杂种鹅掌楸花粉发育过程中细胞游离Ca 2+的动态变化
鹅掌楸属花粉的超微结构研究及系统学意义

鹅掌楸属花粉的超微结构研究及系统学意义
研究发现,鹅掌楸属花粉是异被子植物,具有自身独特的外形结构,其花粉颗粒表面宏观形状具有类似鹅掌状的特征。
鹅掌楸属花粉超微结构受到科学家特别关注,其超微形态特征是异被子植物系统发育的重要方面。
关注鹅掌楸属花粉超微结构的研究,有助于全面了解与评价鹅掌楸属植物的多样性和系统学意义。
为了更好地研究鹅掌楸属花粉的系统学意义,科学家们对花粉形态特征进行了详细的分析和比较,结果发现鹅掌楸属花粉宏观形态和超微形态存在一定的差异,宏观形态及其表面韧皮特征在植物属间有很强的特化性,但超微形态的差异较小,表明被子植物在远古时代就已经形成,后来受微环境影响仅在总形上起到细微差异化作用。
此外,研究还发现,鹅掌楸属花粉的超微结构是其生态和进化之间不断连接的关键点,这表明其适应性能强,耐受性强,可以快速适应不同环境条件。
从系统发育角度看,花粉结构在植物演化史上显示出稳定性,但它们也有一定的可塑性,可以对环境作出响应,这个可塑性是植物演化的保证。
总之,鹅掌楸属花粉的超微结构研究丰富了我们对植物进化史的认识,有助于揭开植物系统发育和环境因素之间的关系。
鉴定和研究各个种类的花粉超微结构不仅有助于了解植物动态发育状况,而且也为识别物种和传播有利信息提供深入的义理根据。
渐危植物鹅掌楸的授粉率及花粉管生长

渐危植物鹅掌楸的授粉率及花粉管生长大多基于人工授粉是否能提高结实率来推断,从而很大程度上忽视了自然居群中可能存在的花粉竞争这一因素(Snow & Spira,...不同来源的花粉在柱头上均可萌发。
从花粉管的长度看,只有少数花粉管可在花柱中继续伸长,大多数的花粉管伸长约20~40 ...黄双全郭友好(武汉大学生命科学学院武汉 430072)陈家宽(复旦大学生物多样性研究所上海 200433)POLLINATION RATES AND POLLEN TUBE GROWTH INA VULNERABLE PLANT,LIRIODENDRON CHINENSE(HEMSL.) SARG.(MAGNOLIACEAE)HUANG Shuang-Quan GUO You-Hao(School of Life Sciences, Wuhan University, Wuhan 430072)CHEN Jia-Kuan(Institute of Biodiversity,Fudan University, Shanghai 200433)Abstract The limited seed production of insect-depended plant, Liriodendron chinense was once considered to be pollen-limited and insufficient cross pollination. In this study, we counted pollen grains deposited on stigmas in three populations in Guizhou, Hunanand Zhejiang provinces of China respectively. Over 61.9% stigmaswere pollinated. From 1994 to 1996, the mean number of pollengrains on each stigma ranged from 4.4 to 42.6, much more thanovules(2) in each pistil. Based on observations of three years,both the pollination rate and pollen quantity on stigmassignificantly affected seed set. When flowers opened without stamens dehiscencing at the early stage of anthesis, stigmas received considerable quantity of pollen grains. Pollen grainsfrom different sources were able to germinate and pollen tubegrowth rates were not greatly variable. It is very likely thatpollen grains arrived first would fertilize eggs. Since onlyseveral pollen tubes went through the stylar canal, the potential pollen competition may exist. In this case, there would be strong selection on floral syndrome which benefit early insect visits. Pollen grains from the early visits would have a greater chanceto fertilize ovules than those from later visits, which impliesthat cross pollination is the predominant breeding system of this plant. The conclusion was also confirmed by following four artificial experiments. Three treatments, including flowers bagged, netted or with the perianth removed, all reduced seed set clearly, but flowers with the stamens removed (emasculation)had a higher seed production by open pollination. As the rates of deposited stigmas in three populations were 6~8 times more than full seed set, we consider that lower seed production in this plant may not mainly be due to pollen limitation.Key words Liriodendron chinense;Pollen deposition;Pollen competition; Breeding system摘要对鹅掌楸Liriodendron chinense 3个自然居群连续3年直接统计落置柱头上的花粉量,其中61.9%以上的柱头接受了花粉。
鹅掌楸苗期生长杂种优势的SSR分析

基 金 项 目 :国家 自然科 学基金 项 目(0731 ; 3929 ) 国家 “ 十一 五 ” 技 支撑课 题 (O6 A oA 6 ; 科 20B D l 1 )江苏省 高 校 自然 科学 重大项 目(0 J 201 ) 1K A207
。
李 火 根 为 通 讯作 者 。
5 8
林
业
科
学
2 . c n ee g r sR sac s tt,C i s c d m r i l g i l rl c ne W n ig5 3 ) ea d B v r eC o e r I tue hn e a e yo To c r ut a i c a nn 7 3 a p e hni e A f p aA c u S e 1 5
J, ) 0 5年 1 月 初 采 种 , 时采 集 上 述 9个 。J L 。2 0 X 1 同
亲本 的 自由授粉 种 子 ( 和 C H 未 采 集 到 自由授 粉 种子 ) 0 6年 1月 播 种 于 南 京 林 业 大 学 校 园 苗 。2 0 圃 。2 0 0 7年 3月 在镇 江扬 子 江 林 业 有 限公 司苗 圃
W an i y ng‘ g X ao a ’ Li Huo n ge
( . e a oaoyo oe r G n t s n e eE gn ei Na n oe r U i r t N n 1 0 7 1 K y L br t r fF r t e e c a d G n n i r g, g F r t n esy s y i e n s y v i a g2 03 ;
第4 7卷 第 4期
20l1年 4 月
林
业
科
学
Vo . 1 47. . No 4
北美鹅掌楸种子老化过程中的生理生化变化

