神经生长因子及其受体在雄性哺乳动物生殖系统中的作用
神经生长因子及受体在肿瘤中的表达及临床意义

神经生长因子及受体在肿瘤中的表达及临床意义神经生长因子(NGF)是一种多功能的神经营养因子,与特异的靶细胞表面的神经生长因子受体结合,介导产生一系列的生物学效应。
目前研究表明NGF 及其受体与肿瘤的增殖、分化、凋亡、血管形成、转移及预后也密不可分,且由NGF介导的信号通路已成为肿瘤治疗新的靶点。
标签:神经生长因子;受体;肿瘤神经生长因子(nerve growth factor,NGF)是一种多功能、多肽性的物质,具有刺激细胞生长活性的细胞因子。
一类通过与特异的、高亲和的细胞膜受体结合,调节细胞生长与其他细胞功能等多效应的多肽类物质。
它通过特异的靶细胞表面的神经生长因子受体(nerve growth factor receptor,NGFR)介导产生生物学效应。
近年来NGF及NG-FR与肿瘤的关系受到人们的关注。
本资料就神经生长因子及其受体与肿瘤的研究及进展作一综述。
1 神经生长因子神经生长因子是神经系统最重要的生物活性分子之一,是由其效应神经元支配的靶细胞合成和分泌,可以促进感觉神经元及交感神经元的存活及分化,参与损伤修复,维持其生物学功能的稳定。
NGF是由α、β、γ 3种亚基构成的多聚体,β亚基是唯一具有NGF的所有生物活性的亚基,现研究发现前列腺癌、乳腺癌和胰腺癌等肿瘤细胞也可合成NGF等细胞生长因子,并通过自分泌和旁分泌作用于周围的基质成分,来调节肿瘤的生长、分化和浸润行为,包括肿瘤的沿神经浸润。
NGF阳性表达随着肿瘤恶性程度的增高而增高,且与肿瘤的淋巴转移密切相关[1]。
2 NGF的受体(TrkA、p75)2.1 TrkATrkA是一种由原癌基因表达,具有酪氨酸激酶活性的跨膜蛋白,由跨膜酪氨酸激酶gp140Trk组成,是酪氨酸蛋白激酶家族成员之一。
由3个部分组成:(1)辨别并结合NGF的细胞外部;(2)跨膜部;(3)含酪氨酸激酶的胞质部。
TrkA称为高亲和力受体,也叫慢受体或I型受体[2]。
生长激素释放肽及其生物学效应

生长激素释放肽及其生物学效应来景辉;范红结【摘要】Ghrelin, a peptide lying widely in animal bodies, is the endogenous ligand for the growth hormone secretagogue receptor(GHSR). Ghrelin, after binding with GHSR in the hypothalamus and pituitary, not only promotes the GH releasing and increases appetite, but also adjusts the functions of digestive system and energy metabolism. This article mainly reviewed the recent developments on structure, distribution and biological effects of ghrelin as well as its potential applications in animal husbandry, which maybe promotes the further research of ghrelin.%生长激素释放肽(Ghrelin)是一种在动物体内广泛存在的生长激素促分泌素受体(GHSR)的内源性配体,Ghrelin与位于下丘脑的GHSR结合后,具有促进生长激素释放、增加食欲、调节消化系统功能及能量代谢等作用.文章就Ghrelin的结构、分布、生物学效应及在畜牧业上的应用前景等方面进行综述,以期促进Ghrelin的进一步深入研究.【期刊名称】《中国畜牧兽医》【年(卷),期】2012(039)011【总页数】4页(P66-69)【关键词】Ghrelin;生长激素促分泌素;生物学效应;展望【作者】来景辉;范红结【作者单位】宿州职业技术学院动物科学系,安徽宿州 234101;南京农业大学动物医学院,江苏南京 210095【正文语种】中文【中图分类】S852.2生长激素释放肽(Ghrelin)是一种生长激素促分泌素受体(GHSR)的内源性配体。
激素对动物生理和行为的调节作用

激素对动物生理和行为的调节作用激素是一类具有特定功能的化学物质,在动物体内起着重要的调节作用。
通过分泌到血液中,它们可以在整个机体范围内传递信号,并影响动物的生理和行为。
本文将讨论激素对动物生理和行为的调节作用,以及它们在不同动物群体中的作用。
一、激素对动物生理的调节作用激素可以通过影响机体的器官和系统来调节动物的生理功能。
下面我们将介绍一些常见的激素及其生理调节作用。
1. 生长激素(GH)生长激素是一种由脑垂体前叶分泌的激素,对动物的生长和发育起着重要的作用。
它可以促进骨骼和肌肉的生长,并影响身高和体重的增长。
在青春期,生长激素的分泌水平增加,促进青少年的骨骼生长和性成熟。
2. 甲状腺激素(T3和T4)甲状腺激素是由甲状腺分泌的激素,对代谢和能量平衡起着重要的调节作用。
它们可以提高基础代谢率,增加能量消耗,并促进脂肪和蛋白质的分解。
甲状腺激素还对神经系统的发育和功能有影响,对动物的认知和行为具有重要作用。
3. 雄激素(睾酮)雄激素是由睾丸分泌的激素,在雄性动物中起着重要的调节作用。
它们可以促进雄性特征的发育,如声音的改变、肌肉的增加和体毛的生长。
雄激素还可以影响动物的行为,如增加攻击性和争斗行为,促进繁殖行为等。
二、激素对动物行为的调节作用激素不仅对动物的生理起作用,还可以直接或间接地影响动物的行为。
下面我们将介绍一些激素在调节动物行为方面的作用。
1. 雌激素(雌二醇)雌激素是由卵巢分泌的激素,在雌性动物中起着重要的调节作用。
它们可以促进雌性特征的发育,调节生殖周期,并对母性行为起重要作用。
雌激素的水平在发情期间升高,促使雌性动物表现出求偶行为和繁殖行为。
2. 催产素催产素是一种由脑垂体后叶分泌的激素,对动物的社会行为和亲子关系具有重要作用。
在哺乳动物中,催产素可以促进产后子宫收缩,促进母乳分泌。
同时,催产素还可以加强动物之间的联系,促进亲子行为的产生。
3. 应激激素(肾上腺素和皮质醇)应激激素是由肾上腺分泌的激素,对动物的应激反应和适应性行为起着重要的调节作用。
bFGF在男性生殖系统的表达及作用研究

17020212312[17] Shao D, Zhai P, Del Re DP, ct al. A functional interactionbetween Hippo-YAP signalling and Fox O1 mediates the oxidative stress response [J]. Nat Commun, 2014, 5: 3315.[18] Lin Z, von Gise A, Zhou P, et al. Cardiac-specific YAP activationimproves cardiac functionand sur vival in an experimental murine MI model [J]. Circ Res, 2014, 115(3): 354-363.[19] Halder G, Johnson RL. Hippo signaling: growth control andbeyond [J]. Development, 2011, 138(1): 9-22.[20] Dorn GW. Parkin-dependent mitophagy in the heart [J]. J MolCell Cardiol, 2016, 95: 42-49.[21] Morikawa Y, Zhang M, Heallen T, et al. Actin cytoskeletalremodeling with protrusion formation is essential for heart regeneration in Hippo-deficient mice [J]. Sci Signal, 2015, 8 (375): 41.[22] Leach JP, Heallen T, Zhang M, et al. Hippo pathway deficiencyreverses systolic heart failure after infarction [J]. Nature, 2017,550(7675): 260-264.[23] Del Re DP, Yang Y, Nakano N, et al. Yes-associated proteinisoform 1(Yap1) promotes cardiomy ocyte survival and growth to protect against myocardial ischemic injury [J]. J Biol Chem, 2013, 288(6): 3977-3988.[24] Meliambro K, Campbell KN. Hippo pathway deficiency andrecovery from heart failure after myoc ardial infarction: potential implications for kidney disease [J]. Kidney Int, 2018, 93(2): 290292.(收稿日期:2021-01-02)bFGF在男性生殖系统的表达及作用研究崔宁宁,刘一朝,姚美静,刘敏袁王宇婧,母慧袁艾庆燕[关键词1碱性成纤维细胞生长因子;生殖系统;睾丸;精子;前列腺中图分类号R697 文献标识码A文章编号 1004-0188(2021)02-0170-03 doi:10.3969/j.issn.1004-0188.2021.02.024成纤维细胞的生长依赖于成纤维细胞生长因子(fib ro b la s t g ro w th fa c to rs,F G F)。
神经调节因子在生长发育中的作用

