蛋白质生物合成
第十五章蛋白质的生物合成

第一位
(5ˊ)
U
U
C
A
G
遗传密码字典 第二位
C
A
G
第三位
(3ˊ)
U C A G
U C A G
U C A G
U C A G
6 4 组 密 码 子 中 , AUG 既 是 甲 硫氨酸的密码,又是起始密码; 有三组密码不编码任何氨基酸, 而是多肽链合成的终止密码子: UAG、UAA、UGA。
• 在原核生物和真核生物中,均存在另一 种携带蛋氨酸的tRNA,识别非起动部位 的蛋氨酸密码,AUG。
第十五章蛋白质的生物合成
核糖体的研究历史
1.早在本世纪30年代后期就发现细胞质和细胞核中都有 核酸存在,不过用1924年福尔根发明的染色法只能使细胞核 中的核酸染色。但两种核酸在260nm的吸收非常相似。
第十五章 蛋白质合成及转运
蛋白质的生物合成
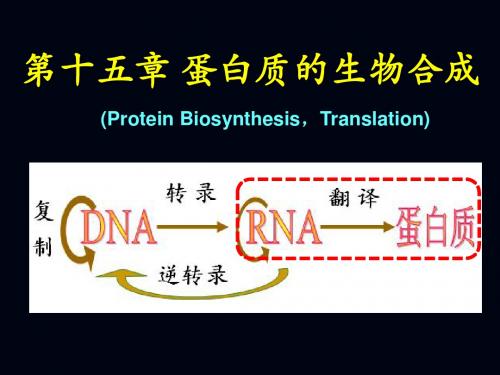
中心法则指出,遗传信息的表达最终是合成出 具有特定氨基酸顺序的蛋白质,这种以mRNA上所 携带的遗传信息,到多肽链上所携带的遗传信息的传 递,就好象以一种语言翻译成另一种语言时的情形 相似,所以称以mRNA为模板的蛋白质合成过程为 翻译(translation)。
2.Crick, F.比较了核酸和氨基酸的大小和形状后,认为不可 能在空间上互补,因此预测:(1) 存在一类分子转换器,使信 息从核酸序列转换成氨基酸序列;(2) 这种分子很可能是核酸; (3) 它不论以何种方式进入蛋白质翻译系统的模板,都必须与 模板形成氢键(即配对);(4) 有20种分子转换器,每种氨基 酸一个;(5) 每种氨基酸必定还有一个对应的酶,催化与特定 的分子转换器结合。
2.1941年,细胞学家J.Brachet和T.Caspersor注意到细 胞质中的核酸与蛋白质的合成有密切的关系。
蛋白质的生物合成

蛋白质的生物合成
蛋白质是生命体中最为重要的分子之一,参与了生物体的很多生命过程。
蛋白质的生物合成是指在细胞内通过核糖体合成多肽链,并将多肽链进一步折叠成特定的三维结构的过程。
这个过程需要包括DNA转录、RNA翻译、蛋白质折叠等多个步骤。
在生物体内,DNA中的基因序列被转录成RNA分子。
RNA分子进一步通过核糖体将多个氨基酸连接成一条多肽链。
在这个过程中,RNA 分子会依据基因序列上的密码子来选择相应的氨基酸,并将它们串联在一起。
这个过程中的每一个密码子都对应着一种氨基酸,这种关系被称为遗传密码。
一条多肽链的生命周期并不仅仅是由其基因序列决定的。
在折叠过程中,这条链会被各种分子和酶修饰和加工,最终形成最终的三维结构。
这个过程中的每一个步骤都非常关键,因为一个错误的步骤都可能导致最终的结构失去功能。
蛋白质的生物合成是生命体中最为复杂的分子合成过程之一。
在这个过程中,细胞需要精确地将基因序列转录成RNA分子,并将氨基酸按照正确的顺序连接成多肽链。
同时,细胞还需要通过各种酶和分子来协助蛋白质的折叠和修饰,最终形成具有特定功能的蛋白质。
这个过程非常关键,因为蛋白质的结构和功能决定了生命体的很多生命过程。
- 1 -。
蛋白质生物合成的方式

蛋白质生物合成的方式
蛋白质生物合成是生物体内制造蛋白质的过程,它是生物体内的重要生化反应之一。
下面介绍蛋白质生物合成的方式:
1. 氨基酸活化:在蛋白质生物合成中,首先需要将氨基酸激活。
这个过程由特定的酶催化,称为氨基酸激酶。
被激活的氨基酸随后会与另一种分子——核糖磷酸结合,形成称为氨酰-tRNA的化合物。
2. 起始复合物形成:第二个步骤是形成起始复合物。
这个过程涉及氨酰-tRNA 与mRNA的结合,其中mRNA是包含蛋白质序列信息的分子。
这个过程需要核糖体起始因子(eIF)的帮助。
3. 肽链合成:一旦起始复合物形成,蛋白质合成就可以开始了。
每个氨基酸通过肽键连接在一起,形成一个连续的肽链。
这个过程由转录延长因子(eEF)和核糖体来催化。
4. 蛋白质折叠:当肽链合成完成后,蛋白质就会开始折叠成其最终的三维形状。
这个过程需要帮助,包括来自分子伴侣蛋白和折叠酶的帮助。
5. 蛋白质修饰:在某些情况下,还需要对蛋白质进行进一步修饰,例如添加糖基或脂质,或者进行磷酸化或乙酰化等化学修饰。
总的来说,蛋白质生物合成是一个复杂的过程,需要多个酶和分子的协同作用。
通过这个过程,生物体能够制造出其生命活动中所需的蛋白质。
【生物化学】蛋白质的生物合成

