分子生物学RNA的生物合成(转录)模板共63页文档
分子生物学RNA的生物合成(转录)模板

6
不对称转录 (asymmetric transcription)
• 转录对DNA链的选择 性称为不对称转录。
不对称转录
structural gene:可以转录出RNA的DNA区段
• 有两方面的含义:
– 在DNA分子双链上某 一区段,一股链用作模 板指引转录,另一股链 不转录 ;
– 模板链并非永远在同一 条单链上。
• 已发现三种RNA聚合酶 分别称为RNA聚合酶Ⅰ、 Ⅱ、Ⅲ,专一性地转录 不同的基因,生成不同 的产物i。
• 鹅膏蕈碱是真核生物 RNA聚合酶的特异性抑 制剂,但敏感性不同。
真核生物RNA聚合酶分子组成
• 真核生物的转录酶由8-12个亚基组成;
• 电泳分析显示:
– 有些亚基是两种酶或者三种酶共有的。
茎环结构使转录终止的机理
• 使RNA聚合酶变构,转录停顿; • 使转录复合物趋于解离,RNA产物释放。
E.coli色氨酸操纵子转录终止信号区的碱基顺序
终止子:
DNA模板上的终止信号或终止序列。 终止子的转录产物RNA可以形成特 殊茎-环结构(发夹)终止转录。 终止信号区的结构特点: 终止信号前的富含GC区后总有富含AT 区跟随。最突出的特点是它的富含GC区 有二重对称性(反转重复序列),使该区 域的转录产物有自身互补性,能形成茎 环(stemloop)或发夹(hairpin)状结 构,使RNA-pol脱落。
22
3. 转录起始前复合物 (pre-initiation complex, PIC)
• 真核生物RNA-pol不与DNA分子直接结合,而 需依靠众多的转录因子。
由RNA-Pol Ⅱ催化转录的PIC
TFⅡF
2个亚基组成:
大亚基有解螺旋酶活性 小亚基与原核生物σ因子同源 。
RNA的生物合成--转录-PowerPointPres
3 启动子( promoter )
DNA上的转录起始序列,称为启动子,有序列保守性, 不仅是转录起始的位置,而且影响转录的活性。
上游
下游
RNA聚合酶结合在启动子上
Pribnow box
注意原核启动子在-35 和-10区域的保守序列
真核转录启动子的保守序列
而真核基因是单顺反子(mono-cistron),其mRNA 的转录比较复 杂, 因为真核通常只转录出一个结构基因,而且其结构基因通常又是断裂 基因, 即是由编码的外显子(exon)和不编码的内含子(intron)间隔 排开组成的。因此, 转录得到的仅仅是mRNA的“毛坯”,称为核不均 RNA(hnRNA),要进行转录后的加工-----------在其5’端加上鸟嘌呤 “帽子”,3’端加上polyA的“尾巴”,再切除内含子,将外显子拼 接,才能成为一个成熟的mRNA。
谢谢大家!
用以转录的单链DNA, 称为模板链, 与复制不同,转录是局部的, 从启动子开始到终止子结束,为一个转录单位;
转录不需要引物;
转录的忠实性相对弱;
转录首先得到RNA前体,然后再进行加工转变为成熟的RNA.
