此反应由丙酮酸羧化酶
河南大学生化简答题

1酮体生成和利用的生理意义。
(1)酮体是脂酸在肝内正常的中间代谢产物,是甘输出能源的一种形式;(2)酮体是肌肉尤其是脑的重要能源。
酮体分子小,易溶于水,容易透过血脑屏障。
体内糖供应不足(血糖降低)时,大脑不能氧化脂肪酸,这时酮体是脑的主要能源物质。
2试述乙酰CoA在脂质代谢中的作用.在机体脂质代谢中,乙酰CoA主要来自脂肪酸的β氧化,也可来自甘油的氧化分解;乙酰CoA在肝中可被转化为酮体向肝外运送,也可作为脂肪酸生物合成及细胞胆固醇合成的基本原料。
3试述人体胆固醇的来源与去路?来源:⑴从食物中摄取⑵机体细胞自身合成去路:⑴在肝脏可转换成胆汁酸⑵在性腺,肾上腺皮质可以转化为类固醇激素⑶在欺负可以转化为维生素D3⑷用于构成细胞膜⑸酯化成胆固醇酯,储存在细胞液中⑹经胆汁直接排除肠腔,随粪便排除体外。
4什么是血浆脂蛋白?试述血浆脂蛋白的分类,来源及生理功能?血浆脂蛋白是脂质与载脂蛋白结合形成球形复合体,是血浆脂蛋白的运输和代谢形式。
.血浆脂蛋白的分类方法有两种:1电泳法:可敬脂蛋白分为乳糜微粒(CM) β-脂蛋白, 前-β脂蛋白和α脂蛋白四类2超速离心法:可将脂蛋白分为乳糜微粒(CM),极低密度脂蛋白(VLDL),低密度脂蛋白(LDL)和高密度脂蛋白(HDL)四类,分别相当于电泳分离的CM、前β-脂蛋白、β-脂蛋白和α-脂蛋白四类。
各种血浆脂蛋白的来源主要生理功能如下:①CM由小肠黏膜细胞合成,功能是转运外源性甘油三酯和胆固醇;②VLDL由肝细胞合成、分泌,功能是转运内源性甘油三酯和胆固醇;③LDL由VLDL在血浆中转化而来,功能是转运内源性胆固醇,即将胆固醇由肝转运至肝外组织;④HDL主要由肝细胞合成、分泌,功能是逆向转运胆固醇,即将胆固醇由肝外组织转运到肝。
1、酶的催化作用有何特点?①具有极高的催化效率,如酶的催化效率可比一般的催化剂高10 8~1020倍;②具有高度特异性:即酶对其所催化的底物具有严格的选择性,包括:绝对特异性、相对特异性、立体异构特异性;③酶促反应的可调节性:酶促反应受多种因素的调控,以适应机体不断变化的内外环境和生命活动的需要。
三羧酸循环(TCA)

TCA 的运转必须通过 O2 条件下才能运转, 实际上O2并不直接参加TCA,那么O2在何处参 加反应呢?
TCA除了产生1个GTP外,另外的能量 均潜在3NADH和1FADH2中,为了TCA的运 转,NAD+和FAD必须再生。NAD+和FAD 的再生则是通过DADH和FADH2进入电子传 递链,将H交给O2,释放潜能生成ATP而实 现。所以,TCA的运转必须有O2。
EMP:高能磷酸基团直接转移给ADP放能 TCA:琥珀酰CoA中的高能键 键水解放能 硫酯
8、琥珀酸氧化生成延胡索酸
第三次脱氢(FAD脱氢) 可逆
生成1FADH2
该酶结合在线粒体内膜上,丙二 酸是竞争性抑制剂
9、延胡索酸水化生成苹果酸
水化作用 可逆 消耗1H2O
10、苹果酸脱氢氧化生成草酰乙酸
第五节 磷酸戊糖途径(HMP PPP)
磷酸戊糖途径的概念:是G分解的另一条途径: 在6—P—G上直接氧化,再分解产生5—P—核糖。
磷酸戊糖途径PPP:Pentose Phosphate Pathway 己糖磷酸途径HMP:Hexose Monophosphate Pathway 磷酸己糖支路HMS:Hexose Monophosphate Shunt G直接氧化途径DOPG:Direct
第四次脱氢 可逆
消耗1NAD+,生成1NADH+H+
总反应式:
乙酰CoA+3NAD++FAD+GDP+Pi+2H2O
2CO2+3NADH+3H++FADH2+GTP
+HS—CoA
四、化学量计算
(一)物质量计算 1mol乙酰CoA (二)能量计算 1、计算1mol乙酰CoA彻底氧化分解产生的ATP的数目 1+3×3+1×2=12molATP 2 molCO2+1molCoA
第十一章 糖类代谢--王镜岩《生物化学》第三版笔记(完美打印版)

反应放能,在生理条件下不可逆(K大于300)。由己糖激酶或葡萄糖激酶催化,需要Mg2+或Mn2+。己糖激酶可作用于D-葡萄糖、果糖和甘露糖,是糖酵解过程中的第一个调节酶,受6-磷酸葡萄糖的别构抑制。有三种同工酶。葡萄糖激酶存在于肝脏中,只作用于葡萄糖,不受6-磷酸葡萄糖的别构抑制肌肉的己糖激酶Km=0.1mM,肝脏的葡萄糖激酶Km=10mM,平时细胞中的葡萄糖浓度时5mM,只有进后葡萄糖激酶才活跃,合成糖原,降低血糖浓度,葡萄糖激酶是诱导酶,胰岛素可诱导它的合成。6-磷酸葡萄糖也可由糖原合成,由糖原磷酸化酶催化,生成1-磷酸葡萄糖,在磷酸葡萄糖变位酶的催化下生成6-磷酸葡萄糖。此途径少消耗1个ATP。6-磷酸葡萄糖由葡萄糖6-磷酸酶催化水解,此酶存在于肝脏和肾脏中,肌肉中没有。
三、能量变化
C6H12O6+2Pi+2ADP+2NAD+=2C3H4O3+2ATP+2NADH+2H++2H2O
有氧时2个NADH经呼吸链可产生6个ATP,共产生8个ATP;无氧时生成乳酸,只有2个ATP。在骨骼肌和脑组织中,NADH进入线粒体要经过甘油磷酸穿梭系统,在细胞质中由3-磷酸甘油脱氢酶催化,将磷酸二羟丙酮还原生成3-磷酸甘油,进入线粒体后再氧化生成磷酸二羟丙酮,返回细胞质。因为其辅酶是FAD,所以生成FADH2,只产生2个ATP。这样其还原当量(2H++2e)被带入线粒体,生成FADH2,进入呼吸链,结果共生成6个ATP。
二、糖的消化和吸收
(一)消化
淀粉是动物的主要糖类来源,直链淀粉由300-400个葡萄糖构成,支链淀粉由上千个葡萄糖构成,每24-30个残基中有一个分支。糖类只有消化成单糖以后才能被吸收。
生化名词解释

重组修复:通过复制后的同源DAN单链之间交换使双链中一条单链上对应损伤的空隙得以修复的方式.合成代谢:从生物体外吸取养料,通过一系列生化反应转变成自己的物质,此过程消耗能量。
又叫同化作用。
分解代谢:将体内原有组分经一系列生化反应分解成不能利用的物质而排出体外,此过程产生能量。
又叫异化作用.生物能学:研究发生在活细胞内的能量转换的定量关系以及支撑这些转换的化学过程的性质.糖酵解:糖酵解途径指糖原或葡萄糖分子分解至生成丙酮酸的阶段,是体内糖代谢最主要途径。
糖异生作用:非糖物质(糖的异生作用的前体,如丙酮酸、乳酸、氨基酸等)转变为葡萄糖的过程。
发酵:葡萄糖在无氧条件下转变成酒精或乳酸的过程。
厌氧有机体把糖酵解生成NADH中的氢交给丙酮酸脱羧后的产物乙醛,使之生成乙醇的过程称之为酒精发酵。
如果将氢交给病酮酸丙生成乳酸则叫乳酸发酵。
巴斯德效应:在厌氧条件下,向高速发酵的培养基中通入氧气,则葡萄糖消耗减少,抑制发酵产物积累的现象称为巴斯德效应。
即呼吸抑制发酵的作用。
底物/无效循环:一对催化两个途径的中间代谢物之间循环的方向相反、代谢上不可逆的反应。
有时该循环通过ATP的水解导致热能的释放。
底物水平磷酸化:在底物被氧化的过程中,底物分子内部能量重新分布产生高能磷酸键(或高能硫酯键),由此高能键提供能量使ADP(或GDP)磷酸化生成A TP(或GTP)的过程称为底物水平磷酸化。
此过程与呼吸链的作用无关,以底物水平磷酸化方式只产生少量A TP。
生物氧化:糖、脂、蛋白质等有机物质在细胞中经过一系列的氧化分解,最终生成CO2和H2O等小分子物质并释放出化学能的总过程称为生物氧化。
柠檬酸循环:发生在线粒体基质内,经由一系列脱氢及脱羧反应将乙酰-CoA最终氧化成CO2 的单向循环途径。
回补反应:酶催化的,补充柠檬酸循环中间代谢物供给的反应,例如由丙酮酸羧化酶生成草酰乙酸的反应。
乙醛酸循环:植物细胞内脂肪酸氧化分解为乙酰CoA之后,在乙醛酸体(glyoxysome)内生成琥珀酸、乙醛酸和苹果酸;此琥珀酸可用于糖的合成,该过程称为乙醛酸循环戊糖磷酸途径:6-磷酸-葡萄糖转变成CO2和5-磷酸核酮糖的过程,也称HMS.高能化合物:指体内氧化分解中,一些化合物通过能量转移得到了部分能量,把这类储存了较高能量的化合物,如三磷酸腺苷(ATP),磷酸肌酸,称为高能化合物生物氧化:有机分子在体内氧化分解成CO2和H2O并释放出能量的过程。
某大学生物工程学院《生物化学》考试试卷(1191)