嘌 呤 霉 素
酯键
(3)转位(translocation)
•转位酶 (translocase): •原核:延长因子G(EF-G),真核:EF-2 • GTP
可结合并水解1分子GTP,促进核 蛋白体向mRNA的3’侧移动
进 位
成肽 转 位
合成
3、肽链终止阶段:
核蛋白体沿mRNA链滑动,不断使多 肽链延长,直到终止信号进入受位。
四、蛋白质生物合成的干扰和抑制
1、抗生素(antibiotics)
名称
作用机制
四环素类 氯霉素类 链霉素类 嘌呤霉素
抑制氨酰-tRNA与原核生物核糖体结合,抑制细菌 蛋白质合成
结合原核生物核糖体大亚基,阻断翻译延长过。高 浓度时,对真核生物线粒体内的蛋白质合成也有阻 断作用 结合原核生物核糖体小亚基,改变其构象,引起读 码错误
,IF)
有抗病毒作
用的蛋白质
1、诱导一种蛋白激酶,使eIF2磷酸 化,从而抑制病毒蛋白质的生物合 成。
2、诱导生成一种寡核苷酸(2’5’A),活化核酸内切酶RNaseL, 可降解病毒RNA。
谢 谢!
(1)识别:释放因子(RF)识别终 止密码,进入核蛋白体的受位。
(2)水解:RF使转肽酶变为水解酶, 多肽链与tRNA之间的酯键被水解, 多肽链释放。
(3)解离:通过水解GTP,使核蛋 白体与mRNA分离,tRNA、RF脱落, 核蛋白体解离为大、小亚基。
进位
肽链的形成 移位
蛋白质 合成过程
肽链合成终止
结构与Tyr-tRNAtyr相似,阻止肽链正常合成
放线菌酮 抑制核糖体转肽酶。且只对真核生物有特异性作用
2、干扰蛋白质生物合成的生物活性物质
名称
蛋白质生物合成(医药经济)

肽链的起始、延伸与终止
总结词
肽链的起始、延伸与终止是蛋白质生物合成中的核心步骤,涉及核糖体、mRNA和 tRNA的相互作用。
详细描述
在起始阶段,核糖体结合到mRNA上,并由起始氨基酰-tRNA进入核糖体的A位。延伸 阶段中,氨酰-tRNA按照mRNA的密码子顺序依次进入核糖体的A位,肽链开始延伸。 终止阶段时,终止密码子出现,肽酰-tRNA进入核糖体的P位,合成终止,新生的肽链
02
蛋白质生物合成涉及多个步骤, 包括氨基酸活化、肽链合成、翻 译后加工等。
蛋白质生物合成的意义
蛋白质是生命活动中不可或缺的分子 ,参与细胞内各种生理和代谢过程, 如细胞信号转导、免疫反应、细胞增 殖和分化等。
蛋白质生物合成是细胞内基因表达的 重要途径,是细胞生长、发育和维持 正常功能的基础。
蛋白质生物合成的历史与发展
疾病诊断与治疗
疾病标志物发现
蛋白质生物合成异常与多种疾病的发 生发展密切相关,通过对其研究有助 于发现新的疾病标志物,提高疾病的 早期诊断率。
个体化治疗
免疫治疗
利用蛋白质生物合成技术可以制备具 有免疫原性的多肽或蛋白质,用于肿 瘤免疫治疗和自身免疫性疾病的治疗。
根据患者的基因型和蛋白质表达谱, 可以为患者量身定制个体化的治疗方 案,提高治疗效果和患者的生存率。
抗体药物技术
抗体药物技术是指利用免疫学原理和技术手段,制备出具有特定结构和功能的抗体, 用于治疗和预防疾病。
该技术广泛应用于医药领域,如单克隆抗体、双特异性抗体、抗体融合蛋白等的制 备和应用,以及免疫诊断和免疫治疗等。
抗体药物技术的优点在于能够精确地靶向病变细胞和分子,提高药物的疗效和安全 性,并且可以降低药物的毒性和副作用。
生物化学第十二章-蛋白质的生物合成

