病毒学9
病毒学第九章病毒与肿瘤课件

一、肿瘤的发生原因
对整个机体而言,肿瘤形成是由于缺乏有效的免疫防御功能; 对细胞而言,则是正常的细胞周期出现障碍。 1、遗传因素:
日本人的胃癌死亡率比美国人高7倍,美国人的肺癌残废率 又比日本人高2倍多,而苏格兰人又比美国人高2倍。
美国黑人前列腺癌的残废率高于美国白人2倍多。 广东人移居上海10年以后,鼻咽癌的发生率仍高于上海本地 居民2.64倍。食管癌高发区的河南省林县居民,移居山西省黎城 后80-100年,移民的食管癌死亡率仍高于当居民5.7-8.6倍。 2.环境因素:
32
2.基因易位 染色体易位在肿瘤组织中屡见不鲜,基因定
位研究证明,在染色体易位的过程中发生了某些 基因的易位(translocation)和重排,使原来无 活性的原癌基因移至某些强的启动基因或增强子 附近而被活化,因而原癌基因表达增强,导致肿 瘤的发生。
33
3、原癌基因扩增 原癌基因扩增(amplification)是原癌基因数量的增加或表达
40
• 低磷酸化型的P105Rb可与转录因子E2F结合形成复合 物, 从而阻止E2F启动某些在细胞增殖过程中起主要作 用的基因的转录, 如c-myc基因等。
• 高磷酸化型的P105Rb则可促使其与E2F转录因子分离, 从而使其呈现活性, 细胞即由G1期进入S期。
24
一、癌基因的概念 在致瘤病毒、人体和动物肿瘤中发现的可以导致细
胞恶性转化的核酸片断。称为癌基因。根据其来源可以 分为两类。
病毒癌基因、原癌基因(或者细胞癌基因) 无论病毒癌基因还是细胞癌基因被激活后均有诱导 肿瘤发生的作用, 所以有时候我们又将肿瘤细胞中的癌 基因称为肿瘤癌基因。
25
病毒癌基因 ( virus-oncogene;v-onco ) 指病毒核酸能够使细胞恶性转化的片断。
病毒学名词解释

1、感染:病毒生存的基本方式2、毒粒:病毒的细胞外颗粒形式,也是病毒的感染性形式。
一团能自主复制的遗传物质+蛋白质外壳+包膜,有保护和载体作用3、螺旋对称壳体:亚基有规律的沿着中心轴呈螺旋状排列,进而形成高度有序、对称的稳定结构4、核衣壳:病毒的蛋白质壳体和病毒核酸构成的复合物5、裸露毒粒:仅有核衣壳构成的病毒颗粒6、包膜:有些病毒的核衣壳包裹的一层脂蛋白膜,是病毒以出芽方式成熟时,由细胞膜衍生而来的7、刺突:包膜或核衣壳上的突起8、裂解量:每个受感染细胞所产生的子代病毒颗粒的平均数目9、隐蔽期:自病毒在受感染细胞内消失至出现新的感染性病毒的时间10、自外裂解:大量噬菌体在短时间内吸附在同一细胞上,使细胞壁产生许多小孔,引起细胞裂解,但是病毒病没有增殖,这种现象称为自外裂解11、脱壳:病毒侵入后,包膜或壳体除去而释放出病毒核酸的过程12、烈性噬菌体:感染宿主细胞后能在细胞内正常复制并且引起细胞死亡,形成裂解循环13、温和噬菌体/溶原性噬菌体:感染宿主细胞后不能完成复制循环,噬菌体基因组长期存在于宿主细胞内,没有成熟的噬菌体产生。
这一现象称为溶原现象14、免疫性:被温和噬菌体感染后,形成的溶源性细菌具有免疫性,其他同类噬菌体虽然也能感染该细胞,但不能导致该细菌裂解,也不能增殖15、遗传:亲代与子代之间形态结构和生理功能的相似性16、变异:亲代与子代之间、子代的各个个体之间的差异性17、株系:同一病毒的不同系或分离物18、体外诱变剂:对病毒的静态核苷酸进行化学修饰,使其在后面的复制中碱基配对发生改变,导致同型碱基置换或异型碱基颠换,如亚硝酸,羟胺,烷化剂等19、体内诱变剂:作用于代谢活跃的核苷酸,为碱基类似物或插入剂20、遗传重组:重组是混合感染中病毒基因组之间的物理