene transcription. FEBS Lett. 579, 899-903.
"for his studies of the molecular basis of eukaryotic transcription".
Roger D. Kornberg, born 1947 (59) in St Louis, MO, USA (US citizen). PhD from Stanford University, CA, USA. Mrs. George A. Winzer Professor in Medicine at Stanford University School of Medicine, CA, USA.
一、核糖核酸的类型、结构和功能
1. 类型 • 编码蛋白质:编码RNA和非编码RNA • 功能:rRNA, tRNA, mRNA • 小RNA (snRNA, snoRNA, scRNA)
2.核糖核酸的结构和功能
4种核苷糖以磷酸二酯键连接的长链, A、U、 C、G;戊糖是核糖。
2.1 mRNA
2.1.1.原核生物mRNA结构的特点:
Bushnell, D.A., Westover, K.D., Davis, R.E. and Kornberg, R.D. (2004) Structural basis of transcription: An RNA polymerase II – TFIIB cocrystal at 4.5 angstroms. Science 303, 983988.
2.4 核酶:
分三类:异体催化的剪切型;自 体催化的剪切型;内含子的自我剪 接型。
二级结构:锤头结构;13(或11) 个保守核苷酸。
• 核酶的发现
1982年Cech在研究低等真核生物四膜 虫的rRNA加工成熟过程中发现其rRNA 前体有自我剪接作用,即酶的作用,故 把有催化活性的RNA称核酶。
• 底物部分含GU,催化部分包括3
Gnatt, A.L., Cramer, P., Fu, J., Bushnell, D.A. and Kornberg, R.D. (2001) Structural basis of transcription: An RNA polymerase II elongation complex at 3.3 Å resolution. Science 292, 1876-1882.
• (1)多顺反子; • (2) mRNA 5′端无帽子结构,3′端无多聚
A尾;
• (3) mRNA一般没有修饰碱基。
2.2.2.真核生物mRNA结构的特点:
(1) 5′端有帽子结构; (2) 大多3′端有多聚A尾; (3)分子中可能有修饰碱基:主要有甲化; (4)分子中有编码区与非编码区。
2.2 tRNA
转录和转录组学优秀课件
Replication
DNA
Transcription
Replication
RNA
Reverse Transcription
Translation
中心法则
(the central dogma) P r o t e i n
The Royal Swedish Academy of Sciences has decided to award the Nobel Prize in Chemistry for 2006 to
Severo Ochoa
1/2 of the prize USA New York University, College of Medicine New York, NY, USA b. 1905 (in Luarca, Spain) d. 1993
Arthur Kornberg
1/2 of the prize USA
• 1.单链小分子; • 2.含有稀有碱基或修饰碱基; • 3. 5′端总是磷酸化, 5′末端往往是pG; • 4. 3′端是CpCpAoH序列; • 5.三叶草结构; • 6.三级结构是倒L型。
三级结构呈倒L形
2.3 rRNA:
• 原核生物:70S--由50S和30S 组成 • 真核生物:80S--由60S和40S 组成
The Nobel Prize in Physiology or Medicine 1959
"for their discovery of the mechanisms in the biological synthesis of ribonucleic acid and deoxyribonucleic acid"
Original scientific articles
Cramer, P., Bushnell, D.A. and Kornberg, R.D. (2001) Structural basis of transcription: RNA polymerase II at 2.8 ångstrom resolution. Science 292, 1863-1876.
Stanford University bS. t1a9n1fo8rd, CA, USA
2006年诺贝氏生理医学奖得主 Dr. Fire A and Dr Me功能 二、RNA的合成 三、真核细胞转录后修饰 四、转录水平的基因表达调控 五、RNA分析 六、转录组学
个茎。1~3个环,含13b保守序 列CAAA,AC,AGUC,GUG
核苷酸链断裂点
槌头状结构,最简单的核酶
Review article
Boeger, H., Bushnell, D.A., Davis, R., Griesenbeck, J., Lorch, Y., Strattan, J.S., Westover, K.D. and Kornberg, R.D. (2005). Structural basis of eukaryotic gene transcription. FEBS Lett. 579, 899-903.