第十二章蛋白质的生物合成一、蛋白质生物合成体系:生物体内的各种蛋白质都是生物体利用约20种氨基酸为原料自行合成的。
蛋白质的生物合成过程,就是将DNA传递给mRNA的遗传信息,再具体的解译为蛋白质中氨基酸排列顺序的过程,这一过程被称为翻译(translation)。
参与蛋白质生物合成的各种因素构成了蛋白质合成体系,该体系包括:1.mRNA:作为指导蛋白质生物合成的模板。
mRNA中每三个相邻的核苷酸组成三联体,代表一个氨基酸的信息,此三联体就称为密码。
共有64种不同的密码。
遗传密码具有以下特点:①连续性;②简并性;③通用性;④方向性;⑤摆动性;⑥起始密码:AUG;终止密码:UAA、UAG、UGA。
2.tRNA:在氨基酸tRNA合成酶催化下,特定的tRNA可与相应的氨基酸结合,生成氨基酰tRNA,从而携带氨基酸参与蛋白质的生物合成。
tRNA反密码环中部的三个核苷酸构成三联体,可以识别mRNA上相应的密码,此三联体就称为反密码。
反密码对密码的识别,通常也是根据碱基互补原则,即A—U,G—C配对。
但反密码的第一个核苷酸与第三核苷酸之间的配对,并不严格遵循碱基互补原则,这种配对称为不稳定配对。
能够识别mRNA中5′端起动密码AUG的tRNA称为起动tRNA。
在原核生物中,起动tRNA是tRNAfmet;而在真核生物中,起动tRNA是tRNAmet。
3.rRNA和核蛋白体:原核生物中的核蛋白体大小为70S,可分为30S小亚基和50S大亚基。
真核生物中的核蛋白体大小为80S,也分为40S小亚基和60S大亚基。
核蛋白体的大、小亚基分别有不同的功能:⑴小亚基:可与mRNA、GTP和起动tRNA结合。
⑵大亚基:①具有两个不同的tRNA结合点。
A位——受位或氨酰基位,可与新进入的氨基酰tRNA 结合;P位——给位或肽酰基位,可与延伸中的肽酰基tRNA结合。
②具有转肽酶活性。
在蛋白质生物合成过程中,常常由若干核蛋白体结合在同一mRNA分子上,同时进行翻译。
蛋白质生物合成体系

核糖体的结构
核糖体是一种高度复杂的超分子结构,由多个蛋白质和RNA分子组装而成。
大、小亚基的形状类似于扁平的椭圆形或球形,大亚基的直径约为70-80 埃,小亚基的直径约为40-50埃。
核糖体中的蛋白质和RNA分子通过相互作用形成了一个稳定的结构,使得 核糖体能够作为一个整体来执行功能。
02
每种氨基酸都有特定的氨酰tRNA合成酶,该酶具有高度的 专一性,只对一种氨基酸起作 用。
03
活化后的氨基酸通过酯键与 tRNA结合,形成氨酰-tRNA, 为接下来的蛋白质合成做准备。
氨基酸在细胞内的转运
氨基酸在细胞内的转运主要依靠细胞内不同的 转运体系来完成,这些转运体系能够识别和结 合相应的氨基酸,并将其转运到需要的地方。
通过调节翻译延长因子EF-Tu、EF-G等,可以影响蛋 白质合成的速率。
调节翻译终止
通过调节翻译终止因子eRFs的活性,可以控制蛋白质 合成的终止。
蛋白质生物合成与疾病的关系
疾病发生
01
当蛋白质生物合成体系出现异常时,可能导致某些疾病的发生,
如癌症、感染性疾病等。
疾病发展
02
蛋白质生物合成体系的变化可能影响疾病的发展进程,如肿瘤
蛋白质的折叠
1
蛋白质折叠是指蛋白质合成后,通过一系列复杂 的化学和物理过程,将其由线性肽链折叠成具有 特定三维结构的构象。
2
蛋白质折叠是一个自发的、动态的过程,需要依 靠分子伴侣、折叠酶等辅助因子来完成。
3
正确的蛋白质折叠对于维持细胞正常功能和生物 体的健康至关重要,而错误的折叠会导致多种疾 病的发生。
核糖体的功能
蛋白质的生物合成

16
(二)氨基酰-tRNA合成酶 (aminoacyl-tRNA synthetase)
氨基酰-tRNA合成酶
氨基酸 + tRNA
氨基酰- tRNA
ATP
AMP+PPi
17
1. 氨基酰-tRNA合成酶对底物氨基酸和tRNA 都有高度特异性:每种氨基酸都只有1种氨基酰tRNA合成酶,生成特定的氨基酰-tRNA(aa-tRNA)
参与蛋白质生物合成的主要物质 蛋白质生物合成过程 翻译后的加工与靶向输送 蛋白质生物合成的抑制剂
3
第一节 参与蛋白质生物合成的 主要物质
蛋白质合成体系:20种氨基酸,mRNA、 tRNA、核蛋白体、酶和蛋白因子、无机离 子、ATP 、GTP 等。
4
一、mRNA:合成蛋白质பைடு நூலகம்模板
(一) mRNA是遗传信息的携带者 遗传学将编码一个多肽的遗传单位称为
14
密码子与反密码子除通过碱基互补结合外, 还具有摆动性,即密码子的第3位碱基与反 密码子的第1位碱基配对不严格,称为摆动 配对。
15
密码子、反密码子配对的摆动现象
tRNA反密码子 第1位碱基
I
U G AC
mRNA密码子 第3位碱基
U, C, A A, G U, C U G
密码子的第3位碱基发生突变时,并不影响tRNA带入 正确的氨基酸。
2.氨基酰-tRNA的表示方法:
Ser-tRNASer
Met-tRNAMet
真核生物起始因子辨认的Met-tRNAiMet 延长因子辨认Met-tRNAeMet
右上角字母可略
18
三、核糖体:合成蛋白质的场所
蛋白质生物合成