Phy il g c la o h m ia. a g n Ar i ca i o e s soo ia nd Bi c e c 1Ch n e i tf ilAgng Pr c s i
o io e d o uii r ed f r d n rn tl f aS e L i pe
维普资讯 Biblioteka 第3 6卷第 3期
西
部
林
业
科 学
Vo _ 6 No 3 l3 . S pt 2 0 e .07
20 0 7年 9月
J u n lo e tC i a F r sr c e c ora f W s h n o e ty S in e
G O Y n .ig , H N Y n .a Y a gya L N a - n U o gqn S E ogbo , U F n .u n , A G N nj u
( . u nnAcd m f oet ,K n igYun n60 0 , R hn ;2 Naj gF r t ie i ,N nig ins 10 7 R C ia 1 Y n a ae yo rsr u m n na 5 2 F y 4 C ia . ni oe r Un rt n s y v sy aj agu20 3 , nJ . hn )
液的相对 电导率与其的 S D酶活性和 C T酶活性呈负相 关 ,与 MD O A A含 量呈正 相关 ;北美 鹅掌楸 种子经人 工老 化处理后 ,其种子膜脂的过氧化作用加强 ,膜 的结构和功能遭到破坏 。 关键词 :北美鹅掌楸种子 ;老化过程 ;生理生化变化 中图分类号 :Q9 9 77 1 4 .4 . 文献标识码 :A 文章编号 :17 62—84 (0 7 3—0 8 o 2 6 20 )0 0 0一 4
用RAPD方法初探鹅掌楸的花粉流-1998

研究简报用RAPD方法初探鹅掌楸的花粉流黄双全 郭友好3 陈家宽 王青锋(武汉大学生命科学学院,武汉430072.3联系人)摘要 花粉流的标记方法是传粉生物学中的关键技术.两性花植物鹅掌楸的花粉传递可能为自花、同株异花和异株异花.从野外居群的传粉者运动观察和去雄等实验得知,落置柱头上的花粉主要是外源的异花花粉,但异株间、远距离的传粉是否存在还不确定.尝试用随机扩增多态DNA(RAPD)方法进行父系分析,标记花粉运动.结果表明这一濒危植物的花粉散布可达100m,初步显示出鹅掌楸的有效居群范围较大.迅速简捷的RAPD标记方法可能成为一种广泛适用的花粉流标记技术.关键词 鹅掌楸 随机扩增多态DNA 花粉流花粉流是导致植物中基因流动的重要因素,花粉运动的走向影响着居群的繁育系统和遗传结构.鹅掌楸(L i riodendron chi nense(Hemsl.)Sarg.)是第四纪冰川遗留下来的孑遗物种,属国家二级保护植物.其自然居群普遍较小,个体间间距大[1].测定花粉流的散布范围是了解其居群遗传结构和确定有效居群大小的重要基础,对制定合理的保护措施有直接的指导意义.1 材料与方法 (ⅰ)实验材料. 1996年在贵州省松桃县长萍乡康金村鹅掌楸自然居群中,观察到3个开花成株记为K1,K2,K3.K1与K2间距约100m,K1与K3间距为14m.1997年8月取成株的嫩叶(硅胶保存带回),和K1株1996年结实的种子在武汉大学苗圃出生的14株幼苗的嫩叶,置-70℃保存备用.(ⅱ)实验方法. 用Murray和Thompson[2]方法稍加改动,提取总DNA.先用3个成株(亲本)的DNA样品进行PCR扩增以筛选引物.随机筛选了Operon公司OPH~OPN和OPQ组中的134个随机引物,其中在3个亲本间OPH207,OPJ213,OPJ219,OPL207,OPQ204和OPQ206共6个引物有较好差异.用这6个引物再扩增亲代和子代的17个DNA样品.扩增反应在PE公司的DNA Thermal Cycler480上进行.反应体积为25μL,内含50ng的引物,约30ng的模板,100mmol/L Tris2HCl(p H=8.3),25mmol/LMgCl2,1U Tag DNA聚合酶(购自Sangon公司).扩增程序为94℃,3min;随后94℃,1min;38℃,1min;72℃,1.5min, 40个循环;72℃延伸8min后转入4℃保存. 1.4%的琼脂糖凝胶(EB+)中电泳.2 结果和讨论 花粉流的标记方法一直困扰着传粉生物学家,至今已有多种方法来尝试研究植物花粉的走向.大致可分为两类:(ⅰ)间接标记方法.如荧染和金属粉尘标记,花粉粒的化学标记,传粉者运动;(ⅱ)直接方法.如人工模拟,统计柱头上花粉粒,子代分析(即在子代中寻找亲本居群中非随机分布的标记基因)[3,4].鹅掌楸为两性花,花粉传递可能为自花、同株异花和异株异花传粉.花部的综合特征适应于虫媒传粉,风散布的花粉基本上在10m以内且数目极7151少.鹅掌楸的风媒传粉作用可以忽略,花粉传递依赖于昆虫.昆虫访花行为的观察和统计柱头上的花粉量[5]表明,鹅掌楸的花在刚开放(此时花药未裂,但花瓣分泌了大量蜜汁),就有昆虫访问且访问频率比花开后期高.检测花开而花药未裂时期的柱头,其上已有较高比例的花粉,显然这些花粉是来自异花的;人工去雄后柱头上也落置了鹅掌楸的花粉,证实了异花传粉的存在[5].连续3年的昆虫访花观察,在所选的贵州居群有19种访花昆虫,包括蜂类8种、蝇类4种和甲虫7种.尽管观察到一些体型较大的蜂类在访问一株鹅掌楸后飞向另一株,但仅通过传粉者的观察,是否导致异株间成功的传粉还不确定.在选取的3个亲本和14个子代中,K1为母本,K1,K2,K3均可提供花粉而作为父本.在随机引物OPQ205扩增产物中(图1),K2样品出现的特征带(箭头所指)在子代中出现.用OPH207扩增17个样品的产物中(图2),K2和K3共同拥有的特征带在选取的14个子代中的9个出现.其他筛选出的有较好差异的4个引物扩增结果与OPH27类似.这些结果证实了鹅掌楸异株传粉的存在,尽管K2与K1相距100m,但RAPD标记表明有花粉携带的基因从K2传递到K1.在已进行传粉生物学观察,分布于贵州、湖南和浙江的3个鹅掌楸自然居群中,贵州居群以蜂类为主要访花者;而湖南和浙江居群以蝇类为主,蜂类表现为偶尔的访问.从昆虫的访问行为来看,异株间的访问主要依赖于体型较大的蜂类,尤其是植株间距较大时.鹅掌楸居群间等位酶分析的结果和花部数量变异的观察也表明贵州居群的遗传变异性较其他居群高[6,7].上述结果表明,用RAPD方法标记贵州居群鹅掌楸花粉的散布范围可达100m,不同距离花粉流的强度和范围的精确测定有待于大量取样.比较传粉效率在居群间的差异(贵州居群最高、湖南居群次之、浙江居群最低),在鹅掌楸的自然或人工居群中引入适当的传粉者(如蜜蜂),不但可提高花粉传送效率,而且可提高花粉流的散布范围.图1 引物OPQ205(5′CCGCGTCTTG3′)扩增鹅掌楸亲代和子代的结果K1,K2,K3为亲本,1~11为子代图2 引物OPH207(5′CTGCA TCGTG3′)扩增鹅掌楸亲代和子代的结果点样顺序同图1自Schaal[8]用同工酶谱带的差异性检测了L upi nus texensis基因流范围的经典性工作,在自然居群中用子代分析法研究花粉散布范围的工作仍然很少.此方法要求在自然居群中找到稀有的基因型且工作量大.最近,Nilsson等人[9]用微型标记的方法为兰科植物花粉流的研究开辟了一条新路,但不太适合其他类群.Lonn等人[10]认为可用于野生居群的基因流和父系分析的两种方法中,人工合成的(TG)n多聚核苷酸序列作为探针标记方法比RAPD方法可靠,但是前者操作难度大、费用高.本文首次在自然居群中用RAPD标记进行父系分析的方8151研究简报研究简报法来研究花粉流.只要亲本个体间有一定差异,避免操作中可能的污染,迅速简捷的RAPD 标记方法将可能成为普遍适用的花粉流标记方法,同时扩增产物的结果提供了评价居群遗传多样性和真正繁育系统的必要信息.致谢 武汉大学遗传研究所朱英国教授、何玉卿博士后、关和新博士、杨征博士等在室内工作中提供了帮助,谢志雄博士提供了有益的建议,谨表谢忱.本工作为国家自然科学基金(批准号:39391500)资助项目.参 考 文 献1 郝日明,贺善安,汤诗杰,等.鹅掌楸在中国的自然分布及其特点.植物资源与环境,1995,4(1):1~62 Murray M G,Thompson W F.Rapid isolation of high molecular weight plant DNA.Nucleic Acids Research,1980,8: 4321~43253 Handel S N.Pollination ecology,plant population structure,and gene flow.In:Leslie Real ed.Pollination Biology.Orlando, Florida:Academic Press Inc,1983.163~2114 Richard A J.Plant Breeding Systems.London:G eorge Allen&Unwin,1986.135~1885 黄双全,郭友好,陈家宽.鹅掌楸的授粉率及花粉管生长.植物分类学报,1998,36(4):待刊6 朱晓琴,马建霞,姚青菊,等.鹅掌楸(L i riodendron chi nense)遗传多样性的等位酶论证.植物资源与环境,1995,5(3): 9~147 黄双全,郭友好,吴艳,等.鹅掌楸的花部数量变异与结实率.植物学报,1998,40(1):22~268 Schaal B A.Measurement of gene flow in L upi nus texensis.Nature,1980,284:450~4519 Nilsson L A,Rabakonandrianina E,Pettersson B.Exact tracking of pollen transfer and mating in plants.Nature,1992,360: 666~66910 Lonn M,Tegelstrom H,Prentice H C.The synthetic(TG)n polydinucleotide:a probe for gene flow and paternity studies in wild plant populations?Nucleic Acids Research,1992,20:1153(1998202205收稿,1998204207收修改稿)兔防御素NP-1基因在转基因烟草中的表达及其对烟草青枯病的抗性研究傅荣昭① 彭于发② 曹光诚① 陈彩霞②马江生① 张丽明① 李文彬① 孙勇如①3(①中国科学院遗传研究所,北京100101;②中国农业科学院植物保护研究所,北京100094.3联系人)摘要 从兔子肝脏细胞中扩增到防御素N P-1成熟肽的编码序列,长约200bp.将此片段插入到切除了GUS编码序列的pB I121质粒载体中,构建成植物表达载体pB IC-35SN P1.通过根癌农杆菌介导的叶盘法,将pB IC-35SN P1导入烟草细胞中,并获得了经Southern杂交证实的转基因植株.进一步的Northern杂交证明防御素N P-1基因能够在转基因植株中正常转录.抗菌实验表明转基因植株增强了对烟草青枯病的抗性并延缓了烟草青枯病的发病.关键词 防御素基因 转基因表达 烟草青枯病植物抗病基因(即R基因)的分离已取得了突破性进展[1].然而,这些R基因都是病源菌9151。
植物透射电镜样品制备

