绿色荧光蛋白在转基因动物中的应用
绿色荧光蛋白GFP的显微观察及其在转基因研究中的应用

报告基因的特点
已被克隆和全序列已测定; 表达产物在受体细胞中不存在,即无背景,在被转染的细 胞中无相似的内源性表达产物;
其表达产物容易观察或能进行定量测定
常用的报告基因
基因名称 GUS 基因编码蛋白
β-D-glucuronidase(β-D葡萄糖苷酶)
检测方法
催化底物形成β-D-葡萄糖苷 酸,它在植物体中几乎无背 景,组织化学检测很稳定
实验仪器:荧光显微镜
实验步骤:
1. 2. 3.
取部分转基因幼苗放入IAA溶液中处理5-10分钟 。 无IAA处理的作为对照。 PI 染色 将植物根部浸泡在1XPI工作液中10秒左右。 用水冲洗30秒。
4. 制片 用擦镜纸擦载玻片 滴一滴水 用镊子取一棵幼苗,将根部在水中展开(可以将叶 片切除) 用镊子夹取盖玻片,从一侧轻轻盖上 5.显微镜观察 放下载物台,将载玻片放上载物台,旋转20X物镜 至光路,滤光片在明场位置(1) 将载物台上升至离物镜很近处,不要碰到物镜 寻找视野中模糊的根,下调载物台,聚焦 移动载物台,寻找根尖 转换至40X物镜,滤光片调至蓝光(3)或绿光(4 ),打开shutter,观察、拍照和保存实验结果。
3:绿色荧光蛋白--GFP
GFP最初从水母,jellyfish Aequorea victoria 中分离出 在分子生物学和细胞学领域,GFP是广 泛使用的报告基因
GFP
GFP蛋白含238 氨基酸残基, 分子量29.6KDa Fluorescent protein (荧光蛋白 ): 在蓝-紫外光的激发下产生强的 绿色荧光
普通生物学实验
GFP报告基因的显微观察及其在
绿色荧光蛋白作为报告基因在分子生物学中的应用

绿色荧光蛋白作为报告基因在分子生物学中的应用绿色荧光蛋白作为报告基因在分子生物学中的应用摘要:随着科学技术的不断更新和发展,绿色荧光蛋白在动物学、植物学、微生物学等领域的应用研究越来越广泛。
绿色荧光蛋白(green fluorescent protein,GFP)可作为报告基因,且具有分子量较小、荧光性质稳定、对生物体无毒性作用、检测时不需要底物等的特点。
本文就对荧光蛋白在分子生物学中的应用做一综述。
关键词:绿色荧光蛋白;报告基因;应用The Application of GFP As Reporter Gene In the Molecular Biology Abstract: With the upgrade and development of science and technology, the application of green fluorescent protein used in Zoology, Botany and microbiology is more extensive. As a reporter gene, GFP have some characteristics, such as low molecular weight, good fluorescent stability, non- toxicity to organisms. This paper reviews the application of GFP in the molecular biology. Key words: green fluorescent protein, reporter gene, application of GFP绿色荧光蛋白(green fluorescent protein,GFP)是一类来自于海洋生物如水母、水螅和珊瑚等腔肠动物内的一种生物发光蛋白,当受到紫外或蓝光激发时,能发射出绿色荧光。
GFP的简介和应用

GFP的简介和应用【摘要】源于多管水母属等海洋无脊椎动物的绿色荧光蛋白(GFP),是一种极具应用潜力的标记物,有着极其广泛的应用前景。
本文就GFP的理化性质、荧光特性、改进以及它在科学研究中发挥的作用进行了综述。
【关键词】绿色荧光蛋白(GFP)、标记物、荧光特性、进展、改进、应用、干细胞移植【正文】一、GFP的简介1. GFP的理化性质,荧光特性及其改进1.1 GFP的理化性质从水母体内分离到的GFP基因,长达2.6kD,由3个外显子组成,分别编码69、98和71个氨基酸。
GFP本身是一种酸性,球状,可溶性天然荧光蛋白。
Aequoria GFP分子量约27×103,一级结构为一个由238 个氨基酸残基组成的单链多肽;而Renilla GFP是分子量为54kD的同型二聚体。
两种GFP有不同的激发光谱,Aequoria GFP在395 nm具有最高光吸收峰,肩峰为473 nm;Renilla GFP在498 nm具有强烈的光吸收,肩峰为470 nm。
两种GFP含有相同的生色团,发射光谱基本相同(λmax= 508~ 509 nm)。
GFP性质极其稳定,易耐受高温处理,甲醛固定和石蜡包埋不影响其荧光性质。
其变性需在90℃或pH<4.0或pH>12.0的条件下用6mol/L盐酸胍处理,一旦恢复中性环境,或去除变性剂,虽然变性的蛋白质并不能完全复性,但是复性蛋白质同天然蛋白质对温度、pH变化的耐受性、抗胰蛋白酶消解的能力是相同的。
更重要的是,它们在很大的pH范围内的吸收、发射光谱也是相同的。
Renilla GFP的稳定性就更为显著。
它在上述一系列的变性条件下都很稳定,不易变性。
根据Sheen等的研究,GFP在受体内表达时,其稳定性并不亚于CAT 蛋白,因而可以得到持续时间较长的荧光。
1.2 GFP的荧光原理GFP的性质和发射光谱的稳定性是同其生色团结构的稳定性密不可分的。
GFP表达后折叠,在氧存在的条件下,使66位氨基酸残基的α、β键间脱氢。
dfhbi 1t类绿色荧光蛋白

