苯丙氨酸氨解酶基因
苯丙氨酸解氨酶_PAL_的研究进展_崔建东

食品工业科技
ScienceandTechnologyofFoodIndustry
综 述
苯丙氨酸解氨酶 (PAL)的研究进展
崔建东 , 李 艳 , 牟德华 (河北科技大学生物科学与工程学院 , 河北石家庄 050018)
苯丙氨酸解 氨酶 (Phenylalanineammonialyase, PAL, EC.4.3.1.5)广泛存在于各种植物和少数微生物 中 , 是植物体内次生代谢的关键酶和限速酶 , 在微生 物中可催化 L-苯丙氨酸脱氨生成肉桂酸和氨 。 纯化 后的 PAL可以用于治疗某些肿瘤 [ 1] , 监控苯丙酮尿 患者血浆中的苯丙氨酸含量 , 治疗苯丙酮尿患者[ 2] , 但其主要用途是催化反式肉桂酸转化合成 L-苯丙氨 酸 (L-phenylalanine, L-phe)。 L-phe是合成阿斯巴 甜 (aspartame, APM)的主要原料 , 随着 APM的全球 热销和反式肉桂酸生产成本的降低 , 利用 PAL转化 反式肉桂酸合成 L-phe的方法成为当前生物化工领 域研究的热点 。本文就 PAL的基本特点及近几年的 研究进展作简要介绍 , 重点介绍微生物 PAL的研究 进展 。
3 微生物 PAL研究
1966年 , Ogata等 [ 9] 在研究芳香族氨基酸的微生 物代谢时发现 , 红酵母属中的某些种能以 L-苯丙氨 酸为唯一碳 、氮源生长 , 并在培养基中积累肉桂酸 。 随后 , 该酶相继在其它微生物中被发现 , 主要有霉菌 与酵母菌 。包括[ 10] :链霉菌属 (Streptomyces)、枝孢霉 属 (Cladosporium)、粘 滑 菇 属 (Hebeloma)、鳞 伞 属 (Pholiola)、 鬼 伞 属 (Coprinus)、 共 头 霉 (Symcephalasilrum)、烟熏离褶伞属 (Lyophyllum)、内 孢霉 属 (Endomuces)、曲 霉属 (Asperhilllus)、地霉 属 (Geotricum)、 从 梗 孢 属 (Monilella)、 小 丛 壳 属 (Glomerella)、卵 孢酵母 属 (Oosporidium)、类酵 母属 (Saccharomycodes)、掷酵母属 (Sporidiobolus)。 目前 , 对微生物 PAL的研究主要集中在菌种选育以及提高 PAL活性和稳定性方面 。
拟南芥苯丙氨酸解氨酶(PAL)基因的研究进展

拟南芥苯丙氨酸解氨酶(PAL)基因的研究进展孙海燕;全雪丽;付爽;吴松权【摘要】苯丙氨酸解氨酶(PAL)是催化苯丙烷代谢途径第1步反应的限速酶,广泛地参与植物生长发育过程中的各种生理活动.本文概述了拟南芥PAL基因的分子生物学和生理学研究进展,主要包括PAL基因的结构、表达特性、调控机制及其参与的植物生理学的意义,为进一步阐明PAL基因的功能提供参考依据.【期刊名称】《延边大学农学学报》【年(卷),期】2016(038)001【总页数】5页(P88-92)【关键词】拟南芥;苯丙氨酸解氨酶;表达;生理作用【作者】孙海燕;全雪丽;付爽;吴松权【作者单位】延边大学农学院,吉林延吉133002;延边大学农学院,吉林延吉133002;延边大学农学院,吉林延吉133002;延边大学农学院,吉林延吉133002【正文语种】中文【中图分类】Q943苯丙烷代谢途径是陆生植物生长和发育所必需的,是植物长期适应自然环境的结果[1-2]。
苯丙氨酸解氨酶(EC 4.3.1 5, PAL)催化L-苯丙氨酸(L-Phe)的非氧化脱氨基作用,生成肉桂酸,是生物合成苯丙烷类天然产物的第1步[3-4],也是第1个被鉴定的植物“防御基因”[5]。
也是苯丙烷类代谢途径中研究最多的酶[6]。
肉桂酸是植物生长、发育和环境适应所必需的各种苯丙烷类物质的合成起始物[7]。
苯丙烷类化合物是植物中大量酚类化合物的前体,包括木质素、黄酮类、异黄酮类、香豆素、芪类和水杨酸等,它们在维持植物结构、抵御紫外线、形成花青素和植保素、保持花粉活力、信号转导与交流等方面发挥着重要作用 [1,5,8]。
自1961年Koukol和Conn首次描述PAL以来,苯丙氨酸解氨酶得到了广泛的研究[9],它是控制苯丙烷途径生物合成去向的关键酶和限速酶[7,10]。
拟南芥是一种十字花科植物,广泛用于植物遗传学、发育生物学和分子生物学的研究,已成为一种典型的模式植物,该植物具有个体小、生长周期快、形态特征简单、生命力强、基因组小、遗传操作简单等优点[11-12]。