神经调节因子在生长发育中的作用在生物体生长发育进程中,神经系统和内分泌系统都扮演了不可或缺的角色。
而人们对于神经系统的了解往往集中在神经元和神经递质方面,而对于神经调节因子的认知则比较少。
本篇文章将介绍神经调节因子的定义、类型、作用及其在生长发育中的具体作用。
一、神经调节因子的定义和类型神经调节因子(neurotrophic factor)是指由神经元、神经胶质细胞、甚至肌肉等细胞产生的一类多样化的细胞因子。
其中最著名的就是神经生长因子(NGF,nerve growth factor)。
目前已知的神经调节因子还包括脑源性神经营养因子(BDNF,brain-derived neurotrophic factor)、神经营养因子3(NT-3,neurotrophin-3)、神经营养因子4/5(NT-4/5,neurotrophin-4/5)等。
这些因子在神经系统的发育、成熟和维持中都起到了重要作用。
二、神经调节因子的作用机制神经调节因子的作用方式主要是通过接触感受器进行信号转导。
由于它们是细胞外介质,因此必须与细胞膜上的受体结合才能起到作用。
在神经系统中,神经调节因子通过与高亲和力受体结合,激活下游信号通路,以此实现多种作用。
这些下游效应通常包括细胞增殖和存活、轴突生长和分支、突触形成和稳定等。
具体来说,神经调节因子可通过以下机制对神经细胞产生影响:1、促进突触的形成和稳定神经调节因子可以通过特定的受体,促进神经元之间的突触形成和稳定。
神经调节因子可使突触上调第一信使,在精细的突触关联中发挥重要的控制作用。
例如,神经生长因子可影响海马的神经递质释放和长时程增强等功能。
2、促进神经细胞的生存和发育神经调节因子不仅能影响成熟神经元的生存,也可以影响神经元的发育。
例如,神经生长因子在神经元发育中起到了重要作用,通过激活Akt、ERK等信号通路促进神经元的生长、骨骼蛋白表达和突触形成等过程。
3、增强神经细胞的功能神经调节因子也可提高神经细胞的功能。
【doc】PPARs信号通路与哺乳动物生殖