作用21、分子内重组:通过核酸断裂与重接22、重配:多分体病毒在混合感染中,来源于不同亲本病毒的基因组片段可能重新组合23、拷贝选择重组:发生在具有一部分单一分子基因组RNA病毒中,仍属于分子内重组,但不涉及核算分子的共价断裂,RNA聚合酶选择性的连接到静止模板链上进行RNA子代链的合成24、多数感染复活现象:以同一株灭活病毒大量感染敏感细胞时,有时会产生活得感染性病毒25:表型混合:在某些具有共同特点的2种病毒混合侵染中,自带的表型具有双方亲本的特征,而基因型无变化26、转壳现象:混合感染产生的子代病毒,是一种病毒的基因组完全被另一种病毒的外壳蛋白包裹,暂时性27、互补作用:一种病毒的增殖促进另一种病毒的增殖,甚至为另一种病毒增殖所必需28、加强作用:在某些混合感染中,一种病毒可以增强第二中病毒的增殖29、缺陷病毒:在任何条件下都不能独立繁殖,必须依靠另一种病毒提供某些必需物质,后者称辅助病毒,前者称卫星病毒30、中间型杂交:用一个病毒的不同血清型或株系作为杂交的亲本31、窗口期:从人体感染HIV后到外周血血液中能检测出病毒抗体这段时间32:单价噬菌体:只能寄生在一个“变种”的细菌细胞上,甚至只能在一定的菌株上寄生33.多效作用:几个明显不相关的表型效应由单个基因所决定。
病毒学[新版]
![病毒学[新版]](https://img.taocdn.com/s3/m/458f431603020740be1e650e52ea551810a6c995.png)
总论第一章1.病毒颗粒的基本结构如何?画出模式图。
2.病毒的核衣壳的对称型有哪些?各有什么特点?3.阐述病毒核酸及蛋白的特点。
4.如何确定病毒是否有囊膜?其依据何在?5.比较病毒与细菌分类体系的异同。
6.简述病毒分类的机构和标准。
总论第二章1.何谓病毒的MOI及一步生长曲线?二者有何关系?2.解释病毒的复制周期及隐蔽期。
3.举例说明病毒的特异性吸附与细胞的受体的关系。
4.双股DNA、单正股RNA、单负股RNA、反转录病毒的生物合成各有何特点?5.有囊膜的病毒如何成熟与释放?总论第三章1.病毒的基因组变异有哪些类型?发生的机率及后果如何?2.举例说明病毒表型变异的特点及应用价值。
3.什么是缺损型干扰突变?有何意义?4.举例说明分子内重组与重配的结果与意义。
总论第四章1.细胞培养与SPF动物或鸡胚相比,培养病毒各有什么优缺点?2.细胞有哪些类型?用于病毒培养各有何适用范围?3.简述各种细胞培养方法的要点。
4.什么是病毒的生产性感染与非生产性感染?对细胞各有什么影响?5.什么是细胞病变?其表现形式及涉及的细胞结构如何?6.什么是空斑试验?有什么用途?7.阐述包涵体的类型、本质及诊断意义。
8.病毒感染的非杀细胞变化通常引致什么后果?9.介绍干扰素的性质、类型及作用。
各论第一章1.痘病毒的结构与形态有何特征?2.举例说明痘病毒感染的宿主谱的类型。
3.对绵羊致病的有哪些痘病毒?如何鉴别?4.研究鸡痘病毒有何意义?5.试述非洲猪瘟病毒形态、基因组结构及传播方式的特点。
6.阐述疱疹病毒科的分类及共同特点。
7.如何解释伪狂犬病感染猪及其它动物的不同后果?8.鸡马立克病毒的致病特点如何?9.试述腺病毒的结构特点。
10.举例说明腺病毒的致病特点及控制手段。
各论第二章1.细小病毒的基因组及病毒颗粒有何结构特点?2.简述细小病毒对猪致病的特点。
3.犬细小病毒病如何诊断与预防?4.简述圆环病毒的主要特征。
5.说明鸡贫血病毒的致病机理及意义。
病毒学-绪论

1908年Ellemuan和Bang发现通过滤器制备的白血病 无细胞滤液能引起小鸡发病.