油松苯丙氨酸解氨酶基因的克隆与序列分析

因的 c D NA 和 D NA 序 列 , c DNA 序列 全长 2 1 5 7 b p , 编码 7 1 8个氨 基 酸 , 包含 有 完整 的 OR F, 命 名
为 P AL( Ge n B a n k登 录 号 : J X 2 8 0 4 6 0 ) 。 通 过 核 苷 酸 序 列 多重 比 较 表 明 , T n P AL 基 因 与 其 他 植 P AL 物P A L基 因 高 度 同 源 , 预 测 的 蛋 白序 列 包 含 与 水 稻 、 玉米 P AL 相 同 的 脱 氨 基 位 点 和 催 化 活 性 位 点 。P AL 系统 进 化 树 显 示 , Ta P AL 与 松 科 类 植 物 P AL 聚 类 关 系 最 近 。 另 外 , 对 比 发 现 基 因组 D NA 序 列 与 c D NA 序 列 完 全 一 样 , 表 明 该 基 因 无 内含 子 。 关键 词 : 油松 ; 苯 丙氨 酸 解 氨 酶 ; 克隆 ; 序 列 分 析 中图分 类号 : ¥ 7 9 1 . 2 5 4 文献标 志码 : A 文 章编 号 : 1 0 0 1 — 7 4 6 1 ( 2 0 1 3 ) 0 3 — 0 0 9 8 — 0 6
c o nt ai ni n g a c o mp l e t e op e n r e a d i ng f r a me a nd 7 1 8 a mi n o a c i d s we r e e n c od e d,d e s i g na t e d a s TaPA L ( Ac c e s —
wi t h o t h e r p l a nt s’v i a mul t i pl e a l i g nme n t . Th e d omi n a t i o n s i t e s a nd c a t a l y t i c a c t i ve s i t e s a p pe a r e d i n PAL p r ot e i n of Or yz a a n d Ze a mays we r e a l s o f o und i n t h a t o f i n f e r r e d pr o t e i n s e q ue nc e . Ph yl o g e ne t i c t r e e a —
苯丙氨酸解氨酶(PAL)检测试剂盒(苯丙氨酸比色法)
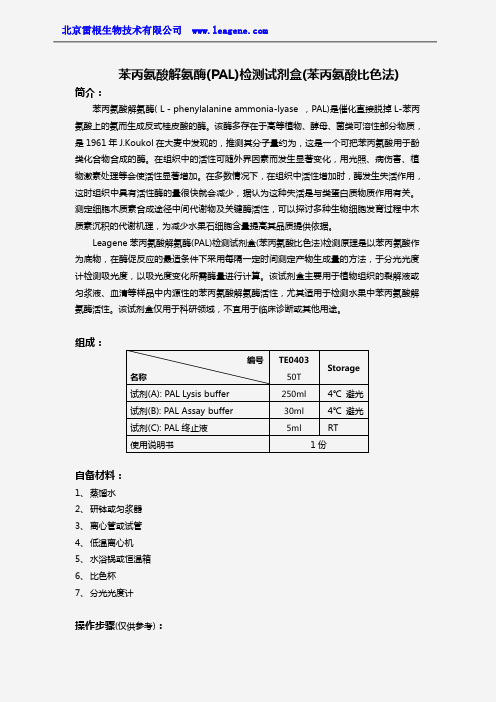
苯丙氨酸解氨酶(PAL)检测试剂盒(苯丙氨酸比色法)简介:苯丙氨酸解氨酶(L-phenylalanine ammonia-lyase,PAL)是催化直接脱掉L-苯丙氨酸上的氨而生成反式桂皮酸的酶。
该酶多存在于高等植物、酵母、菌类可溶性部分物质,是1961年J.Koukol在大麦中发现的,推测其分子量约为,这是一个可把苯丙氨酸用于酚类化合物合成的酶。
在组织中的活性可随外界因素而发生显著变化,用光照、病伤害、植物激素处理等会使活性显著增加。
在多数情况下,在组织中活性增加时,酶发生失活作用,这时组织中具有活性酶的量很快就会减少,据认为这种失活是与类蛋白质物质作用有关。
测定细胞木质素合成途径中间代谢物及关键酶活性,可以探讨多种生物细胞发育过程中木质素沉积的代谢机理,为减少水果石细胞含量提高其品质提供依据。
Leagene苯丙氨酸解氨酶(PAL)检测试剂盒(苯丙氨酸比色法)检测原理是以苯丙氨酸作为底物,在酶促反应的最适条件下采用每隔一定时间测定产物生成量的方法,于分光光度计检测吸光度,以吸光度变化所需酶量进行计算。
该试剂盒主要用于植物组织的裂解液或匀浆液、血清等样品中内源性的苯丙氨酸解氨酶活性,尤其适用于检测水果中苯丙氨酸解氨酶活性。
该试剂盒仅用于科研领域,不宜用于临床诊断或其他用途。
组成:自备材料:1、蒸馏水2、研钵或匀浆器3、离心管或试管4、低温离心机5、水浴锅或恒温箱6、比色杯7、分光光度计操作步骤(仅供参考):编号名称TE040350TStorage试剂(A):PAL Lysis buffer250ml4℃避光试剂(B):PAL Assay buffer30ml4℃避光试剂(C):PAL终止液5ml RT使用说明书1份1、准备样品:①植物样品:取植物组织或水果中层果肉,加入PAL Lysis buffer,冰浴情况下充分捣碎研磨或匀浆,离心,留取上清液,冻存,用于苯丙氨酸解氨酶的检测。
