项目名称-糖脂代谢稳态调控的分子机制-首席科学家-林圣彩厦门大学-
不同品种毛茶水浸出物成分及抗氧化、降血糖活性研究

㊀Guihaia㊀Jun.2023ꎬ43(6):1145-1154http://www.guihaia-journal.comDOI:10.11931/guihaia.gxzw202206062霍华珍ꎬ蔡爱华ꎬ郭春雨ꎬ等ꎬ2023.不同品种毛茶水浸出物成分及抗氧化㊁降血糖活性研究[J].广西植物ꎬ43(6):1145-1154.HUOHZꎬCAIAHꎬGUOCYꎬetal.ꎬ2023.Componentsꎬantioxidantandhypoglycemicactivitiesofwaterextractsfromdifferentvarietiesofrawtea[J].Guihaiaꎬ43(6):1145-1154.不同品种毛茶水浸出物成分及抗氧化㊁降血糖活性研究霍华珍1ꎬ蔡爱华1∗ꎬ郭春雨2ꎬ谢运昌1ꎬ李典鹏1(1.广西壮族自治区中国科学院广西植物研究所ꎬ广西植物功能物质与资源持续利用重点实验室ꎬ广西桂林541006ꎻ2.广西壮族自治区茶叶科学研究所ꎬ广西桂林541004)摘㊀要:为比较7个不同品种毛茶水浸出物活性成分及体外抗氧化与降血糖活性的差异ꎬ确定各成分与活性之间的相关性ꎬ该文选取制作六堡茶常用的7个茶树品种制备的毛茶作为研究对象ꎬ测定毛茶水浸出物及其浸膏中总多酚㊁总黄酮㊁茶多糖的含量ꎬ以DPPH 清除能力㊁ORAC值和α ̄葡萄糖苷酶㊁α ̄淀粉酶抑制作用为指标评价毛茶水浸出物的抗氧化和降血糖活性ꎬ并采用Pearson进行相关性分析ꎮ结果表明:(1)7个茶树品种毛茶水浸出物㊁总多酚㊁总黄酮㊁茶多糖含量均存在显著差异ꎬ含量最高的分别为黄金茶(53.42%ʃ0.14%)㊁桂红4号(40.87%ʃ1.09%)㊁云南大叶种(27.17%ʃ0.26%)㊁福云6号(2.70%ʃ0.02%)ꎮ(2)对DPPH 清除能力㊁ORAC值存在显著差异ꎬ在两种评价方法中均显示较好抗氧化效果的品种为六堡群体种㊁桂红4号㊁宛田种ꎮ(3)对α ̄葡萄糖苷酶㊁α ̄淀粉酶的抑制作用均显著强于阳性对照阿卡波糖ꎬ在两种评价方法中均显示较好降血糖效果的品种为六堡群体种㊁桂红4号㊁桂青种ꎮ(4)抗氧化㊁降血糖活性均与总多酚㊁总黄酮含量有较强正相关ꎮ综上认为ꎬ六堡群体种㊁桂红4号㊁宛田种㊁桂青种的毛茶品质均较好ꎬ其中六堡群体种㊁桂红4号同时具有开发成抗氧化㊁降血糖功能食品的前景ꎬ宛田种㊁桂青种分别具有开发成抗氧化㊁降血糖功能食品的潜力ꎻ总多酚㊁总黄酮对毛茶体外抗氧化㊁降血糖活性的贡献较大ꎬ在毛茶进一步的加工利用过程中应着重注意对这类成分的保护ꎮ该研究结果为开发抗氧化㊁降血糖活性更好的六堡茶产品在毛茶原料筛选和加工方式选择方面提供了科学依据ꎮ关键词:不同品种ꎬ毛茶ꎬ水浸出物成分ꎬ抗氧化活性ꎬ降血糖活性中图分类号:Q946㊀㊀文献标识码:A㊀㊀文章编号:1000 ̄3142(2023)06 ̄1145 ̄10ComponentsꎬantioxidantandhypoglycemicactivitiesofwaterextractsfromdifferentvarietiesofrawteaHUOHuazhen1ꎬCAIAihua1∗ꎬGUOChunyu2ꎬXIEYunchang1ꎬLIDianpeng1收稿日期:2022-12-18基金项目:广西科技重大专项(桂科AA20302018 ̄10)ꎻ广西植物功能物质与资源持续利用重点实验室主任基金(ZRJJ2022 ̄7ꎬZRJJ2020 ̄3ꎬZRJJ2018 ̄2)ꎻ广西植物研究所基本科研业务费项目(桂植业21004ꎬ桂植业22001)ꎻ广西科学院基本科研业务费项目(CQZ ̄C ̄1901)ꎻ桂林市创新平台和人才计划项目(20210102 ̄3ꎬ20220124 ̄13)ꎮ第一作者:霍华珍(1993-)ꎬ硕士ꎬ助理研究员ꎬ研究方向为植物资源开发与利用ꎬ(E ̄mail)hhz5951@foxmail.comꎮ∗通信作者:蔡爱华ꎬ副研究员ꎬ研究方向为植物资源开发与利用ꎬ(E ̄mail)356542930@qq.comꎮ(1.GuangxiKeyLaboratoryofPlantFunctionalPhytochemicalsandSustainableUtilizationꎬGuangxiInstituteofBotanyꎬGuangxiZhuangAutonomousRegionandChineseAcademyofSciencesꎬGuilin541006ꎬGuangxiꎬChinaꎻ2.GuangxiTeaResearchInstituteꎬGuilin541004ꎬGuangxiꎬChina)Abstract:Tocomparethedifferencesintheactivecomponentsꎬtheantioxidantandhypoglycemicactivitiesinvitrobetweensevendifferentvarietiesofrawteawaterextractsꎬandtodeterminethecorrelationbetweeneachingredientandactivityꎬthecontentsoftotalpolyphenolsꎬtotalflavonoidsandteapolysaccharidesinthewaterextractsofrawteaanditsinfusionweredetermined.TheantioxidantandhypoglycemicactivitiesofrawteawaterextractswereevaluatedbyDPPH scavengingabilityꎬORACvalueandα ̄glucosidaseandα ̄amylaseinhibitionasindicatorsꎬandPearsoncorrelationanalysiswasperformed.Theresultswereasfollows:(1)ThereweresignificantdifferencesinthecontentsofwaterextractꎬtotalpolyphenolsꎬtotalflavonoidsandteapolysaccharidesamongthesevenvarietiesꎬwhilethehighestcontentswerefoundinGoldentea(53.42%ʃ0.14%)ꎬGuihongNo.4(40.87%ʃ1.09%)ꎬYunnanbigleafspecies(27.17%ʃ0.26%)andFuyunNo.6(2.70%ʃ0.02%)ꎬrespectively.(2)TherewerealsosignificantdifferencesinDPPH scavengingabilitiesandORACvaluesamongthesevenvarietiesꎬwhilethevarietiesshowingbetterantioxidanteffectsinbothevaluationmethodsweretheLiupaogroupspeciesꎬGuihongNo.4andWantianspecies.(3)Theinhibitionsofα ̄glucosidaseandα ̄amylasebythewaterextractsofsevenvarietiesweresignificantlystrongerthanthoseofacarbosepositivecontrol.ThevarietiesshowingbetterhypoglycemiceffectsinbothevaluationmethodswereLiupaogroupspeciesꎬGuihongNo.4andGuiqingspecies.(4)Theantioxidantandhypoglycemicactivitieswereallstronglyandpositivelycorrelatedwiththecontentsoftotalpolyphenolsandflavonoids.InsummaryꎬrawteaqualityofLiupaogroupspeciesꎬGuihongNo.4ꎬWantianspeciesandGuiqingspeciesarebetterꎬamongwhichLiupaogroupspeciesandGuihongNo.4havetheprospectofdevelopingantioxidantandhypoglycemicfunctionalfoodꎻWantianspeciesandGuiqingspecieshavethepotentialofdevelopingantioxidantandhypoglycemicfunctionalfoodrespectively.Totalpolyphenolsandtotalflavonoidshaveagreatcontributiontotheinvitroantioxidantandhypoglycemicactivitiesofrawteaꎬsothatattentionshouldbepaidtotheprotectionofsuchcomponentsduringthefurtherprocessingandutilizationofrawtea.TheresultsofthisstudyprovideascientificbasisforthedevelopmentofLiupaoteaproductswithbetterantioxidantandhypoglycemicactivitiesintheselectionofrawmaterialsandprocessingmethodsofrawtea.Keywords:differentvarietiesꎬrawteaꎬwaterextractcomponentsꎬantioxidantactivityꎬhypoglycemicactivity㊀㊀六堡茶为广西特有的传统名茶ꎬ属于黑茶类ꎬ不仅具有丰富的功能成分和良好的保健功效ꎬ还给人 红㊁浓㊁陈㊁醇 的感官体验ꎬ越来越受到消费者青睐(黄敏周等ꎬ2020ꎻ马婉君等ꎬ2020)ꎬ已成为广西壮族自治区重点支持发展 千亿元茶产业 中的重要组团(李欣鞠ꎬ2019ꎻ孔妮ꎬ2020)ꎮ由于六堡茶是以鲜茶叶为原料ꎬ经初制工艺制成毛茶ꎬ在此基础上进一步精制陈化(渥堆发酵)而成ꎬ因此六堡茶生产过程中间原料毛茶的品质优劣对六堡茶品质的形成至关重要(马婉君等ꎬ2020)ꎮ然而ꎬ通过前期市场调研发现ꎬ制作六堡茶所采用的毛茶来源于较多的茶树品种且原料质量不一ꎬ从而导致生产的六堡茶成品茶质量参差不齐ꎬ严重阻碍了六堡茶产业的发展ꎬ迫切需要评价这些茶树品种之间的毛茶品质差异ꎬ有利于六堡茶生产企业准确地选择毛茶中间原料以进一步稳定成品六堡茶的品质ꎮ随着人们生活水平的提高和饮食方式的改变ꎬ糖尿病的患病率日益增长且有年轻化趋势(杨玉洁等ꎬ2021)ꎮ抗氧化剂是能捕获并中和自由基ꎬ从而保护人体免受自由基损害的一类物质(张泽生等ꎬ2017)ꎬ能增强机体的抗氧化能力ꎬ是预防和治疗糖尿病及其并发症的常用物质ꎮ随着生活中诱发自由基产生的辐射增多以及人类对健康越来越重视ꎬ抗氧化剂已成为食品以及医学领域研究的热点ꎮ然而ꎬ目前常用的抗氧化剂多为西药ꎬ有较多毒副作用ꎬ并且不能有效地控制并发症(龚艳振等ꎬ2012)ꎮ人们更青睐可长期食用㊁毒副作用小的天然抗氧化剂ꎬ选择茶产品时ꎬ除了注重口感外ꎬ人们也会更多关注它在抗氧化㊁降血糖方面的保健功效ꎮ因此ꎬ开发抗氧化㊁降血糖活性更好的六堡茶产品具有广阔的市场前景ꎮ茶多酚(黄烷醇类㊁黄酮类㊁花青素㊁酚酸)㊁茶6411广㊀西㊀植㊀物43卷多糖是茶叶中重要的活性成分ꎬ不仅具有抗氧化㊁降血糖㊁护肝等保健功效ꎬ还可以调节茶汤的口感ꎬ是决定茶叶品质的关键性物质基础(刘小玲等ꎬ2012ꎻ宋林珍等ꎬ2018ꎻ马婉君等ꎬ2020ꎻ刘淑文等ꎬ2022)ꎮ茶叶中的活性成分通常是通过煮茶或泡茶的方式被人体所吸收利用ꎬ其茶汤中浸出物含量的多少反映出茶汤的厚薄㊁滋味的强弱程度ꎬ在一定程度上还可以反映其品质的优劣(刘小玲等ꎬ2012)ꎮ因此ꎬ测定毛茶水浸出物含量及水浸出物中活性成分的含量对毛茶的品质研究具有重要意义ꎮ尽管已有较多有关六堡茶方面的研究报道ꎬ但主要集中于六堡茶成品茶活性成分(陈小强等ꎬ2008ꎻ林小珊等ꎬ2019ꎻ张均伟等ꎬ2019)㊁香气成分(温立香等ꎬ2021)㊁加工工艺(黎敏等ꎬ2021)和生物活性(叶颖等ꎬ2019ꎻ龚受基等ꎬ2020)等方面ꎬ而对六堡茶中间原料毛茶的相关研究较少报道ꎮ周伟勤等(2013)对不同季节的六堡茶原料茶树品种(六堡群体种㊁桂青种㊁云南大叶种)鲜叶中的茶多酚㊁水浸出物㊁儿茶素含量进行了测定ꎬ并对其毛茶进行感官(外形㊁汤色㊁香气㊁滋味㊁叶底)评价ꎬ结果表明六堡群体种更适制六堡茶ꎬ而该文献却未涉及毛茶中活性成分的比较分析ꎬ并且采用的茶树品种较少ꎮ林国轩等(2012)报道了9个品种不同季节茶鲜叶的活性及营养成分含量ꎬ结果表明茶多酚含量高的茶树品种适制六堡茶ꎬ虽然该文献采用的茶树品种较多ꎬ但未涉及毛茶中活性成分的比较分析ꎮ刘小玲等(2012)对7种六堡茶及单一产地(柳州)六堡毛茶制备的水浸出物㊁总多酚㊁总黄酮㊁总游离氨基酸等含量进行了测定ꎬ结果表明毛茶水浸出物含量㊁总多酚㊁总黄酮含量均比六堡茶成品茶高ꎬ但该文献所用毛茶的茶树品种不详且未涉及不同茶树品种之间活性成分的比较分析ꎮ黄敏周等(2020)对不同干燥工艺的毛茶品质进行感官评价ꎬ结果表明最佳的工艺为萎凋棚下晾晒4h辅助鼓风工艺ꎮ由此可见ꎬ已有的关于毛茶的研究仅侧重于感官品质和生化成分的含量水平方面ꎮ然而ꎬ从其保健价值角度来看ꎬ由于缺少对不同品种毛茶的活性成分及其抗氧化㊁降血糖活性比较的研究ꎬ因此无法科学判断不同品种毛茶的保健价值及成分和活性之间的相关性ꎮ 七分靠原料ꎬ三分靠工艺 ꎬ这说明毛茶原料抗氧化㊁降血糖活性与六堡茶产品的保健价值具有密切关系ꎮ本研究以制作六堡茶常用的主要茶树品种(六堡群体种㊁桂青种㊁宛田种㊁云南大叶种)以及桂红4号㊁黄金茶㊁福云6号茶树品种制备的毛茶为对象ꎬ测定毛茶水浸出物及其浸膏中活性成分的含量和体外抗氧化㊁降血糖活性ꎬ并利用Pearson进行相关性分析ꎬ确定各成分与活性之间的相关性ꎬ从活性成分含量㊁体外抗氧化以及降血糖活性3个方面综合考虑ꎬ优选出品质较好的毛茶原料品种ꎬ以期为开发抗氧化㊁降血糖活性更好的六堡茶产品在毛茶原料筛选和加工方式选择方面提供科学依据ꎮ1㊀材料与方法1.1材料和试剂鲜茶叶来自树龄为6a的7个茶树品种ꎬ于2021年4月采自广西壮族自治区茶叶科学研究所茶园ꎬ并统一按一芽三叶标准进行采摘ꎮ7个茶树品种分别为黄金茶(L1)㊁六堡群体种(L2)㊁桂红4号(L3)㊁福云6号(L4)㊁宛田种(L5)㊁云南大叶种(L6)㊁桂青种(L7)ꎮ芦丁(纯度ȡ98%ꎬ合肥博美生物科技有限责任公司)ꎬ没食子酸㊁水溶性维生素E(Trolox)㊁维生素C(VC)(纯度ȡ98%ꎬ上海士锋生物科技有限公司)ꎬα ̄葡萄糖苷酶㊁α ̄淀粉酶(美国sigma公司)ꎬ福林酚(FC)试剂㊁亚硝酸钠㊁九水合硝酸铝㊁氢氧化钠㊁95%乙醇㊁磷酸二氢钾㊁磷酸氢二钾㊁1ꎬ1 ̄二苯基 ̄2 ̄三硝基苯肼(DPPH)㊁2ꎬ2ᶄ ̄偶氮二异丁基脒二盐酸(ABAP)㊁荧光素钠㊁对硝基苯基 ̄α ̄D ̄吡喃半乳糖苷(PNPG)均为分析纯ꎬ试验用水为实验室自制超纯水ꎮ1.2仪器和设备电子天平(沈阳龙腾电子有限公司)ꎬT6紫外-可见分光光度计(北京普析通用仪器有限公司)ꎬXS205分析天平(梅特勒-托利多仪器有限公司)ꎬRIOS8超纯水系统(美国Millipore公司)ꎬ移液器(美国ThermoFisher公司)ꎬTD5A ̄WS低速离心机(湖南湘仪实验室仪器开发有限公司)ꎬHH ̄S4恒温水浴锅(上海况胜实业发展有限公司)ꎬN1100旋转蒸发仪(上海艾朗仪器有限公司)ꎬTecanSpark多功能酶标仪(上海帝肯贸易有限公司)ꎬBCD ̄213D11D双门冰箱(广东容声电器股份有限公司)ꎬMJ-系列霉菌培养箱(上海一恒科技有限公司)ꎮ74116期霍华珍等:不同品种毛茶水浸出物成分及抗氧化㊁降血糖活性研究1.3方法1.3.1六堡茶毛茶及其水浸出物的制备㊀参考黄敏周等(2020)的六堡茶毛茶制作工艺ꎬ将鲜叶先在萎凋棚下晾晒4hꎬ再经滚筒杀青㊁揉捻后以提香机70ħ烘至茶样足够干ꎬ即得六堡茶毛茶ꎮ将毛茶进一步粉碎后密封置于-20ħ保存ꎬ备用ꎮ按GB/T8305 2013(中华人民共和国国家标准化管理委员会ꎬ2013)的方法对毛茶水浸出物进行提取ꎬ所得提取液分装后置于-20ħ保存ꎬ待用ꎮ1.3.2六堡茶毛茶水浸出物含量的测定㊀按GB/T8305 2013(中华人民共和国国家标准化管理委员会ꎬ2013)的方法对六堡茶毛茶水浸出物含量进行测定ꎬ茶叶中水浸出物含量以干态质量分数(%)表示ꎮ1.3.3六堡茶毛茶水浸出物浸膏中总多酚含量的测定㊀参考GB/T8313 2018(中华人民共和国国家标准化管理委员会ꎬ2018)中的福林-酚法ꎬ并稍作改进ꎮ取毛茶水浸出物提取液0.2mL进行测定ꎬ加水补足至6mLꎬ随后分别加入FC试剂0.5mLꎬ混匀ꎬ10min后加入1.5mL20%Na2CO3溶液ꎬ充分混合后加水定容至25mLꎬ30ħ避光反应30minꎮ以水为空白对照ꎬ在760nm波长下测定吸光度Aꎮ以没食子酸为标准品绘制标准曲线ꎬ水浸出物浸膏中总多酚含量以干态质量分数(%)表示ꎬ按式(1)计算ꎮ总多酚含量(%)=水浸出物提取液中总多酚(g)/水浸出物浸膏质量(g)ˑ100(1)1.3.4六堡茶毛茶水浸出物浸膏中总黄酮含量的测定㊀参考罗磊等(2016)的硝酸铝比色法ꎬ并稍作改进ꎮ取毛茶水浸出物提取液1.0mL进行测定ꎬ用50%乙醇补足至10.0mLꎮ加入5%的亚硝酸钠溶液1.0mLꎬ混匀ꎬ10min后再加入17.6%的九水硝酸铝溶液1.0mLꎬ摇匀ꎬ放置10minꎬ加入4%的氢氧化钠溶液10.0mLꎬ用50%乙醇定容至刻度ꎬ摇匀ꎬ放置30minꎬ同时做试剂空白ꎬ于510nm波长处测定吸光度Aꎮ以芦丁为标准品绘制标准曲线ꎬ水浸出物浸膏中总黄酮含量以干态质量分数(%)表示ꎬ按式(2)计算ꎮ总黄酮含量(%)=水浸出物提取液中总黄酮质量(g)/水浸出物浸膏质量(g)ˑ100(2)1.3.5六堡茶毛茶水浸出物浸膏中茶多糖含量的测定㊀参照GB/T40632 2021(中华人民共和国国家标准化管理委员会ꎬ2021)中的苯酚-浓硫酸比色法进行茶多糖含量测定ꎮ1.3.6体外抗氧化、降血糖活性的测定1.3.6.1DPPH自由基清除能力㊀参考李明杨等(2022)的方法ꎬ并稍作改进ꎮ将毛茶水浸出物稀释成合适的系列浓度的待测样品ꎮ分别精密量取1.0mL待测样品与2.0mL95%乙醇㊁1.0mL0.12mg mL ̄1DPPH溶液混合摇匀ꎬ于室温下避光静置反应20minꎬ作为样品组ꎮ以4.0mL95%乙醇为调零组ꎬ以相同体积的95%乙醇代替DPPH溶液为样品本底组ꎬ空白组为3.0mL95%乙醇和1.0mLDPPH溶液ꎬ在517nm波长下测定吸光度ꎮ以VC为阳性对照物(浓度梯度为5㊁10㊁15㊁20㊁25μg mL ̄1)ꎬ清除率按式(3)计算ꎮ清除率(%)=[1-(T-T0)/C]ˑ100(3)式中:T表示样品组的吸光度ꎻT0表示样品本底组的吸光度ꎻC表示空白组的吸光度ꎮ毛茶水浸出物对DPPH自由基的清除能力以半数清除浓度IC50表示ꎬIC50值越小说明清除自由基的能力越强(吕平和潘思轶ꎬ2020)ꎮ1.3.6.2氧自由基吸收能力(oxygenradicalabsorbancecapacityꎬORAC)㊀参照Wen等(2016)的方法测定水浸出物样品的ORAC值ꎬ并表示为每克水浸出物浸膏(干重)的Trolox当量(μmolTE g ̄1)ꎮORAC值越大说明毛茶水浸出物的氧自由基吸收能力越强ꎬ其抗氧化活性越高ꎮ1.3.6.3α ̄葡萄糖苷酶抑制活性㊀按Pan等(2020)的方法ꎬ测定毛茶水浸出物对α ̄葡萄糖苷酶抑制活性ꎮ抑制50%的酶活所需要的样品质量浓度用IC50值表示(μg mL ̄1)ꎬIC50值越小说明对α ̄葡萄糖苷酶的抑制能力越强ꎮ1.3.6.4α ̄淀粉酶抑制活性㊀参考柳梅等(2017)的DNS法ꎬ并稍作改进ꎮ将不同浓度的样品溶液(1.0mL)和1U mL ̄1α ̄淀粉酶(1.0mL)混合ꎬ在37ħ下孵育10minꎬ随后加入1.0mL1%可溶性淀粉溶液(已煮沸糊化)ꎬ在37ħ下反应15minꎬ取出ꎬ经高温灭酶后分别加入1.5mLDNSꎬ继续煮沸8minꎮ将混合体系定容至25mLꎬ以水代替α ̄淀粉酶溶液㊁1%可溶性淀粉溶液为校零管ꎬ于540nm波长处测定其吸光度A1ꎬ以无样品的反应体系为空白对照ꎬ测定其吸光度A0ꎬ阿卡波糖为阳性对照ꎻ试剂背景以水校零ꎬ以等体积水取代DNSꎬ测定其吸光度A2ꎮ按式(4)计算抑制率ꎮ8411广㊀西㊀植㊀物43卷α ̄淀粉酶抑制率(%)=[1-(A1-A2)/(A0-A2)]ˑ100(4)抑制50%的酶活所需要的样品质量浓度用IC50值表示(μg mL ̄1)ꎬIC50值越小说明对α ̄淀粉酶的抑制能力越强ꎮ1.3.7数据处理和分析㊀每个试验设置3个平行ꎬ采用Excel2019软件进行数据处理ꎬ结果用平均值ʃ标准差(xʃs)表示ꎮ样品间各指标的差异采用SPSS19.0软件ꎬ利用单因素方差分析(one ̄wayANOVA)和Duncan多重范围检验ꎬP<0.05表示差异显著ꎬP>0.05表示无显著差异ꎮ体外活性试验中的IC50值采用SPSS19.0软件中的Probit进行计算ꎮ采用Pearson对各项指标间的相关性进行分析ꎮ2㊀结果与分析2.1不同茶树品种制备的六堡茶毛茶水浸出物及活性成分含量分析由图1可知ꎬ7个不同品种毛茶水浸出物含量变化范围为(45.18%ʃ0.11%)~(53.42%ʃ0.14%)ꎬ其中黄金茶的含量最高(53.42%ʃ0.14%)ꎬ其次是桂红4号ꎬ福云6号的最低ꎮ水浸出物含量不仅都高于国家标准(全国茶叶标准化技术委员会ꎬ2016)对中小叶种绿茶水浸出物的规定(ȡ36.0%)ꎬ而且大于45%ꎬ说明这些茶树品种内容物丰富ꎬ具有较高的耐泡性(纪鹏彬等ꎬ2021)ꎬ满足六堡茶耐冲泡特性所需的物质基础ꎮ7个不同品种毛茶水浸出物的总多酚含量在(34.50%ʃ0.22%)~(40.87%ʃ1.09%)之间ꎬ其中桂红4号的总多酚含量最高(40.87%ʃ1.09%)ꎬ比桂青种高出6.37%ꎬ两者总多酚含量存在显著性差异(P<0.05)ꎬ福云6号㊁桂青种的总多酚含量均显著低于其他品种(P<0.05)ꎬ表明不同茶树品种制备的毛茶水浸出物中总多酚含量存在差异ꎬ这与苏秋芹等(2018)的研究结果基本一致ꎮ7个不同品种毛茶水浸出物的总黄酮含量变化范围为(16.63%ʃ0.32%)~(27.17%ʃ0.26%)ꎬ含量最高的品种为云南大叶种(27.17%ʃ0.26%)ꎬ其次为六堡群体种㊁桂红4号ꎬ含量最低的品种为桂青种(P<0.05)ꎮ7个不同品种毛茶水浸出物的茶多糖含量分布范围为(1.76%ʃ0.04%)~(2.70%ʃ0.02%)ꎬ平均茶多糖含量为(2.23%ʃ0.03%)ꎬ其中福云6号的茶多糖含量最高ꎬ是云南大叶种的1.5倍ꎬ其余品种毛茶水浸出物的茶多糖含量差异也较大(P<0.05)ꎮ以上结果表明不同茶树品种制备的毛茶水浸出物含量及浸膏中活性成分的含量存在显著差异(P<0.05)ꎮ产生显著性差异的原因可能与茶树品种之间遗传基因的差异有关(刘淑文等ꎬ2022)有关ꎮ2.2不同茶树品种制备的六堡茶毛茶水浸出物抗氧化活性比较由图2可知ꎬ不同品种毛茶水浸出物对DPPH自由基的清除能力(IC50)存在显著性差异(P<0.05)ꎬ其中对DPPH自由基的清除能力最强的品种为六堡群体种[(16.73ʃ0.02)μg mL ̄1]ꎬ其次为桂红4号[(17.20ʃ0.25)μg mL ̄1]㊁宛田种[(19.51ʃ0.19)μg mL ̄1]ꎮ由图3可知ꎬ不同品种的ORAC值存在显著差异(P<0.05)ꎮ7个品种毛茶水浸出物的ORAC值在(5387.41ʃ39.71)~(6762.63ʃ50.81)μmolTE g ̄1之间ꎬ其中桂红4号㊁宛田种㊁六堡群体种的ORAC值相对较高ꎮ采用ORAC法与DPPH自由基清除法测定的7个品种毛茶水浸出物的抗氧化活性强弱顺序不完全一致ꎬ这与周金伟等(2014)的研究结果相似ꎬ产生这一结果的原因可能与这两种评价方法的自由基和作用机理不同有关ꎮ由于DPPH在溶液中以自由基形态和正离子态(DPPH+)存在ꎬ因此存在单电子转移和氢转移两种作用机制(李铉军等ꎬ2011)ꎮORAC法是一种经典的氢原子转移反应过程终止自由基链式反应ꎬ具有较高的特异性(续洁琨等ꎬ2006)ꎮ不同品种毛茶水浸出物中含有的大分子抗氧化剂含量可能不同ꎬ而大分子抗氧化剂由于空间位阻不能与DPPH自由基反应(续洁琨等ꎬ2006)ꎬ因此导致抗氧化效果有偏差ꎮ虽然所测样品采用ORAC法与DPPH自由基清除法测定的结果存在差异ꎬ但桂红4号㊁宛田种㊁六堡群体种水浸出物的ORAC值和DPPH自由基清除能力均较高ꎬ说明这3个品种的抗氧化活性优于其余品种ꎮ2.3不同茶树品种制备的六堡茶毛茶水浸出物体外降血糖活性比较由图4可知ꎬ7个品种毛茶水浸出物对α ̄葡萄糖苷酶的抑制作用均显著强于阳性对照阿卡波糖IC50=(606.21ʃ4.30)μg mL ̄1ꎬ说明这些品种均94116期霍华珍等:不同品种毛茶水浸出物成分及抗氧化㊁降血糖活性研究不同字母表示不同茶树品种制备的毛茶水浸出物及其浸膏中活性成分的含量存在显著差异(P<0.05)ꎻ相同字母表示无显著差异(P>0.05)ꎮ下同ꎮDifferentlettersindicatethatthereisasignificantdifferenceinthecontentsofactivecomponentsinthewaterextractandextractofrawteapreparedbydifferentteavarieties(P<0.05)ꎻthesameletterindicatesnosignificantdifference(P>0.05).Thesamebelow.图1㊀不同茶树品种制备的毛茶水浸出物浸膏中活性成分的含量Fig.1㊀Contentsofactivecomponentsinrawteawaterextractspreparedfromdifferentteavarieties图2㊀不同茶树品种制备的毛茶水浸出物对DPPH自由基的清除能力Fig.2㊀DPPHfreeradicalscavengingabilityofrawteawaterextractspreparedfromdifferentteavarieties可作为α ̄葡萄糖苷酶的天然抑制剂ꎬ其中六堡群体种㊁桂青种㊁桂红4号水浸出物对α ̄葡萄糖苷酶的抑制效果更佳ꎬIC50值分别为(9.66ʃ0.11)㊁(10.87ʃ0.07)㊁(11.06ʃ0.11)μg mL ̄1ꎮ福云6图3㊀不同茶树品种制备的毛茶水浸出物的氧自由基吸收能力值Fig.3㊀ORACvaluesofrawteawaterextractspreparedfromdifferentteavarieties号㊁宛田种对α ̄葡萄糖苷酶的抑制作用均显著低于六堡群体种(P<0.05)ꎬ其余品种对α ̄葡萄糖苷酶的抑制作用差异不大ꎮ由图5可知ꎬ部分品种对α ̄淀粉酶的抑制作用有显著差异(P<0.05)ꎮ0511广㊀西㊀植㊀物43卷7个品种毛茶水浸出物对α ̄淀粉酶的半数抑制浓度(IC50)在(89.13ʃ1.07)~(160.15ʃ1.88)μg mL ̄1之间ꎬ均具有显著的α ̄淀粉酶抑制效果ꎬ其中六堡群体种㊁桂红4号㊁桂青种比其他品种有更好的抑制α ̄淀粉酶活效果ꎮ两种体外降血糖评价方法的结果均显示六堡群体种㊁桂红4号㊁桂青种的体外降血糖活性更好ꎮ图4㊀不同茶树品种制备的毛茶水浸出物对α ̄葡萄糖苷酶的抑制作用Fig.4㊀Inhibitionofα ̄glucosidasebyrawteawaterextractspreparedfromdifferentteavarieties图5㊀不同茶树品种制备的毛茶水浸出物对α ̄淀粉酶的抑制作用Fig.5㊀Inhibitionofα ̄amylasebyrawteawaterextractspreparedfromdifferentteavarieties2.4不同茶树品种制备的六堡茶毛茶水浸出物活性成分与体外抗氧化、降血糖活性的相关性分析由于样品活性越强ꎬ其IC50值则越小ꎬ因此进行活性成分与抗氧化㊁降血糖活性的相关性分析时用1/IC50表示样品活性的强弱ꎮ由表1可知ꎬ总多酚含量分别与DPPH自由基清除作用和ORAC值呈显著的正相关ꎬ相关系数分别为0.751和0.817ꎬ表明总多酚含量与体外抗氧化活性有显著的相关性ꎮ总多酚含量与总黄酮含量呈显著正相关(P<0.05)ꎬ这是因为黄酮是多酚的组成成分之一ꎮ总多酚含量与α ̄葡萄糖苷酶㊁α ̄淀粉酶抑制作用有较好的正相关关系ꎬ总黄酮含量分别与α ̄葡萄糖苷酶㊁α ̄淀粉酶抑制作用有中等强度㊁显著正相关ꎬ表明体外降血糖活性与总多酚㊁总黄酮相关ꎮDPPH自由基清除作用与ORAC值具有显著正相关ꎬ表明这两种抗氧化活性的评价方法可以相互佐证ꎮ同理ꎬα ̄葡萄糖苷酶抑制作用与α ̄淀粉酶抑制作用相关系数为0.929(P<0.01)ꎬ表明这两种体外降血糖的评价方法可以互为辅助参考ꎮ而茶多糖含量与体外抗氧化㊁降血糖活性均没有正相关性ꎬ这与余启明等(2021)和宋林珍等(2018)的研究结果不同ꎬ可能是不同品种之间的茶多糖的分子量㊁化学成分㊁结构和立体构象不同从而导致体外降血糖活性不同(宋林珍等ꎬ2018)ꎬ具体影响因素有待进一步研究ꎮ3㊀讨论与结论7个品种毛茶水浸出物及其浸膏中总多酚㊁总黄酮㊁茶多糖的含量之间存在显著差异(P<0.05)ꎬ体外抗氧化㊁降血糖活性也存在差异ꎮ两种抗氧化评价方法结果均表明六堡群体种㊁桂红4号㊁宛田种毛茶水浸出物的抗氧化活性较好ꎬ同时总多酚㊁总黄酮含量也较高ꎮ相关性分析表明总多酚含量与抗氧化活性有显著的正相关ꎬ总黄酮含量与抗氧化活性也有较好的正相关ꎬ因此总多酚㊁总黄酮对毛茶抗氧化活性的贡献较大ꎮ总多酚含量与α ̄葡萄糖苷酶㊁α ̄淀粉酶抑制作用有较好的正相关ꎬ总黄酮含量分别与α ̄葡萄糖苷酶㊁α ̄淀粉酶抑制作用有中等强度㊁显著正相关ꎬ表明总多酚和总黄酮均具有降血糖活性ꎬ同时两者可能还具有协同作用ꎮ两种降血糖评价方法结果均表明六堡群体种㊁桂红4号㊁桂青种的毛茶水浸出物比其余品种的降血糖活性更好ꎬ其中六堡群体种㊁桂红4号毛茶水浸出物的总多酚㊁总黄酮含量均较高ꎬ而桂青种的却低于其余品种ꎮ可能与以下两个因素有关:潘福璐等(2020)研究表15116期霍华珍等:不同品种毛茶水浸出物成分及抗氧化㊁降血糖活性研究表1㊀不同品种毛茶水浸出物的1/IC50值㊁ORAC值与各项指标的相关性分析Table1㊀Correlationanalysisof1/IC50valueꎬORACvalueandvariousindexesofrawteawaterextractsfromdifferentvarieties指标Index总多酚Totalpolyphenols总黄酮Totalflavonoids茶多糖TeapolysaccharideDPPH清除能力DPPHscavengingabilityORAC值ORACvalueα ̄葡萄糖苷酶抑制作用α ̄glucosidaseinhibitionα ̄淀粉酶抑制作用α ̄amylaseinhibition总多酚Totalpolyphenols10.850 -0.4950.751 0.817 0.5010.521总黄酮Totalflavonoids1-0.5370.6600.6140.4490.757茶多糖Teapolysaccharide1-0.419-0.394-0.586-0.839DPPH 清除能力DPPH scavengingability10.855 0.6220.564ORAC值ORACvalue10.2850.312α ̄葡萄糖苷酶抑制作用α ̄glucosidaseinhibition10.929 α ̄淀粉酶抑制作用α ̄amylaseinhibition1㊀注: 表示在0.05级别(双尾)ꎬ相关性显著ꎻ 表示在0.01级别(双尾)ꎬ相关性显著ꎮ㊀Note: indicatessignificantcorrelationat0.05(two ̄tailed)ꎻ indicatessignificantcorrelationat0.01(two ̄tailed).明茶叶中多酚组分ECG分别与EGCG㊁GCG两两组合对抑制α ̄葡萄糖苷酶活性存在协同作用ꎬ且这两种组合及组合内不同含量配比对α ̄葡萄糖苷酶活性抑制作用存在差异ꎬ因此不同品种毛茶水浸出物中ECG㊁EGCG㊁GCG的含量组成差异对降血糖活性产生影响ꎻ茶多糖的单糖组成㊁分子量㊁支链结构㊁立体构象是影响其降血糖活性的重要因素(杨玉洁等ꎬ2021)ꎬ如糖醛酸能明显影响多糖的活性ꎬ而茶树品种间的中性糖㊁糖醛酸含量存在极显著差异(刘思思ꎬ2009)ꎬ因此不同品种毛茶水浸出物的茶多糖的单糖组成㊁分子量㊁支链结构和有效结构含量等方面可能不同ꎬ从而影响其降血糖活性ꎮ桂青种的体外降血糖活性的物质基础及其作用机制有待进一步研究ꎮ综上所述ꎬ六堡群体种㊁桂红4号㊁宛田种㊁桂青种的毛茶品质均较好ꎬ其中六堡群体种㊁桂红4号同时具有开发抗氧化㊁降血糖功能食品的前景ꎬ宛田种㊁桂青种分别具有开发抗氧化㊁降血糖功能食品的潜力ꎻ总多酚㊁总黄酮对毛茶体外抗氧化㊁降血糖活性均有较大贡献ꎻ总多酚具有一定的热稳定性ꎬ但在高湿高温及强光条件下含量迅速下降(沈生荣ꎬ1993)ꎮ为得到抗氧化㊁降血糖活性更好的六堡茶产品ꎬ在毛茶进一步的加工利用过程应着重注意对这类成分的保护ꎬ不宜长时间使用高温㊁强光直射的加工方式ꎮ储存时应密封避光置于干燥阴凉通风处或冷藏ꎮ总黄酮是总多酚的组成成分之一ꎬ因此其保护方法与总多酚的一致ꎮ另外ꎬ为保证同一品种茶树品质的稳定性ꎬ在对茶树进行异地引种栽培时ꎬ宜选择地域㊁土壤㊁气候㊁海拔高度等条件与原产地相近的地理环境(刘淑文等ꎬ2022)ꎬ同时还需采用统一的茶叶采摘标准ꎮ本研究为开发抗氧化㊁降血糖活性更好的六堡茶产品在毛茶原料筛选和加工方式选择方面提供科学依据ꎮ参考文献:CHENXQꎬYEYꎬCHENGHꎬetal.ꎬ2008.StudiesonphysicalandchemicalpropertiesofLiupaotea[J].ChinAgricSciBullꎬ24(7):77-80.[陈小强ꎬ叶阳ꎬ成浩ꎬ等ꎬ2008.六堡茶的理化分析研究[J].中国农学通报ꎬ24(7):77-80.]GONGYZꎬXUYJꎬLIUHWꎬetal.ꎬ2012.Progressofsynergisticantioxidanteffectsofnaturalantioxidants[J].FoodSciTechnolꎬ37(6):264-267.[龚艳振ꎬ徐亚2511广㊀西㊀植㊀物43卷。
我国科学家揭示“节俭基因”的作用机制

