分子生物学第三章
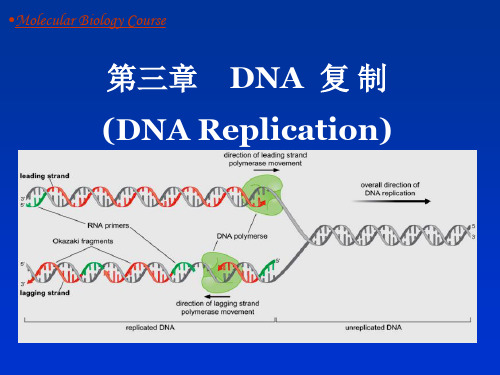
分子生物学 第3章 DNA复制
DNA helicase (DNA解旋酶)
利用ATP供能,解开DNA双链, 可随复制叉 的伸展向前移动
大肠杆菌中解旋酶的种类
种 类
DnaA DnaB DnaC
功 能
辨认起始点,并结合到复制起始部位 解开DNA双链 运送和协同DnaB
single-stranded binding protein (SSB, 单链结合蛋白)
是一类调节DNA分子的超螺旋水平,可改变DNA拓扑性 质的酶。对DNA分子的作用是既能水解、又能连接磷酸 二酯键。 • 拓扑异构酶 I: 切开DNA双链中的一股,使DNA在解链旋 转中不打结,DNA变为松弛状态再封闭切口。 同转录有 关 • 拓扑异构酶 II: 能切断DNA双链,使螺旋松弛。在ATP参 与下,松弛的DNA进入负超螺旋,再连接断端。同复制
3´→5´外切酶活性: 切除错配的核苷酸
5'
3' C T T C A G G A G A A G T C C G G C G 5'
3'
DNA ligase
连接DNA链3-OH末端和相邻DNA链5-P末端,形成磷 反应需要ATP。
酸二酯键,从而把两段相邻的DNA链连接成完整的链。
二、 DNA复制的过程
E. Coli DNA在15N-标记的营养液中生
长多代,使DNA双链充分标记
将15N-标记
细胞在
14N中
细胞在
14N中复
细胞在
14N中复
的E.Coli 加入14N 培 养液中
万有引力
复制1 次
制第2次
制第3次
单林娜 制作
11
DNA半保留复制的生物学意义:
DNA的半保留复制表明DNA在代谢上的稳定性,
《分子生物学教学》第三章可移动的遗传因子

探讨可移动遗传因子在基因工程、基因治疗、生物 育种等领域的应用前景,以及相关的伦理和安全问 题。
02
可移动遗传因子的类型和特性
转座子的类型和特性
80%
插入序列(IS)
是细菌中最简单的转座子,能够 编码自身转座所需的酶,并能在 基因组中随机插入。
100%
转座噬菌体(Tn)
是一种复杂的转座子,带有与噬 菌体相关的基因,能够在细菌之 间水平转移。
分子生物学教学第三章可移动 的遗传因子
目
CONTENCT
录
• 引言 • 可移动遗传因子的类型和特性 • 可移动遗传因子的机制 • 可移动遗传因子的生物学意义 • 研究方法和实验技术 • 实际应用和未来展望
01
引言
目的和背景
阐述可移动遗传因子的概念
本章旨在介绍可移动遗传因子的概念,包括其定义、分类、功能 以及在生物学领域的重要性。
转录调控
可移动遗传因子如转座子和逆转 录病毒可通过插入或删除基因序 列,影响转录因子的结合和基因 表达的调控。
表观遗传学调控
某些可移动遗传因子能够影响染 色质结构和组蛋白修饰,从而参 与表观遗传学调控,改变基因的 表达模式。
在基因组进化和多样性中的作用
基因重组
可移动遗传因子通过介导基因重组事 件,促进基因组的重排和多样性产生 。
对核酸和蛋白质序列进行比对,找出同源序列和保守区域,并进行功能
注释。
02
基因表达与调控分析
利用高通量测序技术分析基因表达谱,研究基因表达的时空特异性和调
控机制。
03
生物信息学数据库与工具
利用生物信息学数据库(如GenBank、UniProt等)和在线分析工具
分子生物学第三章蛋白质大分子结构与功能

49
4.蛋白质四级构造
一个蛋白质由几条多肽链组成1个活性单 位。亚基的相互关系,空间排布,亚基 间通过非共价键聚合成的特定构象。单 一亚基无活性,只有聚合后才有生物活 性。如血红蛋白。
50
51
蛋白质预测网站
Compute pI/WM ://expasy.hcuge.ch Predictprotein :// embl-heidelberg.de/predictprotein/ SOPMA :// ibcp.fr/predict.html Unpredict ://www /
43
44
45
46
构造域
• 多肽链在超二级构造根底上进一步卷 曲折叠成严密的近似球状的构造。对 较小蛋白质分子,构造域往往就是三 级构造,即这些蛋白质是单构造域。
• 许多蛋白质是多构造域。
47
构造域
48
3.蛋白质三级构造
• 多肽链的某些区域氨基酸形成二级构造: α螺旋、β-折叠、 β-转角、无规那么卷曲等构 象单元, 然后相邻二级构造集装成超二级构 造, 进而折叠绕曲成构造域, 由2个或2个以上 的构造域组装成三级构造。
30
二 硫 键
31
一级构造确定的原那么
• 测定蛋白质中氨基酸组成 • 蛋白质N端和C端的测定 • 2种以上方法水解蛋白质,得到一系列
肽段 • 别离提纯所得肽,测其序列 • 从有重叠构造的肽序列中推断蛋白质
的全部氨基酸的排列顺序
32
〔二〕蛋白质的空间构造
• 肽平面:肽腱中的4个原子以及相邻的 2个α-碳原子处在同一平面,使肽链具 有一定的稳定性
20
氨基酸与异硫氰酸苯酯的反响