②血浆、血清和尿液样品:血浆、血清按照常规方法制备后可以直接用于本试剂盒的测定,冻存,用于苯丙氨酸解氨酶的检测。
植物苯丙氨酸解氨酶研究进展

植物苯丙氨酸解氨酶研究进展摘要苯丙氨酸解氨酶(phenylalanine ammonia-lyase,pal,e.c4.3.1.5)是催化苯丙烷代谢途径第一步反应的酶,也是这个途径的关键酶和限速酶,对植物具有非常重要的生理意义。
综述了苯丙氨酸解氨酶的存在与分布、基本特性、生理代谢意义、抗病以及基因表达与调控等,为pal在植物抗病的应用上提供基础数据。
关键词苯丙氨酸解氨酶;生理作用;抗逆境;基因表达与调控中图分类号s661.1文献标识码a文章编号1007-5739(2009)01-0030-04苯丙氨酸解氨酶(phenylalanine ammonia-lyase,pal,e.c4.3.1.5)催化l-苯丙氨酸解氨生成反式肉桂酸,是连接初级代谢和苯丙烷类代谢、催化苯丙烷类代谢第一步反应的酶,也是苯丙氨酸代谢途径的关键酶和限速酶。
莽草酸途径产生的莽草酸通过分枝酸、预苯酸经转氨作用生成苯丙氨酸,从而进入苯丙烷类代谢途径,苯丙烷类代谢可生成反式肉桂酸、香豆酸、阿魏酸、芥子酸等中间产物,这些中间产物可进一步转化为香豆素、绿原酸,也可以形成coa酯,再进一步转化为木质素、黄酮、异黄酮、生物碱、苯甲酸酯糖苷等次生代谢产物[1,2]。
一切含苯丙烷骨架的物质都由该代谢途径直接或间接生成。
在生物次生物质代谢中具有防紫外线伤害、抵抗病原体的侵害、保持花粉生活力及形成植物花青素等多种重要作用。
该酶在不同组织中、不同的内外因素调节下,含量水平及其基因表达的时空方式均有所不同。
1pal的存在和分布1961年koukal和conn首次在绿色植物大麦幼苗中发现了pal,并进行了分离纯化[3]。
随着研究的深入,该酶已被固定化,用于工业化生产苯丙氨酸。
至今,pal已在所有绿色植物中发现,在真菌、细菌、藻类中也有存在。
不同植物中pal活性不同,在同一株植物的不同组织部位,pal活性也不同。
一般越嫩的部位pal活性越高,如杨树的幼叶、顶芽、幼茎中pal活性最高,而老茎和成熟叶中pal则较低[4]。
芥蓝(Brassica alboglabra)苯丙氨酸解氨酶基因BaPAL的克隆、表达与序列分析

u i r ot sng o s, s e s, la e tm e v s, fowe b l r uds op n fowe s a d slq s e , e l r n iiua DN A a PCR t m plt s, s e ae r s e tvey.Re uls i ia e h e p ci l s t ndc t d t atBaPA L n s e pr s e n alor nsw ih t x e in o oo s, ge e wa x e s d i l ga t he e c pto fr t a s hi l xp e s d i pe lw e s Se nd wa gh y e r s e n o n fo r . que e c m pa ion o PA L nd PA Ls f o he l n s nc o rs fBa a r m ot r p a t r v ae ha B. al o ab a, B. n e e ld t t b gl r apu a d s n B . r apa v r Chi nss e e he l es r l tve . B. a. ne i w r t cos t ea i s al gl b a ha l e ea i hi n ha e h a eca n ph og ne i r ew ih .i bo a r d a cos rr ltons p a d s r d t e s m ldei yl e tc te t f ndi tc d go ia an A . t alan . H o e e h i a w v r,i s dit nty r l t d t e nsf he rl t wa s a l e a e o f r or t i ow e e e h s qu nc om o o l gy.