1911年P.Rous证实家禽肉瘤也同样能通过宿主 传递,这种病原体是过滤性的,所以认为是病毒, 即病毒能引起肿瘤。
Rous的发现激发了人们对肿瘤本质及病毒与癌 之间的可能关系的研究,他本人也因该项发现于 1966年获得诺贝尔奖。
认为口蹄疫的致病物是很小的,并指出 那些病因未明的天花、牛痘、麻疹、牛瘟可 能是由类似于口蹄疫致病物那样极小的生物 引起的。
绪论——病毒的发现时期
病毒本身研究
1901年W.Reed等在古巴首次鉴定到人过滤性病毒,
即黄热病毒(Yellow fever virus)。
Walter Reed (1851-1902)
病毒学
Virology
第一章 绪论
绪论
病毒发展简史 分子病毒学研究的主要内容 病毒的定义、命名和分类
一、病毒的发展简史
病毒的发展简史
I. 病毒的发现时期 II. 病毒的化学时期 III. 病毒研究的细胞水平时期 IV. 分子病毒学的研究时期
I. 病毒的发现时期
绪论——病毒的发现时期
病毒病的发现历史很悠久
1905年,获得了诺贝尔医学奖
Robert Koch (1843-1910)
绪论——病毒研究的细胞水平时期
①利用大肠杆菌研究噬菌体的感染过程取得了迅 速发展。
②组织培养技术也在这一时期开始应用于动物病 毒的研究。
我国学者黄桢祥早在1943年就利用鸡胚组织块在试 管内进行病毒传代、定量滴定及中和试验。 许多学者采用这一新技术,相继分离了上百种过去 对动物不敏感的新病毒。 1949年J.J.Enders利用单层细胞培养繁殖脊髓灰质 炎病毒取得成功,并且由于他对脊髓灰质炎病毒的 开创性研究,而于1954年获得诺贝尔奖。
动物病毒学课件第9章 单链DNA病毒

➢WUH-1株全长4.8kb
➢进化分析证实属于博卡病毒
NP1 NS1
VP1/2
PBoV-4F
犊牛的腹泻,给养牛业造成了巨大损失
猪博卡样病毒是不是博卡病毒?
由于以前的研究只是通过部分基因片段和进化系 统树分析推测并命名为猪博卡样病毒,没有从全 基因组水平上证实,因此猪博卡样病毒是否属于 博卡病毒还需要进一步验证。
猪博卡样病毒WH-1株的全基因组序列分析
➢根据报道的1.8kb 的序列从临 床上检测到阳性样品,采用简 并引物方法获得了WH-1株的 全基因组序列
✓75℃以上加热处理后,病毒的血凝效价几乎完全 消失,弱酸性到中性的介质适于血凝特性的保持。
3.培养
➢细胞:
✓猪源cell:原代猪肾、猪睾丸细胞 传代细胞系PK-15、IBRS-2
✓人的某些传代cell系:Hela、Hep-2
➢接种方式: ✓同步接种 ✓细胞长成单层前接种
➢病毒的增殖需要正处于有丝分裂期的细胞的某些机 能的辅助。
第九章 单链DNA(ssDNA)病毒
➢9.1 细小病毒科(Parvoviridae) ➢9.2 圆环病毒科(Circoviridae)
9.1 细小病毒科(Parvoviridae)
DNA replication ξ=10-10-10-9 RNA transcription ξ=10-5-10-4 ssDNA 细小病毒 ξ=8×10-4
犬细小病毒
肠炎、心肌炎、白细胞减少
水貂肠炎病毒 白细胞减少、肠炎
水貂阿留申病毒 慢性免疫复合物疾病、脑炎
鹅细小病毒
肝炎、肠炎、心肌炎
猫泛白细胞减少 大脑发育全、白细胞减少、肠炎
症病毒
牛细小病毒
犊牛腹泻
病毒学复习题(附答案)

一、名词解释1. 复制周期2. 顿挫感染3. 缺陷病毒4. 干扰现象5. ts突变株6. CPE7. 包涵体 8. 干扰素 9.抗原性漂移 10.抗原性转变二、简答题1. 病毒有何主要特点?2. 病毒结构由哪几部分组成?各部分主要功能是什么?3. 简述ts突变株获得的条件及实际意义?4.病毒复制周期包括哪几个阶段?5. 举例列出5个与医学有关的DNA病毒科名称?6. 举例列出5个与医学有关的RNA病毒科名称?7. 病毒感染常用的血清学诊断方法有哪些?8.简答干扰素抗病毒的作用特点?三、论述题1.试述病毒性疾病特异性预防措施和制剂。
2.举例说明病毒持续性感染的种类及其特点。
3.病毒感染对宿主细胞的致病作用是什么?4.试述病毒致病机制与细菌致病机制有何不同。
参考答案一、名词解释1.复制周期:病毒增殖是复制,必须在活细胞内增殖。
病毒复制一般分为吸附、穿入、脱壳、生物合成及装配与释放5个阶段,称为病毒的复制周期,经过复制产生大量的子代病毒。
