山东省科学院情报所检索中心
基于AIDUA框架的生成式人工智能使用意愿研究

2024年第36卷第2期基于AIDUA 框架的生成式人工智能使用意愿研究王伟正1,乔鸿2,李肖俊3,4,王静静5(1.山东师范大学图书馆,济南250358;2.山东师范大学商学院,济南250358;3.齐鲁工业大学(山东省科学院)数字人文研究中心,济南250014;4.齐鲁工业大学(山东省科学院)情报研究所,济南250014;5.山东大学新闻与传播学院,济南250100)摘要:[目的/意义]探究科研人员使用生成式人工智能技术意愿的影响因素,基于此促进生成式人工智能技术在学术研究及高等教育中的应用。
[方法/过程]基于认知评估理论和人工智能接受框架(AIDUA ),开发并实证检验了生成式人工智能使用意愿的理论模型,通过三阶段对用户使用意愿进行评估,旨在解释用户使用意愿的形成过程。
[结果/结论]用户使用生成式人工智能之前,经历了三阶段的决策过程。
社会影响、享乐动机和拟人化对绩效期望和努力期望均呈正相关,拟人化是影响绩效期望和努力期望的最强变量,绩效期望和努力期望对消极情绪呈负相关,而享乐动机对消极情绪的影响不显著,消极情绪与用户的使用意愿呈负相关,结果显示在校身份对努力预期和消极情绪之间具有调节作用。
关键词:生成式人工智能;ChatGPT;拟人化;用户信息行为;使用意愿;数字素养中图分类号:G252;TP18文献标识码:A文章编号:1002-1248(2024)02-0036-15引用本文:王伟正,乔鸿,李肖俊,等.基于AIDUA 框架的生成式人工智能使用意愿研究[J].农业图书情报学报,2024,36(2):36-50.收稿日期:2024-01-18基金项目:国家自然科学基金青年项目“基于多源异构数据的科技关键节点及信息扩散机理研究”(72304169)作者简介:王伟正(1999-),男,硕士研究生,研究方向为用户信息行为。
乔鸿(1973-),女,博士研究生,副教授,研究方向为社会网络、知识管理。
李肖俊(1981-),男,博士研究生,研究员,研究方向为数字人文、知识图谱、推荐系统。
DIALOG联机检索2--查新知识

辅助索引包括了除基本索引字段以为的 所有其他可检字段,即与主题内容无关 的字段,如作者字段(AU=)、出处字段 (SO=)、刊名字段(JN=)、出版年代字段 (PY=)、分类号字段(CC=)等。这些字段 随数据库的不同而有所不同。
DIALOG基本指令
一个最基本的DIALOG系统联机检索流程: 与主机系统联机,并输入用户名和密码 明确查找范围,选库并开库(Begin) 构建准确检索式并检索(Select) 按需输出合适格式记录(Type或Print) 结束检索(Logoff)
Select 或 S
选词指令
Type 或 T
联机打印指令 ? t Sn/打印格式/打印记录范围 ? tn/打印格式/打印记录范围
t s1/5/2-4 t2/ti/all t3/6,k/1,3,5
DIALOG基本指令:打印格式
预定义格式
DIALOG存取号 除文摘及正文外的全记录(包括文献的题目、出处及标引词等内容) 题录(包括文献的题目、作者、来源、出版时间及全文的字数等) 除正文以外的全记录 免费格式,一般只含题目等最基本信息 题录和文摘 短格式,题目和标引词 全记录,包括正文部分 关键词格式,可显示检索词在记录中出现的位置,并显示前后的词 语可供参考
数据库的选择方法
使用《DIALOG数据库说明书》选择数据 库 DIALOG Bluesheets 即蓝页 印刷版 活页型 联机版 DIALOG 415文档(免费) 网络版 /bluesheets/
DIALOG提供的专业信息范围
科学技术 化学工程、机械与民用工程、航空技术、计 算机科学和软件学等应用科学领域; 知识产权 专利、商标和版权信息 能源与环境 石油、天然气、电能、原子能、环境污染 与保护等; 医学 生物医学研究、医学实践、医疗仪器、药物相互 影响和治疗方法等; 制药 追踪药物的发展,包快正在开发中的药物和新药; 药物等级、许可证、临床试验等。 化学 化学物质的合成、最新发现以及商业开发;
国家科技部认可查新机构名单

国家科技部认可查新机构名单国家发明奖励评审委员会1990年公布的国家发明奖项目查新单位名单:中国科技信息研究所军事医学科学院情报研究所(即我站)国家知识产权局专利检索咨询中心中国医学科学院医学信息研究所中国化工信息中公安部科学技术情报研究所机械工业信息研究院天津市科技信息研究所冶金信息标准研究院上海科技情报研究所中国农科院科技文献信息中心辽宁省科技情报研究所中国建筑材料科学研究院技术情报中心湖北省科技情报研究所国土资源部信息中心四川省科技情报研究所中国林业科技信息研究所陕西省科技信息研究所煤炭技术信息研究国家科委《关于公布第一、二批科技查新咨询单位的通知》(国科通[1994]23号)公布的第一、二批科技查新机构名单中国化工信息中心中国农业科学院科技文献信息中心中国国防科技信息中心上海科技情报研究所机械工业信息研究院四川省科技情报研究所中国科技信息研究所天津市科技信息研究所中国航天信息中心辽宁省科技情报研究所中国医学科学院医学信息研究山东省科技情报研究所科技部西南信息中心河北省科技情报研究所中国林业科技信息研究所江苏省科技情报研究所冶金信息标准研究院黑龙江省科技情报研究所中国航空信息中心湖北省科技情报研究所中国林业科技信息研究所江苏省科技情报研究所冶金信息标准研究院黑龙江省科技情报研究所中国航空信息中心湖北省科技情报研究所中国科学院文献信息中心广西区科技情报研究所兵器工业情报研究所陕西省科技情报研究所中国有色金属工业公司技术经济研究院广东省科技情报研究国家科委《关于复审一级科技查新咨询单位的情况和公布第三批科技查新咨询单位的通知》(国科函信字[1997]005号)公布的第三批科技查新机构名单:铁道部科技信息研究国家海洋信息中心国土资源部信息中心中油集团石油经济和信息研究中心中国石化集团公司经济技术研究院山西省科技情报研究所湖南省科技信息研究所甘肃省科技情报研究所煤炭技术信息研究院云南省科技情报研究所国家一级科技查新单位名单中国化工信息中心河北省科技信息研究所中国国防科技信息中心上海市科技情报研究所机械工业信息研究院四川省科技情报研究所中国科技信息研究所广东省科技情报研究所中国航天信息中心辽宁省科技情报研究所中国医学科学院医学信息研究所山东省科技情报研究所科技部西南信息中心天津市科技信息研究所中国林业科技信息研究所江苏省科技情报研究所冶金信息标准研究所黑龙江省科技情报研究所中国航空信息中心湖北省科技情报研究所中国农业科学院科技文献信息中心广西区科技情报研究所中国有色金属工业公司技术经济研究院陕西省科技信息研究所中国航空信息中心山西省科技情报研究所中国矿产信息研究院铁道部科技信息研究所兵器工业情报研究所安徽省科技情报研究所中国石化信息研究所浙江省科技情报研究所中国石油天然气总公司信息研究所湖南省科技信息研究所国家海洋信息中心甘肃省科技情报研究所煤炭科学技术信息研究所云南省科技情报研究所。
山东省科学院情报所检索中心

山东省科学院情报所检索中心科技查新委托书(红色标记为委托人必须填写)项目名称:项目主要负责人(1-3名)委托人:委托单位:联系人:联系电话:E-mail:是否需邮寄(联系地址:邮编:)委托日期:预定完成日期:山东省科学院情报所检索中心说明:(1)查新项目名称、关键词,除国内查新外,国内外查新需要详细提供中、英文关键词,关键词的来源应是课题相关的专业惯用词,不能单纯地从一般词典中选择。
查新人员根据用户提供的主题词进行复核后确定。
(2)委托人信息中包括机构名称,通讯地址,项目负责人姓名,联系人姓名,电话,传真及E-mail。
(3)认真填写待查新项目的科学技术要点一栏,该栏内容反映查新课题的需要确立的新颖点,要着重说明查新项目在科学技术特征、技术参数或指标、应用范围等方面,查新委托人自我判断的新颖性,空间不够时可加附页。
如若报告科技要点或查新点不清楚,最好能给我们提供研究项目的技术报告。
(1、科技内容:完整简要叙述查新项目的研究内容(类似文摘),要着重说明查新项目的主要科学技术特征、技术参数或指标、应用范围、查新委托人自我判断的新颖性等2.研制总结或预研报告:用作鉴定材料。
3.技术背景:支持查新点。
)(4)查新点主要是指需要查证的内容要点。
(本查新项目与已有技术的明显不同之处,即需要查证的内容要点。
要求填写清楚,查新中途不得更改),包括课题的研究内容、特点、结构、工艺、配方以及技术参数、指标和发明点等。
(5)委托书填好后,可以网上委托(发电子邮件,电子邮件的标题及委托书名称请按照,王萍--山东大学—申报科技攻关项目查新方式发送),请务必填好联系电话,发送邮件后请及时打电话予以确认;也可以直接将报告送过来。
(6)、正常时间,课题国内查新需4-5个工作日完成,国内外查新5-6个工作日完成。
(特殊情况除外)(7)我们的地址:山东省科学院情报研究所(济南市科院路19号)科学院机关楼东三楼(从东门进3楼)(9)、附:委托书科学技术要点、查新要求书写示范委托书科学技术要点、查新要求书写示范。
申报山东省科学技术进步奖项目公示材料

申报山东省科学技术进步奖项目公示材料一、项目名称:黄河中下游粉土工程特性与路基长期性能保障技术二、推荐单位意见:黄河冲淤积土占山东省总面积的34%,该项目面向黄河中下游道路建设中存在的环境负荷重、能源效率低、资源瓶颈制约等共性问题,以黄河冲淤积粉土特殊性能判识与表征为基础,经过10多年产学研联合技术攻关和大规模的工程应用,构建了黄河冲淤积地区粉土特征分析方法和理论体系,打破了目前相关研究碎片化、关键科学问题未突破的现状,为黄泛区路基设计、施工和维护提供了理论基础;创造性地提出了黄河冲淤积地区地基工后变形控制和路基填筑和长期性能维护8项技术,研发了全寿命周期黄河冲淤积粉土路基全方位安全监测、诊断与预警系统。
形成了一整套黄河冲淤积粉土路基长期性能保障技术体系。
项目攻克了黄河冲淤积地区粉土路基长期性能保障的技术难题,成果应用于荣乌高速、青银高速等26条新建和拓宽高速公路工程,产生直接经济效益45091.9万元。
对照2017年山东省科学技术奖励条件,山东省交通运输厅推荐该项目申报山东省科技进步一等奖。
三、项目简介:本项目所属领域为交通运输工程。
黄河中下游的频繁泛滥形成了广袤的黄河冲淤积平原,仅在山东就占国土面积的34%。
在该地区建设高速公路面临优质填料匮乏和生态脆弱两大挑战。
作为不良的筑路材料,黄河冲淤积粉土毛细作用和冻敏性强烈,水稳定性、抗冲刷性以及动稳定性差。
传统的路基修建技术使得路基的长期性能得不到有效保障,持续的“桥头跳车”和反复的路基翻浆和边坡滑塌等工程恶疴严重影响了道路的安全运营和快速畅通。
本项目以黄河冲淤积粉土特殊性能判识与表征为基础,以适应黄河冲积平原特有的气候条件和地质地理环境为重点,以提升该地区粉土路基长期使用性能为目标,在多项国家基金和山东省杰出青年基金资助下,经过10多年产学研联合技术攻关,形成了黄河冲淤积粉土路基长期性能保障技术体系。
项目主要创新成果如下:(1)基于自主研发的现场交通荷载模拟装置,揭示了交通荷载作用下黄河冲淤积粉土物性变异规律和液化翻浆机制,提出了交通荷载引起的长期累积沉降现场预测方法,建立了冻融循环引起的动强度损伤演化模型;通过大型模型试验揭示了粉土路基毛细上升引起的路基土强度衰变规律,以及粉土路基边坡冲刷淘蚀机制。
不同日龄毛锤角细蜂寄生黑腹果蝇蛹的生物学特性研究