苯丙氨酸解氨酶综述2

苯丙氨酸解氨酶摘要:本文从前言介绍了PAL基本特性(基本性质、酶学性质和分子结构)、苯丙氨酸解氨酶的研究意义(植物生理代谢上的意义和医疗上的意义)、PAL 研究现状(产PAL微生物的选育及其稳定性研究和植物PAL基因的克隆与表达)和PLA 发展趋势四个方面对苯丙氨酸解氨酶进行综述。
Abstract: The article of phenylalanine ammonia lyase were reviewed from the basic characte- ristics of PAL (the basic nature of the enzymatic properties and molecular structure), the signifi -cance of the phenylalanine ammonia-lyase (the meaning of Plant Physiology and metabolism, and medical significance), PAL Researching (producing PALBreeding and stability of microbial and plant PAL gene cloning and expression) and the development trends of PLA.关键词:苯丙氨酸解氨酶、PAL基本性质、PAL酶学性质、PAL分子结构、植物生理代谢作用、细胞分化和木质化、植物色素形成、根瘤、抗逆境、抗虫性和抗病性、PAL选育、活性和稳定性研究、基因的克隆与表达、苯丙烷类代谢Key words:Phenylalanine ammonia lyase, the basic nature of the PAL, PAL enzymatic properties, the PAL molecular structure, plant physiology and metabolism, cell differentiation and lignification, plant pigments formed nodules, stress resistance, insect resistance and disease resistance, PALbreeding, activity and stability of gene cloning and expression of phenylpropanoid metabolism .一、前言1.PAL的基本特性1.1 PAL的基本性质苯丙氨酸解氨酶( phenylalanine ammonia- lyase, PAL,EC4. 3. 1. 5)是连接初级代谢和苯丙烷类代谢、催化苯丙烷类代谢第一步反应的酶,是苯丙烷类代谢的关键酶和限速酶,也是苯丙烷类代谢途径中研究最多的酶。
杉木苯丙氨酸解氨酶基因ClPAL的克隆与表达分析_徐莉莉

童再康
林二培
黄华宏
临安 311300 )
亚热带森林培育国家重点实验室培育基地
摘
要:
利用 RACE 方法从杉木中分离得到 3 个编码苯丙氨酸解氨酶 ( PAL ) 的全长 cDNA 序列, 分别为 2 455 ,
2 418 , 2 689 bp, 710 , 709 。氨基酸序列分析显示, HAL 编码的氨基酸个数分别为 709 , 推导的氨基酸序列包含 PAL和 PAL 2 个功能域以及酶活性中心序列 ( GTITASGDLVPLSYIA) 。荧光定量 PCR 分析表明, 虽然 3 个 ClPAL 基因在 调查的器官和组织中皆有表达 , 但仍表现出一定的组织特异性 。ClPAL1 在木质化茎中的表达量明显高于针叶 、 雌 ClPAL3 主要在木质化茎中表达 ; 二者均在木质部中优势表达 , 根和非木质化茎中的表达量 , 且在中间材中的 球花、 GA3 和 BR 能促进木质部 表达量高出边材 20 倍以上。ClPAL2 主要在富含韧皮部针叶和皮层中高丰度表达 。 IAA、 ClPAL1 和 ClPAL3 的表达模式相 细胞的分化, 与木质素的生物合成有直接或间接的联系 。 经此 3 种激素处理后, 均受到不同程度的诱导 , 其中 IAA 和 BR 的诱导效应相对明显; ClPAL2 基因的表达则受到相对弱的抑制 。这些 似, ClPAL1 在应压 结果说明 ClPAL1 和 ClPAL3 可能主要参与了杉木木材中木质素的合成过程 。在应压木诱导形成中, 处理 10 天后达对照的 1. 4 倍, 在 对 应 区 木 质 部 中 则 呈 现 下 调 表 达; 区木质部的表达量表现为先下降后上升 , ClPAL2 在应压区木质部中表现为上调表达 , 而在对应区木质部中, 仅处理 10 天后的表达量明显高于对照 ; ClPAL3 ClPAL2 基因在中间材中的表达丰度也明显高于边材 , 的表达量变化不是很明显 。另外, 是否参与杉木心材特殊色 泽的形成值得深入研究 。 关键词: 杉木; PAL 基因; 基因克隆; 基因表达; 木材形成 文献标识码: A 文章编号: 1001 - 7488 ( 2013 ) 12 - 0064 - 09 中图分类号: S718. 46 ; Q943. 2
水稻苯丙氨酸解氨酶的生物信息学分析