PPARs信号通路与哺乳动物生殖细胞生物学杂志ChineseJournalofCellBiology2005.27:1418http://www. PPARs信号通路与nSV#L动物生殖赵越超杨增明(东北农业大学生命科学学院,哈尔滨150030)摘要过氧化物酶体增殖因子活化受/e~(peroxisomeproliferator-activatedreceptors,PPARs1在动物体内有着广泛的生物学作用,可调节脂类代谢,能量收支平衡以及细胞分裂分化等重要生理过程.已经发现,PPARs信号通路与糖尿病和癌症等许多重大疾病的发生有关.随着基因剔除技术的应用以及PPARs人工配体的开发利用,.人们对PPARs的认识不断深入.现对PPARs通路在卵巢周期,黄体形成,胚胎着床,胎盘发育和雄性生殖等哺乳动物生殖系统中的表达,功能及作用机制进行综述.关键词过氧化物酶体增殖因子活化受体:卵巢;着床;胎盘;雄性生殖1PPA信号通路概述PPARs是细胞核激素受体超激素受体家族成员,定位于细胞核上,可由配体激活.在动物体内一般存在3种PPARs,即PPARct,PPAR[~(也称PPARS)和PPARy.PPARs与配体结合后可被激活,然后与目标基因结合,可在转录水平调节目标基因蛋白质产物的活性,进而参与调节脂类代谢,机体免疫,细胞分化及细胞凋亡等生理功能.3种PPARs同各自配体结合后可参与调节不同的生理过程:(1)PPAR0c参与脂肪酸代谢和炎症反应.(2) PPAR[~参与调节胚胎着床,细胞增殖和凋亡.(3) PPARy调节脂肪细胞分化,单核细胞分化及退出细胞周期等….从目前研究进展来看,PPARs在哺乳动物生殖过程中起重要作用.1990年首次克隆并筛选得到了PPARctcDNA, 1992年又获得了PPAR[~和PPARy的cDNA.3种PPARs间同源性很高,它们与甲状腺激素,类维生素A,维生素D,蜕皮激素等分子一样,都起源于细胞核受体超家族.PPARs同配体结合后,作用于过氧化物增殖因子反应元件(peroxisomeprolif- eratorresponseelements,PPREs),从而调节靶基因表达.在结构上,PPREs为同向重复的基因序列,根据3种PPARs对其DNA结合能力的不同,可将天然的PPREs分成3类:强应性的,一般应性的和弱应性的.PPARs可特异性地识别6核苷酸序列AGGTCA,还能与9位顺式视黄酸受体(9一cis—retinoicacidreceptor,RXR)形成二聚体,进而作用于靶基因.RXR是一个常见的DNA结合参与者,它可与包括PPARs在内的许多类固醇/甲状腺受体超家族的核受体协同作用.如果有配体和RXR作用,也可激活PPARs:RXR二聚体信号通路.现已确认,一些不饱和脂肪酸及其衍生物可作为3种PPARs的天然配体.例如,类花生酸类物质是一类主要源白花生四烯酸的脂肪酸,可在环氧合酶(cycloxygnase,cox)作用下生成各种前列腺素(prostaglandins,PGs).PGD2的衍生物15一脱氧一A12,14一前列腺素J2(15一deoxy—A12,14一prostaglandinJ2,15d—PGJ2)即是PPARy的配体.另外还有些人工合成药物也可与PPARs作用,例如花生四烯酸类似物ETY A可同3种PPARs结合;纤维类的低血脂药物(hypolipidemic drugs)n~激活PPARct,进而调节许多和脂肪酸代谢相关的基因;而PPARy可由thiazolidinedione类的低血糖药物激活,然后调节脂肪细胞分化【3】.PPARs的表达主要与线粒体及过氧化物酶体的氧化活性有关,而且3种PPARs在各种组织中经常协同表达.PPARct在肾脏,心脏,肌肉和肝脏等组织中高水平表达,PPAR[~在许多组织都有表达,而PPARy则主要存在于脂肪组织,单核细胞,巨噬细胞以及胎盘组织【4】.在卵巢,子宫和胎盘等雌性生殖系统中,可检测到PPARsmRNA和蛋白质,并且在睾丸等雄性生殖器官组织也可检测到PPARs表达,这提示PPARs在哺乳动物生殖过程中起重要的调节作用.收稿日期:2004.01.29接受日期:2004.08.25通讯作者.Tel:0451.55191416;Fax:0451.55103336;E.mail ****************赵越超等:PPARs信号通路与哺乳动物生殖l52PPARs和卵巢功能卵巢是一个周期性变化的器官,其周期可分为卵泡期,排卵期和黄体期.卵巢的主要功能是排卵和分泌类固醇激素.黄体是卵巢内一个暂时性的内分泌器官,所产生的孕酮是妊娠建立和维持所必需的.黄体的形成和功能涉及到脂类代谢,血管发生,甾类激素合成及前列腺素的产生等许多过程.假孕或发情周期的大鼠卵巢中,在发育卵泡的颗粒细胞内有高水平的PPARymRNA表达.PPARctmRNA在发情周期的大鼠黄体中高水平表达,并且在由上一发情周期来的黄体中表达量升高.PPARct mRNA主要定位于卵泡膜和卵巢基质,其水平在发情期较低.PPAR~mRNA在整个卵巢中都有分布,其水平在假孕或整个发情周期中均保持稳定15】. PPART能参与排卵及黄体生成.PPARct可能在卵泡膜和卵巢基质的脂类代谢过程中起作用.PPAR~可能参与调节维持卵巢的基本功能. PPARymRNA在注射PMSG后的卵巢中高水平表达,提示其在卵泡发育过程中起作用.在hCG处理(模拟LH脉冲)后,PPARymRNA水平降低,暗示PPART~颗粒细胞的黄体化过程可能起抑制作用【6】.在牛的黄体组织中发现PPARy的蛋白质水平在发情周期的黄体早期和中期有所下降.此外, PPART的类似物也能够影响体外培养的大鼠,猪和人颗粒黄体细胞孕酮的产生.在大鼠的发情周期中,LH峰后PPAR7mRNA水平降低.在黄体形成初期,PPAR7表达也很低,而孕酬在这一时期表达增强.随着黄体期的逐步进行,黄体开始退化,孕酮的表达也逐步降低,但PPAR7mRNA的表达却又升高,这些发现表明孕酮和PPARy的水平呈反比关系【5】.但用PPARy激动剂处理体外培养的处于周期中期的牛黄体细胞时,发现孕酬分泌增强,表明PPAR7可通过促进孕酮分泌来影响牛黄体细胞的功能,这很可能是种属差异或黄体分化时不同的细胞反应造成的.在人乳房的脂肪和由颗粒细胞向黄体细胞转变的组织中,芳香酶活性可因PPARy激活而受到抑制,提示PPARy可能也参与黄体组织中类固醇生成酶活性与表达的调节.在体外培养的猪和人的颗粒细胞中,PPAR7对孕酮合成有抑制作用,这可能是由于它能够降低3p.类固醇脱氢酶的活性16】. PPARY可能还与20.羟基类固醇脱氢酶(20一hydroxys—teroiddehydrogenase,20.HSD)的表达相关,该酶可将孕酮转化为非活性的20一二羟基孕酮(20一dihydroprogesterone).在大鼠和,J,鼠中,20一HSD在黄体退化时孕酮分泌降低过程中起重要作用[5t.另外,PPARs也能调节COX一2的表达,而且类花生物质又可激活PPARs,表明PPARs活性和前列腺素合成之间存在着反馈调节系统.前列腺素在黄体形成及退化过程中起重要作用,特别是前列腺素F20c(prostaglandinF2ct,PGF2ct)可通过调节孕酮分泌来诱导黄体退化,这表明PPARs还可能通过调节COX一2一PGs系统来影响黄体功能.PPARy在发育卵泡的颗粒细胞中的高度表达可能与卵泡雌激素的分泌相关.PPARs与雌激素反应元件结合后,可阻止雌激素反应元件和雌激素受体的结合,从而抑制雌激素活性【6】.PPARs还能调节芳香酶的活性和表达,而芳香酶参与调节雌激素的生物合成.此外,MEHP(monoethylhexylphthalate), PPARct和PPART的特异性配体在体外均能够降低雌激素的分泌和芳香酶mRNA的表达水平.MEHP可抑制芳香酶活性并激活PPARs(TJ.因此,MEHP可能通过PPARs介导的信号通路来抑制卵巢雌激素的分泌,从而导致排卵失败.PPARs也能够影响黄体形成中的血管发生和组织重塑等过程.PPARs可以调节一些蛋白水解酶的表达和活性,而这些酶类在许多种动物的卵巢组织中均有分布,说明PPARs可能通过调节它们的活性,来影响黄体的形成和退化等过程.体内及体外研究显示,PPART的激活可抑制血管发生.纤溶酶原激活因子可调节尿激酶型纤溶酶原的活性, PPART又可促进纤溶酶原激活因子mRNA的表达, 并可能进而抑制血管发生.而且,PPART还能下调血管内皮生长因子受体的表达【8】.另外,PPART 的激活可降低巨噬细胞和血管平滑肌细胞中一氧化氮的合成,还可抑制内皮细胞分泌内皮素一l.因此,除调节血管发生外,PPARs还可通过抑制内皮素一l和一氧化氮的合成来影响卵巢血管扩张.总之,3种PPARs在哺乳动物卵巢中都有表达分布,其中PPART可通过调节孕酮分泌,影响雌激素活性,COX.2.PGs系统以及一些血管相关因子等途径来参与排卵及黄体生成等过程.3PPARs和着床COX可将花生四烯酸转化为前列腺素H2 (prostaglandinH2,PGH2),而PGH2是各种前列腺素l6综述合成酶的共同底物.COX以两种亚型存在:COX一1和COX一2.COX一1缺失的雌性小鼠有生育能力, 但分娩时存在一定缺陷;COX一2缺失的雌鼠则表现出很多生殖功能上的障碍,如卵细胞成熟,排卵,着床及蜕膜化的失败.核膜与内质网膜均可表达COX一1和COX.2.内质网合成的PGs可出入细胞, 并通过G蛋白相联的细胞表面受体来行使功能.相反,由细胞核COX合成的PGs能与PPARs结合,从而直接在细胞核内发挥效应.在子宫着床位点处特异性表达COX一2,但检测不到COX一1,并且COX一2基因剔除小鼠不能正常着床和蜕膜化,说明COX一2来源的PGs参与早期妊娠的建立.在检测小鼠早期妊娠子宫的各种前列腺素含量时发现,PGI2的水平最高,而且它在子宫着床位点的水平要明显高于非着床位点,推测COX一2来源的PGI2可能在胚泡着床和蜕膜化过程中起重要作用[io1.与其他前列腺素一样,PGI2可与细胞膜表面G蛋白偶联的PGI2受体(IP)结合.IP激活后可通过刺激腺苷酸环化酶来促使细胞内cAMP水平升高.血管内皮及其下面的平滑肌等血管组织能够通过前列环素合成酶(PGIS)来合成PGI2.通过PGI—IP信号通路,PGI2可作为血管扩张因子和抗血凝剂,作用于血管组织和血小板,而且用PGI2类似物可模拟这些效应.PGI2也可与PPAR~结合来调节特定的细胞功能f】11.COX一2,PGIS,PPAR~和RxR在着床胚泡周围的基质细胞中协同表达,提示这些蛋白质之间可能存在一个信号级联系统在着床过程中起作用.COX一2和PGI2在基质细胞核_J的表达位点相近,表明两者在合成位点处可直接通过与PPAR[I结合来起作用.已知的IL1),PPAR~和PPAR(t 等PGI2受体中,在黏附反应起始阶段以及蜕膜化过程中的子宫中只有PPAR~表达,表明PPAR~与着床关系密切【101.用cPGI(carbarprostacyclin)或L一165,041等PPAR~的特异性配体处理,可恢复cOx一2缺失小鼠中的着床缺陷.尽管这些配体在结构上没有同源性,但它们在调节PPAR~转录方面的活性却相似,而且视黄酸(9一cis—retinoicacid,9-cis—RA)还可显着上调这种活性.PPAR~和RXR配体协同,可上调PPAR~的转录活性.已证实,用L一165,041和9一cis—RA共同处理,可提高COX2一,-小鼠的着床率[1o】.在蜕膜细胞核中,PPAR~/RXR异二聚化的增强或稳定可进一步提高其对PPAR~I配体的反应性,进而促进SRC一1等转录激活因子的募集.由于SRC一1缺失小鼠的着床率降低,SRC一1可能参与子宫蜕膜反应.通过检测一系列血管生成前标记物表达发现,cPGI或PPAR~激动剂能够弥补COX一2缺失小鼠的着床缺陷,同时着床位点的血管生成也得到恢复,但IP或PPAR~都有可能参与PGI2在血管系统中的这种效应.我们的结果也表明,在大鼠子宫着床位点处的腔上皮下基质中可检测到高水平的PPAR[~mRNA和蛋白质,而且在这些部位也可检测到RXR(t蛋白质表达.PPARB表达是由活性胚泡刺激的,因为在假孕第6天大鼠子宫中未检测到其表达信号D21.这些结果和在小鼠早期妊娠过程中发现的类似,表明PPARBxR二聚体在大鼠着床过程中也起重要作用.在着床位点处,许多哺乳动物的子宫腔上皮细胞发生细胞凋亡.由于PPAR~与细胞内源性配体PGI2结合后可以诱导细胞凋亡,推测PPAR~可能还与着床过程中的细胞凋亡相关.尽管已有许多文献报道PPAR~在胚胎发育和着床中起广泛作用,但它的特异性受体是否为PGI2或其他内源性的配体,还需要进一步确定.因为PPAR~可结合多种配体,体内或体外PPAR~的激活并非只涉及到PGI2.在PPARs配体结合域存在一个大腔,PPARs还可被亚油酸和花生四烯酸等多聚不饱和脂肪酸以及一些人工合成药物激活.因此,在胚泡着床过程中,很可能还涉及到其他和PPARs相关的作用因子和信号通路.总之,应用基因剔除以及人工配体,激动剂处理等技术方法发现在哺乳动物着床过程中, PPAR~与RXR形成二聚体,通过细胞核上的cOx. 2一PGI2通路来参与着床过程.4PPA和胎盘发育胎盘由胎儿和母体共同构成,是两者进行物质交换,营养,代谢,激素分泌,防止异源物质入侵以保证胎儿正常发育的一个重要器官.胎盘形成是一个复杂的组织重构过程,包括滋养层侵入,蜕膜反应,细胞外基质(ECM)降解及血管形成等过程.在人的胚胎中,合体滋养层具有多个细胞核,最终分化为覆盖在胎盘绒毛外表面的细胞团,因此直接和母体血液接触.细胞滋养层分化为合胞体滋养层这一过程,对于胎盘功能甚至胎儿发育是极为重要的.赵越超等:PPARs信号通路与哺乳动物生殖l7 PPAR丫在前脂肪细胞(preadipocytes),成肌细胞以及单核细胞等细胞的分化过程中都起作用,也参与乳癌和脂肪肉瘤细胞等细胞的最终分化过程[41. PPARy缺失的小鼠表现出胎盘发育和滋养层分化异常,在胎盘中还出现异常的血管发生现象.这些发育缺陷可导致胚胎在第l0天死亡[131.在人细胞滋养层和合体滋养层中均有PPARy表达.当滋养层细胞在H/W培养基(该培养基已知可抑制滋养层分化)中培养时,PPART的表达减弱.这些研究结果提示PPARy在母体胎盘的滋养层分化过程中起重要作用.在RXR~x或P.XRI3基因剔除的小鼠中,胎体不能形成具有正常功能的尿囊绒膜胎盘,母体.胎儿问的物质交换因而受阻,最终导致流产[131.PPARy缺失的胎儿也因滋养层分化和血管发生异常导致胎盘发育不全.这些发现提示,PPARy/RXR异二聚体对小鼠着床和正常胎盘形成是必需的.近来发现,RXR或PPARy的激活可刺激人绒毛滋养层的分化和内分泌功能.另外,在人妊娠前3个月的胎盘中,PPARy和RXRt~在位于整个锚定绒毛上的绒毛外滋养层细胞核中协同表达.人等哺乳动物的滋养层和蜕膜细胞能够合成反式视黄酸(all—trans—retinoic acid)和其9一顺式同工分子,两者均为RXR的天然配体【14】.人的子宫内膜和蜕膜也表达COX和PGs【l5】, 这些分子很可能成为PPARy的配体.胎盘组织中也表达各种PGS和脂类物质【1引.这些结果表明,PPARy/RXRtx二聚体可作为控制人细胞滋养层分化和侵入的转录水平调控因子.PPARy和RXRt~在蜕膜区的绒毛外滋养层中虽然协同表达,但PPARy和RXRt~不同,它只在绒毛外滋养层中特异性表达,而在蜕膜细胞中则无表达.这表明PPARy作为细胞核受体,可能在绒毛外滋养层的侵入过程中也起重要作用[141.PPARy还可调节基质金属蛋白酶等炎症因子基因的表达,并调节正常或肿瘤细胞的迁移性和侵入性.然而, PPART/RXRt~异二聚体是否能在细胞滋养层侵入过程中调控蛋白水解酶的表达还需要进一步研究. PPARy有许多种天然和人工合成的配体.在一种细胞中,同一受体和不同的配体结合可能产生完全不同的生理效应.PPARy的两种配体曲格列酮(troglitazone)和15d—PGJ2对人滋养层细胞的分化产生截然相反的效应,前者可促进分化,而后者则能抑制分化并促进凋亡.由于曲格列酮可诱导人滋养层细胞的分化,而且PPART基因缺失小鼠的滋养层分化异常,提示在体内可能存在一个内源性的和曲格列酮类似的配体,在滋养层分化过程中起作用I4】. 曲格列酮和15d—PGJ2在滋养层中所起的作用不同, 可能是由于配体和受体蛋白在空问布局上发生了变化,导致在与特定的协同调节子作用时,一种协同调节因子可刺激目标基因表达,而另一种则抑制目标基因的表达.在人分娩期间,羊膜,绒毛蜕膜(choriodecidua)和胎盘中的PPARymRNA水平并未改变;PPARtx mRNA在羊膜中的表达也没有显着改变;而PPAR[3 的表达却显着提高.在绒毛蜕膜中,PPARtx表达在分娩期下降,而PPAR~I水平却升高.在胎盘中, PPARtx和PPAR~的表达都升高[161.这表明PPARs 在维持妊娠或启动分娩过程中起作用.总之,在胎盘组织中,PPARy可能通过体内某种配体激活形成PPART/RXR二聚体,进而调节细胞滋养层分化及侵入等过程.5PPA和雄性生殖过氧化物酶体增殖因子(peroxisomeproliferators, PPs)是一大类工业和药用物质,现已成为普遍存在的污染物质,可激活小鼠和大鼠体内许多种过氧化物酶体.睾丸中Leydig细胞功能和睾酮合成的异常可导致雄激素依赖的雄性生殖系统组织发育缺陷, 并直接影响成年个体的睾丸功能,包括精子发生和生育能力.最近发现,一些PPs对雄激素表现出抗性效应N7].一般认为,类固醇生成是由促激素(trophic hormone)调节的,这些激素可促进胆固醇从贮藏或合成位点向线粒体内膜运输.PPs可阻止由促激素诱导的这一运输作用,并进而影响雄性生殖系统. PPs可以激活外周受体苯并二氮卓(peripheral—type benzodiazepinereceptor,PBR)的基因转录.PBR基因编码一个对线粒体胆固醇具有高亲和性的蛋白质,该蛋白质参与调节胆固醇跨膜运输.研究发现,PPs诱导的PBR基因转录是由PPARtx介导的. PPARtx缺失小鼠的循环系统中,睾酮的水平明显要比野生型低,这表明PPARt~对于正常的雄性类固醇激素的合成是必需的】.PPARy在雄性生殖过程中也起一定作用.用DBP(di—n—butylphthalate)处理睾丸组织后,纤溶酶原激活因子的抑制因子一I(PAI.1)mRNA的水平显着提高,可作为PPARy激活的标志,而PAI—l水平的提高可能和精子发生的破坏有关,从而提示18综述.PPAR丫可能参与调节精子发生[191.在非生殖系统组织中,PPARy在多种癌变细胞中表达,而且其配体可通过细胞凋亡来抑制这些癌细胞的生长.在正常的和癌变的睾丸组织中,均可检测到高水平的,有免疫活性的PPAR~和PPAR~蛋白,但PPARy只在癌变的睾丸组织中显着表达.并且,人工合成的PPARy配体(thiazolidinedione)和内源性的配体(15一deoxy.delta-prostaglandinJ2)均可抑制睾丸癌细胞的生长【20】.在睾丸癌细胞中PPARy表达的上调表明, PPARy配体可能对睾丸癌细胞有抗增殖作用.因此,在睾丸癌症治疗领域,PPAR7已成为新的研尽管上述研究表明PPARs参与雄性激素合成以及睾丸癌变等过程,但目前还很少有直接证据表明PPARs和雄性生殖作用紧密相关.6小结从目前积累的研究结果来看,PPARs广泛参与哺乳动物许多生殖过程.虽然已证实PPARI~在小鼠胚胎着床过程起关键作用,但是将PPAR~基因剔除后,小鼠的生殖过程却未见异常【z”.随着RNA干涉技术的逐步完善和发展,将有利于进一步研究PPARs信号通路在生殖过程中的作用机制.[1】[2】[3】[4】[5】【61[7】【81[9】[10】【l11[13】【14】[15】161718192O21参考文献(References)HihiAKeta1.CellMolLifeSc-.2002.59:790 DesvergneBeta1.EndocrRev.1999.20:649 KerstenSeta1.ExS.2000.89:141SchaiffWTeta1.JClinEndocrinolMetab,20oo.85:3874 KomarCMet口,Reprod.2002.66:l531KomarCMeta1.Endocrinology,2001.142:4831 Lovekamp—SwanTeta1.Em,ironHealthPerspect,2003,111: 139XinXr口,..,BiolCheml999.274:9ll6NegishiMeta1.BiochimBiophysActa,1995,1259:109 LimHeta1.GenesDev.1999.13:1561LinlHeta1.Endocrinology,2002,143:3207DingNZeta1.Reproduction,2003,125:817WendlingOeta1.ProcNatlAcadSciU,1999,96:547TarradeAeta1.JClinEndocrnolMetab.2ool,86:5017ShawKJ口ProstaglandinsLeukotEssentFattyAcids,l99l4.50:239BerryEBeta1.MolPharmaco1.2003,64:l586MooreRWeta1.EnvironealthPerspect.2oo1.109:229GazouliMet口L£ndocrinology.2002.143:2571KobayashiTeta1.ToxicolLett,2003,138:215HaseTeta1.Urology,2002.60:542PetersJM£fa1.MolCellBio1.2000.20:5l19 PPARsSignalingPathwayinMammalianReproductionY ue—ChaoZhao.Zeng—MingY ang (CollegeofLifeSciences,NortheastAgriculturalUniversity,Harbin150030 ,China)Peroxisomeproliferator-activatedreceptors(PPARs)playimportantrolesin manybiologicalprocesses.includingmediationoflipidmetabolism,energybalance,celldiffe rentiationanddivision.Ithasbeen confirmedthatPPARssignalingpathwayisalsorelatedtosomepathologicalp rocesses,suchasdiabetesandcancer.TheunderstandingonPPARshasimprovedastheapplicationofgenek nockouttechnologyandtheartificialligands.ThisarticlereviewsPPARsexpression,functionandmechanisminm ammalianreproductivesystemduring theprocessesofovariancycle,lutealformation,embryoimplantation,placen tationandmalereproduction. Keywordsperoxisomeproliferator—activatedreceptors;ovary;implantati on;placenta;malereproductionReceived:January29.2004Accepted:August25,2004*Correspondingauthor.Tel:86—451—55191416.Fax:86—45l-55103336 ,E—mail:****************.edu-ca。
神经生长因子对雄牲动物生殖系统作用的研究进展