㊀山东农业科学㊀2024ꎬ56(3):115~119ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2024.03.015收稿日期:2023-03-06基金项目:国家重点研发计划项目(2023YFD1400600ꎬ2023YFE0123000ꎬ2023YFD1401200)ꎻ山东省重点研发计划项目(2023LZGC017)ꎻ国家自然科学基金青年科学基金项目(32202313)ꎻ山东省自然科学基金青年基金项目(ZR2021QC184)作者简介:代晓彦(1988 )ꎬ女ꎬ硕士ꎬ助理研究员ꎬ主要从事天敌与授粉昆虫繁育与应用研究ꎮE-mail:151****7554@163.com通信作者:刘艳(1990 )ꎬ女ꎬ博士ꎬ助理研究员ꎬ主要从事天敌与授粉昆虫繁育与应用研究ꎮE-mail:liu8882@126.com不同日龄毛锤角细蜂寄生黑腹果蝇蛹的生物学特性研究代晓彦ꎬ陈浩ꎬ王瑞娟ꎬ苏龙ꎬ高欢欢ꎬ郑礼ꎬ翟一凡ꎬ刘艳(农业农村部天敌昆虫重点实验室/山东省农业科学院植物保护研究所/山东省蜂业良种繁育中心ꎬ山东济南㊀250100)㊀㊀摘要:为了调查不同日龄毛锤角细蜂寄生黑腹果蝇的生物学特性ꎬ本研究对1~10日龄毛锤角细蜂成虫对黑腹果蝇蛹的寄生率及其后代的发育历期和雌性比进行统计ꎬ同时对寄生后黑腹果蝇蛹重的动态变化进行分析ꎮ结果表明ꎬ不同日龄毛锤角细蜂对黑腹果蝇蛹的寄生率有显著差异ꎬ7日龄寄生率最高ꎬ为(58.89ʃ2.94)%ꎬ1日龄寄生率最低ꎬ仅为(30.00ʃ1.94)%ꎬ但对毛锤角细蜂后代的发育历期和雌性比没有显著影响ꎮ黑腹果蝇蛹被毛锤角细蜂寄生后ꎬ重量显著降低ꎮ因此ꎬ1~10日龄毛锤角细蜂成虫均可大量寄生黑腹果蝇蛹ꎬ以7日龄为最佳寄生时期ꎬ这为毛锤角细蜂的大量扩繁和应用提供了理论依据ꎮ关键词:毛锤角细蜂ꎻ黑腹果蝇ꎻ寄生ꎻ日龄ꎻ生物学特性ꎻ生物防治中图分类号:S476.3:Q969.548.3㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2024)03-0115-05BiologicalCharacteristicsofTrichopriadrosophilaeatDifferentAgesafterParasitizingDrosophilamelanogasterDaiXiaoyanꎬChenHaoꎬWangRuijuanꎬSuLongꎬGaoHuanhuanꎬZhengLiꎬZhaiYifanꎬLiuYan(KeyLaboratoryofNaturalEnemyInsectsꎬMinistryofAgricultureandRuralAffairs/InstituteofPlantProtectionꎬShandongAcademyofAgriculturalSciences/ShandongApicultureBreedingCenterꎬJinan250100ꎬChina)Abstract㊀InordertostudythebiologicalcharacteristicsofTrichopriadrosophilaeatdifferentagesafterparasitizingDrosophilamelanogasterꎬthisexperimentwasconductedtoinvestigatetheparasiticrateof1~10 ̄day ̄oldT.drosophilaeadultsonD.melanogasterpupaeꎬthedevelopmentdurationandfemaleproportionofT.drosophilaeoffspringꎬandthedynamicchangeofpupaeweightofD.melanogasterafterparasitizedbyT.dros ̄ophilae.TheresultsshowedthattherewassignificantdifferenceinparasiticratebetweendifferentagesofT.drosophilaeonD.melanogasterpupae.Theparasiticrateof7 ̄day ̄oldT.drosophilaewasthehighest(58.89ʃ2.94)%ꎬandthatof1 ̄day ̄oldT.drosophilaewasthelowest(30.00ʃ1.94)%.HoweverꎬagesofT.dros ̄ophilaehadnosignificanteffectondevelopmentaldurationandfemaleproportionofitsoffspringafterparasiti ̄zingD.melanogasterpupae.InadditionꎬtheweightofD.melanogasterpupaewassignificantlyreducedafterparasitizedbyT.drosophilae.ItwasconcludedthatD.melanogasterpupaecouldbeparasitizedby1~10 ̄day ̄oldT.drosophilaeadultsꎬofwhichꎬtheagein7dayswasoptimum.Theseresultscouldprovidetheoreticalba ̄sesforbreedinginquantityandapplicationofT.drosophilae.Keywords㊀TrichopriadrosophilaeꎻDrosophilamelanogasterꎻParasitizeꎻAgeindaysꎻBiologicalcharac ̄teristicsꎻBiologicalcontrol㊀㊀黑腹果蝇(Drosophilamelanogaster)和斑翅果蝇(Drosophilasuzukii)ꎬ均属双翅目(Diptera)果蝇科(Drosophilidae)ꎬ是葡萄㊁樱桃㊁杨梅等浆果的重要害虫[1]ꎬ但两种果蝇具有生态位分离的特点ꎮ黑腹果蝇虫体小ꎬ繁殖力强ꎬ生活周期短ꎬ世代重叠ꎬ喜取食腐烂果实ꎻ而斑翅果蝇则偏爱于在新鲜的水果中产卵ꎬ孵化为幼虫后取食果肉ꎬ造成果实腐烂ꎬ进而引发病原菌的侵染ꎬ因此斑翅果蝇对水果的危害更为严重[2]ꎮ目前斑翅果蝇的防治主要以化学防治为主ꎬ但由此引起的环境污染和害虫抗药性等问题不容忽视[3]ꎬ采用天敌寄生蜂等生物防治手段则能够有效降低化学农药的使用量ꎬ被广泛应用于害虫防治ꎮ目前ꎬ国内外对果蝇寄生蜂的研究主要集中于布拉迪小环腹瘿蜂(Leptopilinaboulardi)㊁蝇蛹金小蜂(Pachycrepoideusvindemiae)和毛锤角细蜂(Trichopriadrosophilae)[3-5]ꎮ其中ꎬ毛锤角细蜂隶属膜翅目(Hymenoptera)毛锤角细蜂科(Diapri ̄idae)毛锤角细蜂属(Trichopria)[6-8]ꎬ是一种蛹期寄生蜂ꎬ能够轻易在斑翅果蝇蛹上寄生并成功发育[9]ꎬ广泛分布于韩国和中国等亚洲地区[10-11]㊁西班牙和意大利[12]等欧洲地区ꎬ以及墨西哥和美国[13]等北美洲地区ꎮ斑翅果蝇和黑腹果蝇均可被毛锤角细蜂寄生[14]ꎬ因此ꎬ黑腹果蝇可以作为斑翅果蝇的替代寄主ꎬ用于饲养毛锤角细蜂ꎬ从而进行斑翅果蝇的防治ꎮ目前国内对毛锤角细蜂的研究还处于初始阶段ꎬ主要对其种类㊁生物学特性㊁寄生机制及生防潜力等方面进行初步探索ꎬ对不同日龄的毛锤角细蜂对黑腹果蝇蛹寄生特性的影响研究甚少ꎮ本研究以毛锤角细蜂和黑腹果蝇的寄生体系为试验材料ꎬ研究毛锤角细蜂对黑腹果蝇蛹的寄生效率㊁发育历期及后代雌性比等生物学特性ꎬ寻找最合适的寄生日龄ꎬ提高室内繁育和田间应用时的寄生效率ꎻ并对寄生后黑腹果蝇蛹的重量进行测量分析ꎬ以确定规模化繁育毛锤角细蜂时收集的最佳时间ꎬ以期为杨梅㊁樱桃㊁葡萄等果园斑翅果蝇的生物防治提供参考ꎮ1㊀材料与方法1.1㊀供试虫源毛锤角细蜂和黑腹果蝇均采集于山东省济南市仲宫实验基地ꎬ饲养于山东省农业科学院植物保护研究所人工气候室ꎬ室内温度25~27ħꎬ相对湿度为(60ʃ5)%ꎬ光周期14Lʒ10Dꎮ黑腹果蝇蛹的获得:使用100目纱网制作边长30cm的正方体蝇笼饲养成蝇ꎬ在蝇笼中放置装有人工饲料的果蝇产卵盘(长30cmꎬ宽13cmꎬ高2cm)ꎮ人工饲料以玉米粉㊁酵母粉㊁蔗糖㊁琼脂㊁苹果㊁香蕉等为主要材料配制而成ꎮ24h后取出成虫ꎬ待产卵盘中卵发育至预蛹ꎬ用于寄生试验ꎮ毛锤角细蜂的饲养:将30对羽化后的寄生蜂(雌雄比为1ʒ1)放置在饲养瓶(高20cmꎬ直径13cm)中ꎬ用30%的蜂蜜水为成蜂补充营养ꎮ将100头黑腹果蝇蛹置于饲养瓶中供寄生蜂裸寄生ꎬ寄生72h后取出寄生蜂ꎬ用棉布封口ꎬ待下一代羽化成蜂ꎮ挑取羽化24h内的雌㊁雄蜂用于试验ꎮ1.2㊀不同日龄毛锤角细蜂寄生后果蝇蛹的重量变化在上述饲养条件下ꎬ收集1对羽化24h内的雌蜂和雄蜂ꎬ放入塑料养虫杯中(高5.5cmꎬ底部直径7cmꎬ开口直径9.5cm)ꎬ接入30头黑腹果蝇预蛹用于寄生ꎮ24h后转移寄生蜂至装有新鲜果蝇预蛹的塑料养虫杯中ꎬ连续重复此操作至第10天ꎮ每天称量寄生蛹的重量ꎬ直至出蜂ꎬ以未寄生蛹为对照ꎮ每处理设5个重复ꎮ1.3㊀毛锤角细蜂的寄生率、发育历期和雌性比统计将每天收集到的30头寄生蛹继续饲养直至羽化ꎬ分别统计毛锤角细蜂对黑腹果蝇的寄生率以及后代的发育历期(自卵发育至成虫羽化所需时间)和雌性比ꎮ以未寄生蛹作为对照ꎬ每处理设5个重复ꎮ寄生率(%)=被寄生的寄主蛹数量/每处理供试寄主蛹数量ˑ100ꎮ1.4㊀数据处理与分析在SPSS软件中采用方差分析法统计分析数据ꎬ采用Tukey检验进行组间差异性分析ꎬP<0.05表示差异显著ꎮ2㊀结果与分析2.1㊀不同日龄毛锤角细蜂对黑腹果蝇蛹的寄生率不同日龄毛锤角细蜂对黑腹果蝇蛹的寄生率有显著影响(F9ꎬ18=3.737ꎬP<0.05)ꎮ1~10日龄毛锤角细蜂均可寄生黑腹果蝇蛹ꎬ1日龄时寄生率最低ꎬ为(30.00ʃ1.94)%ꎬ1~3日龄寄生率随龄期增加而提高ꎻ4日龄降低ꎬ4~10日龄先增加后降低ꎬ7日龄时达到最高ꎬ为(58.89ʃ2.94)%ꎬ显著高于1日龄ꎬ但与其他日龄无显著差异(图1)ꎮ611山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第56卷㊀柱上不同小写字母表示0.05水平上差异显著ꎬ下同ꎮ图1㊀不同日龄毛锤角细蜂对黑腹果蝇蛹的寄生率2.2㊀不同日龄毛锤角细蜂寄生后代的发育历期和雌性比毛锤角细蜂寄生黑腹果蝇蛹后ꎬ其后代发育历期显著延长(F10ꎬ22=21.065ꎬP<0.01)ꎬ为20.36~22.68dꎬ但不同日龄间差异不显著(图2A)ꎻ后代雌性比没有显著差异(F9ꎬ20=2.170ꎬP=0.072)ꎬ10日龄的后代雌性比最低ꎬ为(51.42ʃ1.52)%(图2B)ꎮ2.3㊀不同日龄毛锤角细蜂寄生后黑腹果蝇蛹的重量变化被寄生蜂寄生后ꎬ果蝇蛹的每日重量变化显著(F=4.927~30.272ꎬP<0.05)ꎮ由图3可见ꎬ随图2㊀不同日龄毛锤角细蜂寄生后代的㊀㊀发育历期(A)和雌性比(B)图3㊀不同日龄毛锤角细蜂寄生后黑腹果蝇的蛹重711㊀第3期㊀㊀㊀㊀㊀代晓彦ꎬ等:不同日龄毛锤角细蜂寄生黑腹果蝇蛹的生物学特性研究着发育时间的延长ꎬ蛹重逐渐减小ꎮ其中ꎬ2日龄寄生蜂产卵后第1天蛹重为(1.11ʃ0.10)mgꎬ而出蜂前的蛹重为(0.39ʃ0.10)mgꎬ失重量较高ꎬ存在显著差异(P<0.05)ꎮ随着发育时间的延长ꎬ未被寄生的果蝇蛹重逐渐降低ꎬ且存在显著性差异(F=4.601ꎬP<0.05)ꎬ但其失重量偏小ꎬ第1天的蛹重为(1.09ʃ0.04)mgꎬ果蝇羽化前蛹体重为(0.91ʃ0.03)mgꎻ寄生蛹重量明显小于未寄生蛹ꎮ3㊀讨论与结论毛锤角细蜂是黑腹果蝇和斑翅果蝇的专性寄生蜂[10-11ꎬ14-16]ꎮ本研究结果表明ꎬ单头果蝇蛹仅能出蜂1头ꎬ使用黑腹果蝇蛹为寄主时ꎬ毛锤角细蜂平均产卵期为10dꎬ与杨健[14]的研究结果一致ꎬ但平均每只毛锤角细蜂雌虫一生可以寄生约100个果蝇蛹ꎬ与杨健的研究结果不同ꎬ这可能与寄主的质量或者饲养条件有关ꎮ不同日龄毛锤角细蜂寄生黑腹果蝇蛹后ꎬ其后代发育历期和雌性比没有显著差异ꎬ说明不同日龄毛锤角细蜂所产卵对后代的发育历期和雌性比没有影响ꎮ当毛锤角细蜂寄生黑腹果蝇后ꎬ蛹的重量明显减小ꎮ有研究表明ꎬ寄主的生长发育延缓和体重变小可能受到其体内的技术平衡[17-18]以及取食行为等因素调节[19]ꎻ毛锤角细蜂寄生黑腹果蝇蛹后ꎬ不同程度上激活了寄主的免疫相关信号通路的表达ꎬ该现象可能与寄生时对黑腹果蝇蛹造成一定程度的损伤有关ꎬ并且延迟黑化反应发生ꎬ降低黑色素对其的侵害[14]ꎮ此外ꎬ通过观察寄生前后果蝇的状态发现ꎬ果蝇蛹被寄生后ꎬ仅余蛹壳和灰黑色的蛹便ꎬ未有成形的蛹组织存在ꎬ可以推测毛锤角细蜂寄生后ꎬ逐渐蚕食寄主组织ꎬ利用寄主充足的营养完成自身的发育ꎮ本研究以毛锤角细蜂和黑腹果蝇寄生体系为对象ꎬ对不同日龄毛锤角细蜂的寄生量㊁寄生率㊁后代发育历期及雌性比等生物学特性进行了研究ꎬ确定1~10日龄的毛锤角细蜂均可用于繁育后代ꎮ此外ꎬ毛锤角细蜂寄生黑腹果蝇后ꎬ寄主的生长发育和体重都受到了显著影响ꎬ结果为后续深入研究寄生蜂调控寄主的分子机制提供了依据ꎬ为毛锤角细蜂规模化繁育及其在杨梅㊁樱桃㊁葡萄等果园斑翅果蝇的防治中的应用提供参考ꎮ参㊀考㊀文㊀献:[1]㊀张治军ꎬ郦卫弟ꎬ贝亚维ꎬ等.温度对黑腹果蝇生长发育㊁繁殖和种群增长的影响[J].浙江农业学报ꎬ2013ꎬ25(3):520-525.[2]㊀MilanNFꎬKacsohBZꎬSchlenkeTA.Alcoholconsumptionasself ̄medicationagainstblood ̄borneparasitesinthefruitfly[J].CurrentBiologyꎬ2012ꎬ22(6):488-493.[3]㊀StacconiMVRꎬBuffingtonMꎬDaaneKMꎬetal.HoststagepreferenceꎬefficacyandfecundityofparasitoidsattackingDro ̄sophilasuzukiiinnewlyinvadedareas[J].BiologicalControlꎬ2015ꎬ84:28-35.[4]㊀KaçarGꎬWangXGꎬBiondiAꎬetal.LinearfunctionalresponsebytwopupalDrosophilaparasitoidsforagingwithinsingleormultiplepatchenvironments[J].PLoSONEꎬ2017ꎬ12(8):e0183525.[5]㊀周思聪ꎬ陈佳妮ꎬ庞兰ꎬ等.布拉迪小环腹瘿蜂的生物学特性及其寄生对黑腹果蝇生长发育的影响[J].昆虫学报ꎬ2018ꎬ61(9):40-46.[6]㊀CiniAꎬIoriattiCꎬAnforaG.AreviewoftheinvasionofDro ̄sophilasuzukiiinEuropeandadraftresearchagendaforinte ̄gratedpestmanagement[J].BulletinofInsectologyꎬ2012ꎬ65(1):149-160.[7]㊀KasuyaNꎬMitsuiHꎬIdeoSꎬetal.EcologicalꎬmorphologicalandmolecularstudiesonGanaspisindividuals(Hymenoptera:Figit ̄idae)attackingDrosophilasuzukii(Diptera:Drosophilidae)[J].AppliedEntomologyandZoologyꎬ2013ꎬ48(1):87-92. [8]㊀WangXGꎬKaçarGꎬBiondiAꎬetal.Foragingefficiencyandoutcomesofinteractionsoftwopupalparasitoidsattackingtheinvasivespottedwingdrosophila[J].BiologicalControlꎬ2012ꎬ96:64-71.[9]㊀仪传冬.斑翅果蝇1种寄生蜂 毛锤角细蜂的生物学及其快繁与冷藏的基础研究[D].福州:福建农林大学ꎬ2019. [10]DaaneKMꎬWangXGꎬBiondiAꎬetal.Firstexplorationofpar ̄asitoidsofDrosophilasuzukiiinSouthKoreaaspotentialclassi ̄calbiologicalagents[J].JournalofPestScienceꎬ2016ꎬ89(3):823-835.[11]WangXGꎬKaçarGꎬBiondiAꎬetal.Life ̄historyandhostpref ̄erenceofTrichopriadrosophilaeꎬapupalparasitoidofspottedwingdrosophila[J].BioControlꎬ2016ꎬ61(4):387-397. [12]GabarraRꎬRiudavetsJꎬRodriguezGAꎬetal.ProspectsforthebiologicalcontrolofDrosophilasuzukii[J].BioControlꎬ2015ꎬ60:331-339.[13]MazzettoFꎬMarchettiEꎬAmiresmaeiliNꎬetal.Drosophilapara ̄sitoidsinnorthernItalyandtheirpotentialtoattacktheexoticpestDrosophilasuzukii[J].JournalofPestScienceꎬ2016ꎬ89(3):1-14.[14]杨健.一种果蝇蛹期寄生蜂的发现㊁生物学特性以及生防潜力的初探[D].杭州:浙江大学ꎬ2017.811山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第56卷㊀[15]ChabertSꎬAllemandRꎬPoyetMꎬetal.AbilityofEuropeanpar ̄asitoids(Hymenoptera)tocontrolanewinvasiveAsiaticpestꎬDrosophilasuzukii[J].BiologicalControlꎬ2012ꎬ63(1):40-47.[16]ZhuCJꎬLiJꎬWangHꎬetal.Demographicpotentialofthepu ̄palparasitoidTrichopriadrosophilae(Hymenoptera:Diapri ̄idae)rearedonDrosophilasuzukii(Diptera:Drosophilidae)[J].JournalofAsia ̄PacificEntomologyꎬ2017ꎬ20(3):747-751.[17]ThummelCS.MolecularmechanismsofdevelopmentaltiminginC.elegansandDrosophila[J].DevelopmentalCellꎬ2001ꎬ1(4):453-465.[18]RiddifordLMꎬHirumaKꎬZhouXFꎬetal.Insightsintothemo ̄lecularbasisofthehormonalcontrolofmoltingandmetamor ̄phosisfromManducasextaandDrosophilamelanogaster[J].InsectBiochemistryandMolecularBiologyꎬ2003ꎬ33(12):1327-1338.[19]MortonGJꎬCummingsDEꎬBaskinDGꎬetal.Centralnervoussystemcontroloffoodintakeandbodyweight[J].Natureꎬ2006ꎬ443(7109):289-295.911㊀第3期㊀㊀㊀㊀㊀代晓彦ꎬ等:不同日龄毛锤角细蜂寄生黑腹果蝇蛹的生物学特性研究。
光纤微震监测系统及其在五阳煤矿的应用研究

