脊索动物的起源和进化
第二十一章脊索动物的起源和进化

附一、脊索动物的起源和进化
六、哺乳类的起源和演化
• 哺乳动物起源于距今2.3亿年前的中生代三叠纪原始的古 爬行动物杯龙类。在石炭纪末期,由其发展出一支似 哺乳类的兽形爬行类,即盘龙类。由它进化出一支较进
步的兽孔类,兽孔类后裔中的一支称兽齿类(被认 为是哺乳类的祖先)。
• 这些动物属于合颞窝类型。它的基本特征都与哺乳类相似, 但下颌除齿骨外,还有其它骨片,所以还属爬行类。 • 最早的兽齿类化石发现于南非三叠纪地层的犬颌兽。我 国云南禄丰的卞氏兽发现于晚三叠纪地层。 • 中生代白垩纪出现了有袋类和早期的有胎盘类,白垩纪末 期(6500万年),古爬行动物逐渐灭绝,哺乳动物兴起并 一直进入新生代。 • 最早的哺乳动物是小型食虫类,从这一主干向各种生态环 境辐射发展,形成了单孔类、有袋类和真兽类。
• 我国近年发现大量早白垩纪具齿鸟类化石,如辽阳中国鸟;玉门甘 肃鸟;辽阳华夏鸟;2002.7中央台报道我国发现热河鸟化石。鸟类 在白垩纪得到了很大的发展,但有牙齿,新生代则与现代鸟类无 明显差别。 • 鸟类起源的两种假说:树栖起源说;奔跑起源说
除始祖鸟外,考古学家查特基(S.Chatterjee)于 1986年从1983年采到的标本中发现了二件原始鸟类的标本, 这2个标本均采自美国得克萨斯的加扎(Garza)县的晚三 叠纪(距今约 2.25亿年前)地层的泥岩中,定名为原鸟 (有人译为新鸟)原鸟大小似鸡,大的个体约长60cm, 似始祖鸟,小的个体长约30cm。
附一、脊索动物的起源和进化
二、圆口纲动物的起源和演化
• 最早的圆口纲动物化石(也就是脊椎动 物的化石)是
甲胄鱼(距今约5亿年
前的古生代奥陶纪);
• 甲胄鱼是适应于向底栖生活发展的一支, 而圆口类是适应于寄生或半寄生生活的 一支;这两类不一定有直接的亲缘关系, 可能来自共同的无颌类祖先。 • 根据甲胄鱼生活于淡水中,推断脊椎动 物起源于淡水。
《脊索动物门》课件 (2)

人类活动导致了脊索动物的栖息地遭到破坏,需要保护和给予更多关注。
鱼类和两栖类的特点
类别 鱼类 两栖类
特点 有鱼鳞和鳍,以腮呼吸。 有蹼和肺,以肺和皮肤呼吸。
哺乳动物的生物学特性和行为
哺乳方式
所有的哺乳动物在幼崽期都以产奶为主要食物。
行为特征
各种哺乳动物会展现出各自不同的行为特征, 如狮子的社交行为和蝙蝠的群居行为。
《脊索动物门》PPT课件 (2)
通过本课件,您将会了解到关于脊索动物门的演化历史、分类、结构、特征 及其对人类的价值和生态环境的影响。
脊索动物门的起源和演化历史
1
古生代
刺胞动物演化为打孔动物,形成了脊索动物的最早形态。
2
中生代
爬行动物开始出现,脊椎动物进化出高级的内脏器官和复杂的神经系统。
3
新生代
在母体内形成胎盘,幼崽通过 胎盘吸收营养。
人类的分类和演化史
1
古人类时期
露西、直立人、尼安德特人等古人类遗留下的化石。
2
早期人类时期
智人创造工具、掌握语言。
3
中期人类时期
农业革命、文明诞生,科技不断进步。
脊索动物门对生态环境的适应及保护 问题
1 生态适应
通过进化,脊索动物与其生态环境适应得越来越好。
哺乳动物逐渐占主导地位,人类的祖先也从猴子和猩猩中演化而来。
脊索动物门的分类及特点
鱼类
有鳞和鳍,以腮呼吸。
爬行动物
有鳞和角质化皮肤,大多是肺呼吸。
两栖类
有蹼和肺,以肺和皮肤呼吸。
鸟类
有羽毛和趾爪,有嘴和肺呼吸。
哺乳动物的起源和分类
嘬奶动物
下颚有尖锐的牙齿,有产奶能 力。有袋类动物胎盘哺乳源自物幼崽会在母亲的育儿袋中生长。
脊索动物门

脊索动物门——最高等的动物门类一、进化地位1.是所有动物中最高等的类群,包括尾索动物、头索动物和脊椎动物2.脊索动物与棘皮动物和半索动物具有共同的祖先:胚胎发育半索动物与棘皮动物相似,半索动物与脊索动物都有咽鳃裂和神经,三者都是后口动物二、生物学特征1.具有脊索、背神经管和鳃裂三大特征;以及具肛后尾、闭管式循环系统、心脏位于身体腹面等次要特征2.脊索在低等脊索动物中终生保留,在脊椎动物中被脊柱所代替。
背神经管在脊椎动物分化为脑和脊髓。
低等脊索动物鳃裂为呼吸器官,进化中消失或演变为其他结构三、名词解释1.神经胚:背部沿中线的外胚层细胞下陷,形成神经板;两侧的外胚层形成一对纵褶,并靠拢愈合,形成中空神经管,并进入胚胎内部与表面分离。
原肠背面中央隆起,形成脊索中胚层,并脱离原肠,形成脊索。
2.脊索:消化道和神经管间的一条棒状结构,内部由液泡细胞组成,围以结缔组织鞘,坚韧而有弹性, 具支持功能。
脊索动物的胚胎期均具有脊索,以后或终生保留,或退化并被脊柱代替3.神经管:位于脊索背面的中空管状的中枢神经系统。
背神经管在高等种类前端膨大成脑,脑后的部分形成脊髓4.鳃裂:咽部两侧成对的裂缝,为鳃裂,直接或间接与外界相通。
低等脊索动物及鱼类的鳃裂终生存在,其他脊椎动物仅在胚胎期有鳃裂5.逆行变态:身体前端粘附在其它物体上,尾部连同脊索逐渐萎缩消失,神经管退化成神经节,感觉器官消失,鳃裂数增加,体壁分泌被囊素形成被囊,开始营固着生活。
