细胞蛋白分选机制整理
细胞生物学 蛋白质分选
而定位于胞质溶胶以及细胞表面的蛋白质是没有
分选信号的,这种定位方式称为违约或欠缺途径
(二)蛋白质的分选信号与运输途径
常见的分选信号 信 号 肽 信号斑
信号肽(signal peptide)
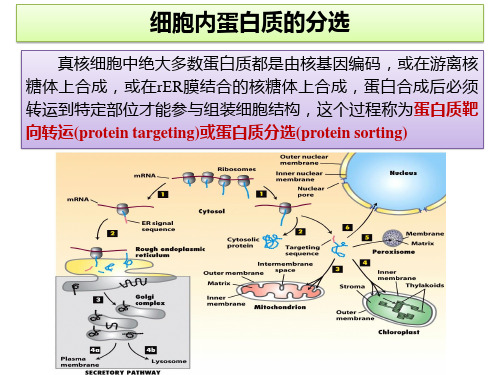
蛋白质中的分选信号
是一段连续的氨基酸序列,长约15-60个氨基酸残基。一旦 完成分选过程,常被一种信号肽酶切除。
1 1 1
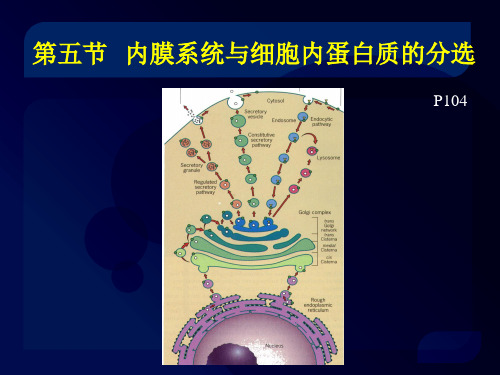
细胞内蛋白质运输路线图
门孔运输
跨 膜
运
输
生物学与遗传学教研室 由膜上的蛋白转运装置(某种膜蛋白充当)运输特异的
蛋白质穿过膜从胞液到各种不同的细胞器。这样运输的 蛋白质通常是不折叠的,运输过程常靠分子伴侣的帮助。
生物学与遗传学教研室
细胞内蛋白质运输路线图
生物学与遗传学教研室
跨 膜 运 输 门 孔 运 输 蛋白质被选择性地包装成膜囊泡的形式(运输小泡), 定向转运到靶细胞器。 生物学与遗传学教研室
信号斑(signal spot)
蛋白质中的分选信号
是指在蛋白质折叠过程中一些氨基酸残基所构成的特异三维 排列。构成信号斑的氨基酸残基在线型氨基酸序列中彼此相 距较远,一般保留在已完成的蛋白质中。
三、蛋白质的运输方式
蛋白质在细胞内的基本运输途径
门孔运输
跨膜运输
囊泡运输
门孔运输
即:蛋白质在核、质之间的运输。该运输需通过核孔复 合体,称为门孔运输或控制运输。核孔复合体具有选择 控制功能,能主动运输特异的大分子和大分子组装物, 1 而小分子物质可自由扩散。亲核蛋白质具有核定位信号 1 (NLS)而核孔复合体上有NLS的受体,二者结合后,亲 核蛋白就能进入核内。 1
囊
泡
运
输
细胞内蛋白质运输路线图
细胞内蛋白质分选的两条途径

细胞内蛋白质分选的两条途径细胞是生命的基本单位,其中蛋白质是细胞最重要的组成部分之一。
在细胞内,蛋白质需要在不同的位置发挥不同的功能,因此需要进行分选。
目前已知有两种主要的细胞内蛋白质分选途径:囊泡转运和直接转运。
一、囊泡转运1. 什么是囊泡转运?囊泡转运是指通过形成、移动和融合小型液滴(即囊泡)来实现蛋白质分选的过程。
这些囊泡可由内质网、高尔基体、溶酶体等细胞器形成。
2. 囊泡转运的过程(1)合成:在内质网上合成的蛋白质被包裹在一个小型液滴中,形成一个囊泡。
(2)移动:这些囊泡随后通过微管道系统向高尔基体或其他目标位置移动。
(3)融合:到达目标位置后,这些囊泡与目标部位上的膜进行融合,并释放出其所携带的蛋白质。
3. 囊泡转运的特点(1)速度快:相对于直接转运,囊泡转运速度更快。
(2)可控性高:囊泡转运可以通过调节囊泡合成、移动和融合等过程来实现对蛋白质分选的精确控制。
(3)适用范围广:囊泡转运可以用于多种类型的细胞内蛋白质分选,例如从内质网到高尔基体、从高尔基体到溶酶体等。
二、直接转运1. 什么是直接转运?直接转运是指蛋白质在没有形成液滴的情况下,通过与其他蛋白质或分子相互作用实现分选的过程。
这些相互作用可能包括靶标蛋白识别、信号传递等。
2. 直接转运的过程(1)靶标识别:特定类型的蛋白质通过与目标位置上的特定靶标结合来实现定向传输。
(2)信号传递:一些蛋白质需要特定信号才能在细胞内进行分选。
例如,磷酸化可以作为一种信号来调节蛋白质在细胞内的分布。
3. 直接转运的特点(1)精确度高:直接转运可以通过靶标识别和信号传递等机制来实现对蛋白质分选的精确控制。