绿色荧光蛋白(Green Fluorescent Protein,GFP)是一种具有绿色荧光的蛋白质,广泛应用于生物学领域的标记和成像技术中。
绿色荧光蛋白的研究和应用已经成为生命科学领域中的热点和前沿课题。
在这篇文章中,我们将深入探讨绿色荧光蛋白的种类、结构、功能和应用。
1. 绿色荧光蛋白的种类绿色荧光蛋白是由Aequorea victoria(水母)发光器官中分离出来的一种蛋白质。
根据不同的来源和结构特点,绿色荧光蛋白可以分为多种类别,包括标准GFP、改良GFP、超变荧光蛋白和环状GFP等。
每种类型的绿色荧光蛋白都具有不同的荧光特性和适用范围。
2. 绿色荧光蛋白的结构绿色荧光蛋白的结构是其功能的基础。
它是一个由238个氨基酸组成的蛋白质,包括一个β桶结构和一个共轭双键序列。
在特定的条件下,它可以通过自发性氧化反应形成荧光色团,并发出绿色的荧光。
绿色荧光蛋白的结构和光学特性为其在生物标记和成像领域的应用奠定了基础。
3. 绿色荧光蛋白的功能作为一种生物标记物,绿色荧光蛋白的主要功能是在转基因生物中标记特定的细胞、器官或组织,以便于研究者对其进行观察和分析。
通过转基因技术,研究人员可以将绿色荧光蛋白基因导入到目标生物体中,从而实现对其活体成像和实时监测。
绿色荧光蛋白在蛋白质定位、蛋白质-蛋白质相互作用和基因表达调控等方面也发挥着重要作用。
4. 绿色荧光蛋白的应用绿色荧光蛋白的广泛应用领域包括但不限于以下几个方面:a. 细胞成像与实时监测:通过转基因技术将绿色荧光蛋白标记到感兴趣的细胞中,可以实现对其活体成像和实时监测,从而揭示生物体内细胞的运动、分化和凋亡等过程。
b. 蛋白质定位与跟踪:通过融合绿色荧光蛋白与感兴趣蛋白质,可以实现对蛋白质在生物体内的定位与跟踪,从而研究其功能和代谢途径。
c. 蛋白质-蛋白质相互作用研究:利用双融合蛋白技术或FRET技术,可以实现对蛋白质-蛋白质相互作用的实时观察和分析,为研究蛋白质分子机制提供了有力工具。
绿色荧光蛋白在转基因动物中的应用

绿色荧光蛋白在转基因动物中的应用杨军廷 201141804054 华大基因摘要来源于水母 (AequoreaVictoria)的绿色荧光蛋白 (Greenfluorescentprotein ,GFP) ,现已成为细胞生物学和分子生物学中应用最广泛的分子标记之一。
其内源性荧光基团在受到紫外或蓝光激发时可发现清晰可见的绿光。
由于检测方便 ,对生物体基本没有毒性 ,在很多领域已有取代LacZ ,荧光素酶等传统标记方法的趋势 ,在制作转基因动物过程中更是如此。
本文综述了GFP在标记目的基因、筛选阳性胚胎等方面的应用关键词绿色荧光蛋白;转基因在研究基因的表达或蛋白质的定位与时序变化时常用荧光物质或报告基因作为标记。
传统的荧光标记是通过纯化蛋白质再共价结合到荧光染料上!此方法难以控制化学剂量和染料附着部位!若该标记蛋白用于活细胞内检测!则难以通过细胞膜。
因此,该方法已基本淘汰现在常用的报告基因如荧光素酶(LUX)基因和β-葡萄糖苷酶(GUS)基因也不尽人意。
LUX所检测到的荧光产生部位不一定反映荧光素酶基因的特异表达部位;GUS则需要昂贵的反应底物且由于其它因素的干扰。
反应颜色的深浅有时不能说明GUS活性的高低或有无.源于水母等海洋无脊椎动物的绿色荧光蛋白(GFP)可吸收蓝光而后发出绿光是生物发光现象中能量传递的受体之一。
它克服了上述缺陷,具有灵敏度高、操作简便,不需要添加任何底物或辅助因子,不使用同位素,也不需要测定酶的活性等优点。
已成为目前最优良的标记基因之一。
1GFP的生化性质及其发光原理1.1 GFP的发光特性GFP吸收的光谱,最大峰值为395nm(紫外),并有一个峰值为470nm的副峰(蓝光);发射光谱最大峰值为509nm(绿光),并带有峰值为540nm的侧峰(Shouder)。
GFP的光谱特性与荧光素异硫氰酸盐(FITC)很相似,因此为荧光素FITC设计的荧光显微镜滤光片组合同样适用于GFP观察。
GFP的发展历程

原来世间真有扫地僧
普瑞泽做出GFP本身就能发荧光,并不需要水母的其它蛋白 处理加工的假设。
他希望能找到并且克隆出编码GFP 的 DNA 序列,并能 将该基因利用基因工程技术在其他生物中表达出来。
验证这一想法的第一步就是克隆GFP的基因。基因是蛋 白的源程序。有了基因,后续工作就容易地多。
结果很让他失望:看来下村修的理论是对的。 GFP自己并不能发绿荧光! 但他还是把克隆GFP基因的过程和结果写成一篇论文,于1992年发表。
普瑞泽1992年发表GFP基因的文章后, 离开科学界。离开前,他将GFP基因 寄给了哥伦比亚大学的查尔菲和加州 大学圣地亚哥分校的钱永健
与普瑞泽不同的是,查尔菲的学生Ghia Euschirken使用PCR技术精 确地将GFP基因插到表达质粒上。她利用T7启动子来起始GFP在大肠杆菌 中的表达,并使用IPTG进行诱导,用长波紫外线(UY)光源照射后,发 现大肠杆菌显示出了明亮的绿色荧光。
酶切后均显示含有1kb的插入物。
A,victoria-gfp mRNA的Northern分析
基因组克隆的分离鉴定
对基因组克隆子λGFP2中的 EcoRI-BamHl和重叠的hindll 片段进行测序,并与gfp10 cDNA进行比较,以检查基 因的结构。gfp mRNA剪接 连接的nt序列与全长序列非 常吻合(如图)。
ChalfiΒιβλιοθήκη et al,1994,science
查尔菲团队迅速地 用GFP标记了线虫,观 察它在线虫体内神经细 胞的表达。
这一成果1994年发 表在《科学》杂志上。
Chalfie et al,1994,science
绿色荧光蛋白(GFP)的基因克隆及表达