这种变态失去了一些重要的构造、形体变得更为简单,称为逆行变态或退化变态。
四、意义脊索出现的意义1.支撑躯体2.保护内脏器官3.肌肉的支点4.有助于动物体有效完成定向运动5.头骨、颌、椎管的出现都基于脊索头索动物在动物进化中的意义具脊索动物的3大特征,同时具脊椎动物的特征,如分节的肌肉,典型的脊椎动物式的血液循环模式,分开的背腹根等原始和特化的结构:无头,无成对附肢,无心脏,无集中肾脏,具特化的口器等是脊椎动物的原始类群其身体结构和胚胎发育特点,为脊索动物的起源和演化提供有力证据五、与无脊椎动物的区别1.肛后尾:即尾在肛门后方2.闭管式循环系统3.心脏位于消化道腹面4.后口5.分节的肌节6.生物化学比较六、与部分无脊椎动物的共同点1.后口(棘皮动物)2.三胚层(扁形动物)3.真体腔(软体动物)4.身体分节(环节动物)5.两侧对称(扁形动物)七、起源脊索动物门的起源棘皮动物的短腕幼虫和半索动物幼虫柱头虫在结构上相似,肌肉中既含有精氨酸又有肌酸,表明这两类动物亲缘关系较近,同时也表明与脊索动物较近的亲缘关系一般认为棘皮动物和脊索动物有共同祖先,但目前尚没发现化石脊索动物的祖先可能类似尾索动物的幼体,一类经变态,成体为固着生活,鳃裂作为取食和呼吸器官;另一类幼体期延长并适应新的生活环境,不再变态,出现生殖腺并进行繁殖(即幼体性成熟),进而发展出具有脊索、背神经管和鳃裂的自由运动的脊索动物脊椎动物门的起源已知最早的脊椎动物是无颌类,化石出现在5亿年前美洲和亚洲的海洋沉积中。
脊索的名词解释动物学

脊索的名词解释动物学在动物学中,脊索(notochord)是指一种特殊的解剖结构,存在于脊椎动物的胚胎阶段,并成为形成脊柱的基础。
脊索是脊椎动物与无脊椎动物之间的主要区别之一,它在进化中起到了关键的作用。
本文将对脊索的起源、结构和功能进行探讨,以及脊索在动物进化中的重要意义。
1. 脊索的起源脊索起源于早期脊索动物,是距今约五亿年前发生在海洋中的一次重要进化事件。
在早期的脊索动物中,脊索是一条柔软而弹性的管状结构,有助于维持身体的形状和支撑内部器官。
随着时间的推移,脊索经历了进化和分化,最终演化成为现代脊椎动物身体结构的基础。
2. 脊索的结构脊索是一条位于神经管背侧的背中线结构,它由一系列细胞沿着胚胎纵轴排列而成。
脊索由外胚层细胞发育而来,同时也与内胚层和中胚层的细胞相互作用。
它通常呈圆柱形,由一层外形成上皮细胞外部的非细胞基质分泌而成,这种分泌物主要是胶原蛋白。
3. 脊索的功能脊索在胚胎发育过程中发挥着重要的功能。
首先,脊索起着身体轴向发育的指导作用。
在胚胎发育的早期阶段,脊索通过和其他发育结构的相互作用,决定了后续身体的左右对称性和结构分化。
其次,脊索在支持体形、运动和保护脊髓等方面起着重要作用。
它作为一条柔软的结构,可以为身体提供稳定的支撑,并允许运动的发生。
同时,脊索也在一定程度上保护和支撑脊髓,为神经系统的正常发育提供条件。
4. 脊索在动物进化中的意义脊索的出现和演化是动物界进化的重要里程碑之一。
脊索的出现使得动物重新定位自己在进化树上的位置,从而成为一种特殊的生物类群——脊索动物。
脊柱的形成以及其他与脊索相关的变化,使得动物在进化过程中获得了更大的适应性和生存优势。
例如,脊柱的存在为动物提供了更好的运动能力和保护内脏器官的能力,使得动物能够在各种环境和生态位中生存和繁衍。
此外,在脊索动物的进化中,脊柱的出现也为神经系统的演化提供了条件。
脊索在神经系统的分布和发展中起到了重要的组织支持和引导作用,为神经元的分化和连接提供基础。
脊椎动物的起源与演化

脊椎动物的起源与演化脊椎动物,又称脊索动物,是指体内有一个脊索和脊柱的动物。
脊椎动物包括鱼类、两栖动物、爬行动物、鸟类和哺乳动物等。
它们具有坚硬的脊椎和神经系统,能够进行更复杂的动作和行为。
但脊椎动物的起源与演化仍存在很多的疑问和争议。
起源的谜团脊椎动物的起源一直是古生物学领域的难题。
科学家们一直在寻找最早的脊椎动物,但是由于化石记录的不完整,我们对于脊椎动物的起源仍存在很多疑问和不确定性。
不过,一些最新的研究表明,脊椎动物的起源可能要比我们想象的要早得多。
最早的脊椎动物在过去的几十年中,科学家们一直在寻找脊椎动物的最早化石记录。
最早的脊椎动物化石可追溯到五亿多年前的寒武纪。
这些化石来自于一些海底生物群落,在这些化石中,我们可以看到一些具有脊椎的生物,如海燕尾鱼、寡肋鱼和东方蟾鱼等。
但是,一些最新的研究表明,脊椎动物的起源可能要比这些化石记录的时间更早。
据科学家们的推断,最早的脊椎动物可能要追溯到六亿五千万年前的早寒武世,而这些生物可能是柔软的、无骨的或半硬骨的动物。
演化的历程脊椎动物的演化历程非常复杂,其中涉及到很多的生态和环境因素。
在过去的几百万年中,脊椎动物经历了许多的变革和进化,它们从最初的鳗鱼和石首鱼逐渐演化成了现代的哺乳动物和鸟类。
脊椎动物的演化过程中,最显著的变化就是体形和行为的改变。
