第二章 DNA复制
染色体与DNA分子生物学

H2A
H2B
H4 H3
真核细胞染色体上的组蛋白成分分析
种类
相对分 子质量
氨基酸 分离难 保守性 数目 易度
染色质 中比例
染色质 中位置
H1
21 000
223
易
不保守 0.5
接头
H2A
14 500
129
较难
较保守 1
核心
H2B
13 800
125
较难
较保守 1
核心
H3
15 300
135
最难
最保守 1
核心
• C值( C-value ): 是指一种生物单倍体基因组DNA的总量
各种生物细胞内DNA总量的比较
在真核生物中,C值一般是随生物进化而增加的,高等 生物的C值一般大于低等生物。
C值反常现象 (C-value paradox)
C值往往与种系进化的复杂程度 不一致,某些低等生物却具有较 大的C值。
H4
11 300
102
最难
最保守 1
核心
组蛋白的特性
• 进化上的极端保守性 • 无组织特异性 • 肽链上氨基酸分布的不对称性
碱性氨基酸分布在N端;疏水基团在C端
• 存在较普遍的修饰作用
甲基化、乙酰化、磷酸化、泛素化等
The core histones share a common structural fold
续性; ③能够指导蛋白质的合成,从而控制整
个生命过程; ④能够产生可遗传的变异。
染色体包括: DNA和蛋白质两大部分。
同一物种内每条染色体所带DNA的量是 一定的,但不同染色体或不同物种之间 变化很大。
真核细胞染色体的组成
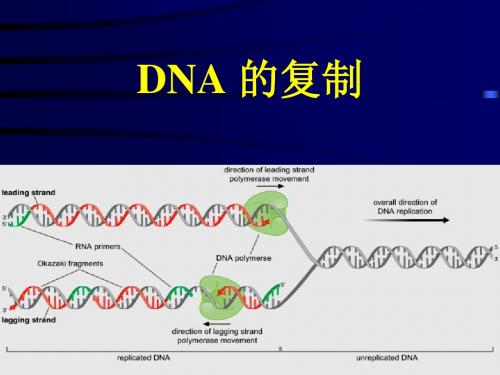
2.3和2.4 DNA的复制和特点

(single-strand binding protein,SSB)
(peimase)
(DNA polymetases)
(DNA ligase) 自由草
16
自由草
17
自由草
18
自由草
(Replication fork)。
• DNA的复制是由固定的起始点开始的。一般把 生物体的复制单位称为复制子(replicon), 一个复制子只含一个复制起点。
自由草
12
正在复制的 部分在电镜 下象只眼睛, 称复制眼 (泡)
自由草
13
原核细胞DNA的半不连续复制过程
自由草
24
2.4 原核生物和真核生物 DNA的复制特点
自由草
25
2.4.1 原核生物DNA的复制特点
大肠杆菌基因组以双链环状DNA分子的形式存在,其DNA复制的中间产物可形成一 个θ,复制从定点开始双向等速进行。复制起始后,两个复制叉在距起始点180°处会 合。
-(可切单链)
突 变 体
突变位点 突变表型
pol A 修复有缺陷
pol B 能修复
polC(dnaE), dnaN, dnaZX, dnaQ, dnaT 阻止复制
自由草
38
DNA聚合酶的共同点:
• 1、都以dNTP为底物。 • 2、都需要Mg2+激活。
• 3、聚合时必须有模板链和具有3‘-OH末端的引
35
链的终止需要Tus蛋白参与
• 当复制叉前移,遇到20bp重复性终止子序列 (Ter)时,Ter-Tus复合物能阻挡复制叉的继 续前移,等到相反方向的复制叉到达后在DNA
拓扑异构酶IV的作用下使复制叉解体,释放子
第二章DNA的复制
DNA Polymerase-palm domain
1. Contains two catalytic sites, one for addition of dNTPs and one for removal of the mispaired dNTP. 2. The polymerization site: (1) binds to two metal ions that alter the chemical environment around the catalytic site and lead to the catalysis. (2) Monitors the accuracy of base-pairing for the most recently added nucleotides by forming extensive hydrogen bond contacts with minor groove of the newly synthesized DNA. 3. Exonuclease site/proof reading site
原核生物中的三种DNA聚合酶
pol Ⅰ 5'→3'聚合酶活性 5'→3'外切酶活性 3'→5'外切酶活性 生理功能 + + +
去除引物,填补缺口 修复损伤 校正错误
pol Ⅱ + +
未知
pol Ⅲ + +
DNA 复制 校正错误
• 在真核生物中,目前发现的DNA聚合酶 有五种,分别命名为DNA聚合酶α(pol α),DNA聚合酶β(polβ),DNA聚合 酶γ(polγ),DNA聚合酶δ(pol δ), DNA聚合酶ε(polε)。 • 参与染色体DNA复制的是polα(延长滞 后链)和polδ(延长前导链),参与线 粒体DNA复制的是polγ,polε与DNA损 伤修复、校读和填补缺口有关,polβ只 在其他聚合酶无活性时才发挥作用。
分子生物学:第二章DNA复制复习题与答案

