铜绿微囊藻
铜绿微囊藻挥发性成分分析

关键 词 : 绿微 囊 藻 ; 铜 同时蒸馏 萃取 ; 气相 色谱 一质谱 法 ; 发性 成分 挥
中图分类号 : 9 9 2 文献标 识码 : 文章 编号 :6 2— 5 3 2 1 )4— 2 5— 5 Q 4 .2 A 17 8 1 ( 0 0 0 0 8 0 An l ss o he Vo a i m po n fM i r c si r gi s a y i f t l tl Co e u ds o c o y tsAe u no a
Kuiz wih t a t t he G s Chr m a o r p y —M a s S cr m e r o tg a h — s pe to t y
温度和光照对铜绿微囊藻生长的影响

第2 6卷
第 2 期 Βιβλιοθήκη 天 津科 技 大 学 学报
J u n l f ini nv ri f cec & T c n lg o r a a j U ies yo in e oT n t S eh oo y
V l 6 NO 2 b- 2 .
Apr 2 1 . 01
2 1 年 4月 01
c l d n i so t i e 0℃ el e st wa b an d i 3 y n c n i o . eo h r a d, i mir ag r w ey so y wh nt ewa e mp rt r o dt n On t t e n t s c o l ag e v r l wl e trt i h h h h e ea e u wa e st a 0 ℃ . i c o l a d d n t e u s ih ri u n t n.n r w e ti 0 x c n i o . e o t l s ls h n 2 Th smi r a g i o q e th g e l mi ai a d g e b s n 4 0 0 l o d t n T p i r l o i h ma
关键词 :温度 ;光照 ;铜绿微囊藻 ;生长
中 图分 类 号 :X1 1 7. 5 文 献标 志码 :A 文 章 编 号 : 17 —5 0 2 1) 20 2 —4 626 1 (0 1 0—0 40
微囊藻

微囊藻水华及其危害业务发展中心黄海平陈根源1. 铜绿微囊藻的分类及特征铜绿微囊藻(Microcysis aeruginosa)属于蓝藻门色球藻科微囊藻属。
微囊藻对磷酸盐的吸收和累积研究表明,某些藻类在吸收磷酸盐时具积累性,藻类能吸收过量的磷酸盐并以多聚磷酸颗粒的形式储存于体内[25]。
高学庆等[26]的研究发现,当外界环境中营养磷浓度较高时,细胞过量吸收磷可以成为微囊藻种群增长的加速剂(这一点对藻类种群在竞争中的生存是有利的)。
较大的生长速率可以使得种群尽可能快地占据较多的生存空间,而能排斥来自其它种群的竞争压力。
当环境中营养磷浓度较低时,过量积累在细胞中的营养磷含量就可以维持种群度过一个较长的时期,以保证种群个体数量不因外界环境中营养磷浓度波动而产生很大的起伏。
从而可以看出,铜绿微囊藻对水体中营养磷过量积累的特点,对微囊藻成为淡水湖泊富营养化发展过程中的一种重要优势种是具有极为重要的作用。
微囊藻内部生理结构水华的形成和扩散也是蓝藻生理生态策略的表现。
其一,形成水华的蓝藻,它们特有的异形胞能够将大气氮固定为可利用氮源,供给其它营养细胞,因此在环境中的当外来氮源不足而水体磷充足时,它们比其它生物更具有竞争优势,容易周期性的大量生长形成水华;其二,水华蓝藻另一个特点是:它们都具有一种调节细胞沉浮的结构体一伪空胞。
伪空胞是中空的蛋白质细胞内含物,气体可透过但不透过水。
当伪空胞以足够的浓度存在时可为细胞提供浮力。
在光学显微镜下可观测到大的伪空胞聚集体,这种伪空胞被称作为气囊。
而气囊的破裂与组装,为微囊藻提供了一个潜在的浮力调节机制[50]。
伪空胞在蓝藻水华的发生、扩散和消失过程中起到非常重要的作用,已有大量的文献报道伪空胞的合成条件和调节与蓝藻水华发生的关系[51-52]。
其三,水华蓝藻具有高效吸收利用外源无机碳的功能—无机碳浓缩机制(CCM)。
在低浓度的二氧化碳介质中,蓝藻可以通过高效地主动吸收浓缩外源无机碳,在细胞内积累比介质高几百到几千倍的二氧化碳浓度,由此能够在其所栖息的环境中最大限度地竞争利用有限的无机碳,保持持续稳定的生长。
铜绿微囊藻在光限制胁迫下的超补偿生长响应

迫后初期增大了 23.71%,分别是对照组和处理组
光 胁 迫 前 的 97.39% 和 96.79%。 Manodharan
在解除光限制胁迫后,处理组藻细胞密度出现 大幅增加,培养至第 10天时的藻细胞密度为胁迫后 初始接种密度的 3.3倍,而对照组的藻细胞密度变 化不大(图 2b)。处理组藻解除胁迫后生长与对照 组比较,表现出显著差异(P<0.05)。 2.3 生长参数的变化 2.3.1 相对生长率 铜绿微囊藻在光限制胁迫培 养过程中,处 理 组 平 均 相 对 生 长 率 低 于 对 照 组 (图 3a),且为负值,这与处理组在光限制胁迫过程中藻 细胞密度下降相对应。在解除胁迫条件后,处理组 的相对生长率均大于各对照组相对生长率(图 3b), 且处 理 组 在 整 个 恢 复 阶 段 平 均 相 对 生 长 率 (01187)为对照组(00034)的 34.6倍,差异极显 著(P<0.01)。但随着培养时间的推移,处理组与 对照组 2组间平均细胞相对生长率的差异逐渐减 小。到了培养 后 期,2组 的 平 均 细 胞 相 对 生 长 率 趋 于接近。
光照是影响藻类生长繁殖的重要生态因子之 一,也是其生长的主要能量来源,在一定 pH、温度和 营养条件下,光照影响着光合作用产物的合成,从而 影响藻类的生长繁殖和密度(孙儒泳等,1993)。研 究表明,微小原甲藻(Prorocernturm minimum)(Ma noharanetal,1999)、蛋 白 核 小 球 藻 (Chlorellapyre noidosa)(刘宁宁和段舜山,2002)、四裂藻(Tetrasel mistetrethele)(郭 羽 丰 等,2004)、眼 点 拟 微 绿 球 藻 (Nannochloropsisocutala)(张珍萍等,2005)和三角 褐 指 藻 (Phaeodactylum tricornutum)(Caietal,
铜绿微囊藻和小球藻对水环境 pH 的影响

中国水产科学 2016年11月, 23(6): 1380-1388 Journal of Fishery Sciences of China研究论文收稿日期: 2016-03-17; 修订日期: 2016-04-17.基金项目: 中央级公益性科研院所基本科研业务费专项资金(中国水产科学研究院东海水产研究所, 2014T04); 上海科技兴农[沪农科攻字(2015)第1-6号].作者简介: 苏发文(1988–), 硕士研究生, 主要从事养殖生理生态研究. E-mail: sufawen1213@ 通信作者: 来琦芳, 研究员, 主要从事盐碱水土资源渔业开发研究. E-mail: laiqf@DOI: 10.3724/SP.J.1118.2016.16040铜绿微囊藻和小球藻对水环境pH 的影响苏发文1, 2, 高鹏程1, 来琦芳1, 么宗利1, 陆建学1, 周凯1, 仲启铖1, 郑亮1, 王慧11. 中国水产科学研究院 东海水产研究所, 咸淡水养殖工程技术中心, 上海 200090;2. 上海海洋大学 水产与生命学院, 上海 201306摘要: 以盐碱池塘优势微藻铜绿微囊藻(Microcystis aeruginosa )、小球藻(Chlorella pyrenoidosa )为研究对象, 采用正交实验方法, 研究不同温度(20℃, 25℃, 30℃)和光照强度(2000 lx, 4000 lx, 6000 lx)组合条件下两种微藻对水环境pH 的影响。
结果显示, 处于对数生长期的铜绿微囊藻和小球藻均能使水环境pH 上升。
在本实验范围内, 不同温度和光照强度组合条件下, 铜绿微囊藻的生长均能使水环境pH 显著上升至9.50以上, 在温度25℃、光照强度2000 lx 条件下, 藻密度达最大值1.1×107 cells/mL, 水环境pH 也达到峰值10.83;小球藻生长亦能使水环境pH 的上升, 并随温度升高、光照强度增强而增大, 在温度30℃、光照强度6000 lx 条件下, 藻密度达最大值8.1×106 cells/mL, 水环境pH 也达到峰值7.73。
滇池铜绿微囊藻不同最适温度的分析
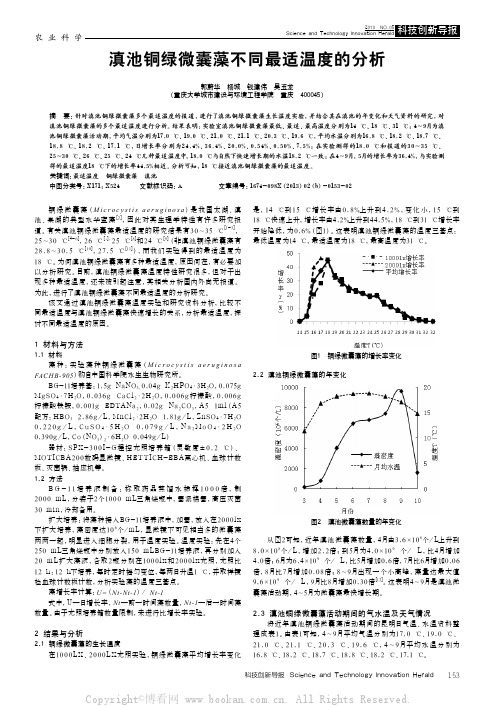
18.8 ℃、18.2 ℃、17.1 ℃,日增长率分别为24.4%、36.4%、20.0%、0.54%、0.50%、7.5%;在实验测得的18.0 ℃和报道的30~35 ℃、
25~30 ℃、26 ℃、25 ℃、24 ℃几种最适温度中,18.0 ℃与自然下快速增长期的水温18.2 ℃一致;在4~9月,5月的增长率为36.4%,与实验测
器 材:S P X-3 0 0 I - G 程 控 光 照 培 养 箱(灵 敏 度±0.2 ℃)、 MO T ICBA 2 0 0 数码显微 镜、H E T T ICH-E BA离心机、血 球 计数 板、灭 菌 锅、抽 虑 机等。 1.2 方法
B G - 1 1 培 养 液 制 备 :称 取 药 品 蒸 馏 水 稀 释 1 0 0 0 倍 ,制 2 0 0 0 m l,分 装于2个10 0 0 m l三角烧 瓶中,塞紧 棉塞,高压 灭菌 30 m i n,冷却备用。
得的最适温度18 ℃下的增长率44.5%相近。分析可知,18 ℃接近滇池铜绿微囊藻的最适温度。
关键词:最适温度 铜绿微囊藻 滇池
中图分类号:X171;X524
文献标识码:A
文章编号:1674-098X(2013)02(b)-0153-02
铜 绿 微 囊 藻( M i c r o c y s t i s a e r u g i n o s a)是 我 国 太 湖 、滇 池 、巢 湖 的 典 型 水 华 蓝 藻 [2],因 此 对 其 生 理 学 特 性 有 许多 研 究 报 道。有关 滇 池 铜 绿 微 囊 藻 最 适 温 度 的 研 究 结 果有 3 0~35 ℃[1-3], 2 5~3 0 ℃[4-6],2 6 ℃[7],2 5 ℃[8]和 2 4 ℃[9](非 滇 池铜 绿微 囊 藻 有 2 8.8~3 0.5 ℃[10],2 7.5 ℃[11]),而 我 们 实 验 得 到 的 最 适 温 度 为 18 ℃。为何滇池铜绿微 囊藻有多种最适温度,原因何在,有必要加 以分析 研究。目前,滇 池铜 绿微 囊 藻 温 度特 性研 究很多,但 对 于出 现多种最 适温度,还未被引起 注意,其相关 分析国内外尚无报 道。 为此,进行了滇 池 铜 绿 微 囊 藻 不同最 适 温 度 的 分 析 研 究。
巢湖市水源地铜绿微囊藻(Microcystis aeruginosa)藻团粒径时空分布规律