某大学生物工程学院《生物化学》课程试卷(含答案)__________学年第___学期考试类型:(闭卷)考试考试时间:90 分钟年级专业_____________学号_____________ 姓名_____________1、判断题(100分,每题5分)1. 主动转运有两个显著特点;一是逆浓度梯度进行,因而需要能量驱动;二是具有方向性。
()答案:正确解析:2. 胆固醇是生物膜的主要成分,可调节膜的流动性,原理是胆固醇是两性分子。
()答案:正确解析:3. RNA聚合酶能以两个方向同启动子结合,并启动相邻基因的转录。
但是,模板链的选择由另外的蛋白因子确定。
()答案:错误解析:核不均一RNA是不成熟的mRNA。
4. 血脂包括甘油三酯、磷脂、胆固醇及其酯、游离脂肪酸和载脂蛋白等。
()答案:错误解析:血脂不包括载脂蛋白。
5. 人体细胞中的核苷酸部分从食物消化吸收而来,部分是体内自行合成。
()答案:错误解析:6. 逆转录只发现在RNA病毒,DNA病毒没有。
()答案:错误解析:7. 电子通过呼吸链的传递方向是从∆Eϴ′正到∆Eϴ′负。
答案:错误解析:8. 一个细菌只有一条双链DNA,人的一个染色体含有46条双链DNA。
()[山东大学2016研]答案:错误解析:人的一个体细胞含有46条双链DNA,一个染色体仅含有1条双链DNA。
9. 阻遏蛋白是阻碍RNA聚合酶与启动子的结合。
()[武汉科技大学2012研]答案:错误解析:阻遏蛋白是基于某种调节基因所制成的一种控制蛋白质,在原核生物中具有抑制特定基因(群)产生特征蛋白质的作用。
阻遏蛋白能识别特定的操纵基因,当操纵序列结合阻遏蛋白时会阻碍RNA聚合酶与启动序列的结合,或使RNA聚合酶不能沿DNA向前移动,阻遏转录,介导负性调节。
10. 人体内若缺乏维生素B6和维生素PP,均会引起氨基酸代谢障碍。
()答案:错误解析:11. 三羧酸循环中,琥珀酸脱氢酶催化琥珀酸氧化成延胡索酸时,电子受体是NAD+。
川北医学院成教学院本科学生《生物化学》自学习题及答案

Qixiaojuan 贡献,答案持续更新中
内含子。 19.核蛋白体循环核蛋白体循环是指活化的氨基酸在核糖体上, 以 mRNA 为模板合成多肽链的 过程。 20.翻译后加工 肽链从核蛋白体释放后,经过细胞内各种修饰处理,成为有活性的成熟蛋 白质的过程。.
二、填空题
1.当氨基酸溶液的 pH=pI 时,氨基酸以 兼性 离子形式存在;当 pH>PI 时,氨基酸 以 阴 离子形式存在。 2.各种蛋白质 N 元素的含量比较相近,平均为 16% 。 3.蛋白质的 一 级结构决定它的空间结构和生物学功能,该结构是指多肽链中 氨基 酸 的排列顺序。 4.蛋白质的基本组成单位是 氨基酸 。 5.蛋白质是由许多 氨基酸 通过 脱水 连接形成一条或多条 肽 链。在每条链的两端 有游离的 COOH 基和游离的 NH2 基,这两端分别称为该链的 C 末端和 N 末 端。 6. 构成蛋白质的 20 种氨基酸中碱性氨基酸有赖氨酸、 精氨酸 、 组氨酸 ,酸性氨基酸有 天冬氨酸 、 谷氨酸 。 7.蛋白质的变性是指在理化因素作用下引起蛋白质的 内部 构象发生改变、 活性 丧失, 没有 H 键 的断裂。 8. 蛋白质亲水胶体稳定的两个因素是蛋白质颗粒表面的 电荷层 和 水化膜 。 9.有紫外吸收能力的氨基酸有 苯丙氨酸 、 酪氨酸 、 色氨酸 ,它们中间以 色氨酸 的 吸收最强。 10.维持蛋白质二级结构最主要的力是 H 键 。 11. 蛋白质二级结构形式主要有α—螺旋、β—折叠、和β—转角,维持蛋白质二级结构的 稳定因素是 H 键 。 12. 核酸的基本结构单位是 核苷酸 。 13. DNA 双螺旋中只存在 两 种不同碱基对。T 总是与 A 配对,C 总是与 G 配对。 14. 核酸的主要组成是 碱基 , 戊糖 和 磷酸 。 15. 双链 DNA 中若 C-G 含量多,则 Tm 值高。 16. 蛋白质和核酸对紫外光均有吸收。蛋白质的最大吸收波长是 250_nm;核酸的最大吸收 波长是_260__nm。 17. 全酶由酶蛋白和辅助因子组成,在催化反应时,二者所起的作用不同,其中 酶蛋白 决定 酶的专一性和高效率, 辅助因子 起传递电子、原子或化学基团的作用。 18. 辅助因子包括 辅酶 , 辅基 和 金属离子 等。其中 辅基 与酶蛋白结合紧密, 需要 变性剂 除去, 辅酶 与酶蛋白结合疏松,可用 超滤、透析方法 除去。 19. 酶是由 生物体内活细胞 产生的,具有催化能力的 有机物 。 20. 酶活性的调节包括酶 激活 的调节和酶 抑制 的调节。 21. 酶的专一性可以分为 绝对 专一性 、 相对专一性和 立体 专一性。 22. 酶 促 动 力 学 的 双 倒 数 作 图 (Lineweaver-Burk) 作 图 法 , 得 到 的 直 线 在 横 轴 上 的 截 距 为 1/[S] ,纵轴上的截距为 1/V 。 23. 磺胺类药物可以抑制二氢叶酸合成酶,从而抑制细菌生长繁殖。 24. 维 生 素 是 维 持 生 物 体 正 常 生 长 所 必 需 的 一 类 微 量 有机物质。主要作用是 作 为 酶 参与体内代谢。
4.08级生化复习题-糖代谢参考答案

糖代谢名词解释:1.糖异生:非糖物质(如丙酮酸乳酸甘油生糖氨基酸等)转变为葡萄糖的过程。
2.Q酶:Q酶是参与支链淀粉合成的酶。
功能是在直链淀粉分子上催化合成(α-1, 6)糖苷键,形成支链淀粉。
3.乳酸循环乳:酸循环是指肌肉缺氧时产生大量乳酸,大部分经血液运到肝脏,通过糖异生作用肝糖原或葡萄糖补充血糖,血糖可再被肌肉利用,这样形成的循环称乳酸循环。
4.发酵:厌氧有机体把糖酵解生成NADH中的氢交给丙酮酸脱羧后的产物乙醛,使之生成乙醇的过程称之为酒精发酵。
如果将氢交给病酮酸丙生成乳酸则叫乳酸发酵。
5.变构调节:变构调节是指某些调节物能与酶的调节部位结合使酶分子的构象发生改变,从而改变酶的活性,称酶的变构调节。
6.糖酵解途径:糖酵解途径指糖原或葡萄糖分子分解至生成丙酮酸的阶段,是体内糖代谢最主要途径。
7.糖的有氧氧化:糖的有氧氧化指葡萄糖或糖原在有氧条件下氧化成水和二氧化碳的过程。
是糖氧化的主要方式。
8.肝糖原分解:肝糖原分解指肝糖原分解为葡萄糖的过程。
9.磷酸戊糖途径:磷酸戊糖途径指机体某些组织(如肝、脂肪组织等)以6-磷酸葡萄糖为起始物在6-磷酸葡萄糖脱氢酶催化下形成6-磷酸葡萄糖酸进而代谢生成磷酸戊糖为中间代谢物的过程,又称为磷酸已糖旁路。
10.底物水平磷酸化(substrate phosphorlation):ADP或某些其它的核苷-5′—二磷酸的磷酸化是通过来自一个非核苷酸底物的磷酰基的转移实现的。
这种磷酸化与电子的转递链无关。
11.柠檬酸循环(citric acid cycle):也称为三羧酸循环(TAC),Krebs循环。
是用于乙酰CoA中的乙酰基氧化成CO2的酶促反应的循环系统,该循环的第一步是由乙酰CoA经草酰乙酸缩合形成柠檬酸。
12.回补反应(anaplerotic reaction):酶催化的,补充柠檬酸循环中间代谢物供给的反应,例如由丙酮酸羧化酶生成草酰乙酸的反应。
13.乙醛酸循环(glyoxylate cycle):是某些植物,细菌和酵母中柠檬酸循环的修改形式,通过该循环可以收乙乙酰CoA经草酰乙酸净生成葡萄糖。
生化复习题简答案 小平

05 糖代谢四、问答题1.糖代谢和脂代谢是通过那些反应联系起来的?答:(1)糖酵解过程中产生的磷酸二羟丙酮可转变为磷酸甘油,可作为脂肪合成中甘油的原料。
(2)有氧氧化过程中产生的乙酰CoA是脂肪酸和酮体的合成原料。
(3)脂肪酸分解产生的乙酰CoA最终进入三羧酸循环氧化。
(4)酮体氧化产生的乙酰CoA最终进入三羧酸循环氧化。
(5)甘油经磷酸甘油激酶作用后,转变为磷酸二羟丙酮进入糖代谢。
2.什么是乙醛酸循环?有何意义?答:乙醛酸循环是有机酸代谢循环,它存在于植物和微生物中,可分为五步反应,由于乙醛酸循环与三羧酸循环有一些共同的酶系和反应,将其看成是三羧酸循环的一个支路。
循环每一圈消耗2分子乙酰CoA,同时产生1分子琥珀酸。
琥珀酸产生后,可进入三羧酸循环代谢,或经糖异生途径转变为葡萄糖乙醛酸循环的意义:(1)乙酰CoA经乙醛酸循环可以和三羧酸循环相偶联,补充三羧酸循环中间产物的缺失。
(2)乙醛酸循环是微生物利用乙酸作为碳源的途径之一。
(3)乙醛酸循环是油料植物将脂肪转变为糖和氨基酸的途径。
3.磷酸戊糖途径有什么生理意义?答:(1)产生的5-磷酸核糖是生成核糖,多种核苷酸,核苷酸辅酶和核酸的原料。
(2)生成的NADPH+H+是脂肪酸合成等许多反应的供氢体。
(3)此途径产生的4-磷酸赤藓糖与3-磷酸甘油酸可以可成莽草酸,进而转变为芳香族氨基酸。
(4)途径产生的NADPH+H+可转变为NADH+H+,进一步氧化产生ATP,提供部分能量。
4.为什么说三羧酸循环是糖、脂和蛋白质三大物质代谢的共同通路?答:(1)三羧酸循环是乙酰CoA最终氧化生成CO2和H2O的途径。
(2)糖代谢产生的碳骨架最终进入三羧酸循环氧化。
(3)脂肪分解产生的甘油通过酵解产生丙酮酸,后者转化成乙酰CoA后再进入三羧酸循环氧化,脂肪酸经β-氧化产生乙酰CoA也需进入三羧酸循环才能氧化。
(4)蛋白质分解产生的氨基酸经脱氨后碳骨架可进入三羧酸循环,同时,三羧酸循环的中间产物可作为氨基酸的碳骨架接受氨后合成必需氨基酸。
名词解释