第二章DNA复制复习题与答案一、名词解释1.中心法则2.半保留复制3.DNA聚合酶4.解旋酶5.拓扑异构酶6. 单链DNA结合蛋白7. DNA连接酶8. 引物酶及引发体9.复制叉10. 复制眼11. 前导链12. 冈崎片段13.半不连续复制14.切除修复15.重组修复16. 诱导修复和应急反应二、问答题1.试述Meselson和Stahl关于DNA半保留复制的证明实验。
2.描述大肠杆菌DNA聚合酶I在DNA生物合成过程中的作用。
3.试述DNA复制过程,总结DNA复制的基本规律。
4.DNA的损伤原因是什么?三、填空题1.Meselson-Stahl的DNA半保留复制证实试验中,区别不同DNA用_______方法。
分离不同DNA用_______方法,测定DNA含量用_______方法,2.DNA聚合酶I(E.coli)的生物功能有_______、_______和_______作用。
用蛋白水解酶作用DNA聚合酶I,可将其分为大、小两个片段,其中_______片段叫Klenow 片段,具有_______和_______作用,另外一个片段具有_______活性。
3.在E.coli中,使DNA链延长的主要聚合酶是_______,它由_______亚基组成。
DNA 聚合酶Ⅱ主要负责DNA的_______作用。
4.真核生物DNA聚合酶有_______,_______,_______,_______。
其中在DNA复制中起主要作用的是_______和_______。
5.解旋酶的作用是_______,反应需要提供能量,结合在后随链模板上的解旋酶,移动方向_______,结合在前导链的rep蛋白,移动方向_______。
6.在DNA复制过程中,改变DNA螺旋程度的酶叫_______。
7.SSB的中文名称_______,功能特点是_______。
8.DNA连接酶只能催化_______链DNA中的缺口形成3’,5’- 磷酸二酯键,不能催化两条链间形成3’,5’- 磷酸二酯键,真核生物DNA连接酶以_______作为能源,大肠杆菌则以作为能源,DNA连接酶在DNA______、________、_______中起作用。
DNA的复制(说课稿).

《DNA的复制》说课稿各位老师:大家好,我今天说课的题目是苏教版高中生物必修2第四章第二节”DNA复制”这部分内容,接下来我就从以下几个方面来说说这一节课。
一、说教材1.教材地位和作用《DNA的复制》这一部分内容是第二章的重点内容之一。
DNA分子的结构和复制是遗传学的基本理论。
“DNA的复制”一课时,在联系DNA结构的基础上,进一步阐明DNA通过复制传递遗传信息的功能。
具体内容有:复制的概念、时间、场所、条件、过程、特点、意义。
学好这一课时,对于学生深刻认识遗传的本质是非常重要的。
二、说教学目标1.根据大纲要求和学生的实际,我确定了以下教学目标:(1)知识目标:1、记住DNA复制的概念2、简述DNA复制的过程,并分析、归纳出DNA复制过程的特点。
3、知道DNA复制在遗传上的意义(2)能力目标:1.通过介绍Meselson、stehl的试验,引导学生分析、比较、推理、归纳,培养科学的思维。
2.通过引导学生观察生活中复印和DNA复制的比较,鼓励学生大胆想象、猜测,培养学生自主探索、合作学习、分析问题、解决问题的能力。
(3)情感目标通过分组探究活动,培养学生的协作意识和科学态度。
2.教学重点、难点(1)教学重点:DNA复制的条件、过程及特点。
(2)教学难点:DNA复制的过程,特别是半保留复制。
3.教材处理及课时安排根据教材的重难点以及学生的实际情况,本节内容只安排一个课时。
教学顺序是“推测-实验证据-复制过程”进行。
三、说教法结合教材的特点和学生实际,本课时主要采用启发式教学法,结合直观、比较、讨论、探究等教学方法。
通过教师的引导,从知识的发生过程入手,教师把教材中的科学活动过程充分展开,着重引导学生自己去探索、观察、思考、分析、归纳、总结,使学生在教学活动中初步学会科学研究的一般方法,即发现问题——提出假说——试验验证——得出结论,培养学生的合作、探索的精神,发展学生的思维能力,为学生铺设符合知识规律的思维轨道。
第二章 DNA结构、复制、 修复

4)DNA序列的异质性及主要序列类型(真核DNA)
■
高度重复序列:重复频率高达几十万到几百万次。
1)卫星DNA:重复单位多由2-10bp组成,成串排列,其碱基 可以用等密度梯度离心法将其与主体DNA分开。根据重复频 率和重复序列长短不同分为小卫星DNA和微卫星DNA(常作 为一种分子遗传标记)
2)分散高度重复序列:短、长散置序列
■影响复性速度:
DNA的大小(小的较大的容易);离子浓度(高浓度); DNA浓度(越大越快)
2) C值反常现象(C-value paradox)
C值矛盾
C值是一种生物的单倍体基因组DNA的总量。
真核细胞基因组的最大特点是它含有大量的重复
序列,而且功能DNA序列大多被不编码蛋白质的非
功能DNA所隔开,这就是著名的“C值反常现象”。
第二章 染色体与DNA
染色体
DNA的结构 DNA的复制 DNA的修复 DNA的转座
三、DNA的复制
RNA 复制 复制
DNA
转录 逆转录
RNA
翻译
蛋白质
内容提要: ● DNA的半保留复制 ●与DNA复制有关的物质 ● DNA的复制过程(大肠杆菌为例) ● DNA复制的其它方式 ●真核生物中DNA的复制特点
染色质是一种纤维状结构,叫做染色质丝,它是由 最基本的单位—核小体(nucleosome)成串排列而成 的。
真核生物染色体的组成
染色体
{蛋白质
DNA
{
组蛋白: H1 H2A H2B H3 H4 非组蛋白
}核小体
(三)染色体的结构和组成
1、组蛋白的一般特性:
■ 进化上的保守性 保守程度:H1 ■无组织特异性 ■肽链氨基酸分布的不对称性 ■H5组蛋白的特殊性:富含赖氨酸(24%) ■组蛋白的可修饰性 H2A、H2B H3 、H4
第2章 DNA的复制