图3所示的多层结构即为药室内壁的胼胝质加厚现象。 可见新方法制备的样品能够提高植物样品的分辨率, 使超薄切片显示出更多的微细结构。
3.2 拟南芥
拟南芥是液泡组织较大的植物,在溶液中叶片难 以下沉,是较难固定的植物透射电镜样品。加之, 细胞壁结构坚硬,药物难以渗透,其样品制备更加 困难。若采用常规制备方法,则会出现叶片内部结 构不清晰、切片有颤痕和开裂等不良现象,不能进 行大视野的结构观察和摄影。笔者采用该方法制备 出了30多个此类透射电镜样品,整体信息丰富,结 构保存良好,没有出现效果不良现象。
3 结果
3.1杂种鹅掌楸花粉
细胞壁厚,组织坚硬,药物难以渗透,是典型 的制备难度大的透射电镜样品。采用常规制备方 法,超薄切片常会出现花粉内部结构不清晰、切 片有颠痕、污染等不良现象,甚至无法观察到样 品的超微结构,造成前功尽弃,只能从头再来, 进一步延长了工作周期。笔者采用延长锇酸固定 时间,减少树脂渗透浓度的新方法制备出了50 多个杂种鹅掌楸花粉样品,整体信息丰富,结构 保存良好,没有出现不良现象。
4 结果
4.1 一般情况下,植物样品用锇酸进行后固定,时 间延长可提高固定效果,但植物的种类较多,性 能区别较大,在后固定时需区别对待,对于迅速 发黑的植物样品,用锇酸固定时不易超过2小时, 否则,会造成样品发脆,难以操作,甚至破坏掉 已保存的结构。
4.2 在812树脂渗透的过程中,采用3:1;3:2; 3:3梯度渗透法,主要是提高样品的渗透效果。 根据植物的细胞壁厚,结构密集,药物渗透较慢 的特点,梯度渗透的时间间隔必须在2小时以上, 否则,影响渗透效果。
研究背景
超薄切片的分辨率高,信息丰富,是透射电镜样品制备方法 中最优秀、最常用、最基本的常规技术,从取材到染色有二十 几个步骤,任何环节出现问题都会影响到样品制备质量甚至导 致样品制备失败。加之,植物的细胞壁厚,结构密集,药物渗 透较慢,其制备难度就显得更大一些。
杂种鹅掌楸花粉败育的结构变异与分析