(2)适用范围窄:相对于囊泡转运,直接转运的适用范围较窄,只适用于特定类型的蛋白质分选。
结论:细胞内蛋白质分选是细胞内复杂的过程之一,目前已知有两种主要的分选途径:囊泡转运和直接转运。
这两种途径在速度、可控性、适用范围等方面存在差异,但都可以通过不同机制来实现对蛋白质分选的精确控制。
2023年高中生物竞赛课件:细胞内蛋白质的分选
新生肽链跨膜取向
新生跨膜蛋白的肽链中没 有N端信号肽,只有内部 信号序列(internal signal sequence)。内部信号序列 的转位方向主要取决于其 侧翼氨基酸残基电荷的分 布。一般而言,带正电荷 的氨基酸残基一侧朝向细 胞质基质一侧
一、信号假说与蛋白质分选信号
Ⅳ型:G蛋白偶联受体、葡 萄糖转运蛋白、电压门 Ca2+通道、CFTR(Cl-)通道
图6-4 内质网膜整合蛋白的拓扑学类型
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
ER单次跨膜 新生跨膜蛋白肽链中既有N端信号肽,又 蛋白的合成 有停止转移信号
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
1、分泌性蛋白在游离核糖体上起始合成 2、多肽链延伸至80个氨基酸残基时,N端内质网信号序列暴露,与 SRP结合,肽链延伸暂停,防止新生肽链N端损伤和成熟前折叠
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
3、SRP与ER膜上SRP受体结合,核 糖体新生肽复合物附着到ER膜,两 分子GTP分别与SRP-p54亚基和SRP 受体α亚基结合,相互作用加强
5、腔面信号肽酶切除信号肽并使之快速降解 6-8、肽链继续延伸,并在ER腔内折叠,移位子关闭
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
➢ 开始转移序列(start transfer sequence) 作为内质网的信号序列,指导新合成的多肽向内
细胞内蛋白质的分选和转运机制

参 考 文 献 : [1] 黄庆洲 ,黎德 斌,伍莉.动物生理 学[M].重庆 :西南师范大 学 出版 社,20151 38—39. [2] 方亮.药剂学[M].北京:人 民卫 生出版社 ,2016:42. [3] 孙红 军.等渗溶 液 中的红细 胞为什 么会涨 破 [J].新课程
学 习:社会综合 ,201lf41:26—27. [4] 金 维 芬 .等 渗 液 与 等 张 液 [J].中 国 医 院 药 学 杂 志 ,1984,4 (2):13—14. [5] 曲宝涵.普通化 学[M].北京 :中国农 业大学 出版社 ,2009:
蛋 白质 作 为 生命 活 动 的执 行 者 和体 现 者 ,与 生物
的 遗 传 、疾 病 等 都有 着 重 要 关 联 。在 细 胞 内 ,有 些 蛋
白质 是 先合 成 再 进 行分 选 转运 ,如 线 粒 体 、叶 绿体 、细
胞 核 等 结 构 中 的 蛋 白质 ;而 有 些 是 边 合 成 边 分 选 转
为 了维 持生 命 ,原核 细 胞 需要 合成 一 些 蛋 白质 分 泌 到
细胞质 或 者转运 到 细胞 外发挥 作用 。原核生 物 中蛋 白
细胞内蛋白质的分选和转运机制

细胞内蛋白质的分选和转运机制作者:陈建坤来源:《中学生物学》2018年第03期蛋白质作为生命活动的执行者和体现者,与生物的遗传、疾病等都有着重要关联。