第3期2018年6月石河子科技不超过100ppm。
风量的多少影响锅炉对应负荷下的氧量,因此风量的多少应根据负荷变化及时调整。
需要注意的是:锅炉负荷低于60t/h时,不应过于追求控制氧量而减少送风,这样会造成一次风量不足导致燃烧器回火以及锅炉壶口流渣不畅,此时应适当增加氧量、增加尿素溶液喷入量来控制NOx。
3.2配风方式燃烬风主要控制对象是CO,CO反映了前置炉燃烧还原性气氛程度。
当锅炉工况稳定时,一般控制燃烬风开度30%~40%之间,CO含量不超过75ppm。
当燃烬风逐渐开大时,NOx会随之降低,CO逐步升高。
一、二次风配比原则:首要是确保一次风压,在此前提下,为保证二次风刚性可尽量关小二次风小风门、开大二次风总风门。
较小的一次室风率能够降低煤粉初始燃烧氧量,从而抑制NOx生成,保证二次风刚性可延长固定碳在一次室内燃烧时间。
当外界负荷波动较大时,燃烬风可以作为调节一、二次风率大小的手段,控制一次室燃烧气氛,从而抑制NOx生成,保证环保参数达标排放。
3.3喷枪运行工况当一只或多只喷枪管路堵塞时,NOx就难于控制,尿素用量大幅上升,氨逃逸大幅增加。
最终导致过量氨与三氧化硫反应生成硫酸氢氨,堵塞空气预热器。
4存在的问题及今后改进的方向利用停炉之际,对旋风炉进行全面检查。
发现两侧二次风口均有较为严重的变形,结焦严重,占风口通道面积约40%。
二次室结焦严重;积灰严重,尤其是催化剂及各处水平烟道。
运行一段时间后内部阻力增加,脱硝效率下降,氨逃逸增加,严重时限制锅炉带负荷能力。
催化剂内积灰板结,难以彻底清理。
针对低氮燃烧的负面影响,提出今后改进的方向:提高二次风口与燃尽风口材质的耐热性;尾部烟道的吹灰器更换成大功率声波吹灰器;对尾部烟道烟气流场进行优化;运行期间加大氨逃逸的考核。
参考文献[1]钟礼金,宋玉宝.锅炉SCR烟气脱硝空气预热器堵塞原因及解决措施[J].热力发电, 2012,41(8):45-48.[2]Hans Jensen-Holm,Nan-YU Topsoe,崔建华.选择催化还原(SCR)脱硝技术在中国燃煤锅炉上的应用(上)[J].热力发电,2007(8):13-18.[3]Hans Jensen-Holm,Nan-YU Topsoe,崔建华.选择催化还原(SCR)脱硝技术在中国燃煤锅炉上的应用(下)[J].热力发电,2007(9):10-15.我国科学家揭示“节俭基因”的作用机制在国家重点研发计划“蛋白质机器与生命过程调控”重点专项的支持下,厦门大学林圣彩教授团队揭示了乙酰转移酶TIP60作为一个“节俭基因”的功能和作用机制。
emuchnet国家自然科学基金生命科学学部食品科学学科食品加工学基础

国家自然科学基金生命科学学部食品科学学科食品加工学基础(C2002)面上项目和青年基金资助项目清单(共133项,累计金额:6,834.0万元)1.基于量子点/NBD荧光比率的高灵敏、高通量功能寡糖筛选和构效关系评估体系构建及其应用负责人:张洪涛参与人:吴剑荣, 许晓鹏, 朱德强, 刘登峰, 杨天怡, 王顺治, 李丹金额:25万申请时间:2012学科代码:制糖(C200202)项目批准号:31201384申请单位:江南大学研究类型:基础研究关键词:功能寡糖筛选;构效关系;可定量;糖芯片;量子点查看摘要收藏负责人:王韧参与人:周星, 牛猛, 徐逸木, 王勇金额:21万申请时间:2012学科代码:食品油脂加工(C200201)项目批准号:31201381申请单位:江南大学研究类型:基础研究关键词:淀粉蔗糖酶;蜡质玉米淀粉;转糖基活力;支链延长修饰;消化特性查看摘要收藏负责人:郭顺堂参与人:徐婧婷, 刘贺, 左锋, 李亮亮, 谢来超, 范玲慧, 姚美伊, 施小迪, 王睿璨金额:82万申请时间:2012学科代码:饮料冷饮(C200211)项目批准号:31271941申请单位:中国农业大学研究类型:基础研究关键词:豆乳;植酸;钙镁离子;结合作用;蛋白粒子结构查看摘要收藏负责人:张公亮参与人:朴美兰, 唐亮, 葛静慧, 村田芳行申请单位:大连工业大学研究类型:基础研究关键词:类胡萝卜素;裂解;硫醚类香料;影响机制;细胞毒性查看摘要收藏负责人:王蓓参与人:徐虹, 孙啸涛, 朱雨晨, 马艳丽, 陈海清, 王超金额:25万申请时间:2012学科代码:乳加工(C200204)项目批准号:31201392申请单位:北京工商大学研究类型:基础研究关键词:传统酸凝奶酪;风味重组;特征风味组分;感官质量评价;风味质量模型查看摘要收藏负责人:王栋参与人:王海燕, 盛嘉, 李鸣, 郑露露, 朱增亮, 吴荣, 郑礼月金额:80万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271920申请单位:江南大学研究类型:基础研究关键词:丝状真菌;华根霉;转录组学;形态工程;脂肪酶查看摘要收藏负责人:王淑军参与人:房耀维, 吕明生, 焦豫良, 刘姝, 蔡阮鸿, 李卫娟, 葛亮金额:78万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271929申请单位:淮海工学院研究类型:基础研究关键词:晶体结构;高温α-淀粉酶热稳定性;超嗜热古菌;钙离子;分子机制查看摘要收藏负责人:吴拥军参与人:周藜, 孟望霓, 韩丽珍, 王嫱, 卢彪, 罗熹, 吴玉俊, 赖茂林, 刘艳敏申请单位:贵州大学研究类型:基础研究关键词:水豆豉;生物安全;控制机制查看摘要收藏负责人:宋洪波参与人:安凤平, 童金华, 仇东朝, 赖彩如金额:68万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31271913申请单位:福建农林大学研究类型:基础研究关键词:过热蒸汽;膨化;水果;蔬菜;动力查看摘要收藏负责人:李晓然参与人:罗义勇, 杨恩, 张忠华, 宫路路, 栾建军, 向新, 李洁, 姜宏章, 姜黎明金额:50万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260397申请单位:昆明理工大学研究类型:基础研究关键词:传统发酵食品;群落结构;元基因组;元转录组查看摘要收藏负责人:李燕萍参与人:何庆华, 季艳伟, 王丹, 唐梦醒, 潘一峰, 王瑶金额:50万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260389申请单位:南昌大学研究类型:基础研究关键词:米曲霉;中性蛋白酶;分子模拟;定向进化;最适pH查看摘要收藏负责人:梁瑞红参与人:陈军, 李燕, 徐雨佳, 袁敏, 余雯, 帅希祥, 王召君, 吴双双, 夏天添申请单位:南昌大学研究类型:基础研究关键词:果胶;高压均质;分子结构;构象行为;功能性质查看摘要收藏负责人:孟昆参与人:石鹏君, 王慧, 罗建杰, 赵亮, 邵弨, 徐晓露, 涂涛金额:80万申请时间:2012学科代码:食品添加剂(C200210)项目批准号:31271937申请单位:中国农业科学院饲料研究所研究类型:基础研究关键词:多聚半乳糖醛酸酶;定向进化;理性设计;DNA改组;定点突变查看摘要收藏负责人:王荣春参与人:程翠林, 赵海田, 秦文信, 王立枫, 李明金额:23万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31201403申请单位:哈尔滨工业大学研究类型:基础研究关键词:亚临界水;植物基质;新生抗氧化物;生成动力学查看摘要收藏负责人:董庆利参与人:张东来, 丁甜, 张宇玲, 吴酉芝, 熊成, 曾静, 梁娜, 邱静, 郑丽敏金额:83万申请时间:2012学科代码:肉加工(C200203)项目批准号:31271896申请单位:上海理工大学研究类型:基础研究关键词:预测微生物学;风险评估;暴露评估;参数优化查看摘要收藏负责人:杨锡洪参与人:章超桦, 卢虹玉, 高加龙, 解万翠, 王林, 张殿军申请单位:广东海洋大学研究类型:基础研究关键词:壳聚糖;牡蛎;麻痹性贝类毒素;结合蛋白;脱除机理查看摘要收藏负责人:张燕参与人:王蓉蓉, 姚佳, 董鹏, 易俊洁, 徐茜, 孔民, 张雅杰, 李瑶金额:78万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31271910申请单位:中国农业大学研究类型:基础研究关键词:高静压;绿色;降解;机制查看摘要收藏负责人:丁重阳参与人:许正宏, 陶冠军, 王玉红, 顾正华, 王琼, 乔双逵, 郑俊丽金额:76万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271918申请单位:江南大学研究类型:基础研究关键词:灵芝多糖;合成途径;代谢调控;菌体形态;液体发酵查看摘要收藏负责人:刘茹参与人:熊善柏, 刘友明, 张亮子, 尹涛, 李俊杰金额:21万申请时间:2012学科代码:肉加工(C200203)项目批准号:31201391申请单位:华中农业大学研究类型:基础研究关键词:肌球蛋白;自组装;流变特性;结构查看摘要收藏负责人:黄业传参与人:严成, 熊双丽, 李凤, 吴照民, 丁建刚申请单位:西南科技大学研究类型:基础研究关键词:高压;肌内脂肪;氧化;酶查看摘要收藏负责人:张丽霞参与人:黄纪念, 芦鑫, 李晋, 从珊, 王瑞萍, 刘晓阳金额:21万申请时间:2012学科代码:食品油脂加工(C200201)项目批准号:31201383申请单位:河南省农业科学院研究类型:基础研究关键词:芝麻林素;芝麻素酚;杂多酸;反应体系;定向转化查看摘要收藏负责人:徐明生参与人:黄占旺, 吴国平, 涂勇刚, 杜华英, 汤群, 王新龙金额:52万申请时间:2012学科代码:蛋加工(C200205)项目批准号:31260385申请单位:江西农业大学研究类型:基础研究关键词:卵转铁蛋白;树突状细胞;免疫调节;分子机制查看摘要收藏负责人:徐为民参与人:王道营, 刘芳, 蒋宁, 张牧焓, 王炜, 李超, 王超, 董晗金额:86万申请时间:2012学科代码:肉加工(C200203)项目批准号:31271891申请单位:江苏省农业科学院研究类型:基础研究关键词:热休克蛋白90;肌内磷脂;水解;氧化;肌肉微结构查看摘要收藏负责人:陆兆新参与人:曹林, 汪晓鸣, 魏照辉, 李远宏, 李金良, 章栋梁, 毕华, 李春燕申请单位:南京农业大学研究类型:基础研究关键词:fengycin;敲除或过表达;信号蛋白;分子调控;蛋白质组学查看摘要收藏负责人:张海德参与人:袁德保, 蒋志国, 蒋欣欣, 周娅, 王伟涛金额:48万申请时间:2012学科代码:食品添加剂(C200210)项目批准号:31260401申请单位:海南大学研究类型:基础研究关键词:木瓜蛋白酶;亲和双水相系统;固定化金属离子;分配行为;机理查看摘要收藏负责人:孔祥珍参与人:华欲飞, 陈业明, 张彩猛, 阮奇珺, 丁秀臻, 赵沙沙, 王金艳金额:26万申请时间:2012学科代码:食品油脂加工(C200201)项目批准号:31201380申请单位:江南大学研究类型:基础研究关键词:大豆球蛋白;酶水解;巯基;二硫键;半胱氨酸查看摘要收藏负责人:张明霞参与人:姜小莹, 高晗, 杨天佑, 魏燕丽, 张怀胜金额:21万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31201410申请单位:河南科技学院研究类型:基础研究关键词:红葡萄酒;香气释放;花色苷;影响;感官分析查看摘要收藏负责人:杨希娟参与人:党斌, 王树林, 邵东燕, 赵媛, 耿贵工, 李萍, 刘新红申请单位:青海省农林科学院研究类型:基础研究关键词:西藏灵菇;抗氧化性;胞外多糖;产生机制;流变学特性查看摘要收藏负责人:朱丽霞参与人:张琴, 张玲, 范瑛阁, 王伟华, 张锐利, 牛荣, 阿依提拉.吾布力卡斯穆金额:55万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260393申请单位:塔里木大学研究类型:基础研究关键词:酿酒酵母;遗传多样性;发酵特性;葡萄酒;种质资源库查看摘要收藏负责人:李春参与人:刘丽波, 郑遂, 孙迪, 李培照, 孙金威金额:23万申请时间:2012学科代码:乳加工(C200204)项目批准号:31201397申请单位:东北农业大学研究类型:基础研究关键词:抗冻干胁迫;保加利亚乳杆菌;氯化钠刺激;糖代谢;biomarkers查看摘要收藏负责人:李崎参与人:田亚平, 郑飞云, 刘春凤, 王金晶, 许维娜, 宋群, 李二娟金额:60万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271919申请单位:江南大学研究类型:基础研究关键词:泡沫活性蛋白质;蛋白质Z;二级结构;理化性质;啤酒查看摘要收藏负责人:汪学德参与人:张振山, 刘玉兰, 魏安池, 马宇翔, 纪俊敏, 刘兵戈, 黄维, 刘日斌, 胡华丽申请单位:河南工业大学研究类型:基础研究关键词:油脂;生产;苯并芘;机理;研究查看摘要收藏负责人:钟秋平参与人:李从发, 蒋志国, 范丽霞, 余雪梅, 张博金额:45万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260398申请单位:海南大学研究类型:基础研究关键词:荔枝酒;酿酒酵母;醋酸;代谢;调控查看摘要收藏负责人:梁琪参与人:甘伯中, 乔海军, 米兰, 张炎, 秦虹, 巨玉佳, 徐瑛金额:50万申请时间:2012学科代码:乳加工(C200204)项目批准号:31260383申请单位:甘肃农业大学研究类型:基础研究关键词:牦牛乳;硬质干酪;成熟期;苦味肽查看摘要收藏负责人:顾振新参与人:周玉林, 沈昌, 王顺民, 杨润强, 顾颖娟, 郭强晖金额:86万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31271912申请单位:南京农业大学研究类型:基础研究关键词:芸薹属芽苗菜;低氧胁迫;热激处理;异硫氰酸酯;富集机理查看摘要收藏负责人:杜金华参与人:金玉红, 张丽敏, 郭萌萌, 王岱杰, 何桂芬, 祝敏, 王聪, 李盼盼, 张庆宣申请单位:山东农业大学研究类型:基础研究关键词:小麦芽;水溶性阿拉伯木聚糖;界面扩张特性;啤酒泡沫;分子结构特征查看摘要收藏负责人:段长青参与人:石英, 蔡建, 段亮亮, 许晓青, 宋文凤, 兰义宾, 蒋仁金额:88万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271926申请单位:中国农业大学研究类型:基础研究关键词:橡木多酚;辅色素;辅色素化效应;呈色机理;红葡萄酒查看摘要收藏负责人:胡旭佳参与人:李晓东, 陈邈, 秦慧, 陈小双, 何文亮, 王佩瑾金额:54万申请时间:2012学科代码:调味食品(C200209)项目批准号:31260399申请单位:昆明理工大学研究类型:基础研究关键词:马槟榔;甜水效应;甜味受体查看摘要收藏负责人:李盛钰参与人:赵玉娟, 张莉, 张雪, 李达, 张健金额:21万申请时间:2012学科代码:乳加工(C200204)项目批准号:31201394申请单位:吉林省农业科学院研究类型:基础研究关键词:益生菌;基因组改组;蛋白组学查看摘要收藏负责人:孙建霞参与人:白卫滨, 刘风霞, 林先哲, 仝雷, 吴珍菊, 刘霞金额:26万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31201402申请单位:广东工业大学研究类型:基础研究关键词:超声波;天竺葵素-3-葡萄糖苷;自由基;降解;机理查看摘要收藏负责人:孟凌华参与人:毕丹, 林琦, 杨锐, 苗平金额:21万申请时间:2012学科代码:食品添加剂(C200210)项目批准号:31201418申请单位:上海交通大学研究类型:基础研究关键词:紫苏花青素;神经保护;氧化应激;β-淀粉样蛋白;构效关系查看摘要收藏负责人:宋诗清参与人:孙涛, 朱广用, 朱建才, 窦银花, 荣志伟金额:22万申请时间:2012学科代码:调味食品(C200209)项目批准号:31201415申请单位:上海应用技术学院研究类型:基础研究关键词:氧化脂肪;特征风味前体;偏最小二乘回归(PLSR);定量预测模型;调控氧化查看摘要收藏负责人:范小平参与人:向红, 宋贤良, 张钦发, 李朗, 邢明, 张丽影, 缪丽华金额:21万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31201401申请单位:华南农业大学研究类型:基础研究关键词:压差膨化;多孔结构;气泡生长;质构;数值模拟查看摘要收藏负责人:方尚玲参与人:陈茂彬, 彭东海, 镇达, 刘超, 李习, 景艳艳金额:15万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271928申请单位:湖北工业大学研究类型:基础研究关键词:异常汉逊氏酵母;基因组文库;转座标签库;关键基因;分子机制查看摘要收藏负责人:刘国艳参与人:张旭, 葛庆丰, 张振芳, 蔡丽丽, 毛文东, 叶群金额:26万申请时间:2012学科代码:食品油脂加工(C200201)项目批准号:31201379申请单位:扬州大学研究类型:基础研究关键词:食用植物油;苯并(a)芘;来源;迁移变化;形成机理查看摘要收藏负责人:黄略略参与人:段续, 乔方, 方长发, 刘芳, 杜晓彦金额:23万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31201399申请单位:深圳职业技术学院研究类型:基础研究关键词:微波真空;干燥;介电特性;果蔬;传热传质查看摘要收藏负责人:罗欣参与人:朱立贤, 毛衍伟, 梁荣蓉, 张一敏, 董鹏程, 卢昊, 侯旭金额:88万申请时间:2012学科代码:肉加工(C200203)项目批准号:31271901申请单位:山东农业大学研究类型:基础研究关键词:牛肉;温度;品质;蛋白质组学;能量代谢查看摘要收藏负责人:胡萍参与人:谢和, 朱秋劲, 曾海英, 王晓宇, 陈韵, 朱建军, 唐玲金额:56万申请时间:2012学科代码:肉加工(C200203)项目批准号:31260379申请单位:贵州大学研究类型:基础研究关键词:传统发酵肉制品;乳酸菌多样性;抗氧化菌株查看摘要收藏负责人:陈勉华参与人:李贞景, 李风娟, 王玉荣, 李伟, 马人杰, 黄玉芳, 马博雅, 鲁丽娟金额:76万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271915申请单位:天津科技大学研究类型:基础研究关键词:红曲米;胆固醇;非他汀类物质;洛伐他汀;高胆固醇血症查看摘要收藏负责人:曹慧参与人:李海洲, 于劲松, 石服鑫, 张成楠金额:21万申请时间:2012学科代码:食品添加剂(C200210)项目批准号:31201421申请单位:上海理工大学研究类型:基础研究关键词:胶原抗冻多肽;制备;分子结构;抗冻机理查看摘要收藏负责人:刘国琴参与人:薛积玉, 韩立娟, 王丽娟, 孙英, 阎乃珺, 巩凡, 荆璐, 范静美金额:83万申请时间:2012学科代码:食品油脂加工(C200201)项目批准号:31271885申请单位:华南理工大学研究类型:基础研究关键词:油脂;热加工;缩水甘油酯;机理;控制方法查看摘要收藏负责人:罗立新参与人:崔堂兵, 林晓珊, 罗剑飞, 劳永民, 卢良坤, 邹晓蕾金额:82万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271924申请单位:华南理工大学研究类型:基础研究关键词:酱油发酵;嗜盐四联球菌;分子伴侣;胁迫应答;甜菜碱运输系统查看摘要收藏负责人:郑志永参与人:詹晓北, 蒋芸, 冯杰, 朱德强, 康晶, 李国顺, 王凤伟金额:80万申请时间:2012学科代码:制糖(C200202)项目批准号:31271888申请单位:江南大学研究类型:基础研究关键词:微生物多糖;信号转导;氧信号;土壤杆菌;热凝胶查看摘要收藏负责人:吴宏参与人:吴洪斌, 宋健, 武冬梅, 刘成江, 王俊刚金额:45万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260396申请单位:新疆农垦科学院研究类型:基础研究关键词:新疆;红曲霉;种质资源;代谢产物查看摘要收藏负责人:田红玉参与人:陈海涛, 孙宇梅, 孙金沅, 刘永国, 梁森, 陈凯, 戴怡凤, 公秀琴, 王亚玲金额:85万申请时间:2012学科代码:食品添加剂(C200210)项目批准号:31271932申请单位:北京工商大学研究类型:基础研究关键词:含硫食品香料;手性;立体异构体;香气特性;天然组成查看摘要收藏负责人:郑先哲参与人:刘成海, 金长江, 姚鑫淼, 牟艳秋, 刘海军, 林甄, 孙井坤, 孙宇金额:85万申请时间:2012学科代码:水果、蔬菜加工(C200206)项目批准号:31271911申请单位:东北农业大学研究类型:基础研究关键词:浆果;微波;泡沫干燥;活性成分;品质查看摘要收藏负责人:张军翔参与人:张惠玲, 李鹏, 田晓菊, 李应龙, 李永山, 郝笑云, 张媛金额:45万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260392申请单位:宁夏大学研究类型:基础研究关键词:红葡萄酒;颜色;多酚;响应机理;稳定性查看摘要收藏负责人:范贵生参与人:董同力嘎, 杨晓清, 田建军, 鲁艳艳, 胡惠敏, 郑策, 王瑞利金额:48万申请时间:2012学科代码:乳加工(C200204)项目批准号:31260382申请单位:内蒙古农业大学研究类型:基础研究关键词:干酪;介电特性;品质特性;成熟度;质构查看摘要收藏负责人:陈忠军参与人:李正英, 殷文政, 张保军, 王新亮, 付倩倩, 范帅, 李海瑄, 高鹤尘金额:45万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260390申请单位:内蒙古农业大学研究类型:基础研究关键词:乳杆菌;抑真菌物质;结构;机理;生物防腐剂查看摘要收藏负责人:廖国周参与人:彭云霞, 李永强, 殷红, 何凡, 石磊申请单位:云南农业大学研究类型:基础研究关键词:葡萄籽提取物;烤羊肉;杂环胺;抑制;机理查看摘要收藏负责人:于淑娟参与人:朱思明, 朱立才, 王晓华, 赵振刚, 管永光, 俞思明金额:60万申请时间:2012学科代码:制糖(C200202)项目批准号:31271889申请单位:华南理工大学研究类型:基础研究关键词:美拉德产物;同位素示踪;5-羟甲基糠醛;4-甲基咪唑查看摘要收藏负责人:于殿宇参与人:胡立志, 富校轶, 魏贞伟, 宋玉卿, 王俊国, 张春艳, 王玥, 宋云花, 刘鑫金额:80万申请时间:2012学科代码:食品油脂加工(C200201)项目批准号:31271886申请单位:东北农业大学研究类型:基础研究关键词:CO2超临界;定向;氢化;TFAs;可控查看摘要收藏负责人:陈从贵参与人:铃木敦士, 王武, 周存六, 方红美, 马飞, 卢秋红, 陈星, 周燕子, 肖雄金额:86万申请时间:2012学科代码:肉加工(C200203)项目批准号:31271893申请单位:合肥工业大学研究类型:基础研究关键词:肌球蛋白;肌动球蛋白;超高压;多糖;响应机制查看摘要收藏负责人:孙为正参与人:赵海锋, 苏国万, 仇超颖, 周非白, 陈楠楠, 杨园媛申请单位:华南理工大学研究类型:基础研究关键词:肌原纤维蛋白质;蛋白质氧化;自由基;乳化特性;构效关系查看摘要收藏负责人:张晖参与人:肖建辉, 钱海峰, 朱松, 齐希光, 申迎宾, 苑华宁, 杨凌霄金额:67万申请时间:2012学科代码:食品添加剂(C200210)项目批准号:31271934申请单位:江南大学研究类型:基础研究关键词:抗菌肽;细胞膜色谱;筛选;食品查看摘要收藏负责人:李锋参与人:唐晓珍, 连玉晶, 姚颖斐, 汪东, 孙田垒, 陈洲金额:26万申请时间:2012学科代码:调味食品(C200209)项目批准号:31201417申请单位:山东农业大学研究类型:基础研究关键词:生姜;II相酶;诱导子;Nrf2/ARE;诱导机制查看摘要收藏负责人:赵改名参与人:柳艳霞, 田玮, 孙灵霞, 刘欣, 常亚楠, 毕姗姗金额:75万申请时间:2012学科代码:肉加工(C200203)项目批准号:31271895申请单位:河南农业大学研究类型:基础研究关键词:酱卤禽肉制品;卤煮;火候查看摘要收藏负责人:肖珊参与人:孙浩, 丁燕, 王波, 张敏, 王菲申请单位:大连工业大学研究类型:基础研究关键词:干腌火腿;脂肪酶;基于活性的蛋白质组学;脂肪甘油三酯脂酶查看摘要收藏负责人:陈霞参与人:顾瑞霞, 李华, 吴琳楠, 徐寅, 刘冬金额:21万申请时间:2012学科代码:乳加工(C200204)项目批准号:31201393申请单位:扬州大学研究类型:基础研究关键词:嗜热链球菌;喹诺酮;生理功能;代谢途径查看摘要收藏负责人:王仲妮参与人:刘峰, 刁兆玉, 周武, 马学彬, 王敏, 郭富民, 魏玲, 王迪, 刘馨金额:83万申请时间:2012学科代码:食品添加剂(C200210)项目批准号:31271933申请单位:山东师范大学研究类型:基础研究关键词:植物多酚;生物相容溶致液晶;释放;抗氧化活性;稳定性查看摘要收藏负责人:徐岩参与人:吴群, 唐柯, 杜海, 张荣, 韩业慧, 陈笔, 朱伟岸, 孔宇金额:86万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271921申请单位:江南大学研究类型:基础研究关键词:清香型白酒;土霉异味;微生物群落;代谢机制;固态发酵查看摘要收藏负责人:韩培培参与人:谭之磊, 闫林, 谭宁, 李艳艳, 李清亮, 孙莹申请单位:天津科技大学研究类型:基础研究关键词:发状念珠蓝细菌;光;多糖;蛋白质;代谢物查看摘要收藏负责人:吴敬参与人:赵丽华, 王英丽, 王越男, 李丽杰, 赵一萍, 高雅罕, 张宇宏, 王美霞, 吴苏日古嘎金额:46万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31260391申请单位:内蒙古农业大学研究类型:基础研究关键词:乳酸菌;肠球菌素;基因;表达;酸马奶查看摘要收藏负责人:许正宏参与人:张晓梅, 吴林寰, 许伟, 王宗敏, 邓晓阳, 陶京兰金额:80万申请时间:2012学科代码:食品发酵与酿造(C200207)项目批准号:31271922申请单位:江南大学研究类型:基础研究关键词:镇江香醋;微生物群落;元基因组分析;风味;代谢途径查看摘要收藏负责人:张万刚参与人:高峰, 祝超智, 郭兵, 殷燕, 郭丽媛金额:75万申请时间:2012学科代码:肉加工(C200203)项目批准号:31271899申请单位:南京农业大学研究类型:基础研究关键词:亚硝基化;钙蛋白酶;一氧化氮;猪肉查看摘要收藏负责人:谭晓荣参与人:王付转, 崔柳青, 李翠香, 徐瑞, 王俊峰, 徐苏丽申请单位:河南工业大学研究类型:基础研究关键词:酵母;乙醇胁迫;自噬;发酵;活性氧查看摘要收藏负责人:彭增起参与人:吴菊清, 靳红果, 王蓉蓉, 白云, 万可慧, 张雅玮, 姚瑶, 田锐花, 王复龙金额:78万申请时间:2012学科代码:肉加工(C200203)项目批准号:31271898申请单位:南京农业大学研究类型:基础研究关键词:焦磷酸酶;三聚磷酸酶;多聚磷酸酶;肌肉蛋白;凝胶特性查看摘要收藏负责人:刘静波参与人:王二雷, 张燕, 王莹, 于志鹏, 刘博群, 赵颂宁, 刘丹, 于一丁金额:86万申请时间:2012学科代码:蛋加工(C200205)项目批准号:31271907申请单位:吉林大学研究类型:基础研究关键词:肽;蛋清;结构;完整吸收查看摘要收藏负责人:孟宗参与人:王风艳, 沈琪, 李满香, 王宇, 沈海燕, 陈芳芳金额:23万申请时间:2012学科代码:食品油脂加工(C200201)项目批准号:31201382申请单位:江南大学研究类型:基础研究关键词:植物油;有机凝胶油;形成机制;纳米结构;微观结构查看摘要收藏负责人:刘贺参与人:何余堂, 李秀霞, 朱丹实, 郭晓飞, 李君。
基于代谢组学分析两种萎凋方式对白茶萎凋过程代谢物与品质的影响