ቤተ መጻሕፍቲ ባይዱ
水平。研究证 明I, 1 雄激素通过与雄激素受体 ( n r e epe , 5 1 A do n eetr g r s A) R结合 , 可促进 N F的表达 。A G R的表达和 N F的表达呈明显 G
的 正 相关 旧。
雄 性 生殖 器官 中广泛 存 在 N F及 其受 体 n re rwhf t 素 。M ub re t的实 验 结 果 显 示 睾酮 能 够 影 响 N F的一 些 G ev o t c r g ao e sugr M1 S 3 1 G rcprr G R, 们 可 能在 雄 性 生 殖 系统 的发 育 、 态 、 能 以 作 用 , 且存 在 复 杂 的交 互 作用 。K th e  ̄4 实验 证 实 , 陆 eet , F ) oN 它 形 功 而 a mb - oS 1 的 雄 及病理过程中起着重要的调节作用H 。而且以往多种研究发现 , 大 鼠阉割后导致血清睾酮含量和脑 、 脊髓 、 下颌下腺 N F明显下 G N F在 多种 实 验 动 物 睾 丸 组 织 中均 有 表 达 日说 明 N F参 与 了 降 , 射 睾酮 后 , 、 G , G 注 脑 脊髓 和下 颌 下腺 N F水 平 又恢 复 至 正 常对 照 G
生理 功能方 面也 有重要 作用 。本 文主要论 述 了 NGF对雄性动 物生 殖 系统的 作用 。
关键 词: 经生长 因子 ; 丸组 织; 激素 ; 神 睾 雄 精子
中 图分类号 : 1 S8 3 文 献标识码 : C 文章编 号:0 1 0 6 (0 )2 0 8 -2 1 0 — 7 92 1 0 — 0 9 1
l前 言
15 年 L v Mot e i 蛇 毒 中首 先 发 现 神 经 生 长 因 子 99 ei na i 在 — ln nr o tfc rN FN F的 发 现 被 认 为 是 神 经 科 学 发 展 史 ev g wh at , G ) G er o , 上 的重 要 里 程碑 ,G N F对 于 神 经 细胞 的 生 长 、 育 和 存 活 是 最 为 发
专业辨析神经生长因子(NGF)的神话