徐柏森 杨静 南京林业大学电子显微镜室
主要内容
1.鹅掌楸败育的细胞学特征及生理生化分析 鹅掌楸败育的细胞学特征及生理生化分析 2.小孢子不能正常发育的形式及原因 小孢子不能正常发育的形式及原因 3.在花药发育初期,绒毡层细胞的败育形态 在花药发育初期, 在花药发育初期 4.鹅掌楸花芽发育时期的 鹅掌楸花芽发育时期的RNA、蛋白质、氨 鹅掌楸花芽发育时期的 、蛋白质、 基酸总量呈下降趋势
结
论
3.杂种鹅掌楸花粉的败育现象严重,胼胝质加厚现象较多, 杂种鹅掌楸花粉的败育现象严重,胼胝质加厚现象较多, 杂种鹅掌楸花粉的败育现象严重 加之,结构密集,药物难以渗透,须延长固定时间,才能保 加之,结构密集,药物难以渗透,须延长固定时间, 存好花粉的微细结构。否则,会降低样品的信息量, 存好花粉的微细结构。否则,会降低样品的信息量,影响对 有关内容的全面把握和分析效果。 有关内容的全面把握和分析效果。 4. 植物在花芽分化时期需要大量的蛋白质去实现多种功能, 植物在花芽分化时期需要大量的蛋白质去实现多种功能, 如果蛋白质含量不足,很难在细胞中构成各种生物酶,同时 如果蛋白质含量不足,很难在细胞中构成各种生物酶, 也不能与细胞核中的DNA构成染色体,与RNA构成核糖体, 也不能与细胞核中的 构成染色体, 构成核糖体, 构成染色体 构成核糖体 以至细胞器在发育过程中不断失去应有功能, 以至细胞器在发育过程中不断失去应有功能,最终导致花粉 败育。 败育。
研究背景
杂种鹅掌楸的自然结实率很低, 一般在15 15% 杂种鹅掌楸的自然结实率很低 , 一般在 15% 以 单株自然结籽率不足1 下,单株自然结籽率不足1%,人工控制授粉虽可提 高结实率( 30% 但不稳定, 在无性繁殖试验中, 高结实率 ( 达 30% ) 但不稳定 , 在无性繁殖试验中 , 成活系数也不高, 成活系数也不高,近年来国内的有关专家在原有的 研究基础上, 研究基础上,积极扩大杂交亲本和加强有关杂种鹅 掌楸的杂种优势及优良无性系选育的研究, 掌楸的杂种优势及优良无性系选育的研究,为生殖 生物学提供了宝贵资料。我们在前人研究的基础上, 生物学提供了宝贵资料。我们在前人研究的基础上, 对败育现象较为严重的雄蕊发育过程进行了动态观 察研究,找出了杂种鹅掌楸花粉败育的主要原因, 察研究,找出了杂种鹅掌楸花粉败育的主要原因, 为保护世界濒危物种和进一步推广该树种提供了确 凿的理论依据。 凿的理论依据。
鹅掌楸种间杂交组合叶芽发育时期基因表达的差异

第3 9卷 第 5期
2 1 年 5月 01
东
北
林
业
大
学
学
报
Vo . 9 No 5 13 . Ma 01 y2 1
J 0UR NAL OF NOR HE T F T AS ORE TR UNI ERST S Y V IY
鹅掌楸种间杂交组合叶芽发育时期基 因表达的差异
i p - ( ad r etno l prns n p—I ( a d rsn i ol bt yr s f ioedo yr s nt e I b nspe n ny ae t)adt e I bn s eetn ny ohh b d )o Lr dnrnhb d y s i y p i i i Hx M adB Ja n ieet o t a ddvlp e t t e ,bt hr e os nf at ieecsi t el ( ad n Mx mogdf r wh n ee m n a s u teew r n i icn df rne p —I b ns ng r o sg e gi ny l pee tnb t hb d n n aet n p. bn s rsn i o l oeprn)o i edo yr sH M ad rsn’ o y r s do e rn)adt e1 ad eetn ny n aet f o n rnhb d x n 、 i h i a p y V( p Hr d i B J in cn ieecs nd eet l e e x rs o at n Mx .Sgi at f rne i rni n pes npt rs(I,Ⅱ ,Ⅲ ,Ⅳ )o r dnrnh b d x n i f d i f ag e i e f ioedo yr s M ad Li i H
杂交鹅掌楸纤维形态和化学组成的纵向变化规律