在早期,脊椎动物主要生活在水中,它们的体型比较长扁,拥有一些适应水环境的特征,如侧线系统和鳃呼吸。
随着时间的推移,一些脊椎动物逐渐进化出了四肢和肺,开始在陆地上生活。
这些动物不再需要通过鳃呼吸来获得氧气,并且发展出了更多适应陆地环境的特征。
同时,它们的智力和行为也逐渐得到了提高,它们开始发展出了社会行为和复杂的思维能力。
结语脊椎动物的起源和演化是一个非常复杂的话题,需要我们综合考虑很多的因素,如环境、生态、遗传和进化等。
虽然我们目前对于脊椎动物的研究还存在很多未知和不确定性,但随着科学技术的不断进步和化石记录的不断完善,相信我们对于脊椎动物的了解会越来越深入和全面。
动物起源与进化

2、趋同进化
发生于2个完全不同的物种或类群,由于 生活在极为相似的环境条件下,经选择 作用而出现相类似的性状,如:蝴蝶翅 和鸟类的翼。
3、平行进化
2个不同类群的动物生活于极为相似的环 境中,具有一些共同的生活习性,而造 成一些对等的器官出现相似的性状或相 似的行为。如:长臂猿和树懒发展相似 的长臂和钩爪。
多结节齿类灭绝,单孔类出现。
现代哺乳动物群在第四纪更新世以后建立 起来。
第二节 人类的起源和发展
一、类人猿的起源和进化 原猴(4000万年前)——类人猿(渐新 世)——埃及古猿(3000万年前)—— 森林古猿(3000万年前)——人、类人 猿
二、人类的起源和进化
1、从猿到人的过渡阶段 森林古猿——拉玛古猿(1200~1500万 年前)——阿法尔南方古猿——LUCY (300~360万年前)
爬行动物的起源与适应辐射
爬行动物的出现是在石炭纪末,造山运动、四 季分明、干燥。
古坚头类
壳椎类 迷齿类
现存两栖类 爬行类
加里冬造山运动
海西运动开始
海西运动高潮
海西运动结束
太平洋运动
古爬行动物化石
最早的爬行类化石叫杯龙类(椎体深凹), 最古的杯龙类叫杯蜥(Cotylosaurus),但 最具过度性特征的叫西蒙龙(Seymouria) 另外一支古爬行动物叫盘龙类(两枕髁,下 颞窝)
2、早期猿人阶段
代表:东非坦桑尼亚能人,1470人 (160~300万年前),能制造工具。
3、猿人阶段;直立猿人(人类)出现在 更新世早期,距今30~180万年前。
4、智人阶段:代表了现代人,分古代类 型(古人,10~30万年前)和现代类型 (新人,3~4万年前)
第三节 动物进化型式和种系发生
第二十一章脊索动物的起源和进化

第二十一章脊索动物的起源和进化第一节脊索动物的起源与演化一、原索动物的起源进化1.“环节动物说”,少有说服力。
2.“棘皮动物说”,认为脊索动物育棘皮动物有共同的祖先。
3. 脊索动物假想的原始祖先,一般公认的最好的解释是由英国科学家加斯坦1928年提出的脊索动物幼体进化假说。
……尾海鞘纲动物可作为幼体性成熟的实例。
4.文昌鱼虽然具有脊索动物的三大主要特征,但一般学者认为文昌鱼不能代表脊椎动物的祖先。
沙隐虫可作为联系原索动物和脊椎动物的代表。
……二、圆口纲的起源和演化1.距今5.25亿年前的古生代早期的甲胄鱼,是现在已经发现的最早的脊椎动物的化石。
2.脊椎动物起源于淡水的说法为更多的人所接受。
三、鱼类的起源和演化1.一般认为颌的出现和最早的鱼类的进化可以追溯到奥陶纪。
2.第583页,图21-6,鱼类演化系统树,也是假说,有些学者有不同的意见3.有人认为软骨鱼、盾皮鱼和棘鱼类有共同的祖先。
软骨鱼在很早就分为二大线系,即全头类和鲨鳐类。
软骨鱼在进化中相当保守,变化不大。
4.硬骨鱼的3大类群似乎是在距今4.5-4.25亿年间从具颌和偶鳍的共同祖先进化而来(1)肺鱼类是一群种类不多的特化淡水鱼。
(2)古总鳍鱼类具有内鼻孔、能作肺用的鳔和能上陆地运动的肉叶状偶鳍……(3)辐鳍鱼类由泥盆纪进化至今大致经历了3个相互继承的阶段……5.四亿年前的泥盆纪世界各地海陆分布发生了巨大变化,通过自然选择,产生了2种适应方式使得鱼类保存下来。
一是由陆地淡水水域迁居海中;二是体内长出能呼吸空气的肺脏。
……四、两栖类的起源和演化1。
两栖类的起源最早可追溯到距今3.5-4亿年前的泥盆纪。
但两栖类究竟起源于哪类动物学者们有不同的意见。
2。
最早的两栖类化石是鱼头螈(或称鱼石螈),生活在距今3.45亿年前的格陵兰地区。
3。
就已有资料,较普遍的看法是鱼头螈分支进化出来的古生代两栖类,可统称为坚头类,在石炭纪和二叠纪适应辐射出迷齿类和壳椎类。
第十三章-脊索动物门

(7)哺乳纲 体外被毛,恒温,胎生(单孔类除外),哺乳(具
乳腺)。
脊椎动物主要类群特征比较
圆口纲 软骨鱼纲 硬骨鱼纲 两栖纲 爬行纲 鸟纲 哺乳纲 上下颌 无颌类 附肢 鱼形类 有 颌 类 (颌 口类) 胚膜 体温
无羊膜类
变温动物
四足类
羊膜类
恒温动物
包括海鞘等大约 2,000 种海生动物,成体多营固着
生活。 个体从几毫米至 30 cm。幼体具脊索动物 3 大
特征, 但脊索仅限于尾部。 幼体经变态至成体只保留
鳃裂。体外具被囊, 由被囊素构成, 又称被囊动物。
尾索动物分类