1、转录:在DNA或RNA分子上合成出与其核苷酸顺序相对应的RNA或DNA的过程。
2、底物水平磷酸化:是指物质在脱氢或脱水过程中,产生高能代谢物并直接将高能代谢物中能量转移到ADP(GDP)生成ATP(GTP)的过程。
3、联合脱氨基作用:氨基酸先与a-酮戊二酸进行转氨基作用,生成相应的a-酮酸及谷氨酸,然后后者在L-谷氨酸脱氢酶作用下脱去原来氨基生成原来的a-酮戊二酸并释放出氢的过程。
4、酮体:在肝脏中,脂肪酸氧化分解的中间产物乙酰乙酸、β-羟基丁酸及丙酮,三者统称为酮体。
肝脏具有较强的合成酮体的酶系,但却缺乏利用酮体的酶系。
酮体是脂肪分解的产物,而不是高血糖的产物。
进食糖类物质也不会导致酮体增多。
5、糖异生:生物体将多种非糖物质转变成葡萄糖和糖原的过程。
6、转氨基作用指的是一种α-氨基酸的α-氨基转移到一种α-酮酸上,生成另一种氨基酸和相应的α-酮酸的过程。
7、解偶联剂:指一类能抑制偶联磷酸化的化合物。
这些化合物能使呼吸链中电子传递所产生的能量不能用于ADP的磷酸化,而只能以热的形式散发,亦即解除了氧化和磷酸化的偶联作用。
8、必需氨基酸指的是人体自身不能合成或合成速度不能满足人体需要,必须从食物中摄取的氨基酸。
9、维生素是人和动物为维持正常的生理功能而必需从食物中获得的一类微量有机物质,在人体生长、代谢、发育过程中发挥着重要的作用。
10、肽键具有一定程度的双键性质,参与肽键的六个原子C、H、O、N、Cα1、C α2不能自由转动,位于同一平面,此平面就是肽平面,也叫氨酰平面。
11、DNA双螺旋是一种核酸的构象,在该构象中,两条反向平行的多核苷酸链相互缠绕形成一个右手的双螺旋结构。
糖酵解是指在氧气不足条件下,葡萄糖或糖原分解为丙酮酸或乳酸的过程,此过程中伴有少量ATP的生成。
这一过程是在细胞质中进行,不需要氧气,每一反应步骤基本都由特异的酶催化。
在缺氧条件下丙酮酸则可在乳酸脱氢酶的催化下,接受磷酸丙糖脱下的氢,被还原为乳酸。
植物磷酸烯醇式丙酮酸羧化酶的研究进展