- 第四节 DNA的复制 真核生物复制的特点
1、复制叉移动速度大约只有50bp/s,不到大肠杆菌得1/20。 2、真核生物每条染色质上可以有多处复制起始点:人类DNA中 每间隔3万-30万个碱基就有一个复制起始点,而原核生物只有 一个起始点; 3、真核生物的染色体在全部完成复制之前,各个起始点上DNA 的复制不能再开始,而在快速生长的原核生物中,复制起始点上 可以连续开始新的DNA复制,表现为虽只有一个复制单元,但 可有多个复制叉。 4、真核生物DNA聚合酶的特性:5种DNA聚合酶 5、端粒酶保证染色体复制的完整性。
“多莉”的衰老 研究端粒丢失的速率,预测人类的寿命 研究推测端粒酶与肿瘤的关系
第五节 DNA复制的调控
原核细胞的生长和增殖速度取决于培养条件,在不同
生长和增殖速度的细胞中DNA链延伸的速度几乎是恒定的, 但复制叉的数量不同。迅速分裂的细胞具较多复制叉,而分 裂缓慢的细胞复制叉较少并出现复制的间隙。
第五节 DNA复制的调控
真核细胞的生活周期可分为4个时期:
(1)G1:复制预备期;
(2)S:复制期;
(3)G2:有丝分裂准备期; (4)M:有丝分裂期。
DNA复制只发生在S期。
第五节 DNA复制的调控
真核细胞中DNA复制有3个水平的调控:
1.细胞生活周期水平调控,也称为限制点调控,即决定细
胞停留在G1期, 还是进入S期。——复制起点点火
5’
5’ 3’
+
3’ 复制叉到达末 3’ 端后,一条单
5’ 链被置换出来
末端碱基配对
5’
形成双链体起
3’
始点
5’
以单链为模板
3’
5’ 的DNA合成
3: 腺病毒DNA的复制
第二章 DNA复制与损伤

一个复制子只含有一个复制起点。
(二)子链DNA延伸方向
新 链 合 成 只 能 从 5’ 到 3’ 方 向
T CA 5’ OH ppp
C OH
T CA C
5’ OH 3’ + ppi
不同细胞的复制参数 原核生物(大肠 杆菌) 1(1) 3~9×106 多为双向 等速(850) 真核生物(人) 多个(103~104) 约105~106 多为双向 等速(60~90)
AP位点的产生
2.核苷酸切除修复(nucleotide-excision repair)
切除酶(excinuclease)也是一种核酸内切酶,但与一般的核酸 内切酶不同,在链的损伤两侧同时切开。编码此酶的基因是Uvr 基因。
五、连接酶(Ligase)
DNA连接酶催化双股链内相邻单链切口的3′-OH与 5′- P酰基形成磷酸酯键。 催化过程:
酶的活化 腺苷酰化DNA 亲核攻击完成DNA连接
需要NAD+或ATP提供腺苷酰基(AMP),与酶活性中心的赖氨酸残 基的ε-NH2以磷酰胺键结合,形成共价中间体酶- AMP; 。 酶- AMP将腺苷酰基(AMP)转移给DNA切口处的5′磷酰基团, 以焦磷酸的形式活化,形成AMP-P-DNA。 通过相邻DNA的3′-OH对活化的P原子进行亲核攻击,生成3′,5′磷酸二酯键,同时释放出AMP。
第三章 DNA的复制
§3-1 DNA复制概貌 §3-2 DNA复制酶和相关蛋白 §3-3 DNA的复制过程 §3-4 DNA的修复 §3-5 DNA的突变
§3-1 DNA复制概貌
DNA复制是一个由多种酶催化和有多种蛋白 质参与的受到精密调控的过程; DNA是细胞中唯一具修复系统的生物大分子。
一、DNA复制的半保留性 (Semiconservative replication)
分子生物学基础第二章DNA的结构、复制和修复第五节 DNA的损伤与修复