{
尾海鞘纲(Appendiculariae)
海鞘纲(Ascidiacea)
(6)羊膜类:在胚胎发育中有羊膜的脊椎动物,包括爬行类,
鸟类,哺乳类 脊索动物起源和进化有多种学说,比较集中的有两种 ,即环节动 物说和棘皮动物说。
4.3 原索动物
——无真正的头和脑,又称无头类。
(1)尾索动物亚门 幼体具上述3大特征,但脊索在 尾部。变态后脊索消失,背神经管退 化成神经节,鳃裂仍存在。成体具被 囊,大多营固着生活。
原索动物
头索动物亚门
脊椎动物
脊椎动物亚门
4.1 分类概述
(1)尾索动物亚门:脊索和N管只存在于幼体 (2)头索动物亚门:脊索和N管纵贯身体全长,终生保留,文昌鱼 (3)脊椎动物亚门:脊索只在胚胎发育中出现,随即为脊柱所代替 圆口纲:鳗形,无颌,无成对附肢 鱼 纲:皮肤被鳞,鳃呼吸,有成对附肢 1室1房 两栖纲:皮肤裸露,幼体鳃呼吸,成体肺呼吸 1室2房
1.脊索动物的进化地位 2.门的生物学特征
3.脊索动物起源
4.门的分类
独特的水沟系统。胚胎发育等方面与其它多细胞动物显著
动物生物学14 脊索动物类群

尾索动物亚门〔Urochordata〕
椎骨:亦称“脊椎骨〞是构成脊柱的单位。根据椎骨前后,两 面的形状,又可分为:双凹〔鱼〕、前凹〔无尾两栖、爬 行〕、后凹〔有尾两栖〕、异凹〔鸟〕、双平〔兽类〕
⑶ 原生的水生种类用鳃呼吸,陆生和次生水生种类 用肺呼吸。
⑷ 除圆口类外,脊椎动物出现了上下颌。 ⑸ 除圆口类外,脊椎动物出现了成对的附肢。 ⑹ 集中的肾脏代替了分节排列的肾管,更有效地排
3.头索动物的地位
1)祖先可能是原始的无头类,与无脊椎动物有共同祖 先;
2)由于适应不同生活方式而演变为两支,一支演变为 原始有头类,导向脊椎动物进化之路,另一支特化 为旁支,演变成头索动物的鳃口科动物;
3)认为头索动物是当前脊椎动物的原始类群,是脊椎 动物的姐妹群。在动物学上占有重要的地位。
脊椎动物亚门
脊索动物类群
最高等的动物门类 ---脊索动物门〔Chordata〕
一、脊索动物门的主要特征: 1.出现了脊索 2.具背神经管 3.出现了咽鳃裂 4.肛后尾 5.脊索动物的心脏等
脊索:由柔软而富有弹性的结缔组织组成,脊索细胞内充满 半液态的胞质,脊索外有1-2层鞘膜组成脊索鞘。
背神经管
•一条位于脊索上方的中空的管状神经索,管内腔为 神经腔。 • 前部膨大成脑,神经腔形成脑室。 • 后部为脊髓,神经腔形成中央管。
不对称。 ⑵脊索:直达体前 ⑶背神经管:脑泡小,管上有眼点; ⑷消化、呼吸:口〔口笠、缘膜、缘膜触手〕——咽
脊索动物的起源和分类【高中生物奥赛讲座专题05】

五、脊索动物门的起源和分类
(一)脊索动物门的起源
关于脊索动物的祖先,比较重要的两个假说: 其一是环节动物论,认为脊索动物起源于环节动物; 其二是棘皮动物论,认为脊索动物起源于棘皮动物。 1. 后口动物 2. 以体腔囊法形成体腔 3. 棘皮动物的幼体——短腕幼虫、半索动物的幼 体——柱头幼虫在形态结构上非常近似 4. 无脊椎动物:仅有精氨酸)↔ 棘皮动物和半索动物:同时含有肌酸和精氨酸↔ 脊索动物:仅具肌酸
脊索动物幼体进化假说: 脊索动物的原始祖先具鳃裂以滤过取食,随 后出现具脊索、背神经管和肛后尾的自由 游泳的幼体阶段(蝌蚪幼体),具幼体性 熟现象。蝌蚪幼体阶段比固着生活的成体 阶段更长,这样,固着生活的成体阶段被 淘汰。具繁殖能力的尾索动物幼体样的蝌 蚪幼体可能发展为脊索动物的祖先
(二)脊索动物Leabharlann 的分类 1.尾索动物亚门 (1) 尾海鞘纲 : 无被囊,也 无围鳃腔。无逆行变态, 终生保持具长尾的幼态状, 故又名幼形纲。代表动物 是住囊虫。
无颌类: 无上下颌的脊椎动物 即 圆口纲
12
一些分类名称的范畴 羊膜类: 胚胎发育中有羊膜的脊椎动物 即 爬行类 鸟 类 哺乳类 无羊膜类:胚胎发育中无羊膜的脊椎动物 即 圆口类 鱼 类 两栖类 变温动物: 体温随环境变化而变化的脊椎动物 即鱼类 两栖类 爬行类 恒温动物: 体温较高相对恒定 不随环境温度的变化而 变化的脊椎动物 即鸟类 哺乳类
(二)脊索动物门的分类 1.尾索动物亚门
(2) 海鞘纲 : 具被囊。 逆行变态。行固着生 活。代表动物是柄海 鞘或菊海鞘。
(二)脊索动物门的分类 1.尾索动物亚门 (3) 樽海鞘纲 : 体呈桶状, 出水孔和入水孔分别开口 于体的前后端。行自由生 活。被囊薄而透明。有世 代交替现象。代表动物是 樽海鞘。
脊索动物发展史

脊索动物的发展—2014年大一下期脊索动物学实验报告总结摘要:脊索动物无疑是从无脊推动物进化来的。
在地球上最早出现的是古代的鱼类,并逐渐进化为原始的两栖类,古两栖类又进化为爬行类,某些爬行类又进化为原始的鸟类和哺乳类.所以脊椎动物进化的大致历程是:原始鱼类→原始两栖类→原始爬行类→原始鸟类和哺乳类。
本文是以实验课上完成从鱼类到两栖类及鸟类与哺乳类动物的解剖实验为基础,论述关于脊椎动物历史过程的综述、概括和发展的主要趋势、方式以及某些基本规律。