山东科学SHANDONGSCIENCE第36卷第5期2023年10月出版Vol.36No.5Oct.2023收稿日期:2023 ̄01 ̄04基金项目:山东省自然科学基金重点项目(ZR2020KC012)ꎻ山东省自然科学基金博士基金(ZR2019BEE019)ꎻ济南市高校20条项目(2020GXRC032)ꎻ济南市新高校20条项目(2021GXRC037)ꎻ济宁市重点研发计划项目(2011AQGX001)作者简介:张华(1992 )ꎬ女ꎬ硕士ꎬ助理研究员ꎬ研究方向为光纤传感器及应用ꎮE ̄mail:187****5976@163.com光纤微震监测系统及其在五阳煤矿的应用研究张华ꎬ胡宾鑫ꎬ朱峰ꎬ王纪强ꎬ宋广东(齐鲁工业大学(山东省科学院)激光研究所ꎬ山东济南250103)摘要:光纤微震监测技术通过观测分析生产活动中产生的微小振动事件ꎬ对其进行监测预警ꎬ具有无源㊁可靠性高等优点ꎮ传感器垂直安装在巷道帮部锚杆上ꎬ监测分站安装在硐室内ꎬ传感器与监测分站通过敷设的光缆形成监测网络ꎮ采用单纯形法进行震源定位ꎬ此方法在定位计算过程中不会出现发散问题ꎬ稳定性高ꎬ在求解过程中不需要求解偏导和逆矩阵ꎬ降低了运算量ꎬ提高了运算效率ꎬ每只传感器可以根据实际情况采用不同波速进行计算ꎬ更加符合实际情况ꎮ光纤微震监测系统安装于山西五阳煤矿ꎬ进行了初步的监测应用ꎬ并对监测结果进行了分析ꎬ结果证明该系统能够监测矿山活动ꎬ发挥预警功能ꎬ对安全生产起到了积极作用ꎮ关键词:光纤加速度传感器ꎻ微震监测ꎻ单纯形法ꎻ监测预警中图分类号:TD326㊀㊀㊀文献标志码:A㊀㊀㊀文章编号:1002 ̄4026(2023)05 ̄0060 ̄07开放科学(资源服务)标志码(OSID):OpticalfibermicroseismicmonitoringsystemanditsapplicationresearchinWuyangCoalMineZHANGHuaꎬHUBinxinꎬZHUFengꎬWANGJiqiangꎬSONGGuangdong(LaserInstituteꎬQiluUniversityofTechnology(ShandongAcademyofSciences)ꎬJinan250103ꎬChina)AbstractʒOpticalfibermicroseismicmonitoringtechnologyisusedtomonitorandalertthemicrovibrationeventsgeneratedduringproductionactivitiesthroughobservationandanalysiswithpassivityandhighreliability.Hereinꎬthesensorsareverticallyinstalledonthesideboltsalongtheroadwayꎬandthemonitoringsubstationisinstalledinthechamber.Thesensorsandthemonitoringsubstationconstituteamonitoringnetworkthroughthelaidopticalcables.Besidesꎬthesimplexmethodisusedtolocatetheseismicsource.Thismethodisfreefromdivergenceproblemsinthelocationcalculationandishighlystable.Moreoverꎬinthismethodꎬthesolutionofthepartialderivativeandinversematrixisnotrequiredꎬwhichreducesthecalculationamountandimprovesthecalculationefficiency.Additionallyꎬeachsensorcanusedifferentwavevelocitiesduringthecalculationbasedontheactualsituation.TheopticalfibermicroseismicmonitoringsystemwasinstalledinShanxiWuyangCoalMineforpreliminarymonitoringandapplicationꎬandthemonitoringresultswereanalyzed.Theresultsshowthatthesystemcanmonitormineactivitiesandwarnearlyꎬtherebyplayingapositiveroleinsafeproduction.Keywordsʒopticalfiberaccelerationsensorꎻmicroseismicmonitoringꎻsimplexmethodꎻmonitoringandearlywarning㊀㊀煤炭长期以来是我国的主要能源ꎬ是经济发展的支柱产业ꎮ随着国民经济持续快速发展ꎬ大规模深部矿产资源开采逐渐成为我国采矿行业的趋势ꎮ随着开采深度不断加大ꎬ地应力明显增大ꎬ同时产生大量的微震活动ꎬ诱发深部巷道的高强度动力灾害ꎬ造成重大人员伤亡和经济损失ꎮ以煤与瓦斯突出㊁突水㊁隧道岩爆为主的动力灾害已成为工业安全领域的主要灾害[1 ̄2]ꎮ这些动力灾害已经成为制约我国煤炭行业发展的关键因素ꎮ岩体微破裂萌生㊁扩展和贯通是煤与瓦斯突出㊁岩爆形成的重要前兆信息ꎬ对前兆信息的精确捕捉㊁测量㊁分析已成为动力灾害监测预警防治亟待突破的关键问题ꎮ研究和实践表明ꎬ微震监测技术是用岩体变形和破坏后本身发出的弹性波来监测工程岩体稳定性的技术方法ꎬ是岩体破裂监测的有效手段ꎮ于群等[3]和王创业等[4]引进加拿大ESG(EngineeringSeisnologyGroup)微震监测系统后ꎬ分别将其应用于锦屏水电站㊁大岗山水电站㊁洋山隧道等ꎬ研究微震事件分析方法ꎬ对动力灾害进行预警ꎮMa等[5]和辛崇伟等[6]对系统软硬件改进ꎬ设计了井下微震定位系统ꎬ在华丰煤矿等处实现了应用ꎮ窦林名等[7]与波兰矿业研究院合作引进波兰SOS(seismologicalobservationsystem)微震监测系统ꎬ在桃山煤矿等进行实时监测ꎮ但是ꎬ上述微震监测系统中前端传感探头均是基于压电式㊁电容式等电学理论ꎬ使用过程需要供电ꎬ易受电磁干扰ꎬ在煤矿等易燃易爆环境应用时受到限制ꎮ而光纤传感器具有灵敏度高㊁频响宽㊁动态范围大㊁本质安全等优点ꎬ由此本文设计了基于悬臂梁式加速度传感器的光纤微震监测系统ꎬ并将其应用在五阳煤矿ꎮ1㊀微震监测技术1.1㊀微震监测原理岩石在受到外力或内力作用时ꎬ其内部将会产生局部弹塑性能集中现象ꎬ当能量积累到临界值之后ꎬ将引起岩体微裂隙的产生与扩展ꎬ微裂隙的产生与扩展伴随着弹性波或应力波的释放并在周围岩体内快速传播ꎬ这种弹性波就称为微震ꎮ微震监测技术是通过观测㊁分析生产活动中产生的微小振动事件ꎬ以监测其对生产活动的影响㊁效果及地下状态的地球物理技术ꎬ可以实现三维空间连续㊁动态监测ꎬ定位精度高㊁可靠性强[8 ̄10]ꎮ当岩石或煤层因为外界因素发生破裂或移动时ꎬ产生微弱的微震信号向周围传播ꎬ微震传感器可以接收这些信号ꎬ记录微震信号的到达时间㊁传播方向等信息ꎬ利用定位算法确定破裂点ꎬ即震源位置ꎮ本文设计悬臂梁式加速度传感器ꎬ其基于惯性原理实现加速度测量ꎬ采用悬臂梁式结构ꎬ光纤布拉格光栅黏贴在悬臂梁上ꎬ当外界发生振动或冲击时ꎬ传感器的外壳和质量块会发生振动ꎬ使黏贴在悬臂梁上的光纤发生应变ꎬ从而导致光纤的中心波长发生移动ꎮ该项目监测设备采用的是自主研发生产的矿山微震监测系统ꎬ系统组成如图1所示ꎮ图1㊀矿山微震监测系统Fig.1㊀Minemicroseismicmonitoringsystem微震监测系统主要包括5支光纤微震传感器㊁微震监测分站㊁光缆㊁交换机㊁同步时钟㊁主站等ꎬ安装示意图如图2所示ꎮ图2㊀安装示意图Fig.2㊀Installationschematic㊀㊀传感器垂直安装在巷道帮部锚杆上ꎬ监测分站安装在硐室内ꎬ传感器与监测分站通过敷设的光缆形成监测网络ꎮ5通道微震监测系统可对8003回风巷的开采面产生的微震事件进行24h不间断监测ꎬ传感器将获得的微震信号通过光缆传送至微震监测分站ꎬ监测分站接收各通道的数据ꎬ将数据通过环网传送至地面主站ꎬ计算机进行数据分析ꎬ形成报告ꎮ1.2㊀微震监测系统定位震源事件的定位是微震监测技术研究的主要内容ꎬ震源事件的定位能够确定煤岩体破裂的时间和坐标ꎬ是煤岩动力灾害监测预警的基础ꎮ微震监测定位方法较多ꎬ通常根据定位原理分为两大类:一是基于三分量传感器的震源定位方法ꎻ二是基于不同到时原理的震源定位方法ꎮ第二种是目前微震定位监测中应用最广的一类震源定位方法ꎬ例如经典的Geiger法㊁粒子群算法㊁Powell算法㊁单纯形法等[11 ̄16]ꎮ经典的Geiger法属于特殊的线性定位算法ꎬ将非线性方程组采用牛顿高斯法进行线性化ꎬ计算出方程组的最小二乘解ꎬ使观测到时间与计算到时间之间的到时残差最小ꎮGeiger法采用多支传感器进行定位ꎬ极大程度上提高了定位的准确性ꎬ但是此算法过度依赖初始值的选取ꎬ若选择不当ꎬ易出现定位不准或无法定位的问题ꎮ单纯形法是一种适用于求解多维无约束优化问题的一种数值搜索方法ꎬ在连续改变几何图形的过程中ꎬ相互比较单纯形各顶点的目标函数值ꎬ逐步以目标函数值较小的顶点取代目标函数值较大的顶点ꎬ在迭代过程中将单纯形逐渐向最优点移动ꎬ从而进行优化ꎮ而单纯形法在定位计算过程中不会出现发散问题ꎬ稳定性高ꎬ在求解过程中不需要求解偏导和逆矩阵ꎬ降低了运算量ꎬ提高了运算效率ꎬ每只传感器可以根据实际情况采用不同波速进行计算ꎬ更加符合实际情况ꎬ所以本文采用的是单纯形法进行定位计算ꎬ算法步骤如下:第一步㊀计算每支传感器之间获取信号的到时差值以及各支传感器的坐标ꎬ选取5支信号完整的传感器ꎬ作为5个顶点构造初始的单纯形ꎮ通过线性定位初步计算微震震源的位置ꎬ得到初始定位坐标p0x0ꎬy0ꎬz0()ꎻ第二步㊀根据微震传感器的坐标ꎬ计算每个传感器的残差ꎮ设微震发生的时刻为t0ꎬ震源坐标为Sx0ꎬy0ꎬz0()ꎬ其中i表示探测到微震有用信号的微震传感器个数ꎬ()ꎬ第i支微震传感器的坐标为Tixiꎬyiꎬzi第i支微震传感器的微震波初至到时时刻为tiꎬ微震波到达第i支微震传感器的波速为viꎬ则震源定位方程可表达为xi-x0()ꎬ(1)()2+yi-y0()2+zi-z0()2=viti-t0根据最小二乘法原理ꎬ震源定位的目标函数可以表示为φx0ꎬy0ꎬz0ꎬt0()=ðmi=1γ2iꎬ(2)其中ꎬγi为微震传感器的残差ꎬ即观测到时和计算到时之间的差值ꎬ可用式(3)表示ꎬγi=ti-t0+tti()ꎬ(3)式中ꎬtti为第i支微震传感器的计算到时ꎻ第三步㊀比较5个点的残差ꎬ选出最大值和最小值ꎬ通过映射㊁扩展㊁压缩㊁收缩等4种形式在误差空间中将原单纯形中的顶点替换成新的单纯形ꎻ第四步㊀不断重复第三步ꎬ得到新的单纯形ꎬ使单纯形朝着空间内目标函数值最小的方向移动ꎬ直到找到最佳的震源位置ꎬ得到定位结果ꎮ算法流程图见图3ꎮ㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀图3㊀单纯形法流程图㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀Fig.3㊀Flowchartofsimplexmethod2㊀微震监测系统数据分析五阳煤矿位于山西省长治市襄垣县王桥镇ꎬ微震监测设备安装在8003回风巷ꎬ地表位于西周村㊁东元垴村㊁西元垴村之间ꎬ地面标高为+925~+958mꎮ8003回风巷位于80采区南部ꎬ工作面标高为+283~+408mꎬ其南部为80 ̄0305底抽巷ꎬ东部为80采区胶带运输巷ꎬ西部为8006放水巷ꎬ北部为实煤体ꎮ煤体结构为原生结构煤ꎬ煤岩类型以亮煤为主ꎬ暗煤次之ꎬ煤质为贫瘦煤ꎮ8003回风巷总设计长度1789mꎬ现已掘进1232mꎬ剩余557mꎮ向前掘进时选择合适位置ꎬ开始留设顶煤ꎬ在相对稳定层位ꎬ沿中煤(顶煤不超1m)掘进至切眼设计位置停掘ꎬ若因煤层酥软或受到80 ̄0305底抽巷造穴孔影响ꎬ无法留住顶煤时ꎬ仍改为沿顶板掘进至切眼设计位置ꎮ2.1㊀微震数据处理光纤微震监测系统将监测到的有效信号进行保存ꎬ在震源定位前首先对信号进行滤波和去噪ꎬ图4是微震传感器采集到的信号ꎮ对这些信号滤波ꎬ进行数据处理和分析之后ꎬ得到震源的位置坐标和能量见表1ꎮ图4㊀微震监测信号波形图Fig.4㊀Waveformofthemicroseismicmonitoringsignal表1㊀部分微震事件定位坐标22022 ̄05 ̄0103:45:25405802.54036364.6336.552878.80.0532022 ̄05 ̄0104:24:00406567.64036897.6213.613625.80.4442022 ̄05 ̄0123:04:10406465.14036738.8257.73090.60.8752022 ̄05 ̄0123:11:58406382.24036491.3324.239266.90.1462022 ̄05 ̄0201:34:29406596.34036991.5303.317065.00.3872022 ̄05 ̄0204:29:07406569.54036992.8182.022753.00.3082022 ̄05 ̄0205:32:56406734.24037062.1277.849330.70.0792022 ̄05 ̄0207:50:10406556.14036785.9323.212585.10.47102022 ̄05 ̄0207:50:30406417.94036206.9133.946056.50.09112022 ̄05 ̄0208:49:48406454.24036330.7343.032740.30.19122022 ̄05 ̄0213:45:44406333.04036356.6284.817311.10.37132022 ̄05 ̄0215:19:04406375.14036508.8330.611802.80.49142022 ̄05 ̄0218:38:39406501.24036806.3492.910352.60.52152022 ̄05 ̄0223:13:59406623.24036858.3198.913298.1-0.45㊀㊀将震源位置在矿图上进行标记ꎬ微震事件基本发生在采煤区ꎬ多是因为煤矿开采引起的ꎬ与矿上开采活动时间对应ꎬ监测效果较好ꎮ2.2㊀微震监测结果分析通过对微震事件的部分数据统计分析ꎬ绘制出折线柱状图ꎬ如图5所示ꎮ图5㊀微震事件数量分布图Fig.5㊀Distributionofmicroseismicevents2022年5月1日至2日ꎬ微震事件频次较低ꎬ5月3日至5月4日ꎬ微震事件数量频次升高ꎬ监测到大量的微震事件信号ꎬ5日至7日微震事件数量下降ꎮ对单日微震事件进行数据统计分析ꎬ绘制柱状图如图6所示ꎮ从图6可以看出ꎬ微震活动主要集中在3个时期ꎬ分别为8~10时㊁11~13时和16~18时ꎮ结合现场实际生产活动情况ꎬ这3个时间段主要为矿山的作业生产事件ꎬ发生小规模爆破活动ꎬ从而引起岩体的扰动和损伤ꎬ导致岩体内部发生微破裂ꎮ这说明微震事件的产生大部分是由矿山开采活动引起的ꎮ图6㊀微震事件日分布图Fig.6㊀Dailydistributionofmicroseismicevents3㊀小结本文采用光纤微震监测系统ꎬ进行了初步的监测应用研究ꎬ结果证明该系统性能良好ꎬ能够监测矿山活动ꎬ发挥预警功能ꎬ满足矿山微震监测的要求ꎬ具有良好的推广应用前景ꎮ光纤微震监测系统对五阳煤矿的地压监测㊁安全生产等提供了技术手段ꎬ对安全生产起到积极作用ꎬ推动了安全生产管理水平的发展ꎮ参考文献:[1]LIUJPꎬSIYTꎬWEIDCꎬetal.DevelopmentsandprospectsofmicroseismicmonitoringtechnologyinundergroundmetalminesinChina[J].JournalofCentralSouthUniversityꎬ2021ꎬ28(10):3074 ̄3098.DOI:10.1007/s11771 ̄021 ̄4839 ̄y. [2]岳东ꎬ刘超ꎬ袁增云.基于微震监测的煤层覆岩关键层判别[J].煤炭科技ꎬ2020ꎬ41(4):40 ̄43.DOI:10.19896/j.cnki.mtkj.2020.04.012.[3]于群ꎬ唐春安ꎬ李连崇ꎬ等.基于微震监测的锦屏二级水电站深埋隧洞岩爆孕育过程分析[J].岩土工程学报ꎬ2014ꎬ36(12):2315 ̄2322.DOI:10.11779/CJGE201412021.[4]王创业ꎬ谷雷ꎬ高照.微震监测技术在矿山中的研究与应用[J].煤炭技术ꎬ2019ꎬ38(10):45 ̄48.DOI:10.13301/j.cnki.ct.2019.10.016.[5]MATHꎬTANGCAꎬLIUFꎬetal.Microseismicmonitoringꎬanalysisandearlywarningofrockburst[J].GeomaticsꎬNaturalHazardsandRiskꎬ2021ꎬ12(1):2956 ̄2983.DOI:10.1080/19475705.2021.1968961.[6]辛崇伟ꎬ姜福兴ꎬ樊硕ꎬ等.矿内-矿间微震监测技术研究[J].煤炭工程ꎬ2021ꎬ53(6):107 ̄112.DOI:10.11799/ce202106021.[7]窦林名ꎬ冯龙飞ꎬ蔡武ꎬ等.煤岩灾变破坏过程的声震前兆识别与综合预警模型研究[J].采矿与安全工程学报ꎬ2020ꎬ37(5):960 ̄968+976.DOI:10.13545/j.cnki.jmse.2020.05.012.[8]MCCREARYRꎬMCGAUGHEYJꎬPOTVINYꎬetal.Resultsfrommicroseismicmonitoringꎬconventionalinstrumentationꎬandtomographysurveysinthecreationandthinningofaburst ̄pronesillpillar[J].PureandAppliedGeophysicsꎬ1992ꎬ139(3):349 ̄373.DOI:10.1007/BF00879942.[9]LIUSCꎬCHENXꎬLILXꎬetal.Microseismicreal ̄timemonitoringsystembasedonvirtualinstruments[J].InternationalJournalofHybridInformationTechnologyꎬ2016ꎬ9(1):413 ̄422.DOI:10.14257/ijhit.2016.9.1.36.(下转第84页)DOI:10.16668/j.cnki.issn.1003 ̄1421.2022.02.19.[4]HALYALS.Forecastingpublictransitpassengerdemand:WithneuralnetworksusingAPCdata[J].CaseStudiesonTransportPolicyꎬ2022ꎬ10(2):965 ̄975.DOI:10.1016/j.cstp.2022.03.011.[5]GUOJYꎬXIEZꎬQINYꎬetal.Short ̄termabnormalpassengerflowpredictionbasedonthefusionofSVRandLSTM[J].IEEEAccessꎬ2019ꎬ7:42946 ̄42955.DOI:10.1109/ACCESS.2019.2907739.[6]赵婧宇ꎬ池越ꎬ周亚同.基于SSA ̄LSTM模型的短期电力负荷预测[J].电工电能新技术ꎬ2022ꎬ41(6):71 ̄79.DOI:10.12067/ATEEE2107053.[7]吴娟ꎬ何跃齐ꎬ张宁ꎬ等.基于VMD ̄GRU的城市轨道交通短时客流预测[J].都市快轨交通ꎬ2022ꎬ35(1):79 ̄86.DOI:10.3969/j.issn.1672 ̄6073.2022.01.013.[8]马超群ꎬ李培坤ꎬ朱才华ꎬ等.基于不同时间粒度的城市轨道交通短时客流预测[J].长安大学学报(自然科学版)ꎬ2020ꎬ40(3):75 ̄83.DOI:10.19721/j.cnki.1671 ̄8879.2020.03.008.[9]何九冉ꎬ四兵锋.EMD ̄RBF组合模型在城市轨道交通客流预测中的应用[J].铁道运输与经济ꎬ2014ꎬ36(10):87 ̄92.DOI:10.3969/j.issn.1003 ̄1421.2014.10.017.[10]朱才华ꎬ孙晓黎ꎬ李培坤ꎬ等.融合车站分类和数据降噪的城市轨道交通短时客流预测[J].铁道科学与工程学报ꎬ2022ꎬ19(8):2182 ̄2192.DOI:10.19713/j.cnki.43 ̄1423/u.t20211018.[11]谢鑫鑫.基于EMD ̄KNN的城市轨道站点客流预测方法研究[D].苏州:苏州科技大学ꎬ2021.[12]张惠臻ꎬ高正凯ꎬ李建强ꎬ等.基于循环神经网络的城市轨道交通短时客流预测[J].吉林大学学报(工学版)ꎬ2023ꎬ53(2):430 ̄438.DOI:10.13229/j.cnki.jdxbgxb.20210720.(上接第66页)[10]LIUFꎬWANGMꎬZHANGMꎬetal.Intelligentmicroseismiceventsrecognitioninfiber ̄opticmicroseismicmonitoringsystemcomparedwithelectronicone[J].IEEEPhotonicsJournalꎬ2022ꎬ14(2):1 ̄5.DOI:10.1109/JPHOT.2022.3148315. [11]GEIGERL.Probabilitymethodforthedeterminationofearthquakeepicentersfromarrivaltimeonly[J].Bull.St.Louis.Univ.ꎬ1912ꎬ8:60-71.[12]张志斌ꎬ金花ꎬ王晓飞.单纯形定位方法在新疆数字地震台网的测定精度分析[J].震灾防御技术ꎬ2019ꎬ14(1):200 ̄209.[13]李健ꎬ高永涛ꎬ谢玉玲ꎬ等.基于无需测速的单纯形法微地震定位改进研究[J].岩石力学与工程学报ꎬ2014ꎬ33(7):1336 ̄1346.DOI:10.13722/j.cnki.jrme.2014.07.005.[14]李楠ꎬ王恩元ꎬ孙珍玉ꎬ等.基于L1范数统计的单纯形微震震源定位方法[J].煤炭学报ꎬ2014ꎬ39(12):8.[15]PRVGGERAFꎬGENDZWILLDJ.Microearthquakelocation:anonlinearapproachthatmakesuseofasimplexsteppingprocedure[J].BulletinoftheSeismologicalSocietyofAmericaꎬ1998ꎬ78(2):799 ̄815.DOI:10.1007/BF00819555. [16]PRUGGERAFꎬGENDZWILLDJ.Microearthquakelocation:Anonlinearapproachthatmakesuseofasimplexsteppingprocedure[J].BulletinoftheSeismologicalSocietyofAmericaꎬ1988ꎬ78(2):799 ̄815.DOI:10.1785/bssa0780020799.。
目前查找国内学位论文可通过以下几个数据库

一、目前查找国内学位论文可通过以下几个数据库:1.清华同方中国优秀博硕士学位论文全文数据库(已经订购,文摘,全文)是目前国内相关资源最完备、收录质量最高、连续动态更新的中国博硕士学位论文全文数据库,每年收录全国300家博士培养单位的优秀博/硕士学位论文约28000篇。
收录范围包括理工A(数理化天地生)、理工B(化学化工能源与材料)、理工C(工业技术)、农业、医药卫生、文史哲、经济政治与法律、教育与社会科学、电子技术与信息科学。
2.中国科学院学位论文数据库(免费文摘)/index_others.jsp?subjectselect=ETD3.国家科技图书文献中心的中文学位论文数据库(免费文摘)/index.html主要收录了1984年至今我国高等院校、研究生院及研究院所发布的硕士、博士和博士后的论文。
学科范围涉及自然科学各专业领域,并兼顾社会科学和人文科学,每年增加论文6万余篇。
每季更新。
4.CALIS高校学位论文库(免费文摘)/ipvalidator.do 目前有大约25万条学位论文文摘索引。
已有约80家大学签订了参加项目建设的协议,有70多家建立了本地学位论文提交和发布系统。
5.中国科技信息所万方数据集团的中国学位论文全文库(免费文摘,未订购)/所收论文主要是科技理工类硕士论文较多,也有经济、法律、和外语、艺术类等论文,但数量较少。
因他的数据来源是国务院学位办规定收缴的理工类全国硕士论文,收藏的数量有40多万篇。
6.国家图书馆学位论文(免费文摘目录)/index_lw.jsp?channelid=75008 国家图书馆学位论文收藏中心是国务院学位委员会指定的全国惟一负责全面收藏和整理我国学位论文的专门机构;也是人事部专家司确定的惟一负责全面入藏博士后研究报告的专门机构。
20多年来,国家图书馆收藏博士论文近12万种。
此外,该中心还收藏部分院校的硕士学位论文,台湾博士学位论文和部分海外华人华侨学位论文。
12个抗稻瘟病基因在山东省地方水稻品种中的分布