第五节 DNA的损伤与修复
图2-13 DNA分子上的胸腺嘧啶二聚体结构
第五节 DNA的损伤与修复
图2-11 甲基介导的错配修复模 型
第五节 DNA的损伤与修复
3.核苷酸切除修复 核苷酸切除修复系统几乎能够修复紫外线照射引起的 各种损伤。包括环丁烷二聚体、6–4损伤、碱基-糖基交联 等引起DNA双螺旋大扭曲(major distortion),而不能修 复由于碱基错配、O6–甲基鸟嘌呤、O4–甲基胸腺嘧啶、8– oxoG或碱基类似物引力是非常重要的。
第五节 DNA的损伤与修复
二、DNA的修复 1.错配修复 E.coli避免突变的主要途径之一就是甲基指导的错配修复系统。 这个系统是非特异性的,它能修复引起DNA双螺旋轻微扭曲的任何扭 伤,包括错配、移码、碱基类似物的掺人和某些类型微小扭曲的烷基 化损伤。 2.碱基切除修复 是一种在细胞中存在较普遍的修复过程。在细胞中都有不同类型、 能识别受损核酸位点的糖苷水解酶,它能特意性切除受损核苷酸上的 N—β-糖苷键,在DNA链上形成去嘌呤或去嘧啶位点(AP位点)。DNA 分子中一旦产生了AP位点,核酸内切酶就会把受损核酸的糖苷-磷酸 键切开,并移去包括AP位点核苷酸在内的小片段DNA,由DNA聚合酶I 合成新的片段,最终由DNA连接酶把两者连成新的被修复的DNA链。
分子生物学DNA复制

二、 DNA双螺旋模型结构(double helix model)
1.双螺旋结构提出背景依据 a、 1938. W. T. Astbury 首次用X-射线分析DNA
b、 1950 Chargaff A + G / T + C = 1 A+T ≠ G + C
c、 1952 Alexander Todd 发现了核苷酸和核苷酸之间由磷酸二酯键联接
☉ 核苷(nucleotide)
嘧啶的1位N原子、嘌呤的9位N原子
糖苷键 Glycosidic bond
9
核糖是戊糖
1’
RNA-核糖核苷(R=OH)
DNA-脱氧核糖核苷(R=H)
☉ 核苷酸(nucleotide acid)
核苷的磷酸酯 脱氧核糖核苷酸 核糖核苷酸
γβα
CMP NMP
dATP dNTP
3、影响 Tm值的因素
(1) 在 A, T, C, G 随机分布的情况下 ,决定于GC含量 GC%愈高 →Tm值愈大;GC%愈低→Tm 值愈小
(2)GC%含量相同的情况下
AT形成变性核心,变性加快,Tm 值小
碱基排列对Tm值具有明显影响
5` GC 3` CG n
136.12 ℃
>> Tm
5` CG 3` GC n 72.55 ℃
Concentration 50μg/ml
Opetical Density
dNTPs A260 = 1.60 S.S DNA A260 = 1.37 D.S DNA A260 = 1
OD 1.37
1.185
1.0
℃
Tm (melting temperature) = midpoint of the temperature range over which DNA is denatured = OD增加值的中点温度(一般为85-95℃)
02 染色体与DNA-1

分子收缩1/7。
人中期染色体中含6.2×109碱基对,其理论长
度应是200cm,这么长的DNA被包装在46个 5μm长的圆柱体(染色体)中,其压缩比约为 104。
分裂间期染色质比较松散,压缩比大约是102~
103。
染色体形成过程中长度与宽度的变化
2.1.3 原核生物和真核生物基因组结构特点比较
核孔复合物蛋白以及肌动蛋白、肌球蛋白、微
管蛋白、原肌蛋白等。
简述真核生物染色体上组蛋白的种类,组蛋白修 饰的种类及其生物学意义
中国科学院2003年硕士研究生入学《生物化学与 分子生物学》试题
2、DNA
1) C值反常现象(C-value paradox)
C值矛盾
C值是一种生物的单倍体基因组DNA的总量。
第二章 染色体与DNA
染色体
DNA的结构
DNA的复制
DNA的修复
DNA的转座
分子生物学研究已经证实,DNA控制了生
物的性状遗传。无论DNA或RNA,都是由 许许多多个核苷酸连接而成的生物大分子, 而每个核苷酸又由磷酸、核糖和碱基3部分 组成。
碱基结构式
2.1 染色体(Chromosome)
X174 D-E-J-F-G-H
mRNA
蛋白J、F、G H D E
E.coli 色氨酸操纵子 9个顺反子 9个酶 ( 第六章 )
基因内基因
部分重叠基因
一个碱基重叠
2、真核生物基因组结构特点
●真核基因组结构庞大 ●单顺反子 ●基因不连续性
3×109bp、染色质、核膜
断裂基因(interrupted gene)、 多于编码序列(9:1)
RNA
翻译
蛋白质
分子生物学基础第二章DNA的结构、复制和修复 第四节原核生物和真核生物DNA的复制特点