目录
目录
Tu TGsTP
Tu GDP
Ts GTP
5'
AUG
3'
目录
(二)成肽
是由转肽酶(transpeptidase)催化的肽键 形成过程。
目录
(三)转位
延长因子EF-G有转位酶( translocase ) 活性,可结合并水解1分子GTP,促进核蛋 白体向mRNA的3'侧移动 。
目录
fMet fMet
目录
原核、真核生物各种起始因子的生物功能
原核 生物
真核 生物
起始因子 IF-1 EIF-2 EIF-3 eIF-2
eIF-2B,eIF-3
IF-4A
eIF--4B eIF-4E eIF-4G eIF-5 eIF-6
生物功能 占据A位防止结合其他tRNA 促进起始tRNA与小亚基结合 促进大小亚基分离,提高P位对结合起始tRNA敏感性 促进起始tRNA与小亚基结合 最先结合小亚基促进大小亚基分离 eIF-4F复合物成分,有解螺旋酶活性,促进mRNA结
目录
遗传密码的特点
1. 连续性(commaless)
编码蛋白质氨基酸序列的各个三联体密 码连续阅读,密码间既无间断也无交叉。
目录
• 基因损伤引起mRNA阅读框架内的碱基发生 插入或缺失,可能导致框移突变(frameshift mutation)。
目录
2. 简并性(degeneracy)
遗传密码中,除色氨酸和甲硫氨酸仅 有一个密码子外,其余氨基酸有2、3、4 个或多至6个三联体为其编码。
目录
1. 核蛋白体大小亚基分离
IF-1 IF-3
目录
2. mRNA在小亚基定位结合
第15章 蛋白质的生物合成
进 位
成肽 转 位
成肽
成肽是由转肽酶催化形成肽键的过程。
Tu GTP
Tu GDP
5'
AUG
3'
fMet fMet
Tu GTP
5'
AUG
3'
(三) 原核生物翻译终止
当mRNA上的终止密码子出现后,多肽链合成 停止,肽链从肽酰-tRNA中脱落,mRNA、核糖体 等解体,此时翻译过程终止。 终止密码子不被任何氨基酰-tRNA识别,只有 释放因子(RF)能够识别它并进入核糖体的A位。 1. 释放因子可引起转肽酶活性的改变,从而使 肽链从tRNA上脱落。 2. 释放因子可引起核糖体结构的改变,从而使 复合体趋于解体。
解码
反密码子(anti-codon) 反密码子是位于tRNA、可与mRNA的三联体密 码子配对的三个相邻核苷酸。在蛋白质的合成中, 起解读密码、辅助将特异的氨基酸引入合成位点。 tRNA
5'
AUG
反密码子
3'
mRNA
密码子
遗传密码的特点
翻译时遗传密码的阅读方向是 5’→3’,即读 码从mRNA的起始密码子AUG开始,按5’→3’ 的方向逐一阅读,直至终止密码子。
COORF
5'
UAG
3'
第四节
蛋白质翻译后加工和靶向输送
Posttranslational Modification and Targeting Transfer of Protein
蛋白质翻译后加工
十三蛋白质的生物合成

转位(移位)
转位酶
5′→3′
5′
AUG
3′
AUG
EFG,GTP
A
OH 21
21
OH
AUG
转位
AUG
进位
3 12
AUG
N→C延
成肽
编辑2p1pt
3
3
21
成肽
四、肽链合成终止
• 需终止因子RF、RR和IF3参与。终止信号 出现,释放因子(release factor,RF, RR) 与其结合。RF有三种RF1,RF2,RF3
编辑ppt
密码子的特点
(1)连续性:两个密码子之间无任何核苷酸加以 隔开和重叠,如插入/删除碱基,可发生移 码突变或框移
5′…. UACGGACAUCUG….3′ 酪甘 组 蛋
插入 缺失
5′….UACCGGACAUCUG….3′ 酪 精 苏 半胱
5′…. UACGACAUCUG….3′ 酪 天 异亮 编辑ppt
编辑ppt
一、氨基酸的活化与转运
氨基酸活化→活化氨基酸的搬运→活化氨 基酸与核蛋白体结合 1.参与活化转运的酶
氨基酰-tRNA合成酶:特异性强,催化特定 的氨基酸与特异的tRNA结合,每种氨基酸 有特异的合成酶催化,此种特异性保证了 遗传信息准确翻译
编辑ppt
氨基酰tRNA的生成--- 氨基酸的活化
编辑ppt
密码子的特点
(四)摆动性:一种氨基酸可有多个密码子
反密码子与mRNA的第三个核苷酸配对时,不 严格遵从碱基配对原则,可出现U-G,I-C,I-A,此种 配对为不稳定配对,又称摇摆性。一般前两个碱 基决定其专一性,第三位碱基可有变异
mRNA5′ ACG
蛋白质的生物合成