巢湖市水源地铜绿微囊藻(Microcystis aeruginosa)藻团粒径时空分布规律范帆;李文朝;柯凡【摘要】于2011年4-8月按月对巢湖市水源保护湖区不同水深及不同区位的铜绿微囊藻藻团粒径进行抽样调查,利用统计分析方法,归纳了巢湖水源保护湖区铜绿微囊藻藻团粒径的时空分布规律.湖区铜绿微囊藻藻团出现在4月中旬至5月中旬之间,S1与S2点位表层的藻团粒径与中、底层的均存在显著差异.粒径小于200 μm的藻团在各水深的分布都比较均匀,没有明显的趋向性;粒径在200~800μm 范围内的藻团更易集中在湖水表层;粒径超过800 μm的藻团更易集中在湖水底层.各月份外湖区S2点位藻团粒径水平均高于内湖湾S1点位.由于易受短时气象条件的影响,藻团粒径按月时间尺度变化的规律性不强.藻团形状在整个空间分布上没有显著差异性,但随着季节变化,逐渐由狭长形向规则形变化.藻团粒径的分布范围表明,大型仿生式蓝藻清除设备的过滤筛网对水源湖区铜绿微囊藻藻团的理论过滤效率为99.81%.%An investigation of Microcystis aeruginosa colony diameters in different water depths and locations was carried out monthly in the protection zone of water source region of Chaohu City from April to August, 2011. The spatio-temporal distribution characteristics of the colony diameter were summarized using statistical analysis. The M. aeruginosa colony appeared between mid-April and mid-May, there was significant difference between the surface layer and lower-middle layers on colony diameters at both sampling sites of SI and S2. Colonies with diameters less than 200 (i.m distributed vertically homogeneously in the water column, colonies with diameters between 200 Ltn and 800 μmtended to gather in the water surface, and colonies with diameters greater than 800 μm were apt to aggregate in the bottom layer. The lev el of colony diameter of outer sampling site (S2) was higher than that of inner sampling site ( SI ) in each month. Because the colony diameters were easily affected by the short-term meteorological condition, the variance of colony diameters didn't show conspicuously regular monthly. There was no significant difference in the spatial distribution of colony shape, while it shifted graduelly from elongated to regular with seasons changing. The distribution range of colony diameters indicated that, in theory, 99.81% of the total M. aeruginosa colonies can be fdtered by the large bionic equipment for clearing cyanobaeteria.【期刊名称】《湖泊科学》【年(卷),期】2013(025)002【总页数】8页(P213-220)【关键词】铜绿微囊藻;藻团粒径;形状因子;分布;巢湖【作者】范帆;李文朝;柯凡【作者单位】中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京210008【正文语种】中文巢湖位于安徽省中部,是我国第五大淡水湖,湖泊面积约780 km2,平均水深4.5 m.巢湖不仅是安徽省重要的渔业基地和航运通道,也是巢湖市及周边地区重要的饮用水水源地.自1970s 以来,巢湖富营养化程度日益加剧,夏、秋季蓝藻水华肆虐,严重影响了当地工业生产用水以及城市居民供水[1].大部分可以形成水华的蓝藻种群易在水体中聚集成藻团,以藻团形式存在的蓝藻占据了整个种群的大部分比例.清除蓝藻藻团可以有效减少水华蓝藻的总生物量,从而降低蓝藻水华的危害程度甚至预防水华的暴发.根据蓝藻上述特性成功研制的大型仿生式水面蓝藻清除设备,能汲取5 cm 富含蓝藻水层,通过高密度超低孔径筛网对湖水中蓝藻进行藻水分离,对肉眼可见蓝藻分离率达100%,处理流量可达1000 m3/h.铜绿微囊藻是巢湖水华蓝藻主要优势种群之一,由于其在水体中分布范围广、持续周期长[2]以及产藻毒素的特性[3],危害性远高于其他藻类.因此,本文着重考察巢湖水源地铜绿微囊藻藻团粒径季节性发育及分布状况,为设备对铜绿微囊藻藻团的过滤效率研究提供科学的理论依据.1 材料与方法1.1 湖区简介巢湖市水源保护湖区位于巢湖东北角(30°34'31″~31°36'45″N,117°47'8″~117°50'54″E),西至龟山,东至裕溪河口.整个湖区是一个西宽东窄的湖湾,面积大约10 km2,平均水深2.7 m,西部湖界横断面长约2.5 km.湖区西部水面开阔,与外湖湖水交换频繁,湖东呈“Y”型岔口,岔口北段为封闭水湾,内有一个小型码头,供渔政船停靠,水深较大,最深处达5 m 左右,南段为裕溪河口.建于河口上的巢湖闸是调节巢湖水量的关键性水利工程,闸门未开时,水源湖区呈半封闭状态.水源地南岸多丘陵,北岸地势较为平坦.巢湖水源保护湖区是整个巢湖目前唯一的水源地,湖区内设有巢湖市第一和第二自来水厂的取水口.该水源地不仅为巢湖市居民提供饮用水水源,同时也为周边企业生产用水以及居民生活用水提供水源,因此水源地水质的优劣对巢湖市的经济与民生有着重大影响.1.2 采样点设置在水源湖区设置2 个采样点(图1),采样点S1(31°35'31″N,117°51'7″E)位于内湖湾内,平均水深约为4.5 m.采样点S2(31°35'59″N,117°45'36″E)位于湖区西部大湖面上,平均水深约为3.4 m.由于南岸丘陵、湖岸大坝以及河口大桥等建筑物的阻隔,内湖湾受风浪的扰动较小,特别是南岸丘陵对西至西南风风力有明显的削弱作用,相比开阔湖面,前者受到的水力扰动明显低于后者.图1 巢湖市水源保护湖区地理区位及采样点位置Fig.1 Location of water protection zone of Chaohu City and sampling sites1.3 水样采集与处理2011年 4-8月,每月中旬分别在 S1、S2点位采集表层(S1:水下0.5 m,S2:水下0.5 m)、中层(S1:水下 2.0 m,S2:水下1.5 m)和底层(S1:水下4.0 m,S2:水下3.0 m)水样.用有机玻璃采水器(容量2.5 L)在每个深度取样多次,倒入大桶(容量10 L)中混合搅匀,再用采样瓶采集1 L 水样,目的是尽量减小因采样不均导致的系统误差.在水样采集后现场加入15 ml 鲁哥试剂固定,用作后续的藻团计数以及粒径测量.用Kestrel 4500 型便携式气象仪测量采样时的瞬时风速,用采水器自带的温度计测量水温.1.4 样品分析与数据采集经鲁哥试剂固定后的1 L 水样在实验室静置24 h 后浓缩至30 ml,转入定量瓶待测.用于藻团计数与粒径测量的仪器为Nikon Ts100 型光学倒置显微镜,显微镜摄像头为DS-Fi1 型.首先进行铜绿微囊藻的鉴别和藻团计数,藻种的鉴别参照文献[4-5],藻团的计数参照文献[6].通过该显微镜自带测量软件(NIS-Elements D,图2)在一定放大倍数(40 ~200 倍)下对各样品中铜绿微囊藻藻团粒径进行测量.测量工作开始之前,先用测微台尺定标各个放大倍数下象素对应的实际长度.每次测量样品时,将定量瓶充分摇匀,从中央部分吸取0.1 ml 样品于0.1 ml 计数框内,随机测量出现在视野中的藻团100 个.在手动测量每个藻团的二维投影面积(S)和拟合椭圆的长短轴(A 轴、B 轴)后,软件会自动生成等圆直径(d)和长短轴比(A 轴/B 轴),在后文中前者简称为粒径,后者的倒数简称为形状因子.形状因子(shape factor)是图像分析以及显微镜检中用来数值化描述颗粒形状的无量纲量,常用来表示微粒形状与理想形状(如圆形、球形、等多边体形)之间的差异,与颗粒大小无关[7].椭圆轴比(axial ratio of an ellipse)是常用的形状因子之一,被大多数图像分析系统所采用,其值为拟合椭圆的短长轴比,比值范围为0 ~1,比值越接近0 颗粒越狭长,越接近1 颗粒越规则、越接近理想形状.计算该量时,对轴的定义有两种:一为颗粒二维投影轮廓的最长轴与最短轴,两轴不一定垂直;二为与颗粒二维投影面积相差最小的理想椭圆的长短轴,两轴相互垂直[8],本实验中对藻团形状因子的测量采用的是后者.采样期间各月份的每日气象数据资料由巢湖气象监测台站(58326)提供.由于5月份内湖湾进行航道疏浚,考虑到扰动底泥及水层对实验结果的影响,未采集S1 的水样.同时因为在4月份水样中没有观测到铜绿微囊藻藻团,因此实际只测量了5月份S2 点位,6、7、8月份S1 和S2 点位的藻团参数.图2 显微镜测量软件工作界面Fig.2 Working interface of the measurement software1.5 实验数据分析用SPSS 17.0 统计分析软件分别对各月份S1、S2 点位3 个水深的藻团粒径和形状因子进行单因素方差分析(One-way ANOVA,α=0.05),比较水源湖区不同水深藻团粒径和形状因子的分布差异.分别对各月份S1、S2 点位的藻团粒径总样本及形状因子总样本(将3 个不同水深的样本混合)进行独立样本t 检验,比较水源湖区各月份下内湖湾与外湖区藻团粒径和形状因子的分布差异.2 结果与分析2.1 铜绿微囊藻藻团粒径的统计描述铜绿微囊藻藻团之间的大小差异很大,小到只有若干个单藻构成,大到肉眼可见.粒径变化范围为20.54 ~1620.28 μm,大部分粒径分布在 30 ~300 μm 范围内,占样本总数的 96.25%,粒径小于30 μm 的藻团占样本总数的0.19%,粒径大于400 μm 的藻团占样本总数的3.56%.由图3、4 可以看出,S1、S2 各月份下不同水深的藻团粒径样本分布均呈正偏态.对各水深粒径样本分布正态性进行单样本K-S 检验的结果显示P 值均小于0.05,即粒径样本不服从正态分布.2.2 不同水深藻团粒径分布的统计分析对7 组不同水深粒径样本进行单因素方差分析的结果显示,6月份S1(df =2297,P <0.05)、8月份S1(df=2297,P <0.01)、5月份 S2(df=2305,P <0.01)和 6月份 S2(df=2297,P <0.01)不同水深的藻团粒径分布差异显著.对7 组不同水深的藻团粒径样本进行多重比较(最小显著差数,LSD)的结果显示,在上述出现显著分布差异的月份及采样点,表层与中层、表层与底层的粒径分布差异显著高于中层与底层,因此水源湖区藻团粒径在水柱中的分布差异主要来自表层与中、底层的差异.图3 6-8月S1 点位不同水深的铜绿微囊藻藻团粒径分布Fig.3 Distribution of M.aeruginosa colony diameter in different depths at S1 site from June to August图4 5-8月S2 点位不同水深的铜绿微囊藻藻团粒径分布Fig.4 Distribution of M.aeruginosa colony diameter in different depths at S2 site from May to August不同粒径水平的铜绿微囊藻藻团在不同水深的分布存在差异,3 组不同粒径水平的藻团在不同水深的比例分布(图5)表明,粒径小于200 μm 的藻团在各水深的分布都比较均匀,在整个水柱中的分布没有明显的趋向性.粒径大于200 μm 的藻团在表层水中的分布比例明显高于中、底层,说明粒径较大的藻团易于在水柱表层聚集.同时也发现,在7、8月S2 采样点,无论是粒径小的藻团还是粒径较大的藻团,在各水深的分布都比较均匀.图5 3 组不同粒径范围的铜绿微囊藻藻团在不同水深的比例分布Fig.5 Ratio distribution of M.aeruginosa colonies in three ranges in different water depths2.3 不同湖区藻团粒径分布的统计分析分别对6、7、8月S1 和S2 采样点整个水柱的粒径样本(将3 个水深的藻团粒径样本混合,作为采样点的月粒径样本)进行独立样本t 检验,检验结果(表1)显示:两个采样点的藻团粒径分布存在显著差异,各月份下S2 的藻团粒径均值均大于S1.两个采样点藻团粒径随月份的变化趋势基本一致,6月份藻团粒径均值明显高于其他月份.粒径处于100 ~200 μm 区间的藻团占总样本比例最高,为44.6%,大于400μm 的藻团占总样本比例最低,为4%,上述两个区间的藻团粒径在S1 和S2 的分布比例大致相当,分别为21.3%和23.3%,1.7%和2.3%.两个采样点粒径分布差异主要来自小于100 μm 和200 ~400 μm 两个区间的藻团,前者在S1 和S2的比例分别为21.2%和11.9%,后者比例分别为5.9%和12.5%(图6).表1 各月份下S1、S2 藻团粒径样本的独立t 检验显著性及均值比较*Tab.1 Significance of independent t-test and comparison of mean values between colony diameter samples at S1 and S2 sites in each month* 显著性水平α=0.05.采样点 5月 6月 7月 8月<0.001 <0.001 <0.001粒径/μm S1- 195.8139 106.3039 114.6498粒径/μm S2 127.3384 249.4594 138.4981145.1884 P-2.4 藻团形状因子的统计分析藻团形状因子最小值为 0.14,最大值为 1.00,形状因子在 0 ~0.50、0.51 ~0.80、0.81 ~1.00 范围内的藻团占总样本比例分别为14.00%、54.63%和31.37%.对各组不同水深藻团形状因子的方差分析以及各月份下S1 与S2 采样点间形状因子的t 检验结果表明,湖区藻团形状在空间分布上没有显著差异.但形状因子均值随月份的增加逐渐增大,S1 点6-8月均值分别为0.6479、0.7400、0.7559,S2 点5-8月份均值分别为0.6439、0.6572、0.7094 和 0.7422.2.5 铜绿微囊藻藻团计数图6 铜绿微囊藻藻团各粒径区间占6-8月份粒径总样本的比例及分别在S1 和S2 的比例Fig.6 Ratios of each diameter interval against the June-to-August diameter sample and their respective ratios at S1 and S2 sites在最早观测到藻团的5月份,S2 水柱表、中、底层藻团数量分别为1545、1077、1114 个/L.6月份S1 的藻团数量略高于S2,自7月份后,S2 的藻团数量开始明显高于S1,两个采样点水柱中藻团的平均数量分别为2515、5137 个/L,这一差距在8月份进一步扩大,两个采样点藻团的平均数量分别为 4060、10183 个/L.3 讨论湖水中的微囊藻藻团不仅来自水体中微囊藻细胞的分裂增殖,同时也来自底泥中过冬藻团的补充[9].当沉积在湖底的微囊藻藻团接受到足够阳光时,细胞开始进行不产氧光合作用并生成气囊,使得藻团能够漂浮[10].水温升高也是促进微囊藻气囊生成的重要条件,当水温超过20℃时,处在黑暗环境下的非漂浮藻团能迅速重新获得浮力[11].水源湖区4月份的平均气温为17.2 ± 0.9℃,采样水温为15.3℃,低水温既不利于湖水中微囊藻的分裂繁殖[12-13],也不利于过冬藻团从底泥向湖水中迁移,因此在4月份较难观测到铜绿微囊藻藻团.水源湖区铜绿微囊藻藻团粒径在水柱中分布的差异性主要来自表层与中、底层的差异,而中层与底层之间的差异并不显著.表层的藻团粒径通常较大,不仅因为表层光照充足,利于藻团细胞的光合作用,还因为粒径大的藻团更容易克服紊流对藻团的裹挟力,从而使其能停留在水体表层[14].水源湖区粒径大于200 μm的藻团在表层的比例明显高于中、底层,因此可以将200 μm 理解为导致水柱中藻团粒径分布存在显著差异的较大藻团的粒径下限.也有研究表明粒径大于120 μm 的藻团较易集中在表层[15],两个结论的差异可能来自对藻团粒径定义的不同.Wallace 等通过模型模拟,认为粒径达到400 μm 的铜绿微囊藻藻团使自己变得足够重后沉入水底,从而能接触到泥水界面[14].Rabouille 等通过模型模拟分析后也认为粒径大于600 μm 的藻团更容易停留在深水层.当大藻团沉入湖底时,由于底层缺少光照和水温较低,减缓了藻团细胞对糖原的消耗,从而延迟了藻团向表层的回迁[16].尽管如此,长时间停留在底层也会为藻团提供一些其他优势,比如能使藻团接触到更多从沉积物中释放出来的营养物质[14,16].本实验中,在各月份下2 个采样点的表层和中层均未观测到粒径超过800 μm 的藻团,这些超大粒径藻团全都出现在底层.因此,水源湖区不同粒径范围的铜绿微囊藻藻团在各水深分布的一般规律是:粒径小于200 μm 的藻团在各水深的分布都比较均匀,没有明显的趋向性;粒径在200 ~800 μm 范围内的藻团更易集中在湖水表层;粒径超过800 μm 的藻团更易集中在湖水底层.大量研究表明风对藻类在水体中的分布有着极其重要的影响.George 等认为当风速大于3.7 m/s 时,紊流会代替层流,导致水柱中藻类趋于均匀性分布[17].Webster 通过构建模型从理论上将该临界值缩小为2 ~3 m/s[18].Cao 等通过在太湖的野外观测,得出的实际临界值为3.1 m/s[19].湖区铜绿微囊藻藻团在水柱中的分布对风速的响应十分敏感,在7月份S1、S2 点位,8月份S2 点位采样时测得的瞬时风速分别为3.6、5.3和4.6 m/s,此时风浪扰动对藻团在水柱中分布的影响已经远大于藻团自身的垂直迁移运动,因此,上述月份和采样点的藻团粒径和数量在整个水柱中的分布都趋向均匀.6月份采样前5 d(6月8-12日)湖区平均气温为26.5℃,平均风速为2.7 m/s,温度较高、风速较小的天气是导致6月份水样的藻团粒径相比5月份有显著增加的一个重要因素.从野外观测和实验模拟都证实风浪的扰动会造成粒径较大的微囊藻藻团破裂[20-21],湖区藻团粒径总体水平在6月份之后显著下降与风浪的扰动密切相关,这也说明藻团粒径的变化易受短时气象条件的影响.O'Brien 等通过实验模拟发现,在经历不同强度扰动后铜绿微囊藻藻团存在一个最大稳定粒径,大小在220 ~420 μm 之间,大于该粒径的藻团易在扰动中破裂[21].在本实验中,大部分藻团粒径分布在300 μm 以内,占粒径总样本的91.17%,粒径在300 ~400 μm 的藻团占总样本的5.27%,而大于400 μm 的藻团仅占总样本的3.56%,属于小概率事件,因此可以认为水源湖区铜绿微囊藻藻团最大稳定粒径在300 ~400 μm 范围内.外湖区的藻团粒径均值在各月份下均高于内湖湾,说明风浪扰动在显著降低湖区水体中藻团粒径总体水平的同时,也会对一定范围内藻团粒径的发育有促进作用,该范围的上限即为最大稳定粒径.这种促进作用可能是因扰动导致的水体中营养盐浓度、光照条件等环境因子变化协同作用的效果,对此机理的探讨需更深入的研究.狭长形藻团其表面积与体积的比率较大,有利于藻团对光的吸收[10],这样有助于处于复苏阶段的铜绿微囊藻提高自身对光的利用效率.而在夏季,过高强度的光照反而会抑制微囊藻的生长,此时较规则形藻团可以减少受光面积.此外,对称形状的藻类在下沉时所受阻力明显大于非对称形状藻类[22],在风浪扰动频繁的夏季,规则形状有利于增加藻团的漂浮能力.以上假设可能是引起湖区藻团形状变化的原因,但对此现象的解释还需要结合更多生物以及非生物因素的影响.大型仿生式水面蓝藻清除设备其筛网的平均孔径为30 μm,仅从藻团粒径大小的角度考虑,该设备对铜绿微囊藻藻团的理论过滤效率达到99.81%.但本文所定义的藻团粒径是对藻团实际大小的一个近似表征,当藻团近似球形时,粒径值与藻团实际大小吻合程度较高,而当藻团呈不规则形态时,上述方法的表征效果较差.这种差别会影响对筛网实际过滤效率的评估,比如两个等二维投影面积且等圆直径大于30 μm 的藻团,一个为理想球形,一个为狭长形,后者在通过筛网表面时可能会穿过网眼.尽管如此,筛网对藻团分离的实际效率仍然保持在一个相当高的水平. 研究藻团粒径分布不仅可以为相关工程应用提供科学的理论支撑,同时也在一些理论研究领域发挥重要作用,比如:在构建模拟微囊藻垂直迁移模型时需要准确的藻团粒径分布数据来支撑[21].尽管与用藻细胞数量来表征微囊藻藻团大小的方法[23]相比,本实验中所使用的图像测量技术更快速、更直观,但这种广泛应用于材料学领域的微粒粒径测量方法应用到微囊藻藻团粒径测量上,仍然存在很大的局限性.因为有的微囊藻藻团呈枝杈状,在水样中不同的悬浮姿态在显微镜视野下对应有不同的二维轮廓,并且在不同的焦距下所观测到的轮廓也不同,因此给实际测量带来相当大的难度.日臻完善的显微三维测量技术有望解决上述问题,同时精确可靠的图像自动识别分析系统也期待在藻类识别和测量领域得到进一步发展[24].4 结论1)巢湖水源湖区铜绿微囊藻藻团最早出现于4月中旬至5月中旬之间.藻团粒径分布范围为20.54 ~1620.28 μm,其中大部分粒径分布在30 ~300 μm 范围内,占总群体的96.25%.2)湖区铜绿微囊藻藻团粒径在各水深的分布差异主要来自表层与中、底层差异.粒径小于200 μm 的藻团在各水深的分布都比较均匀,没有明显的趋向性;粒径在200 ~800 μm 范围内的藻团更易集中在湖水表层;粒径大于800 μm 的藻团更易集中在湖水底层.外湖区藻团数量及藻团粒径较内湖湾均具有明显优势,两者粒径分布差异主要来自小于100 μm 和200 ~400 μm 区间段.3)风浪的扰动不仅对藻团有破坏作用,使藻团粒径整体水平显著降低,而且使藻团粒径以及数量在整个水柱中的分布趋于均匀.湖区藻团最大稳定粒径在300 ~400 μm 之间.4)铜绿微囊藻藻团形状在空间分布上没有显著差异.但随季节变化,藻团形状逐渐从狭长形向规则形演变.5)大型仿生式蓝藻清除设备对湖区铜绿微囊藻藻团的理论过滤效率可达99.81%.5 参考文献【相关文献】[1]殷福才,张之源.巢湖富营养化研究进展.湖泊科学,2003,15(4):277-283.[2]闫海,潘纲,张明明.微囊藻毒素研究进展.生态学报,2002,22(11):1968-1975.[3]Deng DG,Xie P,Zhou Q et al.Studies on temporal and spatial variations of phytoplankton in Lake Chaohu.Plant Biology,2007,49(4):409-418.[4]黎尚豪,毕列爵.中国淡水藻志.北京:科学出版社,1998.[5]Doers MP,Parker DL.Properties of Microcystis aeruginosa and M.flos-aquae (Cyanophyta)in culture:taxonomic impli-cations.Journal of Phycology,1988,24(4):502-508.[6]国家环境保护总局《水和废水监测分析方法》编委会.水和废水监测分析方法:第4 版.北京:中国环境科学出版社,2002.[7]Friel JJ,Grande JC,Hetzner D et al.Practical guide to image analysis.Ohio:ASM International,2000.[8]Nafe R,Schlote W.Methods for shape analysis of two-dimensional closed contours-a biologically important,but widely neglected field in histopathology.Electronic Journal of Pathology and Histology,2002,8.2:022-02.[9]Tsujimura S,Tsukada H,Nakahara H et al.Seasonal variations of Microcystis populations in sediments of Lake Biwa,Japan.Hydrobiologia,2000,434(1/2/3):183-192.[10]Naselli-Flores L,Barone R.Steady-state assemblages in a Mediterranean hypertrophic reservoir.The role of Microcystis ecomorphological variability in maintaining an apparent equilibrium.Hydrobiologia,2003,502(1/2/3):133-143.[11]Thomas RH,Walsby AE.The effect of temperature on recovery of buoyancy byMicrocystis.Microbiology,1986,132(6):1665-1672.[12]孔繁翔,高光.大型浅水富营养化湖泊中蓝藻水华形成机理的思考.生态学报,2005,25(3):589-594.[13]陈宇炜,秦伯强,高锡云.太湖梅梁湾藻类及相关环境因子逐步回归统计和蓝藻水华的初步预测.湖泊科学,2001,13(1):63-70.[14]Wallace BB,Bailey MC,Hamilton DP et al.Simulation of vertical position of buoyancy regulating Microcystis aeruginosa in a shallow eutrophic lake.Aquatic Sciences,2000,62(4):320-333.[15]Wu X,Kong FX.Effects of light and wind speed on the vertical distribution of Microcystis aeruginosa colonies of different sizes during a summer bloom.International Review of Hydrobiology,2009,94(3):258-266.[16]Rabouille S,Salencon MJ.Functional analysis of Microcystis vertical migration:a dynamic model as a prospecting tool.Ⅱ.Influence of mixing,thermal stratification and colony diameter on biomass production.Aquatic Microbial Ecology,2005,39(1):281-292.[17]George DG,Edwards RW.The effect of wind on the distribution of chlorophyll a and crustacean plankton in a shallow eutrophic reservoir.Journal of Applied Ecology,1976,13(3):667-690.[18]Webster IT.Effect of wind on the distribution of phytoplankton cells in lakes revisited.Limnology and Oceanography,1994,39(2):365-373.[19]Cao HS,Kong FX,Luo LC et al.Effects of wind and wind-induced waves on vertical phytoplankton distribution and surface blooms of Microcystis aeruginosa in Lake Taihu.Journal of Freshwater Ecology,2006,21(2):231-238.[20]Robarts RD,Zohary T.Microcystis aeruginosa and underwater light attenuation in a hypertrophic lake (Hartbeespoort Dam,South Africa).Journal of Ecology,1984,72(3):1001-1017.[21]O'Brien KR,Meyer DL,Waite AM et al.Disaggregation of Microcystis aeruginosa colonies under turbulent mixing:laboratory experiments in a grid-stirredtank.Hydrobiologia,2004,519(1/2/3):143-152.[22]Padisák Judit,Soróczki-Pintér éva,Rezner Z.Sinking properties of some phytoplankton shapes and the relation of form resistance to morphological diversity of plankton-an experimental study.Hydrobiologia,2003,171:243-257.[23]Cao H,Zhou Y.Variation in colony size of Microcystis aeruginosa in a eutrophic lake during recruitment and bloom formation.Journal of Freshwater Ecology,2010,25(3):331-335.[24]Walker RF,Kumagai M.Image analysis as a tool for quantitative phycology:a computational approach to cyanobacterial taxa identification.Limnology,2000,1(2):107-115.。
流动影响铜绿微囊藻生长的实验研究