第四节 原核生物和真核生物DNA的复制特点
第四节 原核生物和真核生物DNA的复制特点
二、真核生物DNA的复制特点 1.真核细胞的每条染色体含有多个复制起始点。复制子的大小 变化很大,约5-300kbp。复制可以在几个复制起始点上同时进行,复 制起始点不是一成不变的。在发育过程中,活化的细胞有更多的复制 起始点。例如,果蝇在胚胎发育早期,其最大染色体上有6000个复制 叉,大约每10 kbp就有一个。 2.真核生物染色体在全部复制完成之前,各个复制起始点不能 开始新一轮的复制。而原核生物中,复制起始点上可以连续开始新的 复制事件,表现为一个复制子内套叠有多个复制叉。 3 . 真 核 生 物 DNA 的 复 制 子 被 称 为 自 主 复 制 序 列 ( ARS), 长 约 150bp左右,含有几个复制起始必须的保守区。并且其复制起始需起 点识别复合物(ORC)参与,并需ATP。真核生物复制叉的移动速度大 约 只 有 5 0 bp/s, 还 不 到 大 肠 杆 菌 的 1 / 2 0 。 因 此 , 人 类 DNA 中 每隔 3x104~3x105就有一个复制起始位点。
第四节 原核生物和真核生物DNA的复制特点
4.真核生物有多种DNA聚合酶,分别为在真核细胞中主要有5种 DNA聚合酶,分别称为DNA聚合酶α、β、γ、δ和ε,真核细胞的 DNA聚合酶和细菌DNA聚合酶基本性质相同,均以dNTP为底物,需Mg2+ 激活,聚合时必须有模板链和具有3´–OH末端的引物链,链的延伸方 向为5´→3´。但真核细胞的DNA聚合酶一般都不具有核酸外切酶活性, 推测一定有另外的酶在DNA复制中起校对作用。DNA聚合酶α的功能主 要是引物合成。DNA聚合酶β活性水平稳定,可能主要在DNA损伤的修 复中起作用。DNA聚合酶δ是主要负责DNA复制的酶,参与先导链和滞 后链的合成。而DNA聚合酶ε的主要功能可能是在去掉RNA引物后把缺 口补全。
第二章 DNA的复制-分子生物学

25
错配碱基
切除错配 核苷酸
正确 核苷酸
复制方向
3´ 5´ 3´ 3´ 5´
DNA聚合酶Ⅰ的校对功能(3’-5’ 外切酶活性)
26
5’-3‘外切酶活性: ♦ 从5’-P端依次切除,可 连续切除多个核苷酸; ♦ 只切配对的5’-P末端核 苷酸; ♦ 既可切除脱氧核苷酸 也可切除核苷酸; ♦ 对只有5’末端的切口也 有活性。
Pol C: polymerase
Dimerizing Units
Sliding Clamp the clamp loader
32
33
34
35
B 真核生物聚合酶
五种 • DNA聚合酶α • DNA聚合酶δ • DNA聚合酶γ
• DNA聚合酶β、ε
功能: • 参与随从链的合成 • 参与前导链的合成 • 参与线粒体DNA的合成 • 参与DNA的修复
21
4)DNA聚合酶(DNA Polymerase): • 以dNTP为前体催化合成DNA • 需要模板和引物的存在 • 不能起始合成新的DNA链
• 催化dNTP加到生长中的DNA链的3’-OH末端
• 催化DNA合成的方向是5'→3'
22
A 原核生物聚合酶 • DNA聚合酶有5种
• 具有多种酶活性的多功能酶 • 参与DNA复制的主要是polⅢ和polⅠ。 DNA聚合酶Ⅰ(DNA PolymeraseⅠ, PolⅠ) • Kornberg酶(1956年)
连续的小片段的链称为随从链。
(复制方向与解链方向相反)
61
♦ 冈崎片段(Okazaki): DNA复制时,一股以5’ 3’方向的母链作为模板, 指导新合成的链沿5’ 3’合成1000—2000个核苷酸不连
高中生物第二章DNA的生物合成(复制)

第二章 DNA的生物合成(复制)教学大纲要求1. 描述遗传学中心法则,扩大的中心法则及生物学意义。
2. 记住DNA合成的概念,包括以DNA作为模板指导的DNA合成(复制),以RNA作为模板指导的DNA合成(反转录)及DNA的修复合成,分别描述其概念。
3. 复述DNA复制特点,过程,参与的酶和因子(包括它们的功能)。
简要叙述复制过程及真核DNA复制特点。
4. 结合反转录酶的功能,简要叙述反转录过程及其生物学意义。
记住端粒酶的概念与功能。
5. 列举DNA损伤的几种类型,写出修复合成的几种方式名称。
叙述切除修复过程。
教材内容精要(一)遗传信息传递概述基本概念:1. 遗传:生殖过程中表现出来的子代与亲代的相似性。
2. 变异:生殖过程中表现出来的子代与亲代的差异性。
3. 基因: 能为生物大分子蛋白质,也包括RNA编码的核酸片段。
高等生物的基因是DNA,少数低等生物的遗传物质是RNA。
4. 复制: 即DNA的生物合成,DNA母链为模板,由核苷酸聚合成子代DNA的过程。
5. 转录即RNA的生物合成,DNA贮存的遗传信息作模板,转抄成RNA的碱基序列。
6. 翻译: 把mRNA的遗传信息用遗传密码的方式破读为蛋白质分子上的氨基酸排列次序,即蛋白质的生物合成。
7. 中心法则: 遗传信息从DNA流向RNA,再流向蛋白质的信息传递规律。
DNA有贮存、表达遗传信息功能,因此认为DNA处于生命活动中心。
8. 基因表达贮存在DNA上的遗传信息,通过转录和翻译,指导合成主要执行生命活动功能的蛋白质的过程。
9. 半保留复制亲代的DNA双链解开,各自作为模板,按照碱基配对规律(AT配对,GC配对),指引子链的合成。
因此,子代DNA双链和亲代DNA双链有一致的碱基序列。
DNA是遗传的物质基础。
DNA分子中由4种不同碱基组成的核苷酸的排列顺序(以下简称碱基顺序)即是储藏的遗传信息。
所谓基因,即指DNA分子中碱基组成的功能片段。
DNA分子很大(如人类基因组DNA约含3 109个碱基对),但全部由A、G、C和T四种碱基以不同的排列方式组成。
DNA复制的起始和终止讲述