2.顿挫感染:病毒进入细胞内,细胞不提供酶、能量及必要成分,则不能合成病毒本身成分,或虽能合成部分或全部成分,但不能装配与释放。
3.缺陷病毒:病毒基因组不完整或基因位点改变而不能进行正常增殖的病毒。
如丁型肝炎病毒。
4.干扰现象:两种病毒共同感染同一细胞时,可发生一种病毒抑制另一种病毒增殖的现象,称为病毒的干扰现象。
5.ts突变株:就是温度敏感性突变株。
即在28℃~35℃条件下可增殖,而在37℃~40℃条件下则不能增殖。
这是因为ts 变异的基因所编码的蛋白质或酶在较高温度下失去功能,故病毒不能增殖。
6.CPE:病毒在细胞内增殖造成细胞破坏死亡的作用称其为杀细胞效应,这种效应在体外组织培养时可观察到细胞变园、聚集、脱落等,称为致细胞病变作用(CPE)。
主要见于无包膜病毒,如肠道病毒等。
7.包涵体:是某些受病毒感染的细胞内,用普通光学显微镜可见到与正常细胞结构和着色不同的斑块,一般呈园形或椭园形,称为包涵体。
病毒学(一到六章)

病毒学(一到六章)第一章病毒的结构(1学时)病毒:是一类个体极小,结构容易,只含单一核酸(DNA或RNA),必须在活细胞内寄生并以复制方式增殖的非细胞型微生物。
这种病原能通过滤器,命名为病毒。
病毒的基本特性1. 以颗粒形式存在,颗粒很小、以纳米为测量单位,普通都能通过细菌滤器,因此病毒原叫“过滤性病毒”,必须在电子显微镜下才干看见。
2. 没有细胞构造,其主要成分仅为核酸和蛋白质两种,故又称“分子生物”。
3. 病毒只含一种核酸,DNA或RNA。
4. 既无产能酶系,也无蛋白质和核酸合成酶系,只能利用宿主活细胞内现成代谢系统合成自身的核酸和蛋白质成分,是郑重的细胞内寄生微生物,不能自立生长和繁殖。
5. 在离体条件下,能以无生命的生物大分子状态存在,并持久保持其侵染活力。
6. 病毒无核糖体(rRNA)和转移RNA (tRNA)。
7. 病毒不能长大,不经分裂繁殖。
8. 病毒对普通抗生素和作用于微生物代谢途径的药物均不敏感,但对干扰素敏感。
9.有些病毒的核酸还能整合到宿主的基因组中,并诱发埋伏性感染。
第一节病毒的结构特征一、形态1. 病毒的大小是指病毒体的大小。
测量单位是纳米(nanometer,nm),即毫微米(1/1000μm)。
各种病毒的大小相差很大,普通病毒介于50nm~250nm之间,其中绝大多数病毒都在100nm左右;最大的病毒如痘病毒(poxvirus)为300nm,在普通光学显微镜下勉强可看到;最小的病毒如小RNA病毒和极小DNA病毒直径约在20nm~30nm 之间。
2. 病毒的形态病毒的形态多种多样。
绝大多数动物病毒呈球形或近似球形;植物病毒多呈杆第1 页/共18 页状或丝状(某些动物病毒也呈丝状);此外,还有呈砖形(痘病毒)、子弹形(狂犬病病毒);而噬菌体(bacteriophage)多呈蝌蚪形。
有些病毒的形态比较固定,如小RNA病毒呈球形;但某些病毒的形态则是多形性的,如粘病毒(orthomyxoviridae),有球形、丝状和杆状。
病毒学各章总结

病毒与宿主细胞的关系
总结词
病毒与宿主细胞的关系是互利共生或寄生,病毒通过入侵和复制影响宿主细胞的生理功能。
详细描述
在寄生关系中,病毒利用宿主细胞提供的能量和原料复制自身,并释放出新的病毒粒子感染其他细胞 。在这个过程中,病毒会对宿主细胞造成不同程度的损害,导致疾病的发生。了解病毒与宿主细胞的 关系是研究病毒致病机制和治疗方法的理论基础。
转录酶
转录酶是病毒在转录过程中所需的酶,能够催化病毒核酸的转录。
辅助因子
辅助因子是病毒复制过程中所需的辅助因子,如三磷酸腺苷、无机 离子等,对病毒的复制和转录过程起到重要的调节作用。
03 病毒的复制周期
吸附与脱壳
吸附
病毒通过识别并附着在宿主细胞 表面的特异性受体上,实现与宿 主细胞的初步结合。
脱壳
在工业领域,病毒学研究可用于生物 技术和生物工程领域,如基因克隆、 蛋白质表达和病毒作为载体等。
在农业领域,病毒学研究有助于防治 植物病毒病,提高农作物产量和品质。 例如,利用基因工程技术培育抗病毒 转基因作物。
未来病毒学研究的发展方向与挑战
未来病毒学研究将更加注重跨学 科合作,综合运用多种技术手段 研究病毒与宿主相互作用机制。
病毒学是生命科学领域的重要分支,研究病毒的特性、复制机制和与宿主相互作用 等方面的内容。
病毒学的发展推动了生命科学领域其他学科的发展,如分子生物学、遗传学和免疫 学等。