专业辨析神经生长因子(NGF)的神话命基111 陈俊青10111115摘要:神经生长因子(NGF)是神经营养因子中最早被发现,目前研究最为透彻的,具有神经元营养和促突起生长双重生物学功能的一种神经细胞生长调节因子,它对中枢及周围神经元的发育、分化、生长、再生和功能特性的表达均具有重要的调控作用。
但是神经生长因子在临床应用上尚未成熟,它的诸多作用还是理论上的推测。
关键词:神经生长因子(NGF)、神经营养因子、神经细胞、神经生长因子的中国神话正文:1.概念[3]神经生长因子nerve growth factor 略称NGF。
在将小鼠肉瘤180移植于3日龄鸡胚体壁时,与移植片连接的脊髓感觉神经节及交感神经节增大20%—40%,基于比克尔(E.D.Bueker,1948)的这一发现,科恩(S.Cohen,1954)等从小鼠肉瘤180和37中成功地分离出具有同一活性的核蛋白质,以后从蛇毒中分离出具有千倍活性(Cohen,R.Levi-Montalcini,1956)和从小鼠颚下腺分离出具有万倍活性的蛋白质(Cohen,1960),这种蛋白质被称为神经生长因子。
NGF含于马、猪的颚下腺和小鼠肉瘤、小鼠胚胎及成体的交感神经细胞、小鼠尿和唾液、鸡胚的多种器官和一切哺乳类的血清中。
神经生长因子(NGF)是神经营养因子中最早被发现,目前研究最为透彻的,具有神经元营养和促突起生长双重生物学功能的一种神经细胞生长调节因子,它对中枢及周围神经元的发育、分化、生长、再生和功能特性的表达均具有重要的调控作用。
NGF包含α、β、γ三个亚单位,活性区是β亚单位,由两个118个氨基酸组成的单链通过非共价键结合而成的二聚体,与人体NGF的结构具有高度的同源性,生物效应也无明显的种间特异性。
2.发现人与研究历程[2]、[6]2.1 1953年意大利生物学家丽塔·莱维-蒙塔尔奇尼(Rita Levi-Montalcini)和美国生化学家斯坦利·科恩(Stanley·Cohen)发现并分离提纯出神经生长因子(NGF)。
前列腺增生手术:剜除好还是切除好?

前列腺增生手术:剜除好还是切除好?在临床上,前列腺增生是一种比较常见的男性疾病类型,该疾病多见于50周岁以上的中老年人男性,随着我国老龄化问题的加重,前列腺增生的法神能够率也在不断提高,严重威胁着患者的身体健康,目前,手术是治疗前列腺增生的常用疗法,但广大患者对于前列腺增生手术中,是选择剜除、还是切除存在疑虑。
为此,本文便针对前列腺增生手术患者的术式选择相关知识进行简单的科普,现报道如下。
1.什么是前列腺增生1.1 前列腺前列腺是男性所特有的一种腺体,属于男性生殖器附属腺体中最大的实质性器官,具有分泌前列腺液和控制排尿等作用,可对于男性正常生殖功能和排尿功能的保障有着重要的意义。
1.2 前列腺增生的概念顾名思义,前列腺增生指的便是人体前列腺组织所发生的细胞增生和体积增大情况,是一种常见于中老年男性的生殖系统疾病类型。
1.3 前列腺增生的流行情况在临床上,前列腺增生仅见于男性,且该疾病会随着男性年龄的增长而逐渐提高。
据统计,40岁-50岁的男性发生前列腺增生的概率约为20.00%,51岁-60岁的男性发生前列腺增生的概率则超过50.00%,80岁以上的男性发生前列腺增生的概率更是在90.00%以上,前列腺增生是一种随着男性年龄增长而不断提高发病率的疾病类型。
1.4 前列腺增生的病因目前,医学界尚未对前列腺增生的具体病因进行明确,但学界对于前列腺增生的病因主要有四大学说。
1.4.1 基本病因1.4.1.1 性激素及受体作用现代医学研究发现,雄激素能够作用于前列腺细胞,从而对前列腺细胞的生长、增殖和凋亡造成影响,故雄激素及其受体同前列腺增生的发生有着密切的联系。
1.4.1.2 细胞增殖与凋亡失衡前列腺细胞增长与凋亡的平衡是决定前列腺大小的根本原因,故一旦前列腺细胞的增殖与凋亡失衡,便会导致前列腺增生的发生。
1.4.1.3 生长因子神经递质作用现代医学研究发现,多种生长因子会控制前列腺细胞的增殖与凋亡,故生长因子的神经递质作用同前列腺增生间有着密切的联系。
神经病理性疼痛中神经生长因子的作用及其相关镇痛药物研发进展