在细胞内,有些蛋白质是先合成再进行分选转运,如线粒体、叶绿体、细胞核等结构中的蛋白质;而有些是边合成边分选转运,如分泌蛋白、膜蛋白等。
细胞根据蛋白质是否携有分选信号(信号序列)以及分选信号的性质,有选择地将蛋白质运送到细胞的不同部位。
1分选信号的种类分选信号有两类:①信号肽:蛋白质多肽链上一段连续的特定氨基酸序列,一般位于新肽链的N端,属于一级结构。
完成分选任务后常被切除。
②信号斑:位于多肽链不同部位的几个特定氨基酸序列经折叠后形成的斑块区,是一种三维结构。
完成分选任务后,仍然存在。
2原核细胞中蛋白质分选转运途径原核细胞(如细菌)没有复杂的生物膜系统。
但是为了维持生命,原核细胞需要合成一些蛋白质分泌到细胞质或者转运到细胞外发挥作用。
原核生物中蛋白质的转运分泌途径主要包括3种:①一般分泌途径,即SEC途径;②双精氨酸移位酶途径,即TAT途径;③信号颗粒识别途径,即SRP途径。
除此之外,还有V型分泌途径、TPS分泌途径和分子伴侣引导分泌途径等。
①一般分泌途径,即SEC途径:SEC分泌途径是原核生物中蛋白质主要的跨膜运输机制,主要由SEC移位酶作为介导。
SEC途径可以转运多种蛋白质,包括毒性因子、菌毛、黏附素和蛋白酶等。
SEC途径主要功能是把尚未折叠完成的蛋白质转运到质膜外,在质膜外折叠成有活性的蛋白质。
SEC途径大致可分为3个过程:信号序列的识别与定位、跨膜转运和多肽的释放。
②双精氨酸移位酶途径,即TAT途径:TAT途径识别的肽链N端信号序列通常含有两个连续的精氨酸残基。
TAT途径主要转运已经折叠完成的蛋白质,而尚未折叠完成的蛋白质通常不能通过该系统分泌,从而避免未完成折叠蛋白在胞外被降解的命运,保证了分泌产物的结构和功能的准确性。
此外,TAT途径还可以将少数蛋白质整合到质膜中。
细胞内蛋白质分选的基本途径

细胞内蛋白质分选的基本途径
一、翻译后转运途径
翻译后转运途径是指蛋白质在完成多肽链的合成后,再通过特定的转运途径将其输送到细胞内指定位置。
这一途径主要涉及信号识别颗粒(SRP)的识别和核糖体与内质网之间的相互作用。
通过翻译后转运途径,细胞可以精确地控制蛋白质的合成和分选过程,以满足其特定需求。
二、共翻译转运途径
共翻译转运途径是指蛋白质在合成过程中即开始进行分选转运的途径。
该途径涉及信号肽的识别和引导,以及跨膜运输过程中的信号肽切除。
共翻译转运途径的主要特点是蛋白质在合成过程中就与转运相关的分子结合,从而引导其向特定方向进行转运和定位。
三、膜泡运输途径
膜泡运输途径是指蛋白质在合成过程中被包裹在膜泡内,通过一系列膜泡的转运和融合过程,最终将蛋白质运送到指定位置。
膜泡运输途径的主要特点是能够将蛋白质从粗面内质网合成部位转运至高尔基体,进而再转运至溶酶体、分泌泡等细胞内的不同部位。
四、门控转运途径
门控转运途径是指通过核孔复合体进行的选择性转运过程。
这一途径主要涉及细胞核内外蛋白质的合成与运输,特别是一些核质穿梭蛋白在细胞核与细胞质之间的运动。
门控转运途径对于维持细胞核的正常功能具有重要意义。
五、定位与锚定途径
定位与锚定途径是指蛋白质通过与细胞骨架系统的相互作用,实现其在细胞内的准确定位和锚定。
细胞骨架系统由微管、微丝和中间纤维构成,它们共同维持了细胞的形态并参与物质运输。
通过定位与锚定途径,蛋白质能够在特定的细胞区域发挥其功能,从而维持细胞的正常生理活动。
细胞内蛋白质分选转运的主要方式

细胞内蛋白质分选转运的主要方式细胞内蛋白质分选转运是细胞内重要的生物过程之一,它确保了蛋白质在细胞内的正确定位和功能发挥。