接收日期:2021-12-03 接受日期:2021-12-26基金项目:国家自然科学基金项目(31270735);国家重大农技推广服务试点项目(KNJ-151000);福建省教育厅中青年教师教育科研项目(JAT170885);福建省本科高校人文社科研究基地建设项目(闽教科[2019]37号、闽农大金山研[2019]4号)*通信作者。
E-mail:*************;***************.cn基于代谢组学分析两种萎凋方式对白茶萎凋过程代谢物与品质的影响岳文杰1,洪翠云2,赵 峰3,宁 芊1,陈凌华1,叶乃兴4*,陈明杰5*(1. 福建农林大学金山学院,福建 福州 350002;2. 福建省亚热带植物研究所 / 福建省亚热带植物生理生化重点实验室,福建 厦门 361006;3. 福建中医药大学药学院,福建 福州 350122;4. 福建农林大学园艺学院,福建 福州 350002;5. 信阳师范学院生命科学学院 / 河南省茶树生物学重点实验室,河南 信阳 464000)摘 要:基于代谢组学技术,比较室内自然萎凋和空气能萎凋两种萎凋工艺对白茶萎凋过程代谢物与品质的影响。
结果表明,不同代谢物对2种萎凋工艺的响应存在差异,体现三种变化规律:第一类代谢物受萎凋工艺影响小,如咖啡碱、表没食子儿茶素和2'-羟基二氢大豆素等;第二类代谢物含量变化速率在空气能萎凋时加快,但其最终含量受影响小,如表没食子儿茶素没食子酸酯、聚酯型儿茶素A 和木麻黄素等;第三类代谢物则受萎凋时间影响大,如茶没食子素与奎宁酸等。
两种萎凋工艺对白茶品质影响存在差异,自然萎凋下黄酮(醇)糖苷类、儿茶素聚合体等含量更高,这些物质多在萎凋中后期含量提升;空气能萎凋下酚酸、茶氨酸含量更高,这些物质在自然萎凋后期含量大幅下降。
萎凋时长是两种萎凋工艺导致代谢物含量差异的重要因素。
关键词:白茶;自然萎凋;空气能萎凋;代谢物;品质Doi: 10.3969/j.issn.1009-7791.2021.06.002中图分类号:TS272.5 文献标识码:A 文章编号:1009-7791(2021)06-0430-09Effects of Two Withering Methods on Metabolites and Quality during Withering Process of White Tea Based on Metabolomics TechnologyYUE Wen-jie 1, HONG Cui-yun 2, ZHAO Feng 3, NING Qian 1, CHEN Ling-hua 1, YE Nai-xing 4*, CHEN Ming-jie 5*(1. Jinshan College, Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian China; 2. Fujian Institute of Subtropical Botany / Fujian Key Laboratory of Subtropical Plant Physiology and Biochemistry, Xiamen 361006, Fujian China; 3. School of Pharmacy, Fujian University of Traditional Chinese Medicine, Fuzhou 350122, Fujian China; 4. College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian China; 5. College of Life Sciences, Xinyang Normal University / Henan Key Laboratory of Tea PlantBiology, Xinyang 464000, Henan China)Abstract: Based on metabolomics technology, the effects of different withering technologies on the metabolites and quality of white tea withering were compared. The results showed that the response of different metabolites to the two withering processes was different, reflecting three kinds of changes: the first type of metabolites was less affected by the withering process, such as caffeine, EGC and 2'-hydroxydihydrodaidzein; the second type of metabolites showed accelerated conversion rate under air conditioner withering, but had little effect on the final dry tea content, such as EGCG, theasinensin A, and strictinin; and the third type of metabolites was greatly affected by the withering time, such as theogallin and quinic acid. There were different effects on the quality of white tea by two withering processes. Under natural withering the contents of flavonoids (alcohols), glycosides, catechin polymers, etc. were higher, and the contents of these substances increased in the middle and late stages of withering;2021,50(6): 430~438.Subtropical Plant Science第6期 岳文杰等:基于代谢组学分析两种萎凋方式对白茶萎凋过程代谢物与品质的影响﹒431﹒431under air conditioner withering the content of phenolic acid and theanine was higher, and these substances decreased sharply in the latter stage of natural withering. The withering time was an important factor for the difference in the content of metabolites between the two withering processes.Key words: white tea; natural withering; air conditioner withering; metabolites; quality白茶是中国传统六大茶类之一,主产于福建省宁德市和南平市,以其外形满披白毫,汤色浅黄明亮,滋味鲜醇清甜有毫香等独特品质广受消费者喜爱[1—2]。
单磷酸腺苷激活的蛋白激酶(AMPK)调控机制的研究进展