被转录成单个RNA分子的一段DNA 称为一个转录单位(transcript)
2 RNA聚合酶( RNA polymerase )
沉降系数 S
生物大分子在离心场中沉降,受到三种力的影响,它们是离 心力,浮力和摩擦力。物质在单位离心力场下的沉降速度是个定 值,称为沉降系数(sedimentation coefficient) 。
蛋白质、核酸等的沉降系数在1 X 10-13到 200 X 10-13秒 之间。 为方便将10-13秒作为一个单位,称Svedberg单位,用S表示。
RNA的生物合成(转录)

转录终止修饰点
编码链3’端后有 3’-AATAAA--GT-5’序列
转录因子
TF Ⅱ :RNA-pol Ⅱ转录的转录因子
TFⅡD TFⅡA TFⅡB
辨认TATA盒,唯一能结 合TATA盒的蛋白质
稳定TFⅡD结合
解旋酶
ATPase 促进polⅡ结合
蛋白激酶
TFⅡF TFⅡE TFⅡH
结合顺序:
TFⅡD → TFⅡA →TFⅡB → TFⅡF→RNA-pol→ TF ⅡE → TFⅡH
过程
ρ因子与RNA转录产物 ( 3’富含C)结合
ρ因子和RNA聚合酶变构
RNA聚合酶停止转 录
解螺旋酶活性使DNA: RNA杂化双链解离
转录产物释放
不依赖ρ因子的转录终止
特点
终止信号:A=T 、G≡C密集区(DNA分子上) 转录产物:3’端连续U区(mRNA) U=A配对不稳定
终止区RNA发夹样或鼓槌状茎环结构
四膜虫rRNA内含子的二级结构 5´-端核苷酸序列
转录后的加工
原核生物转录与翻译连续进行 往往转录还未完成,翻译已开始,
原核生物中转录生成的mRNA 没有特殊的转录后加工修饰过程
I :主要存在于线粒体、叶绿体及某些低等真核生物 的 rRNA基因;
II:也发现于线粒体、叶绿体,转录产物是mRNA; III:是常见的形成套索结构后剪接,大多数mRNA
基因有此类内含子; IV:是tRNA基因及其初级转录产物中的内含子,剪
接过程需酶及ATP。
核酸内切酶
并接体(U1 SnRNA, U2 SnRNA)与hnRNA内含 子碱基互补结合于5’ ,3’端→套索→ 二次转酯反应切下内含子
mRNA的加工
RNA-polⅡ 的转录产物
分子生物学第十四章_RNA的生物合成

(二)原料
4种三磷酸核苷(NTP): ATP、GTP CTP、UTP
(三)RNA聚合酶 (RNA POLYMERASE)
• 全称: 依赖DNA的RNA聚合酶(DNA-dependent RNA polymerase DDRP)
• 简称: RNA聚合酶( RNA-pol)
• 作用:它以DNA为模板,4种NTP为原料,按 A=U、CG、T=A碱基配对规则,催化合成 5′→3′方向的RNA链。
(6)酶类 DNA聚合酶 RNA聚合酶
(7)配对 A-T,G-C
A-U,T-A,G-C
第四节 真核生物前体RNA的加工和降解
• 真核生物转录产物RNA都要经过一系列改变才能成为有生物活性的RNA 分子。
几种主要的修饰方式:
1. 剪接(splicing)
2. 剪切(cleavage)
3. 修饰(modification)
甲基转移酶
5' m7GpppGp…
帽子结构:
• 功能:
①可能是真核核糖体小亚基识别与结合位 点,从而参与蛋白质合成的启动
②可能阻止5′外切核酸酶对mRNA的降 解,增加mRNA的稳定性
2.加尾(adding tail):
• 这一过程也是细胞核内完成,首先由核酸外切酶切去3'-端一些过剩的核苷 酸,然后再加入polyA。
图18-1 5种E.coli启动序列的共有序列
(五)终止因子(Ρ)
• 作用:协助RNA聚合酶辨认终止点并终止转录
第二节 原核生物的转录的过程
转录是以DNA为模板合成RNA的过程 起始 延长 终止
(一)起始阶段
转录起始需解决两个问题: • RNA聚合酶必须准确地结合在转录模板的起始区域 • DNA双链解开,使其中的一条链作为转录的模板
第十章RNA转录

(三)转录的单向性
生物化学 Biochemistry
• RNA转录合成时,只能向一个方向进行 聚合,所依赖的模板DNA链的方向为 3'→5',而RNA链的合成方向为5'→3'。
生物化学 Biochemistry
(四)有特定的起始和终止位点