绿色荧光蛋白(GFP)的基因克隆及表达摘要绿色荧光蛋白(GFP)是一类存在于包括水母、水螅和珊瑚等腔肠动物体内的生物发光蛋白。
采用PCR技术,对实验室提供的质粒pEGFP-N1中的目的基因进行扩增。
所得PCR产物和质粒pET-28b经过BamH I和Nde I双酶切后,用琼脂糖凝胶电泳法检测酶切产物的酶切情况并回收凝胶,再利用T4DNA连接酶将目的基因与载体连接起来,得到重组质粒。
将重组质粒导入克隆菌E. coli DH5a中培养扩增,提取阳性菌落质粒进行重组子鉴定,进而导入表达菌E. coLi BL-21大肠杆菌感受态细胞中,经IPTG诱导目的基因表达产生绿色荧光蛋白。
关键词:绿色荧光蛋白 PCR 基因克隆表达1.前言1.1绿色荧光蛋白(green fluorescent protein,GFP)绿色荧光蛋白是一类存在于包括水母、水螅和珊瑚等腔肠动物体内的生物发光蛋白。
当受到紫外或蓝光激发时,GFP 发射绿色荧光[1]。
1.2 GFP 的结构GFP中央是一个圆柱形水桶样结构,如图二。
长420 nm,宽240 nm,由11 个围绕中心α螺旋的反平行β折叠组成,荧光基团的形成就是从这个螺旋开始的,桶的顶部由3个短的垂直片段覆盖,底部由一个短的垂直片段覆盖,对荧光活性很重要的生色团则位于大空腔内。
发色团是由其蛋白质内部第65-67位的Ser-Tyr-GLy自身环化和氧化形成。
1.3 GFP的研究应用GFP可标记细胞和蛋白质,具有广泛的应用前景。
GFP及其突变体已被广泛应用于基因表达调控、蛋白质空间定位、生物分子之间相互作用、转基因动物]2[等方面。
基于新型功能荧光蛋白的光学分子成像技术的发展,为在活细胞乃至活体动物内研究基因表达和蛋白质功能提供了更多的选择空间。
GFP还用于观察微生物、发育机理研究、细胞筛选、免疫学等方面。
本实验是利用实验室提供的质粒pEGFP-N1,其结构如图三所示。
其上有所用酶的酶切位点。
首例绿色荧光蛋白转基因克隆猪降生

CATALOGUE目录•研究背景与意义•转基因克隆技术介绍•绿色荧光蛋白在生物医学中的应用•首例绿色荧光蛋白转基因克隆猪的研究过程•转基因克隆猪的应用前景与挑战•研究结论与展望绿色荧光蛋白克隆技术探索转基因克隆技术的应用01提高猪的品种质量02推进医学研究03转基因技术定义通过人工合成或从其他生物体中提取出目的基因,将其导入到目标生物体的受精卵或胚胎中,从而使后代表现出新的性状。
转基因技术流程转基因技术的意义转基因技术克隆技术克隆技术定义克隆技术的意义转基因克隆猪的研究现状研究背景研究进展前景展望绿色荧光蛋白是一种水母发光蛋白,具有高亮度、稳定性、可诱导发光等特性。
绿色荧光蛋白在紫外光激发下可发出明亮的绿色荧光,具有高灵敏度和低毒性的优点。
绿色荧光蛋白在细胞生物学研究中常被用作标记物,以追踪细胞活动和行为。
绿色荧光蛋白的特性绿色荧光蛋白可以作为标记物,用于监测和鉴定转基因克隆动物的成功率和效率。
转基因克隆技术疾病模型研究药物筛选细胞治疗通过将绿色荧光蛋白与其他基因融合,可以研究疾病的发生和发展过程。
利用绿色荧光蛋白报告基因系统,可以快速筛选出对特定疾病有治疗作用的候选药物。
利用绿色荧光蛋白标记细胞,可以追踪其在体内的分布和存活情况,为细胞治疗提供可视化工具。
绿色荧光蛋白在生物医学中的应用领域研究目标研究方案研究目标与方案实验过程实验结果实验过程与结果意义绿色荧光蛋白转基因克隆猪的诞生是一项重要的生物技术突破,它不仅展示了基因编辑技术在生物医学研究领域的巨大潜力,也为相关领域的发展提供了强有力的工具。
价值绿色荧光蛋白转基因克隆猪可以作为生物医学研究的实验模型,用于模拟和研究人类遗传疾病的发生和发展机制。
同时,它也为药物筛选及移植医学等领域提供了新的实验平台,有助于推动相关领域的发展和创新。
研究成果的意义与价值改善猪的疾病抵抗力生产具有特定性状的猪医学和生物医学研究010203转基因克隆猪的应用前景技术成本和效率目前转基因克隆技术的成本较高,效率也较低,需要进一步的技术突破和改进,才能实现大规模的应用和推广。
绿色荧光蛋白的应用及其最新研究进展

绿色荧光蛋白的应用及其最新研究进展一、关键词:绿色萤光蛋白、酵母双杂交系统、流式细胞仪、下修村、马丁·查尔菲、钱永健二、背景2008年10月8日,三位美国科学家——伍兹霍尔海洋生物学实验室(Woods Hole Marine Biological Laboratory, MBL)的Osamu Shimomura、哥伦比亚大学(Columbia University)的Martin Chalfie以及加州大学圣地亚哥分校(University of California, San Diego)的钱永健(Roger Y onchien Tsien),因在研究和发现绿色荧光蛋白(green fluorescent protein,GFP)方面做出突出贡献而获得诺贝尔化学奖。
绿色荧光蛋白(green fluorescent protein, GFP)最早由日裔科学家下村修于1962年在水母(Aequorea victoria )中发现。
而后马丁·查尔菲则证明了GFP在作为多种生物学现象发光遗传标记方面的应用价值。
钱永健阐明了GFP发光的机制,并且发现了除绿色之外可用于标记的其它颜色。
他对细胞生物学和神经生物学领域的贡献具有划时代的意义。
他的多色荧光蛋白标记技术让科学家能够用不同颜色对多个蛋白和细胞进行标记,从而实现了同时对多个生物学过程进行追踪。
现在,三位科学家的研究成果已经作为标记工具在生物科学中得到广泛应用。
三、GFP的主要性能GFP在蓝色波长范围的光照激发下发出绿色荧光,其发光过程需要冷光蛋白质Aequorin 的帮助,而且,这个冷光蛋白质可与钙离子(Ca2+)相互作用。
GFP的激发光谱在400nm 附近有一个主激发峰,在470nm附近有一个次激发峰。
发射光谱在505nm附近有一尖锐的主发射峰,在540nm附近有一肩峰。
在Aequorea victoria 中发现的野生型绿色荧光蛋白的分子量较小,由238个氨基酸残基组成,仅为27~30kDa,而编码GFP的基因序列也很短,为2.6kb。
绿色荧光蛋白(GFP)标记亚细胞定位与农杆菌瞬时侵染方法