杂 交 鹅 掌 楸L i r i o d e n d r o n h y b r i d s是 以 鹅 掌楸属 L i r i o d e n d r o n两 个 现 存 种 (中 国 马 褂 木 L . c h i n e n s e( He ms l 1 S a r g . 和 北 美 鹅 掌 楸 . , / p / f e r a L . )为亲本 创造 的种 间杂种 ,由我 国著名林 木育 种 学 家 叶培 忠 教授 于 1 9 6 3年 首 次育 成 l l 1 。与其 亲 本 相 比, 该杂 种 在 生 长 速 度 和抗 逆 性状 等 方 面 均 表
d e c l i n e d s l i g h t l y a te f r 8 -9 y e a r s . Th e L. h yb r i d h a d t h e o p t i mu m i f be r mo ph r o l o g y a f t e r 8 - 9 y e a r s ;I n c o n t r a s t t o o t h e r f a s t — g r o wi n g wo o d, s uc h a s t r i p l o i d Pn pu l u s t o me n t o s a a n d Eu c al y pt u s , a n d t h e e x t r a c t i v e c o n t e n t o f L. h y b r i d s wa s l o we r , i t s a s h c o n t e n t wa s l e s s t ha n l %, t h e h o l o c e l l u l os e , l i g n i n a n d p e n t o s n a c o n t e n t s we r e 8 2 . 3 9 %, 2 2 . 7 1 %a n d 1 4. 02 % r e s p e c t i v e l y , wh i c h we r e s u i t a b l e f o r p ul pi n g t o g e t p ul p wi t h h i g he r yi e l d; Me a n wh i l e ,t h e L. h yb r i d s we r e s i mi l a r t o Ma s s o n p i n e i n l e n g t h — wi d t h r a t i o ,c e l l wa l l — l u me n r a t i o a n d
2023北京高三一模生物汇编:遗传与进化(选择题)
2023北京高三一模生物汇编遗传与进化(选择题)一、单选题1.(2023·北京东城·统考一模)某种伞形花科植物叶片含有对昆虫有毒的香豆素,经紫外线照射后香豆素毒性显著增强,乌凤蝶可以将香豆素降解,织叶蛾能将叶片卷起,取食内部叶片。
下列叙述错误的是()A.推测织叶蛾所取食的此种植物内部叶片中香豆素毒性较低B.乌凤蝶对香豆素降解能力的形成是基因定向突变的结果C.织叶蛾对该种植物叶片的取食策略是香豆素对其定向选择的结果D.植物的香豆素防御体系和昆虫避免被毒杀策略是协同进化的结果2.(2023·北京东城·统考一模)研究发现,AGPAT2基因表达的下调会延缓脂肪生成。
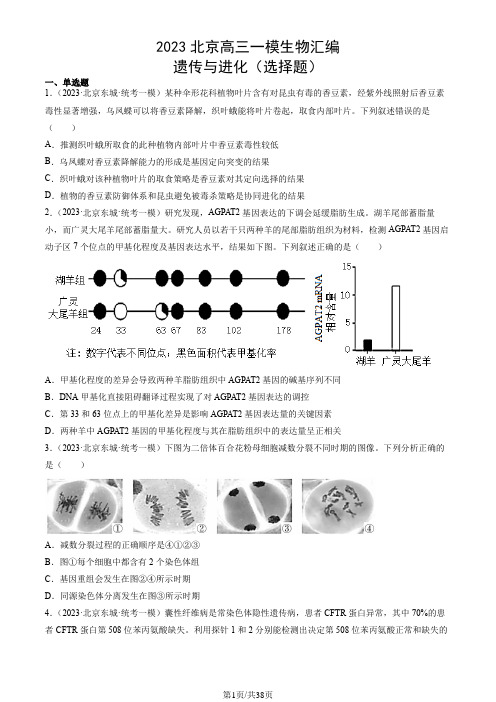
湖羊尾部蓄脂量小,而广灵大尾羊尾部蓄脂量大。
研究人员以若干只两种羊的尾部脂肪组织为材料,检测AGPAT2基因启动子区7个位点的甲基化程度及基因表达水平,结果如下图。
下列叙述正确的是()A.甲基化程度的差异会导致两种羊脂肪组织中AGPAT2基因的碱基序列不同B.DNA甲基化直接阻碍翻译过程实现了对AGPAT2基因表达的调控C.第33和63位点上的甲基化差异是影响AGPAT2基因表达量的关键因素D.两种羊中AGPAT2基因的甲基化程度与其在脂肪组织中的表达量呈正相关3.(2023·北京东城·统考一模)下图为二倍体百合花粉母细胞减数分裂不同时期的图像。
下列分析正确的是()A.减数分裂过程的正确顺序是④①②③B.图①每个细胞中都含有2个染色体组C.基因重组会发生在图②④所示时期D.同源染色体分离发生在图③所示时期4.(2023·北京东城·统考一模)囊性纤维病是常染色体隐性遗传病,患者CFTR蛋白异常,其中70%的患者CFTR蛋白第508位苯丙氨酸缺失。
利用探针1和2分别能检测出决定第508位苯丙氨酸正常和缺失的CFTR基因,对两个家系成员基因的检测结果如下图。
下列叙述错误的是()A.甲家系Ⅰ-1、Ⅰ-2、Ⅰ-1均含有决定苯丙氨酸缺失的CFTR基因B.若甲家系Ⅰ-2表型正常,用探针1、2检测出两条带的概率为1/2C.乙家系成员CFTR蛋白的第508位苯丙氨酸没有缺失D.探针1、2不适用于对乙家系Ⅰ-2的CFTR基因进行产前诊断5.(2023·北京西城·统考一模)在人类性染色体上,除了非同源区域外,还含有2个同源区域(PAR)。
鹅掌楸油细胞发育过程中超微结构的变化与挥发油产生的关系

鹅掌楸油细胞发育过程中超微结构的变化与挥发油产生的关系蔡霞;胡正海;何一【期刊名称】《西北植物学报》【年(卷),期】2002(022)002【摘要】鹅掌楸[Liriodendronchinense(Hemsl.)Sargent.]油细胞的发育过程可依据细胞壁的结构变化依次划分为3个阶段,即仅具初生纤维素壁层阶段、木栓质化壁层形成阶段和内纤维素壁层形成阶段.在发育早期,仅具初生纤维素壁层时,油细胞因其体积大,核仁显著,含极少淀粉粒和质体几乎无类囊体而与周围的组织细胞不同.对其3个发育阶段中内部结构变化分析表明,挥发油合成于细胞质和质体中.细胞质中,挥发油就以小滴形式产生,然后逐渐与油囊融合直接贮入油囊,与此同时,在各种细胞器中,质体的变化最为明显,质体中合成的嗜锇物质,随质体解体进入细胞质中,再经转化通过杯形构造积累入油囊.油囊中积累的油经OsO4染色后呈灰色,且分为2层,外层较内层深,推测与油的2种来源有关.【总页数】6页(P327-332)【作者】蔡霞;胡正海;何一【作者单位】西北大学植物研究所,西安 710069;中国科学院西安光机研究所瞬态光学技术国家重点实验室,西安 710068;西北大学植物研究所,西安 710069;西北大学植物研究所,西安 710069【正文语种】中文【中图分类】Q944.683【相关文献】1.烤烟叶片发育过程中栅栏细胞超微结构的变化 [J], 时向东;焦枫;范豪杰;韩永镜;刘国顺2.蟠桃果实发育成熟过程中果肉细胞超微结构的变化 [J], 李银;张辉;骆建敏;袁海英;薛珊珊;张茜3.玉兰油细胞发育和挥发油产生的超微结构 [J], 蔡霞;何一;胡正海4.杂种鹅掌楸花粉发育过程中细胞游离Ca2+的动态变化 [J], 尹增芳;宁代锋;李玉春;席梦利;施季森5.猫屎瓜乳汁道形成过程中显微和超微结构变化以及与橡胶产生的关系 [J], 胡正海;赵桂仿;田兰馨因版权原因,仅展示原文概要,查看原文内容请购买。
四川省达州市宣汉毛坝职业高级中学高二生物期末试卷含解析