• AA的氨基可与异硫氰酸苯酯(PITC)反 响,生成苯氨基硫甲酰氨基酸(PTC-AA)。 所得PTC-AA经乙酸乙酯抽提→层析鉴 定→ 确定N端氨基酸的种类。 “多肽 顺序自动分析仪〞据此原理。
分子生物学基础第三章遗传与变异 第三节基因、基因组与基因突变

第三节 基因、基因组与基因突变
2.基因的命名 (1)用三个小写英文斜体字母表示基因的名称 (2)在三个小写英文斜体字母后面加上一个大写英文 斜体字母表示其不同的基因座,全部用正体时表示其相应 的蛋白产物和表型。 (3)对于质粒和其他染色体外成分,如果是自然产生 的质粒,用三个英文正体字母表示,第一个字母大写;但 如果是重组质粒,则在两个大写字母之前加一个p,大写 字母表示构建该质粒的研究者或单位。 (4)对果蝇基因命名的例子最繁多,特别是在发育生 物学中。对突变表型的表示用1-4个字母代表。
第三节 基因、基因组与基因突变
图3-23 化学修饰剂改变碱基的配对性质
第三节 基因、基因组与基因突变
3.诱变剂和致癌剂的检测 许多化合物需在体内经过代谢活化才有诱变作用,在 测试时可将待测物与肝提取物一起保温,使其转化,这样 可使潜在的诱变剂也能被检测出来。大肠杆菌的SOS反应 可以使处于溶源状态的λ噬菌体激活,从而裂解宿主细胞 产生噬菌斑。通常引起细菌SOS反应的化合物对高等动物 都是致癌的。Devoret根据此原理,利用溶源菌被诱导产
第三节 基因、基因组与基因突变
三、基因突变
1.基因突变的类型
基因突变有以下多种类型:碱基对置换指DNA错配碱 基在复制后被固定下来,由原来的一个碱基对被另一个碱 基对所取代,又称为点突变。碱基对置换有两种类型:即 转换是在两种嘧啶或两种嘌呤之间的互换;颠换发生在嘧 啶与嘌呤或嘌呤与嘧啶之间的互换。碱基替换通常仅发生 在一个碱基上,偶尔也有几个碱基同时被替换。转换发生 的频率一般比颠换高1倍左右。插入突变指在基因的序列 中插入了一个碱基或一段外来DNA导致的突变。例如,大 肠杆菌的噬菌体Mu-1、插入序列(IS)或转座子都可能诱 发插入突变。插入突变有两种方式:①拷贝或复制移动, 指一个位点上的序列被复制后插入到另一位点。②非拷贝 移换,DNA序列从一个位点直接移动到另一位点。
分子生物学第三章RNA转录

分⼦⽣物学第三章RNA转录第三章 RNA 转录(RNA transcription)3.1. Basic concept3.2. Trancription survey3.3. Promoter in Eukaryotes and Prokaryotes3.4. Transcription Termination3.5. Pre-RNA processing in Eukaryotes3.1. 基本概念(P64) Basic concept●基因表达的第⼀步●以D. S. DNA 中的⼀条单链作为转录的模板某⼀基因只以⼀条单链DNA 为模板进⾏转录(不对称转录)●在依赖DNA 的RNA 聚合酶的作⽤下●按A U ,C G 配对的原则,合成RNA 分⼦●模板单链 DNA 的极性⽅向为3’ → 5’, ⽽⾮模板单链DNA 的极性⽅向与RNA 链相同,均为5’ → 3’.● RNA 的转录包括promotion, elongation, termination 三个阶段●从启动⼦(promoter )到终⽌⼦(terminator )的DNA序列称为转录单位(transcriptional unit )●原核⽣物中的转录单位多为 polycistron in operon真核⽣物中的转录单位多为monocistron, No operon●转录原点记为+1,其上游记为负值,下游记为正值● RNA 的主要种类及功能:mRNA ——携带编码多肽的遗传信息tRNA ——将核苷酸信息转化为aa 信息转运aa 进⼊核糖体rRNA ——参与多肽合成3.2.RNA 转录概况3.2.1转录的基本过程1. 模板识别:RNApol 与启动⼦相互识别并结合的过程(形成封闭的⼆元复合物)启动⼦(promoter ):DNA 分⼦上结合RNApol 并形成转录起始复合物的区域,通常也包括促进这⼀过程的调节蛋⽩结合位点rich A/T ,易发⽣DNA 呼吸现象形成单链区2转录起始:启动⼦区解链,转录起始(封闭的⼆元复合物开放的⼆元复合物三元复合物)通常在这⼀过程中RNApol 移动较慢,且易发⽣脱落——流产式起始 ——决定启动⼦的强弱3延伸:延伸过程中的延宕现象(Eukaryotes ):Euk genome G/C 分布不均匀σ脱离全酶(Pro )/RNApol 脱离转录起始复合物(Euk )4终⽌:在终⽌⼦(terminator )处停⽌转录3.2.2 RNApolymerase1 RNA polymerase in Prokaryotes (以E.coli 为例)1)构成核⼼酶(core enzyme):2αββ’DNA3’----TACTCAT----5’ RNA 