单磷酸腺苷激活的蛋白激酶(AMPK)调控机制的研究进展林志忠;叶志云;林圣彩;林舒勇【摘要】代谢是细胞和有机体最基本、最重要的活动之一.细胞和有机体通过感应系统实时监测代谢过程中的物质和能量状态,并不断地通过错综复杂的代谢调控途径来维持其稳态.代谢调控一旦出现紊乱,机体代谢就会发生异变,从而导致如糖尿病、肥胖、心血管疾病乃至肿瘤等多种人类重大疾病的发生,并影响发育、生长、繁殖、衰老等生命过程.单磷酸腺苷激活的蛋白激酶(AMPK)作为在代谢调控中起核心作用的激酶,自其被发现以来一直是研究的热点.对AMPK调控机制的深入研究为揭示代谢性疾病的发生和发展机制、探索其治疗和预防的策略提供了重要的新思路.围绕近年来国内外AMPK研究领域取得的进展,重点阐述AMPK在体内的激活机制以及本课题组在该领域中的一系列重要成果.【期刊名称】《厦门大学学报(自然科学版)》【年(卷),期】2018(057)002【总页数】8页(P149-156)【关键词】能量稳态;单磷酸腺苷激活的蛋白激酶;葡萄糖感应【作者】林志忠;叶志云;林圣彩;林舒勇【作者单位】厦门大学生命科学学院,福建厦门361102;厦门大学生命科学学院,福建厦门361102;厦门大学生命科学学院,福建厦门361102;厦门大学生命科学学院,福建厦门361102【正文语种】中文【中图分类】Q591三磷酸腺苷(ATP)是几乎所有细胞和有机体中最重要的能量载体,ATP水解成二磷酸腺苷(ADP)所释放出的能量直接用于绝大多数生命活动,当能量进一步缺乏时ADP 可再转变为单磷酸腺苷(AMP)以提供更多的能量;而当能量充足时,细胞又可将AMP 和ADP 重新转化成ATP从而将能量存储起来.在正常情况下,大部分真核细胞中的ATP浓度始终维持在一个非常稳定的水平,因此细胞中必然存在一个可以感知ATP、ADP和AMP水平变化的机制.AMP激活的蛋白激酶(AMP-activated protein kinase,AMPK)目前已被公认是有机体和细胞内感应能量水平并调节代谢稳态的最重要分子.早期的研究发现,向大鼠的肝脏匀浆中加入AMP或ADP,可以有效地抑制2种脂肪酸合成过程中的关键酶——3-羟基-3-甲基戊二酸单酰辅酶A还原酶(HMG-CoA reductase)和乙酰辅酶A羧化酶(acetyl-CoA carboxy-lase,ACC)的活力从而强烈地抑制肝脏的脂肪合成[1-3].进一步的研究表明,这一抑制作用实际上是由AMP引起的,且依赖于某个未知蛋白激酶的催化反应[4].为了找到这个未知的蛋白激酶,苏格兰邓迪大学的Hardie实验室对大鼠肝脏匀浆进行了分离和纯化,并对能够响应AMP抑制ACC 的组分进行了近4年的细致分析,终于在1989年找到了一个由分子质量分别为63,38和35 ku的3个亚基组成的蛋白,并根据该蛋白能被AMP激活的特性将其命名为AMPK[5].后来的研究表明,当细胞内处于能量缺乏状态时,AMPK被激活,进而直接导致了多条信号通路的抑制或激活并引起一系列的生理变化(图1),包括:促进脂肪细胞中甘油三酯的水解[6],促进肝脏对血液中脂肪酸的摄取和氧化[7],抑制脂肪酸和胆固醇的合成以及甘油三酯的形成[8-9],促进肌肉中脂类氧化以及葡萄糖的摄取和分解[10-11],抑制胰岛β细胞分泌胰岛素以及糖原的合成[12],抑制蛋白质的合成并促进自噬作用[13-15]和酮体的生成[16]等.其中,抑制脂肪酸合成并促进其氧化被认为是AMPK最经典的功能,主要是通过磷酸化并抑制其底物ACC的活力实现的,而ACC又是脂肪酸合成的关键酶[10].这些作用的结果是增加体内的产能代谢,减少体内的耗能代谢,稳定体内能量水平,维持能量稳态,从而保证细胞的正常生命活动.由此可见,AMPK是机体内最重要的能量稳态调节者,它的功能和人类的生存与健康密切相关.图1 AMPK参与调控的许多重要细胞生物学过程Fig.1 Multiple important cellular biological processes regulated by AMPK 1 AMPK的结构AMPK是由α、β和γ 3个亚基构成的一个异源三聚体.α亚基的N 端是AMPK 催化活性的核心区域,包含一个保守的丝氨酸/苏氨酸激酶结构域(kinase domain,KD)、C端的β亚基相互作用结构域(β-subunit interacting domain,β-SID)和与C端紧邻的自抑制结构域(autoinhibitory domain,AID),其中AID的作用是通过结合KD来抑制α亚基的激酶活性[17-18];β亚基包含一个中等碳水化合物的结合模块(mid-molecule carbohydrate-binding module,CBM)和一段位于C末端的连接α和γ亚基的结合序列(α,γ subunit-binding sequence,SBS);γ亚基含有4个参与结合ATP、AMP等的胱硫醚β-合成酶(cystathionine-β-synthase,CBS)结构域.当AMP结合到γ亚基上的CBS结构域时会引起该亚基发生变构,随后影响AID的构象,使α亚基上的苏氨酸172(T172)位点暴露出来,导致该位点被AMPK的上游激酶磷酸化[18-22];同时由于AMP的结合导致的构象变化还可阻止该位点的去磷酸化,从而使AMPK保持较高的激活水平.相反地,ATP的结合则能够抑制这一过程[23-24].在哺乳动物细胞中,AMPK的α和β亚基各包含2种不同的亚型(α1、α2和β1、β2),γ亚基包含3种不同的亚型(γ1、γ2和γ3),这些亚型均由不同的基因编码表达,它们可以组合成至少12种不同的AMPK三聚体[25-26].研究发现这些亚型的表达具有组织特异性,γ2在人体的心肌细胞中高度表达,而γ3则集中表达于快速抽动的骨骼肌细胞中;此外,在不同物种之间这些亚型的表达差异很大,如γ2在人的心肌细胞中高表达,但在小鼠的心肌细胞中表达量却很低,而在小鼠的脑组织和脂肪组织中又是高表达的[27-28].因此,AMPK不同亚型之间的关联性及其在不同物种、不同组织细胞中所发挥的功能还有待进一步的深入研究.2 AMPK的调控机制如上所述,AMPK的激活依赖于其T172位点的磷酸化,目前已发现2种主要的上游激酶,分别为肝脏激酶B1(liver kinase B1,LKB1)和钙离子/钙调蛋白依赖蛋白激酶β(Ca2+/calmodulin-activated protein kinase β,CaMKKβ)[29-32].LKB1是一个肿瘤抑制因子,自身处于持续激活的状态,当组织缺氧、缺血或运动导致细胞内能量缺乏时,细胞内AMP/ATP比值上升,AMP与AMPK结合时后者的构象发生改变,使得AMPK的T172位点暴露出来,从而使LKB1能够对该位点进行磷酸化,并可使其活性上升100倍以上[29];而CaMKKβ则需要在细胞内钙离子浓度升高时才能被激活,并进而磷酸化AMPK[30,33].另外,早期的研究表明ADP与AMPK结合亦可引发与AMP相同的效应[34-35];但是较新的一项研究证明,只有当AMP与AMPK结合后LKB1才对其T172位点进行磷酸化,而ADP的结合只参与了抑制该位点的去磷酸化[22].然而,这些发现大都基于体外的生化实验,AMPK在体内被激活的机制,以及AMP对AMPK磷酸化的具体作用长期以来一直不为人知.2013年,本课题组率先揭开了AMPK在体内被激活的“路线图”,发现体轴发育抑制因子(axis inhibitor,AXIN)是LKB1激活AMPK所必需的,其中AXIN是一个多功能的构架蛋白,参与调节许多信号通路,如Wnt、JNK和p53等,对细胞生长起重要的调控作用[36-40].当细胞在饥饿状态下,由于AMP结合至AMPK后增强了后者对AXIN的亲和力,而AXIN自身与LKB1有很强的相互作用,所以此时的AXIN起到了一个桥梁的作用,将AMPK和其上游激酶LKB1连接在一起,三者形成一个复合体,促进了LKB1对AMPK的磷酸化激活,使得AMPK的活性升高[41](图2).以上研究成果完善了AMPK活性调控过程中的关键一环,揭示了AMP促进AMPK磷酸化的分子机制.图2 AXIN激活AMPK的机制[41]Fig.2 Mechanism of AMPK activation by AXIN[41]引自参考文献[42],经修改.图3 v-ATPase-Ragulator复合体调控AMPK和mTORC1活化切换的机制Fig.3 Mechanism of switch between AMPK and mTORC1 activation regulat ed by v-ATPase-Ragulator complex随后,本课题组又通过酵母双杂交的方式找到了和AXIN相互作用的溶酶体相关膜蛋白LAMTOR1,并且证明该蛋白也是能量缺乏情况下AMPK激活所必需的[42].LAMTOR1是一个锚定在溶酶体膜表面的蛋白复合体Ragulator的重要成员,在缺乏LAMTOR1的小鼠或细胞中,饥饿信号不能引起AMPK的激活.在能量充足的情况下,Ragulator通过其自身的鸟苷酸交换因子(guanine nucleotide exchange factor,GEF)激活鸟苷酸酶Rag,从而使哺乳动物雷帕霉素靶蛋白复合体1(mammalian target of rapamycin complex 1,mTORC1)迁移到溶酶体上并被激活,这时合成代谢途径开启(图3(a));而在能量不足的情况下,溶酶体上的能量感知者空泡型ATP酶(vacuolar-type ATPase,v-ATPase)发生变构,通过与其相互作用的Ragulator复合体共同促进AXIN-LKB1迁移到溶酶体上并与Ragulator结合,最后在此激活AMPK,同时AXIN抑制了Ragulator的GEF活力,进一步促进 mTORC1的解离,从而关闭合成代谢途径,开启分解代谢途径(图3(b)).综上,本课题组发现细胞内由于葡萄糖缺乏所引起的AMPK激活是在溶酶体膜上进行的,并且揭示了该过程所涉及的蛋白质机器.这一发现不仅揭示了AMPK在体内生理状态下被激活的方式与过程,也解析了细胞在营养物质缺乏的情况下调节合成代谢与分解代谢的切换机制.此外,除了能量缺乏以外,AMPK还可以响应多种药物的刺激而被激活[43].根据作用机制的不同可以将这些药物分为以下3大类:1) 线粒体复合体抑制剂,通过抑制呼吸链来提高细胞内AMP/ADP水平从而激活AMPK,常见的如二甲双胍(metformin)[44]、黄连素[45]等;2) AMP类似物的前体化合物,如5-氨基-4-甲酰胺咪唑核糖核苷酸(5-aminoimidazole-4-carboxamide ribonucleoside,AICAR)[46]和近几年被发现的一种膦酸类化合物5-(5-hydroxyl-isoxazol-3-yl)-furan-2-phosphonic acid[47],这些化合物可在细胞内转变为AMP类似物,从而直接提高AMP水平使AMPK被激活;3)AMPK激活剂,可直接作用于AMPK使之构象发生变化并随后被激活,包括最早发现的被命名为A769662的化合物[48]以及最近发现的991[49]和MT-63-78[50].在上述这些药物中,二甲双胍是一个重要的代表性药物,已被广泛应用于治疗Ⅱ型糖尿病,它能通过激活AMPK来缓解脂肪肝并降低并发症风险[51].为了进一步探究Ragulator-AXIN在药理情况下对AMPK激活过程的调控作用,本课题组对缺失了LAMTOR1的小鼠胚胎成纤维细胞(mouse embryonic fibroblast,MEF)进行二甲双胍处理,发现二甲双胍不能在这些细胞中激活AMPK;同样地,对于LAMTOR1肝脏特异性敲除的小鼠,二甲双胍也不能在其肝脏中激活AMPK;进而在缺失了AXIN的细胞和小鼠肝脏中检测了二甲双胍对AMPK的激活作用,发现AXIN是二甲双胍激活AMPK所必需的,且二甲双胍激活AMPK也是通过溶酶体途径进行的,并依赖于上述溶酶体上的v-ATPase-Ragulator感应复合体[52].这一发现不仅解决了二甲双胍如何激活AMPK的重要问题,还为人们了解二甲双胍降低糖尿病人的肿瘤发生机制打开了窗口.3 AMPK和葡萄糖感应引自参考文献[59],经修改.图4 细胞感知葡萄糖水平的分子机制Fig.4 Molecular mechanism of glucose sensing in cell葡萄糖是生物界最基本、最主要的供能物质,故其水平和机体的能量代谢状态本身就具有紧密的联系;此外,葡萄糖代谢的中间产物又是几乎所有合成代谢途径最重要的原材料来源.可见,感受葡萄糖的水平并随时作出相应的响应是生物体维持代谢平衡的基本功能.AMPK能够响应机体葡萄糖水平的变化,并在其水平下降的时候被激活,该机制在各种表达有AMPK的真核生物中高度保守,从酵母到哺乳动物中都已被发现[53],是进化上少有的经典表型.饥饿、运动等生理过程都会导致整体或某个器官的葡萄糖水平变化.在葡萄糖水平下降时,激活的AMPK能够通过一系列方式维持代谢平衡和机体的正常生理功能.例如:饥饿能够引起血液和组织液中葡萄糖水平的下降,AMPK在此时被激活,进而促进肝脏等组织中脂肪酸的β-氧化,使之转换为利用脂肪酸[7];与此同时,AMPK还能促进肝脏生成酮体,通过血液供给脑部等不能利用脂肪酸的器官,以满足这些组织的能量需求.许多生物的生命活动有昼夜节律(circadian rhythms),这种周期性也引起了机体内葡萄糖水平的周期性变化[54],而AMPK能够响应这种变化并维持机体能量稳态[55].近年来,人们利用AMPK和葡萄糖之间的密切联系开发了许多药物,来模拟葡萄糖缺失的情况以激活AMPK,如无法被代谢的葡萄糖类似物2-DG和5-TG等[56-57],这些药物能够通过阻断葡萄糖代谢达到激活AMPK的目的,并已应用于某些肿瘤的前期实验中[58].因此,葡萄糖水平的感应对于AMPK的激活至关重要,是打开代谢稳态维持机制之门的“钥匙”.然而,AMPK如何感应机体的葡萄糖水平并被激活的机制长期以来并不清楚,甚至存在重大的误区.由于AMP在AMPK激活过程中的重要而经典的地位,它一直以来被认为是AMPK唯一的激活信号;同时,传统的理论把葡萄糖看作一种“能量信号”,它的下降将引起细胞内ATP水平的下降和AMP水平的上升,后者作为激活剂直接激活AMPK.但实际上,并没有一种生理状态能够验证以上理论.本课题组在对AMPK激活机制的深入研究中发现,无论是在葡萄糖缺乏的细胞培养条件下,还是在饥饿的低血糖动物体内,都不能检测到AMP水平的上升,但与此同时AMPK却处在被激活的状态,这说明葡萄糖水平的降低与“能量缺失”无关,即与AMP水平无关,而是通过一种新的信号和一条独立于能量代谢的通路来调控AMPK 的激活.本课题组之前已经发现了一条响应葡萄糖缺失并激活AMPK的溶酶体途径[42],揭示了内源AMPK的激活过程.在此基础上,进一步研究AMPK感受和响应葡萄糖水平而被激活的机制,成功地鉴定出传递葡萄糖水平的“信使”——果糖1,6-二磷酸(fructose-1,6-bisphosphate,FBP),同时找到了感应这一“信使”的“感受器”——参与糖酵解通路的醛缩酶(aldolase),依靠这一套完整的信号系统,葡萄糖水平就能够作为一种具体的信号调节AMPK的激活乃至整个机体的代谢稳态[59].具体地说,当葡萄糖水平下降时,葡萄糖代谢物FBP的水平也相应地下降,该过程进一步被醛缩酶所感应,后者将启动激活AMPK的溶酶体途径进而介导AMPK的激活(图4).这一发现改变了人们一直以来认为的AMPK活性只被代表低能量的AMP调节的观点,是对传统AMPK激活机制的一次重新认识;并据此提出,葡萄糖水平是机体的一种“状态信号”,通过调节AMPK的激活从而调节机体代谢的总平衡.这对全面了解细胞代谢稳态维持的分子机制、糖代谢和相关应激反应乃至生理、病理进程有重要意义,且有助于为研发治疗人类重大疾病的药物提供全新、有效的靶点.4 AMPK和疾病糖尿病是一种糖脂代谢稳态失调、紊乱的疾病.已有研究发现,在肥胖小鼠和Ⅱ型糖尿病病人的外周组织中,AMPK的活性明显被抑制[60-61].而AMPK的激活能够促进肌肉中的葡萄糖转运蛋白GLUT4转移到细胞膜上[62-63],增加肌肉对血液中葡萄糖的吸收和分解代谢,从而降低血糖[64].在肝脏中,当ATP缺乏引起AMPK 的激活后,AMPK可以通过磷酸化转录辅激活因子CRTC2(也称TORC2)促进其出核,或者通过磷酸化去乙酰化酶HDAC4/5/7促进转录因子FOXO1出核,这两种方式的结果都是抑制肝脏的糖异生途径,从而降低血糖[65-67].除了代谢性疾病,许多肿瘤的发生和代谢紊乱也有着密切的联系.肿瘤以及肿瘤微环境与正常组织相比的一个重要特征就是代谢过程的不同,而代谢紊乱进一步促进了肿瘤的发生和发展[68-69].由于肿瘤组织的快速增殖消耗大量营养,肿瘤微环境是一个营养匮乏的环境,这将引起肿瘤组织中AMPK的激活[70].在细胞水平上,已经有大量的研究表明AMPK的激活可以通过抑制mTORC1复合体而抑制肿瘤细胞的合成代谢,从而阻止其增殖[71];还可以通过促进p53的活性引起肿瘤细胞的生长阻滞和凋亡,从而抑制肿瘤细胞的生长[72-74].因此,在多种肿瘤组织,如黑色素瘤、乳腺癌、结肠癌和肺癌组织中,AMPK的表达或者活性被强烈抑制,进一步打破了这些组织中原有的合成代谢和分解代谢的平衡,加剧了肿瘤的发展[70,75-76].肿瘤微环境对免疫细胞中AMPK的激活则反过来阻止了促炎症免疫细胞的分化和发育以及相关免疫因子的释放,使之处于休眠状态不能被激活并发挥效应,从而抑制了免疫反应对肿瘤组织的识别和杀灭,导致肿瘤的生长更加不受控制[77].5 结语与展望作为能量代谢调节的核心分子,AMPK一直是细胞代谢领域研究的重点.虽然近年来人们已经在该领域获得了一系列的新发现,然而对AMPK精细的调控机制还需要更深层次的研究.目前已知AMPK的激活存在以下两条截然不同的途径:1) 依赖于AMP(AMP-dependent)的经典途径;2) 不依赖于AMP(AMP-independent)而通过糖酵解通路中代谢酶aldolase的途径.那么,进一步需要回答的重要问题就是细胞如何根据生理状况对上述两种途径做出选择.另外,本课题组的研究发现细胞中存在两个激活AMPK的位置——溶酶体和细胞质.那么,在这两个位置上激活的AMPK是否有不同的底物及其生理意义也是后续研究中有待解决的问题,这不仅能够帮助人们更深刻地理解AMPK以及代谢稳态的调控机制,还有助于理解和操控相关的重要生理过程和病理过程,最终为战胜威胁人类健康的重大疾病贡献力量.过去的十几年来,人们也一直在研发以AMPK为靶点用于治疗代谢性疾病的药物,并已筛选出了一些潜在的AMPK激活剂,如国际制药巨头Pfizer公司筛选出的名为991的化合物.然而,这些激活剂虽然在体外实验中可以有效地激活AMPK,但其面临的真正挑战是能否在动物体内以及人体内也发挥同样的功能.随着人们对AMPK激活机制的不断深入研究与解析,或许在不久的将来有望见证以全新的作用机制激活AMPK的药物问世.[1] GIL G,SITGES M,BOVÉ J,et al.Phosphorylation-dephosphorylation of rat liver 3-hydroxy 3-methylglutaryl coenzyme A reductase associated with changes in activity[J] .FEBS Lett,1980,110(2):195-199.[2] HARWOOD H J,JR,BRANDT K G,RODWELL V W.Allosteric activation of rat liver cytosolic 3-hydroxy-3-methylglutaryl coenzyme A reductase kinase by nucleoside diphosphates[J ].J Biol Chem,1984,259(5):2810-2815.[3]CARLSON C A,KIM K H.Regulation of hepatic acetyl coenzyme A carboxylas e by phosphorylation and dephosphorylation[J].J Biol Chem,1973,248(1):37 8-380.[4]CARLING D,ZAMMIT V A,HARDIE D G.A common bicyclic protein kinase ca scade inactivates the regulatory enzymes of fatty acid and cholesterol bios ynthesis[J].FEBS Lett,1987,223(2):217-222.[5]CARLING D,CLARKE P R,ZAMMIT V A,et al.Purification and characterization of the AMP-activated protein kinase.Copurification of acetyl-CoA carboxylase kinase and 3-hydroxy-3-methylglutaryl-CoA reductase kinase activities[J].Eur J Biochem,1989,186(1/2):129-136. [6] GAUTHIER M S,MIYOSHI H,SOUZA S C,et al.AMP-activated protein kinase is activated as a consequence of lipolysis in the ad ipocyte:potential mechanism and physiological relevance[J].J Biol Chem,20 08,83(24):16514-16524.[7] SAVAGE D B,CHOI C S,SAMUEL V T,et al.Reversal of diet-induced hepatic steatosis and hepatic insulin resis-tance by antisense oligonucleotide inhibitors of acetyl-CoA carboxylases 1 and 2[J].J Clin Invest,2006,116(3):817-824.[8]SMITH B K,MARCINKO K,DESJARDINS E M,et al.Treatment of nonalcoholic fatty liver disease:role of AMPK[J].Am J Physiol Endocrinol Metab,2016,311( 4):E730-E740.[9]LI Y,XU S,MIHAYLOVA M M,et al.AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resis-tant mice[J].Cell Metab,2011,13(4):376-388.[10]MERRILL G F,KURTH E J,HARDIE D G,et al.AICA riboside increases AMP-activated protein kinase,fatty acid oxidation,and glucose uptake in rat mus cle[J].Am J Physiol,1997,273(1):E1107-E1112.[11] O′NEILL H M,MAARBJERG S J,CRANE J D,et al.AMP-activated protein kinase (AMPK) β1β2 muscle null mice reveal an essential r ole for AMPK in maintaining mitochondrial content and glucose uptake du ring exercise[J].Proc Natl AcadSci USA,2011,108(38):16092-16097.[12] SALT I P,JOHNSON G,ASHCROFT S J,et al.AMP-activated protein kinase is activated by low glucose in cell lines derived fro m pancreatic β cells,and may regulate insulin release[J].Biochem J,1998,335 (3):533-539.[13]INOKI K,ZHU T,GUAN K L.TSC2 mediates cellular energy response to contr ol cell growth and survival[J].Cell,2003,115(5):577-590.[14]GWINN D M,SHACKELFORD D B,EGAN D F,et al.AMPK phosphorylation of r aptor mediates a metabolic checkpoint[J].Mol Cell,2008,30(2):214-226. [15]EGAN D F,SHACKELFORD D B,MIHAYLOVA M M,et al.Phosphorylation of U LK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy[J].Science, 2011,331(6016):456-461.[16] WINDER W W,HARDIE D G.AMP-activated protein kinase,a metabolic master switch:possible roles in type 2 diabetes[J].Am J Physiol,1999,277(1):E1-E10.[17] PANG T,XIONG B,LI J Y,et al.Conserved α-helix acts as autoinhibitory sequence in AMP-activated protein kinase α subunits[J].J Biol Chem,2007,282(1):495-506. [18]CHEN L,JIAO Z H,ZHENG L S,et al.Structural insight into the autoinhibition mechanism of AMP-activated protein kinase[J].Nature,2009,459(7250):1146-1149.[19]XIAO B,HEATH R,SAIU P,et al.Structural basis for AMP binding to mammali an AMP-activated protein kinase[J].Nature,2007,449(7161):496-500. [20]XIN F J,WANG J,ZHAO R Q,et al.Coordinated regulation of AMPK activity b y multiple elements in the α-subunit[J].Cell Res,2013,23(10):1237-1240. [21] OAKHILL J S,CHEN Z P,SCOTT J W,et al.β-Subunit myristoylation is the gatekeeper for initiating metabolic stress sen sing by AMP-activated protein kinase (AMPK)[J].Proc Natl Acad Sci USA,2010,107(45):19 237-19241.[22]GOWANS G J,HAWLEY S A,ROSS F A,et al.AMP is a true physiological regul ator of AMP-activated protein kinase by both allosteric activation and enhancing net ph osphorylation[J].Cell Metab,2013,18(4):556-566.[23] CARLING D,THORNTON C,WOODS A,et al.AMP-activated protein kinase:new regulation,new roles?[J].Biochem J,2012,445(1 ):11-27.[24]OAKHILL J S,SCOTT J W,KEMP B E.AMPK functions as an adenylate charge-regulated protein kinase[J].Trends in Endocrinology and Metabolism,2012, 23(3):125-132.[25] CARLING D,MAYER F V,SANDERS M J,et al.AMP-activated protein kinase:nature′s energy sensor[J].Nat Chem Biol,2011,7(8): 512-518.[26]MILAN D,JEON J T,LOOFT C,et al.A mutation in PRKAG3 associated with ex cess glycogen content in pig skeletal muscle[J].Science,2000,288(5469):124 8-1251.[27] CHEUNG P C F,SALT I P,DAVIES S P,et al.Characterization of AMP-activated protein kinase g-subunit isoforms and their role in AMP binding[J].Biochem J,2000,346(3):65 9-669.[28]HAWLEY S A,BOUDEAU J,REID J L,et plexes between the LKB1 tumor suppressor,STRAD α/β and MO25 α/β are upstream kinases in the AMP-activated protein kinase cascade[J].J Biol,2003,2(4):28.[29] HAWLEY S A,PAN D A,MUSTARD K J,et al.Calmo-dulin-dependent protein kinase kinase-β is an alternative upstream kinase for AMP-activated protein kinase[J].Cell Metab,2005,2(1):9-19.[30] WOODS A,DICKERSON K,HEATH R,et al.Ca2+/calmodulin-dependent protein kinase kinase-β acts upstream of AMP-activated protein kinase in mammalian cells[J].Cell Metab,2005,2(1):21-33.[31]WOODS A,JOHNSTONE S R,DICKERSON K,et al.LKB1 is the upstream kinas e in the AMP-activated protein kinase cascade[J].Curr Biol,2003,13(22):2004-2008. [32] SUTER M,RIEK U,TUERK R,et al.Dissecting the role of 5′-AMP for allosteric stimulation,activation,and deactivation of AMP-activated protein kinase[J].J Biol Chem,2006,281(43):32207-32216. [33]HURLEY R L,ANDERSON K A,FRANZONE J M,et al.The Ca2+/calmoldulin-dependent protein kinase kinases are AMP-activated protein kinase kinases[J].J Biol Chem,2005,280(32):29060-29066.[34]XIAO B,SANDERS M J,UNDERWOOD E,et al.Structure of mammalian AMPK and its regulation by ADP[J].Nature,2011,472(7342):230-233.[35]OAKHILL J S,STEEL R,CHEN Z P,et al.AMPK is a direct adenylate charge-regulated protein kinase[J].Science,2011,332(6036):1433-1440.ZENG L,FAGOTTO F,ZHANG T,et al.The mouse Fused locus encodes Axin,an inhibitor of the Wnt signaling pathway that regulates embryonic axis form ation[J].Cell,1997,90(1):181-192.[37]RUI Y,XU Z,LIN S,et al.Axin stimulates p53 functions by activation of HIPK2 kinase through multimeric complex formation[J].EMBO J,2004,23(23):4583-4594.[38]LI Q,WANG X,WU X,et al.Daxx cooperates with the Axin/HIPK2/p53 comple x to induce cell death[J].Cancer Res,2007,67(1):66-74.[39]LI Q,LIN S,WANG X,et al.Axin determines cell fate by controlling the p53 ac tivation threshold after DNA damage[J].Nat Cell Biol,2009,11(9):1128-1134.[40] RUI Y,XU Z,XIONG B,et al.A β-catenin-independent dorsalization pathway activated by Axin/JNK signaling and an tagonized by aida[J].Dev Cell,2007,13(2):268-282.[41] ZHANG Y L,GUO H,ZHANG C S,et al.AMP as a low-energy charge signal autonomously initiates assembly of AXIN-AMPK-LKB1 complex for AMPK activation[J].Cell Metab,2013,18(4):546-555. [42] ZHANG C S,JIANG B,LI M,et al.The lysosomal v-ATPase-Ragulator complex is a common activator for AMPK and mTORC1,acting as a switch between cata-bolism and anabolism[J].Cell Metab,2014,20(3):526-540.HARDIE D G.AMPK:sensing energy while talking to other signaling pathway s[J].Cell Metab,2014,20(6):939-952.[44]HAWLEY S A,FULLERTON M D,ROSS F A,et al.The ancient drug salicylate dir ectly activates AMP-activated protein kinase[J].Science,2012,336(6083):918-922.[45] HARDIE D G.AMP-activated protein kinase:maintaining energy homeostasisat the cellular and whole-body levels[J].Annu Rev Nutr,2014,34(34):31-55.[46]GADALLA A E,PEARSON T,CURRIE A J,et al.AICA riboside both activates AM P activated protein kinase and competes with adenosine for the nucleoside transporter in the CA1 region of the rat hippocampus[J].J Neurochem,200 4,88(5):1272-1282.[47] GO′MEZ-GALENO J E,DANG Q,NGUYEN T H,et al.A potent and selective AMPK activ ator that inhibits de novo lipogenesis[J].ACS Med Chem Lett,2010,1(9):478-482.[48]COOL B,ZINKER B,CHIOU W,et al.Identification and characterization of a s mall molecule AMPK activator that treats key components of type 2 diabet es and the metabolic syndrome[J].Cell Metab,2006,3(6):403-416.[49]XIAO B,SANDERS M J,CARMENA D,et al.Structural basis of AMPK regulatio n by small molecule activators[J].Nat Commun,2013,4(1):3017.[50]ZADRA G,PHOTOPOULOS C,TYEKUCHEVA S,et al.A novel direct activator of AMPK inhibits prostate cancer growth by blocking lipogenesis[J].EMBO M ol Med,2014,6(4):519-538.[51]FORETZ M,GUIGAS B,BERTRAND L,et al.Metformin:from mechanisms of act ion to therapies[J].Cell Metab,2014,20(6):953-66.[52]ZHANG C S,LI M,MA T,et al.Metformin activates AMPK through the lysoso mal pathway[J].Cell Metab,2016,24(4):521-522.[53] HARDIE D G,CARLING D.The AMP-activated protein kinase:fuel gauge of the mammalian cell?[J].Eur J Bioche m,1997,246(2):259-273.[54]MARCHEVA B,RAMSEY K M,BUHR E D,et al.Disruption of the clock compon ents CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes[J].Nature, 2010,466(7306):627-631.[55]JORDAN S D,LAMIA K A.AMPK at the crossroads of circadian clocks and m etabolism[J].Mol Cell Endocrinol,2013,366(2):163-169.[56] CRAMER F B,WOODWARD G E.2-Desoxy-D-glucose as an antagonist of glucose in yeast fermentation[J].Journal of theFranklin Institute,1952,253(4):354-360.[57] ZHANG D,LI J,WANG F,et al.2-Deoxy-D-glucose targeting of glucose metabolism in cancer cells as a potential ther apy[J].Cancer Lett,2014,355(2):176-183.[58]ROWE I,CHIARAVALLI M,MANNELLA V,et al.Defective glucose metabolism i n polycystic kidney disease identifies a new therapeutic strategy[J].Nat Me d,2013,19(4):488-493.[59] ZHANG C S,HAWLEY S A,ZONG Y,et al.Fructose-1,6-bisphosphate and aldolase mediate glucose sensing by AMPK[J].Nature,20 17,548(7665):112-116.[60]VIOLLET B,HORMAN S,LECLERC J,et al.AMPK inhibition in health and diseas e[J].Crit Rev Biochem Mol Biol,2010,45(4):276-295.[61]WU Y,SONG P,XU J,et al.Activation of protein phosphatase 2A by palmitate inhibits AMP-activated protein kinase[J].J Biol Chem,2007,282(13):9777-9788.[62] TREEBAK J T,GLUND S,DESHMUKH A,et al.AMPK-mediated AS160 phosphorylation in skeletal muscle is dependent on AMP K catalytic and regulatory subunits[J].Diabetes,2006,55(7):2051-2058. [63] MCGEE S L,VAN DENDEREN B J,HOWLETT K F,et al.AMP-activated protein kinase regulates GLUT4 transcription by phosphorylating histone deacetylase 5[J].Diabetes,2008,57(4):860-867.[64] HUANG S,CZECH M P.The GLUT4 glucose transpor-ter[J].Cell Metab,2007,5(4):237-252.[65] KOO S H,FLECHNER L,QI L,et al.The CREB co-activator TORC2 is a key regulator of fasting glucose metabolism[J].Nature, 2005,437(7062):1109-1111.[66] MIHAYLOVA M M,VASQUEZ D S,RAVNSKJAER K,et al.Class Ⅱa histone deacetylases are hormone-activated regulators of FOXO and mammalian glucose homeostasis[J].Cell, 2011,145(4):607-621.[67] WANG B,MOYA N,NIESSEN S,et al.A hormone-dependent module regulating energy balance[J].Cell,2011,145(4):596-606.[68]PAVLOVA N N,THOMPSON C B.The emerging hallmarks of cancer metabol ism[J].Cell Metab,2016,23(1):27-47.[69]CAIRNS R A,HARRIS I S,MAK T W.Regulation of cancer cell metabolism[J].N at Rev Cancer,2011,11(2):85-95.[70]INOKI K,KIM J,GUAN K L.AMPK and mTOR in cellular energy homeostasis a nd drug targets[J].Annu Rev Pharmacol Toxicol,2012,52(1):381-400. [71]BUDANOV A V,KARIN M.p53 target genes sestrin1 and sestrin2 connect ge notoxic stress and mTOR signaling[J].Cell,2008,134(3):451-460.[72]ADAMOVICH Y,ADLER J,MELTSER V,et al.AMPK couples p73 with p53 in cel l fate decision[J].Cell Death Differ,2014,21(9):1451-1459.[73] JONES R G,PLAS D R,KUBEK S,et al.AMP-activated protein kinase induces a p53-dependent metabolic checkpoint[J].Mol Cell,2005,18(3):283-293.[74] SHACKELFORD D B,SHAW R J.The LKB1-AMPK pathway:metabolism and growth control in tumour suppression[J].N at Rev Cancer,2009,9(8):563-575.[75]PINEDA C T,RAMANATHAN S,FON TACER K,et al.Degradation of AMPK bya cancer-specific ubiquitin ligase[J].Cell,2015,160(4):715-728.[76] VILA I K,YAO Y,KIM G,et al.A UBE2O-AMPKα2 axis that promotes tumor initiation and progression offers opport unities for therapy[J].Cancer Cell,2017,31(2):208-224.[77]O′NEILL L A,HARDIE D G.Metabolism of inflammation limited by AMPK and pseudo-starvation[J].Nature,2013,493(7432):346-355.。
EMS_诱变创制水稻抗乙酰辅酶A_羧化酶抑制剂类除草剂种质