四川省达州市宣汉毛坝职业高级中学高二生物期末试卷含解析一、选择题(本题共40小题,每小题1.5分。
在每小题给出的四个选项中,只有一项是符合题目要求的。
)1. 对胚芽鞘A作如图所示的处理,形成胚芽鞘B,光照后它的弯曲情况是()A.Q 处向左B.Q 处向右C.P 处向右D.不弯曲参考答案:D2. 松林防治松毛虫害的常用方法有招引灰喜鹊和施放松毛虫白僵病菌,灰喜鹊及白僵病菌与松毛虫的关系分别是A.竞争和共生B.捕食和共生C.捕食和寄生D.寄生和捕食参考答案:C3. 基因工程中,需使用特定的限制酶切割目的基因和质粒,便于重组和筛选。
已知限制酶Ⅰ的识别序列和切点是一G↓GATCC-,限制酶Ⅱ的识别序列和切点是—↓GATC—。
根据图示判断下列操作正确的是A.目的基因和质粒均用限制酶Ⅱ切割B.目的基因和质粒均用限制酶Ⅰ切割C.质粒用限制酶Ⅱ切割,目的基因用限制酶Ⅰ切割D.质粒用限制酶Ⅰ切割,目的基因用限制酶Ⅱ切割参考答案:D4. 按自由组合定律遗传的具有两对相对性状的纯合子杂交,F2中出现性状重组类型的个体占总数的 A. 3/8 B 5/8 C.7/16 D. 3/8或5/8参考答案:D5. 如图能正确表示内环境成分间关系的是()A.B.C.D.参考答案:【考点】E8:内环境的组成.【分析】内环境及其各组分之间的关系①内环境的概念:由细胞外液构成的液体环境叫做内环境,包括血浆、组织液和淋巴.②各组分之间的关系如图所示:C【解答】解:组织液、血浆和淋巴之间的关系如图所示:a、血浆和组织液之间的物质交换是双向的,毛细血管壁有一定的通透性,正常情况下除血细胞和大部分血浆蛋白外,其他物质都可以通过毛细血管壁;b、淋巴与组织液之间的物质交换是单向的,毛细淋巴管的功能是回收组织液,组织液中的一些物质一旦进入毛细淋巴管就成为淋巴,经过淋巴循环汇入血浆;c、淋巴和血浆之间的物质交换也是单向的,淋巴液在左右锁骨下静脉处流入血浆.故选:C.6. 下图表示内环境成分间的关系,正确的是参考答案:C7. 下列各组分泌腺产生的分泌物,均直接排放到内环境的是()A.唾液腺、垂体、肠腺 B.肾上腺、甲状腺、胰岛C.睾丸、汗腺、皮脂腺D.卵巢、胃腺、肝脏参考答案:B8. 右图表示某生态系统中4种成分之间的关系。
鹅掌楸属植物花粉生活力的检测

鹅掌楸属植物花粉生活力的检测
唐芸
【期刊名称】《广西林业科学》
【年(卷),期】2002(031)002
【摘要】用中国鹅掌楸(Liriodendron chinensis)和北美鹅掌楸(Liriodendron tulipifera)花粉材料,运用多种检测方法,检测花粉的萌发或生活力的情况,结果表明:(1)不同方法,花粉的萌发情况有很大的差异,其中看护培养的方法效果最佳;(2)看护培养北美鹅掌楸花粉的萌发率约为90%,说明本年度花粉和柱头是亲和的,花粉品质良好.
【总页数】3页(P80-82)
【作者】唐芸
【作者单位】广西林业学校,柳州,545003
【正文语种】中文
【中图分类】S718.3
【相关文献】
1.菊属植物花粉生活力检测方法的比较 [J], 赵宏波;陈发棣;房伟民
2.植物花粉生活力检测技术进展 [J], 左丹丹;明军;刘春;王丽娜
3.鹅掌楸属两种植物花粉品质和花粉管生长的研究 [J], 周坚;樊汝汶
4.木本植物花粉生活力测定及生活力下降原因探讨 [J], 陈品良;金炜
5.多种蔷薇属植物花粉生活力的探究 [J], 李卉;韩禧;陈向前;商阳;姜马兰
因版权原因,仅展示原文概要,查看原文内容请购买。
杂种鹅掌楸及其亲本花部形态和花粉活力的遗传变异

杂种鹅掌楸及其亲本花部形态和花粉活力的遗传变异徐进;王章荣【期刊名称】《植物资源与环境学报》【年(卷),期】2001(010)002【摘要】对鹅掌楸属(Liriodendron L.)种间杂种[L. chinense (Hemsl.)Sarg. × L. tulipifera L.]及其亲本北美鹅掌楸(L.tulipifera)和鹅掌楸[L.chinense(Hemsl.)Sarg.]的花朵大小与颜色、雄蕊花药、雌蕊柱头和花粉活力的变异进行了观测,杂种鹅掌楸花朵较大,花瓣橙黄色,色彩艳丽,观赏价值较高.在固体培养条件下杂种鹅掌楸及其亲本北美鹅掌楸和鹅掌楸的花粉开始萌动大约需40min,而观测花粉发芽率以培养160min为宜.杂种鹅掌楸的花粉发芽率低于鹅掌楸和北美鹅掌楸,种间差异达显著水平.杂种鹅掌楸花粉活力在株间差异达显著水平,两亲本花粉活力的株间差异不显著.树冠上层花的花粉活力较高,与下层花的差异达显著水平.【总页数】4页(P31-34)【作者】徐进;王章荣【作者单位】南京林业大学森林资源与环境学院,江苏南京 210037;南京林业大学森林资源与环境学院,江苏南京 210037【正文语种】中文【中图分类】S792.21;Q321.2【相关文献】1.鹅掌楸属种间杂种苗期生长性状的遗传变异与优良遗传型选择 [J], 李周岐;王章荣2.鹅掌楸属种间杂种与亲本花粉壁的超微结构的比较观察 [J], 徐进;王章荣3.鹅掌楸属种间杂种F1与亲本花果数量性状的遗传变异分析 [J], 李周岐;王章荣4.鹅掌楸属种间F1杂种与亲本的叶下表皮微形态研究 [J], 叶金山;王章荣5.中国鹅掌楸、美国鹅掌楸及其杂种在形态和生长性状上的遗传变异 [J], 刘鸿谔因版权原因,仅展示原文概要,查看原文内容请购买。
广西猫儿山鹅掌楸天然种群动态研究