2010年12月第33卷第4期湖南师范大学自然科学学报Journa l o f N atura l Sc ience ofH unan N o r m al U niversityV o.l33N o.4D ec.,2010水稻苯丙氨酸解氨酶的生物信息学分析康晓慧1*,雷桅2,张梅1(11西南科技大学生命科学与工程学院,中国绵阳621010;21中山大学生命科学学院,国家教育部食品与健康工程研究中心,中国广州510650)摘要苯丙氨酸解氨酶(PAL)是水稻防卫反应中的关键蛋白,采用生物信息学工具和方法对G enBank中的日本晴水稻p al基因的核苷酸序列及其相应氨基酸序列进行预测和分析,获得了其分子结构、理化性质、系统演化和生物学功能等重要参数.结果表明:水稻pal基因序列只有开放阅读框,其编码蛋白亲水性较强,无跨膜转运肽,且定位于细胞基质,二级结构以随机卷曲和A-螺旋为主要构件,三维建模成功,并明确了多个功能位点.关键词水稻;苯丙氨酸解氨酶;生物信息学;防卫基因;抗病性中图分类号Q946.5;S511.2+2文献标识码A文章编号1000-2537(2010)04-0089-06B i o i n f or m ati c Analysi s of Phenyl a l a nine Ammoni a-Lyase Gene i n O ryza S a ti vaKANG X iao-hui1,LE I W ei2,Z HANG M ei1(1.Co llege o f L ife Sc i ence and Eng i neering,Sout hw est U n i versity of Sc i ence and T echno logy,M ianyang621010,Chi na;2.F ood and H ea lth Eng i neering R esea rch Center of State Educa ti onM i n istry,College o f L ife Sc i ence,Sun Y at-sen U n i versity,G uangzhou510650,China)Abst ract Pheny lalan i n e a mm onia-lyase(PAL)p lays a cr ucia l ro l e in plan t defense m echanis m.The nucleo-ti d e and its corresponding a m ino ac i d sequence of pal gene fro m Oryza s ativa(japonica cu lti v ar-g r oup),reg istered i n GenBank,are analysis by the b i o i n f o r m atic m ethod,and subsequentl y so m e i m portant para m eters are ga i n ed i n-clud i n g m o lecu lar structure,physi o che m ica l properti e s,phy logenetic evolution and b iolog ical functi o n.The result is as f o llo w i n g:there is on ly ORF in O s pal fu l-l leng th sequence,and its coding prote i n has no trans m e m brane to-polog i e al str ucture,and located in cytoplas m,w ith A-heli x and rando m coil asm ai n co mponents in secondary struc-ture.The3D m odel is constr ucted,and so m e functionalm otifs are deter m i n ed.K ey w ords Or yza sativa;phenyla lanine a mm on ia-lyase;b i o i n fo r m atics;defense gene;resistance抗病性是植物与其病原生物在长期的协同进化过程中相互适应、相互选择所形成的一种可遗传特性.植物被病原物侵染后,会发生一系列生理变化和防卫反应,包括植保素和木质素等抗病物质的合成和积累,其中苯丙烷类代谢与植物的抗病性密切相关,苯丙氨酸解氨酶(pheny lalan i n e a mm onia-lyase,PAL)是该途径中的第一个关键酶,也是限速酶.它催化L-苯丙氨酸非氧化性脱氨生成反式肉桂酸(c i n na m ic aci d,C A),即是植保素、木质素、类黄酮和某些酚类等生物合成的通用前体.因此,该酶在植物病理学研究中有极重要的地位.P AL为多基因家族所编码,但遗传特性相当保守,目前已从马铃薯(Solanu m tuberos um)、水稻(Or yza sativa)、烟草(N ico tiana t a bacum)和黄瓜(Cucu m is sativus)等多种植物中克隆了pal基因,并将其作为植物抗病机制研究的重要内容.水稻是世界上最重要的粮食作物之一,而病害给水稻的产量和质量造成了巨大的损失,因此水稻抗病的分子机理和遗传改良成为一项重要的课题,而作为关键防卫基因的pal则有望成为理想的调控靶点.粳稻品种日本晴(N i p ponbare)是单子叶植物基因组学研究的模式植物,测序发现其pal基因家族包含9个成员.