关键字:脊索动物两栖类鱼类哺乳类脊索动物门,它构造进步、种类繁多,各大类间的进化关系比较清楚,且最后进化出我们人类自己,因而被人们所重视,并乐于了解它们的进化历程。
脊椎动物分类:圆口纲:无颌,又称无颌类;无成对附肢。
脊索终生存在,并出现雏形脊椎骨。
鱼纲:又分为软骨鱼亚纲硬骨鱼亚纲和辐鳍鱼亚纲前者出现上下颌,体被盾鳞,出现成对的鳍,鳃裂直接开口于体外;后者骨骼一般为硬骨,体被硬鳞、圆鳞或栉鳞,鳃裂不直接开口于体表。
两栖纲:由水上陆的过渡种类,幼体鱼形,以鳃呼吸,成体出现5指(趾)型四肢,皮肤裸露,以肺和皮肤呼吸。
与其他更高等脊椎动物共称为四足类。
爬行纲:完全陆生。
皮肤干燥,被以角质鳞、角龟质骨片或骨板。
肺呼吸。
胚胎发育中出现羊膜,与鸟类、哺乳类共称为羊膜类。
其他各纲脊椎动物称为无羊膜动物。
鸟纲:全身被羽,前肢变为翼,适应空中飞翔生活。
血液循环为完全双循环,恒温,卵生。
哺乳纲:体外被毛,恒温,胎生(单孔类除外),哺乳(具乳腺)。
目前比较占优势的一种说法认为棘皮动物可能是脊索动物的祖先,因为某些棘皮动物幼虫的构造和某些原始脊索动物幼虫的很相似。
可惜的是我们迄今还不知道最早的脊索动物是怎样的。
因为它们是比较细小、简单的动物,不大可能在化石中被保存下来。
不过,我们可以想像,它们可能近似于现生的脊索动物文昌鱼。
文昌鱼虽还没有真正的头和脑子,但已具有脊索。
脊索的背侧是神经管,腹侧是消化管。
脊索动物的起源和进化

答:甲胄鱼类一般在体前部覆盖着盔甲样的外骨骼(例如鳞甲鱼和半环鱼等),和现存的圆口纲动物有许多共同特点:无上下颌,早期类型没有成对的附肢,有鳃笼和单鼻孔,嗅囊与鼻垂体囊相通,两眼间有小的松果体孔,内耳有两个半规管等。说明它们之间有一定的亲缘关系。甲胄鱼是现在已发现的最早的脊椎动物化石,现代生存的圆口纲可能是甲胄鱼类的后裔。甲胄鱼是适应于向底栖生活发展的一支,而圆口类动物是适应半寄生或寄生生活的一支,这两类不一定有直接的亲缘关系,可能是来自共同的无颌类祖先。甲胄鱼到泥盆纪末即告绝灭,现生圆口类(盲鳗与七鳃鳗等)则留存至今,成为脊椎动物中特化的一群过去一般认为海洋是脊椎动物的发源地,但根据最早的脊椎动物化石甲胄鱼所在地层进行岩相、岩性与同层的无脊椎动物化石的综合分析研究,推断5亿年前这些动物都栖息于淡水中。由河、湖移居海洋是在泥盆纪中期之后。这样脊椎动物起源于淡水的说法便为更多的人所接受。
6.试论总鳍鱼类在脊椎动物进化中的地位。
答:古总鳍鱼类具有内鼻孔、能作肺用的鳔和能上陆地运动的肉叶状偶鳍,这就使它们能够爬越干涸的泥洼进入新的水域去生活。有的种类经常这样爬越陆地、呼吸空气,结果发展成为最初的陆生脊椎动物,即两栖动物。总鳍鱼中有一种骨鳞鱼,其脑颅骨片的排列式样、牙齿的类
7.简述两栖类的起源与系统演化。
答:由水上陆,在脊椎动物的进化史上是一次巨大的飞跃。
两栖类动物就是这种由水上陆的过渡类型动物。两栖类的起源可以追索到距今3.5亿年至4亿年前的泥盆纪,在那时候,陆上的气候变得干燥,河流与湖泊周期性的变成污浊的池塘和广阔的泥滩。同时海平面下降,使得一些鱼类只能生活在沿岸边留存的水塘或潮湿的岸边。在如此恶劣的条件下,只有能行气呼吸、具“肺”(鳔)并有较强的偶鳍能在陆上爬行的种类才能更好地适应这种恶劣的环境条件。在泥盆纪的鱼类中只有肺鱼和总鳍鱼能行“肺”呼吸,但肺鱼的偶鳍细弱,为双列式的,不能适应在陆上爬行。只有总鳍鱼类,除具“肺”能行气呼吸外,尚具有类似陆生脊椎动物附肢的偶鳍,具有强壮的肌肉和类似陆生脊椎动物四肢ห้องสมุดไป่ตู้骨骼结构。当然,总鳍鱼的鳍作为陆上活动的运动器官不是很有效的,但终究能使其从一个干涸的池塘爬行到另外有水的池塘。气呼吸可使其在少氧的混浊的池塘和短时间越过陆地而存活。缺乏这些适应能力的鱼类就可能被自然选择所淘汰。因此,由逐渐适应和相对快速的一系列进化改变,具气呼吸与肉鳍的鱼越来越适应陆地生活,最后,某些总鳍鱼类进化成第一个类群的两栖动物,它们的鳍进化为陆生五指型附肢。总鳍鱼有两大类群,即骨鳞鱼与腔棘鱼,现今尚存的矛尾鱼即属腔棘鱼类。腔棘鱼为海生的、是远离进化主干的总鳍鱼类,其偶鳍骨骼以及头骨(特别是颌骨)均趋于退化,而且不具内鼻孔,显然不是两栖类的直接祖先。
脊索动物门
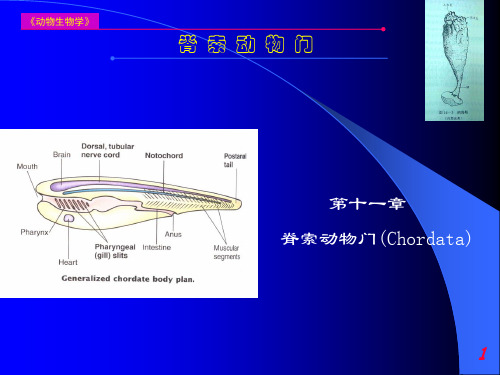
5、神经系统:因固着生活,神经系统退化,仅在入、出水管孔之间有一个神经节(nervus ganglion),没有内腔,圆而坚硬,状如小瘤。其旁(靠出水口)有一无色透明、略为膨大的神经腺(neural gland),相当于高等动物的脑下腺。无专门感觉器官,仅于入水管孔、出水管孔、外套膜上有少量散在的感觉细胞。 6、生殖系统:雌雄同体,生殖腺位于肠环间和外套膜内壁上。精巢大,呈分支状,为乳白色颗粒状小块;卵巢长管状,呈淡黄色,内含有许多圆形的卵细胞,精、卵巢紧贴重叠,分别以单根生殖导管(gonoduct),开口于肛门附近的围鳃腔中。成熟的性细胞输入围鳃腔,然后经出水管孔排出体外,或在围鳃腔内与另一海鞘的生殖细胞相遇受精。精卵生殖细胞不同时成熟,为异体受精。