六. 核糖体
核糖体像一个能沿mRNA模板移动的工厂, 执行着蛋白质合成的功能。它是由几十种蛋 白质和几种核糖体RNA(ribosomal RNA, rRNA)组成的亚细胞颗粒。一个细菌细胞内 约有20000个核糖体,而真核细胞内可达106 个,在未成熟的蟾蜍卵细胞内则高达1012。 核糖体和它的辅助因子为蛋白质合成提供了 必要条件。
蛋白质的生物合成
一. 基因与基因表达的一般概念 二. 遗传密码——三联子 三.密码子和反密码子的相互作用 四.tRNA 五.AA- tRNA合成酶 六. 核糖体 七. 信使核糖核酸 八、蛋白质的生物合成 九、氨基酸及功能蛋白质合成后的修饰 十、蛋白质的运输和降解
AA-tRNA合成酶及其它蛋白质因子的细胞抽提物中加入 mRNA或人工合成的均聚物作为模板以及ATP、GTP、氨基 酸等成分时又能合成新的肽链,新生肽链的氨基酸顺序由外 加的模板来决定。
1961年,Nirenberg等以poly(U)作模板时发现合成了多 聚苯丙氨酸,从而推出UUU代表苯丙氨酸(Phe)。以poly (C)及poly(A)做模板分别得到多聚脯氨酸和多聚赖氨酸。
三.密码子和反密码子的相互作用
蛋白质生物合成过程中,tRNA的反密码子通 过碱基的反向配对与mRNA的密码子相互作 用。1966年,Crick根据立体化学原理提出摆 动假说(wobble hypothesis),解释了反密 码子中某些稀有成分如I以及许多有2个以上 同源密码子的配对问题。
1、tRNA的三叶草型二级结构
受体臂(acceptor arm)主要由链两端序列碱基配对 形成的杆状结构和3’端末配对的3-4个碱基所组成,其3’ 端的最后3个碱基序列永远是CCA,最后一个碱基的3’或 2’自由羟基(—OH)可以被氨酰化。TφC臂是根据3个核 苷酸命名的,其中φ表示拟尿嘧啶,是tRNA分子所拥有 的不常见核苷酸。反密码子臂是根据位于套索中央的三 联反密码子命名的。D臂是根据它含有二氢尿嘧啶 (dihydrouracil)命名的。 最常见的tRNA分子有76个碱基,相对分子质量约为 2.5×104。不同的tRNA分子可有74-95个核苷酸不等, tRNA分子长度的不同主要是由其中的两条手臂引起的。 tRNA的稀有碱基含量非常丰富,约有70余种。每个 tRNA分子至少含有2个稀有碱基,最多有19个,多数分 布在非配对区,特别是在反密码子3'端邻近部位出现的频 率最高,且大多为嘌呤核苷酸。这对于维持反密码子环 的稳定性及密码子、反密码子之间的配对是很重要的。
蛋白质的生物合成名词解释

蛋白质的生物合成名词解释
蛋白质生物合成是指在生物体内利用遗传信息,通过转录和翻译作用,将氨基酸按照一定的顺序和方式组合成蛋白质分子的过程。
具体来说,蛋白质生物合成主要分为三个阶段:转录、RNA加工和翻译。
转录:转录是指将DNA上的基因序列转换为RNA信使分子的过程。
在细胞核中,RNA聚合酶会按照DNA模板进行配对,并合成一个与DNA互补的RNA分子(称为前体mRNA),它包含了DNA中编码蛋白质所需的信息。
RNA加工:在细胞核中,前体mRNA需要经过信使RNA的加工,包括剪接、修饰和成熟,才能成为可用的成熟mRNA。
翻译:翻译是指将mRNA上的编码信息翻译成蛋白质的过程。
这一过程在细胞质中进行,靠着核糖体和tRNA的配合以及氨基酸的加入,从而合成出蛋白质分子。
例如胰岛素,它是由基因表达产生的,并在胰腺中被编码和分泌。
此外,肌肉组织中的肌肉蛋白,以及酶、激素等众多蛋白质都是通过生物合成得到的。
生物学中的蛋白质生物合成

生物学中的蛋白质生物合成蛋白质生物合成是生物学中一个非常重要的过程,它是细胞中的一种复杂的分子机器。
蛋白质是生命的基础积木,是构成细胞器、调节生理活动和实现遗传信息传递的重要分子。
为了揭示蛋白质生物合成的机理,许多科学家做了大量的研究。
本文将从蛋白质的结构、生物合成的过程、影响生物合成的因素等方面,对这个课题进行阐述。
一、蛋白质的结构蛋白质是由氨基酸组成的,大多数蛋白质都含有20种不同的氨基酸。
不同的氨基酸按照不同的序列排列,形成了具有不同结构和功能的蛋白质。
蛋白质分为四个结构层次,包括原始结构、二级结构、三级结构和四级结构。
原始结构指的是由氨基酸形成的多肽链,二级结构包括α-螺旋和β-折叠等,三级结构是指蛋白质的立体构型,四级结构指的是多个立体构型相互作用形成的高级结构。
二、蛋白质生物合成的过程蛋白质的生物合成是一个非常复杂的过程,大致由转录、转移、翻译等步骤组成。
1. 转录转录是指DNA的信息被转录成RNA的过程。
在转录过程中,DNA的一部分序列被复制到RNA分子上。
这个过程是由RNA聚合酶催化完成的。
RNA链具有相同的碱基,但是没有T碱基,而是U碱基。
2. 转移转移是指RNA分子被转移到核糖体中的过程。
核糖体是由RNA和蛋白质组成的复合物,它的主要功能是将RNA分子合成蛋白质。
RNA分子通过与核糖体上的蛋白质结合,形成转移复合体。
3. 翻译翻译是指RNA分子被翻译成蛋白质的过程。
在翻译过程中,核糖体读取RNA分子的信息,并将之翻译成具有特定功能的氨基酸序列。
三、影响蛋白质生物合成的因素蛋白质生物合成过程受到多种因素的影响,包括温度、pH值、适宜性等。
1. 温度温度对蛋白质的合成具有重要的影响。
低温下,酶的催化活性降低,蛋白质折叠发生异常,导致蛋白质的生物合成受到抑制。
2. pH值pH值对蛋白质的合成同样具有重要的影响。
在合适的pH范围内,蛋白质能够正确地折叠成稳定的结构,从而发挥正常的生理功能。
蛋白质的生物合成概述