广西猫儿山鹅掌楸天然种群动态研究李博;冯源恒;杨章旗;袁晓峰;叶靖;李火根【摘要】为进一步探讨鹅掌楸濒危的机制,对广西桂林猫儿山鹅掌楸天然种群进行了调查,以种群生命表及生存分析理论为基础,以林木径级结构代表龄级结构,编制鹅掌楸种群静态生命表,分析种群结构的动态变化特点.结果表明:鹅掌楸种群龄级结构为金字塔型,幼龄个体数量较多,中龄个体数居中,而老龄个体则相对数量较少,表现为增长型种群;在第Ⅳ龄级时该种群出现死亡高峰,其种群存活曲线属Deevey-Ⅱ型;猫儿山鹅掌楸天然种群具有前期种群数量快速减少,中后期稳定,末期衰退的特点.该研究结果对鹅掌楸天然种群的保护提供参考依据.%In order to provide reference and further explore endangered mechanism of Liriodendron chinensis natural populations the population structure dynamic of L. chinensis was analyzed by taking a natural population in Maoer Mountain ofGuilin,Guangxi,as a sample. Based on the theory of population life table and survival analysis,the life table of L. chinensis population was constructed by replacing age classes with size-classes,and the population structure dynamic of L. chinensis was explored. The census data showed that the number of young seedling was larger than that of middle aged and old individuals, indicating that the population was in an incremental status. The survival curve in this population appeared to be a Deevey-Ⅱ type,with a peak of mortality in size class Ⅳ. It can be concluded that the population size of L. chinensis in Maoer mountain declined sharply in early stage, stablized in middle stage and fell rapidly in last stage.【期刊名称】《广西植物》【年(卷),期】2013(033)002【总页数】6页(P242-246,207)【关键词】鹅掌楸;龄级;静态生命表;种群结构;动态【作者】李博;冯源恒;杨章旗;袁晓峰;叶靖;李火根【作者单位】南京林业大学,林木遗传与生物技术/省部共建教育部重点实验室,南京,210037;南京林业大学,林木遗传与生物技术/省部共建教育部重点实验室,南京,210037;广西壮族自治区林业科学研究院,南宁,530001;广西壮族自治区林业科学研究院,南宁,530001;南京林业大学,林木遗传与生物技术/省部共建教育部重点实验室,南京,210037;南京林业大学,林木遗传与生物技术/省部共建教育部重点实验室,南京,210037;南京林业大学,林木遗传与生物技术/省部共建教育部重点实验室,南京,210037【正文语种】中文【中图分类】Q948种群生态学主要研究不同种群数量在时间和空间上的动态变化情况。
武汉植物园揭示中国鹅掌楸结实率低下的原因

武汉植物园揭示中国鹅掌楸结实率低下的原因
佚名
【期刊名称】《生物学教学》
【年(卷),期】2014(39)12
【摘要】据光明网2014年7月2日援引报道,中国科学院武汉植物园博士研究生李明在杨平仿研究员指导下,以鹅掌楸授粉后不同时间点的雌蕊为材料,分别鉴定到51个和468个蛋白质的表达量在授粉前后发生变化。
对这些差异表达的蛋白质进行的功能分类发现,共有66个蛋白质参与有性生殖过程。
雌雄蕊发育和花粉萌发等的形态学观察结果揭示,以蛋白质二硫键异构酶A6和胚胎发育缺陷蛋白质为代表的差异表达的蛋白质与鹅掌楸的低结实率现象高度相关,提示这些参与有性生殖过程的蛋白质的差异表达影响了鹅掌楸的受精过程。
【总页数】1页(P73-73)
【关键词】中国鹅掌楸;武汉植物园;结实率;原因;差异表达;二硫键异构酶;生殖过程;蛋白质
【正文语种】中文
【中图分类】Q786
【相关文献】
1.鹅掌楸的花部数量变异与结实率 [J], 黄双全;郭友好;吴艳;刘琴;张帆;陈家宽
2.中国鹅掌楸与北美鹅掌楸种间杂交的胚胎学研究 [J], 尹增芳;樊汝汶
3.中国鹅掌楸、美国鹅掌楸及其杂种在形态和生长性状上的遗传变异 [J], 刘鸿谔
4.中国鹅掌楸与北美鹅掌楸种间杂交的胚胎学研究 [J], 尹增芳;樊汝汶
5.北美鹅掌楸和中国鹅掌楸种间杂交胚胎学 [J], 樊汝汶;尤录祥
因版权原因,仅展示原文概要,查看原文内容请购买。
鹅掌楸分子标记开发及遗传连锁框架图谱构建的开题报告

鹅掌楸分子标记开发及遗传连锁框架图谱构建的开题报告在生物多样性保护和资源利用方面,分子标记和遗传连锁是非常有效的工具。
本开题报告旨在介绍鹅掌楸分子标记开发及遗传连锁框架图谱构建的研究计划。
研究背景与意义:鹅掌楸是我国南方常见的乔木,因其高质木材和药用功效,已成为重要的林业经济树种。
然而,由于其繁殖受限、生长缓慢、发芽率低等生物学特性,传统的育种方法已不能满足其种质资源分析和改良的需要,因此需要开展分子标记和遗传连锁研究,以促进鹅掌楸育种的进一步发展。
研究内容与方案:1.分子标记的开发根据已有的鹅掌楸的EST序列,我们将筛选SNP位点并设计引物,利用PCR技术对鹅掌楸样品进行扩增,并进行测序以获取SNP情况。
然后,我们将对序列信息进行生物信息学分析,以构建鹅掌楸SNP分子标记。
2.遗传连锁框架图谱的构建还原全亲系群体(纯合系与无杂合性群体),进行二倍体的基因型分析,采用 JoinMap4.1和R/qtl软件来进行QTL和遗传连锁分析,构建鹅掌楸的遗传连锁框架图谱。
最后,通过QTL定位和基因克隆,我们将探讨鹅掌楸木材性状在基因水平上的遗传规律,为鹅掌楸的育种提供科学依据。
研究意义与预期结果:该研究将为鹅掌楸育种提供新的思路和方法,同时向森林资源保护和利用提供了技术支持。
预期结果如下:1.获得鹅掌楸SNP分子标记,为鹅掌楸的种质资源核心采集、资源保护和利用提供有力支撑。
2.构建鹅掌楸遗传连锁框架图谱,探究木材性状遗传规律,为鹅掌楸育种提供科学依据和方法。
3.完善鹅掌楸种质资源的数据库,为鹅掌楸种质资源的利用和共享提供基础支撑。
研究难点与解决方案:1. 鹅掌楸生长缓慢、繁殖受限等生物特性会导致DNA样品质量不佳,因此需要使用精细的样品处理技术,并进行质量控制。
2. 鹅掌楸SNP分子标记开发的设计、扩增、测序以及序列信息处理等方面需要耗费较长的时间和精力。
为此,我们将建立高效的实验平台和操作流程以提高工作效率。
鹅掌楸属树种物候观测和杂种家系苗光合日变化