流动影响铜绿微囊藻生长的实验研究流动影响铜绿微囊藻生长的实验研究高一记叙文流动影响铜绿微囊藻生长的实验研究高一记叙文随着科学技术的发展,各国学者对微囊藻的研究也越来越深入,微囊藻是一种单细胞真核藻类,它具有很强的抵抗环境污染和光照等不利因素的能力,并且能在极端恶劣的环境下正常生长,具有广泛的应用前景。
微囊藻的结构简单,且繁殖快,易于培养,是作为植物生产抗生素、降解农药和清除水中重金属的最佳菌株。
如果把微囊藻大量培养到工业上,可以取代化肥,既减少了污染,又节省了开支,提高了经济效益。
但是,微囊藻对培养基成分和水体环境有着较高的要求,在自然条件下它们一般只能适应低溶解氧的环境,当外界溶解氧含量过低时,藻类的生长速度会变慢,而对铜绿微囊藻的生长起决定性作用的是水中的溶解氧。
根据现有技术,铜绿微囊藻在恶劣水体环境中的培养难题尚未得到解决,目前主要依靠的是研究微囊藻在纯水或是复合培养基中的培养,至于铜绿微囊藻在污染水体环境下的培养与研究还很少,还有待进一步的探讨与研究。
所以,为了研究铜绿微囊藻在恶劣水体环境下的生长情况,本人通过查阅相关资料,根据不同水体的特点,设计出不同的实验方案,采用理论分析法和数值模拟法进行实验研究。
选择颗粒较大的碳酸钙颗粒,控制液面与加液管之间的距离。
当固液比达到一定程度后,添加清水至上液位线处;继续滴加至与原液面平齐时,即为临界状态,保持这一水位不变。
定时测定容器内液面位置,当液面高度降低到0.05时,向系统内缓慢添加碘酒,同时将配置好的培养基倒入定时烧瓶中,一次完成菌种的接种工作。
待容器内液面升至1.5时,打开排气管,使瓶内空气完全排出,在瓶口塞一团脱脂棉,以防止进水。
培养结束后,抽去棉花,称重培养基。
对照实验为用相同浓度的培养基在无菌条件下进行纯水的培养。
每组两个培养皿,各装25g培养基,用7-9层脱脂棉布包裹严密,放入盛有生理盐水的锥形瓶内,置37 ℃恒温箱中培养24h,观察对照组的水体颜色及透明度变化情况,检测铜绿微囊藻的生长情况。
溶藻菌的分离及其对铜绿微囊藻溶藻特性的研究