蛋白质的生物合成概述蛋白质是构成生物体的基本组成部分之一,具有多种重要的功能。
蛋白质的生物合成是指从基因到蛋白质的整个过程,包括转录和翻译两个主要步骤。
在生物体内,蛋白质的合成由细胞核内的遗传物质DNA指导,通过转录将DNA的信息转录成为mRNA,然后通过翻译将mRNA的信息转化为氨基酸序列,最终形成蛋白质。
1. 转录(Transcription):转录是指将DNA中一个基因的信息通过RNA聚合酶酶催化,生成该基因的mRNA分子。
转录的过程包括启动、延伸和终止三个阶段。
-启动阶段:在DNA的启动序列上结合转录因子,形成转录起始复合物。
随后,RNA聚合酶酶催化聚合核苷酸,从而形成mRNA链的起始部分。
-延伸阶段:RNA聚合酶继续在DNA模板上向下移动,并在不断向RNA链中添加新的核苷酸,直到遇到终止信号。
-终止阶段:当RNA聚合酶读取到终止信号时,mRNA链与DNA模板分离,转录过程结束。
2. RNA加工(RNA Processing):转录过程产生的初级转录产物(pre-mRNA)还需要经过一系列修饰和处理步骤,形成成熟的mRNA,以便于翻译过程中正确地合成蛋白质。
- 在RNA加工过程中,先是通过剪接(splicing)去除在初级转录产物中的内含子(intron)区域,保留外显子(exon)区域。
剪接是由剪接体(spliceosome)完成的,它由snRNP(small nuclear ribonucleoprotein)复合物和其他蛋白质组成。
- 随后,在转录加工过程中,mRNA的5'端会经历甲基化修饰,而3'末端则会加上聚腺苷酸尾巴(poly(A) tail)。
这些修饰可以保护mRNA 分子免受降解,同时还有助于mRNA的定位和翻译。
3. 翻译(Translation):翻译是指将mRNA的信息翻译成蛋白质的氨基酸序列,与转录过程不同,翻译发生在细胞的细胞质中,主要通过核糖体来完成。
(生物化学)蛋白质合成

活化后的氨基酸具有更高的反应活性,能够更容易地与tRNA结合, 进而参与蛋白质的合成。
tRNA在氨基酸转运中作用
1 2
tRNA结构特点
tRNA具有三叶草形二级结构和倒L形三级结构, 其3'端CCA序列是氨基酸的结合部位。
tRNA与氨基酸结合
在氨酰-tRNA合成酶的催化下,活化氨基酸与 tRNA的3'端CCA序列结合,形成氨酰-tRNA。
翻译异常
翻译过程中tRNA的错误识别或核糖体的异常导致氨基酸的错误 加入,产生异常蛋白质。
相关疾病举例及临床表现
囊性纤维化
由于基因突变导致CFTR蛋白质合成异常, 使得氯离子通道功能障碍,临床表现为肺部 感染、胰腺功能不全等。
苯丙酮尿症
由于基因突变导致苯丙氨酸羟化酶合成异常,使得 苯丙氨酸代谢障碍,临床表现为智力低下、癫痫等 。
microRNA调控
microRNA是一类小非编码RNA,通过与靶mRNA结合,抑制其翻 译或促进其降解,从而调控基因表达。翻译平调控策略翻译起始调控
通过调控翻译起始复合物的形成和活性,控制蛋 白质合成的起始速率。
翻译延伸调控
通过影响翻译延伸过程中氨酰-tRNA的合成和转 运,调控蛋白质合成的速度和效率。
细胞治疗
通过移植正常的细胞来替代病变细胞,从而 恢复正常的蛋白质合成功能。
THANKS
感谢观看
(生物化学)蛋白质合 成
目录
• 蛋白质合成概述 • 氨基酸活化与转运 • 核糖体循环与多肽链延伸 • 翻译后修饰与蛋白质折叠
目录
• 蛋白质合成调控机制 • 蛋白质合成异常与疾病关系
01
蛋白质合成概述
蛋白质合成定义与意义
蛋白质的生物合成