绿色荧光蛋白(GFP)标记亚细胞定位与农杆菌瞬时侵染方法转基因技术,生物定向改良,分子育种基因工程和育种的最有效途径,转基因技术有农杆菌介导法、花粉通道法、显微注射法、基因枪法、离子束介导法。
农杆菌瞬时侵染方法(1) 转入目的载体的农杆菌EHA105单菌落接入4mL LB-50ug/mL-Kan, 30℃摇床震荡培养20h;(2) 转接3mL菌液进入30 mL LB-50ug/ml-Kan, 30℃摇床震荡培养6-8h;(3) 收集菌体,用10mL 1/2 Ms(1.5% sucrose)液体培养基悬浮菌体,加入5ul 100mM As, 混合均匀;(4) 切洋葱内表皮,撕开后内表皮,切开的宽度控制在0.5-0.7cm, 浸入农杆菌菌液,浸泡30min;(5) 滤纸轻轻吸去洋葱表皮表面可见的农杆菌菌液,放置在1×Ms+1.5%Sucrose固体培养基上,25℃培养18-20h;(6) 用1XPBS (pH7.0)溶液轻轻漂洗洋葱表皮两次,无菌水漂洗1次;(7) 在荧光显微镜下观察,采用激光共聚集显微镜采集图像;绿色荧光蛋白(GFP)标记亚细胞定位一、原理利用绿色荧光蛋白(GFP)来示踪胞内蛋白的技术。
利用GFP融合蛋白技术来进行活细胞定位研究是目前较为通行的一种方法,在光镜水平进行研究,不需要制样,没有非特异性标记的影响。
并且GFP的分子量为27kD,经激光扫描共聚集显微镜激光照射后,可产生一种绿色荧光,从而对蛋白质进行精确定位。
激光扫描共聚焦显微镜(Laser Scanning Confocal Microscope, LSCM, 以下简称共聚焦显微镜)因其独特的设计原理,有效地排除了非焦平面信息,提高了分辨率及对比度,使图像更为精确清晰,因此极其适于进行活细胞内蛋白质、核酸等定位及活体动态研究。
二、主要步骤1.真核表达载体的构建①引物设计利用引物设计软件,根据pEGFP-N1的酶切位点设计目的基因引物:②载体构建将PCR产物酶切后插入pEGFP-N1,得到表达目的基因与EGFP融合蛋白质的真核表达载体。
转基因荧光标记种类

转基因荧光标记种类
转基因荧光标记技术是将荧光蛋白基因导入细胞或生物体内,使其表达荧光蛋白,从而实现对细胞或生物体的可视化标记。
这种技术广泛应用于生物医学研究,如基因表达、细胞迁移、蛋白质相互作用等领域。
常用的荧光蛋白包括:
1. 绿色荧光蛋白(GFP)
来自于绿色发光水母,发射绿色荧光。
GFP是最早发现和应用的荧光蛋白,也是最常用的荧光标记蛋白。
2. 红色荧光蛋白(RFP)
来自珊瑚和海葵,发射红色荧光。
RFP与GFP荧光颜色互补,常用于双荧光标记实验。
3. 黄色荧光蛋白(YFP)
来源于GFP的突变体,发射黄色荧光。
YFP的激发和发射波长与GFP有所区别,可用于多色荧光标记。
4. 蓝色荧光蛋白(BFP)
来源于GFP的突变体,发射蓝色荧光。
BFP与其他荧光蛋白的颜色差异较大,适合多色荧光标记。
5. 红荧光蛋白(mCherry、mRFP)
来源于珊瑚和海葵,发射红色荧光。
相比于传统RFP,这些改良型红荧光蛋白具有更高的亮度和光稳定性。
除了上述常见的荧光蛋白,还有一些新型荧光蛋白被开发出来,如远红外荧光蛋白、近红外荧光蛋白等,可用于深组织成像等特殊应用。
根据实验需求,研究人员可以选择合适的荧光蛋白进行标记和观察。
绿色荧光蛋白在基因工程教学中的应用

摘要基因工程是生命科学相关专业学生的必修课程之一。
然而,由于基因工程的基本知识点与部分分子生物学的原理密切相关,导致教学内容过于抽象,学生不易深入地理解课程内涵。
通过设计绿色荧光蛋白在哺乳动物细胞中的转染表达实验,将基因工程的基本理论应用到实验操作中,一方面可以帮助学生更好地理解基因工程的基本原理和操作流程,另一方面将会激发学生探索未知的浓厚兴趣。
关键词绿色荧光蛋白;基因工程;教学改革Application of Green Fluorescent Protein in the Teaching of Genetic Engineering //XU Yinfeng,ZHOU Lu,WAN Wei Abstract Genetic engineering is one of the compulsory courses for students in the field of life science.However,the basic knowledge of genetic engineering is closely related to some principles of molecular biology,which leads the teaching content to be quite abstract.Almost all the students share the same feeling that the content is hard to understand.By designing the experiment,namely transfection and expression of green fluo‐rescent protein in mammalian cells,we try to apply the abstract theory of genetic engineering to the experimental operation during lecturing the course.On one hand,it can help studentsbetter understand the basic principles and operation process of genetic engineering,on the other hand,it will stimulate stu‐dents’strong interest in exploring the unknown in science.Key words green fluorescent protein;genetic engineering;teaching reform绿色荧光蛋白(Green fluorescent protein,GFP)是一类存在于海洋腔肠动物体内的生物发光蛋白,最早是从水母中分离得到的。
绿色荧光蛋白基因
综合上述资料,请你给基因下个定义?
结论:基因是有遗传效应的DNA 片段,它是生物体遗传的功能单位和 结构单位.
遗传效应:指具有复制、转录、翻译、
重组、突变及调控等功能。
二、DNA片段中的遗传信息
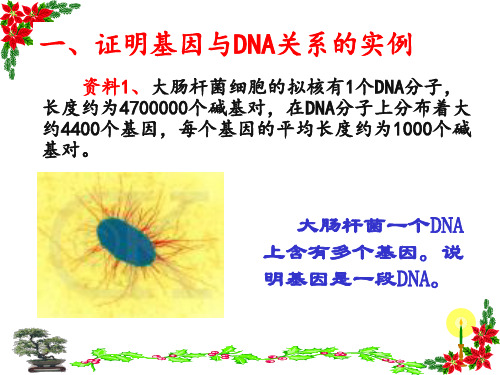
一、证明基因与DNA关系的实例
资料1、大肠杆菌细胞的拟核有1个DNA分子, 长度约为4700000个碱基对,在DNA分子上分布着大 约4400个基因,每个基因的平均长度约为1000个碱 基对。
大肠杆菌一个DNA 上含有多个基因。说 明基因是一段DNA。
资料2、生长在太平洋西北部的一种海蛰能发 出绿色荧光,这是因为海蛰的DNA分子上有一段长 度为5170个碱基对的片段---绿色荧光蛋白基因。 转基因实验表明,转入了海蛰的绿色荧光蛋白基 因的转基因鼠,在紫外线的照射下,也能象海蛰 一样发光。
转基因烟草
美国密苏里大学培育出四只含有荧光水母基因的 小猪。左为荧光小猪,右为正常小猪。
美科学家研制出世界上第一只转基因蝴蝶
中国首批t-PA转基因羊在山东东营诞生
t-PA是目前治疗急性心肌梗塞最好的溶血栓药物,如果从 国外进口,价格非常昂贵,由动物的乳腺生产出含有t-PA的药 物蛋白,价格就会大大降低。
DNA中的特定一段,但是一段DNA并不一定是基
因。
非基因片段
--ATGCATGCATCCATGCTAGCCATCCCTAAGGACAG----TACGTACGTAGGTACGATCGGTAGGGATTCCTGTC---
基因
基因
资料4、不少人认为,人和动物的胖瘦是 由基因决定的。近来的科学研究发现,小鼠体 内的HMGIC基因与肥胖直接相关。具有HMGIC基 因缺陷的实验鼠与作为对照的小鼠,吃同样多 的高脂肪食物,一段时间后,对照组的小鼠变 得十分肥胖,而具有HMGIC基因缺陷的实验鼠 体重仍然保持正常。
绿色荧光蛋白在转基因动物研究中的应用