溶藻菌的分离及其对铜绿微囊藻溶藻特性的研究溶藻菌的分离及其对铜绿微囊藻溶藻特性的研究溶藻菌是一种能够降解藻类细胞壁并释放藻胞内物质的细菌,对于藻类生态系统的稳定性具有重要作用。
本文旨在介绍溶藻菌的分离方法,并研究其对铜绿微囊藻的溶藻特性。
通过这项研究,人们可以更好地认识溶藻菌的作用机制,进一步推动藻类生态系统的保护和修复。
首先,我们需要分离溶藻菌。
采样地点选择在铜绿微囊藻繁殖较为丰富的水域,如湖泊或水库。
采用传统的培养基,将水样接种于琼脂平板培养基上,培养出单菌落后进行纯化。
通过进行形态观察、生理生化检测以及基因测序等方法可以鉴定溶藻菌的种类。
最后,将纯化后的菌株保存以备后续实验使用。
接下来,我们研究溶藻菌对铜绿微囊藻的溶藻特性。
首先,在实验条件下培养出一定浓度的铜绿微囊藻。
然后,将不同种类的溶藻菌接种于含有铜绿微囊藻的培养基中,建立菌藻共生体系。
通过监测铜绿微囊藻生长的光密度、叶绿素含量以及细胞数目等指标,可以评估溶藻菌对藻类的溶解效果。
在实验过程中,研究人员发现不同溶藻菌对铜绿微囊藻的溶藻效果存在差异。
有些溶藻菌能够迅速降解铜绿微囊藻细胞壁,释放出藻胞内物质,限制藻类的生长。
而有些溶藻菌对铜绿微囊藻影响较小,甚至对其生长有促进作用。
这表明溶藻菌的种类和数量对藻类生态系统的影响非常重要。
此外,溶藻菌的溶藻机制也是研究的重点之一。
通过对溶藻菌分泌的溶藻酶进行分析,研究人员发现溶藻菌通过释放酶来分解藻类细胞壁,并吸收其中富含的营养物质。
这不仅揭示了溶藻菌的溶藻机制,也为微囊藻类繁殖的控制提供了一种新的思路。
综上所述,溶藻菌的分离及其对铜绿微囊藻的溶藻特性研究对于藻类生态系统的保护和修复具有重要意义。
通过深入研究溶藻菌的作用机制,人们可以寻找到一种更加有效的方法来控制藻类的繁殖,保护水域环境的健康。
此外,这项研究还为微生物资源的利用提供了新的途径,有助于推动生态学领域的研究和应用综合以上研究结果,溶藻菌对铜绿微囊藻的溶解效果存在差异,有些溶藻菌能够有效降解铜绿微囊藻细胞壁,限制藻类的生长,而有些溶藻菌对其影响较小甚至具有促进作用。
铜绿微囊藻(Microcystic aeruginosa)胞外酸性多糖抗肿瘤活性实验研究