病毒学研究对于理解生命起源、生物进化以及疾病发生机制等方面具有重要意义。
病毒学在医学、农业和工业领域的应用
在医学领域,病毒学研究对于预防、 诊断和治疗病毒性疾病具有重要意义。 例如,疫苗研发和抗病毒药物的研制 等。
病毒的分类与命名
总结词
微生物学 病毒---ZH---v9
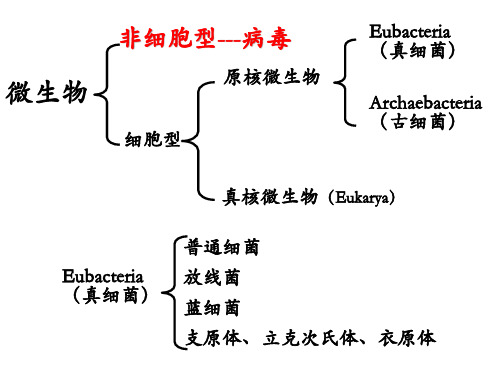
真病毒 至少含核酸和蛋白质二种组分 非细胞生物 类病毒 只含具独立侵染性的RNA组分 亚病毒 拟病毒 只含不具独立侵染性的RNA具有细胞结构,具有一般化学大分子的特征。
Eg. 一些简单的病毒仅由核酸和蛋白质外壳 (coat)构成,故可把它们视为核蛋白分子。
David Baltimore in 2013
七、病毒及其繁殖方式
一切病毒粒并不存在个体的生长过程,只有其两种
基本成分(核酸和蛋白质)的合成和装配。 病毒感染敏感宿主细胞后,病毒核酸进入细胞, 通过其复制与表达产生子代病毒基因组和新的蛋白 质,然后由这些新合成的病毒组分装配(assembly) 成子代毒粒,并以一定方式释放到细胞外。病毒的
(二) 病毒的结构
(三) 病毒的形态 病毒是一类非细胞生物,故称之为病毒 粒或病毒体(virion),有时也称病毒颗粒或病 毒粒子(virus particle),专指成熟的、结构完 整的和有感染性的单个病毒。
毒粒的形状 球形颗粒(或称拟球形颗粒) 杆状颗粒 复杂形状颗粒(如蝌蚪状,卵形)
噬菌体
这种特殊繁殖方式称做复制(replication)。
病毒的繁殖一般分为5个阶段
①吸附
②侵入
③增殖(复制与生物合成)
④成熟(装配)
⑤裂解(释放)
烈性噬菌体所经历的繁殖过程
A typical virus replication cycle
①吸附(adsorption)
病毒吸附蛋白与细胞受体间的结合力来源于空间结构 随机碰撞而接触(静电引力或氢键) 的互补性,相互间的电荷、氢键、疏水性相互作用及 范得华力。 特异性,不可逆吸附,病毒表面蛋白与细胞受体的结 合不同种系的细胞具有不同病毒的细胞受体,病毒受 体的细胞种系特异性决定了病毒的宿主范围。
医学微生物学(第9版)第二篇 病毒学 第二十四章 病毒感染的检查方法与防治原则

第一节
病毒感染的检查方法 diagnosis of viral infection
配套题库请下载 医学猫 APP,执业、三基、规培、主治、卫生资格、正副高等题库都已入库。
医学微生物学(第9版)
一、标本的采集与送检
1. 采急性期标本 2. 抗生素处理易污染标本 3. 低温保存,尽快送检 4. 血清学诊断采双份血清(发病初期、病后 2~3 周)
医学微生物学(第9版)
二、病毒的分离与鉴定
(二)病毒的鉴定
3. 病毒的感染性及数量测定 (3)空斑形成试验 (plaque forming test):
原理:将适量病毒接种单层细胞,病毒增殖使感染的细胞脱落,形成肉眼可见的空斑, 一个空斑即一个空斑形成单位(plaque forming unit,PFU):每个空斑是由一个感染 性病毒颗粒形成的。可测定活病毒数量(感染性病毒量的滴度可用 PFU/ml 表示)
配套题库请下载 医学猫 APP,执业、三基、规培、主治、卫生资格、正副高等题库都已入库。
医学微生物学(第9版)
三、病毒感染的诊断
(一)形态学检查
1. 电镜和免疫电镜检查:病毒形态和大小
(李凡主编的第 8 版医学微生物学 图 27-1,258 页)
配套题库请下载 医学猫 APP,执业、三基、规培、主治、卫生资格、正副高等题库都已入库。
配套题库请下载 医学猫 APP,执业、三基、规培、主治、卫生资格、正副高等题库都已入库。
医学微生物学(第9版)
二、病毒的分离与鉴定
(一)病毒的分离培养
2. 鸡胚培养 (1)孵化 9~14 天的鸡胚 (2)优点:对多种病毒敏感,条件易控制,操作简单,成本较低 (3)缺点:无病毒增殖指标;有些病毒可致鸡胚死亡
病毒学试题(回忆版)