细胞内蛋白质分选转运的主要方式包括囊泡运输、膜蛋白介导的转运和核糖体直接转运。
一、囊泡运输囊泡运输是细胞内蛋白质分选转运的重要方式之一。
它涉及到细胞膜上的囊泡与细胞器之间的运输。
囊泡是由膜蛋白包裹的小泡状结构,可以在细胞内进行蛋白质的转运。
囊泡运输主要分为内质网-高尔基体-溶酶体途径和内质网-高尔基体-细胞膜途径两种。
内质网-高尔基体-溶酶体途径是一种常见的囊泡运输方式。
在这个过程中,新合成的蛋白质被翻译成多肽链后,通过内质网的蛋白质翻译复合物进入内质网腔。
内质网腔中的蛋白质经过修饰和折叠后,被囊泡包裹形成转运囊泡。
这些囊泡随后与高尔基体融合,将蛋白质运输到溶酶体进行降解或分泌到细胞外。
内质网-高尔基体-细胞膜途径是另一种囊泡运输方式。
在这个过程中,蛋白质通过内质网进入高尔基体,然后通过囊泡运输到细胞膜。
这种方式主要用于膜蛋白的转运,以及一些细胞外分泌蛋白的释放。
二、膜蛋白介导的转运膜蛋白介导的转运是细胞内蛋白质分选转运的另一种重要方式。
在这个过程中,膜蛋白起到了关键的作用,它们通过与其他蛋白质相互作用,将目标蛋白质从一个细胞器转运到另一个细胞器。
一个典型的例子是膜蛋白介导的线粒体蛋白质转运。
线粒体是细胞内的重要细胞器,它需要从细胞质中转运蛋白质进入。
这个过程中,线粒体膜上的特定蛋白质与目标蛋白质相互作用,将其引导到线粒体内部。
这种蛋白质介导的转运方式在细胞内的其他膜蛋白转运中也起到了重要的作用。
三、核糖体直接转运核糖体直接转运是一种相对简单的蛋白质分选转运方式。
在这个过程中,蛋白质在合成过程中直接从核糖体转运到目标细胞器。
这种方式主要适用于一些小分子蛋白质的转运,例如核糖体合成的核糖体蛋白质。
细胞内蛋白质分选转运的主要方式包括囊泡运输、膜蛋白介导的转运和核糖体直接转运。
这些方式相互配合,确保了细胞内蛋白质的正确定位和功能发挥。
细胞生物学 蛋白质分选

②穿膜运输 :是指通过结合 在膜上的蛋白质运转体进 行的蛋白质运输。
③小泡运输:是由不同膜性 运输小泡承载的一种蛋白 质运输形式。
蛋白质定向转运取决于两个方面
分选信号:信号序列和信号斑 细胞器上特定的信号识别装置
翻译后转运:在细胞质基质游离核糖体上 完成多肽链的合成,然后转运至膜围绕的 细胞器。
共翻译转运:通过定位信号,一边翻翻译,一 边进入内质网, 然后再进行进一步的加工和 转移。
蛋白质分选的四种基本类型
蛋白质的跨膜转运:主要指在细胞质基质 合成的蛋白质转运至内质网、线粒体、叶 绿体和过氧化物酶体等细胞器。
蛋白质分选
蛋白质分选(protein sorting)主要是指膜结合 核糖体上合成的蛋白质, 通过信号肽,在翻译 的同时进入内质网, 然后经过各种加工和修 饰,使不同去向的蛋白质带上不同的标记, 最 后经过高尔基体反面网络进行分选,包装到 不同类型的小泡,并运送到目的地。也包括 在游离核糖体上合成的蛋白质的定位。
膜泡运输:蛋白质通过不同类型的转运小 泡从其糙面内质网合成部位转运至高尔基 体进而分选运至细胞不同的部位。
选择性的门控转运:指在细胞质基质中合 成的蛋白质通过核孔复合体选择性地完成 核输入或从细胞核返回细胞质。
细胞质基质中的蛋白质的转运
蛋白质分选运输的途径
①门孔运输:是指由特定的
分拣信号介导,并通过核
提高细胞对蛋白质的合成和利用效率 使蛋白质分子能准确定位到功能部位,使
其能准确行使其生物学功能 分选过程中伴随着对蛋白质分子的加工和
修饰,使真核细胞蛋白质分子的结构和功 能更加多样化
Thank you!