㊀山东农业科学㊀2023ꎬ55(10):7~14ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2023.10.002收稿日期:2022-12-13基金项目:济宁市重点研发计划项目(2021NYNS016ꎬ2020NYNS004)ꎻ国家水稻产业技术体系济宁试验站项目(CARS-01-080)ꎻ山东省重点研发计划(乡村振兴科技创新提振行动计划)项目(2022TZXD040)ꎻ山东省农业良种工程项目(2019LZGC003)ꎻ山东省农业科学院农业科技创新工程项目(CXGC2023A04)作者简介:房文文(1988 )ꎬ女ꎬ博士ꎬ助理研究员ꎬ主要研究方向为水稻抗病育种ꎮE-mail:fwenwen1988@163.com通信作者:张士永(1968 )ꎬ男ꎬ研究员ꎬ主要从事水稻育种研究ꎮE-mail:133****1992@163.com12个抗稻瘟病基因在山东省地方水稻品种中的分布房文文ꎬ王海凤ꎬ郭涛ꎬ姜艳芳ꎬ薛芳ꎬ张焕霞ꎬ张士永(山东省农业科学院湿地农业与生态研究所/山东省水稻工程技术研究中心ꎬ山东济南㊀250100)㊀㊀摘要:为进一步明确山东省地方水稻品种中抗稻瘟病基因(以下简称抗瘟基因)的分布情况ꎬ本研究利用Pita㊁Pita2㊁Pia㊁Pi9㊁Pigm㊁Pikm㊁Pi54㊁Pi5㊁Pii㊁Pib㊁Pish和Pi112个抗瘟基因的功能标记ꎬ对78个山东省地方水稻品种进行了分子检测ꎮ结果表明ꎬPi54和Pia在供试品种中的分布频率高达67.95%和50.00%ꎻ其次是Pi5㊁Pikm㊁Pib㊁Pii和Pi1ꎬ分布频率分别为29.49%㊁21.79%㊁20.51%㊁20.51%和16.67%ꎻ分布频率较低的是Pi9㊁Pish和Pita2ꎬ分布频率均为1.28%ꎻ供试品种中都未检出Pita和Pigmꎮ在供试品种中ꎬ检出2个抗瘟基因的品种数量最多ꎬ占供试品种总数的34.62%ꎻ检出1个抗瘟基因的品种数量次之ꎬ占24.36%ꎻ检出3㊁4㊁5㊁6㊁7个抗瘟基因的品种分别有13㊁7㊁4㊁1㊁2个ꎬ另有5个品种未检出任意一个抗瘟基因ꎮ在遗传相似系数为0.80时ꎬ可将78个品种聚为4类ꎬ其中70个品种聚为一类ꎮ本研究初步明确了78个山东省水稻品种的抗稻瘟病基因分布情况ꎬ可为进一步利用地方品种中的稻瘟病抗性基因培育广谱抗稻瘟病新品种提供科学依据ꎮ关键词:水稻ꎻ山东省地方品种ꎻ抗稻瘟病基因ꎻ分子标记中图分类号:S511.034ꎻQ781㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2023)10-0007-08DistributionofTwelveRiceBlastResistanceGenesinLocalRiceVarietiesinShandongProvinceFangWenwenꎬWangHaifengꎬGuoTaoꎬJiangYanfangꎬXueFangꎬZhangHuanxiaꎬZhangShiyong(ShandongInstituteofWetlandAgricultureandEcologyꎬShandongAcademyofAgriculturalSciences/ShandongRiceEngineeringTechnologyResearchCenterꎬJinan250100ꎬChina)Abstract㊀InordertoclarifythedistributionofriceblastresistancegenesinlocalricevarietiesinShan ̄dongProvinceꎬ78localricevarietiesweredetectedwiththemolecularfunctionalmarkersofPitaꎬPita2ꎬPiaꎬPi9ꎬPigmꎬPikmꎬPi54ꎬPi5ꎬPiiꎬPibꎬPishandPi1genes.TheresultsshowedthatthedistributionfrequencyofthetworesistancegenesPi54andPiawerehigheras67.95%and50.00%ꎬfollowedbyPi5ꎬPikmꎬPibꎬPiiandPi1withthedistributionfrequencyas29.49%ꎬ21.79%ꎬ20.51%ꎬ20.51%and16.67%ꎬrespectivelyꎻPi9ꎬPishandPita2hadthelowestdistributionfrequencyof1.28%.Allthesevarietiesdidn tcontainPitaandPigm.Thenumberoflocalvarietieswith2blastresistancegeneswasthemostꎬwhichac ̄countedfor34.62%ꎻthenumberoflocalvarietieswithoneblastresistancegenewassecondandaccountedfor24.36%ꎬwhilethatwiththreeꎬfourꎬfiveꎬsixandsevenblastresistancegeneswere13ꎬ7ꎬ4ꎬ1and2ꎻfivevarietieshadnoblastresistancegenes.The78localvarietiescouldbeclusteredintofourclassesatthegeneticsimilaritycoefficientof0.80ꎬamongwhichꎬ70varietieswereclusteredintooneclass.Inthisstudyꎬthedistri ̄butionofblastresistancegenesinthe78localricevarietiesofShandongProvincewerepreliminarilyidenti ̄fiedꎬwhichcouldprovidescientificreferencesforbreedingnewvarietieswithbroad ̄spectrumblastresistancebyfurtherutilizingtheblastresistancegenesinlocalricevarieties.Keywords㊀RiceꎻLocalvarietiesofShandongProvinceꎻRiceblastresistancegenesꎻMolecularmarker㊀㊀水稻是世界上的主粮作物之一ꎬ近一半的人口以稻米为主食[1-2]ꎮ稻瘟病是水稻生产上最重要的病害ꎬ严重影响水稻产量和品质[3-4]ꎮ山东省一般5年左右会暴发一次ꎬ发病田块一般减产10%~30%ꎬ严重的甚至超过50%[5]ꎮ山东属黄淮海稻区ꎬ以粳稻为主ꎬ年均种植面积达13万公顷[6]ꎮ稻瘟病的发病条件为气温26~28ħ㊁相对湿度90%以上ꎬ临沂㊁日照等稻区水稻抽穗灌浆期的气候特点符合其发生流行的条件ꎬ是山东省稻瘟病发生较重的地区[7]ꎮ目前ꎬ选育和合理种植抗病品种是控制稻瘟病流行最经济有效的途径[8-9]ꎮ由于田间稻瘟病菌小种类型多样ꎬ易发生变异ꎬ一些抗病水稻品种连续种植3~5年后抗性就会丧失ꎬ田间表现为感病ꎬ从而造成减产[10]ꎮ随着水稻基因组学和测序技术的迅速发展ꎬ越来越多的抗稻瘟病基因被定位与克隆ꎬ目前已被定位的抗瘟基因有100多个ꎬ已克隆的抗瘟基因超过30个[11]ꎮ抗瘟基因多为显性基因ꎬ其作用方式多为通过与核苷酸结合位点-富含亮氨酸重复序列蛋白(NBS-LRR)结合ꎬ从而使病原菌失去侵染性[12-13]ꎮ近年来ꎬ抗瘟基因分子标记的开发ꎬ主要为SSR㊁Indel㊁SNP标记ꎬ为高效检测抗瘟基因奠定了基础[14-16]ꎮ张亚玲等利用35个抗瘟基因标记检测了黑龙江省50个水稻主栽品种ꎬ发现抗瘟基因Pish㊁Pi36㊁Pi33和Pi-CO39在供试品种中的出现频率为100%ꎬPi63㊁Ptr㊁Pi37㊁Pi64㊁pi21㊁Pi9㊁Pi54㊁Pia㊁Pikp㊁Pi35㊁Pikm和Pik的出现频率为50%~100%ꎬPita㊁Pib㊁Pii㊁Pi5㊁Piz-t㊁Pi50和Pi2的出现频率为10%~50%ꎬPid2仅在2个品种中被检测到ꎬPigm仅在1个品种中被检测到ꎬ而Pit㊁Pid3㊁Bsr-d1㊁Pi25㊁Pid3-A4㊁Pi56㊁Pi1㊁Pike和Pb1在供试品种中未检测到[17]ꎻ杨林浩等利用一套稻瘟病菌标准菌株和特异性分子标记相结合ꎬ检测了吉林省68个主栽水稻品种ꎬ发现Pi ̄ta㊁Pia㊁Pish㊁Pita2㊁Pib和Pi9为吉林省主栽品种中出现频率较高的抗瘟基因ꎬ推测所有品种可能不含有Pi2㊁Pi54㊁Pikm㊁Pi11和Pi12基因[18]ꎮ山东省作为重要的粳稻产区ꎬ有着丰富的地方品种资源ꎬ但有关这些品种资源的抗瘟基因分布未见报道ꎮ本研究利用Pita㊁Pita2㊁Pia㊁Pi9㊁Pigm㊁Pikm㊁Pi54㊁Pi5㊁Pii㊁Pib㊁Pish㊁Pi1共12个抗瘟基因的分子标记对78个山东省地方品种进行抗瘟基因检测ꎬ以初步明确这些品种的抗瘟基因组成ꎬ为今后进一步挖掘和利用抗瘟基因㊁培育持久广谱抗瘟品种提供基因信息和数据支持ꎮ1㊀材料与方法1.1㊀试验材料供试材料为78个山东省地方水稻品种ꎬ见表1ꎬ均由山东省农业科学院湿地农业与生态研究所种质资源团队收集和保存ꎬ均以常规方法栽培ꎮ试剂及仪器:十六烷基三甲基溴化铵(CTAB)ꎬ购于生工生物工程(上海)股份有限公司ꎻPCR试剂ꎬ北京全式金生物技术(TransGenBiotech)有限公司产品ꎻEvaGreenqPCR核酸染料ꎬ美国Biotium公司产品ꎮSPX智能型生化培养箱㊁RXZ-500C-LED人工气候培养箱ꎬ宁波江南仪器厂产品ꎻBCD-290W型立式冰箱ꎬ青岛海尔股份有限公司产品ꎻ罗氏LightCycler96实时荧光定量PCR仪ꎬRoche公司产品ꎮ1.2㊀水稻DNA提取剪取3段供试水稻品种的叶片(大小为2mmˑ2mm)于2mL离心管中ꎬ采用CTAB法提取DNAꎮ具体步骤如下:(1)向装有供试水稻叶片的离心管中加600μL65ħ预热的2ˑCTABꎬ然后65ħ温浴30minꎻ(2)冷却后加600μL氯仿-异戊醇溶液(氯仿和异戊醇体积比为1ʒ1)混匀ꎬ12000r/min离心10minꎬ吸200μL上清液ꎬ加入等体积的异丙醇ꎬ轻轻颠倒混匀后于-20ħ条件下沉淀30minꎻ(3)12000r/min㊁4ħ条件下离心15minꎬ弃上清液ꎬ加入600μL体积百分含量为70%的乙醇洗涤沉淀1~2次ꎬ放入通风橱内吹干ꎻ(4)加入100μLTE(10mmol/LTris-HClꎬ0.1mmol/LEDTAꎬpH8.0)溶解DNAꎬ使用核酸定量仪测量DNA浓度ꎬ并将浓度调整为约10ng/μLꎬ8㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀㊀㊀表1㊀供试山东省地方水稻品种编号品种名编号品种名编号品种名编号品种名编号品种名编号品种名1小黄稻14塘稻27秋旱稻40北圆1号-153香稻66大红芒-22竹秆青-115旱糯稻28高秸水旱稻-141北圆1号-254水稻67佃户屯乡11号3大青秸-116汪稻29高秸水旱稻-242北圆1号55郯城野生稻68石槐4大青秸-217水牛皮旱稻-130紫皮水旱稻43清水稻56红皮高丽旱稻-169糯米-15大青秸-318水牛皮旱稻-231白芒子44明水香稻57红皮高丽旱稻-270糯米-26竹秆青-219木枪秆稻子32小白屯45旱稻58水旱稻-171小壳旱稻7小红芒20旱稻白芒33香粘稻-146老旱稻-259水旱稻-272旱粘稻-18旱稻子21明水米旱稻-134香粘稻-247白砂米60白米稻73旱粘稻-29紫皮旱稻22明水米旱稻-235香粘稻-348铁耙子61粘稻74白香糯10秋白水稻23粘旱稻-136青旱稻49八月芒62秃头红米稻75紫秆旱稻11蒋庄1号24粘旱稻-237蓬莱稻-150乐陵旱稻63短顶芒白米76紫皮旱稻-112大长芒旱稻25圆粒水稻38蓬莱稻-251葫芦香稻64水斗扬77紫皮旱稻-213紫颖稻26半芒水稻39蓬莱稻-352小雪稻65大红芒-178范李庄1号-20ħ保存待测DNAꎮ1.3㊀抗瘟基因分子标记本研究选用抗瘟基因Pita㊁Pi9㊁Pikm㊁Pigm㊁Pia㊁Pi54㊁Pish㊁Pii㊁Pi1㊁Pib㊁Pita2㊁Pi5的分子标记进行试验ꎬ其PCR特异性引物信息见表2ꎮ所有引物均由华大基因有限公司合成ꎮ㊀㊀表2㊀抗瘟基因的特异性引物引物名称引物序列(5ᶄ-3ᶄ)退火温度/ħPita-FCTTCTTTCTTTCTCTGCCGTGG57Pita-RCATCAAGTCAGGTTGAAGATGPi9-FTGATTATGTTTTTTATGTGGGG57Pi9-RATTAGTGAGATCCATTGTTCCPikm-E-FCAATGCCAGCACTCGAAATCAT57Pikm-E-RCAGTGACGATGCCATCAACAAAPikm-I-FTGAAAATCTCCGTGAGGTGCAT59Pikm-I-RTTGGTTATTGCTTCTGCCCCATPigm-F2CGAACTAATAACTCCAAACGTA60Pigm-R1GGAGGTGCTAGAGTTTGTCGCCPia-FAGATAGTTGTTAAGGTGCACA60Pia-RCGTTCACTGACGCAGCCAGCGCPi54-FCAATCTCCAAAGTTTTCAGG60Pi54-RGCTTCAATCACTGCTAGACCPish-FTCTAGACGAGATGTACTTCC58Pish-RTACATTTTCAAGATTAACGAPii-FAGAAAGATCCAGTGCTTCTG58Pii-RCAATCTCAGCAGGTTCCTCAPi1-FGTGCTGCTGTGGCTAGTTTG58Pi1-RAGTCCCCGCTCAATTTTTCTPib-F4CGTGATGCGTGGACTTTCGC55Pib-R7TAACTCCAAAGGAGCTCAGGPita2-FCAGCGAACTCCTTCGCATACGCA60Pita2-RCGAAAGGTGTATGCACTATAGTATCCPi5-09RL09F2TCTGGTGAAAACTCTGAACG56Pi5-09RL09R2CGGTGAGTTTAGCGAAGACC1.4㊀PCR扩增荧光PCR反应体系:10ˑTaqbuffer(含20mmol/LMg2+)1μLꎬ250μmol/LdNTP0.2μLꎬ0.5UTaqDNAPolymerase0.1μLꎬ20ˑEvagreen0.125μLꎬ10ng/μLDNA模板3μLꎬ0.1mmol/L正㊁反向引物各0.2μLꎬ用ddH2O定容至10μLꎮ荧光PCR扩增程序:94ħ预变性2minꎻ94ħ变性10sꎬ60ħ退火30sꎬ40个循环ꎮ普通PCR反应体系:2ˑESTaqMasterMix(Dye)7.5μLꎬ水稻DNA3.0μLꎬ正㊁反向引物各0.5μLꎬ3.5μLddH2OꎮPCR反应程序:95ħ预变性5minꎻ95ħ变性30sꎬ55~60ħ退火30s(根据梯度PCR的结果选择最佳退火温度)ꎬ72ħ延伸0.5~1.5min(由扩增产物的长度决定)ꎬ32次循环ꎻ72ħ延伸10minꎮ1.5㊀琼脂糖凝胶电泳检测抗瘟基因Pish㊁Pii㊁Pib㊁Pita2㊁Pi5的扩增产物用1%琼脂糖凝胶电泳检测ꎬ150V稳压电泳30minꎬ然后在凝胶成像系统中拍照ꎮ1.6㊀HRM检测抗瘟基因Pita㊁Pi9㊁Pikm㊁Pigm㊁Pia㊁Pi54㊁Pi1的扩增产物经熔解曲线(HRM)检测ꎮ分析程序:95ħ变性15sꎻ60ħ退火15sꎻ以0.07ħ/s的速率升温ꎬ10reading/ħ采集荧光信号ꎬ至90ħ保持1sꎬ采用LightCycler96software软件进行熔解曲线分析ꎮ1.7㊀数据处理与统计分析采用MicrosoftExcel对数据进行整理和作图ꎮ采用NTSYS2.10软件计算78个品种间的遗9㊀第10期㊀㊀㊀㊀㊀房文文ꎬ等:12个抗稻瘟病基因在山东省地方水稻品种中的分布传相似系数ꎬ然后利用非加权类平均法(UPGMA)对遗传相似系数进行聚类分析ꎬ得到聚类树形图ꎮ2㊀结果与分析2.1㊀稻瘟病抗性基因的分子标记检测及在品种中的分布本研究利用Pita㊁Pita2㊁Pia㊁Pi9㊁Pigm㊁Pikm㊁Pi54㊁Pi5㊁Pii㊁Pib㊁Pish和Pi1的功能标记对78个山东省地方水稻品种进行分子检测ꎬ结果(表3)发现ꎬ有53个品种含有Pi54ꎬ有39个品种含有Piaꎬ有23个品种含有Pi5ꎬ有17个品种含有Pikmꎬ有16个品种含有Pib和Piiꎬ有13个品种含有Pi1ꎬ有1个品种含有Pi9㊁Pish和Pita2ꎬ所有品种中均未检测到Pita和Pigmꎮ部分基因分子标记的检测结果如图1㊁图2所示ꎮPi54和Pia在供试品种中的分布频率最高ꎬ分别高达67.95%和50.0%ꎻ其次是Pi5㊁Pikm㊁Pib㊁Pii和Pi1ꎬ分布频率分别为29.49%㊁21.79%㊁20.51%㊁20.51%和16.67%ꎻ分布频率较低的是Pi9㊁Pish和Pita2ꎬ均为1.28%ꎻ检测品种中都不含有抗性基因Pita和Pigm(表3㊁图3)ꎮA为Pi1基因分型结果ꎬ红色曲线代表携带Pi1基因纯合型ꎬ蓝色曲线代表不含Pi1基因的野生型ꎻB为Pi9基因分型结果ꎬ红色曲线代表携带Pi9基因的纯合型ꎬ蓝色曲线代表不含Pi9基因或含有其他等位基因的野生型ꎻC为Pia基因分型结果ꎬ红色曲线代表携带Pia基因的纯合型ꎬ蓝色曲线代表不含Pia基因的野生型ꎮ图1㊀高分辨率熔解曲线检测地方水稻品种的抗瘟基因01㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀A为Pi54基因检测结果ꎬ216bp(抗病基因)/359bp(感病基因)ꎻB为Pib基因(915bp)检测结果ꎻC为Pii基因(1220bp)检测结果ꎮ泳道25~36依次是圆粒水稻㊁半芒水稻㊁秋旱稻㊁高秸水旱稻-1㊁高秸水旱稻-2㊁紫皮水旱稻㊁白芒子㊁小白屯㊁香粘稻-1㊁香粘稻-2㊁香粘稻-3㊁青旱稻ꎮ图2㊀琼脂糖凝胶电泳检测地方水稻品种的抗瘟基因Pi54㊁Pib㊁Pii㊀㊀表3㊀山东省地方水稻品种抗瘟基因检测结果编号品种名抗瘟基因PiaPi9PikmPi54Pi5PiiPibPishPita2Pi1PitaPigm1小黄稻----+-------2竹秆青-1+--+--------3大青秸-1------------4大青秸-2+--+--------5大青秸-3--+---------6竹秆青-2+--+--------7小红芒---+--------8旱稻子+-----------9紫皮旱稻+--+++------10秋白水稻---+--------11蒋庄1号---+--------12大长芒旱稻---++-------13紫颖稻--+---------14塘稻---++-------15旱糯稻+--+--------16汪稻------------17水牛皮旱稻-1------------18水牛皮旱稻-2+-++--+-----19木枪秆稻子---+--------20旱稻白芒---+--------21明水米旱稻-1+--+--------22明水米旱稻-2---+--------23粘旱稻-1---+++------24粘旱稻-2+-+---------25圆粒水稻+--+--------26半芒水稻---++-------27秋旱稻---+++------28高秸水旱稻-1------+-----29高秸水旱稻-2--+++-------30紫皮水旱稻---++++-----31白芒子--+---------11㊀第10期㊀㊀㊀㊀㊀房文文ꎬ等:12个抗稻瘟病基因在山东省地方水稻品种中的分布㊀㊀表3(续)编号品种名抗瘟基因PiaPi9PikmPi54Pi5PiiPibPishPita2Pi1PitaPigm32小白屯+--+--------33香粘稻-1---+--------34香粘稻-2+-----------35香粘稻-3+--+--------36青旱稻+-++--+-----37蓬莱稻-1+---++---+--38蓬莱稻-2+-----------39蓬莱稻-3+--+--+-----40北圆1号-1+--+--------41北圆1号-2---------+--42北圆1号+--+--------43清水稻+--+--------44明水香稻---+----+---45旱稻----++------46老旱稻-2------------47白砂米---++-------48铁耙子+--++++-----49八月芒+--++++-----50乐陵旱稻+--++++-----51葫芦香稻+-++--+-----52小雪稻---+-----+--53香稻+-----+-----54水稻+-++--------55郯城野生稻--+-++------56红皮高丽旱稻-1+-+++++--+--57红皮高丽旱稻-2---+-----+--58水旱稻-1---+-----+--59水旱稻-2+-++-----+--60白米稻+--+--------61粘稻+-+++++--+--62秃头红米稻+--+++++----63短顶芒白米+--++++-----64水斗扬---+-----+--65大红芒-1+-++--------66大红芒-2+-++--------67佃户屯乡11号---+++------68石槐+--+--------69糯米-1---+--------70糯米-2---------+--71小壳旱稻+-++--------72旱粘稻-1+-----+--+--73旱粘稻-2------------74白香糯+-----+--+--75紫秆旱稻--++--------76紫皮旱稻-1----++------77紫皮旱稻-2++--+-------78范李庄1号---------+--㊀㊀注:分子标记检测结果中 + 代表含有抗病基因ꎬ - 代表不含有抗病基因或含有其等位基因ꎮ21㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀图3㊀12个抗稻瘟病基因在山东省地方㊀㊀水稻品种中的分布频率2.2㊀不同地方水稻品种含抗瘟基因的数量分析由表3和图4可知ꎬ竹秆青-1㊁竹秆青-2㊁塘稻㊁明水米旱稻-1㊁明水香稻㊁紫秆旱稻等27个品种检测出2个抗瘟基因ꎬ占检测品种总数的34.62%ꎬ其中ꎬ有23个品种检出Pi54ꎬ有15个品种检出了Piaꎮ小黄稻㊁大青秸-3㊁小红芒㊁旱稻子㊁秋白水稻㊁蒋庄1号等19个品种检测出1个抗瘟基因ꎬ占检测品种总数的24.36%ꎬ其中ꎬ有8个品种检测出Pi54ꎮ粘旱稻-1㊁秋旱稻㊁高秸水旱稻-2㊁蓬莱稻-3㊁香稻㊁郯城野生稻等13个品种检测出3个抗瘟基因ꎬ占检测品种总数的16.67%ꎮ另外ꎬ紫皮旱稻㊁水牛皮旱稻-1㊁青旱稻㊁蓬莱稻-1㊁葫芦香稻等7个品种检测出4个抗瘟基因ꎬ铁耙子㊁八月芒㊁乐陵旱稻㊁短顶芒白米4个品种检测出5个抗瘟基因ꎬ仅秃头红米稻检测出6个抗瘟基因ꎬ红皮高丽旱稻-1㊁粘稻2个品种检测出7个抗瘟基因ꎮ而大青秸-1㊁汪稻㊁水牛皮旱稻-1㊁老旱稻-2㊁旱粘稻-2共5个品种未检测到12个抗瘟基因中的任何一个ꎮ图4㊀山东省地方水稻品种含抗瘟基因数统计2.3㊀山东省地方水稻品种的聚类分析基于遗传相似系数ꎬ利用UPGMA法对78个山东省地方水稻品种进行聚类分析ꎬ结果(图5)显示ꎬ在遗传相似系数0.80处ꎬ可将78个供试材料分为四类ꎮ第一类包括大青秸-1㊁汪稻及水牛皮旱稻-1共3个品种ꎻ第二类仅有高秸水旱稻-1ꎻ第三类包括大青秸-3㊁紫颖稻㊁粘旱稻-2及白芒子共4个品种ꎻ其余70个品种均聚在第四类中ꎮ图5㊀78个山东省地方水稻品种的聚类分析结果3㊀讨论与结论水稻与稻瘟病菌存在协同进化关系ꎬ一个抗病新品种在生产上使用几年后ꎬ由于稻瘟菌变异而突破水稻的防御机制ꎬ使抗病品种的抗病能力下降甚至丧失ꎬ变为感病品种ꎮ因此ꎬ在改良水稻品种的稻瘟病抗性时ꎬ要首先弄清品种中的抗瘟31㊀第10期㊀㊀㊀㊀㊀房文文ꎬ等:12个抗稻瘟病基因在山东省地方水稻品种中的分布基因(组合)ꎬ从而更好地利用不同的抗瘟基因组合对品种进行改良ꎮ有关我国现有水稻品种中抗瘟基因的分布情况ꎬ前人已进行过一些研究ꎬ如岂长燕等研究了抗瘟基因Pib㊁Pita㊁Pi5㊁Pi25和Pi54在我国124份水稻微核心种质中的分布[19]ꎻ范方军等研究了Pib㊁Pita㊁Pi54和Pikm在江苏省迟熟中粳稻预试64份品系中的分布[20]ꎮ本研究利用12个抗瘟基因(Pita㊁Pita2㊁Pia㊁Pi9㊁Pigm㊁Pikm㊁Pi54㊁Pi5㊁Pii㊁Pib㊁Pish㊁Pi1)的分子功能标记检测了78个山东地方水稻品种的抗瘟基因分布情况ꎬ结果表明ꎬ除Pita㊁Pigm未检测到外ꎬ其余10个基因均能在山东省地方水稻品种中检测到ꎮ其中ꎬPi54和Pia在供试品种中的分布频率较高ꎬ达67.95%和50.0%ꎻ其次是Pi5㊁Pikm㊁Pib㊁Pii和Pi1ꎬ分布频率分别为29.49%㊁21.79%㊁20.51%㊁20.51%和16.67%ꎻPi9㊁Pish㊁Pita2的分布频率较低ꎬ仅为1.28%ꎮ各品种含有抗瘟基因的数量为1~7个不等ꎬ抗瘟基因组合种类丰富ꎮ目前ꎬ培育持久广谱抗病品种和合理布局抗病品种仍是控制稻瘟病最经济有效的途径ꎬ而挖掘种质资源中的抗瘟基因是重要基础ꎮ本研究结合山东省稻瘟病菌群体生理小种的变异动态ꎬ挑选出可进一步利用的抗瘟基因Pi54㊁Pia㊁Pi5㊁Pikm㊁Pib和Piiꎬ下一步可作为目的基因利用到主栽品种中ꎬ这为育种家培育抗稻瘟病新品种提供了数据支持ꎬ助力了培育持久和广谱抗稻瘟病水稻新品种ꎮ但需注意有些抗瘟基因与不良性状的 连锁累赘 效应ꎬ如Pigm与小粒性状连锁ꎬ以避免在提高抗病性的同时负向影响产量或品质ꎮ本研究仅检测了12个抗瘟基因在78个山东省地方水稻品种中的分布情况ꎬ但其他抗瘟基因的分布情况还有待进一步研究ꎻ另外ꎬ地方品种中抗瘟基因数量与水稻品种稻瘟病抗性频率间的关系也有待进一步研究ꎮ参㊀考㊀文㊀献:[1]㊀GlobalRiceSciencePartnership.RiceAlmanac[M].LosBañosꎬPhilippines:InternationalRiceResearchInstituteꎬ2013. [2]㊀FAOSTA.FAOStatisticalDatabases[DB/OL].Rome:FoodandAgricultureOrganization(FAO)oftheUnitedNations.2017.http://www.fao.org/statistics.[3]㊀KhanMAꎬBhuiyanMRꎬHossainMSꎬetal.Neckblastdis ̄easeinfluencesgrainyieldandqualitytraitsofaromaticrice[J].ComptesRendusBiologiesꎬ2014ꎬ337(11):635-641. [4]㊀孙国昌ꎬ杜新法.水稻稻瘟病防治研究进展和21世纪初研究设想[J].植物保护ꎬ2000ꎬ26(1):33-35. [5]㊀刘振林ꎬ杨军ꎬ金桂秀ꎬ等.山东水稻稻瘟病发病规律㊁特点及防治措施[J].安徽农业科学ꎬ2015ꎬ43(14):105-106. [6]㊀吴修ꎬ杨连群ꎬ陈峰ꎬ等.山东省水稻生产现状及发展对策[J].山东农业科学ꎬ2013ꎬ45(5):119-125. [7]㊀赵文生.水稻稻瘟病发病规律与防治措施[J].科技创新与应用ꎬ2012(1):187.[8]㊀朱有勇ꎬ陈海如ꎬ范静华ꎬ等.利用水稻品种多样性控制稻瘟病研究[J].中国农业科技ꎬ2003ꎬ36(5):521-527. [9]㊀贺雄ꎬ丁朝辉ꎬ胡立东ꎬ等.湖南48个水稻栽培品种抗瘟性评价及抗性基因鉴定[J].江苏农业科学ꎬ2020ꎬ48(1):108-113.[10]FangWWꎬLiuCCꎬZhangHWꎬetal.Selectionofdifferen ̄tialisolatesofMagnaportheoryzaeforpostulationofblastresist ̄ancegenes[J].Phytopathologyꎬ2018ꎬ108:878-884. [11]高清ꎬ张亚玲ꎬ葛欣ꎬ等.水稻抗稻瘟病基因研究进展[J].分子植物育种ꎬ2022ꎬ20(11):3634-3643.[12]ParkCHꎬChenSꎬShirsekarGꎬetal.TheMagnaportheoryzaeeffectorAvrPiz ̄ttargetstheRINGE3ubiquitinligaseAPIP6tosuppresspathogen ̄associatedmolecularpattern ̄trig ̄geredimmunityinrice[J].PlantCellꎬ2012ꎬ24(11):4748-4762.[13]TakahashiAꎬHayashiNꎬMiyaoAꎬetal.UniquefeaturesofthericeblastresistancePishlocusrevealedbylargescaleretro ̄transposon ̄tagging[J].BMCPlantBiol.ꎬ2010ꎬ10:175. [14]易怒安ꎬ李魏ꎬ戴良英.水稻抗稻瘟病基因的克隆及其分子育种研究进展[J].分子植物育种ꎬ2015ꎬ13(7):1653-1659.[15]BryanGTꎬWuKSꎬFarrallLꎬetal.AsingleaminoaciddifferencedistinguishesresistantandsusceptibleallelesofthericeblastresistancegenePi ̄ta[J].ThePlantCellꎬ2000ꎬ12(11):2033-2046.[16]XiaoGꎬYangJꎬZhuXꎬetal.Prevalenceofineffectivehaplo ̄typesatthericeblastresistance(R)genelociinChineseelitehybridricevarietiesrevealedbysequence ̄basedmoleculardi ̄agnosis[J].Riceꎬ2020ꎬ13(1):6.[17]张亚玲ꎬ高清ꎬ赵羽涵ꎬ等.黑龙江省水稻种质稻瘟病抗性评价及抗瘟基因结构分析[J].中国农业科学ꎬ2022ꎬ55(4):625-645.[18]杨林浩ꎬ王瀚ꎬ赵丹ꎬ等.吉林省主栽水稻品种抗瘟性评价及抗瘟基因型鉴定[J].植物病理学报ꎬ2022ꎬ52(3):404-415.[19]岂长燕ꎬ许兴涛ꎬ马建ꎬ等.抗稻瘟病基因Pib㊁Pita㊁Pi5㊁Pi25和Pi54在我国水稻微核心种质中的分布[J].植物遗传资源学报ꎬ2019ꎬ20(5):1240-1246ꎬ1254.[20]范方军ꎬ王芳权ꎬ刘永峰ꎬ等.Pi-b㊁Pi-ta㊁Pikm和Pi54对水稻穗颈瘟的抗性评价[J].华北农学报ꎬ2014ꎬ29(3):221-226.41㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀。
山东科技情报科学技术奖奖励办法