神经病理性疼痛中神经生长因子的作用及其相关镇痛药物研发进展神经病理性疼痛是躯体感觉神经系统损伤或功能紊乱而导致的疼痛,全球约有6% ~8%的人患有神经病理性疼痛。
神经病理性疼痛病程短则数月长则数年,常伴随睡眠障碍、焦虑、抑郁,严重影响患者的生存质量。
神经生长因子(nerve growth factor,NGF)是神经营养因子家族成员之一,广泛分布于外周及中枢神经系统、骨骼肌以及腺体中,在胚胎发育、免疫调节、造血等方面具有重要作用。
近年来,NGF及其受体已成为治疗神经病理性疼痛的新靶点,人源化NGF单克隆抗体已进入临床试验阶段,具有良好的临床应用前景。
本文就目前NGF参与调节神经病理性疼痛的具体机制和调节NGF的镇痛药物的研发进展进行综述,以期为疼痛研究和相关药物研发提供参考。
1 神经生长因子参与调节神经病理性疼痛的作用机制1.1 神经生长因子的异常表达在胚胎和幼年动物体内,NGF主要由靶组织(包括皮肤、肌肉和血管组织)内神经合成分泌,促进神经纤维生长、分化和损伤修复;正常成年机体内中枢和外周神经纤维的生存不依赖于NGF,神经系统内NGF蛋白水平极低,而损伤、炎症等情况下炎症细胞及神经胶质细胞、施旺细胞(Schwann cell)、成纤维细胞、上皮细胞、内皮细胞、结缔组织、肌肉细胞等可大量分泌NGF,调节免疫反应,促进损伤修复,维持机体稳态。
研究发现,NGF的异常表达与神经病理性疼痛密切相关。
在坐骨神经慢性压迫损伤(chronic instruction injury,CCI)[1]、脊神经结扎(spinal nerve ligation,SNL)[2]、紫杉醇[3]等诱导的神经病理性疼痛动物模型中NGF蛋白水平异常升高,临床神经病理性疼痛患者[4] NGF表达上调。
其中,在CCI诱导的神经病理性疼痛模型中术侧坐骨神经[5]、背根神经节(dorsal root ganglion,DRG)[6]、脊髓背角[7] NGF蛋白或转录水平均显著增高。
2014年四川农业大学本科生科研兴趣培养计划项目题目
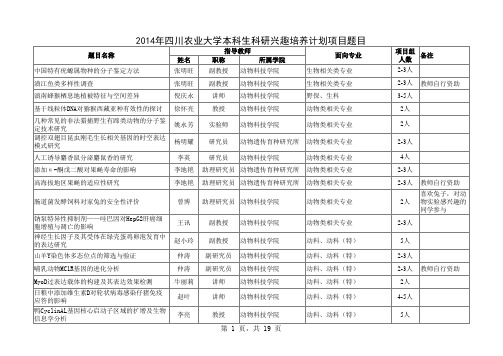
项目组 备注 人数 2-5人 2-4人 2-3人 2-3人 3人 5人 5人 5人 5人 5人 2-3人 2-3人 2-3人 2-3人 2-3人 2-3人 5人 教师自行资助 教师自行资助 教师自行资助 教师自行资助 教师自行资助
动物科技学院 动物科技学院 动物科技学院 动物科技学院 动物医学院 动物医学院 动物医学院
2014年四川农业大学本科生科研兴趣培养计划项目题目
题目名称 中国特有疣螈属物种的分子鉴定方法 濆江鱼类多样性调查 滇南蜂猴栖息地植被特征与空间差异 基于线粒体DNA对猕猴西藏亚种有效性的探讨 几种常见的非法猎捕野生有蹄类动物的分子鉴 定技术研究 调控双翅目昆虫刚毛生长相关基因的时空表达 模式研究 人工诱导麝香鼠分泌麝鼠香的研究 添加α -酮戊二酸对果蝇寿命的影响 高海拔地区果蝇的适应性研究 肠道菌发酵饲料对家兔的安全性评价 钠泵特异性抑制剂——哇巴因对HepG2肝癌细 胞增殖与凋亡的影响 神经生长因子及其受体在绿壳蛋鸡卵泡发育中 的表达研究 山羊Y染色体多态位点的筛选与验证 哺乳动物MC1R基因的进化分析 MyoD过表达载体的构建及其表达效果检测 日粮中添加维生素D对轮状病毒感染仔猪免疫 应答的影响 鸭CyclinAL基因核心启动子区域的扩增及生物 信息学分析 姓名 张明旺 张明旺 倪庆永 徐怀亮 姚永芳 杨明耀 李英 李地艳 李地艳 曾博 王讯 赵小玲 仲涛 仲涛 牛丽莉 赵叶 李亮 指导教师 职称 副教授 副教授 讲师 教授 实验师 研究员 研究员 所属学院 面向专业 生物相关类专业 生物相关类专业 野保、生科 动物类相关专业 动物类相关专业 动物类相关专业 动物类相关专业 动物类相关专业 动物类相关专业 动物类相关专业 动物类相关专业 动科、动科(特) 动科、动科(特) 动科、动科(特) 动科、动科(特) 动科、动科(特) 动科、动科(特) 项目组 备注 人数 2-3人 2-3人 3-5人 2人 2人 2-3人 4人 2-3人 2-3人 2人 2-3人 5人 2-3人 2-3人 2人 4-5人 5人 教师自行资助 教师自行资助 喜欢兔子,对动 物实验感兴趣的 同学参与 教师自行资助 动物科技学院 动物科技学院 动物科技学院 动物科技学院 动物科技学院 动物遗传育种研究所 动物科技学院
BDNF 及其受体在多囊卵巢综合征大鼠中的表达

BDNF 及其受体在多囊卵巢综合征大鼠中的表达∗吴敏敏;蒋立艳;操思凡【摘要】Objective To investigate the expression of Brain-derived neurotrophic factor (BDNF)and its receptors mRNAs in ovary tissues of letrozole-induced polycystic ovary syndrome (PCOS)in rats.Methods The SD rats were divided into two groups (moder group and control group),20 rats in each group.The rat models of PCOS were established by letrozole.Serum sex hormone levels were determined by radioimmunoassay.The histologic changes in ovaries were observed by Hematoxylin-eosin stai-ning,and the expression of BDNF and its receptors gene in ovary tissues was detected by Real-time PCR.Results Although the se-rum testosterone,follicle-stimulating hormone and luteinizing hormone levels in model group were markedly increased more than those in the control group (P < 0.01 ),estradiol and progesterone in model group showed a considerable reduction (P < 0.05 ). When compared with the control group,model group rats showed increased ovarian volume and high incidence of subcapsular ovari-an cysts together with decreased number of corpora lutea.The expression of BDNF and p75 mRNAs was significantly higher in model group than that in the control group(P<0.05),but the expression of TrkB mRNA reduced(P <0.05).Conclusion BDNF/TrkB/p75 expressed in ovarian tissues may play a specific role on follicular developmental disorders in letrozole-induced PCOS rats.%目的:研究脑源性神经营养因子(BDNF)及其受体基因在来曲唑诱导的多囊卵巢综合征(PCOS)大鼠模型卵巢组织中的定量表达。
哺乳动物生殖系统的结构与生理机制

哺乳动物生殖系统的结构与生理机制哺乳动物是一种生殖高度发达的动物,其生殖系统结构复杂、功能完备。
这种结构和功能的发展受到物种生存需求、环境因素和遗传基因等多种因素的影响。
本文将从哺乳动物生殖系统的结构和生理机制两方面来探讨其生殖特点和发展趋势。
一、生殖系统的结构哺乳动物生殖系统由生殖器官、性腺和内分泌系统组成,包括雌性和雄性。
雌性生殖器官主要包括输卵管、卵巢、子宫和阴道等;雄性生殖器官主要包括精子产生器官、输精管、前列腺、射精管和阴茎等。
1.雌性生殖系统雌性生殖系统是指分泌雌激素、孕激素和卵子、产生胎儿的女性生殖系统。
雌性生殖器官相对较为复杂,主要包括输卵管、卵巢、子宫和阴道四个部分。
输卵管是连接生殖器官和卵巢的通道,其内膜质地柔软,运输受精卵经过输卵管进入子宫。
卵巢是产生卵子和女性激素的主要器官,一个卵巢内含有许多小卵泡和沟槽。
卵子成熟后,从其中的卵泡释放出来,进入输卵管。
子宫是胎儿孕育和发育所在地,也是孕激素的主要生产场所。
阴道是通向外部的通道,具有排泄、交配和生产的作用。
2.雄性生殖系统雄性生殖系统由睾丸、附睾、精囊、尿道和阴茎等组成。
睾丸是产生精子和男性激素的主要器官,附睾是精子储存和排泄的通道。
精囊是生产精液成分的主要器官。
阴茎是通向外部的生殖器官,具有射精、排泄和交配等功能。
二、生理机制1.雌性生殖机制生殖周期是指雌性生殖系统中的一系列变化过程,包括卵巢的周期性变化、子宫内膜的增生和脱落等。
生殖周期一般为28天左右。
雌性生殖系统的激素分泌周期性发生变化,引起了一系列生理反应,如月经、排卵等。
月经是一种生理现象,是女性卵巢内的一组细胞经过周期性变化后,排出子宫腔,伴随子宫内膜的脱落而产生的周期性经血。
每个月经周期都有一个排卵期,即女性卵子从卵巢中释放出来进入输卵管。
此时,如果受精卵存在,则会在输卵管内受精,形成胚胎。
2.雄性生殖机制雄性生殖机制是指男性生殖系统中离子的吐出和季风的排出。
神经系统与生殖系统的相互作用