2021年 2月 Journal of Science of Teachers′ College and University Feb. 2021文章编号:1007-9831(2021)02-0054-07植物磷酸烯醇式丙酮酸羧化酶的研究进展于济1,2,沙伟1,2,张梅娟1,2,马天意1,2(齐齐哈尔大学 1. 生命科学与农林学院,2. 抗性基因工程与寒地生物多样性保护黑龙江重点实验室,黑龙江 齐齐哈尔 161006)摘要:磷酸烯醇式丙酮酸羧化酶(Phosphoenolpyruvate carboxylase,PEPC,EC 4.1.1.31)是广泛存在的一种细胞质酶,催化磷酸烯醇式丙酮酸和HCO3-生成草酰乙酸.PEPC作用的产物在植物的生长发育过程和植物应对环境胁迫反应中起到调控的作用,因而被广泛关注.介绍了植物PEPC的种类、结构特征、不同物种中PEPC基因的克隆与分离、在植物逆境反应中的应用、植物PEPC 的活性调节,为深入研究PEPC基因提供理论依据.关键词:磷酸烯醇式丙酮酸羧化酶;植物;植物逆境胁迫中图分类号:Q945 文献标识码:A doi:10.3969/j.issn.1007-9831.2021.02.011Research progress of plant phosphoenolpyruvate carboxylaseYU Ji1,2,SHA Wei1,2,ZHANG Meijuan1,2,MA Tianyi1,2(1. School of Life Sciences,Agriculture and Forestry,2. Heilongjiang Provincial Key Laboratory of Resistance Gene Engineering andProtection of Biodiversity in Cold Areas,Qiqihar University,Qiqihar 161006,China)Abstract:Plant phosphoenolpyruvate carboxylase(PEPC,EC 4.1.1.31)is a ubiquitous cytoplasmic enzyme that catalyzes the production of Oxaloacetate by phosphoenolpyruvate and HCO3-.The products of PEPC play important roles in the growth and development of plants and the responses of plants to environmental stresses,so that the PEPC is widely concerned.Introduced the types,structural characteristics,the cloning and isolation of PEPC genes in different species,the applications in plant stress responses,and the researches of plant PEPC activity regulation,which provides a theoretical basis for further studiesof PEPC genes.Key words:phosphoenolpyruvate carboxylase;plant;plant stress responses磷酸烯醇式丙酮酸羧化酶(Phosphoenolpyruvate carboxylase,PEPC,EC 4.1.1.31)是一种胞质酶,在HCO3-存在的情况下,可以催化磷酸烯醇式丙酮酸(Phosphoenolpyruvate,PEP)的β-羧化反应,以Mg2+为辅因子生成草酰乙酸(Oxobutanedioic acid,OAA)和无机磷酸[1]70,此反应为不可逆反应.PEPC存在于所有植物中,如绿藻、蓝细菌、大多数古细菌、非光合细菌中,但在动物和真菌中不存在[2]15. PEPC以其在C4和景天酸代谢(Crassulacean acid metabolism,CAM)光合作用中的作用而闻名,在此过程中,其初步固定了大气中的CO2[3]274.然而,PEPC在C3植物的非光合和光合组织中也发挥着广泛的作用,它可以通过补充C4-二羧酸进行能量和生物合成代谢,主要起抗衰老作用[4-5].在C3植物叶片和非光合收稿日期:2020-11-15基金项目:齐齐哈尔大学大学生创新创业训练计划项目(202010232817);黑龙江省省属高等学校基本科研业务费青年创新人才项目(135309364);黑龙江省省属高等学校基本科研业务费科研项目植物性食品加工技术特色学科专项(YSTSXK201876);黑龙江省人力资源和社会保障厅2018年省级留学回国人员择优资助项目作者简介:于济(1997-),女,吉林松原人,在读硕士研究生,从事植物逆境分子遗传学研究.E-mail:*****************通信作者:马天意(1989-),男,黑龙江齐齐哈尔人,讲师,博士,从事植物逆境分子遗传学研究.E-mail:********************组织中,PEPC的主要作用是在三羧酸循环中补充中间体,以及后面的氮同化和各种生物合成途径[3]274.此外,PEPC还参与了广泛的生理和发育过程,包括种子萌发和发育、果实成熟、豆科植物根瘤固氮等,在气孔保卫细胞中提供苹果酸以及增强了对渗透压和生物胁迫的耐受性[2,6-9].正是因为PEPC在植物中发挥着非常重要的作用,因此被人们广泛关注.为了更好地了解植物体中的磷酸烯醇式丙酮酸羧化酶基因的作用机制,本文概括分析了植物磷酸烯醇式丙酮酸羧化酶基因的分类、功能、分离与克隆、活性调节以及在植物抵抗逆境过程中的作用等方面的研究结果,介绍了磷酸烯醇式丙酮酸羧化酶基因在基因工程方面的应用,也为进一步研究磷酸烯醇式丙酮酸羧化酶基因提供了理论基础.1 PEPC的基本结构1984年,首先从大肠杆菌(Escherichia coli)克隆的PEPC基因推导了PEPC的一级结构[10].目前,已经分别在植物和细菌中发现了许多PEPC序列,包括同一生物体中的同工型,并且大约500个部分序列已经记录在GenBank中,主要用于系统发育重建[11-12].系统进化树显示,这些PEPC是从相同的祖先进化而来的[3,13].PEPC多肽的大小随生物体的种类而异:细菌、维管束植物、蓝细菌、原生动物(如疟疾病原体)的氨基酸残基数量大约为870(100 kDa),970(110 kDa),1010(116 kDa)或1150(134 kDa)[1]71.尽管已知古细菌的亚基大小非常小(约60 kDa),并且没有常见的变构调节子,但尚无来自古细菌的PEPC 序列数据[14-15].此外,还寻找了藻类PEPC的序列数据,因为硒的纯化亚型之一是由3种不同的亚基组成的[16],这种新的PEPC类型也在发育中的蓖麻(Ricinus communis)种子中被发现[17].在拟南芥(Arabidopsis thaliana)基因组序列中确定了另一种新的进入高等植物PEPC的第4种亚型[18],在分子大小上与蓝细菌的PEPC相似,并且缺少N端磷酸化位点,这是植物PEPC的标志.在植物和微生物界的多样性和广泛分布使得PEPC成为系统发育分析中最有趣的目标之一.对PEPC序列的比对表明,大约100个残基始终是保守的(相同的),另外100个残基是保守的具有相似的氨基酸残基的,对于成对的陆地植物酶,其识别率超过71%,因为C末端是高度保守的,所以长度的任何差异似乎是由于在N末端或内部区域(大约10个基因位点)处添加或插入了额外的序列而引起的.所制备的重组PEPCs的数量和特征仍是有限的,所涉及的物种目前不超过15个[1]71,因此有必要做进一步的工作.2 植物PEPC的分类及生理生化功能PEPC基因已大致分为2个亚家族,分为植物型PEPC(PTPC)和细菌型PEPC(BTPC)[19]11.PTPC表现出高度的遗传保守性,并在其100~110 kDa蛋白质中包含高度保守的N末端丝氨酸磷酸化基序和关键的C末端四肽QNTG[1,20].相比较而言,116~118 kDa的BTPC蛋白质显示低序列相似性并包含1个类似原核的C端(R/K)NTG四肽基序[17,21].PTPC典型地存在于同型四聚体-1类PEPCs中,而BTPCs作为调节和催化亚基存在于超常的异质配合物中(2类PEPCs)[2]26.PTPC又可以分为C4,C3和根特异亚型[21-22],它们在植物细胞中发挥着不同的功能.植物中PEPC的C3,C4类型至少在3个关键方面不同[23]:(1)C4型的PEPC通过吸收C4和CAM物种中的大气CO2参与了CO2浓缩机制的第1步,C3型PEPC存在于所有植物中,它参与多种生理功能,如三羧酸循环中中间体的补给,OAA的合成以及随后的苹果酸及其衍生物的合成[2]15.(2)C4型PEPC存在于叶肉细胞中,其表达方式对实现C4光合循环具有重要意义[24],而C3型的PEPC分布在C3和/或C4植物的不同组织中,起到管家的作用[22]865.(3)C4型PEPC的底物(PEP)饱和常数(Km)高于C3型,并且显示更多对苹果酸的耐受性,但是在一些植物物种中(如南美白菊科(Chenopodiaceae)植物和异子蓬(Suaedaaralocaspica))尚未将不同的PEPC区分为C3或C4类型,仅命名为ppc-1,ppc-2[25],它们的功能需要进一步验证.3 植物PEPC基因的分离与克隆1953年,植物的磷酸烯醇式丙酮酸羧化酶从菠菜(Spinacia oleracea)叶中首次被分离出来.目前,PEPC基因已在多种植物中被鉴定和研究,拟南芥和水稻(Oryzasativa)中分别报道4,6个PEPC家族成员[18,26],在大豆(Glycine max)中共鉴定到10个PEPC基因(GmPEPC1~10),其中GmPEPC6,GmPEPC8,GmPEPC9被铝毒、寒害、盐害等非生物胁迫诱导表达[27].花生(Arachis hypogaea)基因组报道5个PEPC 基因(AhPEPC1~5)[28],涂嘉琦[29]等从蔓花生(Arachis duranensis)基因组中鉴定到9个PEPC基因.马海洋[30]等从菠萝(Ananas comosus)基因组中鉴定出3个PEPC基因.邵姁[31]等从蓝莓(Vaccinium corymbosum)果实中克隆到磷酸烯醇式丙酮酸羧化酶编码基因VcPEPC,该基因开放阅读框全长为2 907 bp,可编码968个氨基酸,分子量为110.59 kD.赵晋锋[32]等从谷子(Setariaitalica)基因组中鉴定出1个SiPEPC基因,进一步研究表明,SiPEPC基因在拔节期和抽穗期正常光照强度下参与了对干旱胁迫的响应,推测SiPEPC基因参与了谷子对非生物逆境的应答,可能在干旱和其它逆境胁迫信号途径中起关键作用.4 植物PEPC基因在植物逆境反应中的作用非生物胁迫(如盐度、干旱、高温)通常会对植物的光合作用产生负面影响[33]901.PEPC已被建议用于支持主要的生理代谢途径(即三羧酸循环中中间体的补给等),以帮助植物和微生物避免和/或耐受极端的环境胁迫[7,34].在水分胁迫下,PEPC介导PEP的羧基化为OAA,随后合成苹果酸和渗透活性化合物(糖、氨基酸、糖醇)以帮助植物耐受水分胁迫[34-35].铝的积累、磷和铁的缺乏导致PEPC的上调,从而增加了包括苹果酸和柠檬酸在内的有机酸合成[36-38].有机酸向土壤中的排泄不仅可以增加磷和铁的可溶性形态并为植物所利用,还可以与Al3+形成稳定的配合物,从而降低毒性[39].生物应激(如病毒感染)也可以诱导PEPC的高活性,推测PEPC可以在防御作用中增强植物抗毒素、氨基酸、蛋白质的合成[40-41].也有研究表明,番茄(Lycopersicon esculentum)PEPC基因在盐、冷、植物激素胁迫下均有不同表达[19]15.可以推测PEPC 对植物的生长发育和抗逆性起一定作用.5 PEPC基因的应用目前,已经在多种植物物种中使用了不同PEPC基因作为改善作物的手段.如在水稻中,C4-PEPC使转基因水稻具有耐旱性,但叶绿体定位的PEPC对铵同化至关重要[42].玉米(Zea mays)C4型PEPC基因在小麦(Triticum aestivum)受体内实现了正确的转录和准确的剪接,也证明了玉米C4型PEPC基因在小麦中表现了一定的光合生理效应[43].以转甘蔗(Saccharum officinarum)PEPC基因的籼稻植株和非转基因的植株为研究材料,发现在光合效率和产量相关性状方面,转甘蔗PEPC基因植株都有较大提高,可以实现增产目的[44].张桂芳[45]等首次将含有稗草(Echinochloacrusgalli)根型的PEPC基因对水稻进行遗传转化,研究结果表明,转基因水稻的PEPC活性最高,为对照的5.85倍,植株叶片的净光合速率(Pn)较对照相比提高了20.0%.尹吴[46]等发现,与对照相比,转玉米PEPC基因的杨树表现出较强的光利用能力,其羧化能力和酶活性与对照相比最高可分别增加62.3%,38.6%.最近有研究表明,许多PEPC基因在增强对各种非生物和生物胁迫的耐受性中起调节作用.在干旱和盐胁迫下,PEPC的过度表达增强了耐受性[8]1513,但抑制导致转基因植物对渗透胁迫的敏感性增加[47].同样,拟南芥AtPPC4也可能在干旱胁迫中发挥作用[33]906.然而,表达PEPC的马铃薯(Solanum tuberosum)转基因植株表现出整体有机氮含量的增加,但淀粉和可溶性糖含量有所损失[48].6 植物组织中PEPC的活性调节PEPC通常由4个相同的亚基组成,相对分子质量约为95~110 kDa.大多数PEPC是变构酶,具有多种变构效应物,具体取决于生物的种类[1]70.此外,维管植物PEPCs通过位于N末端附近的保守丝氨酸处的可逆磷酸化来调节[49].目前解析了来自大肠杆菌和玉米的PEPC三维结构[50-51],这种结构信息以及通过定点诱变获得的信息为长期以来一直深入研究的催化和变构调节的分子机理提供了启示. 大多数PEPC都受变构调节,维管植物的效应仅限于一组效应类型,双子叶植物的PEPCs被葡萄糖6-磷酸(G6P)激活,并被L-苹果酸或天冬氨酸抑制,而单子叶植物的PEPC被甘氨酸或丙氨酸进一步激活[52]154.相反,大肠杆菌PEPC受更复杂的方式调节,被乙酰辅酶A、果糖1,6-双磷酸、长链脂肪酸、鸟苷3′-二磷酸5′-二磷酸激活,并被天冬氨酸或L-苹果酸抑制[53].此外,细菌PEPC中不存在的调节性磷酸化是植物PEPC中固有的[1]73.调节性磷酸化后,PEP的半饱和浓度略有降低,而抑制剂的半饱和浓度在包含底物和Mg2+生理浓度的反应混合物中增加了约2.7倍[52]155.Tovar-Mendez[52,54-55]等采用目前可用的最明确的玉米C4-PEPC制剂进行了全面的动力学分析,从生理条件下的动力学测量清楚地表明了变构效应子和磷酸化的重要性.S形PEP饱和度曲线归因于附加的PEP与G6P位点的结合[1]73,这些研究成功地应用了变构调控的协调过渡模型来描述玉米C4-PEPC的动力学行为,并且通过快速动力学分析观察到伴随变构过渡的构象变化[56].PEPC的活性在多个层次上受到调节.变构控制由调节PEPC活性(尤其是在细胞质pH值下)的正(葡萄糖6-P,甘氨酸)、负(L-苹果酸,天冬氨酸)效应子施加[3]280.此外,C4-PEPC通过光依赖性磷酸烯醇丙酮酸羧化酶激酶(Phosphoenolpyruvate carboxylase kinase,PEPCK)进行调节性磷酸化[57].PEPC的磷酸化形式与去磷酸化形式的动力学性质不同,如更高的最大反应速率对PEP的亲和力更高,对变构效应子的敏感性有所改变,从而减轻了苹果酸对其抑制作用,并增强了其对葡萄糖-6-P和甘氨酸的活化作用[58-59].高粱(Sorghum bicolor)中所有PTPC(SbPPC1-5)均含有带有保守丝氨酸的N末端磷酸化结构域,BTPC (SbPPC6)中不存在此域,PEPCK基因家族包括3个基因(SbPPCK1~3)[60],在叶肉细胞中,SbPPCK1的表达是由光触发的,其转录本在叶肉中比束鞘细胞更为丰富,由于这些原因,它被认为是光合异构体,其中SbPPCK2和SbPPCK3的功能未知[61].另一种可能调控PEPC活性的翻译后修饰(Post-translational modification,PTM)是保守的赖氨酸残基的单泛素化作用,使该酶对苹果酸和天冬氨酸更加敏感[62-64].据报道,该PTM(仅针对C3型PEPCs)存在于蓖麻种子[62]、Hakea prostata种子[63]、高粱种子[64-65]、拟南芥叶片[65-66]中.此外,在不同的植物组织(叶片、保卫细胞、根、果实)[67-70]以及大麦(Hordeum vulgare)、小麦、高粱种子[71-74]中还报道了2条免疫反应性PTPC条带,表明上部条带是下部条带的单泛素化形式.PEPC改变了该酶的动力学特性,从而干扰了其与PEP结合的能力,并增强了其对大多数代谢物效应子的敏感性[62]29656.7 展望植物磷酸烯醇式丙酮酸羧化酶作为C4型光合途径的关键酶,对其结构特点、代谢途径已有较为深入的研究[75].现在PEPC基因已经在越来越多的植物中被发现,并对其功能进行研究探索.在C4和CAM植物中,PEPC的作用以及其基因表征信息比较详细,但在C3植物中PEPC表达的细节报道很少,仍需进一步探索.转PEPC基因植物也在不同方面表达出PEPC基因对植物的调控功能,PEPC作为植物中广泛存在的酶,其在植物应对环境胁迫过程中的作用逐渐被发现,尤其是CAM植物中的大多数物种具有较高抗逆性,而这之中PEPC的相关功能机制还有待进一步挖掘,因此有必要进一步对PEPC基因在植物抗逆性研究中的作用进行详细探索.参考文献:[1] Izui K,Matsumura H,Furumoto T,et al.Phosphoenolpyruvate carboxylase:a new era of structural biology[J].Annual reviewof plant biology,2004,55:69-84[2] O′Leary B,Park J,Plaxton W C.The remarkable diversity of plant PEPC(phosphoenolpyruvate carboxylase):recent insightsinto the physiological functions and post-translational controls of non-photosynthetic PEPCs[J].The biochemical journal,2011(1):15-34[3] Chollet R,Vidal J,O'Leary M H.PHOSPHOENOLPYRUVATE CARBOXYLASE:A Ubiquitous,Highly Regulated Enzyme inPlants[J].Annual review of plant physiology and plant molecular biology,1996,47:273-298[4] Driever S M,Kromdijk J.Will C3crops enhanced with the C4CO2-concentrating mechanism live up to their full potential(yield)?[J].Journal of experimental botany,2013,64(13):3925-3935[5] Masumoto C,Miyazawa S,Ohkawa H.Phosphoenolpyruvate carboxylase intrinsically located in the chloroplast of rice plays acrucial role in ammonium assimilation[J].Proceedings of the National Academy of Sciences of the United States of America,2010,107(11):5226-5231[6] Cousins A B,Baroli I,Badger M R,et al.The role of phosphoenolpyruvate carboxylase during C4 photosynthetic isotope exchangeand stomatal conductance[J].Plant physiology,2007,145(3):1006-1017[7] O′Leary B,Fedosejevs E T,Hill A T,et al.Tissue-specific expression and post-translational modifications of plant-andbacterial-type phosphoenolpyruvate carboxylase isozymes of the castor oil plant,Ricinus communis L[J].Journal of experimental botany,2011,62(15):5485-5495[8] Qin N,Xu W,Hu L,et al.Erratum to:Drought tolerance and proteomics studies of transgenic wheat containing the maize C4phosphoenolpyruvate carboxylase(PEPC)gene[J].Protoplasma,2016,253(6):1513[9] 蔡小宁,陈茜,杨平,等.磷酸烯醇式丙酮酸羧化酶的生物信息学分析[J].安徽农业科学,2008(3):914-916[10] Fujita N,Miwa T,Ishijima S,et al.The primary structure of phosphoenolpyruvate carboxylase of Escherichia coli.Nucleotidesequence of the PPC gene and deduced amino acid sequence[J].Journal of biochemistry,1984(4):909-916[11] BesnardG,Offmann B,Robert C,et al.Assessment of the C4 phosphoenolpyruvate carboxylase gene diversity in grasses(Poaceae)[J].Theor Appl Genet,2002,105(2-3):404-412[12] Gehrig H,Heute V,Kluge M.New partial sequences of phosphoenolpyruvate carboxylase as molecular phylogeneticmarkers[J].Molecular phylogenetics and evolution,2001,20(2):262-274[13] Svensson P,Blasing O,Westhoff P,et al.Evolution of C4 phosphoenolpyruvate carboxylase[J].Archives of Biochemistry andBiophysics,2003,414(2):180-188[14] Sako Y,Takai K,Nishizaka T,et al.Biochemical relationship of phosphoenolpyruvate carboxylases(PEPCs)from thermophilicarchaea[J].Fems Microbiology Letters,1997,153(1):159-165[15] Sako Y,Takai K,Uchida A,et al.Purification and characterization of phosphoenolpyruvate carboxylase from the hyperthermophilicarchaeon Methanothermussociabilis[J].FEBS Letters,1996,392(2):148-152[16] Rivoal J,Trzos S,Gage D A,et al.Two unrelated phosphoenolpyruvate carboxylase polypeptides physically interact in the highmolecular mass isoforms of this enzyme in the unicellular green alga Selenastrumminutum[J].The journal of biological chemistry,2001,276(16):12588-12597[17] Blonde J D,Plaxton W C.Structural and kinetic properties of high and low molecular mass phosphoenolpyruvate carboxylaseisoforms from the endosperm of developing castor oilseeds[J].The journal of biological chemistry,2003,278(14):11867-11873 [18] Sánchez R,Cejudo F J.Identification and expression analysis of a gene encoding a bacterial-type phosphoenolpyruvate carboxylasefrom Arabidopsis and rice[J].Plant physiology,2003(2):949-957[19] Waseem M,Ahmad F.The phosphoenolpyruvate carboxylase gene family identification and expression analysis under abiotic andphytohormone stresses in Solanum lycopersicum L[J].Gene,2019,690:11-20[20] Xu W,Ahmed S,Moriyama H,et al.The importance of the strictly conserved,C-terminal glycine residue in phosphoenolpyruvatecarboxylase for overall catalysis:mutagenesis and truncation of GLY-961 in the sorghum C4leaf isoform[J].The journal of biological chemistry,2006,281(25):17238-17245[21] Gennidakis S,Rao S,Greenham K,et al.Bacterial-and plant-type phosphoenolpyruvate carboxylase polypeptides interact in thehetero-oligomeric Class-2 PEPC complex of developing castor oil seeds[J].The plant journal:for cell and molecular biology,2007(5):839-849[22] Dong L Y,Masuda T,Kawamura T,et al.Cloning,expression,and characterization of a root-form phosphoenolpyruvate carboxylasefrom Zea mays:comparison with the C4-form enzyme[J].Plant & cell physiology,1998,39(8):865-873[23] Gehrig H,Faist K,Kluge M.Identification of phosphoenolpyruvate carboxylase isoforms in leaf,stem and roots of the obligate CAMplant Vanilla planifolia Salib.(Orchidaceae):a physiological and molecular approach[J].Plant molecular biology,1998,38(6):1215-1223[24] Stockhaus J,Schlue U,Koczor M,et al.The promoter of the gene encoding the C4 Form of phosphoenolpyruvate carboxylase directsmesophyll-specific expression in transgenic C4Flaveriaspp[J].The plant cell,1997,9(4):479-489[25] Rosnow J J,Edwards G E,Roalson E H.Positive selection of Kranz and non-Kranz C4 phosphoenolpyruvate carboxylase aminoacids in Suaedoideae(Chenopodiaceae)[J].Journal of experimental botany,2014,65(13):3595-3607[26] Muramatsu M,Suzuki R,Yamazaki T,et al.Comparison of plant-type phosphoenolpyruvate carboxylases from rice:identificationof two plant-specific regulatory regions of the allosteric enzyme[J].Plant & cell physiology,2015,56(3):468-480[27] Wang N,Zhong X,Cong Y,et al.Genome-wide Analysis of Phosphoenolpyruvate Carboxylase Gene Family and Their Responseto Abiotic Stresses in Soybean[J].Sci Rep,2016(6):38448[28] Shanlin Y U,Pan L,Yang Q,et al.Identification and expression analysis of the phosphoenolpyruvate carboxylase gene family inpeanut(Arachis hypogaea L.)[J].Agricultural Sciences in China,2010,9(4):477-487[29] 涂嘉琦,甘璐,冯兰兰,等.蔓花生PEPC基因家族的生物信息学分析[J].热带亚热带植物学报,2018,26(2):107-115[30] 马海洋,赵秋芳,陈曙,等.菠萝PEPC基因家族的生物信息学分析[J].热带作物学报,2020,41(1):97-103[31] 邵姁,王月,应炎标,等.蓝莓磷酸烯醇式丙酮酸羧化酶基因cDNA克隆及表达分析[J].基因组学与应用生物学,2016,35(1):166-171[32] 赵晋锋,王高鸿,杜艳伟,等.谷子磷酸烯醇式丙酮酸羧化酶基因(PEPC)对逆境胁迫的响应[J].华北农学报,2019,34(4):67-74[33] Sánchez R,Flores A,Cejudo F J.Arabidopsis phosphoenolpyruvate carboxylase genes encode immunologically unrelatedpolypeptides and are differentially expressed in response to drought and salt stress[J].Planta,2006,223(5):901-909 [34] Doubnerová V,Rylavá H.What can enzymes of C4 photosynthesis do for C3 plants under stress?[J].Plant science:an internationaljournal of experimental plant biology,2011,180(4):575-583[35] Seki M,Umezawa T,Urano K,et al.Regulatory metabolic networks in drought stress responses[J].Current opinion in plantbiology,2007,10(3):296-302[36] Ma J F,Furukawa J.Recent progress in the research of external Al detoxification in higher plants:a minireview[J].Journal ofinorganic biochemistry,2003,97(1):46-51[37] Gregory A L,Hurley B A,Tran H T,et al.In vivo regulatory phosphorylation of the phosphoenolpyruvate carboxylase AtPPC1 inphosphate-starved Arabidopsis thaliana[J].The biochemical journal,2009(1):57-65[38] López-Millán A F,Morales F,Andaluz S,et al.Responses of sugar beet roots to iron deficiency.Changes in carbon assimilationand oxygen use[J].Plant physiology,2000(2):885-898[39] Rangel A F,Rao I M,Braun H P,et al.Aluminum resistance in common bean(Phaseolus vulgaris)involves induction andmaintenance of citrate exudation from root apices[J].Physiologia plantarum,2010(2):176-190[40] Ryslava H,Muller K,Semoradova S,et al.Photosynthesis and activity of phosphoenolpyruvate carboxylase in Nicotiana tabacum L.Leaves infected by potato virus A and potato virus Y[J].Photosynthetica,2003,41(3):357-363[41] Müller K,Doubnerová V,Synková H,et al.Regulation of phosphoenolpyruvate carboxylase in PVY(NTN)-infected tobaccoplants[J].Biological chemistry,2009,390(3):245-251[42] Liu X,Li X,Zhang C,et al.Phosphoenolpyruvate carboxylase regulation in C4-PEPC-expressing transgenic rice during earlyresponses to drought stress[J].Physiologia plantarum,2017(2):178-200[43] 陈绪清,张晓东,梁荣奇,等.玉米C4型PEPC基因的分子克隆及其在小麦的转基因研究[J].科学通报,2004(19):1976-1982[44] 马宏敏.转PEPC基因籼稻恢复系“N175”产量相关性状分析[J].福建农业科技,2014(8):1-2[45] 张桂芳,丁在松,赵明.稗草(Echinochloacrusgalli)根型PPC基因对水稻的遗传转化及其对光合速率的调节效应[J].作物学报,2015,41(3):507-513[46] 尹吴,李丽莎,王立科,等.转玉米PEPC基因杨树的光合生理特性分析[J].林业科学,2012,48(6):63-71[47] Chen M,Tang Y,Zhang J,et al.RNA interference-based suppression of phosphoenolpyruvate carboxylase results in susceptibilityof rapeseed to osmotic stress[J].Journal of integrative plant biology,2010,52(6):585-592[48] Rademacher T,Häusler R E,Hirsch H J,et al.An engineered phosphoenolpyruvate carboxylase redirects carbon and nitrogenflow in transgenic potato plants[J].The plant journal:for cell and molecular biology,2002(1):25-39[49] Vidal J,Chollet R.Regulatory phosphorylation of C4 PEP carboxylase[J].Trends in Plant Science,1997,2(6):230-237[50] Kai Y,Matsumura H,Inoue T,et al.Three-dimensional structure of phosphoenolpyruvate carboxylase:a proposed mechanismfor allosteric inhibition[J].Proceedings of the National Academy of Sciences of the United States of America,1999,96(3):823-828[51] Matsumura H,Xie Y,Shirakata S,et al.Crystal structures of C4 form maize and quaternary complex of E.coli phosphoenolpyruvatecarboxylases[J].Structure,2002,10(12):1721-1730[52] Tovar-Méndez A,Mújica-Jiménez C,Muñoz-Clares R A.Physiological implications of the kinetics of maize leafphosphoenolpyruvate carboxylase[J].Plant Physiology,2000(1):149-160[53] Izui K,Taguchi M,Morikawa M,et al.Regulation of Escherichia coli phosphoenolpyruvate carboxylase by multiple effectors invivo. II. Kinetic studies with a reaction system containing physiological concentrations of ligands[J].Journal of biochemistry,1981(5):1321-1331[54] Tovar-Méndez A,Muñoz-Clares R A.Kinetics of phosphoenolpyruvate carboxylase from Zeamays leaves at high concentration ofsubstrates[J].Biochimica et Biophysica Acta,2001,1546(1):242-252[55] Tovar-Méndez A,Rodríguez-Sotres R,López-Valentín D M,et al.Re-examination of the roles of PEP and Mg2+ in the reactioncatalysed by the phosphorylated and non-phosphorylated forms of phosphoenolpyruvate carboxylase from leaves of Zeamays.Effects of the activators glucose 6-phosphate and glycine[J].Biochemical Journal,1998(3):633-642[56] Frank J,Clarke R J,Vater J,et al.Influence of allosteric effectors on the kinetics and equilibrium binding of phosphoenolpyruvate(PEP)to phosphoenolpyruvate carboxylase(PEPC)from Zea mays[J].Biophysical Chemistry,2001,92(1-2):53-64 [57] Echevarria C,Vidal J.The unique phosphoenolpyruvate carboxylase kinase[J].Plant Physiology and Biochemistry,2003,41(6):541-547[58] Echevarria C,Pacquit V,Bakrim N,et al.The effect of pH on the covalent and metabolic control of C4 phosphoenolpyruvatecarboxylase from Sorghum leaf[J].Archives of Biochemistry and Biophysics,1994,315(2):425-430[59] Takahashi-Terada A,Kotera M,Ohshima K,et al.Maize phosphoenolpyruvate carboxylase.Mutations at the putative bindings ite for glucose 6-phosphate caused desensitization and abolished responsiveness to regulatory phosphorylation[J].The journal of biological chemistry,2005,280(12):11798-11806[60] Wang X,Gowik U,Tang H,et al.Comparative genomic analysis of C4 photosynthetic pathway evolution in grasses[J].GenomeBiology,2009,10(6):447-477[61] Shenton M,Fontaine V,Hartwell J,et al.Distinct patterns of control and expression amongst members of the PEP carboxylasekinase gene family in C4 plants[J].Plant Journal,2006,48(1):45-53[62] Uhrig R G,She Y M,Leach C A,et al.Regulatory monoubiquitination of phosphoenolpyruvate carboxylase in germinating castoroil seeds[J].Journal of Biological Chemistry,2008,283(44):29650-29657[63] Shane M W,Fedosejevs E T,Plaxton W C.Reciprocal control of anaplerotic phosphoenolpyruvate carboxylase by invivo monoubiquitination and phosphorylation in developing proteoid roots of phosphate-deficient harsh hakea[J].Plant Physiology,2013(4):1634-1644[64] Ruiz-Ballesta I,Feria A B,Ni H,et al.In vivomonoubiquitination of anaplerotic phosphoenolpyruvate carboxylase occurs atLys624 in germinating sorghum seeds[J].Journal of Experimental Botany,2014,65(2):443-451[65] Ruiz-Ballesta I,Baena G,Gandullo J,et al.New insights into the post-translational modification of multiple phosphoenolpyruvatecarboxylase isoenzymes by phosphorylation and monoubiquitination during sorghum seed development and germination[J].Journal of experimental botany,2016,67(11):3523-3536[66] Figueroa C M,Feil R,Ishihara H,et al.Trehalose 6-phosphate coordinates organic and amino acid metabolism with carbonavailability[J].The plant journal:for cell and molecular biology,2016(3):410-423[67] Denecke M,Schulz M,Fischer C,et al.Partial purification and characterization of stomatal phosphoenolpyruvate carboxylase fromVicia faba[J].Physiologia Plantarum,1993(1):96-102[68] Law R D,Plaxton W C.Purification and characterization of a novel phosphoenolpyruvate carboxylase from bananafruit[J].Biochemical Journal,1995(3):807-816[69] Nisi P D,Zocchi G.Phosphoenolpyruvate carboxylase in cucumber(Cucumis sativus L.)roots under iron deficiency:activity andkinetic characterization[J].Journal of Experimental Botany,2000,51(352):1903-1909[70] Rao S,Reiskind J,Bowes G.Light regulation of the photosynthetic phosphoenolpyruvate carboxylase(PEPC)in Hydrillaverticillata[J].Plant and Cell Physiology,2006,47(9):1206-1216[71] González M C,Osuna L,Echevarría C,et al.Expression and localization of phosphoenolpyruvate carboxylase in developing andgerminating wheat grains[J].Plant Physiology,1998(4):1249-1258[72] Osuna L,Pierre J N,Gonzalez M C,et al.Evidence for a slow-turnover form of the Ca2+-independent phosphoenolpyruvatecarboxylase kinase in the aleurone-endosperm tissue of germinating barley seeds[J].Plant Physiology,1999(2):511-520 [73] Nhiri M,Bakrim N,Bakrim N,et al.Posttranslational regulation of phosphoenolpyruvate carboxylase during germination ofSorghum seeds:influence of NaCl and l-malate[J].Plant Science,2000,151(1):29-37[74] Feria A B,Alvarez R,Cochereau L,et al.Regulation of phosphoenolpyruvate carboxylase phosphorylation by metabolites andabscisic acid during the development and germination of barley seeds[J].Plant Physiology,2008(2):761-774[75] 王丽媛,张玉,徐明怡,等.植物磷酸烯醇式丙酮酸羧化酶的研究进展[J].国土与自然资源研究,2017(5):86-89。
生物化学考研精解名词解释答案(下)