绿色荧光蛋白在转基因动物研究中的应用绿色荧光蛋白(GFP)是一种来自水母的蛋白质,具有独特的荧光性质,可以发出绿色荧光。
近年来,GFP被广泛应用于生物学研究中,特别是在转基因动物研究中得到了广泛应用。
利用GFP基因的表达,科学家可以追踪细胞、组织以及整个生物体系的运动和功能。
通过将GFP基因转入目标细胞或组织中,科学家可以用荧光显微镜观察其在生物中的位置和运动轨迹,繁殖情况以及基因表达水平等重要信息。
在转基因动物研究中,GFP的应用尤其重要。
通过将GFP基因转入小鼠、果蝇等模式动物中,科学家可以追踪这些动物的胚胎发育、器官生长、细胞分化以及疾病模型等过程。
此外,还可以利用GFP的荧光特性,观察细胞内各种蛋白质的表达情况,从而了解其在疾病发生发展中的作用,为药物开发提供参考。
总之,GFP在转基因动物研究中的应用,不仅能够促进科学家对于生物体系的认识和了解,还能够为疾病治疗提供新的思路和方法。
随着技术的进步,GFP的应用前景将会更加广阔。
- 1 -。
《2024年用绿色荧光蛋白和肝再生增强因子基因进行转基因绵羊胚胎的研究》范文

《用绿色荧光蛋白和肝再生增强因子基因进行转基因绵羊胚胎的研究》篇一一、引言随着生物技术的飞速发展,转基因技术已成为现代生物学研究的重要手段。
转基因技术通过将外源基因导入动物体内,实现基因的修饰和改造,从而获得具有特定性状或功能的转基因动物。
绿色荧光蛋白(GFP)和肝再生增强因子(HRE)基因作为两种重要的外源基因,在转基因研究中具有广泛的应用前景。
本研究以绵羊为研究对象,利用绿色荧光蛋白和肝再生增强因子基因进行转基因研究,旨在探讨其在胚胎发育及肝再生等方面的应用。
二、材料与方法1. 材料本实验所需的材料包括:绿色荧光蛋白基因、肝再生增强因子基因、绵羊胚胎、培养基、转染试剂等。
2. 方法(1)基因构建与转染:首先构建包含绿色荧光蛋白基因和肝再生增强因子基因的重组载体,然后通过显微注射法将重组载体导入绵羊胚胎中。
(2)胚胎培养:将转染后的胚胎置于培养基中,进行体外培养,观察其发育情况。
(3)转基因动物鉴定:通过PCR、荧光显微镜等方法,检测转基因动物的基因型及表达情况。
(4)肝再生实验:对转基因动物进行肝切除手术,观察其肝再生能力。
三、实验结果1. 胚胎发育情况转染后的胚胎在培养过程中,其发育情况良好,能够正常分化成囊胚和胎膜等组织。
绿色荧光蛋白基因在转基因胚胎中成功表达,使胚胎在荧光显微镜下呈现出明显的绿色荧光。
2. 转基因动物鉴定通过PCR和荧光显微镜等方法,成功检测到转基因动物的基因型及表达情况。
绿色荧光蛋白基因在转基因动物体内稳定表达,而肝再生增强因子基因的表达情况也得到了验证。
3. 肝再生实验结果对转基因动物进行肝切除手术后,发现其肝再生能力较非转基因动物有显著提高。
这一结果表明肝再生增强因子基因的转染确实有助于提高动物的肝再生能力。
四、讨论本研究利用绿色荧光蛋白和肝再生增强因子基因进行转基因绵羊胚胎的研究,探讨了其在胚胎发育及肝再生等方面的应用。
结果表明,通过显微注射法将外源基因导入绵羊胚胎中,可实现基因的稳定表达。
绿色荧光蛋白基因在水稻遗传转化中的应用