病毒学试题(回忆版)一、名词解释:1.spike2.di(defectiveinterfering)3.replicationintermediate(ri)4.plague(噬斑)5.pfu(噬斑构成单位)、cpe(细胞炎症)、隐密期6、信息体(informosome)7、温度敏感性变异株连8、抗原飘移与抗原转型9准种10、互补11、细胞原癌基因12、干扰素13、ribozyme14、dna疫苗15单克隆抗体16法氏囊,网状内皮非政府细胞分裂症等(传染病英文翻译成中文)二、简答题:1、慢病毒感染的概念及原因(两个方面即病毒方面与机体方面)2、病毒载体活疫苗的概念并举例说明病毒载体构建的技术路线3、亚病毒的分类及其定义4、病毒的特性及其疫苗的研究进展(病毒学)5、病毒的溶解和入侵过程(分后两步)(病毒学)6、请设计试验方案,如何证明某一种病毒的核酸具有感染性(病毒学)7、简述病毒对细胞的损害作用方式(病毒学)8、叙述新病毒出现的机制(病毒学)9、叙述反转录病毒的复制步骤(病毒学)10、存有几个就是写下病毒的英文名称使写下分别属于哪科、哪属于(不常用的几个)11、如果一牛场出现布氏杆菌,恳请制订免疫系统计划(传染病)?12、假如出现烈性传染病如何搞?(传染病)三、论述题1、详细叙述禽流感病毒的基因组结构,并写出每段基因组所编码的蛋白及其每种蛋白的功能(病毒学)2、详细叙述sars病毒与禽流感病毒在分子水平上的异同(传染病)。
2021年农科院考博题动物病毒学一、名词解释1di2antigenticdrift3感染性cdna4核酸疫苗5pfu6ribozyme7ltr8温度敏感突变株9envelope二、简答题1类病毒与朊病毒的区别。
2鉴别新城疫病毒毒力的依据。
3病毒血凝、血凝抑制作用及血吸附机理。
4写出下列各科病毒的一种病毒herpesviridae,adenoviridae,coronaviridae,rhabdoviridae,orthomyxoviridae5黄病毒科存有几个属于,各写下一个病毒。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
《病毒学》论文关于朊病毒的研究年级:2011级姓名:韩蕾学号:20114083012日期:2014年5月摘要:朊病毒是一类不含核酸而仅由构成的可能自我复制并具有感染性的亚病毒因子,是造成人类及其他动物的多种致死性中枢神经系统的慢性退化性疾患的元凶。
它不含核酸,所以其特性、结构和致病机理具有特殊性,并且朊病毒的遗传具有多样性,即造成了朊病毒在不同宿主间的传播障碍。
本文将对朊病毒的疾病防控研究阶段性成就与最新进展进行简单介绍关键字:朊病毒、致病型朊病毒蛋白、细胞型朊病毒蛋白、朊病毒的发现人类在18世纪就发现了朊病毒引起的致死性中枢神经系统的慢性退化性疾患,但却一直无法分离出朊病毒病原。
这因为朊病毒不含有核酸和脂类,只是有侵染性的疏水性蛋白颗粒。
直到美国学者S.B.Prusiner于1982年研究羊瘙痒病时发现,这类传染性海绵状脑病是由一种分子质量为27~30ku的不含核酸的糖蛋白质分子感染所引发的,在一定条件下可形成短杆或纤维状结构,被称为朊病毒(Prion)[1]。
由于发现意义重大,S.B.Prusiner在1997年被授予诺贝尔生理或医学奖。
朊病毒是一种新型的蛋白侵染因子,可导致人和动物的神经元退化变性、脑组织海绵体化、胶质细胞增生及细胞内的朊病毒的自身累计等。
许多致命的哺乳动物中枢神经系统机能退化症均与此病原有关,如人的库鲁病(Kuru)、克雅氏症(CJD)、和动物的羊瘙痒病(Scrapic)、疯牛病(Mad Cow Discasc)麋鹿慢性消耗性疾病(CWD)等[2]。
朊病毒的结构纯化后具有传染性的蛋白被称为朊病毒蛋白(PrP)朊病毒蛋白有2种构型,即致病性朊病毒(PrP Sc)和细胞型朊病毒(PrP C)。
致病型朊病毒蛋白是一种相对分子质量为2.7×10⁴∼3.0×10⁴的蛋白质,具有抗蛋白酶K水解的能力,特异地出现在被感染的脑组织中,是患者脑组织呈现淀粉样空斑。
致病型朊病毒蛋白中含有43%的β-折叠和30%的α-螺旋,在细胞型朊病毒的构象改变过程中,二硫键保持完好,主要的转变部位在90~112位的氨基酸残基[3],这里的α-螺旋转变为β-螺旋,二维结晶结构呈六边形,N-端的β-螺旋位于六角单元内,与位于边上的第二α-螺旋、第三α-螺旋及位于单元间的糖基相对称[4]细胞型朊病毒蛋白相对分子质量3.3×10⁴∼3.5×10⁴,含有1对二硫键和2个N型复合寡糖连。
二硫键和糖基化的残基都在C端。
N端含有22个氨基酸残基组成的信号肽序列,C端含有23个氨基酸残基组成的糖基磷酸肌醇锚受体结合位点(GPT)[5],说明它是一种膜糖蛋白。
朊病毒的致病机理[6]关于朊病毒的致病机理,主要有两种观点。