5’----AUGAGUA----3’5’---ATGAGTA----3’ Non-template (sense strand)template (antisense strand)全酶(holoenzyme)2αββ’σα:核⼼酶组建因⼦/ 启动⼦识别β:RNA合成的活性中⼼β’:与β共同构成活性中⼼σ:识别启动⼦,增加酶与DNA的亲和⼒σ因⼦可减少RNApol与⾮启动⼦DNA序列的亲和⼒,⽽增加RNApol与启动⼦的亲和⼒,⼀旦转录起始,σ因⼦将脱离RNApol再次引导新的RNApol进⾏转录ρ:参与转录终⽌2)Rifamycin(利福霉素)及Streptolydigin(利链菌素)对Pro转录的影响Rif可结合β,阻⽌NTP的进⼊I位点(Initiation site )(⼀旦形成三元复合物Rif不再起抑制作⽤);利链菌素结合β的延伸位点(Elongation site),抑制延伸。
分子生物学第三章

Density gradient of sucrose Measure H3-T
H3-T
pulse-labeling
pulse-chase 120’’
60’’
7’’
2’’
(Prok. 400Nt/sec)
15’’
10S (1kb)
40S
DNA replication in Okazaki fragment 1kb
直接证据?
pulse-labeling in dT-H3 ?
Lig (ts)?
(Source:Molecular Biology(2002),Robert F.Weaver,Page643)
DNA semi-discontinuous replication
leading strand , lagging strand 均有 dUMP 的掺入 Okazaki 片段在某种意义上为 dUMP 片段
在富含AT的区域内尤为明显
• replication origin 两侧基因的转导频率高
a c a o o b b d
复制的不同步性 f f h
断裂的随机性
replication origin
g e c a
e
c
a
o o
b b
d d
Most rDNA are located near the origin of replication
( Cairns model , θ form, theda form)
Eukaryote(500-5000bp/min)
Rifampin
有M13 RF
M13
有M13 RF
Conclusion
• M13 RF的形成需要 RNA polymerase发动合成一 段 RNA分子作为引物 • RF启动后,RNA引物已经形成, Rifampin 的抑制无效
分子生物学第三章基因和基因组

第三章基因和基因组第一节基因组的大小与C值矛盾一、相关概念☆基因(gene)是合成一种功能蛋白或RNA分子所必须的全部DNA序列.一个典型的真核基因包括①编码序列—外显子(exon)②插入外显子之间的非编码序列—内合子(intron)③5‘-端和3’-端非翻译区(UTR)④调控序列(可位于上述三种序列中)基因平均由1000个碱基对组成,一个DNA分子可能包含几个或几千个基因。
☆基因组(genome):一特定生物体的整套(单倍体)遗传物质的总和,即生物体维持配子或配子体正常功能的全套染色体所含的全部基因(DNA)。
基因组的大小用全部DNA的碱基对总数表示。
比如人基因组的全长为大约3×109对碱基,编码3-4万个蛋白分子。
细菌或噬菌体、病毒---单个染色体中所含的全部基因(DNA)。
人类基因组计划(human genome project HGP)基因组学(genomics)结构基因组学(structural genomics)功能基因组学(functional genomics)☆C值(C-value):在真核生物中,每种生物的单倍体基因组的DNA总量总是恒定的,称为C-值低等真核生物中与形态学复杂程度相关,但高等真核生物中变化很大所谓C值(C value)即单倍体基因组的DNA总量。
它是每一种活生物的一个性质。
C 值大小有着巨大差异。
小到象支原体那样的不足106bp,大到一些植物及两栖类的1011bp。
进化中不同门的C值范围。
随着复杂度的增加,基因组大小的最小值是增加的。
但是随着高等真核生物DNA绝对量的增长,有些门的基因组大小出现了很大的变化。
每门的一种生物DNA的最小量要使原核生物比低等真核生物更复杂,增加基因组大小是必要的。
盐沼核菌(pyrenomas salina)是现已证明的含有最小基因组的真核生物,它有6.6×105bp,(然而这些生物可能并不是真正的真核生物,但可能是进化的中间阶段,代表了细胞核与叶绿体的原始存在形式。
大学分子生物学经典课件第三章 生物信息传递上从DNA到RNA.ppt
2019/11/23
34
转录单元 (transcription unit)
起点(startpoint) 上游(upstream) 下游(downstream)
2019/11/23