山东省科技情报科学技术奖奖励办法第一章总则第一条为推动我省科技情报事业的发展,贯彻尊重知识,尊重人才的方针,奖励我省从事科技信息工作的科研人员在科技信息研究领域所取得的优秀成果,依据《国家科学技术奖励条例》、《国家科学技术奖励条例实施细则》、国家和省《社会力量设立科学技术奖管理办法》的有关规定,特制定本办法。
第二条山东科技情报学会设立并承办“山东省科技情报科学技术奖”(以下简称“科学技术奖”)。
“科学技术奖”每2年评选一次,设一等奖、二等奖、三等奖,奖项总数不超过100项。
第三条凡申报“科学技术奖”的集体和个人,必须做到坚持以邓小平理论为指导,坚持为社会主义服务、为人民服务的方向,贯彻“三个代表”重要思想,运用马克思主义的立场、观点和方法进行科学研究。
第四条“科学技术奖”贯彻尊重知识、尊重人才的方针。
“科学技术奖”的推荐、评审和授奖工作,坚持“公开、公平、公正”的原则。
第二章奖励范围和条件与标准和奖励等级第五条范围和条件(一)科技情报的分析、研究与预测选题针对性强,能较全面、准确地反映国内外科学技术水平和经济建设或市场需求的发展趋势;有分析对比,有研究预测和建议,为领导部门决策或制定计划、编制规划,为制定有关政策提供重要依据;或对科研、设计、生产部门较有指导意义,经采纳后已取得明显的经济或社会效益。
(二)科技情报的搜集加工与传递报道善于掌握并利用国内外科技情报,搜集加工的科技情报信息量大,针对性强,经用户使用后已产生明显的经济或社会效益,或对技术进步起了重要作用。
科技情报(信息)刊物(包括检索报道刊物)传递报道方向明确,针对性强,并富有自己的特色,影响面较大,对提高技术水平或学术水平、成果推广以及改善经营管理和提高人员素质方面起到了促进作用,并取得了良好效果。
(三)科技情报服务(包括实物、声相、文献、咨询、展览、计算机检索等)经用户采纳,在新技术新产品开发、技术攻关、技术引进、成果推广、开拓技术市场或改善经营管理方面发挥了重要作用,效果显著。
研究项目和成果新颖性查证--科技查新

一直以来,科技查新工作由国家科技部(原国 家科委)主管,所以科技部(原国家科委)直 接授权认定的查新机构一般也被称为一级查新 机构或一级查新单位。 目前为止,全国共有38个科技信息机构获得了 一级查新单位资格。
卫生部授权的查新机构
至2001年,中华人民共和国卫生部授权认定的医药卫生类科技 查新机构共31个: 中国医学科学院医学信息研究所、中国预防医学科学院信息中 心(预防医学部分)、四川省医学情报研究所、广东省医学情报 研究所、天津市医学情报研究所、江苏省医学情报研究所、北 京市医学情报研究所、河北省医学情报研究所、湖北省医学情 报研究所、湖南省医学情报研究所、吉林省医学情报研究所、 上海市医学科技查新委员会、湖南医学大学图书馆、白求恩医 科大学图书馆、同济医科大学图书馆、中国医学科学院华西分 院医学情报研究所、西安医科大学图书馆、中国医科大学图书 馆、北京医科大学图书馆、中山医科大学图书馆、山东医科大 学图书馆、云南省医学信息研究所、海南省医学信息研究所、 浙江省医学情报研究所、黑龙江省医学文献信息中心、甘肃省 医学情报研究所、山东省医学情报研究所、河南省医学情报研 究所、重庆市医学情报研究所、深圳市医学情报所、福建省医 学情报所。
查新曾用定义
科技情报人员利用各种文献资源,通过各种情报 手段(手工检索和计算机检索等),对需查新的 科研成果(项目)广泛搜寻与之相关的文献资料, 运用综合分析和对比分析等方法,比较该成果 (项目)在所检相关文献中所处的特点、水平和 技术状态,以查新报告的形式对该成果(项目) 的新颖性进行综合客观的论证及评价的一种信息 咨询服务形式。
(查新机构)受理查新与订立合同
判断是否属于所承担的查新业务受理范围 可受理的查新委托: 经获准的专业范围内的查新业务 立项查新 研究、开发、转化、技术转移过程中查新 国家和地方规定要求的查新 其他 可拒绝的查新委托: 不能明确列示查新项目的查新点 不能出具与查新内容相关的技术资料
齐鲁工业大学山东省科学院20xx年新增硕士学位研究生指导教师
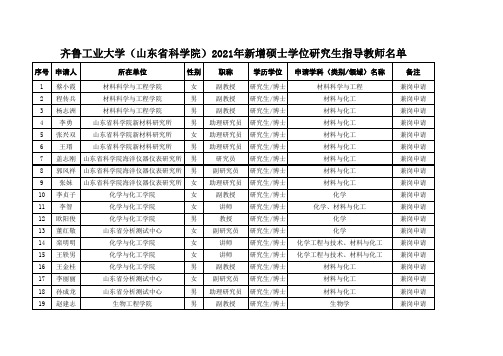
序号 申请人
所在单位
性别 职称
学历学位 申请学科(类别/领域)名称
80 王振 山东省科学院海洋仪器仪表研究所 男 副研究员 研究生/博士
81 刘建翔
山东省科学院自动化研究所
男
研究员 研究生/硕士
82 王学林
山东省科学院自动化研究所
男 副研究员 研究生/博士
83 胡桐 山东省科学院海洋仪器仪表研究所 男 副研究员 研究生/博士
46 王超
山东省科学院情报研究所
男 助理研究员 研究生/博士
47 姜树明
山东省科学院情报研究所
男 副研究馆员 研究生/硕士
48 刘用
山东省科学院情报研究所
男 高级经济师 研究生/博士
49 张玉瑾 电子信息工程学院(大学物理教学部) 女
副教授 研究生/博士
50 李奎龙 电子信息工程学院(大学物理教学部) 男
男 助理研究员 研究生/博士 材料科学与工程、材料与化工
女
副教授 研究生/博士
化学、材料与化工
男
讲师
研究生/博士
化学、材料与化工
男
副教授 研究生/博士
化学、材料与化工
男
讲师
研究生/博士
化学
女
讲师
研究生/博士
化学、材料与化工
男
教授
研究生/博士
化学
备注
兼岗申请 兼岗申请 兼岗申请 兼岗申请 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内 新增校内
化学
14 栾明明
化学与化工学院
女
讲师
研究生/博士 化学工程与技术、材料与化工
国内外网络信息检索资源汇总

NSTL(National Science Technology Library )是 中国目前最大的一个虚拟的科技文献信息服 务机构,它采集、收藏和开发理、工、农、 医等学科领域的科技文献资源,包括:中文 科技期刊约4000种,西文科技期刊11000多种, 西文会议文献3000多种,以及中文会议、中 文学位论文、国内外专利等文献类型。目前 报道的文摘数据总量已超过1100万条。 目前提供的服务项目包括:网上科技文献数 据库免费联机检索服务、面向注册用户的全 文提供、代查代借服务。
山东省科学技术情报研究所
CNKI《中国知识资源总库》 ——检索方法
专业检索用于图书情报专业人员查新、 信息分析等工作,使用逻辑运算符和关 键词构造检索式进行检索。 引文检索以检索参考文献为出发点,根 据文献的引用关系,找到引用文献。引 文数据库中的所有文献都与其它文献具 有引用或被引用的关系,引文检索是通 过这些关系检索到文献。
山东省科学技术情报研究所
Elsevier ScienceDirect——介绍
荷兰Elsevier公司是世界上著名学术期刊出版 公司,是全球最大的出版商。出版的期刊是 世界上公认的高品位学术期刊,大多数都是 核心期刊, 并且被世界上许多著名的二次文 献数据库所收录。 ScienceDirect是它的网络全文期刊数据库平台 系统。该库提供98年以来Elsevier公司收录 1800余种电子期刊(其中1500余种为全文, 200余种为文摘。网站免费提供全科技领域 5900万余篇文献题录检索。
山东省科学技术情报研究所
中国专利信息网
1985年以来的中国专利文摘、全文数据 库 免费用户:免费注册。可检索中国专利 文摘 高级用户:收费。可检索中国专利文摘、 全文数据库 特点:检索方便、灵活,有二次检索功 能。
陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析