江苏农业学报(JiangsuJ.ofAgr.Sci.)ꎬ2023ꎬ39(2):305 ̄312http://jsnyxb.jaas.ac.cn江㊀群ꎬ凌溪铁ꎬ唐兆成ꎬ等.EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质[J].江苏农业学报ꎬ2023ꎬ39(2):305 ̄312.doi:10.3969/j.issn.1000 ̄4440.2023.02.001EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质江㊀群1ꎬ㊀凌溪铁2ꎬ㊀唐兆成2ꎬ㊀周珍珍2ꎬ㊀张保龙1ꎬ2(1.海南大学热带作物学院/三亚南繁研究院ꎬ海南海口570228ꎻ2.江苏省农业科学院种质资源与生物技术研究所/江苏省农业生物学重点实验室ꎬ江苏南京210014)收稿日期:2022 ̄11 ̄25基金项目:江苏省农业科技自主创新基金项目[CX(21)2041]作者简介:江㊀群(1998-)ꎬ女ꎬ四川宜宾人ꎬ硕士研究生ꎬ主要从事水稻抗除草剂育种研究ꎮ(E ̄mail)1692579264@qq.com通讯作者:张保龙ꎬ(E ̄mail)zhbl2248@hotmail.comꎻ周珍珍ꎬ(E ̄mail)zhenzhenzhounj@163.com㊀㊀摘要:㊀创制非转基因抗除草剂水稻种质资源对于稻田杂草防控具有重要价值ꎮ本研究以甲基磺酸乙酯(EMS)水溶液诱变镇糯19水稻种子ꎬ获得1株能稳定遗传的可耐受乙酰辅酶A羧化酶(ACCase)抑制剂类除草剂高效盖草能的M3代水稻幼苗(突变体)ꎮ分别扩增镇糯19野生型和突变体的基因组DNA并进行测序和序列比对ꎬ发现突变体ACCase基因的开放阅读框(ORF)的第5374位碱基发生了点突变ꎬ导致编码的第1792位氨基酸由异亮氨酸突变为亮氨酸ꎮ镇糯19野生型和突变体分蘖盛期大田喷施3种田间推荐剂量的ACCase抑制剂类除草剂后农艺性状调查结果表明突变体对高效盖草能㊁精喹禾灵和唑啉草酯抗性明显高于野生型ꎮ本研究获得了能稳定遗传的非转基因抗AC ̄Case抑制剂类水稻新种质ꎬ具有一定的应用价值ꎬ为抗除草剂水稻育种提供了种质资源ꎮ关键词:㊀水稻ꎻ抗除草剂种质ꎻ甲基磺酸乙酯(EMS)ꎻ乙酰辅酶A羧化酶中图分类号:㊀S335.3㊀㊀㊀文献标识码:㊀A㊀㊀㊀文章编号:㊀1000 ̄4440(2023)02 ̄0305 ̄08EMSmutagenesistocreatericeanti ̄acetyl ̄CoAcarboxylaseinhibitor ̄her ̄bicidegermplasmJIANGQun1ꎬ㊀LINGXi ̄tie2ꎬ㊀TANGZhao ̄cheng2ꎬ㊀ZHOUZhen ̄zhen2ꎬ㊀ZHANGBao ̄long1ꎬ2(1.CollegeofTropicalCrops/SanyaNanfanResearchInstituteꎬHainanUniversityꎬHaikou570228ꎬChinaꎻ2.InstituteofGermplasmResourcesandBio ̄technology/ProvincialKeyLaboratoryofAgrobiologyꎬJiangsuAcademyofAgriculturalSciencesꎬNanjing210014ꎬChina)㊀㊀Abstract:㊀Cultivatingnon ̄transgenicherbicide ̄resistantricegermplasmresourcesisofgreatvalueforweedcontrolinricefields.InthisstudyꎬZhennuo19riceseedsweremutagenizedbyethylmethylsulfonate(EMS)solutionꎬandaM3generationofriceseedlingswithstableinheritanceandtolerancetoacetyl ̄CoAcarboxylase(ACCase)inhibitorherbicideswereobtained.ThegenomicDNAsofwild ̄typeandthemutantwereamplifiedandsequencedrespectively.Itwasfoundthattherewasapointmutationatthe5374thbaseoftheopenreadingframeoftheresistantriceACCasegeneꎬresultinginamutationoftheencoded1792thaminoacidfromisoleucinetoleucine.ThreekindsofACCaseinhibitorherbicidesweresprayedinthefieldandtheagronomictraitswereanalyzed.Theresultsshowedthattheresistanceofthemutanttohaloxy ̄fop ̄R ̄methylꎬquizalofop ̄P ̄ethylandpinoxadenwassignificantlyhigherthanthatofwildtype.Inthisstudyꎬanewnon ̄transgenicricegermplasmwithACCaseinhibitorresistancewasobtainedꎬwhichhadcertainapplicationvalueandcouldprovidegermplasmresourcesforherbicide ̄resistantricebreeding.Keywords:㊀riceꎻherbicide ̄resistantgermplasmꎻethylmethylsulfonate(EMS)ꎻacetylCoAcarboxylase503㊀㊀水稻是中国三大粮食作物之一ꎬ培育高产稳产的优质水稻是解决粮食问题的关键ꎮ稻田杂草严重影响水稻的产量和品质ꎬ杂草导致中国稻谷每年亏损率超过15%ꎬ部分地区甚至超过50%[1]ꎮ化学除草是当今世界使用最多的稻田除草方法ꎮ然而ꎬ过度使用除草剂不仅会导致杂草对除草剂产生抗性ꎬ还会对作物产生药害㊁降低水稻产量和品质ꎬ严重时甚至造成水稻颗粒无收[2]ꎮ因此ꎬ培育抗除草剂的水稻品种可以经济有效地解决稻田的杂草防除问题ꎮ乙酰辅酶A羧化酶(ACCase)是植物初级代谢中脂肪酸合成的关键酶之一ꎬ其主要功能是将乙酰辅酶A羧化为丙二酰辅酶Aꎮ该反应是脂肪酸合成的第一步ꎬ也是限速的关键步骤[3]ꎮ脂肪酸不仅是功能物质甘油三脂的组成成分ꎬ还能转化为作为细胞膜组成成分的磷脂[4]ꎮ自1958年发现乙酰辅酶A羧化酶可作为除草剂的作用靶标后ꎬ针对该靶标已开发了三大类除草剂并商品化应用ꎬ分别是芳氧苯氧基丙酸酯类(APP)[5]㊁环己烯酮类(CHD)[6]和新苯基吡唑啉类(DEN)[7 ̄8]ꎮ其中ꎬAPP类除草剂包括高效氟吡甲禾灵(Haloxyfop ̄R ̄methylꎬ又称高效盖草能)㊁精喹禾灵(Quizalofop ̄P ̄ethyl)㊁精恶唑禾草灵(Fenoxaprop ̄P ̄ethylꎬ又称骠马)㊁恶唑酰草胺(Metamifop)和氰氟草酯(Cyhalofop ̄butyl)等ꎮCHD类除草剂包括烯禾啶(Sethoxydim)㊁噻草酮(Cy ̄cloxydim)和环苯草酮(Profoxydim)等ꎻDEN类除草剂有唑啉草酯(Pinoxaden)ꎮ乙酰辅酶A羧化酶抑制剂类除草剂主要被用于控制禾本科杂草ꎬ具有高效㊁低毒㊁对后茬作物安全等特点[9]ꎮ目前ꎬ水稻生产中登记并许可使用的乙酰辅酶A羧化酶抑制剂类除草剂仅有氰氟草酯㊁恶唑酰草胺和环苯草酮ꎬ这极大限制了水稻生产中杂草的防治ꎮ因此培育抗乙酰辅酶A羧化酶抑制剂类除草剂水稻ꎬ不仅可以拓宽稻田除草剂的选择和使用范围ꎬ还可有效控制稻田杂草的发生与危害ꎮ化学诱变是培育和筛选抗性除草剂作物种质资源的重要方法ꎮEMS是非常有效且负面影响小的化学诱变剂ꎬ被广泛应用于构建优良性状的水稻突变体[10 ̄12]ꎮ顾佳清等利用EMS处理粳稻品种中花11ꎬ从诱变的水稻群体中筛选出高产的突变体ꎬ经过后代的纯化ꎬ得到了一个可以直接推广应用的水稻突变新品系申化一号[13]ꎮ陈忠明等通过EMS处理籼稻9311ꎬ筛选出了大粒的突变体M316和长穗突变体9311eR[14 ̄15]ꎮ本课题组用EMS诱变处理包括9311在内的多个水稻品种ꎬ成功筛选到多个抗咪唑啉酮类除草剂的突变体ꎬ进一步鉴定结果表明突变均发生在编码乙酰乳酸合成酶(ALS)靶标基因上[16]ꎮ本研究通过EMS诱变糯稻品种镇糯19构建突变群体ꎬ用APP类除草剂高效盖草能去筛选诱变处理后的M2代幼苗ꎬ获得能稳定遗传的抗性植株ꎬ并对抗性植株的ACCase基因位点突变㊁氨基酸序列变异进行鉴定ꎬ最后就3种不同ACCase抑制剂类除草剂对获得的抗除草剂材料农艺性状影响进行分析ꎬ旨在为水稻抗除草剂育种提供依据和材料ꎮ1㊀材料与方法1.1㊀材料与试剂供试水稻材料镇糯19由江苏丘陵地区镇江农业科学研究所提供ꎮ试验所用除草剂的种类及相关信息见表1ꎮ表1㊀本试验所用除草剂Table1㊀Herbicidesusedinthisstudy名称㊀类别来源推荐田间施用剂量(g/hm2ꎬa.i.)高效盖草能APP江苏中旗科技股份有限公司64.8精喹禾灵APP天津中农立华农用化学品有限公司60.0唑啉草酯DEN瑞士先正达作物保护有限公司45.0㊀㊀生物试剂甲基磺酸乙酯(EMS)购自美国Sigma ̄Aldrich公司ꎬ2ˑRapidTaqMasterMix㊁PhantaMaxSuper ̄FidelityDNAPolymerase聚合酶购自南京诺唯赞生物科技有限公司ꎬCTAB购自北京鼎国昌盛生物技术有限责任公司ꎮ1.2㊀镇糯19水稻种子的EMS诱变及抗ACCase抑制剂类除草剂突变体的筛选㊀㊀镇糯19种子(M1代)清水浸泡2h后ꎬ用质量浓度5 0mg/ml的EMS水溶液浸种处理14hꎬ硫代硫酸钠中和30min后ꎬ将种子捞出并用清水冲洗5~6遍ꎮ将诱变处理后的种子播种于大田ꎬM1代植株成熟后ꎬ种子混收(M2代)作为突变群体库ꎮ从突变群体库中取M2代种子播种于大棚苗床ꎬ待水稻幼苗长至3~4叶期时喷施64 8g/hm2ꎬa.i.高效盖草603江苏农业学报㊀2023年第39卷第2期能ꎬ施药后21d观察记录水稻表型并将正常生长的水稻苗移栽至盆钵内ꎬ单株收获种子得到突变体种子(M3代)ꎬM3代种子播种后得到M3代幼苗ꎮ1.3㊀抗除草剂突变体ACCase基因的PCR鉴定和碱基序列分析㊀㊀从国家水稻数据中心数据库(https://www.rice ̄data.cn/)获得水稻ACCase基因(OsACCꎬ序列号为LOC_Os05g22940)的碱基序列ꎮ根据OsACC基因的保守序列使用SnapGene6.0.2软件进行特异性引物设计ꎬ共设计了8对引物ꎬ分别是OsACC ̄F1~Os ̄ACC ̄F8和OsACC ̄R1~OsACC ̄R8(表2)ꎮ表2㊀本试验所用引物Table2㊀Primersusedinthisstudy引物名称㊀序列(5ᶄң3ᶄ)PCR产物长度(bp)OsACC ̄F1GTCAGATTTCACACATCTGGG1422OsACC ̄R1CAGGGGCACAAATAATGTACTOsACC ̄F2AAAAAGCTGCGTGAAGTATGC1614OsACC ̄R2TCTCGACTGTGAAGTGCTGCOsACC ̄F3CCCTATTGAAGACATCCTGATTG1597OsACC ̄R3AACAGAAATGGCATGATGGAOsACC ̄F4CAAACGTAGACTACACAGTTGAC1641OsACC ̄R4TGTTTGGCACCATTATGAGAAOsACC ̄F5TTGACAAGGTAAACATCATGTCC1635OsACC ̄R5AAAAGGTCATTGAAAAATTCACGOsACC ̄F6TCTATCCAAATCCTGCTGCC1631OsACC ̄R6AATGGCCAGTTCTAATTGCGOsACC ̄F7AGTTTTCTTCGGGCCAGATT1634OsACC ̄R7GGCTGGTCAAGACGCTGTATOsACC ̄F8CATGGAAGTGCTGCTATTGCCAG1866OsACC ̄R8CAGACTTGCACTTTCATCTGGCA㊀㊀采用CTAB法[14]提取水稻的基因组DNAꎬ取M3代三叶一心期的叶片0 5g放在带有1颗小钢珠的2ml离心管中ꎬ放到液氮中冷冻至叶片组织变脆ꎬ再将离心管放到频率为60Hz的组织研磨机研磨2minꎬ然后加入400μlCTAB提取液ꎬ离心管65ħ水浴30min后ꎬ在通风橱中加入400μl氯仿ꎬ充分混匀至提取液呈乳绿色ꎬ12000r/min离心10minꎬ在离心期间标记好管号ꎬ将600μl无水乙醇加入到已经标记好的1 5ml离心管中ꎬ移液枪吸取上清液300μl加到已经准备好的离心管中ꎬ上下颠倒混匀再沉淀1h以上ꎬ12000r/min离心10minꎬ倒掉上清液ꎬ开盖ꎬ室温下风干12h至离心管底部有明显的白色DNA沉淀ꎬ风干后加入灭菌蒸馏水200μlꎬ于-20ħ保存ꎮ以M3代的基因组DNA为模板ꎬ采用2ˑRapidTaqMasterMix或PhantaMaxSuper ̄FidelityDNAPolymerase聚合酶扩增OsACC基因的8个片段ꎮ用1%琼脂糖凝胶进行电泳检测ꎮ将条带大小正确的PCR产物送南京擎科生物科技有限公司进行测序ꎻ使用SnapGene6.0.2软件分析测序结果ꎬ明确野生型和突变体的OsACC基因碱基序列差异性ꎮ1.4㊀喷施除草剂后水稻农艺性状调查2022年在江苏省农业科学院试验基地进行镇糯19野生型和突变株系对3种乙酰辅酶A羧化酶抑制剂类除草剂耐受性试验ꎮ5月中旬播种ꎬ6月中旬插秧ꎮ试验设分别喷施高效盖草能㊁精喹禾灵㊁唑啉草酯及清水对照4个处理ꎬ每处理2.5mˑ4 0mꎮ移栽行距为0 25mꎬ株距为0 15mꎮ按照常规大田生产进行浇水和施肥等田间管理ꎮ镇糯19野生型和突变体幼苗移栽大田27d后ꎬ进行高效盖草能㊁精喹禾灵㊁唑啉草酯及清水(CK)的喷施处理ꎮ除草剂的用量见表1ꎮ各处理选择连续的20株ꎬ在水稻喷施除草剂前以及喷施除草剂后30d㊁90d进行茎蘖数㊁株高㊁主茎旗叶长度等农艺性状调查ꎮ其中ꎬ喷药后90dꎬ水稻已进入成熟期ꎬ统计的茎蘖数为成穗数ꎮ1.5㊀数据处理与统计分析采用MicrosoftExcel2019进行数据处理ꎬ用GraphPadPrism8.0.1软件进行统计分析ꎮ2㊀结果与分析2.1㊀抗除草剂突变体筛选高效盖草能是一种内吸传导型除草剂ꎬEMS诱变的镇糯19M2代水稻幼苗在3~4叶期喷施高效盖草能7d后ꎬ绝大部分水稻幼苗叶片颜色变成浅绿ꎻ喷施高效盖草能21d后ꎬ敏感植株叶片几乎完全失去绿色㊁部分已经枯死ꎻ具有抗性的植株能继续正常生长ꎮ经大量筛选后ꎬ最终获得1株具有高效盖草能抗性的M2单株(图1)ꎬ成熟后收获单株种子ꎬ得到M3代抗性突变体ꎮ2.2㊀OsACC突变位点已知高效盖草能的作用靶标是ACCaseꎬ植物对703江㊀群等:EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质图1㊀喷施高效盖草能后筛选到的M2代抗性水稻植株Fig.1㊀M2generationresistantriceplantscreenedafterspra ̄yingwith64.8ga.i./hm2haloxyfop ̄R ̄methyl高效盖草能的抗性主要源于ACCase基因的突变[17 ̄19]ꎮ为了确定突变体中靶标基因是否发生突变ꎬ我们用了8对引物对野生型(镇糯19 ̄WT)和抗性M3单株(镇糯19 ̄1792)的基因进行扩增ꎬ全部都获得了与预期大小相符合的条带(图2)ꎮ㊀㊀上述PCR扩增的产物经测序和碱基序列比对ꎬ发现相对于野生型OsACC的ORFꎬ突变体OsACC基因中存在一个点突变ꎬ其开放阅读框(ORF)的第5374位碱基由A突变成Tꎬ从而引起编码的第1792位氨基酸由异亮氨酸(Ile)突变为亮氨酸(Leu)(图3A)ꎮOsACC蛋白的全长有2327个氨基酸ꎬ将Os ̄ACC蛋白全长氨基酸序列在NCBI的ConservedDo ̄main数据库(https://www.ncbi.nlm.nih.gov/Struc ̄ture/cdd/wrpsb.cgi)进行保守结构域分析ꎬ发现其包含了4个结构域(Domain):生物素羧化酶(BC)㊁生物素羧基载体蛋白(BCCP)㊁乙酰辅酶A羧化酶中心(ACCcentral)和羧基转移酶(CT)(图3B)ꎮ进一步的氨基酸序列分析结果表明ꎬ突变体中第1792位氨基酸的突变位于CT结构域ꎬ该突变类型与已报道的大穗看麦娘(Alopecurusmyosuroides)的抗性位点突变类型是一致的ꎬ对应于其ACCase氨基酸序列第1781位点ꎻ突变类型也相同ꎬ均由Ile突变为Leu(图3B和3C)[17]ꎮ因此ꎬ突变体抗除草剂功能的获得是由OsACC氨基酸序列第1792位氨基酸由异亮氨酸突变为亮氨酸引起的ꎮ2.3㊀突变体的农艺性状在分别喷施高效盖草能㊁精喹禾灵和唑啉草酯14d后ꎬ野生型植株生长均受到了显著影响ꎬ大部分植株叶片出现枯黄症状ꎮ突变体植株在分别喷施以上3种除草剂后ꎬ叶片仍然是绿色且可以正常生长ꎬ表明突变体对这3种除草剂均具有抗性(图4)ꎮ㊀㊀分蘖期分别喷施3种不同除草剂后ꎬ野生型和突变体株高㊁分蘖数及旗叶长度的变化如图5所示ꎮ结果显示ꎬ在喷施清水处理的情况下ꎬ野生型和突变体植株的株高在处理前(0dꎬ即幼苗移栽到大田27d)基本没有差异ꎬ但在处理后30d和90dꎬ突变体的株高显著低于野生型的株高(图5A)ꎻ两者在处理前㊁后的单株茎蘖数均无明显差异(图5E)ꎮ在分别喷施3种不同除草剂前(0d)ꎬ野生型和突变体植株的株高和单株分蘖数都没有明显差异ꎬ但是在喷施处理后ꎬ两者受除草剂的影响表现出明显差异(图5B~图5D㊁图5F~图5H)ꎮ其中ꎬ在喷施高效盖草能30d和90d后ꎬ突变体的株高均显著高于野生型(图5B)ꎬ单株茎蘖数也显著多于野生型(图5F)ꎮ野生型对精喹禾灵和唑啉草酯都非常敏感ꎬ喷施田间推荐剂量后水稻植株均死亡ꎬ因此未统计喷药后的株高和分蘖数ꎬ而突变体对这两种除草剂表现出较强的抗性ꎬ所有植株存活且能正常生长ꎬ株高随时间逐渐增加(图5C和5D)ꎮ突变体的单株茎蘖数在精喹禾灵处理后随时间呈先增后减趋势ꎬ但经唑啉草酯处理后变化不明显ꎬ未出现明显增加现象(图5G和5H)ꎮ喷施清水处理的突变体旗叶长度显著短于野生型ꎻ高效盖草能处理后ꎬ突变体的旗叶长度显著长于野生型(图5I)ꎮ由于野生型在喷施田间推荐剂量的精喹禾灵和唑啉草酯后植株已经枯死ꎬ因此未能进行旗叶长度统计ꎮ综合以上结果ꎬ在田间推荐剂量下ꎬ突变体对高效盖草能㊁精喹禾灵和唑啉草酯的抗性水平均高于野生型ꎮ3㊀讨论植物对除草剂的抗性机制包括非靶标和靶标抗性两大类ꎮ其中ꎬ非靶标抗性是由靶标基因以外的突变引起的ꎬ使植物对除草剂的吸收或转运率降低㊁螯合或代谢作用增强ꎻ靶标抗性是由除草剂的靶标基因发生突变引起的[20]ꎮ现在已发现的大部分植物抗ACCase抑制剂类除草剂的抗性机制是由于ACCase基因碱基突变引起氨基酸位点发生变异ꎬ这也是导致杂草抗药性产生的主要原因[21 ̄22]ꎮ截止803江苏农业学报㊀2023年第39卷第2期M表示DNAmarkerꎻ泳道1表示野生型ꎻ泳道2表示突变体ꎮF1~F8㊁R1~R8为引物ꎬ见表2ꎮ图2㊀镇糯19野生型和突变体中OsACC基因的PCR扩增结果Fig.2㊀PCRamplificationofOsACCinZhennuo19wild ̄typeandmutantA:突变体(镇糯19 ̄1792)中OsACC基因的Sanger测序色谱图ꎻB:OsACC蛋白结构域示意图ꎻC:野生型(镇糯19 ̄WT)和突变体(镇糯19 ̄1792)的羧基转移酶(CT)结构域氨基酸序列比对ꎮ图3㊀镇糯19突变体中突变基因OsACC及其编码氨基酸序列分析Fig.3㊀AnalysisofmutantgeneOsACCanditsencodedaminoacidsequenceinZhennuo19mutant903江㊀群等:EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质镇糯19 ̄WT㊁镇糯19 ̄1792分别表示镇糯19野生型和突变体ꎻGCN㊁JK㊁ZL和H2O分别表示喷施高效盖草能㊁精喹禾灵㊁唑啉草酯及清水处理ꎮ图4㊀镇糯19野生型和突变体田间喷施不同除草剂后的表型Fig.4㊀PhenotypesofZhennuo19wild ̄typeandmutantaftersprayingwithdifferentherbicidesinthefield目前ꎬ杂草中已报道了十几种ACCase氨基酸置换与其抗药性相关ꎬ分别对应于大穗看麦娘ACCase的7个氨基酸位点(均位于CT结构域内):第1781位㊁第1999位㊁第2027位㊁第2041位㊁第2078位㊁第2088位和第2096位[22 ̄25]ꎮ在以上这些突变中ꎬ以第1781位氨基酸由Leu突变成Ile最为普遍ꎬ对三大类不同的ACCase抑制剂类除草剂都表现出高抗性ꎬ却没有适合度代价(Fitnesscost)[26 ̄28]ꎮ本研究通过筛选EMS诱变的镇糯19水稻突变体ꎬ鉴定到了1个能稳定遗传的抗除草剂突变体ꎮ对突变体进行了基因鉴定ꎬ确定其编码靶标蛋白OsACC的第1792位氨基酸由Leu突变成Ileꎮ该突变类型与已报道的突变类型一致ꎬ对应于大穗看麦娘ACCase第1781位氨基酸突变ꎮ这是该突变类型使水稻获得多种ACCase抑制剂类除草剂抗性的首次报道ꎮEMS是最常见的化学诱变剂ꎬ在植物的诱变育种中被广泛应用[29]ꎮ本试验通过EMS诱变镇糯19种子ꎬ筛选到了抗ACCase抑制剂类除草剂的水稻植株ꎬ突变体能耐受田间推荐剂量的高效盖草能㊁精喹禾灵和唑啉草酯ꎮ其中ꎬ喷施了田间推荐剂量的唑啉草酯后ꎬ镇糯19野生型植株在处理30d后几乎全部死亡ꎻ喷施了田间推荐剂量的精喹禾灵后ꎬ野生型的植株在喷施30d后全部死亡ꎻ而突变体在分别喷施3种除草剂后ꎬ均未出现死亡现象ꎬ基本可以正常生长ꎮ所获得的抗性突变体对高效盖草能㊁精喹禾灵㊁唑啉草酯的抗性水平均明显强于野生型ꎮ突变体和野生型的最小致死剂量或50%抑制浓度(GR50)㊁OsACC酶活性的差异尚有待进一步明确ꎮ大豆㊁棉花和玉米等转基因作物已在全球范围内进行了商品化生产ꎬ产生了巨大的社会效益和经济效益ꎮ目前为止ꎬ中国虽然有多种转基因作物已经被正式批准商品化生产ꎬ但进行大面积种植的仅013江苏农业学报㊀2023年第39卷第2期H2O㊁GCN㊁JK㊁ZL分别表示喷施清水㊁高效盖草能㊁精喹禾灵㊁ꎬ∗∗表示在0.01水平上极显著ꎮND表示没有数据ꎬns表示没有显著差异ꎮ图5㊀不同除草剂处理下的水稻株高㊁分蘖数和旗叶长度Fig.5㊀Plantheightꎬtillernumberandflagleaflengthofriceunderdifferentherbicidetreatments有番木瓜和棉花ꎮ2009年ꎬ农业部颁发了中国拥有自主知识产权的转Bt基因抗虫水稻生产应用安全证书ꎬ但目前中国尚未批准转基因水稻的商业化生产ꎮ因此ꎬ培育非转基因的抗除草剂水稻品种具有重要价值ꎮ上世纪90年代晚期ꎬ美国路易斯安那州州立大学稻米研究中心通过EMS诱变技术育成了一系列耐咪唑啉酮类除草剂(ALS抑制剂类除草剂)的非转基因水稻品种ꎮ2002年ꎬ巴斯夫公司开发了非转基因抗咪唑啉酮类除草剂的水稻品种Clearf ̄ield在美国进行了商业化推广ꎬ解决了水稻种植的杂草稻危害问题[30]ꎮ2018年ꎬ巴斯夫又在美国上市了非转基因水稻品种Provisiaꎬ可以抗精喹禾灵ꎬ拟与抗咪唑啉酮类除草剂水稻品种Clearfield进行轮作并交替使用两种不同作用机理的除草剂ꎬ实现对杂草稻和其他一年生杂草的可持续性防控[31]ꎮ本研究通过EMS诱变筛选到的抗ACCase抑制剂类除草剂突变体ꎬ具有与抗除草剂精喹禾灵水稻品种Provisia类似的抗除草剂性状ꎬ可为中国非转基因抗除草剂水稻育种提供重要材料ꎮ4㊀结论本研究通过EMS诱变筛选获得了可稳定遗传的抗ACCase抑制剂类除草剂的水稻突变体材料ꎬ可耐受3种不同田间推荐剂量的除草剂ꎬ具有一定的生产应用价值ꎮ野生型在喷施田间推荐剂量的高效盖草能㊁精喹禾灵㊁唑啉草酯后ꎬ株高和分蘖均受到严重抑制甚至死亡ꎬ但突变体基本能正常生长ꎮ突变体中OsACC突变基因编码蛋白质的第1792位氨基酸由Ile变成Leuꎬ使其对ACCase抑制剂类除草剂的耐受性显著提高ꎮ在当前中国转基因水稻尚未放开㊁公众对转基因作物品种存在疑虑的大背景下ꎬ本研究获得的非转基因抗除草剂材料具有良好的应用前景ꎮ113江㊀群等:EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质参考文献:[1]㊀董立尧ꎬ高㊀原ꎬ房加鹏ꎬ等.我国水稻田杂草抗药性研究进展[J].植物保护ꎬ2018ꎬ44(5):69 ̄76.[2]㊀程艳勤.浅析除草剂对水稻的危害及治理[J].农技服务ꎬ2016ꎬ33(6):109 ̄114.[3]㊀KONISHITKUJꎬSHINOHARAKꎬYAMADAKꎬetal.Acetyl ̄CoAcarboxylaseinhigherplants:mostplantsotherthangramineaehaveboththeprokaryoticandtheeukaryoticformsofthisenzyme[J].PlantandCellPhysiologyꎬ1996ꎬ37(2):117 ̄122. [4]㊀王㊀爽ꎬ张荣全ꎬ叶㊀非.乙酰辅酶A羧化酶抑制剂的研究进展[J].农药科学与管理ꎬ2003(10):26 ̄32.[5]㊀蔡靖萱.扬州市小麦田菵草对ACCase抑制剂的抗性研究[D].扬州:扬州大学ꎬ2020.[6]㊀RENDINAARꎬFELTSJM.CyclohexanedioneHerbicidesarese ̄lectiveandpotentinhibitorsofacetyl ̄CoAcarboxylasefromgrasses[J].PlantPhysiolꎬ1988ꎬ86(4):983 ̄986.[7]㊀袁国徽ꎬ王恒智ꎬ赵㊀宁ꎬ等.耿氏硬草对乙酰辅酶A羧化酶类除草剂抗性水平及分子机制初探[J].农药学学报ꎬ2016ꎬ18(3):304 ̄310.[8]㊀董元海.新苯基吡唑啉类除草剂唑啉草酯的合成[D].武汉:武汉工程大学ꎬ2017.[9]㊀刘博宏ꎬ叶㊀非.芳氧苯氧基丙酸酯类除草剂的应用进展[J].农药科学与管理ꎬ2011ꎬ32(2):20 ̄25.[10]吕㊀军ꎬ刘㊀军ꎬ姜秀英ꎬ等.EMS诱导水稻 辽星1号ᶄ突变体的筛选与鉴定[J].分子植物育种ꎬ2022ꎬ20(12):4038 ̄4043. [11]董颖苹ꎬ连㊀勇ꎬ何庆才ꎬ等.植物化学诱变技术在育种中的运用及进展Ⅱ.突变体的筛选及分子检测[J].种子ꎬ2005ꎬ24(8):54 ̄58.[12]黄㊀静.水稻EMS诱变效率和品种内遗传多态性分析[D].福州:福建农林大学ꎬ2015.[13]顾佳清ꎬ张智奇ꎬ周㊀音ꎬ等.EMS诱导水稻中花11突变体的筛选和鉴定[J].上海农业学报ꎬ2005ꎬ21(1):7 ̄11.[14]陈忠明ꎬ王秀娥.水稻强优势恢复系9311粒重的诱变改良[J].分子植物育种ꎬ2005ꎬ3(3):353 ̄356.[15]陈忠明ꎬ王秀娥ꎬ胡兴雨ꎬ等.水稻长穗颈恢复系9311eR的诱变选育[J].江苏农业科学ꎬ2005(4):9 ̄11.[16]陈天子ꎬ余㊀月ꎬ凌溪铁ꎬ等.EMS诱变水稻创制抗咪唑啉酮除草剂种质[J].核农学报ꎬ2021ꎬ35(2):253 ̄261.[17]DÉLYECꎬCALMÈSÉꎬMATÉJICEKA.SNPmarkersforblack ̄grass(AlopecurusmyosuroidesHuds.)genotypesresistanttoacetylCoA ̄carboxylaseinhibitingherbicides[J].TheoreticalandAppliedGeneticsꎬ2002ꎬ104(6):1114 ̄1120.[18]DÉLYECꎬZHANGXꎬCHALOPINCꎬetal.Anisoleucineresi ̄duewithinthecarboxyl ̄transferasedomainofmultidomainacetyl ̄coenzymeAcarboxylaseisamajordeterminantofsensitivitytoary ̄loxyphenoxypropionatebutnottocyclohexanedioneinhibitors[J].PlantPhysiologyꎬ2003ꎬ132(3):1716 ̄1723.[19]DÉLYECꎬZHANGXꎬMICHELSꎬetal.Molecularbasesforsensitivitytoacetyl ̄coenzymeAcarboxylaseinhibitorsinblack ̄grass[J].PlantPhysiologyꎬ2005ꎬ137(3):794 ̄806.[20]POWLESSBꎬYUQ.EvolutioninAction:plantsresistanttoher ̄bicides[J].AnnualReviewofPlantBiologyꎬ2010ꎬ61(1):317 ̄347.[21]袁国徽ꎬ田志慧ꎬ高㊀原ꎬ等.上海市水稻田千金子对3种乙酰辅酶A羧化酶抑制剂的抗性现状及酶突变机制[J].农药学学报ꎬ2022ꎬ24(3):492 ̄500.[22]BECKIEHJꎬTARDIFFJ.Herbicidecrossresistanceinweeds[J].CropProtectionꎬ2012ꎬ35:15 ̄28.[23]DENGWꎬCAIJꎬZHANGJꎬetal.MolecularbasisofresistancetoACCase ̄inhibitingherbicidecyhalofop ̄butylinChinesespran ̄gletop(Leptochloachinensis(L.)Nees)fromChina[J].PesticBiochemPhysiolꎬ2019ꎬ158:143 ̄148.[24]PENGYꎬPANLꎬLIUDꎬetal.Confirmationandcharacterizationofcyhalofop ̄butyl ̄resistantChinesesprangletop(Leptochloachinensis)populationsfromChina[J].WeedScienceꎬ2020ꎬ68(3):253 ̄259.[25]张㊀怡ꎬ陈丽萍ꎬ徐笔奇ꎬ等.浙江稻区千金子对氰氟草酯和噁唑酰草胺的抗药性及其分子机制研究[J].农药学学报ꎬ2020ꎬ22(3):447 ̄453.[26]VILAAIUBMMꎬNEVEPꎬPOWLESSB.ResistancecostofacytochromeP450herbicidemetabolismmechanismbutnotanAC ̄CasetargetsitemutationinamultipleresistantLoliumrigidumpopulation[J].NewPhytologistꎬ2005ꎬ167(3):787 ̄796. [27]MENCHARIYꎬCHAUVELBꎬDARMENCYHꎬetal.Fitnesscostsassociatedwiththreemutantacetyl ̄coenzymeAcarboxylaseallelesendowingherbicideresistanceinblack ̄grassAlopecurusmy ̄osuroides[J].JournalofAppliedEcologyꎬ2008ꎬ45(3):939 ̄947. [28]WANGTꎬPICARDJCꎬTIANXꎬetal.Aherbicide ̄resistantACCase1781setariamutantshowshigherfitnessthanwildtype[J].Heredity(Edinb)ꎬ2010ꎬ105(4):394 ̄400.[29]SERRATXꎬESTEBANRꎬGUIBOURTNꎬetal.EMSmutagene ̄sisinmatureseed ̄derivedricecalliasanewmethodforrapidlyobtainingTILLINGmutantpopulations[J].PlantMethodsꎬ2014ꎬ10(1):5.[30]SHAXYꎬLINSCOMBESDꎬGROTHDE.Fieldevaluationofimidazolinone ̄tolerantClearfieldrice(OryzasativaL.)atnineLouisianalocations[J].CropScienceꎬ2007ꎬ47(3):1177 ̄1185. [31]CAMACHOJRꎬLINSCOMBESDꎬSANABRIAYꎬetal.Inherit ̄anceofProvisiariceresistancetoquizalofop ̄p ̄ethylunderlabora ̄toryandgreenhouseenvironments[J].Euphyticaꎬ2019ꎬ215(4):83.(责任编辑:石春林)213江苏农业学报㊀2023年第39卷第2期。
芒果苷对胰岛素抵抗HepG2细胞糖脂代谢的影响