一个是由于PrP C正常功能的缺失,另一个是由于PrP Sc的过度增殖而产生神经毒性。
除此之外,还有拟病毒假说与联合假说等等。
1.1 PrP C正常功能的缺失PrP C的编码基因在小鼠中位于2号染色体,在人类位于20号染色体。
PrP C在神经元、神经胶质细胞、小胶质细胞、肌细胞、白细胞等多种细胞中表达。
关于PrP C生理功能的研究进展较为缓慢,目前发现其可能在神经系统、T细胞信号转导及核酸代谢等方面发挥一定作用(王小凡,2005)。
1.2 PrP Sc的神经毒性PrP Sc具有潜在的神经毒性,其中PrP106-126称为神经肽,单独这一段小肽也能使在体外培养的神经细胞发生凋亡。
而大量PrP Sc在CNS尤其是在脑内的积累可抑制Cu2+与SOD或其它酶的结合,从而使神经细胞的抗氧化作用下降,PrP Sc还可抑制星形细胞摄入能诱导其增殖的Glu。
此外,细胞内的PrP Sc可能还抑制tau调节的微管蛋白的聚合,导致L-型钙通道发生改变,进而使细胞骨架失去稳定性,最终都可使神经细胞发生凋亡并形成空泡状结构,进而使各种信号传导发生紊乱。
外在表现为自主运动失调、恐惧、生物钟紊乱等症状(乔俊文,2006)。
1.3 酵母菌朊病毒假说[7]Cox(1965)发现在某些酵母菌株内存在一种引起终止密码抑制的成分,表现为显性,且不遵守孟德尔遗传规律,故命名为[PSI+]。
Lacrout发现某些酿酒酵母菌中存在另外一种非孟德尔成分,与[PSI+]具有相似的遗传特性,该成分被命名为[URE3]。
Wickner提出用酵母菌朊病毒假说来解释[PSI+]和[URE3]的遗传行为。
认为[PSI+]和[URE3]可能和朊病毒一样,是由细胞内正常成分Sup35和Ure2P经构型改变而来,因为蛋白质的C端失去其正常功能,所以能产生Sup35和Ure2P突变相同的表现型;失去正常结构和功能的Sup35和Ure2P又可与新的Sup35和Ure2P相互作用,诱导他们发生同样的构型改变,从而使[PSI+]和[URE3]表现出显性且不遵守孟德尔遗传规律。
因此,将[PSI+]和[URE3]称为酵母菌朊病毒(yeast prion)。
与哺乳动物朊病毒不同的是:[PSI+]和[URE3]不在细胞间传播,而是由细胞母代传给子代;也不会导致其存在的细胞发生死亡,而是使其产生新的细胞代谢型。
因此,对于哺乳动物和人类,朊病毒是一种致病因子,对于酵母菌,则可视作一类可遗传的代谢表现型决定因子。
1.4 拟病毒假说Dickinson和Outran首次提出,后由Kimberlin阐述。
拟病毒假说即核蛋白论,认为TSEs 病原因子是拟病毒,由蛋白质和核酸组成。
核酸由宿主酶催化复制,不编码病原因子的蛋白质,但其必需成分与宿主成分(如PrP Sc)紧密结合,并受其保护。
病原因子蛋白质由宿主基因组编码,并形成核酸的外被,但至今未能证实TSEs病原因子有特异性核酸(Bruce,1997)。
1.5 联合学说[8]Weissmann认为感染因子是完全朊病毒(holoprion),它由2种成分组成:一种是分离朊病毒(apoprion),即PrP Sc,由宿主基因组复制,本身就可致病;另一种是协同朊病毒(coprion),即核酸,它决定毒株的株特异性。
这种核酸可能连结于PrP Sc上,也存在于正常宿主细胞中。
PrP Sc单独侵入细胞后,可激活某种细胞核酸,使其呈现协同朊病毒的作用。
协同朊病毒借助细胞正常的聚合酶复制,这一过程由PrP Sc激发,且可能依赖于PrP Sc的存在。
这一假说实际上是朊病毒假说和拟病毒假说的综合,使该病原因子的结构理论更趋完善,颇具吸引力,但目前还没有可靠的试验证据(Hornemann,1997)。
朊病毒的检测[9]双色强荧光目标扫描法:双色强荧光目标扫描法可用来检测极微量的朊病毒。
基本原理是运用共聚焦双色荧光相关分光镜技术,利用致病性PrP Sc在适当条件下自我复制和自发聚集的特点,将重组PrP标记上绿色荧光,PrP Sc与正常构象的PrP结合,通过变构复制出大量异常构象的PrP,由于PrP Sc在一定条件下可自发聚集,从而使PrP Sc周围聚集大量的PrP,使其荧光强度大大增加,同时又加入了能增强试验特异性的标记上红色荧光的特异性单抗,使聚集体又标记上红色荧光。
只有同时发出高强度红、绿荧光的颗粒,才能被检测仪器检测到。
该方法灵敏度高、特异性好,可用于区别其他的朊病毒斑点印迹法(Dot-Blot):斑点印迹法是由Serban等提出,其灵敏度较低,但是操作简单,而且对仪器设备要求低,适合大批量样本的筛查。