35
启动子区的基本结构
细菌启动子特征:
1、起点通常是一个嘌呤; 2、起点上游10 bp处,有一约6 bp的保守区域,称为10 区(也叫Pribnow Box),共有序列为:
21
2019/11/23
细胞中不同状态RNA聚合 酶的数量
每个E.coli细胞中含约 7000个RNA 聚合酶;
核心酶主要以松散的闭 合复合体为主;
足量的σ因子使三分之 一的聚合酶以全酶形式 存在,主要是在非特异 位点的松散复合体和启 动子处的紧密(开放) 复合体;
约2500个核心酶正在进
行转录。
2019/11/23
33
转录单元(transcription unit):是一段 从启动子开始到终止子结束的DNA序列,RNA 聚合酶从转录起始位点开始沿着模板前进,直 到终止子为止,转录出一条RNA链。
转录起始位点:是指与新生RNA链第一个 核苷酸相对应DNA链上的碱基,研究证实通常 为一个嘌呤。
2019/11/23
6
随机扩散
模板识别:RNA聚合 酶与启动子DNA双链 相互作用并与之相结合 的过程。
随机行走
定向取代
2019/11/23
7
启动子(promoter):是基因转录起始所必需的一段 DNA序列,是基因表达调控的上游顺式作用元件之一。
在原核生物中, σ 因子辨认-35区,全酶与该区结 合形成疏松复合物,继而全酶向-10区及起始位点移动, 到起始位点后全酶与DNA结合紧密。
分子生物学3生物信息的传递(上)——从DNA到RNA

分子生物学3生物信息的传递(上)——从DNA到RNA第三章生物信息的传递(上)——从DNA到RNA重点:1.启动子与转录起始2. 原核生物和真核生物mRNA的特征比较3. 内含子的剪接、编辑及化学修饰难点:1.启动子与转录起始2. 终止和抗终止3. 内含子的剪接、编辑及化学修饰第四节启动子与转录起始大肠杆菌RNA聚合酶与启动子的相互作用主要包括启动子区的识别、酶与启动子的结合及因子的结合与解离等。
1. 原核启动子的基本结构(1)启动子:是一段位于结构基因5 ′端上游区的DNA序列,能活化RNA聚合酶,使之与模板DNA准确的相结合并具有转录起始的特异性。
基因的特异性转录取决于酶与启动子能否有效地形成二元复合物,所以,RNA聚合酶如何有效地找到启动子并与之结合是转录起始过程中首先要解决的问题。
我们知道,转录的起始是基因表达的关键阶段,而这一阶段的重要问题是RNA聚合酶与启动子的相互作用。
启动子的结构影响了它与RNA聚合酶的亲和力,从而影响了基因表达水平。
(2)转录单元:是一段从启动子开始至终止子结束的DNA序列。
RNA聚合酶从转录起点开始沿着模板前进,直到终止子为止,转录出一条RNA链。
在细菌中,一个转录单元可以是一个基因,也可以是几个基因(3)转录起点:是指与新生RNA链地一个核苷酸相对应DNA链上的碱基。
常常把起点前面,即5′末端的序列称为上游,而把其后面即3′末端的序列称为下游。
在描述碱基的位置时,一般用数字表示,起点为+1,下游方向依次为+2、+3等,上游方向依次为-1、-2、-3等。
启动子区是RNA聚合酶的结合区,其结构直接影响到转录的效率。
那么,启动子区有什么结构特点呢?(4)绝大部分原核启动子都存在-10区和-35区-10区:在-6~-13bp之间,共同序列为TATAAT,又称pribnow 框,酶在此处与DNA结合成稳定的复合物,在转录方向上解开双链形成开放型起始结构。
-35区:共同序列为TTGACA,是RNA聚合酶起始识别区,这一识别过程与σ因子有关。
分子生物学课件 第3章 基因与基因组

实际应用中“基因组”这个词既可以特指储存在细胞核中 的整套DNA(即核基因组),也可以指储存在细胞器中的 整套DNA(即线粒体基因组或叶绿体基因组),还可以指 一些非染色体的遗传元件,如病毒基因组、质粒基因组和 转座元件等。
不同基因家族各成员之间的序列 相似度也不同:
序列高度相似:经典的基因家族,如rRNA基因家族和组蛋 白基因家族。 保守性较低,但是编码产物具有大段的高度保守的氨基酸 序列。
序列保守性很低,编码产物之间也只有很短的保守氨基酸 序列,但通常由于具有保守的结构和功能区域,因而编码产 物具有相似的功能。
基因家族的成员在染色体上 的分布形式不同:
成簇存在的基因家族(clustered gene family)或称基因簇 (gene cluster),如人类类α链基因簇和类β链基因簇。 散布的基因家族(interspersed gene family),如肌动蛋白 基因家族和微管蛋白基因家族。
基因间隔区较短且内含子较少,基因排列紧密。
3.2.7 沉默基因
沉默基因( Silent Gene)也叫隐蔽基因(Cryptic gene), 是处于不表达状态的基因。它可能是假基因,也可能是被关闭的 基因。这些基因以隐性的方式埋藏在染色体中,但遇到特殊因子 的刺激,有可能解除关闭变成显性基因。
3.2.8 RNA基因