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
题目:1.用自己的语言复述课堂列出的四组关于信号肽的实验,分析其产物为何有所不同;根据这些实验结果构建的信号肽学说要点有哪些?2.请整理线粒体、质体、内膜系统、膜泡系统、细胞核等章节有关蛋白分选内容,详细描述细胞内蛋白分选机制。
1.共四组实验,在第一组(对照组)中加入含编码信号序列的mRNA,第二组中加入含编码信号序列的mRNA和SRP,第三组中加入含编码信号序列的mRNA和SRP,DP,第四组中加入含编码信号序列的mRNA和SRP,DP,微粒体。
实验结果:第一组产生含信号肽的完整多肽,第二组合成70~100氨基酸残基后,肽链停止延伸,第三组产生含信号肽的完整多肽,第四组信号肽切除,多肽链进入微粒体中。
产物不同的原因:组2:SRP 有Alu和S 两个结构域,它们同RNA 相互连接。
其中Alu结构域由SRP9 和SRP14 组成,结合到7S RNA的5'端和3'端序列。
SRP 能识别并结合在游离核糖体上新合成蛋白质的信号肽。
当它与信号肽结合后,多肽合成就暂时中止,所以会只形成70~100氨基酸残基。
组3:DP与SRP结合后,解除了SRP 对核糖体肽链合成的抑制,新生链继续合成延长。
组3:微粒体中含有内质网和核糖体,加入之后,多肽链会进入其中被加工,信号肽则被信号肽酶水解。
信号肽学说要点:分泌蛋白先在游离核糖体上开始合成-----当其N端的信号肽延伸出核糖体后,被胞质中的SRP识别并结合-----rER膜上的SR识别并结合SRP----信号肽的疏水核心与膜结合-----新形成的多肽链进入内质网----信号肽被信号肽酶水解-------新生肽链通过蛋白转运子进入内质网腔中--------核糖体移到mRNA的终止密码子,蛋白质合成结束,核糖体重新处于游离状态。
2.线粒体:线粒体中有1000 多种蛋白质,它本身的DNA 及核糖体只能合成其中少数蛋白质,其余的线粒体蛋白质都是由核DNA编码的,在胞质游离核糖体上合成后运输到线粒体中由线粒体的核糖体合成的蛋白,以共翻译运输(co-translational transport)的方式插入到线粒体内膜,在细胞质核糖体上合成的蛋白,以翻译后运输(post-translational transport)的方式转运到线粒体中。
(1)在胞质核糖体上合成的蛋白质,大都以前体形式存在。
多由N端的一段导肽和成熟形式的蛋白质组成。
(2)蛋白质通过膜时,在外膜上有专一性不很强的受体参与作用。
(3)蛋白质通过膜需要水解ATP和利用质子动势的能量过程。
(4)导肽引导蛋白质前体,在受体及转运子的作用下,通过内、外膜的接触点,运输到线粒体的基质中。
(5)导肽对所牵引的蛋白质无特异性。
(6)蛋白质运送时需要一些分子伴侣使蛋白进行折叠状态与解折叠状态的转变。
(7)前体蛋白运入线粒体后,需要蛋白酶切除导肽,再折叠成成熟蛋白。
线粒体膜上存在前体蛋白转运子,外膜上的TOM、SAM,内膜上TIM23、TIM22、OXA转运子。
TOM 转运子(translocase of the outer mitochondrial membrane)介导几乎所有线粒体蛋白通过外膜。
它能识别细胞质中的前体蛋白,并使之与胞质中的分子伴侣分离,以未折叠的构象通过TOM 的转运孔运入。
同时它还可以介导一些外膜蛋白插入外膜。