芒果苷对胰岛素抵抗HepG2细胞糖脂代谢的影响作者:黎梓霖金惠杰方佳刘奕明林爱华来源:《中国药房》2021年第09期中圖分类号 R285.5 文献标志码 A 文章编号 1001-0408(2021)09-1082-07DOI 10.6039/j.issn.1001-0408.2021.09.10摘要目的:分析芒果苷(MGF)对胰岛素抵抗HepG2细胞(IR-HepG2细胞)糖脂代谢的影响,并探讨潜在机制。
方法:以人肝癌HepG2细胞为对象,以1 mmol/L棕榈酸+2 mmol/L油酸联合培养建立IR-HepG2细胞模型。
以盐酸二甲双胍为阳性对照,分别检测低、中、高浓度MGF(125、250、500 μmol/L)作用24 h对IR-HepG2细胞中校正葡萄糖耗氧量和三酰甘油(TG)、总胆固醇(TC)含量的影响;采用实时荧光定量聚合酶链式反应技术检测细胞中腺苷一磷酸活化蛋白激酶(AMPK)通路上游关键因子脂联素(APN)、脂联素受体2(AdipoR2)、APPL1、AMPK以及下游胰岛素信号通路关键因子胰岛素受体底物1(IRS-1)、蛋白激酶B(Akt)、葡萄糖转运体4(GLUT4)mRNA的相对表达量;采用Western blot 法检测AMPK蛋白的磷酸化水平。
结果:与对照组比较,模型组细胞校正葡萄糖消耗量和APN、AdipoR2、APPL1、AMPK、IRS-1、GLUT4 mRNA的相对表达量以及AMPK蛋白磷酸化水平均显著降低,TG、TC含量均显著升高(P关键词芒果苷;胰岛素抵抗;人HepG2细胞;糖脂代谢;腺苷一磷酸活化蛋白激酶信号通路;脂联素Effects of Mangiferin on Glucose and Lipid Metabolism of Insulin-resistant HepG2 CellsLI Zilin1,JIN Huijie1,FANG Jia1,LIU Yiming1,2,LIN Aihua3(1. Phase Ⅰ Clinical Trial Center, Guangdong Provincial Hospital of TCM/the Second Affiliated Hospital of Guangzhou University of TCM, Guangzhou 510120, China; 2. Guangdong Provincial Key Laboratory ofClinical Research on Traditional Chinese Medicine Syndrome, Guangdong Provincial Hospital of TCM/the Second Affiliated Hospital of Guangzhou University of TCM, Guangzhou 510120,China; 3. Dept. of Pharmacy, Guangdong Provincial Hospital of TCM/Zhuhai Hospital, the Second Affiliated Hospital of Guangzhou University of TCM, Guangdong Zhuhai 519000,China)ABSTRACT OBJECTIVE: To analyze the effects of mangiferin (MGF) on glucose and lipid metabolism in insulin resistance (IR) HepG2 cells, and to explore the potential mechanism. METHODS: Using human hepatoma HepG2 cells as research objects, 1 mmol/L palmitic acid and 2 mmol/L oleic acid were used to establish the IR-HepG2 cell model. Using metformin hydrochloride as positive control, the effects of low-concentration, medium-concentration and high-concentration MGF (125, 250,500 μmol/L) on the corrected glucose consumption, the contents of triglyceride (TG) and total cholesterol (TC) in IR-HepG2 cells were detected. The mRNA expression of APN, AdipoR2, APPL1, AMPK in the upstream of AMPK signaling pathway and IRS-1, Akt and GLUT4 in the downstream insulin signaling pathway were detected by RT-PCR. The phosphorylation level of AMPK protein was detected by Western blot assay. RESULTS:Compared with control group, corrected glucose consumption, mRNA expression of APN,AdipoR2, APPL1, AMPK, IRS-1 and GLUT4, as well as the phosphorylation level of AMPK protein were decreased significantly in model group, while the contents of TG and TC were increased significantly (PKEYWORDS Mangiferin; Insulin resistance; Human HepG2 cells; Glucose and lipid metabolism; AMPK signaling pathway; APN2型糖尿病(T2DM)多由胰岛素抵抗(IR)和B细胞功能障碍所致[1],其中IR主要表现为肝脏、肌肉、脂肪等靶组织对胰岛素的敏感性下降以及对葡萄糖的摄取及利用减少[2]。
“继糖感受机制”或可治疗肥胖症

62绿农业科学工程3卷疫反应。
他们发现,在进行两次注射后的35#至50#内,4头牛的血液中便产生了针对H I V的广泛中和抗体,其中一种被称为)C-C o w l的抗 体能中和约2/3的H I V。
研究人员指出,虽然少数H IV 感染者体内会产生广泛中和抗体,但这些抗体都是在感染很长一段时间后产生,此时体内的病毒已进化到能抵制这些抗体了。
而新研究表明,牛能在极短时间内产生广泛中和抗体,并在与H I V的对抗中获得优势。
虽然目前牛抗体还不太适合在人类中临床使用,但弄清楚这种快速产生抗体的机制,对于抗艾疫苗的设计非常有用,也有助于开发新的艾滋病防治手段。
(《自然》杂志)“繼糖感受机制”或可治疗肥胖症近期,《自然》杂志发表了厦门大学林圣彩教授课题组的研究成果,他 们发现了生理状态下机体感受葡萄糖水平并调节代谢模式的机制,对开 发用于治疗肥胖症,乃至延长寿命的药物具有深远意义。
生活中的生物体内,葡萄糖水平的波动十分常见,睡一大觉、剧烈运动几个小时或者太忙了没时间吃饭,都会引起葡萄糖水平的显著下降。
这 时,机体能够触发一套有效的过程应对这类“不利情况”,林圣彩课题组研究发现,其中最为关键的就是激活“代谢的核心调节者”一A M P K。
此前研究认为,葡萄糖水平下降引起细胞内的能量分子A T P含量下降,进而引起另一代表低能量状态的分子A M P水平的上升,并由A M P直接激活A M P K,维持新陈代谢平衡。
无论在不含葡萄糖的细胞培养条件下,还是在饥饿的低血糖的动物体内,都不能观测到A M P水平的上升,这充分说明了机体有一套不为人知的、独立于A M P的感应葡萄糖水平的机制。
林圣彩课题组进一步研究揭示了这一完整过程:葡萄糖水平下降将引起葡萄糖代谢物下降,并被糖酵解通路上的代谢酶感应,后者启动激活A M P K的溶酶体途径进而介导A M P K的激活。
林圣彩教授进一步地把葡萄糖水平总结为一种“状态信号”,以区别于传统的“能量信号”。
厦门大学发现“葡萄糖感受机制”,有助于治疗肥胖症

厦门大学发现“葡萄糖感受机制”,有助于治疗肥胖症
佚名
【期刊名称】《生物学教学》
【年(卷),期】2017(42)12
【摘要】据2017年7月20日《科技日报》报道,《自然》杂志发表了厦门大学林圣彩教授课题组发现了生理状态下机体感受葡萄糖水平并调节代谢模式的机制,对开发用于治疗肥胖症,乃至延长寿命的药物具有深远意义.
【总页数】1页(P72-72)
【关键词】厦门大学;葡萄糖水;肥胖症;机制;感受;治疗;《科技日报》;《自然》杂志【正文语种】中文
【中图分类】TS972.161
【相关文献】
1.“繼糖感受机制”或可治疗肥胖症 [J], ;
2.研究单基因糖尿病有助于发现2型糖尿病发病机制及治疗靶点 [J], 王晓晶; 肖建中
3.厦门大学发现一种基因或有助于治疗老年痴呆症 [J],
4.日本发现白血病癌细胞增殖机制或有助于开发治疗癌症药物 [J],
5.新发现:有助于开发新的肥胖症治疗方法 [J],
因版权原因,仅展示原文概要,查看原文内容请购买。
2015年纳入973计划(含重大科学研究计划)结题验收项目清单
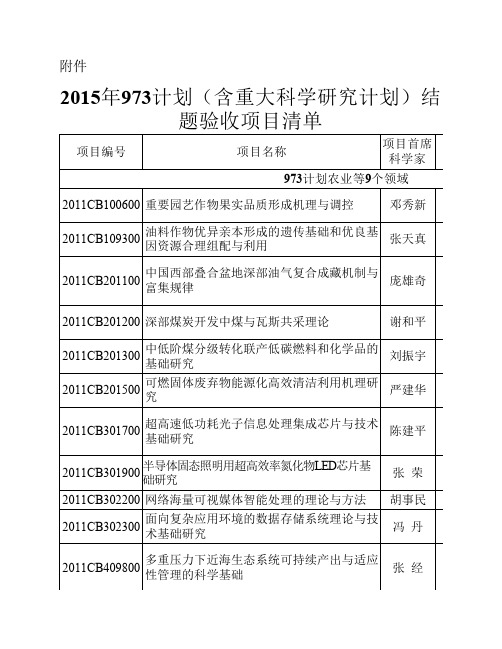
人多能干细胞多能性维持和发育潜能差异的 系统研究
康九红
2011CB965300
干细胞分化与重新编程中蛋白质的结构与功 能研究
2011CBA00300 全量子网络
2011CBA01000
诱导多功能干细胞(iPS)猪与小型猪疾病 模型
孙方霖 姚期智 刘忠华
孟安明
2011CB944500
雌性生育力维持调节机制研究及生殖资源库 建立
乔杰
2011CB944600
植物减数分裂过程中染色体相互作用的分子 机理
马红
2011CB946100 胸腺的起源、发生、维持与退化
2011CB952000
大尺度土地利用/覆盖变化对区域气候影响 的研究
张毓 郭维栋
2011CB965100
庞雄奇
2011CB201200 深部煤炭开发中煤与瓦斯共采理论
谢和平
2011CB201300
中低阶煤分级转化联产低碳燃料和化学品的 基础研究
刘振宇
2011CB201500
可燃固体废弃物能源化高效清洁利用机理研 究
严建华
2011CB301700
超高速低功耗光子信息处理集成芯片与技术 基础研究
陈建平
2011CB301900
2011CB932500
ቤተ መጻሕፍቲ ባይዱ
功能导向的纳米超分子组装体结构调控与可 控制备
刘育
2011CB932900
新型图像传感器及并行图像处理芯片的研究 与集成
2011CB933000 碳基无掺杂纳电子器件和集成电路
2011CB933300
基于纳米结构的新型柔性纤维基可编织光伏 器件重要基础问题研究
郑厚植 彭练矛 邹德春
中枢神经损伤修复与功能重建中胶质细胞的 作用及意义
国家重大科学研究计划2012年立项项目清单

国家重大科学研究计划2012年立项项目清单项目编号项目名称项目首席2012CB910100 代谢相关蛋白质修饰在肿瘤发生发展过程中的作用及机制赵世民复旦大学教育部上海市科学技术委员会2012CB910200 天然免疫应答相关蛋白的鉴定、结构与功能舒红兵武汉大学教育部湖北省科学技术厅2012CB910300 泛素-蛋白酶体:系统性发现其底物、发掘新作用机制及其生物学意义秦钧军事医学科学院放射与辐射医学研究2012CB910400 重要G蛋白偶联受体的结构与功能研究及配体发现刘明耀华东师范大学教育部上海市科学技术委员会2012CB910500 植物表观遗传机制与重要调控蛋白质的功能和结构研究沈文辉复旦大学教育部上海市科学技术委员会2012CB910600 蛋白质定量新方法及相关技术研究张丽华中国科学院大连化学物理研究所中国科学院2012CB910700 肿瘤发生发展中关键蛋白的功能与调控肖智雄四川大学教育部四川省科学技术厅2012CB910800 炎症诱导肿瘤的分子调控网络研究林安宁中国科学院上海生命科学研究院上海市科学技术委员会中国科学院2012CB910900 植物表观遗传调控及其在重要发育过程中的作用机制及结构基础研究邓兴旺北京大学教育部2012CB911000 蛋白质的生成、修饰与质量控制 Sarah Perrett 中国科学院生物物理研究所中国科学院2012CB911100 病毒与宿主细胞相互作用分子机制的研究于晓方吉林大学教育部2012CB911200 端粒相关蛋白对人类重大疾病作用机制的研究刘俊平杭州师范大学浙江省科学技术厅2012CB921300 极端条件下量子输运的研究和调控牛谦北京大学教育部2012CB921400 异质界面诱导的新奇量子现象及调控龚新高复旦大学教育部上海市科学技术委员会2012CB921500 人工微结构材料中光、声以及其它元激发的调控彭茹雯南京大学教育部2012CB921600 受限空间中光与超冷原子分子量子态的调控及其应用贾锁堂山西大学山西省科学技术厅2012CB921700 功能关联电子材料及其低能激发与拓扑量子性质的调控研究鲍威中国人民大学教育部2012CB921800 全固态量子信息处理关键器件的物理原理及技术实现肖敏南京大学教育部2012CB921900 光场调控及与微结构相互作用研究王慧田南开大学教育部天津市科学技术委员会2012CB922000 氧化物复合量子功能材料中的多参量过程及效应陆亚林中国科学技术大学中国科学院2012CB922100 囚禁单原子(离子)与光耦合体系量子态的操控詹明生中国科学院武汉物理与数学研究所中国科学院2012CB932200 纳米金属材料的多级结构制备及优异性能探索研究卢柯中国科学院金属研究所中国科学院2012CB932300 纳米材料功能化宏观体系的构筑和性能研究姜开利清华大学教育部2012CB932400 光功能导向的硅纳米结构高效、可控制备及其应用的基础研究张晓宏中国科学院理化技术研究所中国科学院2012CB932500 肝癌治疗的新型纳米药物研究杨祥良华中科技大学教育部2012CB932600 纳米界面生物分子作用机制的基础研究及其在前列腺癌早期检测中的应用樊春海中科院上海应用物理研究所中国科学院2012CB932700 新型高性能半导体纳米线电子器件和量子器件徐洪起北京大学教育部2012CB932800 高比能直接甲醇燃料电池关键纳米材料与纳米结构研究杨辉上海中科高等研究院中国科学院上海市科学技术委员会2012CB932900 纳米结构材料在先进能源器件应用中的表界面问题研究王春儒中国科学院化学研究所中国科学院2012CB933000 基于扫描探针技术的纳米表征新方法研究白雪冬中国科学院物理研究所中国科学院2012CB933100 高频磁性纳米材料的电磁性能调控及其在磁性电子器件中的应用薛德胜兰州大学教育部2012CB933200 高效节能微纳结构材料体系研究杨振忠中国科学院化学研究所中国科学院2012CB933300 基于纳米技术的肺癌早期检测研究赵建龙中国科学院上海微系统与信息技术研究所中国科学院上海市科学技术委员会2012CB933400 石墨烯材料的宏量可控制备及其应用基础研究石高全清华大学教育部2012CB933500 面向高性能计算机超结点的关键微纳光电子器件及其集成技术研究郑婉华中国科学院半导体研究所中国科学院2012CB933600 多级微纳结构生物活性材料促进骨组织快速修复的研究刘昌胜华东理工大学教育部上海市科学技术委员会2012CB933700 新型铜基化合物薄膜太阳能电池相关材料和器件的关键科学问题研究肖旭东中国科学院深圳先进技术研究院中国科学院2012CB933800 仿生可控粘附纳米界面材料张广照中国科学技术大学中国科学院2012CB933900 纳米材料在骨、牙再生修复中的生物学过程研究林野北京大学教育部2012CB934000 基于肿瘤微环境调控的抗肿瘤纳米材料设计和机制研究聂广军国家纳米科学中心中国科学院2012CB934100 微纳惯性器件运动界面纳米效应基础问题研究刘晓为哈尔滨工业大学工业和信息化部2012CB934200 新型微纳结构硅材料及广谱高效太阳能电池研究李晋闽中国科学院半导体研究所中国科学院2012CB934300 基于纳米材料的太阳能光伏转换应用基础研究戴宁中国科学院上海技术物理研究所上海市科学技术委员会中国科学院2012CB944400 雌性生殖细胞减数分裂的分子基础孙青原中国科学院动物研究所国家人口和计划生育委员会中国科学院2012CB944500 心脏与肝脏发育和再生的遗传调控研究彭金荣浙江大学教育部浙江省科学技术厅2012CB944600 生殖细胞基因组结构变异的分子基础金力复旦大学上海市科学技术委员会教育部2012CB944700 排卵障碍相关疾病发生机制研究陈子江山东大学教育部山东省科学技术厅2012CB944800 植物胚乳发育及储藏物质累积的分子调控机制研究薛红卫中国科学院上海生命科学研究院上海科学技术委员会2012CB944900 辅助生殖诱发胚胎源性疾病的风险评估和机制研究黄荷凤浙江大学教育部浙江省科学技术厅2012CB945000 上皮组织的形成、更新及其调节机理朱学良中国科学院上海生命科学研究院中国科学院上海市科学技术委员会2012CB945100 血管发育和稳态维持的遗传及表观遗传机制杨晓中国人民解放军军事医学科学院生物工程研究所2012CB955200 东亚季风区年际-年代际气候变率机理与预测研究刘征宇北京大学教育部2012CB955300 全球典型干旱半干旱地区气候变化及其影响黄建平兰州大学教育部2012CB955400 全球变化与环境风险关系及其适应性范式研究史培军北京师范大学教育部2012CB955500 气候变化对人类健康的影响与适应机制研究刘起勇中国疾病预防控制中心卫生部2012CB955600 太平洋印度洋对全球变暖的响应及其对气候变化的调控作用谢尚平中国海洋大学教育部2012CB955700 气候变化对社会经济系统的影响与适应策略黄季焜中国科学院地理科学与资源研究所中国科学院2012CB955800 气候变化经济过程的复杂性机制、新型集成评估模型簇与政策模拟平台研发王铮中科院科技政策与管理科学研究所2012CB955900 全球气候变化对气候灾害的影响及区域适应研究宋连春国家气候中心中国气象局2012CB956000 全球变暖下的海洋响应及其对东亚气候和近海储碳的影响袁东亮中国科学院海洋研究所中国科学院2012CB956100 湖泊与湿地生态系统对全球变化的响应及生态恢复对策研究沈吉中国科学院南京地理与湖泊研究所中国科学院2012CB956200 全球典型干旱半干旱地区年代尺度气候变化机理及其影响研究马柱国中国科学院大气物理研究所中国科学院2012CB966300 神经分化各阶段细胞命运决定的调控网络研究及其转化应用章小清同济大学上海市科学技术委员会教育部2012CB966400 人多能干细胞向胰腺β细胞和神经细胞定向分化的机制研究邓宏魁北京大学教育部2012CB966500非整合人诱导性多能干细胞(iPS)及相关技术用于β地中海贫血治疗的研究潘光锦中科院广州生物医药与健康研究院中科院2012CB966600 中胚层干细胞自我更新分化的机制与功能研究冯新华浙江大学教育部浙江省科学技术厅2012CB966700 多能干细胞定向分化的表观遗传学调控网络沈晓骅清华大学教育部2012CB966800 干细胞分裂模式和干细胞干性维持的机制研究高维强上海交通大学教育部上海市科学技术委员会2012CB966900 体内间充质干细胞自我更新、分化及其调控相关组织干细胞的机制研究李保界上海交大教育部上海科学技术委员会2012CB967000 肿瘤干细胞的动态演进及干预研究刘强中山大学教育部2011年生命科学部资助重点项目清单。
厦门大学发现“葡萄糖感受机制”有助于治疗肥胖症

窑72窑生物学教学2017年(第42卷)第12期2017年第1耀12期总目录题目姓名页期【生物科学综述】表观遗传及表观遗传学概述..........王丽娟等2 (1)缓步动物的隐生现象................张嘉卉等7 (1)淡水水质的浮游生物评价研究进展.......袁聿军9 (1)长链非编码RNA及其生物学功能概述……李法君2 (2)痤疮发病机制及对青少年的影响.......郭焕焕等4 (2)常用鱼类麻醉剂及其作用机理研究进展…李博岩等7 (2)中波紫外辐射对两栖动物的生物学效应研究进展…….............................陈智强等2 (3)新型DNA识别受体cGAMP合成酶的发现及其功能…….............................何枝芳等6 (3)降解组测序技术及其研究进展...........李法君2 (4)中华鳖的热生物学研究进展概述.......张利双等4 (4)蜱的危害及防制措施概述..............梁裕芬2 (5)噬菌体抗体库技术概述................蔡尽忠4 (5)黑木耳多糖药理学效应研究进展.......张志强等7 (5)家族性髙胆固醇血症及其病理、诊断和治疗研究进展….............................王丽娟等2 (6)盘基网柄菌——研究胞外诱捕起源和机制的模型…….............................薛德明等4 (6)乳腺癌耐药蛋白的C421A多态性对肿瘤易感性及疗效影响研究概述..................杜静仪等6 (6)髙亲和力的人源化CD20单克隆抗体的研究进展.....[医疗卫生]厦门大学发现“葡萄糖感受机制”,有助于治疗肥胖症据2017年7月20日《科技日报》报道,《自然》杂志发表了厦门大学林圣彩教授课题组发现了生理状态下机体感受葡萄糖水平并调节代谢模式的机制,对开发用于治疗肥胖症,乃至延长寿命的药物具有深远意义。
转录调节蛋白TP53基因敲除增加斑马鱼耐寒能力与运动行为的研究