tRNA、rRNA; 核仁小分子RNA(small nucleolar RNA, snoRNA) 微小分子RNA(microRNA, miRNA); 小分子干扰RNA(small interfering RNA, siRNA); 核内小分子RNA(small nuclear RNA, snRNA);
分子生物学第三章 基因与基因组的结构与功能

3.1 基因的概念
基因(gene):是原核、真核生物以及病毒的
DNA和RNA分子中具有遗传效应的核苷酸序
列,是遗传的基本单位和突变体及控制性状
的功能单位。
结构基因
包括:
(编码蛋白质、tRNA、rRNA)
调控基因
(编码调控蛋白)
• 基因通过复制、转录和翻译合成蛋白质以及
• 有关基因的命名方法现在并没有严格的统一。
随着分子生物学的飞速发展。许许多多的基 因组都已大规模被测序,更多的基因也不断 的被鉴定。因而十分需要一个统一的命名方 法。
• 为便于学习理解,根据现代分子生物学中目
前使用最多的方法暂归纳如下:
• 1)用三个小写英文斜体字母表示基因的名
称,例如涉及乳糖(lactose)代谢相关的酶 基因lac;涉及亮氨酸(Leucine)代谢相关 的酶基因leu。
7)植物基因的命名
目前还没有适用于植物的惯用命名法 多数用1~3个小写英文斜体字母表示。 如:hsp90,热激蛋白基因
Oryza sativa,Arabidopsis thaliana
OsAthsp90;
Athsp90;Athsp90.3; Athsp90.6
• 8)脊椎动物基因的命名 • 用描述基因功能的1~4个小写字母和数字
• 2)在三个小写英文斜体字母后面加上一个斜体大写
字母表示其不同的基因座。全部用正体时表示蛋白 产物和表型
• 例如,对于大肠杆菌和其他细菌,用三个小写字母
表示一个操纵子,接着的大写字母表示不同基因座,
lac 操纵子的基因座:lacZ,lacY,lacA;其表达
产物蛋白质则是lacZ,lacY,lacA。
分子生物学-03复制

Biotechnology Institute Hu Dongwei hudw@
第三章 DNA的复制
一、半保留复制
Semi-conservation replication
以每条链为模板,按碱基互补配对原则由DNA 聚合酶催化合成新的互补链。
DNA polymerases in human and SV40
6 DNA连接酶 (DNA lygase) A.原核生物
催化DNA链的5'-PO4与另一 DNA链的3'-OH生成磷酸二酯键。 (1) 大肠杆菌的DNA连接酶
75kD,对胰蛋白酶敏感,每个 细胞中约有300个分子。在DNA复 制、修复和重组中起着重要的作用。
2 单链DNA结合蛋白(SSBP)
E. coli的SSBP为四聚体, 可结合32 bp。 SSBP使单链DNA呈伸展 状态,有利于单链DNA作 为模板。 SSBP防止单链DNA重新 配对或被降解。
3 DNA拓扑异构酶 (Topisomerase)
催化DNA不同超螺旋状 态之间的转变。 A. 拓扑异构酶I :双链解旋 切断形成“酶-DNA“共 价中间物 DNA连接。不 需辅助因子。 B. DNA旋转酶(DNA gyrase): 拓扑异构酶II,引入DNA分 子负超螺旋 。需要ATP。
在DNA合成延伸过程中主要是DNA聚合酶III的作用。当 冈崎片段形成后,DNA聚合酶I通过其5'→3'外切酶活性切 除冈崎片段上的RNA引物,同时,利用后一个冈崎片段作 为引物由5'→3'合成DNA。最后两个冈崎片段由DNA连接 酶将其接起来,形成完整的DNA后续链。
DNA复制的终止
DNA上也存在着复制终止位点,DNA复制将在复制终 止位点处终止,并不一定等全部DNA合成完毕。当RNA 引物被切除后,中间所遗留的间隙由DNA聚合I所填充。 但目前对复制终止位点的结构和功能了解甚少。线性DNA
分子生物学笔记完全版第三、四章

分子生物学笔记完全版第三、四章--------------------------------------------------------------------------------作者: tonyloveyou 收录日期: 2006-07-13 发布日期: 2006-07-13第三章基因表达的调控基因表达:DNA→mRNA→蛋白质的遗传信息传递过程基因表达的调控第一节基因的活化基因的“开关”-染色质的活化一、活性染色质的结构间期核染色质:异染色质(heterochromatin),高度压缩(不转录);常染色质(euchromatin),较为松散,常染色质中约10%为活性染色质(更开放疏松)。