3、两栖纲(Amphibia):皮肤裸露,湿润,幼体用鳃呼吸,成体用肺呼吸,出现五趾型附肢。 4、爬行纲(Reptilia):皮肤干燥,具角质鳞或骨板,羊膜出现。 5、鸟纲(Aves):体被羽毛(feather),前肢变为翼,恒温,卵生。 6、哺乳纲(Mammalia):身体被毛,恒温,胎生,哺乳。
第一节 脊索动物门的主要特征 脊索动物门(Chordata):是动物界中最高等的一个门,包括全部脊椎动物和一些没有脊椎骨的脊索动物。 主要特征(共同特征、三大特征): 1、脊索(notochord): 是背部起支持体轴作用的一条棒状结构,介于消化道和神经管之间。来源于胚胎发育过程中,由原肠背侧的一部分细胞离开肠管而形成(内胚层)。 A、低等脊索动物终生具有脊索; B、有的种类仅见于幼体; C、高等脊索动物,只有胚胎期间出现脊索,发育完全时,即被分节的脊柱所取代,称这类动物脊椎动物。
动物学课件:_脊索动物

• 发生:发生时,由胚胎背中部的外胚层下陷卷拢
而形成的。
• 演化发展:原索动物不发达,脊椎动物的背神经
管前部膨大形成脑。脑以后的神经管发育成为脊 髓(内腔在脑中形成脑室、在脊髓中成为中央 管)。
第一节 概 述
鳃裂(gill slits)
• 定义:即消化管的前段(咽部)两侧有一系列成 对的裂缝,称鳃裂。直接或间接地外界相通。
胚膜
体温
无羊膜类 变温动物
羊膜类 恒温动物
第一节 概 述
尾索动物亚门(Urochordata):幼体具上述3大特征,但脊索在尾部。 变态后脊索消失,背神经管退化成神经节,鳃裂仍存在。成体具被囊 (tunic),大多营固着生活。分为尾海鞘(Appendiculariae)、海鞘纲 (Ascidiacea)、樽海鞘纲(Thaliacea)。
• 无羊膜类(Anamniotes ):胚胎发育中不出现羊膜的脊椎动物,
包括圆口类、鱼类、两栖类。
• 羊膜类(Amniotes) :胚胎发育中出现羊膜的脊椎动物,包括爬行
类、鸟类、哺乳类。
• 四足类(etrapoda):包括两栖类、爬行类、鸟类和哺乳类。 • 变温动物(Ectotherm):鱼类、两栖类和爬行类。 • 恒温动物(Endotherm):鸟类和哺乳类。
注意点:一切脊索动物都具有脊索,但并不是所有的脊索动物都终 生保留。例:人类,只在胚胎发育早期出现脊索,到了胚胎发育后期 即被脊柱和脑颅基部所取代,因此不终生保留。只有低等脊索动物才 终生具有脊索。
Байду номын сангаас
第一节 概 述
背神经管(dorsal tubular nerve cord)
• 定义:脊索动物的中枢神经系统位于脊索的背面,
12第十四章脊索动物

次要特征:心脏位于消化管腹面,闭管式循 环。多数脊索动物为肛后尾。
二、脊索动物的起源与进化
1、环节动物说:有人认为脊索动物起源于环节动物, 理由是:这二类动物身体都是二侧对称,都分节,又 有发达的体腔和分节的排泄器官。
2、棘皮动物说:认为脊索动物和棘皮动物起源于共 同的祖先。 1)半索动物的成体有接近于脊索动物的 特征,即,具背血管、消化管的前端有鳃裂。 2)柱 头幼虫的形态和短腕幼虫相似。3)海胆和柱头虫的 肌肉中都同时含有肌酸和精氨酸。
(2) 肌肉:具“<”形分 节
(3) 骨骼:脊索,另外 口笠触手、缘膜触手、 轮器内部由角质物支持; 鳍条、鳍、鳃条由结缔 组织支持。
(4) 消化系统:口、咽(围 鳃腔、腹板、背板和围咽 沟)、肠、肝盲囊和肛门。
水流经口入咽,食物被内 柱细胞分泌物粘结成团,纤 毛运动由后向前流经围咽沟 至咽上沟,送入肠。未消化 的物质由肝盲囊重返肠中, 在后肠进行消化吸收。肠未 端开口于身体左侧的肛门。
海鞘的内部解剖
2.消化系统:口、咽(鳃裂、 内柱、背板、围咽沟)、食 道、胃、肠和肛门。
3. 循环方式:具鳃血管和 肠血管,可逆性开管式血液 循环。无专门的排泄器官
4. 神经:退化,有一无内 腔的N节和神经腺。
5. 生殖系统:雌雄同体, 具生殖腺和生殖导管。
二、幼体和变态 逆行变态:由
于幼体与成体的生 活方式不同,经过 很大的变化,使幼 体与成体的结构不 同,结果幼体一些 重要的结构在成体 中消失,成体的结 构反而比幼体简单。 如海鞘。
三. 脊索动物分类概述
尾索动物亚门 头索动物亚门
原索动物(无头类):没有明显的头部结构
脊椎动物亚门:又称有头类
无颌类 有颌类
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
答:最早的脊椎动物是甲胄鱼。现代生存的圆口纲可能是甲胄鱼类的后裔。甲胄鱼是适应于向底栖生活发展的一支,而圆口类动物是适应半寄生或寄生生活的一支,这两类不一定有直接的亲缘关系,可能是来自共同的无颌类祖先。甲胄鱼到泥盆纪末即告绝灭,现生圆口类(盲鳗与七鳃鳗等)则留存至今,成为脊椎动物中特化的一群。过去一般认为海洋是脊椎动物的发源地,但根据最早的脊椎动物化石甲胄鱼所在地层进行岩相、岩性与同层的无脊椎动物化石的综合分析研究,推断5亿年前这些动物都栖息于淡水中。由河、湖移居海洋是在泥盆纪中期之后。这样脊椎动物起源于淡水的说法便为更多的人所接受。