1. 核糖体:肽链合成的场所
在原核细胞中,它可以游离形式存在,2000 个 /细胞 真核细胞中的核糖体位于胞浆内,可分为两类,一类与细胞内质网相 结合,形成粗糙内质网,主要参与白蛋白、胰岛素等分泌性蛋白质的 合成。另一类游离于胞浆,主要参与细胞固有蛋白质的合成。 106~107 个 / 细胞 。线粒体、叶绿体也有自己的核糖体。
We found that the codon UGA specifies insertion of selenocysteine and cysteine in the ciliate Euplotes crassus, that ……….
纤毛虫
Thus, the genetic code supports the use of one codon to code for multiple amino acids.
(7)偏倚性:蛋白质生物合成时对简并密码子的使用频率不同。
Science 9 January 2009: Vol. 323. no. 5911, pp. 259 - 261 DOI: 10.1126/science.1164748
Sicence:颠覆经典遗传学理论
Genetic Code Supports Targeted Insertion of Two Amino Acids by One Codon
2. tRNA —— 翻译的接合体(adaptor)
同功受体tRNA (isoaccepting tRNAs):携带相同氨基酸而 反密码子不同的一组tRNA 。一种氨基酸可有2~6种特异 tRNA。
tRNA中与蛋白质合成有关的四个位点:
核糖识别部位
氨基酸结合部位
氨酰-tRNA合 成酶识别部位
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
There are sporadic alterations of the universal code
Changes in the genetic code in mitochondria can be traced in phylogeny. The minimum number of independent changes is generated by supposing that the AUA=Met and the AAA=Asn changes each occurred independently twice, and that the earlyAUA=Met change was reversed in echinoderms.
(一)三联子密码定义
遗传密码: DNA(或mRNA)中的核苷酸序列与蛋白质 中氨基酸序列之间的对应关系称为遗传密码。 密码子(codon):mRNA上每3个相邻的核苷酸编码蛋白 质多肽链中的一个氨基酸,这三个核苷酸就称为一个密 码子或三联体密码(triplet coden) 。
mRNA
5’ GCU AGU ACA AAA CCU 3’
CAT
CAT
CAT
CAT
ATC
CAT CAC ATC CAT CAC AXT
ATC ATC CAT
CAT CAT
CAX
TXC
ATX
CAT
CAT
CAT
以均聚物为模板指导多肽的合成
Poly U 为模板,产生的多肽链为Poly Phe
Poly C 为模板,产生的多肽链为Poly Pro Poly A 为模板,产生的多肽链为Poly Lys
甲硫氨酸本身, 而是甲酰甲硫氨酸,起始AA-tRNA为
fMet-tRNAfMet
(三) tRNA的种类
2、同工tRNA
代表同一种氨基酸的tRNA称为同工tRNA。 同工tRNA既要有不同的反密码子以识别该氨基酸 的各种同义密码,又要有某种结构上的共同性, 能被相同的氨基酰-tRNA合成酶识别。
(三) tRNA的种类
tRNA contains modified bases that influence its pairing properties
All of the four bases in tRNA can be modified.
摆动配对
U
mRNA
5' tRNA
AUU UAI
5' 3' 3'
AUC UAI
反密码子环
图 14-15 tRNA 由三叶草型折叠成 L 型三维结构
(二) tRNA的功能
1、解读mRNA的遗传信息 2、运输的工具,运载氨基酸
tRNA有两个关键部位: ● 3’端CCA:接受氨基酸,形成氨酰-tRNA。 ●与mRNA结合部位—反密码子部位
3’
CCAOH
5’
I
5’
CCG GGC
数学的计算:
核甘酸序列 mRNA 5′
mRNA 5′ 氨基酸序列
AUCGACCUGAGC
4 20 (×)
3′
AUCGACCUGAGC
42=16 20 (×)
3′
mRNA 5′
AUCGACCUGAGC
43=64 20 (√)
3′
缺失或插入核苷酸引起三联体密码的改变
CAT -1
-1,+1 +3
CAT
5' 3'
AUA UAI
5'
3'
Ile
Ile
Ile
密码子、反密码子配对的摆动现象
tRNA反密码子 第1位碱基 mRNA密码子 第3位碱基
I
U
G
A
C
U, C, A
A, G U, C
U
G
Modification to inosine allows pairing with U, C, and A.
二、tRNA的结构、功能与种类