g a u l stete t n t o t u s a dtehg e tn a in fe t c u o he6 r8 a . AP dEAP I r d a ya h r ame ti c ni e. l me n n h i h s i bt gefcso c r nt 血o md y E S I a h i n SI
h n t t FU. h r wt f l . P -o 5 c l s l e i e t e t i e y t e a d t ei a i n f e t g p t a a f - h o 5 T eg o h o a S 2 r He o U2 el i a l v d n l r s a n d b m n 1 s y r h h n b t g e f cs ou h i
维普资讯
※营养卫生
良 晶 科 学
20,o 2, o0 31 07V1 8N .6 1 .
铜绿微囊藻( co yt eu ioa胞外酸性 Mir cs cargn s) i
多糖抗肿瘤 活性 实验研 究
张 玲1 ,梅 秋 红 1 薛 华 杰 1 周 耀 明 2 陆长 梅 1 吴 国荣 , , , , ,
tmo e s dc c r c . sls n iae a AP a dE SI aealo i e s f I u rc l a e e Reut dc tdt t n a n i m i h E S I n AP I l l txct c l EK23 u etxct s o oF 9b th ii i l s t o y se
Ab ta t EAP dEAPSI tatdfo M .e u io ah dih bt gefcs nt mo el i ut aign r l d sr c : S Ia n e rce m a r gn s a a i n fe t Ix r n i o u r l c l v t o ma c sn i n n a
臭氧氧化铜绿微囊藻及其对水质的影响

・
第3 6卷
第 6期
21 0 0年 1 2月
水污 染 防治 ・
臭氧氧化铜绿微囊藻及其对水质的影响
Std n t e Oz nz t n o c o y t r gn s n f c n W a e ai u y o h o ia i f o Mirs s i Ae u io a a d E e to t r c Qu ly t
K y wo d E t p i t n Mir s s i A r gn s Oz e Oxd t e rs u r hc i o ao c o y t e u io a s n o i i ao n
世界 上淡水 湖泊 藻类 水华 发 生 的频 率 与严 重
程度 都呈现 增 长 的趋 势 , 中蓝 藻 是 引 起 藻 类 水 其 华 污染 的主要藻 类… 。水 体 富营 养 化而 导致 的蓝 藻水 华暴发 给世 界各 国 带来 重大 的 环境 问题 。藻 类繁 殖会 释放 臭 味 物 质 , 起 饮 用 水 感 官 性 能 的 引 下降, 加大 了处 理 含 藻 水 的 难 度 。藻 类 的 繁 殖 还 对饮 用水 的安 全 性 造 成 了威 胁 , 放 藻 毒 素 引 起 释
o v o ai c bni r s. a cladcloy l ocnri epe e y 40 1 0adcnettno c o p y ) sl r n ro c ae gl e n h r blcnettn《xr sdb D 4 ,038 n ocna o l ohla d e g ca ne l op ao s O 6 r i fh r l
d ce e s n t f ro ia o e ra sd t cl at xd t n.F r e m r , e p s i i y e i u t r o e t H,tr ii n V2 4 o e w tr u l y h d b e r v os me e tn . h h u bdt a d U 5 f h ae a i e n i o e t y t q ta mp d o x t e
铜绿微囊藻(Microcystis aeruginosa)次生代谢物对普通小球藻(Chlorella