㊀山东农业科学㊀2023ꎬ55(4):11~23ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2023.04.002收稿日期:2022-07-19基金项目:山东省农业良种工程项目(2020LZGC002)ꎻ国家自然科学基金项目(31901422)ꎻ山东省农业科学院农业科技创新工程项目(CXGC2021A46)作者简介:陈义珍(1990 )ꎬ女ꎬ助理研究员ꎬ主要从事棉花抗衰老及遗传育种研究ꎮE-mail:chenyizhen0@126.com通信作者:柳展基(1972 )ꎬ男ꎬ研究员ꎬ主要从事棉花遗传育种研究ꎮE-mail:scrcliuzhanji@sina.com陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析陈义珍ꎬ李浩ꎬ傅明川ꎬ王立国ꎬ刘任重ꎬ柳展基(农业部黄淮海棉花遗传改良与栽培生理重点实验室/山东省农业科学院经济作物研究所ꎬ山东济南㊀250100)㊀㊀摘要:膜结合脂肪酸去饱和酶(fattyaciddesaturaseꎬFAD)是生物合成不饱和脂肪酸的关键酶ꎮ本研究以陆地棉基因组数据为基础ꎬ利用生物信息学方法对棉花FAD基因家族进行全基因组鉴定和进化分析ꎮ结果表明ꎬ陆地棉基因组中共含有37个FAD基因ꎬ进化分析发现这些基因分为4个亚家族ꎬ各亚家族成员数量不一ꎬ但相同亚家族成员含有相似的基因结构和保守基序ꎮ共线性分析发现28对复制基因全为片段复制基因ꎬ且都经历了严格的纯化选择作用ꎮ此外ꎬ在陆地棉FAD基因的启动子区域ꎬ发现了丰富的响应植物激素信号和逆境胁迫的顺式作用元件ꎮ转录组分析表明ꎬ陆地棉FAD基因家族响应干旱和盐胁迫ꎬ定量PCR分析表明施加外源褪黑素显著影响了FAD基因的表达ꎮ本研究结果为进一步解析FAD家族基因的功能奠定了基础ꎮ关键词:陆地棉ꎻ膜结合脂肪酸去饱和酶ꎻ全基因组鉴定ꎻ生物信息学分析ꎻ基因表达ꎻ干旱和盐胁迫ꎻ褪黑素中图分类号:S562:Q78㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2023)04-0011-13Genome ̄WideIdentificationandExpressionAnalysisofMembrane ̄BoundFattyAcidDesaturaseGeneFamilyinUplandCotton(GossypiumhirsutumL.)ChenYizhenꎬLiHaoꎬFuMingchuanꎬWangLiguoꎬLiuRenzhongꎬLiuZhanji(KeyLaboratoryofCottonBreedingandCultivationinHuang ̄Huai ̄HaiPlainꎬMinistryofAgricultureandRuralAffairs/InstituteofIndustrialcropsꎬShandongAcademyofAgriculturalSciencesꎬJinan250100ꎬChina)Abstract㊀Membrane ̄boundfattyaciddesaturases(FADs)arethekeyenzymesforthebiosynthesisofunsaturatedfattyacids.InthisstudyꎬthewholegenomeofFADgenefamilywasidentifiedanditsevolutionwasanalyzedbybioinformaticsmethodbasedonthegenomicdataofuplandcotton(GossypiumhirsutumL.).Theresultsshowedthattherewere37FADgenesintheuplandcottongenome.PhylogeneticanalysesindicatedtheseFADgenesweredividedintofoursubfamiliesꎬandeachcontaineddifferentnumbersofFADgenesꎬbutthemembersinthesamesubfamilycontainedsimilargenestructureandconservedmotifs.Collinearanalysisrevealedthatallthe28pairsofreplicatorswerefragmentreplicatorsandhadundergonerigorouspurificationandselection.Inadditionꎬabundantcis ̄actingelementsrelatedtophytohormonesignalingandstressresponseswereidentifiedinthepromoterregionsofFADgenes.TranscriptomeanalysisshowedthattheFADgenefamilyofuplandcottonrespondedtodroughtandsaltstressꎬandquantitativePCRanalysisshowedthatexogenousmelatoninsignificantlyaffectedtheFADgeneexpression.TheresultsofthisstudylaidafoundationforfurtherfunctionalanalysisofFADfamilygenes.Keywords㊀UplandcottonꎻMembrane ̄boundfattyaciddesaturaseꎻGenome ̄wideidentificationꎻBioin ̄formaticsanalysisꎻGeneexpressionꎻDroughtandsaltstressꎻMelatonin㊀㊀脂肪酸去饱和酶(fattyaciddesaturaseꎬFAD)能够在脂肪酸链的特定位置引入双键ꎬ是产生不饱和脂肪酸的关键酶[1ꎬ2]ꎮ植物中的不饱和脂肪酸主要包括油酸㊁亚油酸㊁亚麻酸和花生四烯酸等ꎬ不仅是储藏油脂的重要组分ꎬ还是植物细胞膜脂的主要成分[3]ꎮFAD包括FAD2㊁FAD3㊁FAD4㊁FAD5㊁FAD6㊁FAD7和FAD8ꎬ其中FAD2和FAD6为ω-6(也称Δ12)去饱和酶ꎬ能够在油酸脂肪酸链的Δ12处引入第二个氢键ꎬ催化油酸转变为亚油酸ꎻFAD3㊁FAD7和FAD8为ω-3(Δ15)去饱和酶ꎬ能够在亚油酸脂肪酸链的Δ15处添加第三个氢键ꎬ进而产生亚麻酸ꎻFAD4㊁FAD5和FAD6的催化底物为棕榈酸[4ꎬ5]ꎮ1992年ꎬArondel等从拟南芥中图位克隆了FAD3基因[6]ꎬ其后FAD基因相继在油菜㊁水稻㊁大豆㊁玉米等作物中被分离[7]ꎮ棉花是世界上最重要的天然纤维作物ꎬ同时也是重要的食用油和蛋白来源[8]ꎮ棉仁中油分含量高达30%~40%ꎬ棉籽油富含多种不饱和脂肪酸ꎬ其中油酸和亚油酸含量约占80%ꎮFAD2是脂肪酸去饱和反应的限速酶ꎬ决定了油脂中油酸和亚油酸的比例和油脂品质ꎬ抑制FAD2基因的表达ꎬ能够降低Δ12-脂肪酸去饱和酶的活性ꎬ致使种子中油酸含量增加ꎮ利用TALENs技术靶向敲除大豆FAD2-1A和FAD2-1B基因ꎬ纯合双基因突变体的油酸含量显著上升ꎬ由20%提高到80%ꎬ而亚油酸含量大幅下降ꎬ由50%降至4%[9ꎬ10]ꎮLiu等利用RNAi技术抑制棉花FAD2-1基因的表达ꎬ获得的棉花高油酸品系High-Ole ̄ic和Mono-Cott的油酸含量分别达到77%和81%[11]ꎮ近年来ꎬ棉花基因组研究进展迅速ꎬ二倍体棉种雷蒙德氏棉(D5)和亚洲棉(A2)以及四倍体棉种陆地棉(AD1)和海岛棉(AD2)的基因组相继被破译[12-15]ꎬ为利用生物信息学在全基因组范围内进行重要基因家族的鉴定和分析提供了可能ꎮ目前ꎬ已知拟南芥基因组中含有17个FAD基因ꎬ水稻中有10个ꎬ油菜中多达84个[16]ꎮ在雷蒙德氏棉基因组中ꎬLiu等鉴定了19个FAD基因[4]ꎮ然而ꎬ陆地棉中FAD基因家族的系统分析尚未见报道ꎮ本研究利用最新发布的陆地棉标准系TM-1的基因组数据[14]ꎬ对膜结合FAD基因家族进行全基因组鉴定ꎬ分析其基因结构㊁染色体定位㊁保守基序㊁基因复制和顺式作用元件ꎬ并分析其在干旱胁迫㊁盐胁迫及施加外源褪黑素下基因的表达模式ꎬ为进一步研究棉花FAD基因家族各成员的功能奠定基础ꎮ1㊀材料与方法1.1㊀供试材料供试陆地棉(GossypiumhirsutumL.)品种为K836ꎬ种子由本实验室保存ꎮ采用1/2Hoagland营养液水培法将棉花幼苗培养至两叶一心ꎮ用100μmol/L褪黑素进行处理ꎬ分别于处理后0㊁3㊁6㊁12h对棉花叶片进行取样ꎬ每个样品3次生物学重复ꎬ置于液氮中冻存备用ꎮ1.2㊀陆地棉FAD基因鉴定与注释陆地棉标准系TM-1的基因组序列数据来自于CottonFGD数据库[17](http://www.cottonfgd.org/)ꎮ利用拟南芥的17个FAD基因的氨基酸序列作为查询序列ꎬ检索棉花基因组蛋白数据库(BlastpꎬEɤe-10)ꎮ同时ꎬ从Pfam数据库(http://pfam.xfam.org/)下载FAD蛋白保守结构域FA_desaturase(PF00487)的隐马尔可夫模型文件ꎬ利用HMMER3.0软件搜索陆地棉的蛋白数据库[18]ꎮ将获得的候选序列提交Pfam和SMART(http://smart.embl-heidelberg.de/)数据库ꎬ确认是否具有FAD蛋白保守结构域ꎮ利用ExPASy(https://web.expasy.org/compute_pi/)分析棉花FAD基因家族成员的分子量和等电点[19]ꎮ利用CELLO(http://cello.life.nctu.edu.tw/)对棉花FAD基因家族成员进行亚细胞定位预测[20]ꎮ1.3㊀陆地棉FAD的系统进化和染色体定位分析利用ClustalW软件对拟南芥和陆地棉FAD蛋白序列进行多重序列比对ꎬ利用MEGA-X软件中的邻接法(neighbor-joiningꎬNJ)构建系统进化树ꎬBootstrap值设置为1000[21]ꎮ根据陆地棉基因组提供的基因注释信息ꎬ获得FAD基因在各染色体上的位置ꎬ利用MapChart2.32软件绘制其染色体分布图[22]ꎮ21山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀1.4㊀陆地棉FAD基因结构、保守基序和上游顺式作用元件分析首先从棉花基因组注释文件(gff文件)获得FAD基因的DNA和cDNA信息ꎬ利用GSDS2.0软件(http://gsds.cbi.pku.edu.cn/)绘制棉花FAD基因家族成员的基因结构图[23]ꎻ利用在线软件MEME(http://meme-suite.org/tools/meme)分析FAD蛋白的保守基序[24]ꎬ基序数值设定为10ꎮ利用陆地棉基因组数据ꎬ提取FAD基因转录起始位点上游1500bp序列ꎬ在PlantCARE数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)中检索ꎬ鉴定上游启动子区域可能存在的顺式作用元件[25]ꎮ1.5㊀陆地棉FAD基因复制分析利用MCScanX软件分析陆地棉基因组内的同源基因[26]ꎬ如比对上的序列长度超过长序列长度的75%且序列的相似性超过75%ꎬ则认为此同源基因为复制基因ꎬ并利用可视化工具Circos(ht ̄tp://circos.ca/)展示复制的同源关系[27ꎬ28]ꎮ采用KaKs_Calculator2.0计算复制基因的Ka(非同义替换率)和Ks(同义替换率)值ꎬ通过Ka/Ks值分析环境选择压力[29]ꎮ1.6㊀RNA-seq分析利用前期试验得到的陆地棉干旱(PEG)和盐胁迫(NaCl)处理3㊁6㊁12㊁24h后的转录组数据(未发表)ꎬ筛选得到FAD基因表达量数据ꎮ利用FPKM(fragmentsperkilobaseofmillionmappedreads)对原始表达数据进行标准化处理ꎬ并利用TBtools软件绘制表达量热图ꎮ以log2(FC)绝对值大于1为差异基因的筛选标准ꎮ1.7㊀基因表达分析采用北京艾德莱生物科技(Aidlab)有限公司植物RNA快速提取试剂盒(RN3802)提取叶片总RNAꎬ分别取1μg总RNA进行反转录ꎬ反应体系和操作程序参照TRUEscriptRTMasterMix(On ̄eStepgDNARemoval)一步法基因组清除反转录试剂盒(PC70)说明ꎮ将反转录产物稀释10倍ꎬ选择Ubiquitin(GenBankNo.AY189972)基因作为内参基因ꎬ进行定量RT-PCRꎬ所用引物见表1ꎬ由北京六合华大基因科技有限公司合成ꎮ选用㊀㊀表1㊀定量PCR引物序列序号基因正向引物(5ᶄ-3ᶄ)反向引物(3ᶄ-5ᶄ)1GH_A01G1380GGGAGCTTTATCGACCGTGGACGAGCCACGTGTATGCGGGTTAT2GH_A01G1382GTTTCCAGCGCTCCGTTTTACGGACCCAAACGCCGGTCAAAATC3GH_A04G1111ACCTTTGGAACAGAAGTCCGGGTCTTTTCGTTCGGTCGGGACAA4GH_A05G0424GCCAAGGTGCCATCTCAGGAAATGGGTTGCTGAGAATCCTTGCC5GH_A05G1248AAACTCTGGCCATCCAGACTCGAAGCCGACACTTTCCCATTGCT6GH_A07G1144CATGGGTTCCGTTGCCTGAGAAACACGGGGAATGCAAATAGGGG7GH_A07G1522AGTGCGGAAGAGCTGAAAGAGCCCAGGGTGATACGCCAGAAAGG8GH_A09G1076GGGACCATGTTTTGGGCTGTCTCGGACATCGGAACCCAAGACTC9GH_A10G2629TCACCATGGTCATGACGAGAAGCATCCCGATCAAGCGTCGTAAGC10GH_A11G0968GGCTTGTCACGGAGTTCTCCAACCCACACGCTATCGCATCTCAA11GH_A11G3588ATGCAGAAGAGCGATCCGTGACTCTTTCGTCAGCTTGCTCCCAC12GH_A12G1258ACCCCACTCTTAAATCTCGCCGAGGTATTGCCAAGCTGTGGTGG13GH_A13G0678CACCAGTACGCCATTACTGCGAAACACGTGGGATGGACCAACTG14GH_A13G2126TATAGGCCACGATTGCGCTCACGTGCCTGTCATGCTTAAACCGC15GH_D01G1464GCGCTCCGTTTTACGCTCATTCACGTTGGAGAGAGCCTGAGGAA16GH_D04G1454ATCATGGCACCCGTTGTCTGAGGGTCGAAGTGCGAACCACTCTT17GH_D05G0426GTTTCCAAGGTTGCCAAGGTGCCTGCAAGGCTGCTGATCGAAGA18GH_D06G1474AGGTGAAGGAGATTGCTCCCGAAGGGCCGATACTCCGATCCATT19GH_D07G1125GAAACTCGACACCAGTACGCGACTTTGCCTGGGCTTCTATGCCA20GH_D07G1526GTATCACCCTGGAACGGCTTGGGTGCCCACACGCTATCACATCT21GH_D08G2811CGACAAGAAAGGACACGGGACACACCATCCCATGCTGATCCCAG22GH_D09G1028ACAGCAACTTGTTTGCTCCCCACTTCTGCTCGTATCCGTGGTGG23GH_D10G2734GAAGCTACCGAAGCAGCAAAGCACACGACATCACCAATGTCACTCA24GH_D11G0999AGGCTTGTCACGGAGTTTTCCACCCACACGCTATCGCATCTCAA25GH_D11G3608TGCAACACACTCACCCTGCATTGCCTTTGTTGCTTCCATTGCGT26UbiquitinAAGACCTACACCAAGCCCAAGACACTCCGCATTAGGACACTC31㊀第4期㊀㊀㊀㊀陈义珍ꎬ等:陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析Aidlab公司的2ˑSybrGreenqPCRMix试剂盒ꎬ反应体系为20μL:2ˑSYBRqPCRMix10μLꎬ基因特异性上下游引物各0.4μL(10μmol/L)ꎬ模板cDNA0.8μLꎬ补ddH2O至20μLꎮ使用Ther ̄moFisherScientificQuantStudio5荧光定量PCR仪ꎬ反应程序:95ħ预变性3minꎻ95ħ15sꎬ60ħ15sꎬ40个循环ꎻ熔点曲线程序为95ħ15sꎬ60ħ1minꎬ95ħ15sꎮ每个样品进行3次重复试验ꎬ采用2-әәCt法计算基因的表达量ꎮ利用Graphpad进行差异显著性分析ꎮ2㊀结果与分析2.1㊀陆地棉FAD基因家族成员的鉴定在陆地棉基因组中共鉴定出37个FAD基因(表2)ꎬ均含有保守的FA_desaturase结构域(PF00487)ꎮFAD蛋白的氨基酸序列长度差异较大ꎬGH_A13G0678最短ꎬ为308个氨基酸ꎬ而GH_A10G2629和GH_D10G2734最长ꎬ均为450个氨基酸ꎻGH_A13G0678的分子量最小ꎬ为35.41kDaꎬGH_A12G1258的分子量最大ꎬ为51.87kDaꎻFAD蛋白的等电点差异较大ꎬGH_D06G1474的等电点最小ꎬ为6.95ꎬ其余FAD蛋白的等电点均在7.29及以上ꎬ为碱性ꎬ以GH_D05G1245的等电点最大(9.37)ꎮ大多数FAD蛋白定位在内质网和质膜上ꎬ少数FAD蛋白位于叶绿体中ꎬ仅GH_D12G1274位于溶酶体中(表2)ꎮ㊀㊀表2㊀陆地棉FAD基因家族基本信息基因染色体定位亚家族外显子数量氨基酸序列长度分子量(kDa)等电点亚细胞定位GH_A01G1380A01:52437258-52438409Omega138344.378.97内质网GH_A01G1382A01:52793676-52794827Omega138344.379.06内质网GH_A01G2453A01:117661885-117664879Omega842749.538.17叶绿体GH_A04G1111A04:76679855-76681857Omega844650.968.47叶绿体GH_A05G0424A05:3932284-3933627Front-end144751.168.43质膜GH_A05G1248A05:11342468-11344453First540546.289.26质膜GH_A06G1435A06:89146264-89147952Sphingolipid233038.407.29质膜GH_A07G1144A07:18014579-18016280Omega837643.508.84内质网GH_A07G1522A07:30432512-30433855Front-end144751.188.36质膜GH_A08G2818A08:124577528-124578871Front-end144751.208.64质膜GH_A09G1076A09:62957866-62959614Omega838845.039.05内质网GH_A10G0260A10:2147390-2149449Sphingolipid233138.628.82质膜GH_A10G2629A10:114411673-114414288Omega845051.198.91叶绿体GH_A11G0968A11:8764840-8766183Front-end144751.388.94质膜GH_A11G3588A11:119680980-119682134Omega138444.258.96内质网GH_A12G1258A12:80872680-80875034Front-end244951.877.98质膜GH_A13G0678A13:14253433-14255056Omega830835.418.69质膜GH_A13G2126A13:105045570-105049386Omega1044551.599.09叶绿体GH_A13G2423A13:108609376-108610533Omega138544.069.09内质网GH_D01G1463D01:30783164-30784315Omega138344.288.94内质网GH_D01G1464D01:30824751-30825902Omega138344.189.04内质网GH_D01G2529D01:64245406-64248408Omega843550.407.42叶绿体GH_D04G1454D04:47832920-47834919Omega844650.728.52叶绿体GH_D05G0426D05:3419790-3421133Front-end144751.208.68质膜GH_D05G1245D06:10327204-10329142First538644.249.37质膜GH_D06G1474D07:44774892-44776562Sphingolipid233038.356.95质膜GH_D07G1125D07:14029866-14031568Omega837643.568.83内质网GH_D07G1526D07:22737990-22739333Front-end144751.198.55质膜GH_D08G2811D08:68604570-68605913Front-end144751.278.61质膜GH_D09G1028D09:35497454-35499216Omega838845.119.05内质网GH_D10G0273D10:2157515-2159474Sphingolipid232337.797.30质膜GH_D10G2734D10:66260614-66263295Omega845051.158.91叶绿体GH_D11G0999D11:8288982-8290325Front-end144751.378.94质膜41山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀㊀㊀表2(续)基因染色体定位亚家族外显子数量氨基酸序列长度分子量(kDa)等电点亚细胞定位GH_D11G3608D11:69715461-69716615Omega138444.279.04内质网GH_D12G1274D12:39933619-39935960Front-end339545.137.30溶酶体GH_D13G2107D13:59015821-59019626Omega1044251.329.17叶绿体GH_D13G2414D13:62456454-62457605Omega138343.898.95内质网2.2㊀陆地棉FAD基因的系统进化和染色体定位分析为了研究陆地棉FAD基因家族成员之间的进化关系ꎬ我们利用陆地棉和拟南芥FAD蛋白序列构建了系统进化树ꎮ按照亲缘关系的远近ꎬ将陆地棉FAD基因家族分为4个亚家族ꎬ分别为First㊁Sphingolipid㊁Front-end和Omega亚家族(图1)ꎮ4个亚家族中的FAD成员数量差异较大ꎬ其中ꎬFirst亚家族中陆地棉FAD成员最少ꎬ仅有2个ꎬ而拟南芥FAD成员多达9个ꎻSphingolipid亚家族中仅含有1个拟南芥FAD成员ꎬ却含有4个陆地棉成员ꎻFront-end亚家族中陆地棉和拟南芥的FAD成员分别为10个和2个ꎻOmega亚家族中陆地棉FAD成员数量最多ꎬ为21个ꎬ拟南芥FAD成员为5个ꎮ图1㊀陆地棉FAD基因家族的系统进化树51㊀第4期㊀㊀㊀㊀陈义珍ꎬ等:陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析㊀㊀根据陆地棉标准系TM-1的基因组注释信息ꎬ将37个FAD基因定位在22条染色体上ꎬ其中A01㊁A13和D01染色体上分别含有3个FAD基因ꎻA05㊁A07㊁A10㊁A11㊁D05㊁D07㊁D10㊁D11和D13染色体上分别含有2个FAD基因ꎻ其余10条染色体上分别含有1个FAD基因(图2)ꎮ图2㊀陆地棉FAD基因家族的染色体分布2.3㊀陆地棉FAD基因结构和保守基序分析陆地棉FAD基因4个亚家族呈现不同的结构特征(图3)ꎮFirst亚家族的FAD成员具有相同的基因结构ꎬ均含有5个外显子ꎻSphingolipid亚家族的FAD基因亦具有相同的基因结构ꎬ均含有2个外显子ꎻFront-end亚家族的基因结构也比较保守ꎬ10个FAD成员中ꎬ8个FAD基因含有1个外显子ꎬ其余2个FAD成员含有2个或3个外显子ꎻOmega亚家族的基因结构存在3种方式ꎬ11个FAD成员含有8个外显子ꎬ2个成员含有10个外显子ꎬ剩余的8个成员含有1个外显子ꎮ利用MEME分析陆地棉FAD蛋白的保守基序ꎬ共发现3个保守组氨酸基序(Histidine-box1㊁Histidine-box2和Histidine-box3)ꎬ但不同亚家族成员的组氨酸基序的序列不同(图4)ꎮOmega亚家族的Histidine-box1为H[D/E]C[G/A]HꎬHis ̄tidine-box2为H[R/D][R/T/I]HHꎬHistidine-box3为H[V/I][I/A/P]HHꎬ尽管中间的氨基酸残基不同ꎬ但两侧均为保守的组氨酸残基ꎮFront-end亚家族的Histidine-box1和Histidine-box2序列十分保守ꎻHistidine-box3为Q[L/I/V]EHHꎬ其起始氨基酸为谷酰胺ꎮSphingolipid亚家族成员的3个组氨酸基序都十分保守ꎬ且基序两侧皆为保守的组氨酸ꎮFirst亚家族Histidine-box3为典型的组氨酸基序ꎬHistidine-box1的最后一个氨基酸残基为亮氨酸ꎬ而Histidine-box2不具有典型的组氨酸基序特征ꎮ2.4㊀陆地棉FAD基因复制分析基因复制在基因家族扩张过程中发挥重要作用ꎮ利用MCScanX对陆地棉基因组中的基因复制事件进行分析ꎬ发现FAD基因家族共存在28对片段复制基因ꎬ无串联复制基因ꎮ其中ꎬ21对基因复制发生在A和D亚基因组之间ꎬ4对发生在A亚基因组之内ꎬ剩余3对发生在D亚基因组之内(图5)ꎬ表明片段复制是棉花FAD基因家族扩张的主要方式ꎮ随后ꎬ利用KaKs_Calculator分析复制基因的Ka/Ks值ꎬ结果显示所有复制基因的Ka/Ks值均小于0.55(表3)ꎮ61山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀图3㊀陆地棉FAD基因结构分析图4㊀陆地棉FAD基因家族的3个保守组氨酸富集区71㊀第4期㊀㊀㊀㊀陈义珍ꎬ等:陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析图5㊀陆地棉FAD基因家族的共线性分析㊀㊀表3㊀陆地棉FAD家族基因的复制分析复制基因对KaKsKa/Ks复制类型GH_A01G1380:GH_D01G14630.0090.0220.413片段复制GH_A04G1111:GH_D04G14540.0110.0560.199片段复制GH_A05G0424:GH_D05G04260.0090.0310.277片段复制GH_A05G1248:GH_D05G12450.0100.0200.517片段复制GH_A06G1435:GH_D06G14740.0080.0400.214片段复制GH_A06G1435:GH_A10G02600.0710.6300.113片段复制GH_A06G1435:GH_D10G02730.0710.6640.106片段复制GH_A07G1144:GH_D07G11250.0100.0320.322片段复制GH_A07G1144:GH_A09G10760.1550.7100.218片段复制GH_A07G1144:GH_D09G10280.1550.7430.208片段复制GH_A07G1522:GH_D07G15260.0010.0370.026片段复制GH_A07G1522:GH_A12G12580.1652.1820.076片段复制GH_A09G1076:GH_D09G10280.0020.0240.093片段复制GH_A09G1076:GH_D07G11250.1450.7560.192片段复制GH_A10G0260:GH_D10G02730.0040.0580.069片段复制GH_A10G2629:GH_D10G27340.0060.0580.100片段复制GH_A10G0260:GH_D06G14740.0750.6700.111片段复制GH_A11G0968:GH_A12G12580.1361.9490.070片段复制GH_A11G0968:GH_D07G15260.0930.8290.112片段复制81山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀㊀㊀表3(续)复制基因对KaKsKa/Ks复制类型GH_A11G0968:GH_D11G09990.0050.0200.237片段复制GH_A11G0968:GH_D12G12740.1421.8910.075片段复制GH_A12G1258:GH_D07G15260.1671.9670.085片段复制GH_A12G1258:GH_D11G09990.1391.9000.073片段复制GH_A12G1258:GH_D12G12740.0160.0840.194片段复制GH_A13G2126:GH_D13G21070.0030.0310.092片段复制GH_D06G1474:GH_D10G02730.0730.7050.103片段复制GH_D07G1125:GH_D09G10280.1450.7920.184片段复制GH_D11G0999:GH_D12G12740.1451.8850.077片段复制2.5㊀陆地棉FAD基因上游顺式作用元件分析利用陆地棉FAD基因转录起始位点上游1500bp序列分析顺式作用元件ꎬ结果发现棉花FAD基因的上游序列除了存在启动子核心元件TATA-box和CAAT-box外ꎬ还存在许多激素和逆境胁迫响应元件(图6)ꎮ其中ꎬ植物激素响应元件有9种ꎬ分别是ABRE㊁AuxRE㊁CGTCA-mo ̄tif㊁TGA-element㊁ERE㊁GARE-motif㊁P-box㊁TCA-element和TGACG-motifꎻABRE㊁ERE和TCA-ele ̄ment分别响应脱落酸㊁乙烯和水杨酸ꎬTGACG-motif和CGTCA-motif为茉莉酸甲酯响应元件ꎬAuxRE和TGA-element为生长素响应元件ꎬGARE-motif和P-box为赤霉素响应元件ꎮ逆境胁迫响应元件有6种ꎬ包括ARE㊁DRE㊁LTR㊁MBS㊁WUN-motif和TC-richꎬ分别是缺氧胁迫㊁冷害㊁低温㊁干旱㊁损伤和防御响应元件ꎮ图6㊀陆地棉FAD基因启动子区域顺式作用元件示意图91㊀第4期㊀㊀㊀㊀陈义珍ꎬ等:陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析2.6㊀陆地棉FAD家族基因在盐胁迫和干旱胁迫下的表达特征为明确FAD基因在盐胁迫和干旱胁迫下的表达模式ꎬ基于转录组数据对37个FAD基因在两种胁迫处理不同时间点陆地棉叶片中的表达量进行聚类分析ꎬ结果如图7所示ꎮ图中log2(FC)代表log2(FoldChange)ꎻNaCl-3h㊁NaCl-6h㊁NaCl-12h㊁NaCl-24h分别代表盐处理3㊁6㊁12㊁24h后的样品ꎻPEG-3h㊁PEG-6h㊁PEG-12h㊁PEG-24h分别代表干旱处理3㊁6㊁12㊁24h后的样品ꎮ图7㊀陆地棉37个FAD基因在盐胁迫和㊀㊀㊀干旱胁迫下的表达分析㊀㊀盐胁迫下ꎬ37个FAD基因呈现不同的表达模式ꎬ其中有28个FAD基因在不同处理时间均有表达ꎬ7个FAD基因未检测到表达信号ꎬ而GH_A06G1435基因仅在处理后6h检测到表达量增加ꎬGH_D01G2529仅在处理后24h检测到表达量增加ꎮ另外ꎬ28个FAD基因呈现不同的表达模式ꎬ17个FAD基因表达量降低ꎬ7个FAD基因表达量增加[log2(FC)>1]ꎬ4个基因的表达量先增后降ꎬ2个基因的表达量先降后增(GH_D08G2811和GH_D11G0999)ꎮ干旱胁迫下ꎬ37个FAD基因中有27个在不同处理时间均有表达ꎬ有6个未检测到表达量变化ꎬ有4个仅在两个处理时间点下表达量增加ꎮ另外ꎬ27个FAD基因中仅GH_D04G1454表达量增加ꎬ17个基因在干旱胁迫下表达量降低ꎬ9个基因仅在一个或两个处理时间(6h或12h)点下表达量增加ꎬ其他时间点的表达量降低ꎮ褪黑素作为一种重要的多效性抗氧化分子ꎬ能帮助植物抵御不利环境的危害ꎬ而且外源褪黑素能显著缓解逆境胁迫引起的伤害[30ꎬ31]ꎮ因此ꎬ本研究基于干旱胁迫和盐胁迫下FAD基因的转录组结果ꎬ选出25个FAD基因(包括Front-end亚家族的8个ꎬOmega亚家族的15个ꎬFirst亚家族的1个ꎬSphingolipid亚家族的1个)ꎬ通过在棉花苗期施加外源褪黑素ꎬ进一步探究FAD基因在外源褪黑素下的表达特征ꎮ定量分析结果显示ꎬ在褪黑素处理后ꎬ1个FAD基因(GH_A01G1380)未检测到表达量的变化ꎬ另外24个FAD基因与对照相比ꎬ有7个基因表达量增加ꎬ17个基因表达量降低ꎬ且上调表达的FAD基因均在处理后6h表达量增加最多ꎮFront-end亚家族8个FAD基因中有6个基因在褪黑素处理后表达量增加ꎬ2个基因表达量降低ꎻOmega亚家族表达的14个FAD基因中仅有1个在处理后表达量增加ꎬ其他基因都呈现表达量降低趋势ꎻFirst㊁Sphingolipid亚家族的各1个基因则均在处理后表现为表达量降低(图8)ꎮ3㊀讨论与结论植物脂肪酸作为储藏油脂和细胞膜脂的重要组分ꎬ既为植物的生长发育提供能量ꎬ也在响应多种逆境胁迫中发挥重要作用[3]ꎮFAD是植物脂肪酸进一步去饱和反应中的关键酶ꎬ目前已在拟南芥㊁水稻㊁花生㊁大豆和油菜等物种中相继完成了全基因组水平上的鉴定和分析[16]ꎮ本研究从陆地棉基因组中鉴定出37个FAD基因ꎬ而二倍体雷蒙德氏棉仅含有19个FAD成员[4]ꎬ是其1.95倍ꎬ造成这种差异的原因可能是棉花进化过程中的异源多倍化现象ꎮ棉属植物进化分析表明ꎬ在170万~190万年前ꎬ陆地棉由A基因组供体亚洲棉和D基因组供体雷蒙德氏棉种间杂交加倍而成[14]ꎮ另外ꎬ陆地棉FAD基因数量远多于模式植物拟南芥和水稻ꎬ分别是其2.18倍和3.70倍ꎮ02山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀柱上∗㊁∗∗分别表示同一基因在不同处理时间与处理0h相比差异显著(P<0.05)㊁极显著(P<0.01)ꎮ图8㊀陆地棉24个FAD基因在褪黑素处理不同时间后的表达分析㊀㊀进化分析将陆地棉和拟南芥的FAD基因分为4个亚家族ꎬ且每个物种的FAD基因在4个亚家族中均有分布ꎬ表明FAD基因的分化可能早于棉花和拟南芥的物种分化ꎮ除了First亚家族ꎬ其他3个亚家族中陆地棉FAD基因的数量远多于拟南芥ꎬ预示Omega㊁Front-end和Sphingolipid亚家族中的陆地棉FAD基因可能在进化过程中发生了物种特异性扩张ꎮ这一结果与之前二倍体雷蒙德氏棉中的报道一致[4]ꎮ基因复制是植物进化的主要动力ꎬ主要包括12㊀第4期㊀㊀㊀㊀陈义珍ꎬ等:陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析串联复制㊁片段复制和全基因组复制[28]ꎮ本研究在陆地棉FAD基因家族中发现了28对复制基因ꎬ来源于4个亚家族ꎬ且全为片段复制ꎬ表明在陆地棉进化过程中FAD基因由于片段复制发生了基因家族扩张ꎮKa/Ks值常用来检验同源基因是否经历了选择作用ꎬKa/Ks>1ꎬ认为有正选择作用ꎻKa/Ks<1ꎬ为纯化或负选择作用ꎻKa/Ks=1ꎬ则认为存在中性选择作用ꎮ本研究中所有复制基因的Ka/Ks值均小于1ꎬ表明陆地棉FAD基因在进化过程中经历了纯化选择作用ꎮ顺式作用元件分析发现ꎬ陆地棉FAD基因的启动子区含有植物激素和逆境胁迫响应元件ꎬ预示其可能在激素信号响应和防御逆境胁迫中起作用ꎮ目前ꎬ已知雷蒙德氏棉中有7个FAD基因受低温诱导表达ꎬ5个FAD基因受低温抑制表达[4]ꎮ部分FAD基因呈现组织特异性表达ꎬ如FAD2-1在发育的种子中特异表达[32]ꎬFAD2-2和FAD2-3在种子发育过程中组成型表达ꎬ在叶片中少量表达[33]ꎮ植物中位于质膜上的脂肪酸大部分是不饱和脂肪酸ꎬ而植物对环境胁迫的抗性与质膜上脂肪酸的不饱和程度密切相关ꎬ膜结合基因对维持植株在多种胁迫下的正常生长起着非常重要的作用ꎮ在拟南芥中ꎬFAD2和FAD6是提高幼苗早期生长耐盐性的重要基因[34ꎬ35]ꎬFAD7基因的反义表达则降低了对盐胁迫和干旱胁迫的耐性[36]ꎮ在番茄中超表达LeFAD3基因增强幼苗早期生长的耐盐性[37]ꎮ为探究陆地棉FAD基因在逆境胁迫下的表达模式ꎬ利用盐胁迫和干旱胁迫的转录组数据对陆地棉37个FAD基因进行分析ꎬ发现70%以上的FAD基因对两种胁迫均有应答ꎬ其中下调表达的FAD基因分别占54%和63%ꎮ值得注意的是ꎬ本研究Front-end亚家族包含的10个基因中除了亚细胞定位预测在溶酶体的GH_D12G1274基因外ꎬ其余9个基因全部定位于质膜ꎬ干旱胁迫后7个基因都是下调表达ꎬGH_A12G1258和GH_D11G0999基因也仅在6h或24h表达量增加ꎬ说明干旱胁迫可能会抑制质膜上FAD基因的表达ꎮ褪黑素在植物非生物胁迫防御系统中发挥重要作用ꎬ且几乎所有非生物胁迫引起的过氧化伤害都能被褪黑素缓解ꎮ褪黑素提高植物对环境胁迫抗性机制主要是通过提高抗氧化物质含量和抗氧化酶活性ꎬ改善光合系统ꎬ调控激素合成和代谢ꎬ调控植物细胞中初级和次级代谢ꎬ刺激细胞合成更多的保护物质等方式[38]ꎮ通过定量PCRꎬ进一步分析了25个FAD基因在施加外源褪黑素后的表达特征ꎬ发现17个基因下调表达ꎬ7个基因上调表达ꎬ1个基因没有表达变化ꎮ值得注意的是ꎬFront-end亚家族中的5个基因在干旱胁迫后下调表达而在褪黑素处理后上调表达ꎬ且在处理后6h表达量显著增加ꎬ它们可作为候选基因用于后续基因功能的进一步探究ꎮ本研究结果可为进一步揭示陆地棉FAD基因逆境响应机理提供参考ꎮ参㊀考㊀文㊀献:[1]㊀戴晓峰ꎬ肖玲ꎬ武玉花ꎬ等.植物脂肪酸去饱和酶及其编码基因研究进展[J].植物学通报ꎬ2007ꎬ24(1):105-113. [2]㊀阮建ꎬ单雷ꎬ李新国ꎬ等.花生FAD基因家族的全基因组鉴定与表达模式分析[J].山东农业科学ꎬ2018ꎬ50(6):1-9. [3]㊀曹福亮ꎬ王欢利ꎬ郁万文ꎬ等.高等植物脂肪酸去饱和酶及其编码基因研究进展[J].南京林业大学学报(自然科学版)ꎬ2012ꎬ36(2):125-132.[4]㊀LiuWꎬLiWꎬHeQꎬetal.Characterizationof19genesenco ̄dingmembrane ̄boundfattyaciddesaturasesandtheirexpres ̄sionprofilesinGossypiumraimondiiunderlowtemperature[J].PLoSONEꎬ2015ꎬ10(4):e0123281.[5]㊀LeeKRꎬLeeYꎬKimEHꎬetal.Functionalidentificationofoleate12 ̄desaturaseandω ̄3fattyaciddesaturasegenesfromPerillafrutescensvar.frutescens[J].PlantCellReportsꎬ2016ꎬ35(12):2523-2537.[6]㊀ArondelVꎬLemieuxBꎬHwangIꎬetal.Map ̄basedcloningofagenecontrollingomega ̄3fattyaciddesaturationinArabidop ̄sis[J].Scienceꎬ1992ꎬ258(5086):1353-1355. [7]㊀杨志刚ꎬ郭子好ꎬ姚琴琴ꎬ等.脂肪酸去饱和酶基因的研究进展[J].生物技术通报ꎬ2013(12):21-26. [8]㊀宋俊乔ꎬ孙培均ꎬ张霞ꎬ等.棉仁高油分材料筛选及其脂肪酸发育分析[J].棉花学报ꎬ2010ꎬ22(4):291-296. [9]㊀HaunWꎬCoffmanAꎬClasenBMꎬetal.Improvedsoybeanoilqualitybytargetedmutagenesisofthefattyaciddesaturase2genefamily[J].PlantBiotechnologyJournalꎬ2014ꎬ12(7):934-940.[10]王福军ꎬ赵开军.基因组编辑技术应用于作物遗传改良的进展与挑战[J].中国农业科学ꎬ2018ꎬ51(1):1-16. [11]LiuQꎬSinghSPꎬGreenAG.High ̄stearicandhigh ̄oleiccot ̄tonseedoilsproducedbyhairpinRNA ̄mediatedpost ̄transcrip ̄tionalgenesilencing[J].PlantPhysiol.ꎬ2002ꎬ129:1732-1743.22山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀[12]PatersonAHꎬWendelJFꎬGundlachHꎬetal.RepeatedpolyploidizationofGossypiumgeneomesandtheevolutionofspinnablecottonfibres[J].Natureꎬ2012ꎬ492:423-427. [13]LiFꎬFanGꎬWangKꎬetal.Genomesequenceofthecultivat ̄edcottonGossypiumarboretum[J].NatureGeneticsꎬ2014ꎬ46(6):567-572.[14]HuYꎬChenJꎬFangLꎬetal.GossypiumbarbadenseandGos ̄sypiumhirsutumgenomesprovideinsightsintotheoriginande ̄volutionofallotetraploidcotton[J].NatureGeneticsꎬ2019ꎬ51(4):739-748.[15]WangMꎬTuLꎬYuanDꎬetal.ReferencegenomesequencesoftwocultivatedallotetraploidcottonsꎬGossypiumhirsutumandGossypiumbarbadense[J].NatureGeneticsꎬ2019ꎬ51(2):224-229.[16]XuYꎬChenBꎬWangRꎬetal.Genome ̄widesurveyandchar ̄acterizationoffattyaciddesaturasegenefamilyinBrassicana ̄pusanditsparentalspecies[J].AppliedBiochemistryandBio ̄technologyꎬ2018ꎬ184(2):582-598.[17]ZhuTꎬLiangCZꎬMengZGꎬetal.CottonFGD:anintegratedfunctionalgenomicsdatabaseforcotton[J].BMCPlantBiolo ̄gyꎬ2017ꎬ17:101.[18]FinnRDꎬCoggillPꎬEberhardtRYꎬetal.ThePfamproteinfamiliesdatabase:towardsamoresustainablefuture[J].Nu ̄cleicAcidsResearchꎬ2016ꎬ44:D279-D285.[19]ArtimoPꎬJonnalageddaMꎬArnoldKꎬetal.ExPASy:SIBbioinformaticsresourceportal[J].NucleicAcidsResearchꎬ2012ꎬ40:W597-W603.[20]YuCSꎬChenYCꎬLuCHꎬetal.Predictionofproteinsubcel ̄lularlocalization[J].Proteins:StructureꎬFunctionandBioin ̄formaticsꎬ2006ꎬ64(3):643-651.[21]KumarSꎬStecherGꎬLiMꎬetal.MEGAX:molecularevolu ̄tionarygeneticsanalysisacrosscomputingplatforms[J].Molec ̄ularBiologyandEvolutionꎬ2018ꎬ35(6):1547-1549. [22]VoorripsR.MapChart:softwareforthegraphicalpresentationoflinkagemapsandQTLs[J].JournalofHeredityꎬ2002ꎬ93(1):77-78.[23]HuBꎬJinJꎬGuoAYꎬetal.GSDS2.0:anupgradedgenefea ̄turevisualizationserver[J].Bioinformaticsꎬ2015ꎬ31(1):1296-1297.[24]BaileyTLꎬJohnsonJꎬGrantCEꎬetal.TheMEMESuite[J].NucleicAcidsResearchꎬ2015ꎬ43:W39-W49.[25]LescotMꎬDehaisPꎬThijsGꎬetal.PlantCAREꎬadatabaseofplantcis ̄actingregulatoryelementsandaportaltotoolsforinsilicoanalysisofpromotersequences[J].NucleicAcidsRe ̄searchꎬ2002ꎬ30(1):325-327.[26]WangYꎬTangHꎬDeBarryJDꎬetal.MCScanX:atoolkitfordetectionandevolutionaryanalysisofgenesyntenyandcol ̄linearity[J].NucleicAcidsResearchꎬ2012ꎬ40(7):e49. [27]KrzywinskiMꎬScheinJEꎬBirolIꎬetal.Circos:aninforma ̄tionaestheticforcomparativegenomics[J].GenomeResearchꎬ2009ꎬ19(9):1639-1645.[28]LiuZꎬFuMꎬLiHꎬetal.SystematicanalysisofNACtran ̄scriptionfactorsinGossypiumbarbadenseuncoverstheirrolesinresponsetoVerticilliumwilt[J].PeerJꎬ2019ꎬ7:e7995. [29]ZhangZꎬLiJꎬZhaoXQꎬetal.KaKs_Calculator:calculatingKaandKsthroughmodelselectionandmodelaveraging[J].GenomicsProteomicsBioinformaticsꎬ2013ꎬ4(4):259-263. [30]LiHꎬChangJꎬChenHꎬetal.Exogenousmelatoninconferssaltstresstolerancetowatermelonbyimprovingphotosynthesisandredoxhomeostasis[J].FrontiersinPlantScienceꎬ2017ꎬ8:295.[31]LiHꎬGuoYꎬLanZꎬetal.MelatoninantagonizesABAactiontopromoteseedgerminationbyregulatingCa2+effluxandH2O2accumulation[J].PlantScienceꎬ2021ꎬ303:110761. [32]LiuQꎬBrubakerCLꎬGreenAGꎬetal.EvolutionoftheFAD2 ̄1fattyaciddesaturaseintronandthemolecularsystemat ̄icsofGossypium(Malvaceae)[J].AmericanJournalofBota ̄nyꎬ2001ꎬ88(1):92-102.[33]PirtleILꎬKongcharoensuntornWꎬNampaisansukMꎬetal.MolecularcloningandfunctionalexpressionofthegeneforacottonΔ ̄12fattyaciddesaturase(FAD2)[J].BiochimicaetBiophysicaActaꎬ2001ꎬ1522(2):122-129.[34]ZhangJTꎬZhuJQꎬZhuQꎬetal.Fattyaciddesaturase ̄6(Fad6)isrequiredforsalttoleranceinArabidopsisthaliana[J].BiochemicalandBiophysicalResearchCommunicationsꎬ2009ꎬ390(3):469-474.[35]ZhangJꎬLiuHꎬSunJꎬetal.ArabidopsisfattyaciddesaturaseFAD2isrequiredforsalttoleranceduringseedgerminationandearlyseedlinggrowth[J].PLoSONEꎬ2012ꎬ7(1):e30355. [36]ImYJꎬHanOꎬChungGCꎬetal.AntisenseexpressionofanArabidopsisomega ̄3fattyaciddesaturasegenereducessalt/droughttoleranceintransgenictobaccoplants[J].MolecularCellꎬ2002ꎬ13(2):264-271.[37]WangHSꎬYuCꎬTangXFꎬetal.Atomatoendoplasmicre ̄ticulum(ER) ̄typeomega ̄3fattyaciddesaturase(LeFAD3)functionsinearlyseedlingtolerancetosalinitystress[J].PlantCellReportsꎬ2014ꎬ33(1):131-142.[38]ArnaoMBꎬHernández ̄RuizJ.Melatoninagainstenvironmen ̄talplantstressors:areview[J].CurrentProteinandPeptideScienceꎬ2021ꎬ22(5):413-429.32㊀第4期㊀㊀㊀㊀陈义珍ꎬ等:陆地棉膜结合脂肪酸去饱和酶基因家族的全基因组鉴定及表达分析。
科技报告的撰写及常见错误