一些移码抑制基因可以识别超过 4个碱基的密码。例如,一种细菌 tRNALys 抑制因子可以识别AAAA或者AAAU,而不是通常状况下的AAA。另一种抑 制基因可以阅读任何一个以ACC为前三位置的4碱基密码子。它与第四位碱 基是什么无关,在这里,这第四位碱基不需要符合Wobble规则。这使得在下 一个tRNA能够找到一个密码子之前,一个碱基被跳过去。 在噬菌体和病毒中,移码经常发生。这也许可以影响蛋白质合成的继续或 终止,且是由于mRNA的内在的性质。
(二)三联体密码的破译
1954年Gamov确认核酸分子中三个碱基决定一 个氨基酸 1961年Crick 等用遗传学方法也证实三联体密码 子学说是正确的 Nirenberg以均聚物共聚物为模板指导多肽的合 成,寻找到了破译遗传密码的途径 Khorana以共聚物指导多肽的合成,加快了破译 遗传密码的步伐
减少了变异对生物的影响
编码某一氨基酸的密码子越多,该氨基酸在蛋白质 中出现的频率就越高。Arg例外
The number of codons for each amino acid does not correlate closely with its frequency of use in proteins.
(三)遗传密码的性质 3、 连续性
编码蛋白质氨基酸序列的各个三联体密 码连续阅读,密码间既无间断也无交叉。
•基因损伤引起mRNA阅读框架内的碱基发生插入或缺 失,可能导致框移突变(frameshift mutation)。
4、方向性 从 mRNA 5 端起始密码子 AUG 到 3 端终止密码 子之间的核苷酸序列,各个三联体密码连续排列 编码一个蛋白质多肽链,称为开放阅读框架(open reading frame, ORF)。
遗传密码
• 定义:遗传密码(Genetic code)是联系核酸的碱基序列和蛋白质的 氨基酸序列的方式。由三个碱基代表一种氨基酸,称为密码子 (Codon)。 • 遗传密码的发现: • 性质: • 简并 • 不同位置的碱基特异性 • 通用与例外 • 阅读方向 • 不重叠 • 无逗点
一、遗传密码——三联子
线粒体与核DNA密码子使用情况的比较
生物
所有 酵母 果蝇 哺乳类
密码 子 UGA CUA AGA
AUA
线粒体DNA编 码的氨基酸 色氨酸 苏氨酸 丝氨酸 终止子
甲硫氨酸
核DNA编码的 氨基酸 终止子 亮氨酸 精氨酸 精氨酸
异亮氨酸
哺乳类 AGA/G
在一个支原体中,UGA编码色氨酸;但在某些特殊物种中,如纤毛纲中的四 膜虫属及草履虫属,UAA和UAG编码谷氨酰胺。
பைடு நூலகம்
3’
tRNA凭借自身的反密码子与mRNA链上的密码子相 识别,把所带氨基酸放到肽链的一定位置。
(三) tRNA的种类
1、起始tRNA和延伸tRNA
能特异地识别mRNA模板上起始密码子的tRNA称起 始tRNA,其他tRNA统称为延伸tRNA。
真核生物:起始密码子AUG 所编码的氨基酸是Met, 起始AA-tRNA为Met-tRNAMet。 原核生物:起始密码子AUG 所编码的氨基酸并不是
3、校正tRNA
无义突变:在蛋白质的结构基因中,一个核苷酸的改变可
能使代表某个氨基酸的密码子变成终止密码子(UAG、UGA、 UAA),使蛋白质合成提前终止,合成无功能的或无意义的 多肽,这种突变就称为无义突变。 错义突变:由于结构基因中某个核甘酸的变化使一种氨基 酸的密码子变为另一种氨基酸的密码子,这种基因突变叫 错义突变。 GGA(甘氨酸) AGA(精氨酸)
移码抑制恢复了原来的阅读框。最简单的外移码抑制因子修正阅读框的例子 是:当在一系列残基中缺失或插入一个附加碱基而发性突变时。例如,G可 以插入到一系列相邻的G碱在当中。此移码抑制基因是一个有额外碱基插入 到它的反密码环中的tRNAGly。它能转化通常的三联体顺序,这个抑制基因 tRNA能识别一个4碱基密码。
以特定的共聚物为模板指导多肽的合成
(1)以多聚二核苷酸作模板可合成由2个氨基 酸组成的多肽 ,如以Poly UG 为模板,合成产物 为Poly Lys-Val。 (2)以多聚三核苷酸作为模板,可得三种氨 基酸组成的多肽。
核糖体结合技术
技术要点:以人工合成的三核苷酸为模板+核糖体+AA-tRNA 保温
5、碱基特异性
一共有8个密码子家族,每个家 族的4个密码子的前2个碱基是 相同的,而且4个密码子具有相 同的意义,所以它们的第3个碱 基在识别氨基酸过程中不起作 用。一共有7个密码子对,只要 在第3个位置上出现的是嘧啶碱 基,它们将表达相同的意义。 一共有5个密码子对,只要在第 3个位置出现的是嘌呤碱基,它 们所编码的那个氨基酸将不会 改变
无义突变
无义突变使 UUG 变为 UAG
AUG UUG UAA AAC AUG UAG UAA
Tyr-tRNA 阅读 UAG 密码子
AUG UAG AUC UAA UAC AUG
释放因子
抑制突变
Leu
Tyr
Tyr
图 14-17 带有突变反密码子的 tRNA 可抑制无义突变
表 14-6 由反密码子突变而产生的无义抑制基因 基因 tRNA 野生型 识别的密码子 SupD(su1) SupE(su2) SupF(su3) SupC(su4) SupG(su5) SupU(su7) Ser Gln Tyr Tyr Lys Trp UCG CAG UAC, UAU UAC/UAU AAA/AAG UGG 反密码子 CGA CUG GUA GUA UUU CCA 抑制基因 反密码子 CUA CUA CUA UUA UUA UCA 识别的密码子 UAG UAG UAG UAA/UAG UAA/UAG UGA/UGG