M 】 2 3
究表明 , G F P基因能够 在转基 因甜 橙 、 转基 因 玉米原 生质
体 J 、 病 毒感染 的烟草叶 片 一 以及水 稻 I 9 等植 物 中瞬 间
表达。本研究将绿色荧光蛋 白基 因通过农杆菌介 导法导入水
江苏农业科学
2 0 1 3年第 4 l卷第 4期
一 3 5一
马旭俊 , 刘春娟 , 吕世博, 等.绿色荧光蛋 白基 因在水稻遗传转化 中的应用[ J ] . 江苏农业科学 , 2 0 1 3 , 4 1 ( 4 ) : 3 5— 3 7
绿色荧光蛋 白基 因在水稻遗传转化 中的应 用
马旭 俊 ,刘春娟 ,吕世 博 ,张 超
因植株根 中绿色荧光蛋 白基 因的表达 。在荧 光显微镜 下观察到 了绿色荧光 蛋 白基 因的表达 , 转基 因水稻愈 伤组织和 根具有很高 的绿色荧光信号 。结果表 明, 在水稻遗传转化研究 中 , 绿 色荧光 蛋 白基 因既可用 于检 测基 因的瞬 间表达 ,
也可用于检测基 因在细胞 中的稳定表达 。
( 林木遗传育种 国家重点实验室/ 东北林业大学 , 黑龙江哈尔滨 1 5 0 0 4 0 )
摘要 : 本研究构建 了含有 编码 绿色荧光蛋 白基 因的植物表 达载体 , 以愈伤组织作 为外植 体 , 采 用农杆菌 介导法将 携带绿色荧光蛋 白的植物表达载体转入水 稻品种秀水 1 1中。分 析 了转化 1个月 的抗 性愈伤组 织 以及 2个月 的转基
1 4 8 9b p 9 2 5 b p
p C A MB I A1 3 0 0 、 p E G F P— N 1 、 农杆菌 E HA 1 0 5 。
绿色荧光蛋白转基因小鼠来源肌卫星细胞的体外培养及体内示踪

第 1作 者 E ma : k l 8 eh nt - i d l l @y a . e l △通 讯 作 者 , - i y n: t E mal ag mmu e u c :z @ . d .n
血 清 ( ic ) 马 血 清 ( c n ) Ha F 0培 养 基 Gbo , Hyl e , m- 1 o
1 2 原代 细胞 的 分 离培 养及 纯化 .
取对 数生 长期 P 0代细 胞 , 酶 消化 收 集 , B 1 胰 P S洗 涤并 离心 ( 0 / n mi) 收 集 后 , 入 至 4 10 0rmi X5 n , 加 ℃预 冷 7 乙醇 固定 约 3 n 以浓度 为 1mg ml RNA O 0 mi , / 的
gnc u e C 7 L e imo s ( 5 B /Ka ̄ei-GF ) adte efr i iot c ig 一at E P , n np r m vv r kn .Meh d : ru hdf rni d einmeh d n h o n a to s Tho g iee t l h s tl C , . n e k f r e gijce t h ii i a tr r sl o u emo s l ae t c P I e d MS s1 2 3 a d 4 w e s t i t i o t et a s n ei c f d u e c O r a e a e b n ne d n b l o mu e n
肌卫 星细 胞 ( sl stlt el, C ) mu ce ael ec i MS s 是骨 骼 i s
色荧 光蛋 白( re u rse t rti , P 转基 因小 ge nf oec n oe GF ) l p n
肌 内最 主 要 的 干 细胞 成 分 , 于肌 纤 维 的 肌 膜 和 基 底 位 膜 之 间 , 不仅 负责 骨 骼 肌 生后 的生 长 发育 , 它 也是 骨 骼 肌损 伤后 再生 修 复 的 主 要 承 担 者 。近 期 的研 究 表 明 ,
利用绿色荧光蛋白GFP作为报告基因检测转基因植物

第2 7卷 第 2期
20 0 8年 3月
食 品 与 生 物 技 术 学 报
J r a fFo d S in e a o e hn l g ou n lo o c e c nd Bi t c o o y
Vo _ 7 NO 2 l2 .
Usng Gr e u r s e t Pr t i s a Re r e n o i e n Fl o e c n o e n a po t r Ge e t
De e tTr n g ni a t t c a s e c Pl n
Y A NG i — M ng yu,
Ma. r 2 8 00
文 章 编 号 :6 318 (0 8 0—0 80 1 7—6 92 0 )20 9 —5
利用绿色荧光蛋 白 GF P作为报告基 因 检 测 转 基 因植 物
杨 明瑜 , 刘 翔 , 褚 华 硕 ,品科 学 与营养 工程 学院 ,北 京 1 0 8 ) 0 0 3 摘 要 : P作 为报 告 分子 被 广 泛 地 运 用在 转基 因植 株 检 测 中。利 用 P R技 术 扩 增得 到番 茄 GF C B xihbtr1 因( e I ) 亚 克隆到含 GF a ii - 基 n o L B一 , 1 P基 因的 p GF E P载体 上 , 一 步将 中间载 体构 建 到 进 植 物表 达载 体 p I2 上 。重 组 p I2 - e I1E P经农 杆 菌介导 的 叶盘转 化 法转 化烟 草 , B1 1 B 1 1L B-- GF 分化
LI Xi n U a g,
CH U ua s uo, W A NG a — a g, H —h Xio gu n
Z U e — u , H W n h a
绿色荧光蛋白在转基因动物研究中的应用