• RNA转录合成时,只能以DNA分子中的某一 段作为模板,故存在特定的起始位点和特定的 终止位点。
• 与模板链互补的另一条DNA链称为编码链(coding strand),也称为有意义链或Crick链。
5’ 3’
5’
模板链 编码链
5’ 3’ 5’
生物化学 Biochemistry
编码链 (coding strand)
5 ' 3 '
模板链 (template strand)
3 ' 5 '
生物化学 Biochemistry
原核生物只有一种酶,转录三种RNA(mRNA、rRNA、tRNA) 前体。
生物化学 Biochemistry
E. coli RNA聚合酶组分
亚基 ω
分子量
功能
36512 决定哪些基因被转录
150618 催化磷酸二酯键形成
155613 70263 11000
结合DNA模板 辨认起始点 未知
核 心 酶 ( coreenzym e)
生物化学 Biochemistry
全酶 (转录起始)
+
σ
核心酶 (转录延长)
全酶: 2ββˊ σ五个亚基组成 核心酶: 2ββˊ四个亚基组成 σ因子:与启动子结合,参与基因转录的起始,一旦引发RNA 的合成,就与核心酶分离。
15第15章RNA生物合成转录
二、tRNA的转录后加工
DNA
TGGCNNAGTGC
RNA pol Ⅲ
GGTTCGANNCC
tRNA前体
RNAaseP、 内切酶
tRNA核苷酸转移酶、 连接酶
ATP ADP
(2) (1)
碱基修饰
(1)甲基化
(1)
如:A Am
(2)还原反应 如:U DHU
(3) (4)
(3)核苷内的转位反应 如:U ψ
POL-Ⅱ
TFⅡF
ⅡH ⅡE
ⅡE
ⅡH
TFTⅡPBFOPTALT-TAⅡAFⅡB
ⅡA
CTD- P
PIC组装完成,TFⅡH使CTD磷酸化
4. 拼板理论(piecing theory)
一个真核生物基因的转录需要3至5个转 录因子。转录因子之间互相结合,生成有活 性,有专一性的复合物,再与RNA聚合酶搭 配而有针对性地结合、转录相应的基因。
5-pppG -OH + NTP 5-pppGpN - OH 3 + ppi
转录起始复合物:
RNApol (2) - DNA - pppGpN- OH 3
(二)转录延长
1. 亚基脱落,RNA–pol聚合酶核心酶变构, 与模板结合松弛,沿着DNA模板前移;
2. 在核心酶作用下,NTP不断聚合,RNA链 不断延长。 (NMP) n + NTP (NMP) n+1 + PPi
转录 (transcription)
生物体以DNA为模板合成RNA的过程 。
DNA
转 录
RNA
复制和转录的区别
模板 原料 酶 产物
配对
复制
转录
两股链均复制 模板链转录(不对称转录)
第六讲RNA的生物合成与转录后加工

1、RNA合成的酶学基础
真核和原核细胞内都存在有DDRP,迄今发现的DDRP的有 以下特点:①以DNA为模板: 在DNA的两条多苷酸链中只有其 中一条链作为模板,这条链叫做模板链,又叫反义链。DNA 双链中另一条不做为模板的链叫做编码链,又叫有意义链 ,编码链的的序列与转录本RNA的序列相同,只是在编 码链上的T在转录本RNA为U,由于RNA的转录合成是以DNA的 一条链为模板而进行的,所以这种转录方式又叫做不对称 转录。
指RNA聚合酶在DNA模板上停顿下来不 再前进,转录产物RNA链从转录复合物上 脱落下来。
分类 • 依赖ρ因子的转录终止 • 非依赖ρ因子的转录终止
当前第7页\共有94页\编于星期五\23点
编码链
模板链
5´……G C A 3´…… c g
G
t
T
c
ACAT
atg
G t
T a
Cc…a…g3´……5}´
DNA
转录
5´……G C A G U A C A U G U C……3´
mRNA 翻译
N …… Ala · Val · His · Val ……C
肽
RNA聚合酶的组成
亚基组成:2'w = 2'w +
全酶 核心酶
亚基 —— 解开前方的DNA双螺旋、恢复后面 的DNA双螺旋
亚基 —— 催化磷酸二酯键的形成 '亚基 —— 与DNA的非模板链结合
当前第16页\共有94页\编于星期五\23点
亚基:识别DNA上转录的起始部位,从而 引导全酶结合上去
核心酶:解开前方的DNA双螺旋、RNA链的 延伸、恢复后面的DNA双螺旋