3.试述无颌类在脊椎动物进化中的地位。
答:甲胄鱼类一般在体前部覆盖着盔甲样的外骨骼(例如鳞甲鱼和半环鱼等),和现存的圆口纲动物有许多共同特点:无上下颌,早期类型没有成对的附肢,有鳃笼和单鼻孔,嗅囊与鼻垂体囊相通,两眼间有小的松果体孔,内耳有两个半规管等。说明它们之间有一定的亲缘关系。甲胄鱼是现在已发现的最早的脊椎动物化石,现代生存的圆口纲可能是甲胄鱼类的后裔。甲胄鱼是适应于向底栖生活发展的一支,而圆口类动物是适应半寄生或寄生生活的一支,这两类不一定有直接的亲缘关系,可能是来自共同的无颌类祖先。甲胄鱼到泥盆纪末即告绝灭,现生圆口类(盲鳗与七鳃鳗等)则留存至今,成为脊椎动物中特化的一群过去一般认为海洋是脊椎动物的发源地,但根据最早的脊椎动物化石甲胄鱼所在地层进行岩相、岩性与同层的无脊椎动物化石的综合分析研究,推断5亿年前这些动物都栖息于淡水中。由河、湖移居海洋是在泥盆纪中期之后。这样脊椎动物起源于淡水的说法便为更多的人所接受。
在石炭纪与二叠纪,由于坚头类的大量辐射进化而形成种类繁多的两栖动物,可将它们分为二大类群;迷齿类或称块椎类和壳椎类,鱼头螈即属迷齿类。迷齿类是古生代两栖类的系统演化的主干,其脊椎骨的形成为软骨原骨,椎体由前、后二部分组成,前为间椎体后为侧椎体,这类似于其他陆生脊椎动物,四足脊椎动物是由它演化而来。而壳椎类的则为膜原骨,椎体不分前后部分,而是线轴状,它在系统演化中是一个侧支。
7.简述两栖类的起源与系统演化。
答:由水上陆,在脊椎动物的进化史上是一次巨大的飞跃。
两栖类动物就是这种由水上陆的过渡类型动物。两栖类的起源可以追索到距今3.5亿年至4亿年前的泥盆纪,在那时候,陆上的气候变得干燥,河流与湖泊周期性的变成污浊的池塘和广阔的泥滩。同时海平面下降,使得一些鱼类只能生活在沿岸边留存的水塘或潮湿的岸边。在如此恶劣的条件下,只有能行气呼吸、具“肺”(鳔)并有较强的偶鳍能在陆上爬行的种类才能更好地适应这种恶劣的环境条件。在泥盆纪的鱼类中只有肺鱼和总鳍鱼能行“肺”呼吸,但肺鱼的偶鳍细弱,为双列式的,不能适应在陆上爬行。只有总鳍鱼类,除具“肺”能行气呼吸外,尚具有类似陆生脊椎动物附肢的偶鳍,具有强壮的肌肉和类似陆生脊椎动物四肢的骨骼结构。当然,总鳍鱼的鳍作为陆上活动的运动器官不是很有效的,但终究能使其从一个干涸的池塘爬行到另外有水的池塘。气呼吸可使其在少氧的混浊的池塘和短时间越过陆地而存活。缺乏这些适应能力的鱼类就可能被自然选择所淘汰。因此,由逐渐适应和相对快速的一系列进化改变,具气呼吸与肉鳍的鱼越来越适应陆地生活,最后,某些总鳍鱼类进化成第一个类群的两栖动物,它们的鳍进化为陆生五指型附肢。总鳍鱼有两大类群,即骨鳞鱼与腔棘鱼,现今尚存的矛尾鱼即属腔棘鱼类。腔棘鱼为海生的、是远离进化主干的总鳍鱼类,其偶鳍骨骼以及头骨(特别是颌骨)均趋于退化,而且不具内鼻孔,显然不是两栖类的直接祖先。
软骨鱼在很早就分为二大线系,即全头类与鲨鳐类。鲨鳐类又因适应不同的生活方式而向2个方向演化,即迅速游泳的鲨类和底栖、少活动的鳐类。
软骨鱼在进化中相当保守,有的种类历经约2亿年的时日而很少变化,如现代的噬人鲨与其古代化石种古棘鲨类相比,变化不大,现代的扁鲨和六鳃鲨与其侏罗纪的祖先相比,变化也不大。早期软骨鱼类,采用了第一种适应方式,即由淡水迁入海水,过海洋的生活。这样,虽然得到了丰裕的水环境,但是由淡水转入海水,是由低渗的水环境转入高渗的水环境,入海的软骨鱼首先遇到的问题,就是体内的水分要不断向外渗透出来。能防止这种渗透的种类才能留存后代,这样长期适应的结果,软骨鱼就有了一种调节自身渗透压的能力,能将含氮的代谢废物转变成尿素,以2%一2.5%的浓度保存在血液中,基本上维持了体内外水液调节的平衡,因此能在海中延续生命,繁殖种族。
5.试述辐鳍鱼类的演替关系。
答:辐鳍鱼类由泥盆纪进化至今大致经历了3个相互承替的阶段:最早的辐鳍鱼类为软骨硬鳞类,以古鳕鱼类为代表,出现于泥盆纪的淡水水域,石炭纪达全盛期,到三叠纪为全骨类所替代,全骨类来自软骨硬鳞类,大部分中生代时期为全骨类繁盛时期,在淡水与海水中进行分支进化,现代的全骨类代表有雀鳝与弓鳍鱼。在中生代早期由全骨类发展出真骨类,开始了辐鳍鱼进化的第三阶段,真骨类自中生代后期至今进行了大量的适应辐射,形成各种生态类型。尽管真骨类鱼种类与形态繁多,但大多数鱼类学家相信,各种真骨鱼类是来自全骨类中的一个目。
ห้องสมุดไป่ตู้73楼
4.简述软骨鱼类的起源与演化。
答:由于软骨鱼类化石保留较硬骨鱼少。根据化石鲨与同时代的棘鱼类和盾皮鱼类的解剖结构比较,三者似源自共同祖先。
最早的软骨鱼化石是泥盆纪的一些食肉鲨,如裂口鲨。在泥盆纪时期软骨鱼大量辐射发展,到石炭纪时(距今约2.8亿年至3.5亿年)在淡水与海水中都很普遍,随后许多祖先类群灭绝,代之以现代软骨鱼。