本文利用生物信息学的方法,选取其中一个成员基因,对其核苷酸及相应氨基酸序列的理化性质、结构特征、生¹收稿日期:2010-10-22基金项目:国家/十一五0科技支撑计划基金资助项目(2006BAD08A01)*通信作者,E-m a i:l xhu i k09@126.co m化功能和系统演化等进行了预测和分析,为今后深入研究PAL 蛋白的酶学特性和植物病害胁迫的分子机理提供一定的理论参考.1 材料与方法粳稻品种日本晴的苯丙氨酸解氨酶(PAL)核苷酸与氨基酸序列均来自于GenBank 中已登录的数据信息:水稻Oryza sativa (japonica cu ltivar -group)(A ccession :AY224546).利用Vector NT I Su it 8等序列分析软件和www .ncb.i nl m .nih .gov 、www .expasy .org 等网站提供的各种在线生物信息学工具进行预测和分析.核苷酸和氨基酸序列的分子结构和理化性质分析使用ProtPara m 工具和V ecto r NT I Su it 8软件完成[1];序列同源性比对使用Vector NTI Su it 8完成;分子系统发生树使用C lusta lX 软件比对后在M EGA3的Phy logeny 中以N eighbor -Jo i n i n g 方法构建[2];亚细胞定位情况的考察使用Tar getP 1.1Server 在线完成[3];跨膜结构域和疏水性分析分别使用TMHMM [4]和ProtSca l e [5]完成;m o tif 和蛋白质功能分别用Scanprosite [6]和Pr o tf u n [7]分析;蛋白质二级结构预测和三级建模分别使用GOR [6]和S W ISS -MODEL[8-9]完成,3D 模型在W ebLab V ie w er L ite 4.2软件中编辑.2结果与分析图1 水稻PAL 的核苷酸及其编码氨基酸序列2.1 水稻pal 基因的核酸及相应氨基酸序列的分子结构和理化性质分析水稻pal 基因的全长mRNA 序列及其推导的氨基酸序列如图1所示,利用Vector NT I Su it 8软件分析得到的Os p al 基因的核苷酸序列及其相应氨基酸序列的结构和性质数据,发现Os pal 基因全长为2136bp ,其中开放阅读框(ORF)长为2133bp ,起始密码子为ATG ,终止密码子为TGA ,3c 和5c 非编码区(UTR)均不存在.水稻Os pal 基因编码蛋白的分子式为C 3188H 5129N 891O 973S 24,相对分子质量为72273,等电点为6.12,摩尔消光系数为37590,肽链长为671aa ,其中含带电氨基酸27.72%,极性氨基酸23.85%,疏水性氨基酸38.15%,酸性和碱性氨基酸比例为11.33%和10.13%,而含量最丰富的4种氨基酸占比为A la :10.88%;Leu :10.13%;Va:l 8120%;G ly :7.90%.90 湖南师范大学自然科学学报 第33卷2.2 水稻pal 基因的氨基酸序列的比对分析选取水稻(japonica cu ltivar -group)等6条植物PAL 氨基酸序列,在Vector NT I 8软件中进行多重比对分析.分析结果用不同颜色显示(图2),颜色越深的表示同源性越高,最深的区域则表示可能存在的重要功能图2 水稻PAL 和其他植物PAL 的氨基酸序列多重比对注:一致位点显示为黑底白字,保守位点显示为灰底白字,其他显示为白底黑字.活性位点标注*,M I O 区用方框表示.S alvia m iltiorrh i za (GenPept accessi on N o .:ABR14606);Tri foli um p retense (GenPept accession No .:BAE71252);Dauc u s carota (GenPept accessi on No .:BAA23367);Ipo m oe a ba t a t a s (GenPept access i on No .:BAA11459);N ic otiana t a bac um (GenPept access i on N o .:BAA22947);O ryza sa tiv a (Gen-Pep t access i on No .:AAO72666).域.分析结果显示所选植物PAL 具有较高的同源性,一致性达到57.4%,可见PAL 编码区高度保守,但是在N 端也表现出较明显的长度和组成上的变异性.推测这可能是由于苯丙烷类途径是植物初生代谢转向次生代谢的重要通路,广泛存在于植物体中,而作为苯丙烷类化合物生物合成途径中的第一个关键酶P AL 必须保持足够的遗传稳定性和演化趋同性.此外,在保守的PAL 比对图谱中,发现水稻PAL 具有典型的苯丙氨酸91第4期 康晓慧等:水稻苯丙氨酸解氨酶的生物信息学分析和组氨酸解氨酶的标志性模式,该保守基序(GT I T ASGDLVPLSY I A G )被包埋在酶活性中心;此外,也存在已查明的M I O 域,它通过A la -Ser -G ly 三肽的自动环化合脱水生成,毗邻底物结合位点,并负责产物E -肉桂酸酯/氨的催化形成.2.3 水稻pal 基因的氨基酸序列的分子系统进化树分析将9条来自不同植物的P AL 氨基酸序列在C lustal X 软件中进行完全比对后,在MEGA3软件中按照N eighbor -Jo ining 方案进行分子进化树分析,获得PAL 氨基酸序列的分子进化树分析结果(如图3).进化树用M EGA3软件基于Ne i g hbor -Jo i n i n g 方法(重复1000次)构建.遗传距离显示在分支上.9种植物的P AL 氨基酸序列聚成3支:甘薯、番薯和烟草聚成第1支,它们的多酚类物质含量较高;粘毛黄芩、丹参、豌豆和胡萝卜聚成第2支,前二者是主产黄酮活性成分的草药,后二者是含有丰富的香豆素和花青素等的蔬菜;马蹄竹和水稻聚成第3支,这两种植物的木质素合成能力较强,且同属禾本科.