绿色荧光蛋白在转基因动物研究中的应用
李振林
【期刊名称】《第一军医大学分校学报》
【年(卷),期】2004(27)2
【摘要】来源于水母(Aequorea Victoria)的绿色荧光蛋白(Green fluorescent protein, GFP),现已成为细胞生物学和分子生物学中应用最广泛的分子标记之一.其内源性荧光基团在受到紫外或蓝光激发时可发现清晰可见的绿光.由于检测方便,对生物体基本没有毒性,在很多领域已有取代LacZ,荧光素酶等传统标记方法的趋势,在制作转基因动物过程中更是如此.本文综述了GFP在标记目的基因、筛选阳性胚胎等方面的应用.
【总页数】5页(P207-211)
【作者】李振林
【作者单位】第一军医大学分校解剖学教研室,广东省广州市,510315
【正文语种】中文
【中图分类】R32.9.27
【相关文献】
1.绿色荧光蛋白转基因大鼠骨髓间充质干细胞在急性脊髓损伤大鼠中的迁移和分化[J], 高瑞;袁文;王新伟;杨立利;陈华江
2.绿色荧光蛋白在转基因研究中的应用 [J], 张雨丽;张桂征;苏红梅;蒙艺英;闭立辉
3.绿色荧光蛋白基因(GFP)在抗虫转基因植物研究中的应用 [J], 朱生伟;秦红敏;孙敬三;田颖川
4.绿色荧光蛋白及其在转基因动物研究中的应用 [J], 李夏;陈素文;喻达辉
5.绿荧光蛋白及其在转基因动物研究中的应用 [J], 刘建忠;李宁
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
绿色荧光蛋白在转基因动物中的应用杨军廷 201141804054 华大基因摘要来源于水母 (AequoreaVictoria)的绿色荧光蛋白 (Greenfluorescentprotein ,GFP) ,现已成为细胞生物学和分子生物学中应用最广泛的分子标记之一。
其内源性荧光基团在受到紫外或蓝光激发时可发现清晰可见的绿光。
由于检测方便 ,对生物体基本没有毒性 ,在很多领域已有取代LacZ ,荧光素酶等传统标记方法的趋势 ,在制作转基因动物过程中更是如此。
本文综述了GFP在标记目的基因、筛选阳性胚胎等方面的应用关键词绿色荧光蛋白;转基因在研究基因的表达或蛋白质的定位与时序变化时常用荧光物质或报告基因作为标记。
传统的荧光标记是通过纯化蛋白质再共价结合到荧光染料上!此方法难以控制化学剂量和染料附着部位!若该标记蛋白用于活细胞内检测!则难以通过细胞膜。
因此,该方法已基本淘汰现在常用的报告基因如荧光素酶(LUX)基因和β-葡萄糖苷酶(GUS)基因也不尽人意。
LUX所检测到的荧光产生部位不一定反映荧光素酶基因的特异表达部位;GUS则需要昂贵的反应底物且由于其它因素的干扰。
反应颜色的深浅有时不能说明GUS活性的高低或有无.源于水母等海洋无脊椎动物的绿色荧光蛋白(GFP)可吸收蓝光而后发出绿光是生物发光现象中能量传递的受体之一。
它克服了上述缺陷,具有灵敏度高、操作简便,不需要添加任何底物或辅助因子,不使用同位素,也不需要测定酶的活性等优点。
已成为目前最优良的标记基因之一。
1GFP的生化性质及其发光原理1.1 GFP的发光特性GFP吸收的光谱,最大峰值为395nm(紫外),并有一个峰值为470nm的副峰(蓝光);发射光谱最大峰值为509nm(绿光),并带有峰值为540nm的侧峰(Shouder)。
GFP的光谱特性与荧光素异硫氰酸盐(FITC)很相似,因此为荧光素FITC设计的荧光显微镜滤光片组合同样适用于GFP观察。
尽管450~490nm(蓝光)是GFP的副吸收峰,但由于长波能量低,细胞忍受能力强,因此更适合于活体检测.1.2GFP的性质特点GFP荧光极其稳定,在激发光照射下,GFP抗光漂白(Photobleaching)能力比荧光素(fluorescein)强,特别在450~490nm蓝光波长下更稳定。
GFP需要在氧化状态下产生荧光,强还原剂能使GFP转变为非荧光形式,但一旦重新暴露在空气或氧气中,GFP荧光便立即得到恢复。
而一些弱还原剂并不影响GFP荧光.中度氧化剂对GFP荧光影响也不大,如生物材料的固定,脱水剂戊二酸或甲醛等。
GFP融合蛋白的荧光灵敏度远比荧光素标记的荧光抗体高,抗光漂白能力强,因此更适用于定量测定与分析。
但因为GFP不是酶,荧光信号没有酶学放大效果,因此GFP灵敏度可能低于某些酶类报告蛋白。
由于GFP荧光是生物细胞的自主功能,荧光的产生不需要任何外源反应底物,因此GFP作为一种广泛应用的活体报告蛋白,其作用是任何其它酶类报告蛋白无法比拟的。
1.3GFP的发光原理GFP之所以能产生绿色荧光是由于其蛋白分子多肽链内含有特殊的生色基团结构。
即第65-67位氨基酸Ser-Tyr-Gly链内环化加氧形成对羟苯甲基咪唑环酮和周围其它3个氨基酸形成六肽生色团。
但上述3个氨基酸残基是如何控制光谱的特征目前还不十分清楚。
体外试验时,当Ca2﹢与来自水母的发光蛋白质结合时,便可以观察到一种蓝光。
这种分子内的反应受一种在结合了Ca2﹢后转化为荧光素酶蛋白质的催化,产生CO2和蓝色荧光蛋白(BFP)在反应中也是一种发光体!如果在反应体系中加入GFP即可以观察到绿光!这一体外模拟试验与水母自然的发光现象一致。
总的过程可用下式表示1.4GFP作为标记的优点1.4.1体积小只有LacZ的1/5,表达无种属限制,这是它能广泛应用的重要基础之一。
一系列与GFP的N-端、C-端融合的蛋白质的性质研究已证明:融合蛋白质同时具有GFP的荧光特性和结合蛋白质的功能。
1.4.2 荧光表达稳定即使是在甲醛或戊二醛固定的标本中,GFP的荧光仍能保持稳定但固定过程中使用了酸或过量的有机溶剂,则会破坏荧光。
在各种制备切片的方法中,冰冻切片是效果最好的。
标本可在﹣70℃保存6周以上而荧光不受影响1.4.3有多种突变体不仅可与其它标记物,也可用不同突变体同时进行多种标记。
1.4.4检测方便常用荧光显微镜,激光扫描共聚焦显微镜(Iascer-scanning confocal microscope,ISCM)观测。
若在体表等容易观测的部位且荧光够强,则可直接用长波长紫外灯照射(如手提式长波长紫外灯)后观察。
可观测荧光的仪器越来越多。
1.4.5无放大作用因而,很多情况下需要强启动子。
这使536在研究一些弱启动子功能时受到限制。
随着检测手段的不断改进,这一限制已被逐渐缩小。
正是由于以上这些特性,使536在问世后不久,便被广大科研工作者广泛应用。
2在转基因动物中的应用2.1 基因打靶和胚胎干细胞技术基因打靶和胚胎干细胞技术的出现为小鼠基因组操作引进了新的方法。
胚胎干细胞是来源于后胚泡期胚胎的多能干细胞系,经体外繁殖和处理后可再导入胚胎。
自人们从胚胎干细胞克隆出小鼠生殖细胞,并证明将亲本的基因组传递给后代后,胚胎干细胞就成为常用的工具。
现在,只要在体外进行基因打靶和转基因操作,就可通过生殖细胞将亲本的基因组传递给后代。
在胚胎干细胞中可进行两种基因组改造:定向改造和非定向改造。
定向改造就是预先确定并精确设计所要做的改变,基因打靶就是定向改造。
非定向改造是随机的,包括转基因和突变。
研究中,这两种方式可混合使用以达到不同的目的。
同源重组和点特异重组可对基因组进行任何想要的改变。
可通过多种方式进行随机突变,利用外源基因陷阱载体可在基因组中随机定位,从而“劫持”基因表达并破坏基因功能。
胚胎干细胞的另一种应用是转基因,它与经典的原核注射类似。
在体外,将载体自身的驱动目的基因(通常为报告基因)的启动子整合入基因组,然后导入体内。
因此,任何改造都能够导入胚胎干细胞,随后通过生殖细胞传递给后代,这样就可以建立小鼠突变品系或嵌合体,从而进行原位研究了。
由于转基因细胞系能在体外增殖,所以与DNA注射相比,用ES细胞制备转基因动物的优势在于,可在动物模型体外使用试剂进行研究。
2.2标记基因直接检测外源基因在动物体内的表达和分布常常是困难的。
因此,在制作转基因动物时,一般在目的基因旁边加上一段表达产物易于检测的基因,即标记基因。
标记基因的表达与分布提示目的基因的表达与分布。
LacZ,Iuc等都是广泛使用的标记基因,其产物能催化一定的底物生成有色产物或荧光。
由于GFP发光时不需底物,能对活体检测等优点,越来越多的人倾向于用它代替传统的分子标记物。
GFP与蛋白融合时,既可融合于蛋白的N-端,也可融合与蛋白的C-端,甚至能让蛋白插入到编码区之间而保持荧光特性不变由于GFP体积较小,只有27KD,融合后常能保持蛋白的正常分布与功能。
不同的是现在能发光了。
发光的蛋白能给研究者有关基因时间、空间表达的有用信息。
这方面成功的例子很多,被成功标记的蛋白目前已超过100种.Morilz OL将视紫红质蛋白(rhodopsin,RHO)基因与GFP的cDNA融合制作转基因爪蟾。
作者根据荧光观察了RHO表达随时间变化的规律,并将转基因动物表达的RHO-GFP与天然RHO作了对比:RHO-GFP能正确地定位,虽然被磷酸化的速度减慢,但对其行使光传导功能影响很小。
磷酸化速度减慢的原因是GFP的占位。
因为27KD虽然不大,但仍会影响某些蛋白的折叠。
寻找分子量更小的GFP突变体,是今后GFP研究的一个重要目标。
GFP也可直接接在启动子后,观测该启动子的表达、分布情况或特异性标记某种细胞、组织或器官。
这是GFP在转基因动物研究中应用最早、最多的一个方面。
Chiocchclli等在血红素结合蛋白(hcmopcxin,HPX)启动子和β1结合素启动子之后加上GFP或LacZ报告基因(共4个质粒),制作转基因小鼠。
其中,HPX启动子能指导下游基因在肝脏强表达和部分脑区弱表达,而β1结合素启动子能指导下游基因在胚胎发育过程中全身表达。
作者比较了这9个报告基因的表达情况。
实验中发现:用GFP作报告基因,不仅检测方便,而且不论是强表达还是弱表达,全身表达还是区域表达,都与启动子指导的天然蛋白的原位杂交结果更加一致。
2.3筛选阳性胚胎制作转基因动物的一个很大的问题便是阳性率低。
从大量的动物中筛选出转基因阳性动物,是一个耗时耗力的过程。
若在胚胎移入假孕母体以前就能用可靠的方法筛选出转基因阳性胚胎,弃去基因未整合或表达不好的胚胎,将大大降低出生后动物个体筛选的工作量,并降低成本。
在桑葚胚或胚泡阶段用活组织检查结合PCR或RT-PCR的方法需要复杂的显微操作技术,并可能损伤胚胎,耗时较长,并可能有假阳性或假阴性。
Thompsom等用荧光素酶作标记基因,在移植前检测。
这种方法需要在培养基中加入一些辅助因子,检测过程较复杂;Takada等将EDFP的cDNA置于EF1α启动子和CMV增强子之下,注入小鼠和牛的受精卵内,在体外培育至桑葚胚阶段或胚泡阶段。
此时,GFP表达,用CLSM可检测出荧光。
用能表达荧光的胚胎作移植,出生后的小鼠发育正常,能表达外源基因(GFP)。
牛的胚胎移植后未能成活。
这种方法对胚胎没有损伤,检测方便。
虽然此次实验只注射了标记基因,但从实验过程来看,加入外源基因是没有技术困难的。
此实验还证明了每天用,405mm的紫外照射胚胎,并不影响植入和发育。
GFP不仅可与外源基因构成双顺反子,也可单独与外源基因共注射。
以摩尔比1:1共注射后,80%的转基因鼠同时携有两种基因,20%的转基因小鼠只携有其中一种基因。
对于大动物来讲,由于受精卵来源有限,实验周期长,费用高等限制,用此法有更大的优越性。
不仅在制作转基因动物时,而且用于F0代之后扩大转基因群体。
因为对大动物来讲,传代也是一个耗时的过程。
但如果能将雄性found动物的生殖细胞取出,于雌性野生动物的卵子在体外受精并培养,并在植入前用GFP筛选,也将大大缩短扩大转基因动物群体的时间。
在某些全身透明的鱼类,用能在生殖细胞特异表达的启动子与GFP融合后,可方便地根据荧光标记筛选出有生殖系整合的转基因动物3问题和展望从1994年Chalfie等首次在果蝇、大肠杆菌等生物体内成功表达GFP基因到现在已有十几年的历史,越来越多的研究人员将其作为又一快速高效的研究工具。
虽然GFP在分子生物学的众多研究领域有着广泛的应用,GFP本身也将不断地被改进及优化,GFP的应用在技术上将出现更多的创新,其应用范围将会越来越开阔。
但在研究中仍然存在着一系列的问题,包括:①检测灵敏度还有待提高,而且其荧光信号强度方面的非线性性质使得定量非常困难也有待于进一步研究。