D O I 10. 1 8 3 0 7 /2 0 1 8 .0 3 1 5
© 2 0 1 8 b y Journal o f Lake Sciences
铜绿微囊藻次生代谢物对普通小球藻 ( . // o $ / /a i+ / g a $ s ) 生 长 及 有 效 量 子 产 率 的 影 响 !
A b s tr a c t: A lle lo p a th ic interactions
among
p h y to p la n k to n
species are re
im portant factors
dom inance and succession
FANG Tingxuan1,2 & M A Zengling1** ( 1 : ZhejiangProvincial Key Laboratory for Subtropical Water Environment and Marine Biological Resources Protection,
方 婷 轩 “ 2 , 马 增 岭 1!! ( 1 $温州大学生命与环境科学学院,浙江省亚热带水环境与海洋生物资源保护重点实验室,温 州 3 2 5 0 3 5 " ( 2 : 浙江省环境监测中心,杭 州 3 1 0 0 1 2 )
摘 要 :浮游植物间的交互化感作用被认为是自然水体中浮游生物群落演替及优势种转换的主要因素之一. 铜绿微囊藻
o logica l role o f th is m olecule
in the
aquatic ecosystem is s till
铜绿微囊藻磷代谢过程研究_杨柳燕

;? 作为空白样供比色用# 另 " ;? 样品中加入 " ;? - ;<1 ’ ?@" *L1 水浴加热 ". ;BC$ 取出后将 )* 值调至 中性$进行比色# "#-#. 藻中糖原的测定 ’ ;BC@" 离心 ". ;BC$ 弃上 取 -0 ;? 藻液$, 000 : 清液$加入 " ;? 三氯乙酸抽提$, 000 : ’ ;BC@" 离心 ". ;BC$取上清液$加入 ’ ;? 无水乙醇$ 在冰箱中放置# 隔夜取出后 "- 000 : ’ ;BC@" 离心 ’0 ;BC$弃上清液# 烘 干后$加入 " ;? 蒸馏水$溶解后加入 ! ;? 0#-%蒽酮 溶液$水浴加热 ". ;BC# 在 ,-0 C; 处比色# 糖原的标 准曲线&取 " ;> ’ ;?@" 的葡萄糖标准储备液配制成不 同浓度的葡萄糖标准溶液$加入 ! ;? 蒽酮溶液$水浴 加热 ". ;BC 后$在 ,-0 C; 处比色# "#-#, 水中可溶性磷的测定 藻液离心后取上清液$放入比色管比色# "#-#G 生长曲线 本试验用 3BP<C"0- 型显微镜$取 "0 个不重叠的 视野$计数藻细胞数目$求其平均值$再计算出培养液 中藻密度#
第 !" 卷第 !期
农
业
环
境
科
学
学
报
,+G
" 材料与方法
"#" 试验藻种 铜绿微囊藻 !!"#$%#&’("’ )*$+,"-%’)" 由中科院武 汉水生生物研究所提供# 用改良的 $% 培养基培养&’($ 初始 )*+#,$培养在 -. #和 - -/0 12 光照培养箱中# "#- 试验方法 "#-#" 试验设计 采用 $% 培养基预培养铜绿微囊藻至对数生长 期末$试验用培养基为用 34*-56! 代替甘油磷酸钠的 改良 $% 培养基$并设不同的磷浓度梯度$ 将铜绿微 囊 藻 加 入 到 这 种 改 良 的 $% 培 养 基 中 $-. # 时 在 789:;<=<:;4!.+, 光照控温培养箱中进行 "- 8 $"- 8 光照黑暗交替培养# 隔日取样$测定铜绿微囊藻的生 长曲线$以及藻中总磷%可溶性磷%聚磷 %糖原和水中 可溶性磷浓度$了解铜绿微囊藻磷代谢过程# 不同磷 形态的标准曲线绘制方法如下& 用 .0 ;> ’ ?@" 磷标准 储备液配制不同浓度的磷酸盐标准溶液$并按不同磷 形态的预处理方法进行处理$最后显色测定# 试验设 -个平行$平均值为试验结果# "A-#- 藻中总磷的测定 取 ’ ;? 藻液$, 000 : ’ ;BC @" 离心 ". ;BC$取上清 液用于测定水体中可溶性磷# 将沉淀物从离心管中用 蒸馏水取出 $ 加入 " ;? .D !;EF" 过硫酸钾 $"-" # 消 化 ’/ ;BC# 放入 "/ ;? 比色管中加水至 "/ ;?$加入 /A-. ;? "/D抗坏血酸和 /A. ;? 钼酸盐溶液后 $显色 ". ;BC$在可见光 G// C; 处$用 G.- 紫外光栅分光光 度计比色# "A-A’ 藻中可溶性磷酸盐的测定 ’ ;BC H" 离心 ". ;BC$弃上清 取 ’ ;? 藻液$, /// : 液$ 加入 " ;? "0%三氯乙酸抽提$, 000 : ’ ;BCI" 离心 ". ;BC$取上清液# 加入 " 滴酚酞$加 346* 直到溶液 呈粉红色 $然后加 " 滴 *-J6! 使溶液呈无色 $ 此时溶 液呈中性$再进行比色# "#-#! 藻中聚磷的测定 取 ’+ ;? 藻液$, 000 : ’ ;BC@" 离心 ". ;BC$ 弃上 清液 $ 加入 .#!D 碱性高氯酸 $ 在 ’0 # 恒温培养箱中 放置 K0 ;BC# 然后 $", 000 :’ ;BC @" 离心 0#. 8$ 用 " ;?"#. ;<1 ’ ?@"34L1M. ;;<1 ’ ?I" NO7% 溶液洗涤 - 次$ ’ ;BC@" 离心 -. ;BC# 加入 0#. ;? 蒸 每次均以 ", 000 : 馏水抽提 - 次$每次 , 000 : ’ ;BC@" 离心 ". ;BC# 取出 上清液$ 加入 ’ ;? 无水乙醇$"- 000 : ’ ;BC@" 离心 ’0 ;BC$弃上清液$烘干后加入 - ;? 蒸馏水溶解# 取出 "
一种水中铜绿微囊藻快速定量方法与流程

一种水中铜绿微囊藻快速定量方法与流程以一种水中铜绿微囊藻快速定量方法与流程为标题,写一篇文章。
水中铜绿微囊藻是一种常见的蓝藻,它对水体生态环境产生了严重的影响。
因此,快速准确地定量水中铜绿微囊藻是非常重要的。
本文将介绍一种有效的方法和流程来进行水中铜绿微囊藻的定量。
准备样品。
需要采集水样,并将其保存在透明的玻璃瓶中,避免暴露在阳光下。
样品的体积应该足够大,以确保结果的准确性。
同时,还需要准备一些标准品,用于后续的定量分析。
接下来,进行样品处理。
将采集到的水样倒入一个容器中,并加入一定量的溶液用于稀释。
通过稀释可以减少样品中的杂质对分析结果的影响。
同时,还需要进行一些预处理步骤,如过滤、沉淀等,以提高分析的准确性。
然后,进行样品测量。
可以使用光谱仪、显微镜等设备来对样品进行测量。
通过测量,可以获取到样品中铜绿微囊藻的光谱特征,进而进行定量分析。
在测量过程中,需要注意避免其他因素的干扰,如杂质的吸收等。
接下来,进行数据处理。
根据测量结果,可以使用一些统计方法来进行数据处理。
可以计算出样品中铜绿微囊藻的浓度,并与标准品进行对比,从而得到准确的定量结果。
同时,还可以进行一些数据分析,如相关性分析、趋势分析等,以了解铜绿微囊藻在水体中的分布情况。
进行结果分析和报告。
根据定量结果,可以分析水体中铜绿微囊藻的分布情况和变化趋势。
如果结果显示铜绿微囊藻的浓度超过了一定的阈值,那么需要采取相应的措施来控制其生长和扩散。
同时,还可以将定量结果制作成报告,用于后续的研究和管理。
水中铜绿微囊藻的快速定量是一项重要的研究工作。
通过合理的方法和流程,可以准确地定量水中铜绿微囊藻,并对其在水体中的分布情况进行分析和报告。
这对于保护水体生态环境和维护人类健康具有重要意义。
希望本文介绍的方法和流程能够对相关研究和管理工作提供一定的参考和指导。
洱海水华铜绿微囊藻生长特性的初步研究

【 A b s t r a c t J 0b j e c t i v e : T o s t u d y o n t h e g r o w t h l a w a n d s o m e p h y s i o l o g i c a l c h a r a c t e r i s i t c s o f Mi c r o c y s t i s a e r u g i n o s a i s o l a t e d f r o m E r h a i
N I Z h a o l i n 。 , WA N G T a o , Z Ha , Y A N G G u a n y i n g ‘ , Z HA N G Y i , S HE N Y u a n y i n g
[ 关键词 ]洱海; 铜绿微囊藻; 分离 ; 生长 曲线; 叶绿素a
[ 中图分类号 ] Q 9 3 9 . 9 [ 文献标 志码 ] A [ 文章编号 ]1 6 7 2 — 2 3 4 5 ( 2 0 1 3 ) 0 4 — 0 0 4 8 — 0 3
A S t u d y o n t h e Gr o wt h a n d S o me P h y s i o l o g i c a l Ch a r a c t e r i s t i c s o f Mi c r o c y s t i s a e r u g i n o s a I s o l a t e d f r o m Er h a i La k e
( 1 . C o l l e g e o f P u b l i c He a l t h , D l a i U n i v e r s i t y , D l a i , Y u n n a n 6 7 1 0 0 0 , C h i n a ;
铜绿微囊藻生长特性研究