无论单细胞或多细胞真核生物都存在各自特异的端 粒酶RNA模板。
细胞永生化
5’ 端粒
端粒维持
染色体DNA
端粒消耗
端粒 5’
细胞衰老
复制忠实性的保证
去除RNA引 物
补充空缺
连接
DNA聚合酶Ⅰ的小片段 5’ 3’外切酶
DNA聚合酶Ⅰ的大片段 3’ 5’外切酶 5’ 3’聚合酶
端粒酶
以自身含有的RNA作为模板,合成和补充端粒 的重复序列,具有逆转录酶性质,是一种RNA 依赖的DNA聚合酶,又称为端粒末端转移酶。
可以催化端粒3’ 端的游离-OH基上连接新的 dNTP以延长端粒序列。
端粒酶的功能
端粒酶以其自身的RNA组分为模板,以原有的一条 DNA单链的3’端游离-OH为引物,在端粒末端添加新 的端粒重复序列使端粒延长。端粒酶的作用解决了 “末端复制问题”,保证真核细胞线性染色体DNA复 制得以完全。
选择=结果
汇报结束 谢谢观看! 欢迎提出您的宝贵意见!
材料: E.coli 或噬菌体
一、复制的起始 二、复制的延伸 三、复制的终止
一、复制的起始
黑色点: oriC electron microscopy.
Antibodies of dnaA
一、复制的起始点 oriC
♦ 三个13bp的重复序列 ♦ 四个9bp的重复序列 dnaA结合位点
若干GATC位点
第二章 DNA复制 (DNA Replication)
第一节 基本概念 第二节 复制概况 第三节 DNA复制的酶学 第四节 DNA复制的半不连续性 第五节 DNA复制 第六节 DNA损伤修复
第五节 DNA的复制
(DNA replication in Prokaryote)
第二章DNA结构复制与修复

第一章 绪论 (2节) 第二章 DNA的结构、复制和修复(4节)
第三章 遗传与变异(6节) 第四章 遗传信息的转录(4节) 第五章 遗传信息的翻译——从mRNA到蛋白质(4节) 第六章 原核生物基因表达调控(5节) 第七章 真核生物表达与调控(4节) 第八章 分子生物学实验技术(3节)
核小体的结构(后图)
第二节 DNA的组成和结构
一、DNA的组成
DNA
磷酸
核酸→核苷酸
碱基(嘌呤或嘧啶)
核苷
RNA
脱氧核糖或核糖
(戊糖)
HOH 2C O H
H OH
OH H
H H
β-D-2-脱氧核糖
NH2
N
N
H
H N
9
N
H
腺嘌呤
HOH2C 5′ O OH
4′
1′
3′ 2′
OH OH 核腺苷糖
损伤来源:
1. DNA分子的自发性损伤 2. 物理因素引起的DNA损伤
UV照射引起DNA损伤主要是嘧啶二聚体;电离辐 射对DNA损伤有直接疚和间接效应。
3. 化学因素引起的DNA损伤
烷化剂,碱基类似物对DNA的损伤。
二、DNA的修复
1. 错配修复 2. 碱基切除修复 3. 核苷酸切除修复 4. DNA的直接修复
(2)基因种类和数量较少 DNA分子大都 带有单拷贝基因,多以重叠基因形式存在。
(3)以操纵子为转录单元 以多顺子形成 翻译成各种蛋白质。
三、真核生物染色体的组成
从单细胞的酵母菌到高等哺乳动物,真核生 物细胞都有一个共同特点: 。
真核生物基因表达调控复杂不仅因为真核生 物基因组的复杂性,也与染色体结构有关。
第二章染色体与DNA结构与复制

2.3染色体中的DNA
2.3.1基因组大小与C值矛盾
基因组(genome):
生物有机体的单倍体细胞中的所有DNA,称为该物种的基因组。
C值(C value):
概念: C值特点: 1.不同物种之间C值差异较大,一般来讲物种越进化C值就越大。 2.在结构、功能很相似的同一类生物中,甚至在亲缘关系十分 接近的物种之间,它们的C值可以相差数十倍乃至上百倍。
10%—60%,由6—100个碱基组成,在DNA链上串联重复几 百万次。常含有一些A· T,A· T浮力密度小; 将DNA切断成数百 个碱基对的片段进 行超离心时,常会 在主要的DNA带的 上面有一个次要的 DNA带相伴随,这 就是所谓的卫星 DNA(satellite DNA)。
2.3.3 原核生物和真核生物基因组结构特点比较
O
OH OH 三磷酸腺苷 (AT P )
AMP ADP ATP
ATP
分子的最显著特点是 含有两个高能磷酸键。ATP 水解时, 可以释放出大量 自由能。 ATP 也是一种很好的磷酰 化剂,是许多生物化学反 应的激活步骤。
ATP的性质
2、GTP (鸟嘌呤核糖核苷三磷酸)
GTP是生物体内游离存在的另一种重要的核苷酸衍 生物。它具有ATP 类似的结构, 也是一种高能化合
中科院硕士学位研究生入学分子遗传学试题
中国科学院上海生化与细胞所招收硕士研究生分子 遗传学入学考试: 简述真核细胞内核小体与核小体核心颗粒的结构。
染色体的二级结构:螺线管
DNA double helix
Nucleosome (10 nm fiber)
30 nm Fiber
Loops I
Loops II
分子生物学基础第二章DNA的结构、复制和修复 第三节DNA的复制