生化考研精解名词解释答案(下)温馨提示:部分解释不是采自教材,如有疑问,请参考课本!第十章糖代谢(P124-125)1.糖酵解(glycolysis):由10步酶促反应组成的糖分解代谢途径。
通过该途径,一分子葡萄糖转化为两分子丙酮酸,同时净生成两分子A TP和两分子NADH。
2.发酵(fermentation):营养分子(Eg葡萄糖)产能的厌氧降解。
在乙醇发酵中,丙酮酸转化为乙醇和CO2。
3.巴斯德效应(Pasteur effect):氧存在下,酵解速度放慢的现象。
4.底物/无效循环(substrate/futile cycle):一对酶催化的循环反应,该循环通过ATP的水解导致热能的释放。
Eg葡萄糖+A TP=葡萄糖6-磷酸+ADP与葡萄糖6-磷酸+H2O=葡萄糖+P i 反应组成的循环反应,其净反应实际上是ATP+H2O=ADP+Pi。
6.底物水平磷酸化(substrate-level phosphorylation):ADP或某些其它的核苷-5′—二磷酸的磷酸化是通过来自一个非核苷酸底物的磷酰基的转移实现的。
这种磷酸化与电子的传递链无关。
7.糖原分解(glycogenolysis/glycogen breakdown):从糖原解聚生成葡萄糖的细胞内分解过程,由糖原磷酸化酶等催化完成。
8.糖原合成(glycogen synthesis):体内由葡萄糖合成糖原的过程。
主要合成场所为肝和肌肉。
包括UDPG途径和三碳途径。
9.磷酸解作用(phosphorolysis):通过在分子内引入一个无机磷酸,形成磷酸脂键而使原来键断裂的方式。
实际上引入了一个磷酰基。
10.糖异生作用(gluconeogenesis):由简单的非糖前体转变为糖的过程。
糖异生不是糖酵解的简单逆转。
虽然由丙酮酸开始的糖异生利用了糖酵解中的七步近似平衡反应的逆反应,但还必需利用另外四步酵解中不曾出现的酶促反应,绕过酵解过程中不可逆的三个反应。
《生物化学原理》张洪渊主编 课后习题及答案(二)