该进化树除反映了这9种植物间的亲缘关系外,也反映了不同植物类群所含苯丙素类化合物的种类和含量差异,即说明该指标与植物的系统演化进程密切相关.苯丙烷类代谢作为植物次生代谢的起始途径,广泛存在于所有植物,其衍生途径随着生命进化而分化出黄酮类、木质素类、植保素类等某些植物的主流次生代谢通路,而PAL 作为苯丙烷类途径的第一个关键酶,显然在植物的功能分化和系统进化过程发挥着重要作用.因此,以PAL 序列分析作为分子指标进行的遗传聚类,对于系统分类和自然进化研究具有一定的参考价值,同时也反映出植物的抗病性与植物本身的种质特性和进化水平密切相关.图3 水稻PA L 家庭和其他植物PAL 的分子进化分析2.4 水稻pal 基因编码蛋白的亚细胞定位和功能结构域分析TargetP 1.1Server 在线工具分析结果显示,水稻PAL 定位于细胞质基质,且无转运肽,预测可靠性等级均为2.可见水稻P AL 在游离核糖体上起始合成后,不进行蛋白转运,而是继续保留在细胞质基质中,直接催化苯丙素类化合物的生成.PAL 的多个重要保守结构域在O sP AL 蛋白中也被找到,包括S 213、Y 120、L 148、N 269、Q 357、Y 360、R 363、F 409、Q 497和4-m ethy li d enei m idazo le -5-one(M I O),这些区域对于PAL 的底物结合、催化活性和M I O 形成都至关重要.这与欧芹的实验结果一致,表明水稻PAL 也属于PAL 蛋白家族.图4 水稻PAL 蛋白的跨膜结构域预测2.5 水稻pal 基因编码蛋白的跨膜结构域和疏水性预测和分析用T MHMM Server v .2.0对水稻PAL氨基酸序列的跨膜结构域进行预测,结果如图4所示,水稻P AL 整条肽链都位于细胞膜内,也即是水稻P AL 不存在跨膜结构域,结合上述转运肽的预测,可以推断,水稻PAL 在细胞质基质中合成后,不经蛋白转运,直接锚定于细胞质基质中的特定部位行使催化功能.这与上述亚细胞定位分析结果相一致.92 湖南师范大学自然科学学报 第33卷图5 水稻PA L 蛋白的疏水性预测由ProtSca le 进行水稻pal 基因编码蛋白的氨基酸序列的疏水性预测结果表明(图5),多肽链具有最低分值-2.778,亲水性最强;而最高分值为2.389疏水性最强.整条多肽链表现为亲水性,但没有明显的亲水区域.结合跨膜结构域的预测结果,可以推断,水稻PAL 不存在明显的疏水区域,与Os PAL 不存在跨膜结构域的特征相吻合.2.6 水稻pal 基因的氨基酸序列二级和三级结构预测和分析GOR4在线工具预测水稻PAL 氨基酸序列的二级结构,结果如图6所示,水稻PAL 多肽链中,随机卷曲占44.02%,A -螺旋占43.88%,延伸链占12.10%,即随机卷曲和A -螺旋是OsPAL 多肽链中的主要结构元件,延伸链散布于整个蛋白质中.图6 水稻PAL 蛋白的二级结构预测在PROST I E 中通过与已知物种PAL 功能位点的比对和计算,成功预测了水稻P AL 的M oti,f 发现OsPAL 含有5个重要的基序,分别是苯丙氨酸与组氨酸标记(Pheny lalan i n e and histi d i n e a mm on i a -l y ases signature)(153~168GT I T ASGDLVPLSY I A ,153~169GTI TASGDLVPLSY I A G),氨基化位点(183~186DGKK ),蛋白激酶C 磷酸化位点(Prote i n kinase C phosphory lation site)(16~18SLR,74~76SHR,172~174TGR,260~262T HK,452~454SAR ,513~515SAR,517~519SEK,648~650SER),酪氨酸激酶II 磷酸化位点(Case i n k-i nase II phosphory lation site)(335~338SVND,523~526TA I D,532~535SYAD,575~578TAFE ,635~638SPGE ),N-糖基化基序(N-g l y cosy lati o n m otif)(99~102NGSD )和N-肉豆蔻酰化位点(N-m yristoy lation site)(20~25GQVAAV,65~70GVTTGF ,95~100GI FGNG ).再将水稻PAL 氨基酸序列上传到S W ISS -MODEL 的建模服务器中进行PAL 结构的三维建模,通过同源搜索和比对,以殴芹(Petroselinum crispu)为模板初步构建了O sP AL 的三维模型,然后在V ie w er L ite 4.2软件中进行序列编辑和位点设置,获得OsPAL 的三级结构模型,相关功能位点见标示于图7.图7 水稻PA L 的三维建模及重要基序93第4期 康晓慧等:水稻苯丙氨酸解氨酶的生物信息学分析94湖南师范大学自然科学学报第33卷3结论与讨论苯丙氨酸解氨酶在木质素、植保素等植物抗病物质的代谢途径中起着重要作用,在水稻的抗病机制研究中也被公认为是重要的生理指标.生物信息学是后基因组时代,以计算机科学技术为平台,分析并挖掘生物学数据,从而为实验研究提供理论依据的新兴学科.本文利用生物信息学软件和方法,对日本晴水稻防卫基因pal的结构性质、生化功能和系统进化等进行了预测和分析,发现O s pal基因的核苷酸序列只包含ORF,缺少两端UTR,无跨膜结构域和质体转运肽,定位于细胞质基质.近年的研究也发现,细胞间质部分P AL活性最高,电镜技术显示PAL分散在细胞的基质中.这些实验结果也间接地验证了本定位预测的可能性.