图 1 pH 对铜绿微囊藻生长的影响 Fig. 1 The effect of pH on the growth
of M icrocystis aeruginosa
的生长[ 13] , 与在该试验中得到的结果不尽相同。 试验利用光照恒温生物摇床模拟扰动条件, 转速
为 30 rPmin。除静止与摇动的差别外, 2 组实验其他条 件均保持一致, 试验结果见图 2。由图 2 可见, 无论从 生物量还是从比增长率上看, 扰动对铜绿微囊藻的生 长影响的唯一表现为生长滞后, 扰动组达到最大现存 量和最大比增长率的时间比静止培养延迟约2~ 3 d, 但从最大现存量与最大比增长率的量值上看并无显著 差异。产生滞后效应的原因可能是由于藻种的培养一 直处于静止状态, 因此在扰动组中经过 2~ 3 d 的适应 阶段后, 铜绿微囊藻仍保持与静止类似的生长过程。
在分析营养盐浓度的影响时, 应用了 Monod 模型 进行动力学分析, 动力学参数的计算方法如下[ 12] :
采用 Leneweaver- Burk 作 图 法, 将 Monod 方 程 式 L= Lmax CP( C + K s ) 变成以下形式:
1 L=Βιβλιοθήκη Ks Lm ax1 C
+
1 Lmax
式中, L 为微生物的比增长率, d- 1 ; Lmax 为限制底物浓
第 17 卷 增刊
环境 科学研究 Research of Environmental Sciences
Vol. 17, Suppl. , 2004
铜绿微囊藻生长特性研究
金相灿1 , 李兆春2, 郑朔方1 , 杨苏文1, 胡小贞1 , 储昭升1
( 1. 中国环境科学研究院 湖泊生态环境创新基地, 北京 100012; 21 中国纺织工业设计院, 北京 100037)
红球菌对铜绿微囊藻的裂解动力学

红球菌对铜绿微囊藻的裂解动力学
红球菌对铜绿微囊藻的裂解动力学是一种有效利用发酵技术去处理水中的有害物质的方法,其主要作用是通过红球菌的代谢产物去分解铜绿藻的微囊壁来降低污染物的浓度。
研究表明,红球菌可以分解微囊壁中的有机物质,如半乳糖、葡萄糖和碳水化合物,从而减少水体中污染物的浓度。
红球菌对铜绿藻微囊壁的裂解动力学可以用一种叫做离子交换的方法来实现。
这种方法是在红球菌膜上形成一个弱酸性的环境,使得铜绿藻微囊壁中的有机物质能够被红球菌所吸收、分解和转化。
当红球菌的细胞壁上的离子交换酶(如磷酸酶和糖酶)开始工作时,细胞内的H+离子会和铜绿藻微囊壁中的有机物质结合,从而将铜绿藻微囊壁中的有机物质分解成葡萄糖和其他简单的有机物质,从而降低水体中有害物质的浓度。
红球菌对铜绿藻微囊壁的裂解动力学也可以利用其他方法实现,比如使用微生物的膜外释放机制来改变铜绿藻微囊壁的结构,使得红球菌能够有效地分解微囊壁中的有机物质。
此外,还可以使用特定的生物酶,如葡萄糖苷酶和磷酸脱氢酶,来加速铜绿藻微囊壁的分解过程,从而减少污染物在水中的浓度。
红球菌对铜绿藻微囊壁的裂解动力学是一种很有效的处理水中污染物的方法,因为它能够有效地减少污染物在水体中的浓度,并且不会对水体的质量产生副作用。
此外,红球菌对铜绿藻微囊壁的裂解动力学还可以提高水的质量,因为它能够把有害物质转化为有益的生物物质,这些生物物质可以作为水体的营养物质,促进水体的生物多样性。
铜绿微囊藻生物钟蛋白的节律检测及其相互作用蛋白的筛选的开题报告