大小可能是决定启动子强度的因素之一。
第12章RNA的生物合成-转录final

第12章RNA的⽣物合成-转录final转录泡的形成转录过程动画帽⼦结构mRNA前体的加⼯动画剪接作⽤即在细胞核内,剪切hnRNA 中内含⼦序列,将各外显⼦序列连接为成熟mRNA的过程(结构基因)(内含⼦、外显⼦)(只有外显⼦)mRNAExon1Exon2Exon3Intron1Intron2蛋⽩质DNA成熟mRNADNA转录hnRNA加⼯mRNA剪接去除内含⼦,连接外显⼦5’末端加帽(5’cap)3’末端加尾甲基化修饰及核苷酸编辑(⼀)真核⽣物mRNA前体的加⼯(3’polyAtail)5''末端帽⼦:转录后加⼯中加⼊的m7GpppNmp-7-甲基鸟嘌呤核苷三磷酸保护新合成的mRNA免被降解增加翻译活性协助mRNA转移⾄胞浆参与第⼀个内含⼦的剪接3’末端加多聚A“尾”:增加mRNA的稳定性加强翻译活性参与最后⼀个内含⼦的剪接RNA+nATPRNA(AMP)n+nPPi多聚A 聚合酶功能:第⼗⼀章RNA的⽣物合成概述DNA和RNA异同点D-2脱氧核糖(dR)胸腺嘧啶(T)胞嘧啶(C)鸟嘌呤(G)腺嘌呤(A)磷酸DNAD-核糖戊糖尿嘧啶(U)碱基胞嘧啶(C)鸟嘌呤(G)腺嘌呤(A)磷酸磷酸RNARNA⽣物合成转录(Transcription):DNA指导的RNA合成反转录DNA复制RNA复制蛋⽩质转录翻译RNA复制(RNAreplication):RNA指导的RNA合成DNA转录RNA前体加⼯成熟RNA DNARNA转录体系:DNA模板DNA指导的RNA聚合酶转录过程起始阶段、链的延长、链的终⽌转录后的加⼯过程信使RNA(mRNA)的加⼯转运RNA(tRNA)的加⼯核蛋⽩体RNA(rRNA)的加⼯本章主要内容转录(transcription)⽣物体以DNA为模板合成RNA的过程。
转录RNADNARNA转录体系TemplatesandEnzymes第⼀节1.转录作⽤的特点部位:细胞核内DNA-AGCT碱基配对规律:RNA-UCGARNA核苷酸连接⽅式:3’,5’-磷酸⼆酯键⽅向:5’?3’,合成过程是连续的,不需引物⽅式:不对称转录(asymmetrictranscription)DNA⽚段转录时,双链DNA中只有⼀条链作为转录的模板,这种转录⽅式称作不对称转录。
分子生物学RNA转录
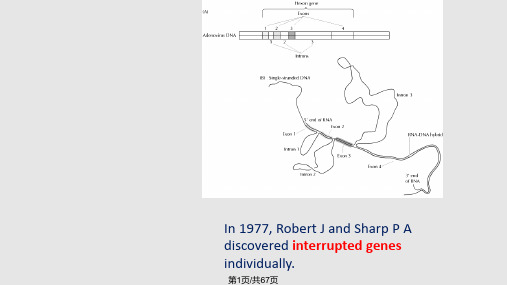
四种内含子的边界序列各有一定共同的特征
第6页/共67页
3、拼接方式 Class I:由拼接复合物完成(核mRNA内含子) 可供识别的特异序列 拼接复合物由多种蛋白质和snRNA组成
Class II:自我拼接(GroupⅠ、Ⅱ ) 形成特定的二级结构 RNA具有催化拼接的能力
Class III:需要蛋白质酶参与的拼接(酵母tRNA) 前两种拼接都属于转酯反应
lariat
第29页/共67页
第30页/共67页
第31页/共67页
第32页/共67页
Trans-splicing
反式剪接的情况较为稀少,较典型的例子是锥 虫 表 面 糖 蛋 白 基 因 VSG(variable surface glycoprotein), 线 虫 的 肌 动 蛋 白 基 因 ( actin genes), 和 衣 藻 ( chlamydomonas) 叶 绿 体 DNA中含有的psa基因。
第46页/共67页
2. Many genes have apparently arisen during evolution in part via exon duplication and divergence.
第47页/共67页
3. Related exons are sometimes found in otherwise unrelated genes.
1. Major and minor splicesome
第38页/共67页
2. nonsense-mediated decay
第39页/共67页
3. A novel strategy used by Dscam Exon 6
Drosophila Dscam gene- encodes cell surface proteins of the Ig superfamily