基本步骤是将组织样本置于冷的裂解缓冲液中匀浆,然后经离心去除不溶性残渣,将上清液与含SDS样本缓冲液等量混合,混合液点样于干燥的NC膜上,经空气干燥、PK消化、硫氰酸胍处理、封闭,最后进行免疫学检测。
朊病毒的防治致病性朊病毒有顽强的抵抗力[10]:(1)耐高温。
疯牛病的脑组织匀浆经134~138摄氏度持续1小时后,对实验动物仍有感染力。
患瘙痒病羊的脑组织经冷冻干燥后,干热到360摄氏度仍有部分感染力,不经冷冻干燥的脑组织匀浆干热到180摄氏度持续1小时不能完全灭活,要干热到200摄氏度才能完全灭活。
(2)耐福尔马林。
羊瘙痒病的致病性朊病毒蛋白不仅能耐0.35%~0.40%福尔马林数月以上,还发现经10%福尔马林固定的羊瘙痒病脑组织,仍有感染力。
人库鲁病、克雅氏症的脑组织经10%福尔马林固定后也可以感染实验动物。
(3)耐强碱或某些消毒剂。
疯牛病的脑组织能耐受2mol/L的NaOH2小时。
因此人们设想了许多较为合理的防治方法,如通过一种药物与细胞型朊病毒蛋白的结合,稳定细胞型朊病毒蛋白的结构,使细胞型朊病毒蛋白不转变为致病型朊病毒蛋白。
或通过修饰作为分子伴娘的蛋白X的作用,干扰细胞型朊病毒蛋白向致病型朊病毒蛋白转变。
或者寻找一种能破坏致病型朊病毒蛋白结构稳定性的药物。
结语病毒病是继癌症、艾滋病后对人类提出的又一大挑战,到底人们在将来如何对抗病毒病还有待研究,但是对朊病毒的有蛋白质复制蛋白质机理的研究也将带来分子生物学理论上的新突破。
参考文献[1] Coles H. Nobel rewards prion theory after years of heated debate [J]. Nature, 1997,389:529.[2] Prusiner S B. Prion disease and the BSE. crisis [J]. Science.1997,278:245 251[3] Prusiner S B. Novel proteinaceous infectious particles cause Scrapie [J]. Science.1982,216(4542):136 140[4] Detlev Riesner . Biochemistry and structure of PrP c and PrP SC. British Medical Bulletin,203,6:21~33[5] Stahl N, et al. Glycosy linositol phospholipid anchors of the scrapie and cellular prion protein [J]. Biochemistry, 1992,31:5043 5053.[6] Cohen F E, et al. Structural clues to prion replication [J].Science,1994,264:530 531.[7] Clover J R, et al. Self-seeded fibers formed by Sup35, the protein Determinant of [PSP],a heritable prion-like factor of S. cerevisiae[J].Cell1997,89:811 819.[8] Sparrer H E, et al. Evidence for the prion Hypothesis: Induction of the Yeast [PSP] Factor by in vitro-Converted Sup35protein [J].Science,200,289:595 599[9] Sun G, Guo M, Shen A, Mei F, Peng X, Gong R, Guo D, Wu J, Tien P, Xiao G. Bovine PrPC directly interacts with alphaB-crystalline. FEBS Lett, 2005, 579:5419–5424.[10] Tuite M F, Sowing the protein seeds of prion propagation [J].Science,2000,289:556 557.。