神经系统与生殖系统的相互作用神经系统和生殖系统是人体两个重要的系统,它们之间有着密切的相互作用。
神经系统通过神经传递信息,调节和控制生殖系统的功能;而生殖系统则对神经系统产生影响,调节和影响神经功能。
本文将探讨神经系统与生殖系统的相互关系,以及其在人体内的重要作用。
一、神经系统对生殖系统的调节神经系统通过中枢神经系统的传导和外周神经系统的调节,对生殖系统的功能产生调控作用。
中枢神经系统包括大脑和脊髓,它通过下丘脑、垂体和神经系统的传导途径,对生殖系统进行直接和间接的调节。
1. 下丘脑对生殖系统的调控下丘脑是大脑中控制体内内分泌系统的重要部分,它直接对垂体腺体产生调节作用。
下丘脑通过分泌促性腺激素释放激素(GnRH),刺激垂体前叶分泌促性腺激素,进而调节性腺激素的合成和释放。
促性腺激素与性腺中的激素分泌密切相关,它们共同调节和控制生殖系统的发育和功能。
2. 神经系统传导对生殖系统的调控外周神经系统通过神经传导作用,对生殖系统的功能进行调节。
例如,自主神经系统的交感神经和副交感神经分别对生殖系统产生不同的影响。
交感神经兴奋可以促使生殖器官的充血和勃起,而副交感神经的兴奋则可以通过降低交感神经活性来增加生殖系统的血流。
二、生殖系统对神经系统的影响生殖系统对神经系统的影响主要体现在性激素对神经功能的直接和间接调节。
1. 性激素的影响性激素对神经系统有着重要的影响。
雌激素能够影响神经细胞的生长和发育,调节神经递质的合成和释放,影响神经电活动的表现形式。
睾丸激素则通过对神经系统的影响间接调节神经系统的功能。
2. 生殖周期对神经系统的影响女性生殖周期的变化对神经系统的功能也有一定的影响。
例如,在月经周期中,雌激素和孕激素的水平的波动会引起一系列的生理和心理变化,如情绪的波动、注意力和记忆力的变化等。
三、神经系统与生殖系统的相互作用在疾病中的影响神经系统与生殖系统的相互作用在某些疾病的发生和发展中起着重要的作用。
生殖系统中神经突触和神经递质的作用及其分子机制探究

生殖系统中神经突触和神经递质的作用及其分子机制探究人类生殖系统是由男性和女性生殖器官组成的,它们在生殖过程中起到了重要的作用。
而神经系统是我们身体内控制行为和生理功能的重要部分,包括神经元、神经突触和神经递质等。
近年来的研究表明,神经突触和神经递质在调节生殖系统的生理过程中也起着关键的作用。
神经突触是神经元用来传递信号的结构。
在中枢神经系统和周围神经系统中,神经突触将电信号转化为化学信号,使神经元之间能够相互通信。
在生殖系统中,神经突触的存在与生殖细胞的形成、发育、调节和运动都有关系。
一些研究表明,雄性小鼠的睾丸和前列腺中含有丰富的神经元和神经突触,这些神经元和神经突触在生殖行为的发生和维持中发挥着重要作用。
神经递质是神经元用来传递信号的分子,它在中枢神经系统和周围神经系统中起着至关重要的作用。
神经递质可以产生兴奋或抑制行为,并且会在一些神经突触中起到调节作用。
在生殖系统中,神经递质也发挥着重要作用。
生殖系统中含有多种神经递质,如去甲肾上腺素、多巴胺、5-羟色胺、神经肽Y等。
这些神经递质与生殖细胞的发育和调节密切相关。
那么,神经突触和神经递质如何调节生殖系统的生理过程呢?最近的研究表明,生殖系统内的神经元和神经突触可以通过多种途径调节生殖细胞的发育和运动。
其中最常见的途径是通过胆碱能和儿茶酚胺能神经元对生殖器官进行调节。
胆碱能神经元可以直接影响生殖细胞的发育和代谢,而儿茶酚胺能神经元则可以间接地影响生殖细胞的发育和代谢,主要是通过影响生殖系统中的性腺激素水平。
此外,生殖系统底部的神经元和神经突触还可以与中枢神经系统的神经元和神经突触进行交流,也就是说,生殖系统和中枢神经系统之间存在着密切的联系。
而在体内发生上述调节过程时,神经递质则是执行调节作用的分子。
不同类型的神经递质会直接影响性腺激素和GnRH的分泌,进而调节生殖细胞的发育和代谢。
例如,多巴胺和去甲肾上腺素可以影响GnRH和生殖激素的分泌,从而调节生殖细胞的发育;而神经肽Y则可以直接影响生殖细胞的代谢和运动。
昆虫生殖系统中生长因子互作的分子机制分析

昆虫生殖系统中生长因子互作的分子机制分析随着生物学研究的不断深入,越来越多的生物过程的分子机制被揭示出来。
其中,生殖系统的研究一直是一个热门的话题。
在昆虫生殖系统中,生长因子是一个极其复杂的系统,对昆虫生殖过程起着重要的调控作用。
因此,本文将对昆虫生殖系统中生长因子的互作分子机制进行较为详细的分析。
生长因子是什么?生长因子是一类具有生物活性的多肽分子,在昆虫中广泛存在。
这些分子可以促进细胞的增殖和分化,以及调节胚胎发育过程。
同时,它们还对调节昆虫生殖过程起着重要的作用。
昆虫中常见的生长因子包括表皮生长因子(epidermal growth factor, EGF)、肝细胞生长因子(hepatocyte growth factor, HGF)、脑源性神经营养因子(brain-derived neurotrophic factor, BDNF)等。
生殖系统中生长因子的作用在昆虫生殖系统中,生长因子可以发挥着以下几个作用:1. 促进生殖细胞增殖。
生长因子能够刺激生殖细胞的增殖,从而增加其数量。
这一作用在生殖过程中尤其重要,因为生殖细胞数量的增加是昆虫繁殖成功的基础。
2. 调节生殖细胞分化。
一旦生殖细胞增殖到一定限度,就需要分化为雄性或雌性细胞,为后续配对做好准备。
生长因子可以在这一过程中发挥着调节作用,使得雄性或雌性细胞能够适时地分化。
3. 配对成功之后,生长因子仍可以继续作用于昆虫卵的发育过程中,从而保证胚胎的正常发育。
生长因子的互作分子机制在昆虫生殖系统中,生长因子之间的互作关系是非常复杂的。
不同的生长因子之间可以通过多种途径相互作用,从而协同调节生殖过程。
以下是一些常见的机制:1. 线粒体线路。
一些生长因子可以通过激活线粒体来促进细胞增殖和分化。
这些生长因子包括胰岛素样生长因子(insulin-like growth factor, IGF)、神经元生长因子(neurotrophic growth factor, NGF)等。
生长因子和受体的作用机制及其应用

生长因子和受体的作用机制及其应用在人体生长和发育过程中,生长因子和受体起着至关重要的作用。
生长因子是一种蛋白质,能够在细胞分裂和增殖时发挥调节作用;而生长因子受体则是一类膜蛋白,负责与生长因子结合并传递信号,从而调节细胞的增殖和分化。
近年来,生长因子和受体的作用机制以及其应用在医学领域中的诸多进展引起了广泛关注。
一、生长因子和受体的作用机制生长因子和受体之间的相互作用是一个非常复杂的过程。
生长因子分泌到细胞外,与受体结合形成配对,启动受体磷酸化,进而激活酪氨酸或丝氨酸/苏氨酸激酶信号通路,从而促进细胞增殖和分化。
另外,生长因子和受体还可以与胞质酶和细胞凋亡通路产生相互作用,引发凋亡或促进存活。
在人体生长发育过程中,生长因子和受体的作用机制非常重要。
比如,胰岛素样生长因子(IGF)和IGF受体在胚胎发育和儿童生长中起着非常重要的作用,它们能够促进胎儿的正常发育,并在生长期沉积骨骼。
而在成年后,生长因子和受体的调节作用则与肌肉发育、创伤修复等方面相关。
二、生长因子和受体的应用尽管生长因子和受体的作用机制非常复杂,但是随着研究深入,它们在医学领域中的应用也越来越广泛。
下面列举一些目前应用较为广泛的生长因子和受体:1.人类生长激素(HGH)人类生长激素是生长激素家族内的一种,是组织生长的主要荷尔蒙。
HGH治疗可用于身材过矮和免疫系统失调等多种情况,具有促进增高、刺激骨骼和肌肉生长、增强免疫力的作用。
2.表皮生长因子(EGF)表皮生长因子在皮肤细胞中表现出很强的细胞增殖和分化能力,被广泛应用于各种皮肤疾病的治疗中,比如溃疡、烧伤、创伤等,具有促进创口愈合、皮肤再生、修复扩张性瘢痕等作用。
3.肿瘤坏死因子α(TNF-α)受体TNF-α是一种促炎细胞因子,可以通过与其受体结合来诱导细胞凋亡。
目前,一些TNF-α受体拮抗剂被广泛应用于治疗风湿性关节炎、炎症性肠病等自身免疫性疾病。
4.血小板衍生生长因子(PDGF)PDGF是一种促进细胞增殖和修复骨骼和软组织的生长因子,具有刺激组织生长、促进伤口修复等作用。
神经内分泌因子调控鱼类生殖和生长的相互作用