么所产生的乳酸分子中哪个碳原子将是含 14C 标记的?如果将此肝匀浆通以氧气,则乳酸将
继续被氧化,所含标记碳原子在哪步反应中脱下的 CO2 含 14C?
5.下列物质各 1mol 经完全氧化分解,产生多少 ATP 和 CO2?
(1)棉籽糖;
(2)磷酸二羟丙酮;
(3)丙酮酸;
(4)琥珀酸;
(5)核糖。
6.虽然氧分子并不直接参与 TCA 循环,但该循环的运行必需在有氧的情况下才能发生,为什
1.(1)有机物在生物体内氧化过程中所脱下的氢原子,经过一系列有严格排列顺序的传 递体组成的传递体系进行传递,最终与氧结合生成水,这样的电子或氢原子的传递体系称为 呼吸链或电子传递链。(2)线粒体生物氧化体系中,两类典型的呼吸链都由五类组分组成, 并按一定的顺序定位于线粒体内膜。NADH 呼吸链由 NADH 还原酶(复合体Ⅰ)、泛醌、细胞
行计算得△G=-42090 J·mol-1。
4.根据△E。=氧化电极电位-还原电极电位公式,计算(1)(3)(4)(5)可按箭头所指方向
反应。
5.已知 1 个 NADH 经电子呼吸链可以产生 2.5 个 ATP。 NADH +H++3ADP+3Pi+1/2O2→NAD++3ATP+4H2O,△G。=-217.57 kJ/mol, 成人基础代谢为每
6.在充分供给底物、受体、无机磷及 ADP 的条件下,并在下列情况中肝线粒体的 P/O 值各为
多少(见下表)?
底物
受体
抑制剂
P/O
苹果酸
O2
----------
琥珀酸
O2
----------
琥珀酸
O2
生物化学:名词解释&简答题