TIM转运子(translocase of the inner mitochondrial membrane)其中最主要的一个为TIM23。
TIM23 能够转运所有的线粒体基质前体蛋白,大多数内膜蛋白,以及许多膜间隙蛋白。
组成TIM23 的亚基分两类:膜组分和输入马达(import motor)。
膜组分包括Tim21、Tim50、Tim23和Tim17。
SAM 复合物位于外膜上,当线粒体外膜上的β桶状结构的前体蛋白(如孔蛋白),经TOM 转运进入膜间隙后,SAM复合物可以将它们正确折叠并插入到外膜上。
TIM22复合物位于内膜上,结构与TIM23类似,由伸入膜间隙的受体亚基和嵌入内膜的孔道亚基组成。
它负责介导部分定位于内膜的转运蛋白的插入。
包括运输ATP、ADP、磷酸进出线粒体的转运蛋白,以及其他转运蛋白。
OXA复合物位于内膜上,负责介导由线粒体自身合成的内膜蛋白的插入,以及一些运入基质的内膜蛋白的插入。
蛋白质运进线粒体的路线主要有以下4个途径:1、外膜蛋白的运输途径定位于外膜的蛋白质一般没有导肽,其末端的氨基酸序列就有识别受体的能力。
在胞质中它们与分子伴侣Hsp70 结合,呈伸展状态,到达线粒体外膜后,Hsp70 与之脱离。
接着,蛋白质通过TOM复合物进入外膜。
对于外膜上丰富的膜孔蛋白,首先由TOM 运入膜间隙,与其中的分子伴侣结合防止凝聚,然后由SAM 插入到外膜。
2、内膜的蛋白质的运输途径这类蛋白质的运输途径有3种。
第一类内膜蛋白,无导肽,但存在内部信号序列。
第二类内膜蛋白,有导肽和疏水(停止)序列。
第三类内膜蛋白,有导肽,停止序列被第二段信号序列取代。
3、膜间隙蛋白的运输途径这类蛋白质的运输途径有两种:一类蛋白质含有导肽和疏水序列,它们由TOM 和TIM23运入膜间隙,同时导肽穿入基质,被基质中的蛋白酶水解,后面的疏水序列停留在内膜,并被膜间隙的蛋白酶水解,释放成熟蛋白到膜间隙中;另一类蛋白质含有导肽和第二段信号序列,它们先经TOM、TIM23 进入基质,并切掉导肽,随后,蛋白又经OXA运送到内膜和膜间隙,其第二段信号序列最后被膜间隙的蛋白酶水解,使得成熟蛋白留在膜间隙4、基质蛋白的运输途径基质蛋白都带有导肽。
在内外膜的接触点处,与Hsp70结合的前体蛋白在ATP作用下解离,并在导肽引导下经TOM复合物进入膜间隙。
随后通过TIM23 复合物进入内膜。
最后,导肽被基质中的线粒体多肽酶水解,在基质Hsp60的帮助下最终折叠成为成熟蛋白。
质体:叶绿体蛋白的转运主要是向基质和类囊体的转运,叶绿体蛋白的转运依赖于叶绿体内外膜上的转运复合体TOC和TIC(translocase of the outer and inner chloroplast)。
定位于叶绿体基质的蛋白质,N端有基质信号序列,定位于类囊体的蛋白质,N端有两个信号序列:基质信号序列和类囊体信号序列。
蛋白质进入类囊体。
至少包括4种途径:①cpSec途径。
②cpTat途径(twin argininetranslocase,TAT),即双精氨酸转运酶途径。
③cpSRP途径。
④自发插入途径。
内膜系统:跨膜运输:蛋白运入内质网、线粒体、叶绿体、过氧化物酶体;膜泡运输:内质网、高尔基体、质膜、溶酶体、胞内体之间的蛋白运输;膜泡系统:网格蛋白包被小泡:介导蛋白质从高尔基体TGN向胞内体、溶酶体、质膜及液泡运输COPII包被小泡:内质网到高尔基体COPI包被小泡:高尔基体向内质网COPⅡ衣被小泡介导内质网到高尔基体的物质运输。