㊀收稿日期:2023-01-15基金项目:国家自然科学基金面上项目(81770165)ꎻ上海市科技兴农重点攻关项目[沪农科创字(2019)第1-2号]作者简介:贾惠莲(1996-)ꎬ女ꎬ山东德州人ꎬ硕士研究生ꎬ研究方向:分子生物学.㊀∗通讯作者:张俊芳ꎬE ̄mail:jfzhang@shou.edu.cnꎻ范纯新ꎬE ̄mail:cxfan@shou.edu.cn.㊀㊀辽宁大学学报㊀㊀㊀自然科学版第50卷㊀第4期㊀2023年JOURNALOFLIAONINGUNIVERSITYNaturalSciencesEditionVol.50㊀No.4㊀2023转录调节蛋白TP53基因敲除增加斑马鱼耐寒能力与运动行为的研究贾惠莲1ꎬ2ꎬ秦彦筠1ꎬ2ꎬ杨倩婷1ꎬ2ꎬ韩兵社1ꎬ2ꎬ范纯新1ꎬ2ꎬ∗ꎬ张俊芳1ꎬ2ꎬ∗(1.上海海洋大学水产种质资源发掘与利用教育部重点实验室ꎬ上海201306ꎻ2.上海海洋大学水产科学国家级实验教学示范中心ꎬ上海201306)摘㊀要:转录调节蛋白TP53(Tumorprotein53)调控多种重要生物学过程ꎬ包括细胞周期㊁DNA损伤㊁修复和细胞凋亡等.在各种应激条件下tp53会被激活并发挥与应激相关的调控作用ꎬ但tp53在鱼类低温应激下的作用尚不明了.本文首先检测在低温应激条件下斑马鱼(Daniorerio)中tp53的表达变化ꎬ然后利用tp53-/-斑马鱼敲除模型研究了敲除tp53对斑马鱼低温耐受能力和运动的影响.RT-qPCR结果显示:在8ħ低温条件下野生型斑马鱼全鱼和鳃组织中tp53的mRNA水平显著升高.低温耐受实验显示:8ħ低温下tp53-/-斑马鱼的耐受能力增强ꎬ半数致死时间(lethaltimeto50%mortalityꎬLT50)显著高于WT(Wildtype)对照组(P<0.0001).运动行为学实验发现:与28ħ相比ꎬ18ħ低温下WT和tp53-/-斑马鱼的移动距离㊁平均速度和活跃度均显著降低(P<0.0001)ꎻ同时发现在28ħ和18ħ条件下ꎬtp53-/-斑马鱼的移动距离㊁平均速度和活跃度均显著高于WT组.研究结果说明tp53参与斑马鱼的低温响应过程ꎬ敲除tp53后斑马鱼的耐寒能力与运动能力增强.本文为鱼类的低温耐受机制和运动行为学研究提供了新思路.关键词:斑马鱼ꎻtp53ꎻ基因敲除ꎻ低温耐受ꎻ运动能力中图分类号:S917㊀㊀㊀文献标志码:A㊀㊀㊀文章编号:1000-5846(2023)04-0348-11KnockoutofTranscriptionalRegulatoryProteinTP53IncreasesColdToleranceandAthleticAbilityofZebrafishJIAHui ̄lian1ꎬ2ꎬQINYan ̄jun1ꎬ2ꎬYANGQian ̄ting1ꎬ2ꎬHANBing ̄she1ꎬ2ꎬFANChun ̄xin1ꎬ2ꎬ∗ꎬZHANGJun ̄fang1ꎬ2ꎬ∗(1.KeyLaboratoryofExplorationandUtilizationofAquaticGeneticResourcesꎬMinistryofEducationꎬShanghaiOceanUniversityꎬShanghai201306ꎬChinaꎻ2.NationalDemonstrationCenterforExperimentalFisheriesScienceEducationꎬShanghaiOceanUniversityꎬShanghai201306ꎬChina)Abstract:㊀TranscriptionalregulatoryproteinTP53(Tumorprotein53)regulatesavarietyof㊀㊀crucialbiologicalprocessesincludingcellcycleꎬDNAdamageꎬrepairꎬapoptosisꎬandsoon.Asaregulatoryfactorꎬtp53isinducedundervariousstresses.Howeverꎬtheroleoftp53undercoldstressinfishstillremainelusive.InourstudyꎬmRNAleveloftp53inzebrafish(Daniorerio)duringcoldtreatmentat8ħwasexaminedꎬandtheeffectoftp53knockoutoncoldtoleranceandathleticabilityofzebrafishwasinvestigatedbyusingatp53knockoutmodel(tp53-/-).RT-qPCRshowedthattp53mRNAlevelinlarvaeandgillsincreasedsignificantlyaftercoldexposure.Coldtolerancetestshowedthattheknockoutoftp53hadenhancedcoldtoleranceꎬindicatedbysignificantlyincreasedLT50(lethaltimeto50%mortality)(P<0.0001)at8ħforcoldtreatment.BehaviorstestshowedthatthemoveddistanceꎬaveragespeedandmobilityweresignificantlydecreasedinbothWT(WildType)andtp53-/-zebrafishat18ħꎬcomparedwiththoseat28ħ.Howeverꎬtheathleticabilityoftp53-/-zebrafishwassignificantlyhigherthanWTzebrafishincludingmoveddistanceꎬaveragespeedandmobilityatbothtemperatures.Thisworkindicatestheinvolvementoftp53undercoldstressꎬandenhancementofcoldtoleranceandathleticabilitybytheknockoutoftp53inzebrafish.Thefindingsprovidenewinsightsforfurtherunderstandingthemechanismofcoldtoleranceandbehaviorsinfish.Keywords:㊀zebrafishꎻtp53ꎻgeneknockoutꎻcoldtoleranceꎻathleticability0㊀引言温度是影响动物生理活动的环境因子之一ꎬ研究动物应对温度变化的适应机制具有重要的生物学意义.低温是自然界普遍存在的环境压力ꎬ哺乳动物在遭受冷应激时会增加食物摄入ꎬ并提高产热机能来维持体温恒定[1].作为变温动物ꎬ鱼类更易受水体温度变化影响.抗冻(糖)蛋白㊁分子伴侣㊁代谢酶类和膜通道蛋白改变等多种机制参与了鱼类响应低温胁迫的过程[2].鱼类温度适应调控不同于恒温动物[3]ꎬ目前学界对鱼类低温胁迫的适应机制的了解比较有限.研究低温耐受相关基因对耐寒能力的调控作用ꎬ是探究鱼类低温耐受机制需要解决的问题之一.作为一种转录因子ꎬtp53可通过激活或抑制多种下游靶基因的转录来调控细胞凋亡㊁胚胎发育和应激等过程ꎬ这对维持生物体内环境稳定至关重要[4-5].近年来ꎬ越来越多的研究表明tp53在鱼类应对外界温度变化带来的压力方面发挥了重要作用.在斑马鱼(Daniorerio)中ꎬtp53高表达与低温应激相关[6].在低温胁迫下ꎬ大黄鱼p53信号通路中相关基因的表达水平升高[7].在小于40ħ的亚热激状态下ꎬ斑马鱼胚胎中的P53蛋白可通过抑制过度热激反应而维持细胞稳态ꎬ促进细胞存活ꎻ但在43ħ的高热激状态下ꎬ磷酸化的P53可促进凋亡蛋白Bax(B-celllymphoma-2AssociatedXproteinꎬBAX)的表达ꎬ诱导细胞凋亡[8].这说明P53在不同的压力状态下可以通过不同的调控机制对细胞产生影响.温度会影响鱼类的游泳㊁摄食和生殖洄游等行为ꎬ由于温度变化导致鱼类行为发生改变时ꎬ游泳943㊀第4期㊀㊀㊀㊀贾惠莲ꎬ等:转录调节蛋白TP53基因敲除增加斑马鱼耐寒能力与运动行为的研究㊀㊀行为就成为鱼类此时最重要的环境适应指标[9].运动行为改变会导致tp53的表达在不同组织细胞中出现差异.丁树哲等[10]发现ꎬ运动训练后小鼠骨骼肌中tp53的表达具有肌纤维类型特异性和运动强度敏感性.对tp53与运动关系的研究中ꎬElabd等[11]敲除斑马鱼tp53基因ꎬ4dpf和5dpf的斑马鱼对振动的反应能力下降ꎬ逃避能力降低.DUX4(Doublehomeobox4)基因过度表达不利于斑马鱼肌肉的发育ꎬ在小鼠中可导致肌肉退化ꎬ研究表明DUX4对肌肉的影响具有tp53依赖性[12].目前已有研究均提示ꎬtp53基因与鱼类在极端环境下的存活情况相关ꎬ且在不同环境下的调控作用有所差异[8ꎬ13].以往研究中ꎬ通过Morpholino法敲降斑马鱼胚胎在特异性㊁作用持续时间和敲降效率方面存在一定限制[14].本文采用由CRISPR/Cas9(Clusteredregularlyinterspacedshortpalindromicrepeats/CRISPR-associatedprotein9ꎬCRISPR/Cas9)基因编辑技术获得的tp53基因敲除斑马鱼ꎬ探究tp53对斑马鱼耐寒能力的影响ꎬ发现tp53基因敲除增加了斑马鱼的低温耐受能力和运动能力.本文为进一步研究鱼类耐寒机制和运动行为调节机制提供了依据.1㊀材料与试剂1.1㊀主要试剂本文的主要试剂:2ˑTaqMasterMix(P111)购于Vazymeꎻ1MTris-HClꎬpH8.0ST780购于BeyotimeꎻTRIzolReagent(15596018)购于Ambionꎻ反转录试剂盒(RR047A)购于TaKaRaꎻSYBRGreen(1725124)购于BIO-RAD.1.2㊀斑马鱼来源本文所用WT斑马鱼为AB品系ꎬF1代tp53基因敲除杂合子斑马鱼tp53+/-由上海海洋大学水产与生命学院范纯新老师课题组赠予:由CRISPR/Cas9基因编辑技术获得F0代tp53基因敲除型斑马鱼ꎬF0代与野生型斑马鱼交配获得F1代tp53+/-杂合子斑马鱼.本实验将F1代tp53+/-进行自交ꎬ筛选F2代中的tp53-/-纯合子斑马鱼ꎬ为排除母体效应的影响ꎬ将筛选出的F2代tp53-/-进行自交以获得F3代tp53-/-ꎬF3代tp53-/-用于实验.斑马鱼的敲除靶点位于tp53基因的第五外显子ꎬ缺失的8个碱基为5ᶄ-CCCTCCAC-3ᶄ.所有实验用鱼均养殖在专用养殖循环水系统中ꎬ斑马鱼传代以人工成对交配的方式进行ꎬ养殖水温控为28ħꎻ通过自动光照系统控制光照周期ꎬ光周期为7:30 21:30ꎬ暗周期为21:30 次日7:30ꎻ以新鲜孵化的丰年虾和草履虫对斑马鱼进行投喂.1.3㊀仪器与设备本文所使用的主要仪器:PCR核酸扩增仪(C1000TouchThermalCycle)购于BIO-RADꎻ斑马鱼胚胎孵化箱(MIR-154-PC)购于Panasonicꎻ恒温培养箱(1000B1)购于江南仪表厂(宁波)ꎻ荧光定量PCR仪(CFX96)购于BIO-RADꎻ12孔板(F603202-9001)购于BBIꎻ斑马鱼幼鱼行为轨迹分析系统(Daniovision)购于Noldusꎻ斑马鱼专用养殖循环水系统为上海海圣水族养殖设备.053㊀㊀㊀辽宁大学学报㊀㊀自然科学版2023年㊀㊀㊀㊀2㊀实验方法2.1㊀低温处理降温方案本文采用的梯度降温方案(如图1所示):水温以0.83ħ/h的速度从28ħ匀速降至18ħꎬ水温在18ħ维持12h后ꎬ再以1ħ/h的速度匀速降至12ħꎬ在12ħ维持12h后ꎬ继续以1ħ/h的速度匀速降温至8ħꎬ此后不再降温.在整个降温过程中停止对斑马鱼投喂食物ꎬ避免因食物残留导致水质破坏影响实验结果.图1㊀斑马鱼低温处理降温方案2.2㊀实时荧光定量PCR检测tp53表达量本文按照图1对斑马鱼进行降温处理ꎬ斑马鱼全鱼和鱼鳃的取样点分别为28ħꎬ8ħ-0㊁2㊁4和6h.在每个取样点收取3尾全鱼(2月龄)样本混样ꎬ收取6尾斑马鱼(3.5月龄)鳃组织样本混样ꎬ用于提取RNAꎬRNA提取采用TRIZOL法.本文提取总RNA后ꎬ取1μg总RNA进行反转录ꎬ以获得的cDNA作为模板ꎬ以斑马鱼β-actin作为内参基因ꎬ通过RT-qPCR法检测tp53基因的mRNA表达量.引物设计在IDT网站(https://sg.idtdna.com)完成.RT-qPCR引物需要跨越内含子进行设计ꎬ产物长度在100~200bp之间ꎬ引物序列见表1.RT-qPCR程序:预变性95ħ3minꎬ1个循环ꎻ变性95ħ10sꎬ退火60ħ30sꎬ39个循环.每个样品设置3个生物学重复ꎬ采用2-әәCt运算方法进行数据分析.表1㊀RT-qPCR和基因型鉴定引物序列PrimernamePrimersequence(5ᶄң3ᶄ)Size/bpRT-β-actin-FCTTTGAGCAGGAGATGGGAACC22RT-β-actin-RGATTCCATACCCAGGAAGG19RT-tp53-FCCCGGCGATCATGGATTTAG20RT-tp53-RCCACATGCTCGGACTTCTTATAG23tp53-FGCTAAACTATAACAACTGGGTGAAA25tp53-RAAATGTCTGTACTATCTCCATCCG242.3㊀F2代斑马鱼基因分型鉴定本文将F1代杂合子斑马鱼进行自交ꎬ获得F2代斑马鱼ꎬ剪取F2代斑马鱼尾鳍ꎬ加入20μL50mmolNaOH溶液ꎬ95ħ裂解20minꎬ涡旋振荡5sꎬ加入2μL1molTris-HCl(pH8.0)ꎬ振荡混匀ꎬ即获得DNA模板ꎬ在靶点上下游设计引物进行PCR扩增ꎬ基因型鉴定引物见表1.PCR反应程序:94ħ预变性5minꎻ94ħ变性30sꎬ55ħ退火15sꎬ72ħ延伸1minꎬ34个循环ꎻ72ħ再延伸5minꎬ扩增产物约240bp.PCR产物进行3%琼脂糖凝胶电泳ꎬ并通过Sanger测序验证ꎬ筛选出F2代敲除型纯合子tp53-/-斑马鱼.本文提取tp53-/-斑马鱼的总RNAꎬ以WT作为对照ꎬ通过RT-qPCR分析tp53基因的mRNA表达量ꎬ引物㊁样品处理和数据分析方法同2.2.2.4㊀低温耐受实验本实验将2.5月龄WT(42尾)和tp53-/-(50尾)斑马鱼进行低温处理(方案见图1)ꎬ统计斑马153㊀第4期㊀㊀㊀㊀贾惠莲ꎬ等:转录调节蛋白TP53基因敲除增加斑马鱼耐寒能力与运动行为的研究鱼的生存时长和半数致死时间(Lethaltimeto50%mortalityꎬLT50).半数致死时间按8ħ冷处理开始至每组斑马鱼死亡数量达到一半时的时间来计算.斑马鱼在低温下会逐渐失去平衡ꎬ不再游动.参考相关文献对鱼类冷处理后的恢复方法[15]ꎬ每间隔0.5hꎬ将不再游动的斑马鱼转移到28ħ养殖水中ꎬ观察其能否恢复ꎬ若不能恢复ꎬ则判定斑马鱼死亡.实验中记录每一尾斑马鱼的生存时长(实验开始降温的时刻记为0)ꎬ绘制生存曲线ꎬ并测量每一尾斑马鱼的体重和体长.2.5㊀运动行为学监测本实验随机选取生长状况良好的1月龄WT和tp53-/-斑马鱼幼鱼转移至12孔板中ꎬ每孔1尾ꎬ每组6尾ꎬ每孔加入4mL养殖水ꎬ并做好标记.本文采用Noldus公司的DanioVision斑马鱼行为轨迹分析系统对斑马鱼进行运动行为学监测ꎬ先将斑马鱼转移至暗室中适应15minꎬ以消除应激反应对实验造成的干扰ꎬ再进行数据采集ꎬ采集过程中ꎬ周围环境保持安静ꎬ以排除噪音对实验的干扰ꎬ先对28ħ条件下斑马鱼的行为活动进行监测ꎬ然后调节温控系统将水温直接降至18ħꎬ监测斑马鱼在18ħ条件下的行为活动ꎬ导出斑马鱼的运动轨迹图和热图ꎬ导出数据并分析斑马鱼的移动距离㊁平均速度㊁最大加速度和活跃度.单次实验监测时长为10minꎬ每个温度条件设置3组平行实验.2.6㊀数据分析本实验在GraphPadPrism8软件中进行数据分析及作图:采用T-test法分析独立样本显著差异性ꎬ采用Log-rank(Mantel-Cox)test法分析生存曲线差异ꎬ采用MannWhitneytest法分析半数致死时间差异.P<0.05表示差异具有统计学意义.3㊀结果与分析3.1㊀低温下斑马鱼tp53表达水平变化为探究低温处理后WT斑马鱼tp53基因的表达变化ꎬ本实验通过RT-qPCR法检测了样品中tp53的mRNA表达水平ꎬ结果如图2所示.低温处理后全鱼和鳃中tp53的mRNA表达水平均显著升高ꎬ说明低温处理诱导了tp53的转录.图2㊀低温胁迫下WT斑马鱼中tp53基因的mRNA表达注:T检验ꎬ∗P<0.05ꎬ差异显著ꎻ∗∗P<0.01ꎬ差异非常显著ꎻ∗∗∗∗P<0.0001ꎬ差异极其显著.253㊀㊀㊀辽宁大学学报㊀㊀自然科学版2023年㊀㊀㊀㊀㊀㊀3.2㊀F2代斑马鱼基因分型筛选为了对F2代斑马鱼进行快速筛选ꎬ本实验剪取F2代斑马鱼尾鳍ꎬ在敲除靶点上游和下游分别设计引物进行PCR扩增.各基因型对应的琼脂糖凝胶电泳图如图3(a)所示ꎬ泳道1为tp53-/-ꎬ泳道2为tp53+/-ꎬ泳道3为tp53+/+.为进一步确认筛选个体基因型正确ꎬ通过测序法对F2代进行鉴定ꎬ序列比对如图3(b d)所示ꎬ结果表明成功筛选出了缺失8bp的F2代纯合子突变体斑马鱼tp53-/-.为了检测斑马鱼中tp53基因的表达ꎬ本实验对tp53-/-和WT斑马鱼的tp53表达量进行RT-qPCR检测ꎬ结果如图3(e)所示ꎬ与WT相比ꎬtp53-/-斑马鱼的tp53基因表达水平显著降低(P<0 001)ꎬ具有统计学差异.从图3(f)看出ꎬ缺失8个碱基导致移码突变ꎬ翻译过程提前终止.图3㊀对F2代tp53基因敲除斑马鱼进行筛选注:T检验ꎬ∗∗∗P<0.001ꎬ差异十分显著.3.3㊀敲除tp53基因对斑马鱼低温耐受能力的影响为了探究敲除tp53基因对斑马鱼低温耐受能力的影响ꎬ本实验对WT和tp53-/-斑马鱼进行低温处理ꎬ分别统计这两种基因型斑马鱼的生存时长ꎬ绘制生存曲线如图4(a)所示.结果显示tp53-/-斑马鱼在8ħ低温下的生存率比WT显著增加(P<0.0001).图4(b)中ꎬWT斑马鱼的LT50为353㊀第4期㊀㊀㊀㊀贾惠莲ꎬ等:转录调节蛋白TP53基因敲除增加斑马鱼耐寒能力与运动行为的研究㊀㊀4hꎬtp53-/-斑马鱼的LT50为24.5hꎬ敲除tp53基因后斑马鱼的LT50显著增加(P<0.0001).本实验分别测量这两种基因型斑马鱼的体重和体长ꎬ数据分析表明体重㊁体长与斑马鱼的生存时长不具有相关性(图4(c d)).显著性差异分析显示体长和体重均不具有显著性差异(图4(e f)).以上结果说明ꎬ敲除tp53基因后ꎬ斑马鱼的低温耐受能力增强.图4㊀斑马鱼低温耐受实验注:nsꎬ没有显著性差异.3.4㊀敲除tp53基因对斑马鱼运动行为的影响为了探究敲除tp53基因是否会影响斑马鱼的运动能力ꎬ本文利用斑马鱼行为轨迹分析系统在28ħ和18ħ条件下对1月龄WT和tp53-/-斑马鱼进行运动行为学监测ꎬ针对斑马鱼的移动距离(图5(a))㊁平均速度(图5(b))㊁最大加速度(图5(c))和活跃度(图5(d))进行统计学分析.数据分析发现:在28ħ条件下ꎬtp53-/-斑马鱼的移动距离㊁平均速度和活跃度均比WT斑马鱼显著升高(P<0.0001)ꎬ最大加速度没有发生显著变化.在18ħ低温条件下WT和tp53-/-斑马鱼的移动距离㊁平均速度和活跃度均显著降低(P<0.0001)ꎬWT斑马鱼的最大加速度没有发生显著变化ꎬ但tp53-/-斑马鱼的最大加速度显著降低(P<0.05).在18ħ条件下ꎬ与WT相比ꎬtp53-/-斑马鱼的移动距离(P<0.0001)㊁平均速度(P<0.0001)和活跃度(P<0.001)显著升高ꎬ最大加速度没有发生显著变化.453㊀㊀㊀辽宁大学学报㊀㊀自然科学版2023年㊀㊀㊀图5㊀WT与tp53-/-斑马鱼的运动能力监测注:T检验ꎬ∗P<0.05ꎬ差异显著ꎻ∗∗∗P<0.001ꎬ差异十分显著ꎻ∗∗∗∗P<0.0001ꎬ差异极其显著.4㊀讨论与结论已有研究发现ꎬ经18ħ低温驯化处理后ꎬ甲基化DNA免疫共沉淀测序(MethylatedDNAimmunoprecipitationsequencingꎬMeDIP-seq)结合qPCR分析结果显示ꎬ斑马鱼胚胎成纤维细胞ZF4中tp53表达水平升高[16].低温下斑马鱼和罗非鱼RPL11/MDM2/P53通路中P53㊁核糖体蛋白(ribosomalproteinsꎬRPs)RPL11和mdm2(murinedoubleminutes2)㊁bad(BCL2associatedagonistofcelldeath)的表达水平上调[17].大黄鱼肝脏㊁肌肉㊁脑㊁脾脏㊁鳃㊁肾㊁肠和心脏中tp53的mRNA表达水平在14ħ低温处理一定时间后也显著上调[7].本研究中WT斑马鱼全鱼和鳃中tp53的mRNA表达水平在8ħ低温处理后显著升高ꎬ与已有研究结果一致.上述研究均提示ꎬtp53在鱼类抵抗外界低温胁迫方面发挥了重要作用ꎬ具体调控作用还需深入探究.为排除母体干扰ꎬ本文在得到F2代tp53-/-斑马鱼后ꎬ将F2代中tp53-/-斑马鱼自交ꎬ获得F3代tp53-/-斑马鱼进行实验研究.线粒体主要来自母体提供的卵细胞ꎬ属于母系遗传.已有研究发现ꎬP53参与了线粒体相关生物学过程的调控ꎬ包括线粒体介导的凋亡㊁自噬和活性氧生成等过程[18].本文对WT和tp53-/-斑马鱼进行低温处理ꎬ在8ħ低温下ꎬWT斑马鱼LT50为4hꎬtp53-/-斑马鱼553㊀第4期㊀㊀㊀㊀贾惠莲ꎬ等:转录调节蛋白TP53基因敲除增加斑马鱼耐寒能力与运动行为的研究㊀㊀㊀LT50为24.5hꎬ敲除tp53基因后斑马鱼在8ħ低温下的LT50增加ꎬ低温耐受能力增强ꎬ斑马鱼的生存时长没有受到体重和体长的影响.鱼类在低温下运动能力减弱ꎬ持续暴露于低温下身体会逐渐失去平衡ꎬ最终死亡[19].在低温耐受实验中观察到ꎬ当温度降至12ħ时ꎬWT斑马鱼可保持身体平衡ꎬ但伏在水底ꎻtp53-/-斑马鱼仍可游动ꎬ但活动迟缓.基于此现象ꎬ本文探究了敲除tp53基因对斑马鱼游泳行为的影响ꎬ在28ħ和18ħ条件下ꎬ与WT相比ꎬtp53-/-斑马鱼的移动距离㊁平均速度和活跃度均显著升高ꎬtp53基因敲除后ꎬ斑马鱼运动能力增强ꎻ当温度降至18ħ后ꎬ与28ħ相比ꎬ两种基因型斑马鱼的移动距离㊁平均速度和活跃度均显著降低ꎬ运动能力减弱.斑马鱼的简单运动行为学分析包括对逃避反应和游泳行为的研究ꎬ对斑马鱼逃避行为研究较多的是听觉刺激诱发的快速逃跑行为[20]ꎬ而游泳行为是斑马鱼自发的运动行为.当外界环境温度发生变化时ꎬ包括鱼类在内的绝大多数物种可通过热调控行为(如生境温度选择行为㊁游泳行为㊁摄食行为和生殖洄游行为等)或偏好来应对环境压力ꎬ其中游泳行为是最重要的环境适应指标[9].在tp53与运动能力调节的研究中发现ꎬ运动会影响tp53基因的表达ꎬ这与诱发应激的刺激类型㊁刺激强度和持续时间相关[21].田振军等[22]发现对大鼠进行为期8周的有氧跑台训练会显著降低tp53的表达.本文中ꎬ缺失tp53基因后ꎬ在避免外界光线和声音刺激的条件下ꎬ斑马鱼的运动能力增强ꎬ说明运动能力的增强与tp53基因的表达水平降低相关.神经系统负责接收和处理各种信息ꎬ并通过对周围环境和机体状态进行评估来调控机体行为以适应生存环境[23].在对脊髓性肌萎缩症(SpinalmuscularatrophyꎬSMA)小鼠模型的研究中ꎬP53蛋白积累会抑制运动神经元和脊髓中间神经元的功能ꎬ这可能是导致小鼠出现肌无力和肌萎缩症状的原因之一[24].tp53基因敲除的小鼠可以抵抗神经元退化ꎬ缺失tp53基因还能增加由诱导多能干细胞分化而来的运动神经元的存活率[25].Amson等[26]在圆形旷场实验中发现ꎬtp53基因敲除小鼠会出现焦虑样行为ꎬ而Campana等[27]在高架十字迷宫中的研究结果与此相反.焦翠等[28]在方形旷场实验中发现ꎬtp53基因缺失不会导致小鼠出现焦虑样行为ꎬ也不影响其自发运动能力ꎬ这与Campana等[27]观点一致ꎬ这表明开阔旷场的形状可能对测试结果有一定影响.上述研究结果显示ꎬtp53与肌肉运动和神经系统之间存在一定联系ꎬ详尽的作用机制有待进一步阐明.Hu等[29]发现随着低温暴露时间延长ꎬ在罗非鱼和斑马鱼鳃中的凋亡信号逐渐增强.在应激条件下ꎬP53过度积累激活凋亡相关基因表达ꎬ这可能会造成组织发生不可逆损伤ꎬ进而导致鱼类死亡[17].野生型tp53的表达会增加缺失tp53的小鼠成纤维细胞对44ħ高温的热敏感性ꎬ这可能是由于P53依赖的细胞凋亡被激活所导致的[30].Langheinrich等[31]发现与野生型相比ꎬtp53基因敲除斑马鱼胚胎经紫外线处理后凋亡水平减弱ꎬ这与P53通过调节P21表达促进细胞凋亡这一过程被阻断相关.当面临生存压力时ꎬ适度的自噬有利于细胞存活[32].对人㊁小鼠和线虫的研究中ꎬ将tp53进行敲除㊁敲降或使用抑制剂抑制tp53表达ꎬ都能诱发自噬ꎬ这提高了细胞在缺乏氧气和营养物质条件下的存活率[33].将ZF4细胞[34]和HeLa细胞[35]在10ħ低温短期处理后自噬增强ꎬ使用BafilomycinA1阻断细胞自噬后ꎬ细胞存活率降低.对小鼠脑卒中模型和原代神经元体外氧糖剥夺模型进行处理ꎬ结果均显示tp53缺失后神经元的凋亡和自噬水平较低ꎬP53表达被抑制后ꎬ通过p53/PRAS40/mTOR途径保护脑卒中后的神经元免受损伤ꎬ该途径调节的神经元自噬和凋亡过程可能是这种保护作用的潜在机制[36].在低温应激下tp53表达上调可能引发机体损伤进而导致斑马鱼死亡.敲除tp53基因后斑马鱼耐寒能力增强ꎬ在同等低温处理时长下ꎬ由tp53导致的机体损伤减弱ꎬ这种损伤可能与tp53调节的细胞凋亡和自噬过程相关.综合本文和前人研究结果ꎬtp53可以促进细胞凋亡发生ꎬ同时653㊀㊀㊀辽宁大学学报㊀㊀自然科学版2023年㊀㊀㊀㊀抑制抗凋亡基因的转录ꎬ过量的P53积累加速斑马鱼在低温下的组织损伤进程ꎬ这可能是导致鱼类在低温胁迫下死亡的原因.缺失tp53后ꎬ斑马鱼对低温的耐受能力增强ꎬ这提示tp53对斑马鱼低温应激可能发挥负调控作用ꎬ涉及的具体调节机制还需深入研究.本文验证了在斑马鱼中tp53基因与低温耐受能力和运动能力相关.斑马鱼在经受低温胁迫后ꎬtp53表达水平上调ꎻ敲除tp53基因后斑马鱼的低温耐受能力和运动能力增强.这为进一步研究鱼类低温适应和运动行为的调控机制提供了研究基础.参考文献:[1]㊀LuDLꎬMaQꎬWangJꎬetal.Fastingenhancescoldresistanceinfishthroughstimulatinglipidcatabolismandautophagy[J].JournalofPhysiologyꎬ2019ꎬ597(6):1585-1603.[2]㊀刘丽丽ꎬ朱华ꎬ闫艳春ꎬ等.鱼类低温耐受机制与功能基因研究进展[J].生物技术通报ꎬ2018ꎬ34(8):50-57.[3]㊀SoyanoKꎬMushirobiraY.Themechanismoflow ̄temperaturetoleranceinfish[J].AdvancesinExperimentalMedicineandBiologyꎬ2018ꎬ1081:149-164.[4]㊀LevineAJ.p53:800millionyearsofevolutionand40yearsofdiscovery[J].NatureReviewsCancerꎬ2020ꎬ20(8):471-480.[5]㊀StorerNYꎬZonLI.Zebrafishmodelsofp53functions[J].ColdSpringHarborPerspectivesinBiologyꎬ2010ꎬ2(8):a001123.[6]㊀LongYꎬLiLCꎬLiQꎬetal.Transcriptomiccharacterizationoftemperaturestressresponsesinlarvalzebrafish[J].PLoSOneꎬ2012ꎬ7(5):e37209.[7]㊀QianBYꎬQiXꎬBaiYꎬetal.Thep53signalingpathwayofthelargeyellowcroaker(Larimichthyscrocea)respondstoacutecoldstress:Evidenceviaspatiotemporalexpressionanalysisofp53ꎬp21ꎬMDM2ꎬIGF-1ꎬGadd45ꎬFasꎬandAkt[J].PeerJꎬ2020ꎬ8:e10532.[8]㊀GongLꎬZhangQHꎬPanXꎬetal.p53protectscellsfromdeathattheheatstrokethresholdtemperature[J].CellReportsꎬ2019ꎬ29(11):3693-3707.[9]㊀吕为群ꎬ袁明哲.温度变化对鱼类行为影响的研究进展[J].上海海洋大学学报ꎬ2017ꎬ26(6):828-835.[10]㊀丁树哲ꎬ陈彩珍ꎬ漆正堂ꎬ等.不同强度训练对大鼠骨骼肌p53和细胞色素氧化酶I亚基基因和蛋白表达的影响[J].中国运动医学杂志ꎬ2008ꎬ27(4):454-457.[11]㊀ElabdSꎬJabeenNAꎬGerberVꎬetal.Delayindevelopmentandbehaviouralabnormalitiesintheabsenceofp53inzebrafish[J].PLoSOneꎬ2019ꎬ14(7):e0220069.[12]㊀WallaceLMꎬGarwickSEꎬMeiWYꎬetal.DUX4ꎬacandidategeneforfacioscapulohumeralmusculardystrophyꎬcausesp53 ̄dependentmyopathyinvivo[J].AnnalsofNeurologyꎬ2011ꎬ69(3):540-552.[13]㊀DuffyKTꎬWickstromE.Zebrafishtp53knockdownextendsthesurvivalofirradiatedzebrafishembryosmoreeffectivelythanthep53inhibitorpifithrin ̄α[J].CancerBiologyTherapyꎬ2007ꎬ6(5):675-678.[14]㊀张美玲ꎬ郝鑫ꎬ周成杰ꎬ等.哺乳动物雌性生殖细胞及早期胚胎基因敲除或敲减的方法[J].生理学报ꎬ2020ꎬ72(1):31-47.[15]㊀王金凤.低温胁迫诱导罗非鱼和斑马鱼鱼鳃差异凋亡的分子机制[D].上海:上海海洋大学ꎬ2016.[16]㊀HanBSꎬLiWHꎬChenZZꎬetal.VariationofDNAmethylomeofzebrafishcellsundercoldpressure[J].PLoSOneꎬ2016ꎬ11(8):e0160358.[17]㊀刘明丽ꎬ杨文意ꎬ王金凤ꎬ等.低温胁迫下鱼类鳃中RPL11/MDM2/P53信号通路相关基因及蛋白表达差异分析[J].大连海洋大学学报ꎬ2021ꎬ36(1):51-56.753㊀第4期㊀㊀㊀㊀贾惠莲ꎬ等:转录调节蛋白TP53基因敲除增加斑马鱼耐寒能力与运动行为的研究㊀㊀[18]㊀LiuYQꎬGuW.Thecomplexityofp53 ̄mediatedmetabolicregulationintumorsuppression[J].SeminarsinCancerBiologyꎬ2022ꎬ85:4-32.[19]㊀FuimanLAꎬBattyRS.Whatadragitisgettingcold:Partitioningthephysicalandphysiologicaleffectsoftemperatureonfishswimming[J].JournalofExperimentalBiologyꎬ1997ꎬ200:1745-1755.[20]㊀TranSꎬNowickiMꎬMuraleetkaranAꎬetal.Neurochemicalfactorsunderlyingindividualdifferencesinlocomotoractivityandanxiety ̄likebehavioralresponsesinzebrafish[J].ProgressinNeuro ̄psychopharmacologyBiologicalPsychiatryꎬ2015ꎬ65:25-33.[21]㊀BeyfussKꎬHoodDA.Asystematicreviewofp53regulationofoxidativestressinskeletalmuscle[J].RedoxReportꎬ2018ꎬ23(1):100-117.[22]㊀田振军ꎬ蔡梦昕ꎬ邢维新.有氧运动对心肌细胞增殖/凋亡的影响及其机制探讨[J].体育科学ꎬ2012ꎬ32(3):60-66.[23]㊀尚春峰ꎬ穆宇ꎬ杜久林.以透明脑ꎬ观澄明心 斑马鱼神经功能研究进展[J].中国科学:生命科学ꎬ2015ꎬ45(3):223-236.[24]㊀SimonCMꎬDaiYꎬvanAlstyneMꎬetal.Convergingmechanismsofp53activationdrivemotorneurondegenerationinspinalmuscularatrophy[J].CellReportsꎬ2017ꎬ21(13):3767-3780.[25]㊀Maor ̄nofMꎬShiponyZꎬLopez ̄gonzalezRꎬetal.p53isacentralregulatordrivingneurodegenerationcausedbyC9orf72poly(PR)[J].Cellꎬ2021ꎬ184(3):689-708.[26]㊀AmsonRꎬLassalleJMꎬHalleyHꎬetal.Behavioralalterationsassociatedwithapoptosisanddown ̄regulationofpresenilin1inthebrainsofp53 ̄deficientmice[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmericaꎬ2000ꎬ97(10):5346-5350.[27]㊀CampanaALꎬRondi ̄reigLꎬTobinCꎬetal.p53inactivationleadstoimpairedmotorsynchronizationinmice[J].EuropeanJournalofNeuroscienceꎬ2003ꎬ17(10):2135-2146.[28]㊀焦翠ꎬ王俭妹ꎬ周雪颖ꎬ等.p53基因敲除对小鼠神经行为及桶状皮层神经元数目的影响[J].中国实验动物学报ꎬ2021ꎬ29(1):35-41.[29]㊀HuPꎬLiuMLꎬLiuYMꎬetal.Transcriptomecomparisonrevealsageneticnetworkregulatingthelowertemperaturelimitinfish[J].ScientificReportsꎬ2016ꎬ6:28952.[30]㊀MatsumotoHꎬTakahashiAꎬWangXꎬetal.Transfectionofp53 ̄knockoutmousefibroblastswithwild ̄typep53increasesthethermosensitivityandstimulatesapoptosisinducedbyheatstress[J].InternationalJournalofRadiationOncologyBiologyPhysicsꎬ1997ꎬ38(5):1089-1095.[31]㊀LangheinrichUꎬHennenEꎬStottGꎬetal.Zebrafishasamodelorganismfortheidentificationandcharacterizationofdrugsandgenesaffectingp53signaling[J].CurrentBiologyꎬ2002ꎬ12(23):2023-2028.[32]㊀WhiteE.Autophagyandp53[J].ColdSpringHarborPerspectivesinMedicineꎬ2016ꎬ6(4):a026120.[33]㊀TasdemirEꎬMaiuriMCꎬGalluzziLꎬetal.Regulationofautophagybycytoplasmicp53[J].NatureCellBiologyꎬ2008ꎬ10(6):676-687.[34]㊀胡春兰.自噬在斑马鱼细胞冷应激过程中的作用机制研究[D].上海:上海海洋大学ꎬ2017.[35]㊀胡春兰ꎬ陈良标ꎬ胡成枫ꎬ等.自噬在细胞冷应激中的作用研究[J].中国细胞生物学学报ꎬ2016ꎬ38(9):1077-1083.[36]㊀ZhaoJLꎬDongYHꎬChenXYꎬetal.p53inhibitionprotectsagainstneuronalIschemia/ReperfusionInjurybythep53/PRAS40/mTORPathway[J].OxidativeMedicineandCellularLongevityꎬ2021ꎬ2021:4729465.(责任编辑㊀李㊀超)853㊀㊀㊀辽宁大学学报㊀㊀自然科学版2023年㊀㊀。
我国科学家揭示“浪费基因”的作用机制