神经内分泌因子调控鱼类生殖和生长的相互作用林浩然摘要:脊椎动物的生长与生殖活动有着密切的联系并相互作用。
许多调节生长和代谢活动的内分泌因子对青春期或者性腺的发育产生影响。
同样,调节生殖活动的许多激素亦同时对生长和代谢产生影响。
近年来,我们和其他学者对鱼类生长和生殖的神经内分泌调节的相互作用进行了研究,主要的进展是:①在促进性腺的激素影响生长方面,发现促性腺激素释放激素(GnRH)和多巴胺都能和脑垂体生长激素细胞的特异性受体结合而刺激生长激素释放,并能提高生长速率;而性类固醇激素能调节脑垂体生长激素的分泌活动。
②在生长激素对性腺的作用方面,发现生长激素能促进鱼类性腺发育和成熟;在鱼类性腺有直接结合生长激素以及传递生长激素作用的胰岛素样生长因子(IGF)的作用位点,对性腺产生作用。
总的看来,鱼类生殖与生长的神经内分泌调节有着密切联系,这和高等哺乳类很相似。
但是,在鱼类研究取得的一些新发现是在哺乳类和其他脊椎动物研究中还没有阐明的,因而值得进一步深入研究。
关键词:鱼类;生长激素;促性腺激素;生长;生殖;神经内分泌调节中图分类号:Q453,Q959.4 文献标识码:A 文章编号:0254-5853(2000)01-012-05THE INTERACTIONS OF NEUROENDOCRINE REGULATIONON REPRODUCTION AND GROWTH IN FISHLIN Hao-ran(Institute of Aquatic Economic Animals,Zhongshan University,Guangzhou 510275 ls32@)Abstract:Interaction between growth and reproduction occurs in many vetebrates.The endocrine factors which regulate growth and metabolism are also affected on puberty and gonadaldevelopment.Similarly,the hormones which effect on reproduction are also influ-enced on growth and metabolism.Recent studies onfish revealed the neuroendocrine interactions between the gonadotropic axis and the somatotropic axis.For example,gonadotropin-releasing hormone (GnRH) and dopamine could stimulate the release of growth hormone,sex steroid could modulate growth hormone secretion;also,growth hormone could accelerate gonadal development andmaturation.Some new findings from studies on fish have not yet been obtained in mammals and other vertebrates are remains to be further investigated.Key words:Fish;Growth hormone;Gonadotropin;Growth;Reproduction;Neuroendocrine regulation许多脊椎动物的生长与生殖活动有着密切的联系并相互作用,在鱼类生活史的一些阶段尤为明显。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
蛋 白, 于肿瘤坏死受 体家族 J 属 。在 高亲和力受体的形成的过程
中 , N F与 t A 的 结 合 外 , 需 要 p5的 作 用 , 者 与 N F 的 除 G r k 还 7 后 G 结 合 可 提 高 细 胞 表 面 N F的 有 效 浓 度 , 两 种 受 体 均 表 达 的 细 G 在 胞 表 面 可 使 N F的亲 合 力 提 高 2 。 另 外 ,7 G 5倍 p5的 存 在 可 以 促 进 t A 的 白磷 酸 化 , 而 导 致 t A 与 配 基 结 合 能 力 和 信 号 转 r k 。t| 。
神 经 生 长 因 子 及 其 受 体 在 雄 性 哺 乳 动 物 生 系 中 的 作 用 殖 统
李彝 马晓婷 翁冀 翁强 北京林 业大 学生物科 学与技 术学院 ( 北京 10 8 ) 0 0 3
【 摘要 】 神经生长 因子 ( ev rwhf t , G ) 于神 经 营养 因子 家族 , 家族主要 对神 经元的 生长 , n r g t c rN F 属 e o ao 该 发育和 分化起 到一 定作
导 功 能 的 改 变 J 。
碑 J 。近年来 的研 究表明 : G N F不 仅作 用于神 经系统 , 生殖 系 对
统 , 疫 系统 , 环 系 统 等 也 存 在 重 要 调 节 作 用 。 。 在 雄 性 生 免 循 、 。 殖 系 统 中广 泛 存 在 N F及 其 受 体 , 们 通 过 自分 泌 和 旁 分 泌 并 G 它 且协 同 生 殖 激 素 和 其 它 细 胞 因子 , 精 子 形 成 和 成 熟 以及 雄 性 生 对
中外 医 学 研 究
一
21 0 0年 8 月 第 8卷
第1 8期
C I E EA D F R I N ME IA E E R H H N S N O EG D C LR S A C
曩 。 一 | 曩 再 曩 曩露 一 I 誊 ≯。 澎
≯一 曩≥≯ 。 ≯
【 b t c】 N rego t fc r( G )blns oa a i f e t rtn q i df esr vl ma tnne addvl m n o A sr t a ev wh at N F eog ml o r a dpoe s eur r h u ia, i eac , ee p et f r o t f y le i r e ot v n n o
熟和 生 殖 系统 的 生 长发 育有 重要 调 节 作 用 。
【 关键词 】 神 经生长 因子 ; 受体 ; 雄性 生殖 系统
Fun i n fne v owt f c or an is r c p or n e o cto s o r e gr h a t d t e e t s i r pr duc i s s e f m ae m a m as tve y t m o l m l.L/Be MA Xi — t n, ao i ng WE NG J , i
用。N F通过与其 受体 结合 来发挥 自分泌与旁分泌的作用 , G 并且其作用不仅 限于神经 系统 , 对其 它系统也 有重要调 节作用。在雄性 哺 乳动物 生殖系统 中, G N F及其受体在精 巢、 附睾、 列腺 、 囊腺等组 织都有 不 同程度 的表 达 , 示 N F及 其 受体 对精 子形成 、 前 精 提 G 精子成
,
W NGQa g C lg il i l c ne n enl yB n oet nvrt,ei 00 3 C /a E in. ol e fBo gc i c dTcoo e gF r r U i sy B in 10 8 , h . e o o aSe a g sy ei jg n
t en .
【 yw r ] N F R cpos Ma erd c v yt Ke o d G ; ee t ; r l rpo ut ess m e i e
发 育 中的 神 经 元 的存 活 依 赖 于 称 为 生 长 因 子 的 特 殊 蛋 白质 的供 给 , 而其 中第 一个 被鉴 定 出 来 的 生 长 因 子 就 是 神 经 生 长 因 子 (e ego hf t , G ) 它 也 是 目前 为 止 研 究 最 清 楚 的 一 个 神 nr rw a o N F , v t cr 经 营 养 因 子 … 。 B c e 在 14 uk r 9 8年 的实 验 中首 先 提 示 存 在 着 神 经 营养 因 子 , 后 在 15 随 9 9年 L v —M na ii 蛇 毒 中 首 先 发 现 了 ei otl n 在 c 神 经 生 长 因 子 , 发 现 被 认 为 是 神 经 科 学 发 展 史 上 的 重 要 里 程 该 的 受体 ; p5属 于 低 亲 和力 受 体 , 分 子 量 为 7 K 的 I 跨 膜 而 7 是 5D 型
adsm n ei e f mm l T eersl ugs dta N F adi eet sp ya m ot teua v o aerpout ess n e ia vs lso ma as h s eut sget t G n srcpo I ni pr n glt e l i m l erdci y— l c . s e h t r a a r i r en v