山东省科学技术情报研究院
二、科技报告基本结构
前置部分
封面 题名页 辑要页(信息表) 序或前言
致谢 摘要 目录 插图和附表清单 符号和缩略语说明
正文部分
引言部分(研究概述) 主体部分 结论部分 建议部分 参考文献
结尾部分
附录 索引 发行列表 封底
山东省科学技术情报研究院
(一)前置部分——结构组成
•封 面
根据授权码登录科技 报告呈交系统,上传 报告
• 山东省科技报告呈 交系统 http://202.110.20 3.75:8088/Report Submit/
确认审核通过后,注 册、登录山东省科技 报告共享服务系统,
领取证书 • 山东科技报告服务 系统
http://www.sdstrs. cn/BaogaoCenter /CertificateApply. aspx
山东省科学技术情报研究院
科技报告撰写、呈交流程
根据合同规定撰写科 技报告
• 最终报告至少在结 题验收前2-3个月撰 写呈交,建议项目 结束后呈交报告。
填写基本信息表,发 送至 sdkjbg@, 以获取授权码
• 关于开展山东省科 技计划科技报告工 作的通知 . cn/0b/47/c328a2 887/page.htm
▲参考文献置于报告 正文部分的后面,宜另 起页。
山东省科学技术情报研究院
(三)结尾部分——附录
▲ 内容:附录是正文的辅助材料和补充内 容,由于篇幅过大等原因不便置于正文中;或 对一般读者并非必要但对本专业同行具有参考 价值的材料。
(二)“引言”、“结论”可以作为章标题,“主体”、“正 文”等措词不能作为章标题。
(三)建议文中不使用“本项目”、“本课题”、“项目(课 题)组”等字眼,改用“本研究”或“本报告”等措辞。
山大认可的核心期刊

心理科学
中国心理学会
108
宗教学研究
四川大学宗教学研究所
109
当代宗教研究
上海社会科学院宗教研究所
110
中国宗教
国家宗教事务局
111
世界宗教文化
中国社会科学院世界宗教研究所
112
周易研究
山东大学
113
社会
上海大学文学院
114
社会工作研究
中国民政理论和社会福利研究会
115
民族研究
中国社会科学院民族研究所
中华医学会
73
中华实验外科杂志
中华医学会
74
中华医院感染学杂志
中国人民解放军总医院
75
中华医院管理杂志
中华医学会北京分会
76
药物生物技术
中国药学会
77
中药材
国家药品监督管理局
78
中国药学(英文版)
中国药学会
79
中国药物化学
沈阳药科大学
80
中国医学科学杂志
中国医学科学院
81
解放军护理杂志
第二军医大学
82
中国药科大学学报
中国药科大学
83
中国儿童保健
中华预防医学会
84
护士学杂志
同济医科大学
85
医学与哲学
中国科协自然辩证法协会
86
口腔颌面修复杂志
首都医科大学
87
护士进修杂志
卫生部
88
中国现代普通外科进展
山东大学
人文部分
特类
序号
期刊名称
主办单位
1
中国社会科学(中英文版)
中国社会科学院
2
2010年山东省科技情报科学技术奖评审结果