名词解释1、等电点(PI):在某一pH的溶液中,氨基酸或蛋白质解离成阳离子和阴离子的趋势或程度相等,成为兼性离子,呈电中性,此时溶液的pH称为该氨基酸或蛋白质的等电点。
2、模体(序)(motif):在蛋白质分子中,有两个或三个具有二级结构的肽段在空间上相互接近,形成一个具有特殊功能的空间结构,称为motif3、结构域(domain):在多肽链上相邻的模序结构紧密联系,形成二个或多个在空间上可以明显区别的局部区域,各自行使其功能。
4、DNA变性:在一定理化因素作用下,DNA双螺旋的空间构象破坏解体,但其一级结构仍完整的现象称变性。
5、DNA复性:变性DNA经过一定处理重新形成双螺旋DNA的过程称复性。
6、Tm值:融解温度,在DNA发生热变性时,紫外光吸收值达到最大值50%时的温度称为融解温度(Tm值)。
在Tm值时,DNA分子内50%的双螺旋结构被解开。
7、Km:米氏常数,单底物反应中酶与底物可逆地生成中间产物和中间产物转化为产物这三个反应的速度常数的综合,是酶的特征性常数之一,其值等于反应速度为最大速度一半时的底物浓度。
8、酶的竞争性抑制作用:有些抑制剂与酶的底物结构相似,可与酶的底物竞争酶的活性中心,从而阻断酶与底物结合形成中间产物。
由于抑制剂与酶的结合是可逆的,抑制程度取决于抑制剂与酶的相对亲和力和与底物浓度的相对比例,这种作用称为竞争性抑制。
9、同工酶(Isoenzyme):具有相同催化作用,但酶分子结构,理化性质和免疫学性质不同的一类酶。
10、糖酵解(glycolysis):在缺氧状况下,葡萄糖或糖原分解为乳酸的过程称为糖酵解。
11、乳酸循环(Cori cycle):在肌肉中葡萄糖经酵解生成乳酸,乳酸经血循环运到肝脏,肝脏又将乳酸异生成葡萄糖,葡萄糖释放释入血液后又被肌肉摄取,这种代谢循环途径称为乳酸循环。
12、三羧酸循环(tricarboxylic acid cycle):由活性二碳化合物(乙酰CoA)与草酰乙酸缩合成柠檬酸开始,以四次脱氢、二次脱羧再生成草酰乙酸完成循环反应过程,成为三羧酸循环,又称kerb 循环和柠檬酸循环。
2013生化试卷附答案
华南农业大学期末考试试卷(A卷)2010学年第一学期考试科目:生物化学B考试类型:(闭卷)考试考试时间:120 分钟学号姓名年级专业一、名词解释(本大题共10 小题,每小题 2 分,共20 分)1、蛋白质等电点(pI):当溶液在某一特定的pH值时,使蛋白质所带的正负电荷恰好相等,即静电荷为零,这时溶液的pH值称为该蛋白质的等电点。
2、结构域:蛋白质多肽链上存在的一些紧密的、相对独立的区域,它们是在超二级结构基础上形成的具有一定功能的结构单位,称为结构域。
3、酶活性中心:酶分子中能直接与底物分子结合,并催化底物化学反应的部位。
4、米氏常数(km值):,是研究酶促反应动力学最重要的常数。
根据米氏方程,其值是当酶促反应速度达到最大反应速度一半时的底物浓度。
它是酶的特征常数,可以表示酶和底物之间的亲和能力,Km值越大,亲和能力越弱,反之亦然。
5、底物水平磷酸化:指在分解代谢过程中,底物因脱氢、脱水等作用而使能量在分子内部重新分布,形成高能磷酸化合物,然后将高能磷酸基团转移到ADP形成ATP的过程。
6、呼吸链:在生物氧化过程中,从代谢物上脱下的氢由一系列传递体依次传递,最后与氧形成水的整个体系7、半不连续复制:DNA复制时,以3’ 5’单链为模板合成的新链是5' 3’方向是连续的,称为前导链;而另一条模板链是5' 3’方向,以它为模板,合成的新链是不连续的DNA 片段,称为冈崎片段,DNA的这种复制方式称为半不连续复制。
8、不对称转录:有两方面含义,一是DNA分子双链上的某一区段,一股链用作模板指引转录,另一股链不转录;其二是模版链并非永远在同一单链上。
9、启动子:RNA聚合酶识别、结合并开始转录所必需的一段DNA序列。
10、逆转录:逆转录是以RNA为模板,在逆转录酶的作用下合成出DNA,而后再以此DNA 为模板,通过转录—翻译的过程,实现遗传物质的生物学功能。
二、写出下列英文缩写代表的中文名称(本大题共5小题,每小题1分,共5 分)SSB:单链结合蛋白NAD+:烟酰胺腺嘌呤二核苷酸(氧化型辅酶I)FAD:黄素腺嘌呤二核苷酸;TPP :焦磷酸硫胺素CoA:辅酶A三、填空题(本大题共12 小题,每空1 分,共30 分)1、本学期实验中测定蛋白质含量的方法是考马斯亮蓝G-250染色法,以小牛血清蛋白为标准蛋白质,该染料与蛋白质结合后变成青蓝色,最大吸收波长是595 nm。
三羧酸循环过程
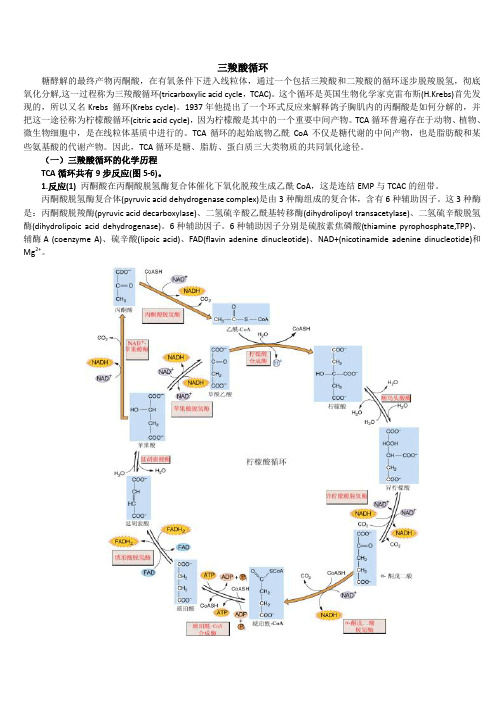
三羧酸循环糖酵解的最终产物丙酮酸,在有氧条件下进入线粒体,通过一个包括三羧酸和二羧酸的循环逐步脱羧脱氢,彻底氧化分解,这一过程称为三羧酸循环(tricarboxylic acid cycle,TCAC)。
这个循环是英国生物化学家克雷布斯(H.Krebs)首先发现的,所以又名Krebs 循环(Krebs cycle)。
1937年他提出了一个环式反应来解释鸽子胸肌内的丙酮酸是如何分解的,并把这一途径称为柠檬酸循环(citric acid cycle),因为柠檬酸是其中的一个重要中间产物。
TCA循环普遍存在于动物、植物、微生物细胞中,是在线粒体基质中进行的。
TCA循环的起始底物乙酰CoA不仅是糖代谢的中间产物,也是脂肪酸和某些氨基酸的代谢产物。
因此,TCA循环是糖、脂肪、蛋白质三大类物质的共同氧化途径。
(一)三羧酸循环的化学历程TCA循环共有9步反应(图5-6)。
1.反应(1)丙酮酸在丙酮酸脱氢酶复合体催化下氧化脱羧生成乙酰CoA,这是连结EMP与TCAC的纽带。
丙酮酸脱氢酶复合体(pyruvic acid dehydrogenase complex)是由3种酶组成的复合体,含有6种辅助因子。
这3种酶是:丙酮酸脱羧酶(pyruvic acid decarboxylase)、二氢硫辛酸乙酰基转移酶(dihydrolipoyl transacetylase)、二氢硫辛酸脱氢酶(dihydrolipoic acid dehydrogenase)。
6种辅助因子。
6种辅助因子分别是硫胺素焦磷酸(thiamine pyrophosphate,TPP)、辅酶A (coenzyme A)、硫辛酸(lipoic acid)、FAD(flavin adenine dinucleotide)、NAD+(nicotinamide adenine dinucleotide)和Mg2+。
图5-6 三羧酸循环的反应过程上述反应中从底物上脱下的氢是经FAD→FADH2传到NAD+再生成NADH+H+。
生化名词解释

一.名词解释1. Tm(解链温度):当核酸分子加热变性时,其在260nm处的紫外吸收会急剧增加,当紫外吸收达到最大变化的半数值时,此时对应的温度称为溶解温度,用Tm表示。
热变性的DNA解链到50%时的温度。
2. 增色效应:DNA变性时,其溶液A260增高的现象。
3. 退火:热变性的DNA经缓慢冷却后即可复性,这一过程称为~。
4. 核酸分子杂交:这种杂化双链可以在不同的DNA单链之间形成,也可以在不同的RNA单链形成,甚至还可以在DNA单链和RNA单链之间形成,这一现象叫做核酸分杂交。
5. DNA复性:当变性条件缓慢去除后,两条解链的互补链可以重新配对,恢复到原来的双螺旋结构。
这一现象称为DNA复性。
6. Chargaff规则:包括 [A] = [T],[G] = [C];不同生物种属的DNA的碱基组成不同;同一个体的不同器官或组织的DNA碱基组成相同。
7. DNA的变性: 在某些理化因素作用下,DNA双链解开成两条单链的过程。
8. 核酸酶:所有可以水解核酸的酶。
9. 糖酵解:在机体缺氧条件下,葡萄糖经一系列酶促反应生成丙酮酸进而还原生成乳酸的过程称为糖酵解(glycol sis),亦称糖的无氧氧化10. 糖异生:是指从非糖化合物转变为葡萄糖或糖原的过程。
11. 丙酮酸羧化支路:糖异生过程中为绕过糖酵解途径中丙酮酸激酶所催化的不可逆反应,丙酮酸需经丙酮酸羧化酶和磷酸烯醇式丙酮酸羧激酶作用而生成丙酮酸的过程称为~。
12. 乳酸循环(Cori循环):肌收缩(尤其是供氧不足时)通过糖酵解生成乳酸。
肌内糖异生活性低,所以乳酸通过细胞膜弥散进入血液后,再入肝,在肝内异生为葡萄糖。
葡萄糖释入血液后又可被肌摄取,这就构成了一个循环,此循环称为~,也称Cori循环。
13. 糖原合成:指由葡萄糖合成糖原的过程。
14. 糖原分解:习惯上指肝糖原分解成为葡萄糖的过程。
15. 血糖:血液中的葡萄糖。
16. 脂肪动员:储存在脂肪细胞中的脂肪,经脂肪酶逐步水解为甘油和脂肪酸,并释放入血供全身组织氧化利用的过程称为脂肪动员。
三羧酸循环过程
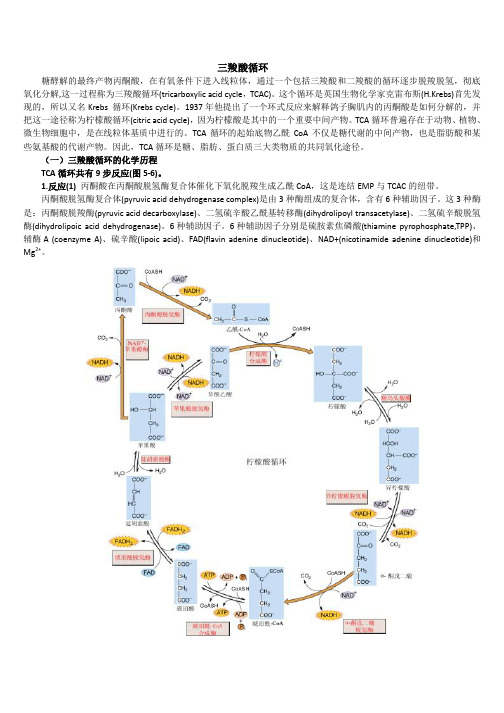
三羧酸循环糖酵解的最终产物丙酮酸,在有氧条件下进入线粒体,通过一个包括三羧酸和二羧酸的循环逐步脱羧脱氢,彻底氧化分解,这一过程称为三羧酸循环(tricarboxylic acid cycle,TCAC)。
这个循环是英国生物化学家克雷布斯(H.Krebs)首先发现的,所以又名Krebs 循环(Krebs cycle)。
1937年他提出了一个环式反应来解释鸽子胸肌内的丙酮酸是如何分解的,并把这一途径称为柠檬酸循环(citric acid cycle),因为柠檬酸是其中的一个重要中间产物。
TCA循环普遍存在于动物、植物、微生物细胞中,是在线粒体基质中进行的。
TCA循环的起始底物乙酰CoA不仅是糖代谢的中间产物,也是脂肪酸和某些氨基酸的代谢产物。
因此,TCA循环是糖、脂肪、蛋白质三大类物质的共同氧化途径。
(一)三羧酸循环的化学历程TCA循环共有9步反应(图5-6)。
1.反应(1)丙酮酸在丙酮酸脱氢酶复合体催化下氧化脱羧生成乙酰CoA,这是连结EMP与TCAC的纽带。
丙酮酸脱氢酶复合体(pyruvic acid dehydrogenase complex)是由3种酶组成的复合体,含有6种辅助因子。
这3种酶是:丙酮酸脱羧酶(pyruvic acid decarboxylase)、二氢硫辛酸乙酰基转移酶(dihydrolipoyl transacetylase)、二氢硫辛酸脱氢酶(dihydrolipoic acid dehydrogenase)。
6种辅助因子。
6种辅助因子分别是硫胺素焦磷酸(thiamine pyrophosphate,TPP)、辅酶A (coenzyme A)、硫辛酸(lipoic acid)、FAD(flavin adenine dinucleotide)、NAD+(nicotinamide adenine dinucleotide)和Mg2+。
图5-6 三羧酸循环的反应过程上述反应中从底物上脱下的氢是经FAD→FADH2传到NAD+再生成NADH+H+。
东北农业大学研究生入学考试生物化学真题部分答案