而骨鳞鱼的头骨、脊柱和偶鳍结构等,均表现出与原始两栖类有惊人的相似,因而一直被公认为是四足动物的近亲。
最早的两栖类化石是鱼头螈(或称鱼石螈),生活在距今约3.45亿年前的格陵兰地区。它具有鱼类和两栖类的特点:如有鱼样的身体,体长约1.2m,具带鳍条的尾鳍,具残余的前鳃盖骨;其头骨骨片的数目与排列和总鳍鱼类十分相似,牙也同于总鳍鱼,为迷齿鱼头螈又具一些适应陆地生活的结构,如有具关节的五指型四肢,缩短的头骨,具双枕髁,具耳裂借以支持陆生动物特有的鼓膜。头骨的吻部较长,显示了其嗅觉能力的增强。其脊椎骨具陆生脊椎动物所特有的前、后关节突,肩带与头骨失去联系,使头部有活动余地,所有这些适应陆地生活的进步性特征证明了鱼头螈已步人两栖动物的范畴但仍保留了一些鱼类特征,说明它确实源自鱼类。从鱼头螈分支进化出来的古生代两栖类,由于头骨均有膜原骨形成的硬骨所覆盖,可统称为坚头类。
一个侧支进化为棘皮动物,这从近来发现的一类棘皮动物化石得到更好地证明,它们具一系列类似鲨鱼样的鳃裂,具肛后尾和一个背神经索,它们是一类用鳃裂滤食的动物,十分类似现代的原索动物,另一侧支进化为半索动物;主干进化为脊索动物。并将半索动物与棘皮动物作为从无脊椎动物向脊椎动物演化之过渡类型。
某些具柄的棘皮动物,如已灭绝的棘皮动物中的海果类以及腕足类在这方面都有相似处,也许这些动物间均有亲缘关系,并与早期脊索动物有共同祖先。也许这些动物均各自独立地经适应辐射而形成这些相似性。而它们的循环系统和按节分布的神经系统与肌肉系统均类似于脊椎动物,消化管中的内柱与脊椎动物的甲状腺同源,尤其是其胚胎发育的中胚层体腔囊的形成方式,在前14对体节的形成方式同于棘皮动物与半索动物,14对体节之后的中胚层是从一条独立的细胞带形成,这种方式又与脊椎动物是—致的。另外,文昌鱼的受精卵在卵裂过程中的染色体具明显的双层膜结构,这又与棘皮动物的海胆等相似,而不同于脊椎动物。但文昌鱼又是十分特化的动物,它们的脊索向前超过神经管,按节排列的肾管和生殖腺均与脊椎动物不同。因此,一般动物学者认为文昌鱼类不能代表脊椎动物的祖先。故文昌鱼类或许是脊索动物进化中离开主干的一个侧支,与脊椎动物有共同祖先。
6.试论总鳍鱼类在脊椎动物进化中的地位。
答:古总鳍鱼类具有内鼻孔、能作肺用的鳔和能上陆地运动的肉叶状偶鳍,这就使它们能够爬越干涸的泥洼进入新的水域去生活。有的种类经常这样爬越陆地、呼吸空气,结果发展成为最初的陆生脊椎动物,即两栖动物。总鳍鱼中有一种骨鳞鱼,其脑颅骨片的排列式样、牙齿的类型、偶鳍骨骼的结构等,均表现出与原始两栖类有惊人的相似,因而一直被公认为是四足动物的近亲。另外,根据美遗传学家李特曼等最近对矛尾鱼的基因组构的研究以及对总鳍鱼类进化的分析,他们发现矛尾鱼的免疫球蛋白重链的可变区基因密码群集围绕着其他免疫球蛋白的基因,这样的基因组构正像鲨鱼而不像硬骨鱼和陆栖脊椎动物。但是在胚胎发育时,这些基因又重新连接起来,这又像硬骨鱼和陆栖脊椎动物,而不像鲨鱼。据此,作者认为腔棘鱼可能是软骨鱼与硬骨鱼之间的过渡类型有可能是在4亿年前由鲨鱼或其他的软骨鱼演化来并由它演化为硬骨鱼。此研究结果似乎将硬骨鱼与陆栖脊椎动物也联系在一起,基因组构和胚胎发育的共同点也提供了由总鳍鱼类演化为陆生脊椎动物的线索。两栖类非常近似,因此,可以认为像骨鳞鱼这样的鱼类很可能就是两栖类的直接祖先。
8.简述西蒙龙的特征及其在进化中的地位。
答:蜥螈(或称西蒙龙)具有一系列近于两栖类的特征和似爬行类的特征。蜥螈是体长51cm的蜥蜴形动物,它的头骨形态和结构很像坚头类,颈特别短,肩带紧贴于头骨之后,脊柱分区不明显,具有迷齿和耳缺等,都与古两栖类相似。而且有些与蜥螈亲缘关系很近的化石种类,尚可见侧线管的痕迹。这些都是似两栖类的特征。但是蜥螈还存在着爬行类特征,例如头骨具单个枕骨髁,肩带具有发达的间锁骨,有2枚荐椎,前肢五指(而不似古坚头类以及现存两栖类的四指),各指的骨节数目也比两栖类多(指式为2、3、4、5、3、或4),腰带与四肢骨均较粗壮,更适于陆生爬行。所有这些都指明了它是两栖类与爬行类之间的过渡类型。由于蜥螈发现于下二叠纪,真正的爬行动物化石在石炭纪末已有发现,故蜥螈不是爬行类的直接祖先,其祖先应从石炭纪杯龙类中去寻找,但目前尚不清楚。近年来,根据对中耳的进化的比较研究,提出了两栖类与爬行类在进化上或多或少是以各自独立的线系,均从总鳍鱼类祖先演化而来。但最终如何回答爬行类源自何种动物的问题仍然是动物学家和古生物学家探索的问题之一。
1.试述脊索动物的起源。
答:有人认为脊索动物与棘皮动物有共同祖先。此说根据半索动物的成体有接近于脊索动物的特点,而胚胎发育和幼体形态却和棘皮动物的极为相似,加以对肌肉的肌蛋白生化成分的分析,可以说半索动物,棘皮动物和脊索动物有明显的共同点,均具肌酸,而半索动物与棘皮动物的肌蛋白中除含有肌酸,尚含有精氨酸,无脊椎动物的肌蛋白含精氨酸不含肌酸。故主张半索动物、棘皮动物与脊索动物源自共同祖先,由此共同祖先分为3支演化: