脂肪酸合成酶
脂肪酸合成酶系的名词解释

脂肪酸合成酶系的名词解释脂肪酸合成酶系是一组参与脂肪酸合成的酶的集合。
脂肪酸是构成人体内脂肪的重要组成部分,也是能量的主要来源之一。
脂肪酸合成酶系的功能是通过一系列化学反应将碳链延长,最终合成出长链脂肪酸。
脂肪酸合成酶系的主要成员包括乙酰辅酶A羧化酶(ACC)、甘油3-磷酸脱羧酶(G3PDH)、NADPH脱氢酶-异戊二烯酮醛还原酶(NADPH-DH-I)、正戊二烯酸羧化酶(ECA I)、磷酸乙醋酸羧化酶(ACAS I)等。
这些酶通过催化不同的化学反应,协同合作完成脂肪酸的合成过程。
脂肪酸合成酶系的关键步骤包括转运、羧化、还原、水解等。
首先,乙酰辅酶A(Acetyl-CoA)在胞浆内转运至线粒体内。
乙酰辅酶A通过乙酰辅酶A羧化酶(ACC)催化转化为丙酮辅酶A(Malonyl-CoA)。
丙酮辅酶A是脂肪酸合成的起始物质。
接下来,丙酮辅酶A与一分子的NADPH和一分子的H+在各自的酶的催化下经过一系列反应,形成长链的饱和脂肪酸。
甘油3-磷酸脱羧酶(G3PDH)催化丙酮辅酶A羧化为羟乙簇烷辅酶A(Hydroxyethyl-CoA),然后通过NADPH脱氢酶-异戊二烯酮醛还原酶(NADPH-DH-I)催化转化为β-羟基丁簇烷辅酶A(β-Hydroxybutyryl-CoA)。
接着,正戊二烯酸羧化酶(ECA I)将β-羟基丁簇烷辅酶A羧化为肌酸簇烷辅酶A(Crotonyl-CoA)。
在脂肪酸合成过程中,还需经过一系列连续的羟基化、水解、水合等步骤。
磷酸乙醋酸羧化酶(ACAS I)催化肌酸簇烷辅酶A的羟化,使其形成戊二烯酸簇烷辅酶A(trans-Enoyl-CoA)。
之后,戊二烯酸簇烷辅酶A进一步经过一系列水解反应,形成各种不同的脂肪酸。
总而言之,脂肪酸合成酶系是一组协同作用的酶,参与了脂肪酸合成的各个关键步骤。
它们通过转运、羧化、还原、水解等多种反应,细致地调控着脂肪酸合成的过程。
脂肪酸的合成不仅对人体能量供应至关重要,也在人体的各种生理功能中发挥着重要作用。
脂肪酸合成酶(FAS)的研究综述

畜牧·水产春秋在A上放牧,夏冬在B上放牧,保证草场不被破坏。
另外需要严格控制牲畜数目,在草场能承受的范围内进行合理放牧。
3 蒙古国羊绒业可持续发展的对策3.1 积极扶持技术创新和羊绒品牌建设技术创新是一个行业发展的不竭动力。
蒙古国政府应该大力加强科技创新,提高羊绒产业生产中清洗,剃毛,编织,成品的技术,并且对各大企业的产品研发和科技引进予以资金帮助,提高生产中的科技成分,提高羊绒生产效率和质量,适当降低羊绒生产成本。
此外还应提高羊绒行业品牌竞争力,建立起完善的品牌机制。
必要情况下,小众羊绒品牌应该合作共赢,形成规模,推出具有影响力的联合品牌,提升竞争力,占据一定的市场份额。
目前,蒙古国政府申请加入了国际羊毛局,并且增加了和各方的合作,这其中包括营销商标的合作。
同时,为了更进一步的提高蒙古国羊绒产品的质量,又申请了在国际羊毛局成立羊绒保证许可的专门实验室,开始实施“建立羊绒质量保证系统”项目的第二阶段工作,这预计将会给蒙古国羊绒业带来积极效应,有助于建设一个羊绒大国形象。
3.2 规范蒙古国羊绒交易市场合理完善的市场交易规则是促使羊绒交易市场健康发展的基础和关键。
所以国家应该对羊绒交易市场进行整顿,通过经济、法律和行政手段加强宏观调控,制定严格的交易秩序,颁布行业行规,明确经营者与消费者的相关权利、义务及责任。
在维护好市场秩序的同时,帮助羊绒企业实现管理规范化、产品标准化,不允许残次产品进行销售,杜绝假货,树立诚信优质的羊绒品牌形象。
4 结论目前蒙古国羊绒业的发展情况较为可观,具有一定的资源禀赋优势,但主要靠量取胜。
随着羊绒产业的不断发展和经验的积累,一批又一批的羊绒企业不断崛起,羊绒的生产工业水准不断提高。
现今工业发展过程中,技术、生产力、生产水平成为了企业能否快速成长的稳定因素。
在羊绒行业,走可持续的绿色发展道路是未来羊绒产业的发展趋势,企业要获得竞争优势,分得市场大蛋糕,就应当以生产技术和科技创新为保证。
脂肪酸合成酶系名词解释生物化学

脂肪酸合成酶系名词解释生物化学
脂肪酸合成酶系是一组参与合成脂肪酸的酶的集合。
脂肪酸是生物体内重要的能量储存形式,也是细胞膜的组成成分之一。
脂肪酸合成酶系在细胞质中催化一系列化学反应,将乙酰辅酶A (Acetyl-CoA)转化为长链脂肪酸。
脂肪酸合成酶系包括多个酶,其中关键的酶包括乙酰辅酶A羧化酶(Acetyl-CoA Carboxylase)、脂肪酰辅酶A合成酶(Fatty Acid Synthase)和脂肪酸合成酶复合物(Fatty Acid Synthase Complex)。
这些酶协同作用,通过一系列的反应步骤,将乙酰辅酶A逐渐扩大成为长链脂肪酸。
乙酰辅酶A羧化酶是脂肪酸合成的起始酶,它将乙酰辅酶A转化为马尿酸。
脂肪酰辅酶A合成酶是核心酶,它通过多个反应步骤将马尿酸逐渐延长为长链脂肪酸。
脂肪酸合成酶复合物是由多个酶组成的大分子复合物,它在细胞质中形成一个功能完整的酶体系,促进脂肪酸的合成。
脂肪酸合成酶系的活性受到多种调控机制的影响,包括激活蛋白磷酸化、抑制蛋白磷酸化、底物浓度和代谢产物的反馈调节等。
这些调控机制保证了脂肪酸的合成与细胞内能量需求和代谢状态之间的平衡。
总结而言,脂肪酸合成酶系是参与合成脂肪酸的一组酶的集合体,通过一系列的反应步骤,将乙酰辅酶A转化为长链脂肪酸。
它在细胞内发挥重要的生物化学功能,并受到多种调控机制的影响。
脂肪酸代谢-2-xin

(二)合成原料
胆固醇的合成代谢 1分子胆固醇 18乙酰CoA + 36ATP + 16(NADPH+H+)
葡萄糖有氧氧化 乙酰CoA通过柠檬酸-丙酮酸循环出线粒体
磷酸戊糖途径
(三)合成基本过程 1. 甲羟戊酸的合成
胆固醇 的合成
代谢
合成胆固醇 的限速酶
2. 鲨烯的合成
胆固醇 的合成
代谢
HMG-CoA还原酶
脂肪酸的全合成与β-氧化的比较
β-氧化 线粒体内 CoA-SH 相互独立的多酶体系 乙酰CoA NAD+,FAD β-羟酰基为L-构型 C18以下脂肪酸
脂肪酸全合成 胞液
酰基载体蛋白ACP 复合体多酶体系 丙二酸单酰CoA NADPH·H+ β-羟酰基为D-构型 C16的软脂酸
脂肪酸合成的碳源 高等动物脂肪酸合成最活跃的组织是脂肪组织、肝脏和乳腺
二软脂酰胆碱--肺泡
三、磷脂甘油的合成与降解 (二)甘油磷脂的降解
磷脂酶 (phospholipase , PLA)
磷脂酶A2(磷脂-2-酰基水解酶)
磷脂酶B:溶血 磷脂酰基水解酶
O R2-C-O-
磷脂酶A1(磷脂-1-酰基水解酶) O
CH2—O—C-R1 C-H
CH2—O—P-O-O X
磷脂酶C(磷脂酰胆碱 磷酸胆碱水解酶)
软脂酸合成的总反应:
CH3COSCoA +
7 HOOCH2COSCoA +
14NADPH+H+
脂肪酸的合成
CH3(CH2)14COOH +
7 CO2 +
6H2O +
8HSCoA +
生物化学试题脂类代谢

第六章脂类代谢.1. 脂肪酸合成酶复合体存在于细胞的(1995年生化试题)A.胞液B. 微粒体C. 粒体基质D. 线粒体内膜E. 溶酶体答案 A2。
胆固醇合成的限速酶是(1996年生化试题)A.HMG-CoA还原酶B.HMG-CoA合成酶C. 鲨烯环化酶D.p酮硫解酶E.HMG-CoA裂解酶(答案) A3.在线粒体中,脂肪酸碳链延长是以什么为原料的·A.丙二酰B.乙酰CoAC.既用乙酰OA,也用丙二酰CoAD。
一碳单位E.甘油(答案) B4.脂肪大量动员时肝内生成的乙酰CoA主要转变为(2001年A.葡萄糖B.胆固醇C.脂肪酸·D.酮体 E.丙二酰CoA(答案) D5.脂肪酸氧化过程中不需要下列哪种化合物参与A.肉碱B。
NAD+ C.NADP+ D.FAD E. CoASH(答案) C6. 脂肪酸在肝脏进行p氧化不生成下列哪一种化合物(1997年研究生考题)A. H2O B.乙酰CoA C. 脂酰CoA D. NADH+H+ E.FADH2答案 A7. 下列关于原核生物脂肪酸合成酶复合体的说法哪种是正确的(1998年研究生考题)A. 催化不饱和脂肪酸合成B. 催化脂酰CoA延长两个碳原子;C. 含一个酰基载体蛋白和七种酶活性D. 催化乙酰CoA生成丙二酰CoA的反应E. 催化脂肪酸活化成脂酰CoA的反应答案C8. 脂肪酸生物合成(2001年研究生考题)A. 不需要乙酰CoA B.中间产物为丙二酰CoAC. 在线粒体内进行D.以NADH为还原剂*E. 最终产物为10碳以下脂肪酸答案 B9. 下列关于酮体的叙述错误的是A. 肝脏可以生成酮体,但不能氧化酮体B.酮体是脂肪酸部分氧化分解的中间产物C. 合成酮体的起始物质是乙酰CoAD.酮体不包括p-羟丁酸E,机体仅在病理情况下才产生酮体(答案) E10.内源性甘油三酯主要由下列哪一种血浆脂蛋白运输A.CM B.LDL C.VLDL D. HDL E.IDL(答案) C(二)X型题1.人体必需脂肪酸包括(1995年生化试题)A.软脂酸B.花生四烯酸 C. 油酸 D. 亚麻酸(答案) B D2.关于酮体的正确说法是(1996年生化试题)A.酮体包括乙酰乙酸、p-羟丁酸和丙酮B.酮体过多可从尿中排出,称酮尿C.饥饿时体内酮体可以增高O.与糖尿病患者的多尿有密切关系(答案) A\B\C3. 胞浆脂肪酸生成具有以下特点A. 需酰基载体蛋白(ACP)运载脂酰链B. 利用NAD+—NADH+H+C. 利用NADPH+H+—NADP+D. 能由柠檬酸促进答案 A C D4. 由乙酰CoA可合成(1997年生化试题)A. 胆固醇B.酮体 C. 脂肪酸D.甘油答案 A B C5. 通过高脂蛋白血症中,下列哪种脂蛋白可能增高(2000年研究生考题)A. 乳糜微粒B.极低密度脂蛋白C. 高密度脂蛋白D.低密度脂蛋白答案 A B D四测试题(一)A型题1.催化体内储存的甘油三酯水解的脂肪酶是A. 激素敏感性脂肪酶B. 脂蛋白脂肪酶C. 肝脂酶D.胰脂酶; E. 组织脂肪酶2.下列关于脂蛋白脂肪酶(LPL)的叙述错误的是:A.LPL是一种细胞外酶,主要存在毛细血管内皮细胞表面,B,它催化脂蛋白中的甘油三酯水解C. 脂肪组织、心肌,脾及乳腺等组织中该酶活性较高-D.apoCⅢ可抑制LPL E.aPOAI能激活LPL3.能促进脂肪动员的激素有A.肾上腺素B.胰高直糖素 C. 促甲状腺素D.ACTH E.以上都是4.下列激素具有抗脂解作用的是A.肾上腺素B.胰高血糖素 C. ACTHD.前列腺素E2 E.促甲状腺素5.下列关于激素敏感脂肪酶的叙述错误的是A.催化贮存的甘油三酯水解的脂肪酶B.胰高血糖素可促使其磷酸化而激活C. 胰岛素则使其去磷酸化而失活D.其所催化的反应是甘油三酯水解的限速步骤E.此酶属于脂蛋白脂肪酶类6.蛋白脂肪酶(LPL)催化A.脂肪细胞中甘油三酯水解 B. 肝细胞中甘油三酯水解C. CM和VLDL中甘油三酯水解D。
脂肪酸合成酶(FAS)的研究综述

脂肪酸合成酶(FAS)的研究综述作者:张芮陈斯钰来源:《农村经济与科技》2018年第13期[摘要]脂肪酸合成酶(FAS)作为一种合成脂肪酸的关键酶,具有丰富的酶系统功能,在高低等动物身上它的存在形式不同,并且它在影响生物的能量代谢中发挥着极大作用。
近年来,有关于脂肪酸合成酶的研究成果越来越多。
从FAS 的生理功能、结构特性和未来应用等方面,梳理了近几年关于FAS的研究成果。
[关键词]脂肪酸合成酶;基因结构;生理功能;应用[中图分类号]F301.24 [文献标识码]A1 脂肪酸合成酶的结构特性脂肪酸合成酶(fatty acid synthase, FAS)的概念是在1957年由Wakil等初次提出并命名。
研究发现FAS基因有Ⅰ型和Ⅱ型2种类型。
植物中的存在的FAS属于Ⅱ型。
而哺乳类动物的FAS是Ⅰ型,它是由一条相对分子质量为250kDa的多肽链首尾相连,形成的以二聚体为主要存在形式的多功能酶,存在于细胞浆中。
脂肪酸合成酶的结构因物种不同而存在差异。
Clemente等通过研究得出人、鹅、鼠的FAS基因中都存在不翻译的外显子I。
人与鹅的相同序列为41%,人和鼠相同序列为65%,得出其外显子有较小的保守性。
Roy等发现鸡的FAS基因在18号染色体上。
后来Joshi等学者发现了由7512个核苷酸编码成2504个氨基酸组成的位于17q25上的人的FAS基因。
同年研究学者发现猪的FAS基因存在于12p1 .5。
Kameda等通过实验也测出鹅中的FAS基因全序列大约长度有50kb。
据东方网2008年9月9日《科技日报》报道,瑞士苏黎世工学院的科学家们通过利用保罗谢勒研究所所采集的数据,了解了哺乳动物中脂肪酸合成酶的原子构成,并在《科学》杂志上发表相关的研究成果。
科学家已经研究了很久,关于哺乳动物中FAS难于令分子合成的机理。
然而,截至目前,科学家们一直在努力通过使用孤立的细菌酶来探究脂肪酸的合成过程中的每个步骤。
脂肪酸合成酶

脂肪酸合成酶(FAS)基因的研究进展以及日粮成分对其表达的调控随着人们生活水平的提高, 对动物性产品的品质提出了“低脂高蛋白且无各种有毒有害物质残留”的新要求。
为此,营养学家们开始把研究重点转移到体脂沉积的营养调控上。
自从1957年,脂肪酸合成酶由S.J.Wakil等人在鹅肝匀浆中首次发现以来,人们对脂肪酸生物合成途径进行了大量研究,初步阐明了脂肪酸合成规律,并在此基础上从分子生物学水平对脂肪酸合成酶,尤其是脂肪酸合成酶的基因表达进行了探索性的研究。
脂肪酸合成酶在动物体脂生成、沉积中发挥重要作用。
熊文中等发现,猪脂肪组织中脂肪酸合成酶活性与胴体脂肪量、胴体的脂肪率呈极显著正相关。
还发现脂肪酸合成酶是一种新的肿瘤标志物,癌细胞的生长依赖于脂肪酸合成酶的活性,许多肿瘤如乳腺癌、前列腺癌等都有脂肪酸合成酶的高度表达。
人的脂肪酸合成酶基因位于17q25、牛的位于19q22、鸡的位于18号染色体上,正常情况下,在肝和脂肪组织中表达。
随着分子生物学的迅速发展,人们对脂肪酸合成酶进行了大量深入的研究。
1 动物体脂合成的调控动物体脂一方面靠直接摄入,更多的则是在体内自身合成。
脂肪酸合成酶是体内合成脂肪途径中一个关键酶,它通过催化乙酰CoA和丙二酰CoA而生成长链脂肪酸。
通过对能量物质体内生化代谢途径进行分析可以发现,在能量代谢中,乙酰CoA是一个重要而又特殊的物质,主要能量物质碳水化合物、脂肪以及部分氨基酸的初级代谢终产物都是乙酰CoA,其可进入三羧酸循环进行完全氧化,为生物细胞提供能量来源;同时,乙酰CoA也是合成脂肪的起点,先羧化为丙二酰CoA,再由脂肪酸合成酶进行一系列的反应合成为脂肪酸,完成合成脂肪的主要工作。
实际上乙酰CoA是个能量汇集点,能量物质摄入体内后被氧化产生能量还是合成脂肪,均在乙酰CoA这个“关节点”进行分配。
该分配的比率取决于这两大通路的“通畅”程度。
任一通路被阻滞或去阻滞都会影响生能和生脂的分配,造成体脂水平的变化。
脂肪代谢合成

花生四烯酸(20C:4 Δ5,8,11,14 )
四、脂肪酸合成的调节
1. 代谢物的影响(1)
进食糖 糖代谢 NADPH+乙酰CoA+ATP
(—)
柠檬酸、异柠檬酸
透出线粒体
(+) 柠檬酸裂解 乙酰CoA
异柠檬酸 脱氢酶
乙酰CoA 羧化酶 合成脂肪有关 的酶的活性
脂肪酸合成
1. 代谢物的影响(2)
饥饿、糖供应不足 透出线粒体 的柠檬酸 脂肪动员
磷脂酶 B1 : 水解溶血磷脂1
磷脂酶 B2 : 水解溶血磷脂2
磷脂酶 C : 存在于细胞膜、蛇毒及某些细菌
磷脂酶 D : 主要存在于高等植物,动物脑组织亦有。
三、甘油磷脂的合成代谢
1. 合成部位:全身各组织内质网,肝、肾、肠最 活跃。
2. 合成原料及辅因子:
脂酸、甘油:由糖代谢提供 多不饱和脂酸:从植物油摄取 磷酸盐:由ATP提供 含氮化合物:从食物摄取或体内合成 CTP:构成活化的中间物
(KS)的催 化下,完成乙酰基 与丙二酸单酰基缩 合并脱羧形成β-酮 丁酰-ACP。
④ 还原
结果:在β-酮 酰-ACP还原酶 (KR)的催化下, β-酮丁酰加氢 还原为D-β-羟 丁酰-ACP。
⑤ 脱水
结果:在β-羟 酰-ACP脱水酶 (HD)的催化下, β-羟丁酰基脱 水形成α,β – 烯丁酰ACP。
• 线粒体中的乙酰 CoA,需通过三羧酸
转运体系(也称拧檬酸穿梭系统)运
到胞浆中,才能供脂肪酸合成所需,
此过程是耗能的。
乙酰CoA转运出线粒体过程图解
线粒体基质
HSCoA 柠檬酸合酶 H2O + 乙酰CoA NADH + H+ 草酰乙酸 柠檬酸
脂肪酸合成酶的相关研究

多种中草药及其中的黄酮化合物
有文献陆续报道了多种对FAS有强抑制作用的中草 药, 在动物实验中表现了良好减重抑食功能的中草 药.其中比较典型的有桑寄生[13]、何首乌[14]、高良 姜[15]、银杏叶[16]、夜交藤[17]、枫叶[18]等.其共同特 点是抑制能力很强,最佳溶剂提取物的IC50值在 0.4—5ug/ml,抑制能力明显高于茶和其它已知FAS 抑制剂 经测定表明, 嶰皮素、山奈酚、桑色素、异鼠李素 以及芸香试等对FAS有强弱不等的抑制, 其中最强 的IC50值可达接近1um/ml,(约2umol/l),抑制活性强, 但基本没有不可逆抑制
FAS抑制剂的创新研究
与单一靶点的抑制剂相似,传统的FAS 抑制剂也 常常因不稳定、用量大等因素而限制其在临床上 的广泛使用,因此开发设计多靶点结合的新型抑 制剂将有很大潜力。此外,FAS 活性位点的多样 性以及功能区域的广泛性,使得开发多靶点相结 合的新型抑制剂成为可能。
总结和展望
FAS 抑制剂在减肥、抗癌方面的神奇功效引起了 人们的广泛关注,特别是开发多靶点相结合的新 型FAS抑制剂已成为研究热点。随着FAS 晶体结构 的不断揭示以及计算机模拟结构设计药物的出现 ,使得在FAS活化机制和新型FAS 抑制剂开发等领 域取得了部分突破性的进展。 尽管人们在FAS 抑制剂研究取得长足的进展,但 作为一种新型治疗方法,FAS抑制剂在临床上的成 功应用还有一段相当漫长的道路。
葱属植物中的多硫醚
百合科葱属植物大多数可供食用, 因其特殊的香味 成为日常生活中的调味蔬菜,如葱、蒜、韭菜、洋 葱、蒜苗和韭黄等。近年来葱属植物在防治心脑 血管疾病, 降压降脂及预防和治疗癌症等方面的功 能受到广泛关注。如有报道食用大蒜可以显著降 低高脂病人与动物的胆固醇及甘油三酯水平[19], 而洋葱除具有降低甘油三酯的作用之外[20],在防治 癌症上也有着一定的作用
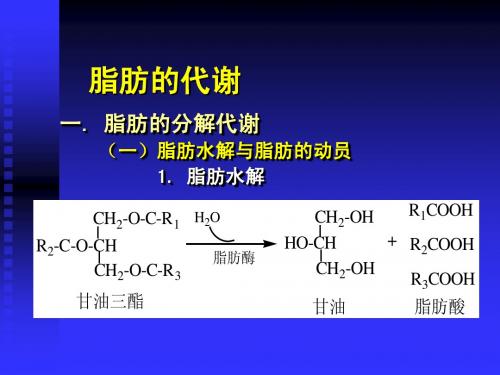
脂肪的代谢
O 消旋酶
L-甲基丙二酰CoA
异构酶
TCA
CH 2-COOH CH 2-COOH 琥珀酸
(五)不饱和脂肪酸的氧化
机体中脂肪酸约一半以上为不饱和脂肪酸, 它也在线粒体中进行β-氧化。不同的是: 天然不饱和脂肪酸中的双键均为顺式结构, 而生物体内催化的反应是反式结构。因此 β-氧化进行到顺式时受阻。必须将顺式转 变为反式结构才能进行。它需2种酶参与, 一种是△3顺→ △2反烯脂酰CoA异构酶或 D(-) - β-羟脂酰表构酶。
二、三脂酰甘油的合成代谢
(一) 脂肪酸的合成——饱和脂肪酸 1. 脂肪酸的合成部位 主要是在肝脏和肌肉组织中, 在胞液中进行。
2. 合成原料 : 乙酰CoA、ATP、NADPH、HCO3- 。 乙酰CoA是合成脂肪酸的主要原料,它主 要来自葡萄糖。细胞内的乙酰CoA全部在 线粒体内产生,而合成脂肪酸的酶体系存 在于细胞液。因此,线粒体内的乙酰CoA 须进入胞液才能作为合成脂肪酸的原料。 实验证明,乙酰COA不能自由透过线粒体 内膜,需借用柠檬酸一丙酮酸循环 (citrate pyruvate cycle)来完成。
+
硬脂酰CoA + HSCoA +NADP
4.酮体生成的生理意义 ⑴酮体是肝输出能源的一种形式。 它是易溶于水的小分子物质,能 通过血脑屏障及肌肉毛细血管壁。 脑组织 不能氧化脂肪酸,却能利 用酮体。
⑵ 正常情况下,血中含有少量酮体,为 0.03— 0.5mmol/L(0.3—5mg/dl)。也就是 肝脏生成的酮体被及时利用。在饥饿、高 脂低糖膳食及糖尿病时,脂肪酸动员加强, 酮体生成增加。如果酮体生成超过肝外组 织利用的能力时,方可引起血中酮体升高, 导致酮症酸中毒,并随尿排出,引起酮尿。
生物化学第九章脂代谢

SH
H2O
HOOCCH2CO-S CH3CO-S CH3COCH2CO-S
SH
③
CO2
④
NADP+ NADPH
2.线粒体中的合成
碳链的延长发生在线粒体和内质网中。与脂肪酸β-氧化的逆 向过程相似,使得一些脂肪酸碳链(C16)加长。 延长是独立于脂肪酸合成之外的过程,是乙酰单元的加长和 还原,恰恰是脂肪酸降解过程的逆反应。光面内质网中的延 长更为活跃。
酮体的生成
HMGCoA裂 解酶 CH3COCH2COOH
乙酰乙酸 脱氢酶
HMGCoA 合成酶
NADH+H+ NAD+
脱羧酶 CO2
OH | HOOCCH2-C-CH2COSCoA | CH3 羟甲基戊二酸单酰CoA (HMGCoA)
CH3CHOHCH2COOH
--羟丁酸
CH3COCOOH
丙酮
酮血症?
5.不饱和脂肪酸的氧化
与脂肪酸的β-氧化相同,但需增加异构酶 和 还原酶:
(三)脂肪酸氧化的其它途径
1.奇数碳原子脂肪酸的氧化 如17个碳直链脂肪酸: 先经β-氧化至3碳的丙酰-CoA ,产生7个乙酰CoA和一个丙酰-CoA 。 丙酰-CoA经3步反应转化为琥珀酰-CoA然后进入 三羧酸循环进一步进行代谢。
acc合酶名词解释

acc合酶名词解释1.引言1.1 概述在生物化学领域中,ACC合酶是一个关键的酶类分子,它扮演了脂肪酸合成途径中的重要角色。
ACC合酶全名为乙酰辅酶A羧化酶(Acetyl-CoA Carboxylase),是一种催化乙酰辅酶A羧化的酶。
乙酰辅酶A是一种在生物体中广泛存在的代谢物,在脂肪酸合成途径中起着重要作用。
脂肪酸是生物体中最重要的能量来源之一,并在细胞膜的构建等方面发挥重要作用。
而乙酰辅酶A则是脂肪酸的前体分子,它通过一系列的反应催化合成脂肪酸。
ACC合酶通过将乙酰辅酶A转化为丙酰辅酶A,对脂肪酸合成途径进行调控。
这个过程涉及到乙酰辅酶A羧化的反应,即将乙酰辅酶A的一个羧基转化为碳酰基。
ACC合酶在生物体中广泛存在,并在不同类型的生物中具有多种不同的同功型。
它在植物中特别重要,因为植物需要合成大量的脂肪酸来提供能量和构建细胞膜。
此外,对于一些细菌和其他微生物而言,ACC合酶也是重要的代谢途径。
ACC合酶的结构和功能一直受到广泛的研究和关注。
研究人员发现,在不同类型的生物中,ACC合酶的结构和催化机制存在一定的差异。
这种差异可能与生物体的适应环境和功能有关。
因此,深入了解ACC合酶的结构和功能对于揭示其作用机制以及在其他领域的应用具有重要意义。
本文将对ACC合酶的定义、作用、结构和功能进行探讨,并总结ACC 合酶在生物体代谢中的重要性。
同时,还将展望ACC合酶在未来的研究方向,期望促进对ACC合酶的深入理解和应用。
文章结构部分的内容可以如下编写:1.2 文章结构本文分为引言、正文和结论三个部分。
引言部分概述了acc合酶的重要性和研究的目的,介绍了本文的研究背景和动机。
通过引人入胜的方式引导读者对acc合酶的认识和关注。
正文部分主要分为两个小节,分别是acc合酶的定义和作用以及acc 合酶的结构和功能。
在第二节中,将详细介绍acc合酶的分子结构、组成以及其在细胞内的功能机制等方面的内容。
这部分将深入探讨acc合酶在生物体中的重要作用,并解释其在脂肪酸合成等代谢过程中的具体功能。
脂肪酸氧化的过程

脂肪酸氧化的过程如下:
1.活化:脂肪酸在脂酰CoA合成酶的作用下,与CoA结合生成
脂酰CoA,并释放出自由能。
2.转移:脂酰CoA进入线粒体基质,在线粒体内膜外侧的肉碱脂
酰转移酶I催化下,脂酰CoA与肉碱(即L-3-羟基-4-三甲基铵丁酸)结合,生成脂酰肉碱。
3.β氧化:脂酰肉碱在线粒体内膜内侧的肉碱脂酰转移酶Ⅱ催化
下,将脂酰肉碱重新转移回线粒体基质内,并释放出肉碱。
4.氧化:在脂酰CoA脱氢酶催化下,脂酰CoA的α、β碳原子各
加一个氧原子生成乙酰CoA。
5.彻底氧化:乙酰CoA进入三羧酸循环彻底氧化分解为CO2和
H2O,并释放大量能量。
乙酰辅酶 a 羧化酶基因

乙酰辅酶 a 羧化酶基因乙酰辅酶A羧化酶(acetyl-CoA carboxylase)是一种参与脂肪酸生物合成的关键酶。
在脂肪酸生物合成过程中,乙酰辅酶A羧化酶将乙酰辅酶A转变为驱动酰辅酶A,进而驱动合成长链脂肪酸。
乙酰辅酶A羧化酶的基因编码乙酰辅酶A羧化酶蛋白,这个基因在人类基因组中被称为ACACA。
乙酰辅酶A羧化酶是一种双亚基酶,由两个亚单位组成:一个大小为280 kDa的亚单位(β-亚单位),以及一个大小为12.5 kDa的亚单位(α-亚单位)。
这两个亚单位是由乙酰-CoA羧化酶的基因编码的。
乙酰辅酶A羧化酶在脂肪酸生物合成过程中起着重要的作用。
它转换乙酰辅酶A为驱动酰辅酶A,并将二氧化碳加到乙酰辅酶A的羧基上,形成β-羧基乙酰辅酶A。
驱动酰辅酶A是合成长链脂肪酸的前体,它用作脂肪酸的合成起始物。
乙酰辅酶A羧化酶通过催化乙酰辅酶A的羧化反应,调控脂肪酸的合成速度。
乙酰辅酶A羧化酶在脂肪酸合成途径中有多个调控点。
首先,乙酰辅酶A羧化酶的催化活性受到乙酰辅酶A浓度的调控。
当细胞中乙酰辅酶A浓度增加时,乙酰辅酶A羧化酶的活性也会增加。
此外,乙酰辅酶A羧化酶的活性还受到营养素和代谢物的调节。
例如,胰岛素可以通过激活乙酰辅酶A羧化酶来促进脂肪酸的合成,而AMP激活的蛋白激酶则可以通过抑制乙酰辅酶A羧化酶来减少脂肪酸的合成。
乙酰辅酶A羧化酶在人类体内的功能异常可能导致一系列疾病。
例如,乙酰辅酶A羧化酶活性的下调可能导致脂肪酸合成不足,使人体无法合成足够的脂肪酸去满足各种生理需要。
这可能导致脂肪酸缺乏性疾病,如低血脂症。
另外,乙酰辅酶A羧化酶的过度活化也可能导致异常的脂肪酸合成,进而引起高脂血症和其他与脂肪代谢相关的疾病。
了解乙酰辅酶A羧化酶的基因结构和功能有助于我们更好地理解脂肪酸生物合成的调控机制,并为相关疾病的诊断和治疗提供线索。
此外,乙酰辅酶A羧化酶也是一个潜在的药物靶点,研究其调控机制和开发抑制剂有望为治疗相关疾病提供新的策略。
胰岛素对脂肪酸合成酶的作用机制

胰岛素对脂肪酸合成酶的作用机制胰岛素对脂肪酸合成酶的作用机制一、胰岛素的作用机制1. 胰岛素的分泌与作用胰岛素是由胰岛中的β细胞分泌的一种蛋白质激素,其主要作用是降低血糖浓度,促进葡萄糖的利用和储存。
胰岛素通过结合胰岛素受体,调控多种代谢途径,具有广泛的生理作用。
2. 胰岛素与脂肪酸合成酶脂肪酸合成酶是一个重要的酶,它参与了脂肪酸的合成过程。
胰岛素通过调控脂肪酸合成酶的活性和表达,影响脂肪酸的合成和代谢过程。
下面我们就来具体探讨胰岛素对脂肪酸合成酶的作用机制。
二、胰岛素对脂肪酸合成酶的作用机制1. 胰岛素调控脂肪酸合成酶的基本机制在细胞内,胰岛素通过激活胰岛素受体,并介导一系列信号传导途径,最终调控脂肪酸合成酶的活性和表达。
这些信号传导通路涉及多种激酶、磷酸化蛋白和转录因子的活化,最终影响脂肪酸合成酶的功能状态。
2. 胰岛素与脂肪酸合成途径的关系脂肪酸的合成是一个复杂的生物化学过程,涉及多种酶和代谢途径。
胰岛素作为主要的代谢调控激素,参与调节葡萄糖和脂肪的代谢平衡。
胰岛素通过抑制脂肪酸合成酶的活性,减少脂肪酸的合成和储存,从而影响了脂肪代谢通路的平衡。
三、个人观点和理解从分子生物学角度来看,胰岛素对脂肪酸合成酶的作用机制是一个复杂而精密的调控过程。
了解这一调控机制,有助于更好地理解脂肪代谢的调控原理,为疾病的防治提供理论依据。
我认为胰岛素在脂肪代谢中的作用不仅仅限于调节葡萄糖代谢,还涉及了更广泛的代谢调控。
这一理解为我们深入研究肥胖、糖尿病等代谢性疾病提供了新的思路和途径。
四、总结和回顾胰岛素作为重要的代谢调控激素,在脂肪代谢中发挥了重要作用。
通过调控脂肪酸合成酶的活性和表达,胰岛素影响了脂肪酸的合成和代谢过程。
深入了解胰岛素对脂肪酸合成酶的作用机制,有助于我们更好地理解脂肪代谢的调控原理,为相关疾病的防治提供理论依据。
在未来的研究中,我们需要进一步探讨这一调控机制,探索相关疾病的治疗策略,并为代谢性疾病的防治提供新的思路和途径。
脂肪酸合成酶系

脂肪酸合成酶系脂肪酸合成酶系是机体内一个重要的代谢途径,其功能是合成脂肪酸。
脂肪酸是一类生物体中广泛存在的重要能量储备物质,也是细胞膜的主要组成成分。
脂肪酸合成酶系通过一系列的反应步骤,将二氢甘油三磷酸(NADPH)和丙酮酸转化为长链脂肪酸。
在哺乳动物细胞中,脂肪酸合成酶系主要存在于细胞质内。
该酶系由多个酶组成,包括乙酰辅酶A羧化酶(ACC)、乙酰脱羧酶(ACLY)、乙酰辅酶A羧基转移酶(ACACA)、脂肪酸合酶(FASN)等。
这些酶在脂肪酸合成途径中起着关键作用。
脂肪酸合成酶系的反应步骤相对复杂,但可以简化为以下几个主要阶段:1. 乙酰辅酶A的合成:乙酰辅酶A是脂肪酸合成的起始物质,它由乙酰辅酶A合成酶(ACC)催化乙酰辅酶A的羧化反应得到。
2. 乙酰辅酶A的转化:乙酰辅酶A经乙酰辅酶A羧基转移酶(ACACA)催化乙酰辅酶A的转化反应,生成丙酮辅酶A。
3. 丙酮辅酶A的合成:丙酮辅酶A通过乙酰脱羧酶(ACLY)催化丙酮酸的脱羧反应得到。
4. 长链脂肪酸的合成:丙酮辅酶A通过脂肪酸合酶(FASN)的催化,经过一系列的加成、脱水和还原反应,最终合成长链脂肪酸。
脂肪酸合成酶系的活性受到多种因素的调控。
例如,内源性信号分子如胰岛素、糖皮质激素等可以促进脂肪酸合成酶系的活化,而其他一些因子如AMP激活蛋白激酶(AMPK)等则具有抑制作用。
这些调控机制保证了脂肪酸的合成与细胞内能量状态的匹配。
值得注意的是,脂肪酸合成酶系的异常活化与多种疾病的发生密切相关。
例如,脂肪酸合成酶过度活化会导致脂肪积聚、肝脏脂肪肝的发生。
因此,对脂肪酸合成酶系的研究不仅有助于深入了解细胞代谢的调控机制,还为相关疾病的治疗提供了新的思路。
总结起来,脂肪酸合成酶系是机体内合成脂肪酸的重要途径,其活性受到多种调控机制的影响。
对于脂肪酸合成酶系的研究有助于我们更好地理解生物体的能量代谢和与相关疾病的关系。
未来的研究将继续揭示脂肪酸合成酶系的详细调控机制,并探索其在疾病治疗中的应用前景。
胰岛素对脂肪酸合成酶的作用机制

# 胰岛素对脂肪酸合成酶的作用机制## 1. 胰岛素和脂肪酸合成酶的基本概念在人体的新陈代谢过程中,胰岛素和脂肪酸合成酶是两个十分重要的元素。
胰岛素是由胰腺分泌的激素,它在调节人体血糖水平方面发挥着至关重要的作用。
而脂肪酸合成酶则是参与脂肪酸合成的酶类,在维持脂质代谢平衡方面起着不可或缺的作用。
本文将从以下几个方面来探讨胰岛素对脂肪酸合成酶的作用机制。
## 2. 胰岛素如何调节脂肪酸合成酶的活性胰岛素对脂肪酸合成酶的作用机制主要是通过调节酶的活性来实现的。
当胰岛素水平升高时,它会促进脂肪酸合成酶的活性,进而加速脂肪酸的合成过程。
具体来说,胰岛素通过促进脂肪酸合成酶的磷酸化来提高其活性,从而增加脂肪酸的合成速度。
胰岛素还可以通过调节脂肪酸合成酶基因的表达来影响其活性,进一步加强对脂肪酸合成的调控作用。
## 3. 胰岛素和脂肪酸合成酶的相互作用除了上述的直接调节作用外,胰岛素还可以通过与其他信号通路相互作用来影响脂肪酸合成酶的活性。
胰岛素与糖原合成酶的相互作用会影响糖原的合成和分解,从而进一步影响脂肪酸的合成过程。
胰岛素还可以通过与其他代谢调节相关的激素和信号通路相互作用,来调控脂肪酸合成酶的活性,从而在脂质代谢过程中发挥着更为复杂和精细的调节作用。
## 4. 个人观点和总结从以上的探讨可知,胰岛素对脂肪酸合成酶的作用机制是多方面的、复杂的,并且与其他代谢调节因素有着密切的相互作用。
理解胰岛素对脂肪酸合成酶的作用机制,有助于我们更加全面地认识胰岛素在脂质代谢调节中的重要作用,对于预防和治疗代谢性疾病具有重要意义。
总结起来,胰岛素对脂肪酸合成酶的作用机制是一个复杂而精妙的过程,需要结合多方面的因素来进行全面的认识和分析。
希望通过本文的探讨,能够对读者对此有所启发和帮助。
也希望我们能够进一步深入探讨和研究,以便更好地认识和利用胰岛素在脂质代谢调节中的重要作用。
胰岛素是一种由胰腺分泌的激素,在调节血糖水平方面发挥着重要作用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
④第一次还原反应(加氢): β-羟脂酰-泛-E2的生成
⑤脱水反应:α, β-烯脂酰-泛-E2的生成
⑥第二次还原反应(加氢):脂酰-泛-E2的生成
每循环一次,增加两个碳原子,经7 次循环,生成16C的软脂酰-泛-E2,经硫酯酶的水解作用,生成软脂酸。(图6-7)
软脂酸总反应:
哺乳动物中的脂肪酸合成酶含有两个等同的多功能单链(形成同源二聚体),每一条氨基酸链的N端区域含有三个催化结构域(酮脂酰合成酶、脱水酶和单酰/乙酰转移酶]]),而C端区域则含有四个结构域(醇还原酶、酮脂酰还原酶、酰基载体蛋白和硫酯酶),这两个区域被中间600个氨基酸残基组成的核心区域所分隔。
提取方法
⑥第二次还原反应(加氢): 丁酰-泛-E2的生成
丁酰-泛-E2是第一轮产物,经酰基转移、缩合、还原、脱水、再还原,碳原子由2增加至4个。
(2)第二轮
①丁酰基转移:由丁酰-泛-E2转移生成丁酰-半胱-E1
②丙二酰基转移:生成丙二酰-泛-E2
③缩合反应:β-酮己酰-泛-E2的生成
结构
脂肪酸合成酶组构的传统模型(“头对尾”模型)大部分是基于双功能试剂1,3-dibromopropanone(DBP)能够将一个脂肪酸合成酶单体上的酮脂酰合成酶结构域活性位点上的半胱氨酸(Cys161)的巯基和另一个单体上的载体蛋白结构域中的磷酸泛酰巯基乙胺辅基联接在一起的现象。
但对脂肪酸合成酶二聚体所进行的突变研究发现酮脂酰合成酶和单酰/乙酰转移酶结构域可以与二聚体中任何一个单体上的载体蛋白共同作用;而对于DBP联接实验结果的再分析显示酮脂酰合成酶的活性位点Cys161的巯基可以被联接到任一单体中载体蛋白4'-磷酸泛酰巯基乙胺的巯基上。而且,近来发现只含有一个完整单体的异源二聚化的脂肪酸合成酶能够进行棕榈酸酯的合成。以上的这些实验结果与之前的“头对尾”模型并不相符,于是另一个模型被提出:两个单体上的酮脂酰合成酶和单酰/乙酰转移酶结构域位于接近脂肪酸合成酶二聚体中心的位置,在这一位置上,它们能够与任一单体中的载体蛋白接触。
乙酰CoA+ 7丙二酰CoA + 14 NADPH+14H+→软脂酸+7 CO2+6H2O+8CoASH+14NADP+
在低等生物中,脂肪酸合成酶系是一种由1分子脂酰基载体蛋白(ACP)和7种酶单体所构成的多酶复合体;但在高等动物中,则是由一条多肽链构成的多功能酶,通常以二聚体形式存在,每个亚基都含有一ACP结构域。
在脂酸合成酶系内各种酶的催化下,依次进行酰基转移、缩合、还原、脱水、再还原等连续反应,每次循环脂酸骨架增加2个碳原子,7次循环后即可生成16碳的软脂酸,经硫酯酶水解释出。
(1)第一轮:
①乙酰基转移:由乙酰转移酶催化生成乙酰-半胱-E1
②丙二酰基转移:生成丙二酰-泛-E2
③缩合反应:β-酮丁酰-泛-E2的生成,同时有CO2脱落
④第一次还原反应(加氢): β-羟丁酰-泛-E2的生成
⑤脱水反应: α ,β-烯丁酰-泛-E2的生成
脂肪酸合成酶
脂肪酸合成酶 fatty acid synthetase 乙酰CoA+7丙=酸CoA+14NADPH+14H+→棕榈酸+7CO2+
8CoA+14NADP++6H2O。催化上述反应的酶称为脂肪酸合成酶,不过酵母酶的最终产物是棕榈酸CoA。如图所示,在丙二酸基和乙酰基缩合时,在每次延长C2单位的同时发生还原反应,在这个复杂的反应中,各有相应的酶参与作用。而酰基以CoA转移到酰基载体蛋白(ACP)上,以与此蛋白质结合的形态进行反应。通过如图所示的反应反复进行可生成棕榈酸。在大肠杆菌中,各部分反应的酶以及ACP是不结合在一起的,但在动物和酵母中,各种酶是结合型,形成所谓多酶复合体。
④第一次还原反应(加氢): β-羟己酰-泛-E2的生成
⑤脱水反应:α, β-烯己酰-泛-E2的生成
⑥第二次还原反应(加氢):己酰-泛-E2的生成
(3)第n轮
①丁酰基转移:由丁酰-泛-E2转移生成脂酰-半胱-E1
②丙二酰基转移:生成丙二酰-泛-E2
③缩合反应:β-酮己脂酰-泛-E2的生成
概述 脂肪酸合成酶是一个具有多种功能的酶系统,在哺乳动物中,其分子量高达272kDa。在脂肪酸合成酶中,底物和中间产物分子在各个功能结构域(可以位于同一酶分子,也可以位于不同酶分子)中传递直到完成脂肪酸的整个合成过程。
在低等生物中,脂肪酸合成酶系是一种由1分子脂酰基载体蛋白(ACP)和7种酶单体所构成的多酶复合体;但在高等动物中,则是由一条多肽链构成的多功能酶,通常以二聚体形式存在,每个亚基都含有一ACP结构域。
在脂酸合成酶系内各种酶的催化下,依次进行酰基转移、缩合、还原、脱水、再还原等连续反应,每次循环脂酸骨架增加2个碳原子,7次循环后即可生成16碳的软脂酸,经硫酯酶水解释出。
功能
脂肪酸是脂肪族类酸,在能量运输和储存、细胞结构、提供激素合成的中间物等多个方面发挥着关键作用。脂肪酸的合成需要将乙酰辅酶A和丙二酸单酰辅酶A通过一系列的克莱森缩合反应然后脱羧(生物素作辅酶)来完成。在脂肪链的延伸过程中,通过连续的酮还原酶、脱水酶以及烯脂酰ACP还原酶的作用,加入的酮基(酰基)被还原为完全饱和的脂肪链。延伸中的脂肪链在这些酶活性位点之间循环传递时,共价连接在酰基载体蛋白的磷酸泛酰巯基乙胺(phophopantetheine)辅基上,并通过硫酯酶的作用而被释放
脂肪酸合成酶是一个具有多种功能的酶系统,在低等生物中,脂肪酸合成酶系是一种由1分子脂酰基载体蛋白(ACP)和7种酶单体所构成的多酶复合体;但在高等动物中,则是由一条多肽链构成的多功能酶,通常以二聚体形式存在,每个亚基都含有一ACP结构域。在脂肪酸合成酶中,底物和中间产物分子在各个功能结构域(可以位于同一酶分子,也可以位于不同酶分子)中传递直到完成脂肪酸的整个合成过程。