东北农业大学研究生入学考试生物化学真题部分答案1.核酸的变性:DNA 分子受某些理化因素影响,使分子中氢键、碱基堆积力等被破坏,双螺旋结构解体,分子由双链变单链的过程。
2.蛋白质等电点:对某一蛋白质而言,在某一pH 值时,其所带正负电荷恰好相等,称此pH 值为该蛋白质的等电点。
3.酶原激活:有些酶在生物体内合成的只是它无活性的前体即酶原,酶原在一定条件下能转化成有活性的酶的过程。
4.呼吸链:由供氢体,传递体,受氢体及相应的酶系统组成的生物氧化还原链。
5.糖异生:由非糖物质合成糖原的过程。
6.酮体:丙酮,乙酰乙酸,β-羟丁酸的统称。
7.一碳单位:某些氨基酸在体内进行分解代谢时产生的含一个碳原子(CO 2,CH 4除外)的基团。
8.生酮氨基酸:氨基酸经脱氨代谢产生的酮能生成酮体,乙酰CoA 或乙酰乙酰CoA ,称为生酮氨基酸。
9.生糖氨基酸:代谢后能生成丙酮,α-酮戊二酸,草酰乙酸,琥珀酸的氨基酸,称为生糖氨基酸。
10.半保留复制:DNA 复制时两条互补链分开,每条都按碱基互补配对原则形成新DNA 分子,这样使子代DNA 分子与亲代完全相同,且一条来自亲代,一条是新合成,这种DNA 复制原则称为半保留复制。
11.启动子:RNA 聚合酶识别,结合和开始转录的一段DNA 序列。
12.终止子:转录的终止控制元件,是基因末端一段特殊的序列,它使RNA 聚合酶在模版上移动缓慢,停止RNA 的合成。
13.冈崎片段:DNA 半不连续复制过程中首先以5’~3’为模板合成的较短DNA 片段。
14.多核糖体:在生物体细胞内,一个RNA 分子常与一定数目的单个核糖体结合成念珠状,称多核糖体。
15.必须氨基酸:维持哺乳动物生长所必须且自身无法合成,需从每日膳食中供给以维持机体氮平衡的氨基酸。
16.转氨作用:在转氨酶作用下,一种氨基酸α-氨基转移到另一种酮酸上生成新的氨基酸,原来的氨基酸则转变为α-酮酸.17.诱导契合学说:由于底物诱导而引起酶蛋白空间结构发生某些精细的改变,与适应的底物相互作用,从而形成正确的催化部位,使酶发挥其催化功能。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
歐亞書局
CH 16 檸檬酸循環
p.656
圖 16-5(a)
歐亞書局
CH 16 檸檬酸循環
p.656
圖 16-5(b)
歐亞書局
CH 16 檸檬酸循環
p.656
圖 16-5(c)
歐亞書局
CH 16 檸檬酸循環
p.656
圖 16-5 (續)
圖16-5 丙酮酸去氫酶複合體的結構。(a) 由牛腎臟分離出的 PDH 複合體 的冰凍電子顯微鏡圖。冰凍電子顯微鏡的分析可在極低溫度下視察生物 樣本,如此可避免經由使用脫水、固定及染色過程後可能潛在的人為誤 差。(b) PDH 複合體的三度空間影像,顯示出次單元結構:E1,丙酮酸去 氫酶;E2,二氫硫辛醯轉乙醯酶;以及 E3,二氫硫辛醯去氫酶。影像是 藉由分析大量如 (a) 所示的圖像,結合各次單元結晶圖的研究所重新建構。 核心(綠色)部分包含 60 個 E2 分子,以 20 個三體排列形成五角形十二 面體。E2 的硫辛醯功能區(藍色)向外延伸接觸到排列在 E2 核心上的 E1 分子的活化區(黃色)。
歐亞書局
CH 16 檸檬酸循環
p.654
圖 16-2
圖16-2 丙酮酸去氫酶複合體催化的整個反應。有五個輔酶參與此反應, 與三個酵素組成的酵素複合體,於本文中敘述。
歐亞書局
CH 16 檸輔酶
輔酶 A(圖 16-3)具有反應性巰基(─SH),其對於 在數個代謝反應中以 CoA 作為醯基攜帶者的角色而 言是極具關鍵性的。醯基會與巰基共價連結形成硫酯 (thioesters)。
p.655
丙酮酸去氫酶複合體由三種不同的酵素所組成
PDH 複合體含有三種酵素—丙酮酸去氫酶(pyruvate dehydrogenase;E1)、二氫硫辛醯轉乙醯酶 (dihydrolipoyl transacetylase;E2),以及二氫硫辛醯 去氫酶(dihydrolipoyl dehydrogenase;E3)。
PDH 複合體的第五個輔因子硫辛酸(lipoate)(圖 16-4)具有兩個巰基,能受可逆性的氧化作用成為雙 硫鍵(─S─S─),與蛋白質中兩個半胱胺酸殘基之間 類似。
歐亞書局
CH 16 檸檬酸循環
p.655
圖 16-3
圖16-3 輔酶 A(CoA)。泛酸的羥基藉由磷酸酯鍵連接上修飾的 ADP, 其羧基以醯胺鏈連上 β-巰基乙胺。在 ADP 部分 3’ 位置上的羥基具有自由 態 ADP 所沒有的磷醯基。巰基乙胺的-SH 基與乙醯輔酶 A(左下方)中 的乙酸形成硫酯。
歐亞書局
CH 16 檸檬酸循環
p.656
在受質引導過程,中間產物不會離開酵素表面
圖 16-6 綱要性地描述丙酮酸去氫酶複合體如何在丙 酮酸脫羧及去氫反應中執行五個連續的反應。
在圖 16-6 所顯示的五個反應序列就是一個受質引導 (substrate channeling)的例子。
歐亞書局
CH 16 檸檬酸循環
歐亞書局
CH 16 檸檬酸循環
p.655
圖 16-4
歐亞書局
圖16-4 與離胺酸殘基醯 胺鍵結的硫辛酸。硫辛離 胺酸部分為二氫硫辛醯轉 乙醯酶(PDH 複合體的 E2)的輔基。硫辛醯基具 有氧化(雙硫鍵結)和還 原(雙巰基)型態,且可 作為氫基與乙醯(或其他 醯類)基的攜帶者。
CH 16 檸檬酸循環
(2) 在第二階段中,乙醯基被投入檸檬酸循環,經由酵 素作用將它們氧化成二氧化碳。
(3) 在呼吸作用的第三階段,這些還原性輔酶會自行氧 化而產生質子(H+)和電子。
歐亞書局
CH 16 檸檬酸循環
p.653
圖 16-1
歐亞書局
CH 16 檸檬酸循環
p.654
圖 16-1 (續) e−
歐亞書局
CH 16 檸檬酸循環
歐亞書局
CH 16 檸檬酸循環
p.654
16.1 乙醯輔酶 A(活化態乙酸)的生成
如何經由丙酮酸去氫酶複合體〔pyruvate dehydrogenase(PDH)complex〕氧化而成乙醯輔酶 A 及二氧化碳。
丙酮酸被氧化成乙醯輔酶 A 和二氧化碳
藉由丙酮酸去氫酶複合體催化的整體反應稱為氧化脫 羧反應(oxidative decarboxylation),為不可逆的氧 化步驟,其將丙酮酸上的羧基移開,產生二氧化碳分 子,所剩下的兩個碳會成為乙醯輔酶 A 上的乙醯基( 圖 16-2)。
生物化學
【第5版】
CH16 檸檬酸循環
歐亞書局
16.1 乙醯輔酶 A(活化態乙酸)的 生成
16.2 檸檬酸循環的反應 16.3 檸檬酸循環的調節 16.4 乙醛酸循環
歐亞書局
CH 16 檸檬酸循環
p.653
細胞呼吸經由三個主要階段:
(1)第一階段,有機燃料分子(葡萄糖、脂肪酸和某些胺 基酸)經氧化而產生以乙醯輔酶 A(acetyl-CoA)的 乙醯基型式存在的兩個碳的分子片段。
歐亞書局
CH 16 檸檬酸循環
p.656
圖 16-5 (續)
圖16-5 丙酮酸去氫酶複合體的結構。有些 E3 次單元(紅色)也會結合 上核心,在此 E2 上的搖擺臂可接觸到它們的活化區。星號標示處為硫辛 醯基貼附在 E2 硫辛醯功能區的位置。為使結構表示得更為清楚,約略有 半個複合體由前方移除。此模型由 Z. H. Zhou 等人所製備(2001);另 一個模型則由 J. L. S. Milne 等人所製備(2002),其 E3 所在位置則更接 近酵素周邊(見延伸閱讀)。(c) E2 包含由短多胜肽連接器所連結的三種 型式的功能區:一有催化活性的醯轉移酶功能區;一結合功能區,包含 結合 E2 與 E1 和 E2 與 E3;以及一個或以上(視物種而定)的硫辛醯功能 區。
p.654
圖 16-1 (續)
歐亞書局
CH 16 檸檬酸循環
p.654
圖 16-1 (續)
圖16-1 細胞呼吸作用三階段中的蛋白質、脂肪和醣類的代謝作用。階段 1:脂肪酸、葡萄糖和一些胺基酸氧化產生乙醯輔酶 A。階段 2:檸檬酸 循環中氧化醯基,包括取得電子的四個步驟。階段 3:電子被 NADH 和 FADH2 攜帶注入粒線體(在細菌則為漿膜)電子攜帶者──呼吸鏈──最 後將氧還原為水。此電子流最後會產生 ATP。