活性染色质→←非活性染色质二、活性染色质的结构特点(一)DNaseI敏感性转录活性(或有潜在转录活性)的染色质对DNase I更敏感.DNase I超敏感位点(DNase I HyperSensitive Sites,DHSS)(二)组蛋白H3的CyS110上巯基暴露,三、活性染色质结构的形成(一)、核小体位相(Phased positioning)1.核小体的旋转定位(rotational positioning)指核小体核心与DNA双螺旋在空间结构中的相互关系,主要包括DNA双螺旋的大沟是面向还是背向核心结构.‘2.核小体的平移定位(translational positioning)指核小体与特定DNA序列的结合位置和方式,特别是转录活性相关的DNA调控元件(启动子、增强子等)序列与核小体的相互位置关系。
(二)、组蛋白修饰1.H1组蛋白磷酸化促进染色体包装,影响转录活性,2.核心组蛋白修饰乙酰化:常发生在组蛋白的Lys,一般活性染色质是高度乙酰化的。
(三)HMG蛋白结合HMG(high mobility group)蛋白—高迁移率蛋白, 如HMG14/HMG17.与核小体核心颗粒结合,有利转录。
分子生物学第三章核酸的结构与功能

分子生物学第三章核酸的结构与功能核酸是生物体内重要的生物大分子,在维持遗传信息传递、调控基因表达和蛋白质合成等生物学过程中起着重要的作用。
本文将介绍核酸的结构和功能,包括DNA和RNA的结构、功能以及细胞中的DNA重复序列和嵌合DNA的现象。
核酸是由核苷酸单元组成的大分子。
核苷酸由一糖分子(核糖或脱氧核糖),一个含有一键磷酸基的磷酸基团和一个含有碱基的碱基组成。
DNA的碱基包括腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)和胞嘧啶(C),而RNA的碱基包括腺嘌呤(A)、鸟嘌呤(G)、尿嘧啶(U)和胞嘧啶(C)。
DNA(去氧核糖核酸)是双链结构,由两条互补的单链以互补碱基配对(A和T,G和C)的方式相互连接而成。
这种双链结构被称为双螺旋结构,其中的两个链通过氢键相互链接。
DNA在细胞中起着存储遗传信息的作用,是遗传物质的主要组成部分。
DNA通过转录过程产生RNA分子,进而通过翻译过程合成蛋白质。
RNA(核糖核酸)有多种类型,包括信使RNA(mRNA)、核糖体RNA (rRNA)和转运RNA(tRNA)。
mRNA是由DNA转录得到的,其中的密码子序列编码蛋白质的氨基酸序列。
rRNA是核糖体的组成部分,参与蛋白质合成的过程。
tRNA将氨基酸带入核糖体与mRNA相匹配的密码子上,完成蛋白质合成的过程。
在细胞中,存在许多DNA重复序列。
其中,基因是密集编码蛋白质的DNA序列,它们在核酸的遗传信息传递和基因表达中起着重要作用。
除了基因,还存在大量的非编码DNA序列,如内含子和调控序列,它们对基因表达的调控起着重要作用。
此外,DNA重复序列还包括微卫星、线粒体DNA和细胞质DNA等。
总之,核酸是生物大分子,在维持遗传信息传递和调控基因表达等生物学过程中起着重要作用。
DNA和RNA具有不同的结构和功能,包括存储遗传信息、编码蛋白质序列、调控基因表达和蛋白质合成等。
此外,细胞中存在多种形式的DNA重复序列和嵌合DNA现象,对维持细胞功能和遗传多样性具有重要作用。
分子生物学第三章DNA的复制知识总结

分子生物学第三章DNA的复制知识总结.doc分子生物学第三章:DNA的复制知识总结引言DNA复制是生物体细胞分裂过程中的一个关键步骤,确保遗传信息的准确传递给下一代细胞。
在分子生物学的第三章中,我们深入探讨了DNA 复制的机制、参与的酶类、复制过程以及复制后的修复机制。
本文将对这些内容进行详细的总结。
第一节:DNA复制的基本概念1.1 DNA复制的定义DNA复制是指在细胞分裂前,DNA分子精确复制自身,生成两份相同的DNA分子的过程。
1.2 DNA复制的重要性遗传信息的传递:确保子代细胞获得与亲代相同的遗传信息。
细胞增殖:为细胞分裂提供必要的遗传物质。
1.3 DNA复制的特点半保留复制:每个新生成的DNA分子都包含一个原始链和一个新合成的链。
高度保守:在不同的生物体中,DNA复制的基本机制相似。
第二节:DNA复制的酶类和蛋白质2.1 DNA聚合酶功能:在DNA复制中添加新的核苷酸,形成新的DNA链。
类型:包括DNA聚合酶I、II、III等。
2.2 解旋酶功能:解开DNA双链,为复制提供模板。
2.3 SSB蛋白功能:保护解开的单链DNA,防止其结构被破坏。
2.4 引物酶功能:合成RNA引物,为DNA聚合酶提供起始点。
第三节:DNA复制的过程3.1 起始阶段解旋酶在复制起点处解开DNA双链。
引物酶合成RNA引物。
3.2 延伸阶段DNA聚合酶III沿着模板链添加核苷酸,合成新的DNA链。
两条新链分别在前导链和滞后链上合成。
3.3 终止阶段当复制达到DNA末端时,复制过程终止。
RNA引物被移除,由DNA聚合酶I填补。
第四节:DNA复制的调控4.1 复制的起始点特定的DNA序列作为复制的起始点。
4.2 复制的调控蛋白多种蛋白质参与调控复制过程,确保复制的准确性和效率。
4.3 复制的周期性细胞周期中,DNA复制发生在特定的时期。
第五节:DNA复制的修复机制5.1 错配修复修复复制过程中发生的碱基错配。