我国科学家揭示“浪费基因”的作用机制
佚名
【期刊名称】《新疆交通运输科技》
【年(卷),期】2018(000)003
【摘要】在国家重点研发计划“蛋白质机器与生命过程调控”重点专项的支持下,厦门大学林圣彩教授和林舒勇副教授团队揭示了“浪费基因”编码蛋白AIDA限制肠道吸收膳食脂肪进而抵御肥胖的分子机制。
人类遗传学家James V.Neel于1962年首次提出了“节俭基因”的概念,认为人类生理系统筛选出了“节俭基因”,可以让远古人类在食物富足的短暂时期中快速增肥,以应对随时将到来的食物缺乏时期。
【总页数】1页(P21-21)
【正文语种】中文
【中图分类】Q255
【相关文献】
1.我国科学家首次揭示西瓜与甜瓜"甜蜜基因" [J],
2.我国科学家首次揭示西瓜甜瓜“甜蜜基因” [J],
3.我国科学家首次揭示西瓜甜瓜“甜蜜基因” [J],
4.我国科学家揭示小麦重大品种基因组演变规律 [J],
5.我国科学家揭示“节俭基因”的作用机制 [J],
因版权原因,仅展示原文概要,查看原文内容请购买。
219316076_植物多酚在消减牛乳蛋白致敏性作用中的研究进展

农丽艳,唐道邦,刘学铭,等. 植物多酚在消减牛乳蛋白致敏性作用中的研究进展[J]. 食品工业科技,2023,44(12):422−429. doi:10.13386/j.issn1002-0306.2022080301NONG Liyan, TANG Daobang, LIU Xueming, et al. Research Progress of Polyphenols in Reducing Lactoprotein Sensitization[J].Science and Technology of Food Industry, 2023, 44(12): 422−429. (in Chinese with English abstract). doi: 10.13386/j.issn1002-0306.2022080301· 专题综述 ·植物多酚在消减牛乳蛋白致敏性作用中的研究进展农丽艳1,2,唐道邦1,刘学铭1,王旭苹1,程镜蓉1,林耀盛1,邹金浩1,杨怀谷1,*(1.广东省农业科学院蚕业与农产品加工研究所,农业农村部功能食品重点实验室,广东省农产品加工重点实验室,广东广州 510610;2.广东海洋大学食品科技学院,广东湛江 524088)摘 要:牛乳蛋白过敏是主要由IgE 介导的病理性免疫反应,在易感体质婴幼儿群体中有较高的发生风险。
当前消除牛乳蛋白致敏性的技术手段包括热处理、高压处理、酶水解、糖基化修饰等,但这些方法可能会降低蛋白的营养功能价值、或产生不良风味。
植物多酚作为一种来源广泛的植物次级代谢产物,具有抗衰老、抗菌、抗炎和抗氧化等多重生物学活性。
多酚对蛋白质等大分子物质有显著的亲和作用,能通过共价和非共价结合方式有效降低不同食物蛋白的致敏性。
本文主要阐述酚-蛋白互作关系对牛乳致敏性的影响及其相关作用机制,并提出研究设想,为新型低致敏乳制品的开发工作提供思路和理论参考。
脂滴——细胞脂类代谢的细胞器

3H@<AEBA?@<6 EJDBHAE1E
SAIK?GDA6 K1@OADAE1E
T?GFF1CU1DO
3H@<AEBA?@<6 H@IA@EBGE1E
图 ;9 9 脂滴的结构与功能
脂滴由极性单分子层 9如磷脂和胆固醇 :、非极性核心 9如胆固醇酯和甘油
三酯:,以及表面的许多功能蛋白构成。脂滴表面蛋白研究表明,脂滴可能参与细胞的脂质合成与代 谢、物质的运输及贮存,以及细胞信号传导过程 6?D4;9 9 +@EFG@FE79 H=A9 IF=G@?/=J9 /I9 0?K?A9 AE/K07@J6 6 ;6 <1=1>6 >?@=<AB6 C@DE1EBE6 @F6 G6 =H@E=H@<1=1>6 I@D@<GJA?6 BHGB6 E2??@2D>E6 G6 HJ>?@=H@K1C6 C@?AL 6 MACADB6 EB2>J6 ?ANAG<E6 BHGB6 <1=1>6 >?@=<ABE6 IGJ6 KA6 G6 >JDGI1C6 @?OGDA<<A6 1D6 IABGK@<1EIP6 B?GDE=@?B6 GD>6 E1ODG<6 B?GDE>2CB1@D
Q1C@EGD@1>6 OADA?GB1@D
R1ODG<6 B?GDE>2CB1@D R1ODG<6 =?@BA1DE
RBA?@1>@OADAE1E
01=1>6 EJDBHAE1EVSABGK@<1EI ADA?OJ6 KG<GDCA
3H@<AEBA?@< EJDBHABGEA
基于亚组分析探讨糖脂代谢对甲状腺结节的影响

中国临床保健杂志 2024年2月第27卷第1期 ChinJClinHealthc,February2024,Vol.27,No.1·论著·基于亚组分析探讨糖脂代谢对甲状腺结节的影响魏灵荣1,王蔚陆2,王屹3,高冰1,姜天乐1,云虹11.北京大学医院检验科,北京100871;2.北京中医药大学;3.北京博爱医院检验科[摘要] 目的 将体检者划分不同亚组,阐释糖脂代谢对甲状腺结节(TN)的影响。
方法 选取2018年1月至2023年10月在北京大学医院体检的有甲状腺B超检查和总胆固醇(TC)、三酰甘油(TG)、高密度脂蛋白胆固醇(HDL C)、低密度脂蛋白胆固醇(LDL C)、血葡萄糖(GLu)资料的人群。
首先比较TN阳性和甲状腺未见明显异常者的糖脂代谢指标,结合性别、年龄、体重指数(BMI)、血压(BP)划分亚组,比较不同年龄组性别比,计算各亚组在总人群中的占比和TN检出率,选取基数大、检出率具有差异的典型亚组。
然后将同龄、同性别、BMI、BP均正常的TN者作为对照组,横向与同龄、同性别、BMI、BP均正常的非TN者比较;纵向与同龄、同性别、甲状腺结节发生率存在明显差异的亚组比较。
最后参考网络生物学解释TN患病的关键因素。
结果52182名体检者,TN阴性24040例,TN阳性28142例。
不同年龄组男女比例有差异。
54个亚组中49~64岁的人群样本含量和TN患病率均较高。
男性正常BMI正常BP组与肥胖高血压组比较,后者TN患病率显著升高,女性亦然(P<0.05)。
其中男性,与对照组相比,正常BMI正常BP的非TN体检者TG、GLu、糖化血红蛋白(HbA1c)、总蛋白(TP)、白蛋白(ALB)和甲胎蛋白(AFP)数值偏低。
肥胖高血压组的TN患者的ALB、AFP、HDL C、载脂蛋白A1(Apo A1)、总胆红素(TBil)和间接胆红素(IBil)降低;TG、GLu、HbA1c、小密低密度脂蛋白胆固醇(sdLDL C)、天冬氨酸氨基转移酶(AST)、丙氨酸氨基转移酶(ALT)、谷氨酰转肽酶(r GGT)、血清总胆汁酸(TBA)、尿酸(UA)、球蛋白(GLB)、β2微球蛋白(β2 MG)、癌胚抗原(CEA)、碱性磷酸酶(ALP)、C反应蛋白(CRP)升高。
身体代谢的奥秘

生命科学学院教授课题组近期的一项研究,找到了体内细胞调控代谢的一个“开关”,由它可以“下达”细胞合成代谢或分解代谢“命令”,从而解开了细胞能量代谢研究领域的一个谜底。
能量代谢是细胞中最基本、最重要的活动之一。
当能量水平下降时,细胞能通过其感应因子加快能量产生;当能量充裕时,细胞则通过另一感应因子加快耗能的活动,从而维持总体能量平衡。
林圣彩介绍,近年来的研究已经发现,能量代谢平衡调控是由多个与之相关的信号通路所介导,其中最为重要也最被广泛研究的有两条:AMPK信号通路和mTOR信号通路。
简单说来,AMPK信号通路开启的是分解代谢通路,mTOR信号通路开启的则是合成代谢通路。
随之而来的问题是,下达合成代谢和分解代谢“命令”的“指挥官”是谁,又何时下达?林圣彩课题组破解了这个谜底。
7月3日,这一研究成果在国际顶尖学术杂志《细胞》子刊《细胞·代谢》在线发表。
他们发现了控制这两个截然相反的代谢路径的“开关”。
让人诧异的是,它还竟然是同一个“开关”。
这是一种分布在细胞内膜的名为“v-ATPase-Ragulator”的蛋白质复合体。
通俗点儿说,当细胞内能量水平降低时,这个蛋白质的形状会发生变化从而让能激活AMPK的复合体与其相互作用,使之被激活。
激活后的AMPK最终下达分解代谢“命令”。
反之,当细胞内能量水平较高时,mTOR将与“v-ATPase-Ragulator”的蛋白质复合体相结合并被激活,开启合成代谢通路。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
项目名称-糖脂代谢稳态调控的分子机制-首席科学家-林圣彩厦门大学-项目名称: 糖脂代谢稳态调控的分子机制首席科学家: 林圣彩厦门大学起止年限: 2011.1至2015.8依托部门: 教育部二、预期目标1. 总体目标确定机体和细胞在不同生理状况和环境因素下维持糖脂代谢稳态的分子机制~阐明在细胞生长和应激反应中起重要作用的调节因子调控细胞代谢的信号通路网络~为糖脂代谢紊乱造成的肥胖、脂肪肝、糖尿病和癌症的早期诊断和治疗提供理论依据。
2. 五年预期目标(1) 建立对实验动物代谢相关的生理生化指标分析的技术平台~发现相关基因敲除或转基因小鼠造成糖脂代谢紊乱的信号通路。
(2) 较系统地描述在逆境下机体和细胞调控糖脂代谢的分子网络以及调控过程中关键蛋白质和蛋白质复合体的动态调控机制。
(3) 发现新的参与代谢调控的基因~为代谢性疾病和肿瘤的防治提供新的分子靶标。
(4) 培养高质量博士研究生20-30名~培养3-5名享有国际知名度的专家和5-8名中青年学术带头人。
(5) 在国际重要刊物发表SCI论文15-25篇~其中争取在Cell、Nature、Science或其子刊等影响因子10以上杂志发表研究论文5-10篇~申请发明专利3-5项。
三、研究方案1. 总体研究方案细胞能量代谢是细胞最基本、最重要的活动之一~与细胞的繁殖、分化、凋亡、运动、信号转导及多种重要疾病的发生密切相关~是生命科学的一个重要领域。
细胞要通过能量感应系统随时监测其能量水平状态~在不同的物质和能量状态下要不断地通过细胞内的代谢调控途径来调节其代谢水平以达到一种稳态。
同时~细胞在面对内外界一些不良因素时也会做出相应的代谢变化~这些应激反应对细胞正常的生长和功能是极其重要的。
如果这些应激反应失调~就会使细胞代谢发生异变~导致如前所述的多种人类重大疾病的发生。
本项目的总体研究方案拟利用我们在蛋白质科学、细胞代谢、细胞信号转导等研究领域的研究优势和技术手段~结合细胞生物学、动物生理学等学科的研究方法~集中力量多层次、多角度地研究与细胞代谢调控相关的信号通路网络~分离和鉴定参与细胞代谢调控的新的基因和信号通路~探讨各个信号通路之间的动态调控机制~并研究细胞异常代谢的信号通路~揭示代谢异常与糖尿病、肿瘤等重大疾病的关系。
项目总体研究方案如下图1: 内外环境因素(缺氧、营养缺乏或过剩、癌基因突变等)内外环境因素(缺氧、营养缺乏或过剩、癌基因突变等)糖脂转运调控因子糖脂转运调控因子细胞生长调节因子细胞生长调节因子应激反应因子应激反应因子(Glu4(Glu4、、TankyraseTankyrase、、CideCide等等))(Wnt(Wnt、、TR3TR3、、ckip-1ckip-1等等))(p53(p53、、AMPKAMPK、、HIF1HIF1等等))糖代谢糖代谢脂代谢脂代谢(转运、酵解、储存、异生等)(转运、酵解、储存、异生等)(储存、分解、分泌等)(储存、分解、分泌等)图1. 项目总体研究方案2(技术路线由于代谢调控常常涉及多种组织、器官乃至整个机体~因此利用基因敲除小鼠模型来确证基因在代谢过程中的生理功能已成为“金标准”。
本项目组已经拥有或正在构建各种基因敲除小鼠和转基因小鼠模型~包括TNKS2、PTEN、CKIP-1、cideb、p53、Lkb1、Tip60基因敲除小鼠以及cidea转基因小鼠。
我们将利用这些小鼠模型~比较研究代谢调控在正常生理状况以及不同内外因素的刺激下细胞能量代谢的变化及其调控的信号通路网络。
同时~我们也将借助体外细胞培养~特别是比较研究正常细胞以及肿瘤细胞在低氧状况下和目前常规培养状况下代谢调控的异同~并与机体内生理条件下代谢调控的情况相比较~寻找适合代谢调控研究的细胞培养模型。
在此基础上验证从动物模型得到的结果~并在分子水平上进一步深入研究影响细胞代谢的信号通路网络和调控机制。
此外~我们将通过酵母双杂交、分子筛层析、免疫共沉淀和质谱等等蛋白质研究手段捕捉并鉴定与代谢调控相关的蛋白质复合体及其组份~从而为阐明代谢调控中各种蛋白质复合体的动态组装奠定基础。
同时~利用蛋白质结构解析来揭示各种蛋白质复合体中蛋白质的相互作用关系。
故~项目的总体技术路线如图2:课题课题1/2/31/2/3课题课题2/1/32/1/3正常生理状况正常生理状况逆境状况逆境状况分子水平分子水平细胞水平细胞水平动物模型动物模型((蛋白质功能、复蛋白质功能、复((细胞培养细胞培养))((小鼠、大鼠小鼠、大鼠))合体及结构分析合体及结构分析))细胞能量代谢调细胞能量代谢调控信号网络控信号网络代谢的调控机制代谢的调控机制图2. 项目的总体技术路线3. 创新与特色本项目拟利用已有或正在进行的各种基因敲除小鼠模型~在动物个体的基础上阐明代谢调控的机理和生理作用。
同时~借助我们在脂质组学、代谢组学、蛋白质组学、功能基因组学方面的研究基础和优势~对调控糖脂代谢稳态的分子机制进行研究~探索细胞在不同逆境,低氧、缺氧、高脂等,下的代谢方式的转变及其调控信号通路网络的动态变化~特别是深入探讨细胞生长和应激反应的重要调节因子对细胞代谢的调控作用~将在分子水平、细胞水平以及动物个体水平上增进我们对代谢调控以及异常代谢与细胞异常增殖之间的关系的认识。
为最终阐明细胞糖脂代谢稳态调控的分子机制和相关重大疾病的防治提供理论基础。
4(课题设置课题1 糖脂转运及其稳态调控的的分子机制研究内容:细胞内糖脂代谢的稳态调控是维持细胞或机体基本生命活动的基础~糖脂代谢的紊乱与糖尿病、肥胖、脂肪肝、心血管疾病、细胞异常增殖以及癌症的发生和发展密切相关。
本课题将在分子、细胞和基因敲除小鼠水平上研究Axin、AMPK、TNKS2、PTEN、Cideb、Cidea等基因对葡萄糖转运、脂肪合成和储存、脂滴的形成等代谢途径的调节作用及其与肥胖症、脂肪肝和细胞异常增殖的关系。
从细胞学的角度来看~肥胖的发生是由于脂肪细胞数量的增加以及脂肪细胞中脂滴变大两个方面引起的。
研究表明~伴随着脂滴的变大~脂肪细胞变大~导致细胞炎症因子分泌的变化~例如resistin、TNFα和游离脂肪酸等分泌的增加~最终导致机体内胰岛素抵抗和糖尿病的发生。
因此~对脂滴的形成过程的研究~不仅有助于我们了解脂滴的生物学功能~而且对肥胖及肥胖引起的其它代谢综合症疾病~有深远意义。
课题1拟开展以下几个方面的研究: 1. 糖代谢的稳态调控通过前期工作~我们发现Axin和AMPK,能相互作用并增强AMPK响应能量缺失的刺激~即AMPKα的第172位苏氨酸的磷酸化。
Axin敲低的细胞在受到低糖刺激的情况下~AMPKa的第172位苏氨酸的磷酸化水平增加幅度与正常细胞相比大幅降低。
这表明Axin是应对糖稳态变化的重要因子~且在AMPK活性调控的过程中扮演了重要角色。
我们拟进行以下实验~深入阐明Axin、AMPK 和糖稳态调控的关系:(1) 研究Axin调控AMPK活性的分子机制研究Axin调节AMPK的激活是通过影响AMPK上游激酶与AMPK的相互作用还是影响了AMPK三个亚基之间的相互作用。
(2) 应用小鼠模型研究Axin缺失或突变导致的AMPK活性的变化利用腺病毒感染系统特异性地敲低小鼠肝脏或肌肉中的Axin~或利用条件性基因敲除技术敲除小鼠肝脏或肌中的Axin~对这些小鼠施以饥饿等影响能量水平的刺激~观察动物体内AMPK活性的变化。
(3) 应用小鼠模型研究Axin缺失或突变导致的糖稳态平衡的改变对上述小鼠进行糖代谢相关生化指标的测定以及AMPK参与糖代谢相关基因的表达水平的检测。
(4) 研究Aurora在Axin调控AMPK活性中的作用 Aurora是在生长中起重要作用的激酶~能调控Axin在中心粒上的定位~因此~我们拟研究Aurora是否在Axin调控AMPK活性中也起作用。
2. 研究葡萄糖转运和脂肪细胞生成的分子机制以及糖脂代谢异常与肥胖和胰岛素抵抗的关系(1) 研究Axin和TNKS2对葡萄糖转运的调节机理我们通过酵母双杂交实验发现Axin能与TNKS2相互作用,TNKS2与Kif3a之间相互作用,Axin与Kif3a之间也存在相互作用。
Kif3是由Kif3a、Kif3b和KAP3构成的异源三聚体。
它是一种依赖于微管的马达动力蛋白质复合体~可以向微管正极定向移动~在细胞内承担膜性细胞器或生物大分子复合物的运输功能。
已有的研究表明在3T3-L1脂肪细胞中Kif3参与胰岛素调节的Glut4向细胞膜的转运。
TNKS2与GSV (Glut4 storage vesicle)上的IRAP有相互作用。
我们将通过免疫荧光实验确定Kif3a、TNKS2、Axin和Glut4之间是否有共定位~而且在胰岛素刺激下是否有向细胞膜共转移的现象。
同时~我们将用siRNA 干扰C2C12细胞中TNKS2、Axin及Kif3a的表达~观察细胞对胰岛素刺激下葡萄糖吸收的反应。
另外~用TNKS2抑制剂XAV939,由厦门大学生命科学学院沈月毛教授合成,刺激C2C12细胞~观察该抑制剂是否能降低胰岛素刺激引起的葡萄糖的吸收。
通过上述实验我们将证明Kif3a、TNKS2、Axin 是否通过形成复合体来调控胰岛素刺激的Glut4的转运~从而调节葡萄糖的吸收。
(2) 分离TNKS2的新的蛋白质复合体为了更全面的找到以TNKS2为核心的调节糖脂代谢的分子网络~我们拟构建TNKS2不同片段的饵~使其覆盖TNKS2全蛋白质序列~进行大规模的酵母双杂交实验。
另一方面~我们拟以小鼠的肌肉、脂肪组织为原料~采用以高效液相色谱为核心的生化分离手段结合蛋白质谱分析技术~分离鉴定出不同生理水平下TNKS2复合体中的蛋白质组份~并用免疫共沉淀、免疫荧光共定位及 GST- pulldown等方法进一步验证这些蛋白质与TNKS2的相互作用。
(3) 研究TNKS2的多聚ADP核糖化酶活性在调节葡萄糖转运及脂肪细胞生成中的作用。
首先我们将通过质谱手段检测胰岛素是否调控TNKS2的翻译后修饰~进而研究该修饰是否影响TNKS2的多聚ADP核糖化酶的活性及TNKS2和其相互作用蛋白质的结合。
此外~由于TNKS2具有多聚ADP核糖化酶的活性~我们将考察TNKS2是否介导与其相互作用的蛋白质的多聚ADP核糖化修饰。
在此基础上我们将构建TNKS2的多聚ADP核糖化酶活性缺失的表达载体~研究其形成复合体的能力及调节葡萄糖转运和脂肪细胞生成的作用。
(4) 在动物水平上研究调控葡萄糖转运及脂肪细胞生成的分子机制。
全面分析TNKS2基因敲除小鼠与代谢相关的表型~包括小鼠在不同生长时期、不同生长条件,饥饿或饱食、缺氧等,下的体重、体温、不同部位脂肪组织的重量和脂肪细胞的数量、内脏器官的重量等。