铜绿微囊藻生物钟蛋白的节律检测及其相互作用蛋白的筛选的开题报告题目:铜绿微囊藻生物钟蛋白的节律检测及其相互作用蛋白的筛选背景:铜绿微囊藻是一种单细胞蓝藻,它具有昼夜节律,可以适应不同的环境条件。
其节律是由生物钟基因控制的。
生物钟基因编码的蛋白质包括时钟基因、时钟蛋白和调节蛋白。
这些蛋白质的相互作用是维持生物钟节律的关键。
因此,研究铜绿微囊藻生物钟蛋白的节律检测及其相互作用蛋白的筛选,对于深入了解其节律调节机制具有重要意义。
研究目的:本研究旨在探索铜绿微囊藻生物钟蛋白的节律检测方法,并筛选其相互作用的蛋白。
研究内容:1. 铜绿微囊藻生物钟蛋白的节律检测方法建立。
采用实时荧光定量PCR技术检测不同时间点的RNA表达量,建立铜绿微囊藻生物钟蛋白的节律检测方法。
2. 铜绿微囊藻生物钟蛋白的相互作用蛋白筛选。
采用酵母双杂交技术筛选铜绿微囊藻时钟蛋白与调节蛋白的相互作用蛋白。
预期结果:1. 建立一种可靠的铜绿微囊藻生物钟蛋白的节律检测方法。
2. 筛选出铜绿微囊藻生物钟蛋白的相互作用蛋白,深入了解其节律调节机制。
研究意义:1. 对于深入了解铜绿微囊藻的生物学特性具有重要意义。
2. 为揭示其节律调节机制提供了重要的实验依据。
3. 可为其他相关领域的研究提供参考。
研究方法:1. 采用铜绿微囊藻培养技术,培养铜绿微囊藻。
2. 采用实时荧光定量PCR技术检测不同时间点的RNA表达量,建立铜绿微囊藻生物钟蛋白的节律检测方法。
3. 酵母双杂交技术筛选铜绿微囊藻时钟蛋白与调节蛋白的相互作用蛋白。
4. 对筛选出的相互作用蛋白进行验证实验。
研究计划:1. 第一年:建立铜绿微囊藻生物钟蛋白的节律检测方法。
2. 第二年:筛选铜绿微囊藻生物钟蛋白的相互作用蛋白。
3. 第三年:对筛选出的相互作用蛋白进行验证实验。
4. 第四年:数据分析和论文撰写。
预期困难:1. 铜绿微囊藻的生长条件比较苛刻,容易受到环境条件的影响。
2. 酵母双杂交技术的误差率较高,需要进行精细优化。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
淡水铜绿微囊藻中无机砷的释放,转化,富集的研究引言:砷不仅是一种潜在的有毒类金属而且是一种环境污染物,人类经常接触到含有这种污染物食物,水,空气和土壤。
世界上,超过1.5亿都接触到含有超过国际卫生组织推荐标准10ug/L的砷。
天然水中砷浓度的变化是很大的,变化范围0.5ug/L~5000ug/L。
一些污染的淡水中砷的浓度高达20mg/L。
水体砷污染是一个普遍的、急切的问题,它需要我们采取立刻行动来改善水质。
生物体中砷的毒性和生物富集不仅取决于砷的总量而且也取决于砷的形态。
虽然自然环境中砷存在几种氧化形态,但是淡水中三价砷和五价砷的含氧离子比有机砷更常见。
人们普遍认为,除了三价甲基化代谢产物(如:单甲基胂酸和二甲基胂酸)和含巯基的五价甲基化代谢产物(如:巯基二甲基砷酸),在哺乳动物体内,无机砷比有机砷毒性更大。
水生系统内,主要的无机砷进入到微生物中,例如:浮游植物,并且进而被转化成甲基胂或者像As这样更高价态的有机砷。
由于藻类具有很高的从水环境中富集砷的能力,因此他们对水环境中砷旳生物富集和生物转化具有重要作用,并且决定着高等生物可利用砷的形态以及他们随后的转化。
最近的研究表明:不同的藻类对无机砷的吸收能力具有很大的不同。
在水生系统中,蓝藻细菌是最大、最重要的能够进行氧化光合作用的原核自养生物菌群之一。
,正如富营养化淡水中最常见的有毒蓝藻一样,铜绿微囊藻能够形成引起其他动物中毒的水花并且对人类的健康具有一定的风险。
由于藻类的不同生长阶段砷的主要形态不同,故砷的吸收随着藻类生长阶段不同而异。
至今,关于浮游植物对砷的吸收、富集和生物转化,由于五价砷的降解以及其代谢与磷的关系的详尽研究,多数研究已经关注海洋生物,尤其是真核藻类。
鲜为人知的是,淡水中有毒藻类的水花不同无机砷形态的生物转化之间的比较,特别是在由蓝藻引起的水花期间,形态转化和释放过程的比较。
如果这种水被饮用,它将对人类和其他生物存在潜在的环境风险,因此,我们有必要了解它们的相关性。
因此,我们研究的主要目的是调查砷在铜绿微囊藻中的生物转化和生物富集以及它们向不同浓度砷污染水体的释放。
用生长研究来确定无机砷对铜绿微囊藻的毒性效应。
对于了解砷的生物转化机制以及预测因砷污染引起的蓝藻爆发的风险,藻类和环境中砷的形态的变化是重要的指标。
结果:藻类的生长和无机砷的毒性无机砷对藻类(铜绿微囊藻)的毒性效应是明确的。
藻类生长的抑制率随砷浓度的增加而增加。
这种藻类对砷表现出极强的耐受性,对三价砷的72-h IC50值为3.582uM,五价砷72-h IC50值为133uM,这表明对于铜绿微囊藻三价砷的毒性比五价砷的毒性更强。
72-h IC50比预期的环境中砷的浓度具有更高的数量级。
图表一反映了在不同浓度下,铜绿微囊藻的叶绿素a浓度和藻类细胞密度随时间的变化。
在15天的培养期内,空白组与不同浓度砷处理组叶绿素a浓度和藻类细胞密度有很大的差异。
叶绿素a浓度和藻类细胞密度都随对数生长期孵化时间的延长而渐渐地增加,这表明:在此浓度范围内,五价砷这种藻类没有明显的生长刺激作用或者致毒效应。
在空白对照组和5~20uM不同浓度的三价砷处理下,叶绿素a浓度和藻类细胞密度没有明显的变化。
然而,50uM的三价砷处理组在起始阶段就抑制了藻类的生长。
虽然与空白组和其他处理组相比,细胞数量和叶绿素a的总量明显的减少,但是它在第七天开始微弱的重新生成并且随后成倍数增长。
在100uM三价砷处理7天之后藻类基本全部死亡,这表明:藻类对三价砷是极其的敏感。
铜绿微囊藻对砷的富集藻类细胞内砷的浓度在15天的三价砷处理下比其在五价砷处理下要高很多。
对照组,藻类中砷的浓度只有4ug/g,这表明最初的藻类并没有被砷污染。
在15天的孵化期内,铜绿微囊藻富集的总砷量是与添加在环境中砷的浓度的增加成比例的。
细胞内砷的量在五价砷处理下最大值为96ug/g,而在三价砷处理下其最大值为260ug/g,明显要高很多。
在15天之后,细胞内的砷占添加在环境中总砷的百分数,100uM五价砷处理的能够达到12%而50uM三价砷处理的为21%。
由于在100uM三价砷处理下,藻类在5天之后就死亡了,故没有此条件下细胞内砷的数据。
砷吸收的动力学常数用拟合双曲线模型来表示,而砷富集的数据用非线性回归模型来表示。
砷吸收能够用米氏方程来很好的描述,三价砷的R²值为0.940,五价砷的R²值为0.973。
米氏计算表明:三价砷的最大吸收率是五价砷的两倍还多。
三价砷的半饱和浓度为27.66±6.484而五价砷的为25.15±6.241。
砷的四种形态五价砷、三价砷、甲基胂和二甲基胂在不同的三价砷处理下在藻类细胞内都能够检测到。
然而,五价砷处理的没有发现甲基胂,而三价砷处的有0.10~0.15ug/g的甲基胂。
砷的形态分析表明:三价砷或者是五价砷处理下五价砷是主要的形态占总砷的78-93%,而另一种形态的无机砷三价砷是次要的形态,占总砷的6-21%。
另外,在不同的处理下少量的有机砷占总砷的量不到1%。
这种藻类从三价砷溶液中富集五价砷的量比从五价砷溶液中富集的相对较多。
细胞对砷的富集与添加在环境中三价砷和五价砷的量呈比例的。
细胞中而甲基胂的量在三价砷和五价砷处理下是相同的,并且是一个稳定的值10uM。
与五价砷的富集相反,五价砷处理下的三价砷的富集变化很大8.9~20.3ug/g,与空白对照组也明显不同。
然而,在三价砷处理下,三价砷富集的趋势与细胞内五价砷浓度很相似,它随三价砷的增加而明显增加。
生长环境中砷形态的变化尽管在含有三价砷(约15%)的生长培养基中发现了少量的ASV,在后15天的潜伏期空白对照组的藻类中没有发现任何砷形态的转化。
这证实,在整个测试期间,它们在生长培养基中的相对稳定性。
只有微量三价砷分别存在于含有铜绿微囊藻试验介质,并在五价砷处理组前5天随时间增加而减少。
三价砷从最初的五价砷还原而来,并且随着最初的五价砷浓度的增加而增加。
然而,在空白组没有发现三价砷,这表明:含有铜绿微囊藻的培养基中的三价砷来自于生物还原作用。
在其他日子里,培养基中三价砷的还原也微弱的发生着。
至于五价砷的处理组,五价砷的浓度随着时间的增加而减少。
虽然最低值是在第13天出现的,五价砷在15天的内保持相对稳定并在培养基中占主导地位。
在这些培养基中甲基砷包括单甲基胂和二甲基胂也能检测到更低的浓度。
另外,它们都随时间的增加而增加,并且在不同的浓度下没有显著的差异。
在含有三价砷的培养基中,砷的形态发生着明显的变化。
随着藻类的指数增长,三价砷的浓度随着时间而迅速减少,并且由于生物和化学氧化作用使三价砷被氧化成五价砷,所以三价砷在第七天几乎就检测不到了。
然而,培养基的最初阶段五价砷迅速出现,然后在第七天成为主要的形态。
实验最后,溶液中99%的砷以五价砷的形式存在。
砷的另外两种形式,甲基胂和而甲基胂,在第五天可以同时检测到,在含有三价砷不同浓度的培养基的15天潜伏期内,它们都随时间的增加而增加。
另外,培养基中而甲基胂的浓度比甲基胂的高。
实验中,我们发现两种还原产物的总量与最后培养基中砷的总量有很大的差异,这表明:还存在其他砷的不明形态向溶液中释放。
很明显,有很少的挥发性砷的损失,我们可以用古老的砷斑法测得。
从图6a中可以看出,三价砷的释放率从4.7×10-³to 4.3×10-¹μM/g/天,随着五价砷的浓度的增加而急剧增加。
这表明,三价砷的主要是由生产五价砷处理过的铜绿微囊藻产生和释放的。
单甲基砷的平均释放率在10uM的五价砷下达到峰值 2.6×10−2 μM/g/天,而而甲基胂的平均释放率在10uM的五价砷下达到峰值7.7×10−2μM /g/天。
显然,我们在五价砷处理的溶液中前五天仅发现了三价砷,其平均释放率比有机砷要高,尤其是在第二天。
从图6b中可以看出,在15天的潜伏期内,五价砷,甲基胂和二甲基胂的平均释放率都随三价砷浓度的增加而增加,特别是三价砷的浓度为50uM时。
这表明砷的甲基化与生物氧化是同时发生的。
讨论铜绿微囊藻对砷的富集在此研究中,测试培养基中铜绿微囊藻对砷的富集随着三价砷和五价砷的浓度的增加而增加。
主动吸收在藻类对砷的富集有主要的作用。
确实,相似的研究已经记录了聚球藻和小球藻的富集作用。
三价砷的最大值是五价砷的两倍多,这表明:作为磷的类似物,在过量的磷存在的条件下,铜绿微囊藻对砷的吸收很难进行。
由于三价砷在pH为6.0时不溶的中性分子存在,所以它能够通过一些植物的水通道蛋白进行转化。
因此,这项研究发现在高浓度的磷溶液中,三价砷的吸收率比五价砷的高。
随后,与五价砷处理的相比,三价砷处理组细胞内砷的浓度明显提高了。
另外,铜绿微囊藻比其他淡水藻类对砷具有更强的生物富集能力。
结果显示:铜绿微囊藻细胞内超过7%的砷是在第十五天从培养基中吸收的。
的确,以前的报告表明:聚球藻和小球藻能够从被砷污染的环境中富集更多的砷。
结论是:由于水花中砷的生物富集对人类和动物具有潜在的危害,所以我们应该给予更多的关注。
铜绿微囊藻中砷的形态细胞内砷形态的分布清楚的证明:用5~100uM的三价砷或者五价砷处理15天之后,五价砷是主要的形态,接着依次是三价砷,二甲基胂和甲基胂。
此研究也表明:在富含三价砷和磷的淡水中,藻类细胞内的主要转化形式是三价砷到五价砷的生物氧化。
:聚球藻和小球藻也存在相似的转化。
Meada有相似的发现:淡水中的聚球藻95%的砷是以无机态的形式而富集,甲基胂化合物则主要是二甲基胂。
在我们的研究中,用不同浓度的三价砷或者五价砷处理15天之后,细胞内砷的形态发生了明显的变化。
随着添加的砷浓度的增加,细胞内砷的主要形态五价砷也迅速增加。
正如Maeda在聚球藻的调查中推断的,甲基胂的含量是藻类富集的限制因素。
细胞内的砷在三价砷处理后有一个相似的趋势,而五价砷处理后急剧波动。
然而,与三价砷处理的相比,五价砷处理组的细胞内三价砷的富集量占的比例更大。
这些结果表明:铜绿微囊藻把五价砷生物转化为还原态的能力并作为甲基化的反应物。
培养基中砷形态的变化含有无机砷的培养基中砷形态的迅速变化表明:铜绿微囊藻能够氧化、还原、甲基化并且向培养基中释放砷。
虽然一部分砷能够进入到细胞里,但是大部分的砷明显的被代谢并被释放到周围的水中。
正如Sanders和Cullen记录的那样,最初阶段,五价砷到三价砷的还原是非常迅速的,它与接触到的五价砷的浓度呈正相关。
培养液中三价砷的一个峰值出现在最初阶段,比Hellweger 记录的水花出现的要早。
人们普遍认为,培养基中有机砷的量取决于甲基化/二甲基化率以及释放过程。
培养液中发现了甲基胂而细胞中几乎没有,这表明:无机砷的甲基化过程也是含五价砷的培养基中甲基胂和二甲基胂的释放过程一样迅速。