5.2 核苷酸切除修复移除并替换受损的核苷酸。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
转座子的分类(续)
• 复制型转座子
在复制过程中,整个转座子被复制,所移动的是 原转座子的拷贝,转座酶(transposase)和解离 酶(resolvase)分别作用于原始转座子和复制 转座子,如TnA类转座子。
• 非复制型转座子
在复制过程中,原始转座子作为一个可移动 的实体直接被移位,如IS序列、Mu、Tn5等。
三、从mRNA到蛋白质
翻译的基本过程
将mRNA链上的核苷酸从一个特定的起始位点开始,按每3个 核苷酸代表一个氨基酸的原则,依次合成一条多肽链的过程。
翻译的起始
核糖体与mRNA 结合并与氨基酸tRNA 生 成 起 始 复合物
肽链的延伸
核 糖 体 沿 5’ 端 向 3’ 端 移 动 , 多肽从N端向C 端合成
开放复合物:在RNA转录的模板识别阶段, 随着DNA结构变化,聚合酶与启动子形成 的封闭复合物转变成开放复合物,此时 DNA序列中有一小段双链被解开。 三元复合物:在RNA转录的模板识别阶段,开 放复合物与最初的2个NTP相结合并在它们之间 形成磷酸二酯键后即转变成包括RNA聚合酶、 DNA和新生RNA的复合物。
真核生物启动子的特点
Hogness区( Hogness box)
在真核生物中,位于转录起始点上游-30—-25bp 处存在共同序列TATAAA,也称TATA区。
将TATA区上游的保守 序列称为上游启动子元 件(UPE)或上游激活 序列(UAS)
CAAT区( CAAT box)
在真核生物中,位于转录起 始点上游-78—-70bp处存在 一段共同序列CCAAT,它 与原核生物中-35区相对应。 GC区、增强子区
tRNA、5SrRNA、snRNA
α-鹅膏覃碱
低浓度:动植物中,RNA聚合酶II被抑制, RNA聚合酶I没影响,RNA聚合酶III在各生物 中差异大。 高浓度:动物细胞中抑制转录,但昆虫中无 影响。
启动子的结构
你还记得什么 是启动子吗?
转录单元(transcription unit): 是一段
从启动子开始至终止子结束的DNA序列。
真核生物
转录起始前复合物 转录调控因子 promoter
(preintiation transcription complex,PIC)
PIC
转 录 起 始 前 复 合 物
封闭复合物:在RNA转录的模板识别阶段, 聚合酶与启动子可逆性结合形成的复合物, 此时DNA处于双链状态。 强启动子
快速/不可逆 封闭复合物 开放复合物
模板识别
转录起始
转录延伸
转录终止
模板识别(template recognition):
指RNA聚合酶与启动子DNA双链相互作用并 Promoter是基因转录起始所必需的 与之相结合的过程。
一段DNA序列,是基因表达调控的 上游顺式作用元件之一。
原核生物 RNA聚合酶 直接识别并结合 promoter RNA聚合酶
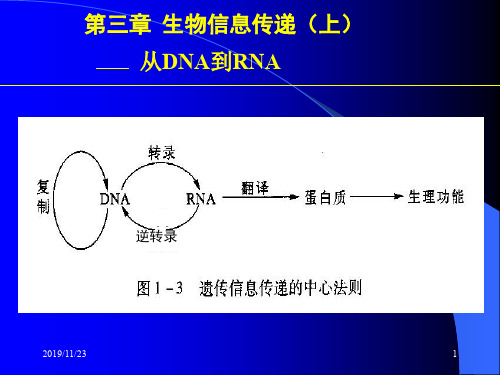
复制(DDDP) 转录(DDRP) DNA 反转录(RDDP) RNA 复制(RDRP) RNA 翻译 蛋白质
肽链的终止及释放
核糖体从mRNA 上解离,准备新 一轮的合成
• 三联子密码(codon)的性质 AUG • 起始密码子(initiation codon)和终止密码子 (termination codon) UAA Ochre • tRNA的特点 UAG Amber • 比较原核与真核细胞核糖体的组成 UGA Opal • 核糖体的活性中心 • 信号肽 • 叶绿体蛋白质的跨膜转运 • 蛋白质的翻译后加工 • 抗生素与蛋白质合成
DNA转座的遗传学效应
1、引起插入突变 各种IS、Tn转座子都可以引起插入突变。 2、产生新基因 3、产生染色体畸变 4、引起生物进化
二、从DNA到RNA
RNA转录的基本过程
RNA链合成特点:RNA是按 5’→3’方向合成,在 RNA聚合酶催 化下,以DNA链中的 反义 链为模板,以 4种核苷三磷酸 为原料, A-U、T-A、 G-C 根据碱基配对原则 ,各核苷酸间通过磷酸二 酯键相连,(需要/不需要)引物的参与,合成的RNA带有与 DNA编码链(有意义链)相同的序列(A-U)。
1983年诺贝尔 生理医学奖
“如果你能脚踏实地从事研究并如实报告观察结果,同时保持健 康长寿,那么,你有可能获诺贝尔奖。” Barbara McClintock
DNA的转座(DNA的移位 transposition)
是由可移位因子(transposable element)介导 的遗传物质重排现象。
第三章 生物信息传递
一、DNA复制
• • • • • • DNA的半保留复制 DNA复制的主要方式 原核生物和真核生物DNA复制的特点 DNA复制的调控 DNA的修复 1 DNA的转座
自学
“倘若你认为自己迈开的 步伐是正确的,并且已经 掌握了专门的知识,那么, 任何人都阻挠不了你,.. 不必去理会人们的非难和 评头品足。” Barbara McClintock