第三节 DNA的复制
表2-2 部分生物复制子的比较
第三节 DNA的复制
图2-5 放射性实验证明DNA的复制是从固定的起始 点双向等速进行的
第三节 DNA的复制
三、DNA复制的几种主要方式
1.线性DNA双链的复制
复制叉生长方向有单一起点的单向(如腺病毒)及双向(如噬菌体), 和多个起始点的双向几种,DNA双向复制时复制叉处呈“眼”型。线 性DNA复制中RNA引物被切除后,留下5′端部分单链DNA,不能为DNA 聚合酶所作用,使子链短于母链。T4和T7噬菌体DNA通过其末端的简 并性, 使不同链的3′端因互补而结合,其缺口被聚合酶作用填满, 再经DNA连接酶作用生成二联体。这个过程可重复进行直到生成原长 20多倍的多联体,并由噬菌体DNA编码的核酸酶特异切割形成单位长
度的DNA分子。制时,5′端首先与末端蛋白共价结合,开始互补链的合成。当另 一条链完全被置换后,两端通过发卡结构相连,形成一个大部分序列 互补的单链环形DNA分子,复制从其内部的起始位点开始按前导链方 式双向进行,经过环形结构到达分子的另一部分,经双链结构交错切 割后生成完整的子链病毒。除了环形部分发生重排之外,所生成的新 DNA分子带有母链的全部遗传信息。
分子生物学基础
第二章 DNA的结构、复制和修复
第三节 DNA的复制
一、DNA的半保留复制机理 二、DNA复制的起点、方向和速度 DNA在复制时,首先在一定位置解开双链,这个复制起点呈现叉 子的形式,称为复制叉。一般把生物体能独立进行复制的单位称为复 制子。实验证明,复制在起始阶段进行控制,一旦复制开始,就连续 进行下去,直到整个复制子完成复制。每个复制子由一个复制起点控 制。 原核生物的复制起始点通常在它染色体的一个特定位点,并且只 有一个起始点,因此,原核生物的染色体只有一个复制子。真核生物 染色体的多个位点可以起始复制,有多个复制起始点,因此是多复制 子(表2-2)。且多个复制子不是同时起作用,而是在特定时间,只 有一部分复制子(不超过15%)在进行复制过程。 关于DNA复制的方向和速度,最为普遍的就是双向等速进行(图 2-5)。某些环状DNA偶尔从一个复制起始点形成一个复制叉,单向复 制。而腺病毒则从两个起始点相向进行复制。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
1、 DNA的半不连续复制的发现 2、引物、引发酶和引发体 3、原核生物半不连续复制的过程 4、几种噬菌体的复制过程
河南师范大学
DNA的半不连续复制
1、DNA的半不连续复制的发现
DNA链的生长方向是从5’-3’的,那么DNA 的 复制就可能有几种方式:
3’ 5’ 5’ 5’ 5’ 3’
子代链
3’
河南师范大学
DNA 的复制
本章内容
第一节 第二节 第三节 第四节 第五节 第六节
DNA的半保留复制 复制原点、方向、和方式 DNA复制的酶学 DNA的半不连续复制 真核生物DNA的复制 DNA复制的错误校对
河南师范大学
DNA的半保留复制
第一节 DNA的半保留复制
单独复制的DNA单元称为复制子(Replicon ),原核 细胞的复制子就是细菌染色体本身,每个真核细胞染色 体都包含有大量的复制子。
河南师范大学
DNA的半不连续复制
河南师范大学
1、一个RNA序列(约10bp),RNA链 的3’-OH自由端会被DNA聚合酶延伸。 细菌DNA和一些病毒复制时使用。 2、 一个事先形成的RNA和模版配对, 允许它的3’-OH端被用来引导DNA的 合成。这种机制在反转录病毒中被 用来引导RNA的反转录。 3 、一个引导点在双链DNA中产生。 最常见的机制是通过引入一个切刻。 在这种情况下,事先形成的链由新 生成的链替代。(注意这和切刻平 移的不同,切刻平移中,DNA聚合酶 I 在切刻同时生成并且降解DNA。) 少数病毒使用 4、一个蛋白质可以直接引导反应, 它可以直接向DNA聚合酶提供一个核 苷。这种方式有一些病毒采用。
河南师范大学
DNA复制的酶学
Single-stranded DNA is needed for replication
SSB :single-strand DNA binding protein 作用:阻止已经解开的双链链间或者链 内退火河南来自范大学DNA的半不连续复制
第四节
DNA的半不连续复制
河南师范大学
复制原点、方向、和方式
The electron micrograph of the replication intermediate of a plasmid DNA is -shaped.
河南师范大学
复制原点、方向、和方式
Replicated DNA is seen as a replication eye flanked by nonreplicated DNA.
河南师范大学
1、 Polymerase
DNA复制的酶学
DNA polymerase Ⅲ
PolⅢ是DNA链延长的主要的聚合酶,是一个复杂的多 亚基酶,它的活性主要有: 1、 Polymerase (DNA replication ); 2、 3`→ 5` exonuclease (proofreading) ; 3、 5`→ 3` exonuclease ( damaged DNA fragments)
河南师范大学
DNA复制的酶学
(二)、DNA连接酶
作用:将相邻的3’--OH和5’--P形成磷酸二酯键
条件:必须是相邻的3’--OH和5’—P;
必须是各自的碱基处于配对状态下
OH
Nick
P 3’ 5’ 5’ 3’
Nick
3’ 5’
5’ 3’
Nick
5’ 3’ 3’ 5’ 5’ 3’
A C
3’ 5’
河南师范大学
DNA的半不连续复制
2、引物、引发酶和引发体