2010年山东省科技情报科学技术奖评审结果(排序不分先后)一等奖:29项1、青岛市重点产业领域技术路线图研究完成单位青岛市科学技术信息研究所完成人员谭思明于升峰房学祥蓝洁王志玲宋福杰李汇简檀壮吴宁王远征2、关于山东省“十一五”科技发展情况和当前科技发展形势的研究完成单位山东省科学技术情报研究所完成人员王坚韩彬彬唐飞闫法义黄立业姜常梅原顺梅门伟莉刘洁3、组织领域国际卫生发展资料收集与情报学分析完成单位山东大学医院与卫生管理处山东省医药卫生科技信息研究所完成人员李士雪刘岩张秀芹何有琴颜建华程艳敏4、国外临床执业医师监管与考核评价的情报学分析完成单位山东省医药卫生科技信息研究所完成人员刘岩姜学军梁晋英孙竟瀚张红丽颜建华李小涛郭全胜5、青岛市2002-2008年科技统计分析研究报告完成单位青岛市科学技术信息研究所完成人员谭思明于升峰吴宁蓝洁李汇简刘祥檀壮郭琳琳6、山东省粮、饲产业结构演变规律与发展趋势及对策研究完成单位山东省农业科学院信息中心完成人员刘延忠贾春林封文杰韩伟赵维王富军沈军辉7、国内外特低渗透油藏勘探开发配套技术及成功实例调研(试油测试技术)完成单位胜利石油管理局井下作业公司完成人员王海波宗丽琴张震姝杨成孙玉卞士举王建东吴平张太斌李现民杜发勇8、山东省情报学研究生教育的创立与理论实践创新完成单位山东理工大学完成人员葛敬民刘文云王效岳邓尚民李长玲吴红王克平9、菏泽煤化工产业发展战略研究与项目策划完成单位菏泽生产力促进中心完成人员李卫东刘凤莲陆劲松刘志安赵治海刘刚义10、《山东科技年鉴》2009卷完成单位山东省科技厅《山东科技年鉴》编辑部完成人员张怀文宋伟高巍赵青张冠南丁娜11、临沂市大型科学仪器设备协作共享平台建设完成单位临沂市科学技术合作与应用研究院完成人员华冰姜芳李遐尚梅花怀涛万海峰12、2009年国内外石油新技术跟踪调研(石油钻井)完成单位胜利石油管理局完成人员杨利蔡西茂侯芳赵晓波李加娜吴帅飞张敏13、“卫计联手”“育医结合”开展唐氏综合征筛查诊断工作研究完成单位潍坊市妇幼保健院完成人员王爱婷于维新赵桂凤徐继秀于学信14、海洋风能专利数据库建设及专利地图研究完成单位青岛市科学技术信息研究所完成人员谭思明李汉清张卓群赵霞姜静秦洪花朱延雄15、张扭性构造油气成藏规律与勘探实例调研完成单位胜利油田地质科学研究院完成人员程敏周德勇陈晓东高德君张友振16、济南市电子政务培训教育网络应用平台研究完成单位济南市信息中心完成人员钟捷程玲齐砚伟张超王越许青井伟伟沈飞刘芃闻冬洪大阳17、医院质量管理体系的应用研究完成单位山东省医药卫生科技信息研究所完成人员王忠儒李国严岱奕曹先平郭秀文18、农业数字图书馆建设及延伸服务完成单位山东省农业科学院图书馆完成人员唐研徐淑良仝雪琴黎香兰曾燕房毅19、温室环境测控系统智能控制算法研究完成单位山东省农业科学院科技信息工程技术研究中心完成人员王风云秦磊磊刘淑云封文杰刘延忠尚明华20、竞技体育信息服务模式创新与应用研究完成单位山东省体育科学研究中心完成人员钟炼李皿卢成义程静静钟亚平魏红段金花田雪文张翠陈勇宋丽严家高21、青岛市主导产业技术预见(家电与电子信息领域)完成单位青岛市科学技术信息研究所完成人员谭思明于升峰蓝洁李汇简王志玲檀壮吴宁22、肛肠病小针刀综合疗法新技术推广工作报告完成单位山东省医药卫生科技信息研究所完成人员宋玮史晓峰周楠赵芳姜学军23、2010年国外科技与产业发展战略译丛完成单位青岛市科学技术信息研究所完成人员谭思明管泉王淑玲王云飞王春玲初志勇24、《心脑疾病电生理诊断》完成单位山东省医药卫生科技信息研究所完成人员刘康马立华马涛孙卫红鲍冉25、家源性病媒生物相关传染病危险因素调研完成单位胜利石油管理局疾病预防控制中心完成人员曹官时孙宝春姜文琴王郁赵春梅都爱武吴新颖贾秀敏肖太钦付春祥26、发挥省城科技资源优势,建设创新型城市对策研究完成单位济南金源科技信息中心完成人员张岩张人广高鹏霄张勐朱国栋27、现代家禽业中科技期刊传播推广作用研究完成单位山东省农业科学院家禽研究所完成人员殷若新连京华李惠敏孙凯王生雨蔡蕊28、黄河三角洲地区科技信息资源综合服务平台建设完成单位山东省东营生产力促进中心完成人员徐自文程合庆王海兴贾天祥周建团李焕芝张文刘英立袁萍刘志鑫吴敬芝二等奖:50项1、科技型中小企业竞争情报系统平台的建设与研究完成单位山东省科学技术情报研究所完成人员田小元孙莹丛培虎孙丽君李树强邱晓东2、高校职称评审中的学术查新探索完成单位青岛科技大学图书馆完成人员高祀亮顾海明李海兰徐仲谭淑琴周晓梅陆银梅肖东张岩3、面向中小企业应用的Web集成框架完成单位山东生产力促进中心完成人员宋峰孔霞吕峰王磊陈艳秋王春光张宁4、论文《Competitive Intelligence between Government and Enterprises》完成单位山东省科学技术情报研究所完成人员甄丽红张国良刘文5、山东省科学院科研重点领域动态信息服务完成单位山东省科学院情报研究所完成人员赵燕清于俊凤王蕾朱世伟冯海洲王克波6、中小型机构档案管理系统研究与开发完成单位滨州市科学技术情报研究所完成人员宋峰高鹏刘珊刘长明王娟王立新蒋海妞徐新建韩群孙凤鸣徐慧岩7、2008年国内外石油新技术跟踪调研(采油工艺)完成单位中国石油化工股份有限公司胜利油田分公司采油工艺研究院完成人员魏亚峰陈文秋于学信左家强王观华黄伟吴家松8、威海市新产品快速设计制造服务平台完成单位山东生产力促进中心完成人员宋峰吕峰王磊孔霞陈艳秋9、植物水分供应动力模型在节水农业上的示范与应用完成单位菏泽市农业科学院完成人员曲杰汪胜军刘志安陈成君刘凤莲王素芳10、卫生检验创新人才培养体系的构建与实践完成单位济宁医学院完成人员陈廷张凯杨金玲张璟李晶杨海霞陈雁君王长芹郭建丽11、震动性神经损伤评价指标的研究完成单位济宁医学院完成人员张凯邢良红张磊郭玉金张璟林立12、消除麻疹时期麻疹疫苗免疫效果及影响因素研究完成单位济宁医学院完成人员王文军张璟刘琥聂尚丹李晶宋烨周军燕杜珍隋桂英13、国内外实验室发展水平调研完成单位山东省胜利油田地质科学研究院完成人员马士坤王秋语高德君张冬玉盖瑜张娜14、临沂市科技信息检索与咨询服务规范化建设完成单位临沂市科学技术合作与应用研究院完成人员华冰姚艳伟任晓林黄思良葛大伟万海峰15、连续管钻井技术调研完成单位胜利石油管理局钻井工艺研究院完成人员彭军生蔡西茂侯芳杨利李庆胥林16、山东省2006-2010中医药新技术推广实施研究完成单位山东省医药卫生科技信息研究所完成人员王忠儒李国严岱奕曹先平郭秀文17、国内外水平井压裂酸化及修井新技术调研完成单位胜利石油管理局井下作业公司完成人员杜发勇李俊成陈东王从严姜立辉黄琼冰孙文森林惠星潘勇薛仁江暴志娟18、电化学治疗前列腺增生新技术工作报告完成单位山东省医药卫生科技信息研究所完成人员史晓峰宋玮周楠赵芳姜学军19、国内外水平井生产测试技术调研完成单位中国石油化工股份有限公司胜利油田分公司采油工艺研究院完成人员张峰皇甫洁郝金克田玉刚伊伟锴宋相隆杨慧温盛魁20、山东省疾病预防控制中心学术论文汇编2008完成单位山东省疾病预防控制中心完成人员刘鸣江丽丽孙成玺胡滨黄静龚文杰严敏张红田勇孙晓黎21、稠油化学蒸汽驱技术调研完成单位中国石化股份胜利油田分公司地质科学研究院完成人员曹嫣红张冬玉周征刘桂芳苏海芳王志全22、妇科常用小手术完成单位山东大学齐鲁医院完成人员张师前王晓红于云海董延磊赖海清沈春燕于浩23、国内外石油工程装备技术水平调研(采油工艺装备)完成单位中国石油化工股份有限公司胜利油田分公司采油工艺研究院完成人员徐文庆郭海滨马珍福杭发琴吕长杰刘常福马恩才24、国内外普通稠油油藏水驱后转蒸汽驱技术调研完成单位中国石化股份胜利油田分公司地质科学研究院完成人员曹嫣红张冬玉马士坤蒋红红高德君25、《当代尘肺病学》完成单位泰安市中心医院分院完成人员李光杰26、充分利用科技情报资源,促进芦笋分子标记技术的应用研究与开发完成单位山东省潍坊市农业科学院完成人员张元国柴树桔刘英杨林李萌张东起27、2009年度山东省自然基金与博士基金项目查新统计报告完成单位山东省科学技术情报研究所完成人员邢文杨德祥李莎蔡馨燕刘洁张莉聂静田文香陈薇28、多网融合的农村科技服务热线系统研发与应用完成单位山东省农业科学院科技信息工程技术研究中心完成人员封文杰赵佳刘延忠刘峰张秀兰29、2009年山东省农业科学院科学抗旱专题片完成单位山东省农业科学院科技信息工程技术研究中心完成人员孔庆富董暐邵历刘锋30、山东省科技查新质量控制综合体系的研建完成单位山东省农业科学院图书馆完成人员黎香兰徐淑良唐研曾燕仝雪芹王福军31、温室蝴蝶兰栽培管理知识模型的研制完成单位山东省农业科学院科技信息工程技术研究中心完成人员张晓艳刘锋刘淑云封文杰王风云尚明华32、农业信息获取与发布有效途径研究完成单位山东省农业科学院科技信息工程技术研究中心完成人员赵佳封文杰刘锋王艳红刘延忠秦磊磊王剑非徐刘璐33、优惠制原产地政策查询系统完成单位聊城检验检疫局完成人员魏艳霞马国良赵君琦孙艺健王岩于立欣王炜玮刘青青李望涛34、青岛市大型德尔菲调查分析平台的开发与应用完成单位青岛市科学技术信息研究所完成人员谭思明于升峰檀壮蓝洁王志玲李汇简房学祥吴宁35、青岛市“十一五”科技发展规划评价研究完成单位青岛市科学技术信息研究所完成人员谭思明于升峰蓝洁李汇简吴宁檀壮36、青岛市蓝色经济区海洋科研资源调查完成单位青岛市科学技术信息研究所完成人员谭思明于升峰蓝洁刘祥吴宁李汇简檀壮37、青岛市“十二五”社会发展愿景调查与科技支撑研究完成单位青岛市科学技术信息研究所完成人员谭思明于升峰房学祥王志玲宋福杰檀壮38、2009青岛市市南区科学技术发展报告完成单位青岛市科学技术信息研究所完成人员谭思明管泉王淑玲王春玲初志勇39、基于数据挖掘的个性化网页推送服务模式研究完成单位青岛市科学技术信息研究所完成人员周文鹏40、NSTL服务站系统升级,服务山东半岛科技创新完成单位青岛市科学技术信息研究所完成人员谭思明赵霞姜静张卓群李汉清周文鹏王静王春莉牛海萍41、网络资源采集技术在科技型中小企业创新平台中的应用完成单位青岛市科学技术信息研究所完成人员谭思明赵霞姜静李汉清张卓群秦洪花梁明李晓东42、2009青岛市科学技术发展报告完成单位青岛市科学技术信息研究所完成人员谭思明管泉苏红王淑玲初志勇王云飞王春玲43、国内外特低渗透油藏勘探开发配套技术及成功实例调研(钻井工程)完成单位胜利石油管理局完成人员杨宁宁赵力国侯芳吴帅飞李加娜44、济南青年科技明星计划可持续发展研究完成单位济南科学技术信息研究所完成人员李丹王文平李海波周延山45、创新信息服务模式打造决策服务品牌完成单位济南科学技术信息研究所完成人员朱月卿王颖莉白美李丹李英46、基于.NET的科技查新管理信息系统完成单位济南科学技术信息研究所完成人员武张亮韩轶王莹莹陈静王伟华毕玉晓王鹏47、医院网络考试平台的研制完成单位山东省医药卫生科技信息研究所完成人员岳媛贾培民谷景亮段永璇张睿王鹏范军48、省管科技类社会组织科学发展长效机制建设完成单位山东省科学技术情报研究所完成人员程凡照高巍邱晓东49、VPN技术在信息共享平台建设中的应用完成单位山东省科学技术情报研究所完成人员许洪光庞怡刘涛50、山东省科技基础条件平台建设研究完成单位山东省科学技术情报研究所完成人员毛原宁程凡照蔡馨燕董亚倩李锐张玉华杨红春李树国三等奖:29项1、临沂市农村科技信息“村村通”平台建设完成单位临沂市科学技术合作与应用研究院完成人员华冰孙继成尚梅花邢艳霞怀涛黄思良2、大化工市场快讯完成单位山东化工网完成人员张福田李海静张晓谦韩学岗李学军3、科技信息检索查新用户服务系统的研究与开发完成单位滨州市科学技术情报研究所完成人员庄波赵舰刘颖州许可军刘建华韩群王梦升韩美娟许春晖王立新范晓明4、事业单位人事管理系统研究与开发完成单位滨州市科学技术情报研究所完成人员孙继磊蒋海妞王爱芹郭永强白勇杨涛张永辉王晓亮李超侯磊孙红梅5、“智慧城市”国内外发展现状及青岛市发展对策与建议完成单位青岛市科学技术信息研究所完成人员秦洪花李汉清赵霞孙琴舒飞涛6、地方特色资源数据库建设完成单位菏泽市科学技术情报研究所完成人员刘志安李卫东彭林赵鲁军孔庆运孙辉7、开展科技情报定题服务,建设特色专题网站完成单位济南市科学技术信息研究所完成人员陈静罗军武张亮王鹏毕玉晓李军8、山东化工网完成单位山东化工网完成人员张福田李海静张晓谦韩学岗李学军9、腹膜代阴道成形术完成单位山东大学齐鲁医院完成人员张师前司红卫于云海王立杰董延磊赖海清于浩10、2008年国内外石油新技术跟踪调研(地震采集)完成单位中国石化胜利石油管理局完成人员段卫星何京国卢湘鹏宁鹏鹏张在武田根海赵国勇潘家智张志林兰文新11、国内外特低渗透油藏勘探开发配套技术及成功案例调研(地质勘探开发)完成单位胜利油田分公司地质科学研究院完成人员张敬轩姜婷蒋红红解曙巍康元勇吴锦莲12、氨甲喋呤治疗子宫内膜异位囊肿的临床研究完成单位日照市中医医院完成人员百安芬13、自制中药乌龙祛斑膏合并针灸治疗外阴白色病变的临床研究完成单位日照市中医医院完成人员杜鑫殷世美李新春范丰田魏玲14、充分利用科技情报资源,促进极早熟大果优质毛桃新品种选育完成单位山东省潍坊市农业科学院完成人员张东起柴树桔刘英李志萍邢利庆徐广宾15、农业科技期刊在现代农业信息化发展中的作用完成单位《山东农业科学》编辑部完成人员王丽丽梅林谷卫刚房毅刘炳福张玉英张丽荣陈庆禹赵文祥16、不同尺度农业信息的空间变异研究完成单位山东省农业科学院科技信息工程技术研究中心完成人员杨玉建仝雪芹王殿昌封文杰刘淑云王利民张晓艳王风云尚明华17、青岛市“十二五”科技发展理念、思路、目标及重点任务研究完成单位青岛市科学技术信息研究所完成人员谭思明于升峰李汇简蓝洁吴宁檀壮郭琳琳18、婴幼儿静脉穿刺专用轮椅的研制与临床应用完成单位菏泽市立医院完成人员智莉马敏李莹王攀茹袁培军19、青岛市城市科技发展路线图研究完成单位青岛市科学技术信息研究所青岛市科技发展战略研究所完成人员谭思明于升峰房学祥宋福杰王志玲20、国外海洋产业发展战略汇编完成单位青岛市科学技术信息研究所完成人员谭思明王淑玲王春玲21、山东省科技多媒体资源共享系统研建完成单位山东省科学技术情报研究所完成人员张俊莹刘斌魏美勇姚斐曲鹏22、青岛市工业企业科技创新活动特征分析报告完成单位青岛市科学技术信息研究所完成人员谭思明于升峰吴宁蓝洁李汇简檀壮刘祥23、山东省农村科技专题教材制播大型系列课件《一村一品》完成单位山东省科学技术情报研究所完成人员姚斐米可宫宇林慧芳杨斌孙媛媛张国良张俊莹刘斌魏美勇24、电视专题片《春风化雨》完成单位济南市科学技术信息研究所完成人员胡立伟王正君石磊张琦25、纤支镜下高频电刀联合气管支架加后装放疗治疗食管癌气道狭窄的疗效完成单位日照市中医医院完成人员张华闫培清葛长胜乔秀丽卜宪聪冯辉孙中美26、科技查新业务拓展研究完成单位济南市科学技术信息研究所完成人员王伟华武张亮陈静毕玉晓王鹏李军27、“山东省优质畜禽品种资源”系列专题片完成单位山东省科学技术情报研究所完成人员林慧芳姚斐张国良杨斌李绮斌刘斌魏美勇李哲28、基于网络的科技查新综合业务平台的设计开发完成单位青岛市科学技术信息研究所完成人员谭思明赵霞张卓群姜静王春莉朱延雄王静舒飞涛孙琴牛海萍29、青岛市“十二五”科技发展预测分析报告完成单位青岛市科学技术信息研究所完成人员谭思明于升峰吴宁蓝洁李汇简檀壮。
科研工作流程OA系统的设计分析——以某省级科研单位为例

科研工作流程OA系统的设计分析——以某省级科研单位为例郑业萌;朱军;丁华【摘要】Through reviewed the research and development situation of Office Automation system at domestic and abroad, on the basis of analysis and interpretation of research business process, this paper takes a medium and small scale research institute as the empirical object. Through the following such aspects as system login, task notification, task sched- uler, document retrieval, task perform, results storage, system maintenance and rights management, this paper proceeds the empirical design of Office Automation.%在综述OA系统的国内外研究与开发现状基础上,突出科研单位的科研工作特点,对科研业务流程进行分析和解读,以一个中小规模的科研院所单位为实证对象,着重从科研工作的角度,分任务提醒、任务安排、资料检索、完成任务、成果存储等方面进行科研业务流程OA系统的实证设计。
本文研究可作为科研单位实施科研工作OA的部分设计依据。
【期刊名称】《科学与管理》【年(卷),期】2012(032)003【总页数】6页(P70-75)【关键词】科研单位;OA系统;科研业务流程;设计【作者】郑业萌;朱军;丁华【作者单位】山东轻工业学院/山东省农业管理干部学院;山东出版集团;山东省科技发展战略研究所/山东省科学院科学决策支持重点实验室,济南市250014【正文语种】中文【中图分类】G311OA(Office Automation)即办公自动化,它利用先进的技术,使人们的各种办公业务活动逐步由各种设备、各种人机信息系统来协助完成,达到充分利用信息,提高工作效率和工作质量,提高生产率的目的[1]。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
山东省科学院情报所检索中心
科技查新委托书(红色标记为委托人必须填写)
项目名称:
项目主要负责人(1-3名)
委托人:委托单位:
联系人:联系电话:
E-mail:
是否需邮寄(联系地址:邮编:)委托日期:预定完成日期:
山东省科学院情报所检索中心
说明:
(1)查新项目名称、关键词,除国内查新外,国内外查新需要详细提供中、英文关键词,关键词的来源应是课题相关的专业惯用词,不能单纯地从一般词典中选择。
查新人员根据用户提供的主题词进行复核后确定。
(2)委托人信息中包括机构名称,通讯地址,项目负责人姓名,联系人姓名,电话,传真及E-mail。
(3)认真填写待查新项目的科学技术要点一栏,该栏内容反映查新课题的需要确立的新颖点,要着重说明查新项目在科学技术特征、技术参数或指标、应用范围等方面,查新委托人自我判断的新颖性,空间不够时可加附页。
如若报告科技要点或查新点不清楚,最好能给我们提供研究项目的技术报告。
(1、科技内容:完整简要叙述查新项目的研究内容(类似文摘),要着重说明查新项目的主要科学技术特征、技术参数或指标、应用范围、查新委托人自我判断的新颖性等2.研制总结或预研报告:用作鉴定材料。
3.技术背景:支持查新点。
)
(4)查新点主要是指需要查证的内容要点。
(本查新项目与已有技术的明显不同之处,即需要查证的内容要点。
要求填写清楚,查新中途不得更改),包括课题的研究内容、特点、结构、工艺、配方以及技术参数、指标和发明点等。
(5)委托书填好后,可以网上委托(发电子邮件,电子邮件的标题及委托书名称请按照,王萍--山东大学—申报科技攻关项目查新方式发送),请务必填好联系电话,发送邮件后请及时打电话予以确认;也可以直接将报告送过来。
(6)、正常时间,课题国内查新需4-5个工作日完成,国内外查新5-6个工作日完成。
(特殊情况除外)
(7)我们的地址:
山东省科学院情报研究所(济南市科院路19号)科学院机关楼东三楼(从东门进3楼)
(9)、附:
委托书科学技术要点、查新要求书写示范
委托书科学技术要点、查新要求书写示范。