鹅掌楸属树种物候观测和杂种家系苗光合日变化季孔庶;杨秀艳;杨德超;王章荣【期刊名称】《南京林业大学学报:自然科学版》【年(卷),期】2002(26)6【摘要】成年鹅掌楸属树种萌动、发芽或展叶期的先后次序依次为反交杂种、北美鹅掌楸、正交杂种、鹅掌楸,但正交杂种展叶速度快于其它被观测植株。
正反交杂种花期持续时间均比双亲的长。
杂种和亲本的花期之间有较长交叉时间(14d)。
参试4个杂种家系和1个鹅掌楸家系苗期光合日变化特点有异:回交家系呈现单峰型变化曲线,峰值下降缓慢;正交家系及F1个体间杂交家系出现双峰曲线;鹅掌楸与反交家系均为单峰曲线,且峰值出现后净光合速率下降迅速。
其净光合速率由高至低的排序依次为回交家系、正交家系、反交家系、鹅掌楸、F1个体间杂交家系。
多数家系的净光合速率与胞间CO2浓度、蒸腾作用、气温、叶温、光合有效辐射等因子密切相关,但不同家系与这些因子相关关系不尽相同。
反交杂种表现出成年树具有萌动早、展叶早、开花早、花期持续时间长的特点;苗期净光合速率与胞间CO2浓度相关不显著。
【总页数】5页(P28-32)【关键词】鹅掌楸属树种;物候观测;树苗;杂种家系苗;光合日变化【作者】季孔庶;杨秀艳;杨德超;王章荣【作者单位】南京林业大学林木遗传与基因工程重点实验室;河北廊坊市农林科学院;湖北省林业厅【正文语种】中文【中图分类】S792.21;S718【相关文献】1.优良生态树种杂种鹅掌楸引种研究 [J], 卜基保2.绿化观赏树种——杂种鹅掌楸 [J], 李冬玲3.中国鹅掌楸、美国鹅掌楸及其杂种在形态和生长性状上的遗传变异 [J], 刘鸿谔4.优良生态树种杂种鹅掌楸引种研究 [J], 卜基保5.杂种鹅掌楸体胚苗根尖染色体核型分析 [J], 刘光欣;陈金慧;孙杰;愈天驰;施季森因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
A src : Fu rset rb ctxme y etr f u 一 Fu 一/ M)w sl d dit tecl f ioe do l i r x b ta t loec n oeaeoy t l s o3( o3 A p h eoF l l a a e o h el o r dn rnt if a o n s Li up e
Dy a i a e o e 2 d rn o ln D e eo m e to n m c Ch ng fFr e Ca u i g P l v l p n f e
Lr d n rntl i r × L. hn ne io edo if a i up e c ie s
尹 增 芳 宁代 锋 李 玉 春 席 梦 利
摘
施 季 森
南 京 2 03 ) 107
( . 京林 业 大学 森林 资 源 与环 境 学 院 南 京 2 03 ; 2 国 家 林 业 局 林 木 遗 传 与 基 因 工 程 重 点 实 验 室 1南 107 .
要 : 利 用 钙 离 子 荧 光 探 针 F o /M 低 温 装 载 技 术 , 激 光 扫 描 共 聚 焦 显 微 镜 下 观 察 杂 种 鹅 掌 楸 花 粉 发 育 不 l A u3 在
i N , t y a
2 K yL b r oyo F r tT eG nt s n n n i e n Sa o t d iirt n N mi 10 7 . e a oa r f o r ee c a G e E gn r g, t eF r r m n t i a n 2 0 3 ) t s e e i d e e i t s e yA sao g
Yi e ga g n Z n fn Nig Daf n n i g e L c u iYu h n XiMe gi・ nl S iJs n ・ h ie
203 ; 10 7
( . o eeo oe e u e a dE v o m n , a n o sy u 1 C lg l fF r t s r s n n i n e t N mi F r t sR o c r g er
L.c ies nh ru d rlw e eaue a ℃ t b ev h i rb t n o re C 2 d rn h oln d v lp n sn hn n e a te n e o tmp rtr t4 o o s re te dsi u i ff a u gt ep l e eo me tu ig t o e i e
维普资讯
第 4 卷 第 1 期 3 1
2007年 1 月 1
林
业
科
学
Vo . 3, 1 4 No. 11 No , v. 2 0 0 7
S ENTI CI A
SL I VAE
S NI I CAE
杂 种 鹅 掌 楸 花 粉 发 育 过 程 中细 胞 游离 C 2 的动 态 变 化 * a+
c 2 荧 光强 度 较 强 , 胝 质 壁 无 荧 光 ; 期 细 胞 质 中 的 c 2 光 减 弱 , 胼 胝 质 壁 处 C 2 光 增 强 。 小 孢 子 内 a 胼 后 a 荧 而 a 荧
c 分 布 不 均 匀 , 胞 质 中 的 C 荧 光 较 弱 , 胞 壁 处 c 荧 光 较 强 , 细 胞 花 粉 时 期 , 胞 呈 现 较 强 的 c 荧 a 细 a 细 a 二 细 a 光 。在 花 药 壁 组 织 内 c 分 布 也 呈 现 规 律 性 的 变化 : 孢 组 织 时期 , 药 壁 组 织 c 荧 光 强 度 在 不 同 壁 层 组 织 中 a 造 花 a 分 布 均 匀 ; 孢 子 母 细 胞 时 期 , 壁 中层 细 胞 C2 光 最 强 , 毡 层 细 胞 次 之 ; 核 小 孢 子 时 期 , 毡 层 细 胞 呈 解 体 小 药 a 荧 绒 单 绒
cnoa lsr cn igm cocp ( L M) h y a i i r uino reC 2 w sm aue i h lt eca gso ofcl ae ann irso y C S .T ed nm cds b t ff a a esrd wt ter ai h ne f s t i o e h e v
育 过 程 的重 要 转 变 环 节 密 切 相 关 。
关键 词 : 杂 种 鹅 掌 楸 ;花 粉 发 育 ;游 离 c 2 ; 温 装 载 a 低
中 图分 类 号 :7 8 4 ¥ 1 .3 文献标识码 : A 文 章 编 号 :0 1 4 8 20 1 0 7— 5 10 —7 8 ( 分 布 , a 的 以其 相 对 荧 光 强 度 的 强 弱 来 衡 量 细 胞 内游 离 C2 平 的 动态 变 化 。研 究 发 现 在 a 水 初 生 造 孢 细 胞 中 c2 光较 弱 , 生 造孢 细 胞 和 小 孢 子 母 细 胞 时 期 c 2 光 增 强 。 早 期 四 分 体 小 孢 子 细 胞 质 中 a 荧 次 a 荧
状态 ,a c2 光 最 强 , 保 持 到 二 核 花 粉 时 期 直 至 绒 毡 层 完 全 消 失 , 此 时 花 药 纤 维 层 发 育 形 成 , 现 出 较 强 的 荧 并 但 表
c 荧 光 。 花 药 组 织 中 c 分 布 动 态 显 示 出 c 2 由外 而 内 流 动 的 迹 象 , 且 细 胞 内游 离 c 分 布 特 征 与 花 粉 发 a a a 而 a