•复合型转座子(composite transposon)
带IS序列 不带IS序列 带有某些抗药性基因(或 其它宿主基因)的转座子, 两翼往往是相同或高度同 源的IS序列。
末端常带有倒 置重复序列
TnA家族:不带IS序列体积 庞大(>5Kb)的转座子,带 有β-内酰胺酶(AmpR) 基因和两个转座必需基因。
转录终止(termination):
当RNA链延伸到转录终止时,RNA聚合酶不再形 成新的磷酸二酯键,RNA-DNA杂合物分离,转录 泡瓦 解 , DNA恢 复 双链状 态 , 而 RNA 聚合酶和 RNA链都被从模板上释放出来。
RNA聚合酶
原核生物
E.coli. RNA聚合酶 2个α亚基 1个β亚基 1个β’亚基 1个ω亚基 1个σ亚基 可能与核心酶组装及启动子识 别有关,并参与RNA聚合酶和 部分调节因子的相互作用。 与模板DNA、新生RNA链及核 核心酶 苷酸底物结合。
转录起始位点
terminator
RNA
原核生物启动子的特点
Pribnow区(Pribnow box):
在RNA聚合酶与DNA结合的区域内有一个由6个核苷 酸组成的序列,是RNA聚合酶的紧密结合点,该位 点的中央约位于起点上游10bp 处,又称-10区。
-35区:
在Pribnow box外存在 的与RNA聚合酶对启动 子识别有关的共同序列 TTGACA,它位于起点 上游35bp处。 -10区和-35区是RNA聚合酶与启动子的结合位点,能与σ因子 相互识别,具有很高的亲和力。
加工过程
加帽 加polyA尾
推测的生理功能
mRNA 从细胞核向细胞质运转,翻译起始 转录终止,翻译起始和mRNA降解
RNA的剪接 从mRNA、tRNA和rRNA前体分子中切除内含子 RNA的切割 从前体RNA中释放tRNA和rRNA分子
DNA转录生成的原始转录产物— 核不均一RNA(hnRNA) 即 mRNA的前体,经过 5’加帽 和 3’加多聚腺苷酸 ,再经 过 RNA的剪接 ,编码蛋白质的外显子部分就连接成为一个 连续的 开放阅读框(ORF) ,通过核孔进入细胞质,即可作 为蛋白质合成的模板了。
核酶 (ribozyme)
ribonucleic acid
enzyme
• 是一类具有催化功能的RNA分子,通过催化靶位点 RNA 链 中 磷 酸 二 酯 键 的 断 裂 , 特 异 性 剪 切 底 物 RNA分子,从而阻断基因的表达。 酶不仅是蛋白质 T4 RNA、一些植物类病毒和拟病毒也可自我剪接 核酶 是继反转录之后对中心法则的又一重要修正, 遗传物质 酶 说明RNA既是 又是 。
模板识别
转录起始(initiation)
• 不需要引物 RNA聚合酶合成9个以上核苷酸并离 开启动子区,转录进入延伸阶段。 RNA聚合酶通过启动子的时间代表一 个启动子的强弱。
转录延伸(elongation):
RNA聚合酶释放σ因子离开启动子后,核心 酶沿模板DNA链移动并使新生RNA链不 断伸长的过程。 E.coli. RNA聚合酶 50-90个核苷酸/秒
自学
氯霉素—可能阻止mRNA与核糖体结合
竞争性抑制剂
四环素类—可能阻止AA-tRNA与核糖体结合 链霉素、新霉素、卡那霉素等—可能干扰 AA-tRNA与核糖体结合而产生错读
The End of Chapter III
DNA是遗传信息的携带者
DNA通过复制将遗传信息由A polymerase——DDDP DNA dependent RNA polymerase——DDRP RNA dependent RNA polymerase——RDRP RNA dependent DNA polymerase——RDDP
原核与真核生物mRNA的特征比较
原核生物的mRNA
1、半衰期短 2、可能以多顺反子形式存在 3、5’端无帽子结构,3’端没 有或只有较短的polyA结构 单顺反子mRNA :
只编码一个蛋白质的 mRNA。
多顺反子mRNA :
编码多个蛋白质的mRNA。
真核生物的mRNA
1、5’端有“帽子”结构
2、3’端有polyA“尾巴”结 常被甲基化 构 帽 子 结 构 是 GTP 和 原 mRNA5’ 三 磷 酸 腺 苷 • “帽子”是什么? (或鸟苷)缩合反应的产物。
全酶 (holoenzyme) 催化核心
?
负责模板链的选择和转录起始, 使酶专一性识别模板上的启动 子。存在多个σ因子,用以识 别不同的启动子。
一种RNA聚合酶几乎 合成所有RNA
真核生物
一般由8-16个亚基组成
转录产物 RNA聚合酶I RNA聚合酶II RNA聚合酶III rRNA hnRNA
mRNA
• 所有真核mRNA都有“尾巴” 组蛋白基因 吗? 转录后 • “尾巴”是何时产生的? 真核基因转录终止位点上游15• 什么是加尾和初级转录产物 30bp处的保守序列AAUAAA 准确切割所必需的? RNA聚合酶II至尾巴下游0.5-2Kb •RNA聚合酶都在“尾巴”终止 核苷酸处 吗?