Priming is required to start DNA synthesis
DNA聚合酶不能初始一个脱氧核糖核酸链,必须要由一 个引导行为提供一个自由的3’-OH端开始前导链和后随链的 合成。前导链只需要一次这样的初始事件,是在复制起点 开始的。但是后随链必须要有一系列的初始事件。因为每 一个冈崎片段都需要自己重新开始。每一个冈崎片段都是 由一个引物开始的。这个引物可以具有多种形式:
Severo Ochoa New York University; College of Medicine New York, NY, USA
Arthur Kornberg Stanford University Stanford, CA, USA
河南师范大学
DNA复制的酶学
一 、脱氧核苷三磷酸前体的来源
第二章
DNA 的复制
DNTP
能量
看守
生产
检验
胶水
河南师范大学
DNA 的复制
问
题
1、原核生物和真核生物DNA的复制会一样吗? 2、DNA复制时是两条链都做模板吗?如果是的 话如何解决两条链的方向问题? 3、DNA链为什么不选择3’---5’的方向延伸呢? 4、DNA复制都需要那些酶?他们是如何起作用 的? 5、线性DNA复制的5’末端隐缩问题是如何解决的 6、DNA的复制是如何保证其准确性的?意义何 在?
河南师范大学
DNA复制的酶学
DNA Pol I catalyzes the 5` to 3` elongation of the new strand
河南师范大学
The Nobel Prize in Physiology or Medicine 1959 "for their discovery of the mechanisms in the biological synthesis of ribonucleic acid and deoxyribonucleic acid"
5’
C
A
3’ 5’
3’
外切酶作用位点
河南师范大学
DNA复制的酶学
3、5`→ 3` exonuclease: PolⅠ和PolⅢ都具有该活性但是区别较大: (1) PolⅢ 5`→ 3`外切活性只能作用于单链DNA (2) PolⅠ的外切活性不能作用于单链DNA,只要是具有5’-磷酸末端的 DNA PolⅠ就可以起作用
废物利用:体内核酸降解或者直接来自培养 基
新合成:(参考生物化学) 注意dTTP的合成,dUTP的渗入阻止
河南师范大学
DNA复制的酶学
二、酶
(一)、DNA聚合酶
1、一个能够在模板链上合成新的DNA链的酶叫做DNA 聚合酶。原核和真核生物中都包含有多种DNA聚合 酶的活动。只有它们中的一部分是真正参与了复制 的;有时候把这些酶叫做DNA复制酶。 2、一个所有DNA聚合酶都具有的特点是:它们不能够 从自由的核苷开始初始化DNA链的合成。它们需要 一个引物提供一个自由的3’-OH端可以加入核苷。 3、原核生物的DNA聚合酶目前发现的主要有三种:分 别是PolⅠ、PolⅡ和PolⅢ
河南师范大学
复制原点、方向、和方式
从两个复制原点开始的单向单链复制
从一个复制原点开始的单向双链复制
从一个复制原点开始的双向双链复制
河南师范大学
DNA复制的酶学
第三节
DNA复制的酶学
DNA 复制是核苷酸链的不断生长,那就需要将 四种脱氧核苷三磷酸在酶的作用下通过磷酸二酯键 连接在一起。
一、脱氧核苷三磷酸的来源(生物化学 ): 二、酶
河南师范大学
DNA复制的酶学
PolⅠ和PolⅢ活性比较
1、Polymerase : PolⅠ主要是DNA的修复和RNA引物 的替换; PolⅢ主要是负责链的延伸 2、3`→ 5` exonuclease: PolⅠ和PolⅢ都具有该活性, 主要是保证聚合作用的正确性,起校对作用,他的 意义还在于它对于DNA作为遗传物质的稳定性和极 高的保真度是非常重要的。
5’
3’
5’
3’ 5’
3’ 5’
A
B
C
河南师范大学
DNA的半不连续复制
几个概念
Lagging strand of DNA must grow overall in the 3′-5′ direction and is synthesized discontinuously in the form of short fragments (5′-3′) that are later connected covalently. Leading strand of DNA is synthesized continuously in the 5′3′ direction. Okazaki fragments are the short stretches of 1000-2000 bases produced during discontinuous replication; they are later joined into a covalently intact strand. 1968年冈琦发现的因此称为Okazaki fragments Semidiscontinuous replication is mode in which one new strand is synthesized continuously while the other is synthesized discontinuously.
河南师范大学
DNA的半保留复制
DNA的半保留复制的证明
1958 年Meselson-Stahl experiment using 15N (the Heavy isotope) and 14N (the Light isotope) to grow E. coli cells 证 明了DNA的复制是半保留复制
DNA 可能的三种复制模式
河南师范大学
DNA的半保留复制
15N 15N
河南师范大学