形成于内质网无核糖体处的出芽位点。
主要亚基:Sar1-GTP、Sec23、Sec24(双酸信号受体)、Sec13、Sec31。
多数跨膜蛋白直接与COPⅡ结合,少数跨膜蛋白和多数可溶性蛋白通过受体与COP Ⅱ结合。
分选信号:位于跨膜蛋白胞质面,形式多样,常包含双酸性基序Asp-X-Glu。
COPⅠ包被膜泡•在哺乳动物细胞中,抑制从内质网出芽的抗体对高尔基体膜囊之间膜泡运输没有抑制效应,说明介导两种运输的蛋白不同。
•COPI有被小泡是第一个被分离鉴定的运输小泡,GTP类似物可以促使细胞积累COPI有被小泡并可通过离心分离。
•COPI包被膜泡介导由高尔基体膜囊间及至内质网的反向运输。
•功能:回收、转运内质网逃逸蛋白(escaped proteins)返回内质网;也可介导高尔基体不同区域间的蛋白质运输。
•组成:由7种蛋白组成,αβ(KKXX膜蛋白回收信号受体)β’γδεδ。
•回收信号:膜蛋白Lys-Lys-X-X(KKXX),可溶性蛋白Lys/His-Asp-GluLeu(KDEL或HDEL)。
•KDEL信号序列受体存在于高尔基体反面管网区、COPI 、COPII 有被小泡膜上。
而KKXX信号可被COPI的包被αβ亚基识别。
•COPI包被的组装和解聚由一种GTP结合蛋白ADP核糖基化因子(ADPribosylation factor,ARF) 调控。
•内质网驻留蛋白回收模式:•KDEL受体捕获可溶性内质网蛋白,内质网跨膜蛋白KKXX被COPI包被α、β亚基识别,通过COPI有被小泡运回内质网。
•正常pH:从KDEL序列上解离;•低pH:受体结合KDEL序列。
网格蛋白有被小泡•网格蛋白包被膜泡介导高尔基体管网区向胞内体、溶酶体、液泡运送物质。
也通过胞吞从质膜向胞内体运送物质。
•高尔基体反面管网区是网格蛋白有被小泡来源。
•网格蛋白有被小泡组成:•网格蛋白----形成外层结构网架。
•结合素蛋白(AP)---- 形成内壳,待转运膜蛋白受体。
•G蛋白ARF调控网格蛋白有被小泡的组装及解离。
·当衣被小泡形成时,可溶性GTP结合蛋白dynamin(发动蛋白)聚集成一圈围绕在芽的柄部,使柄部的膜尽可能地拉近(小于1.5nm),导致膜融合,使网格蛋白有被小泡缢缩下来。
其他两种有被小泡缢缩还没发现由发动蛋白参与。
•发动蛋白突变可能导致不能产生网格蛋白有被小泡,只产生与膜连接的长颈部结合发动蛋白的芽体。
细胞核:门控通道运输:通过核孔运输;亲核蛋白的核输入信号:核定位信号(NLS) ;10 个氨基酸的短肽,指导亲核蛋白完成核输入后并不切除(NLS 、NES、信号肽和信号斑) (importinα/β、nucleoporin、Ran—GTP/GDP)亲核蛋白的入核转运:①核蛋白通过NLS识别importinα,与可溶性NLS受体importinα/β异二聚体结合,形成转运复合物;②importinβ的介导下,转运复合物与核孔复合体的胞质纤维结合;③转运复合物通过改变构象的核孔复合体从胞质面被转移到核质面;④转运复合物在核质面与Ran-GTP 结合,并导致复合物解离,亲核蛋白释放;⑤受体的亚基与结合的Ran 并与importinβ解离,Ran-GDP 返回核内再转换成Ran-GTP 状态。
mRNA 、tRNA和核糖体亚基的核输出:核输出信号nuclear export signal (NES)。