本项研究明确了P AL蛋白的二级结构以A-螺旋和随机卷曲为主,三维建模成功,并明确了多个功能基序.这些重要生物学参数的获得为进一步研究PAL的酶学性质和水稻的抗病机制奠定了理论基础.PAL作为抗病性指标蛋白的水稻生物学研究已有诸多报道,但大多是以PAL酶活变化表征抗病能力强弱的生化研究.本文解析了OsPAL蛋白的分子结构,并描述了其细胞学特性,将有助于深入阐述水稻病理生理与抗性获得机制;而对O s pal基因的序列和系统学演化分析结果,将为基于O s pal的基因操作和遗传工程提供重要的调控靶点.因此,本项研究对于水稻抗病性分子育种也有重要参考价值.参考文献:[1]LEIW,LUO K M,S HU I X R,et al.Co m pute si m ulati on to charac terize structure and functi on o f chalcone synt hase from scute-llaria ba ica lensis georg i[J].M o l ecular B i o logy,2009,43(6):1012-1017.[2]KUM AR S K,TMAM URA K,J AKO BSEN I I B,et al.M EGA2:M o l ecular evo l u tionary gene ti cs ana l ys i s so ft wa re[J].B i o i n-for m atics,2001,17:1244-1245.[3]O LOF E,HENR I K N,SªREN B,et al.P red i cti ng subce ll u lar l o ca li zati on o f prote i ns based on t he ir N-ter m i na l a m i no ac i d se-quence[J].JM o l B i o,l2000,300:1005-1016.[4]I K EDA M,ARA I M,LAO D M.T rans m e m brane t opo l ogy pred icti on m ethods:A reassess m ent and i m prove m ent by a consensusm ethod usi ng a da taset o f exper i m enta lly character ized trans m e mbrance topo l og i es[J].In Silico B i o,l2002,2:19-33.[5]GA S T EI G ER E,HOOGLAND C,GATT I K ER A,et al.P rote i n Identifica ti on and ana l y si s t oo ls on the ExPA Sy Serv er[C]//John M W a l ker.The Proteom ics Protoco lsH andbook.N ew Y ork:H u m ana P ress,2005.[6]COM BET C,BLANCHET C,GEOUR J ON C,et al.NPS@:N e t w ork pro tei n sequence ana l ysis[J].T IBS,2000,25:147-150.[7]J EN SEN J L,ST¶R FELDT H H,BRUNAK S.P red icti on o f hu m an pro te i n f unc ti on accord i ng t o G ene On t o logy categor i es[J].B i o i n f o r m atics,2003,19:635-642.[8]ARNOLD K,BORDOL I L,KOPP J,et al.The S W ISS-MODEL W orkspace:A w eb-based env iron m en t for prote i n structure ho-m o l ogy modelli ng[J].B io i nfor m atics,2006,22:195-201.[9]SC HW EDE T K J,GUEX N,PEITSCH M C.S W ISS-M ODEL:an au t om ated prote i n ho m o l ogy-m ode li ng server[J].N uc l e icA cids R es,2003,31:3381-3385.(编辑王健)。
苯丙氨酸解氨酶的研究进展

No.7.2006trehalose-6-phosphontessynthase(TPS1)genefromSac-charomycescerevisiae[J].MolCells,2000,10(3):263-268[16]AjayK.Garg,JukonKim,ThomasG,etal.Trehaloseac-cumulationinriceplantsconfershightolerancelevelstodifferentabioticstresses[J].ProcNallAcadSci,2002,99(25):15898-15903[17]张树珍,郑学勤.基因枪介导甘蔗遗传转化研究初报[J].热带作物学报,2001,22(1):35-41[18]王自章,张树珍,杨本鹏,等.甘蔗根癌农杆菌介导转化海藻糖合酶基因获得抗渗透胁迫能力增强植株[J].中国农业科学,2003,36(2):140-146[19]赵恢武,陈杨坚,胡莺雷,等.干早诱导性启动子驱动的海藻糖-6-磷酸合酶基因载体的构建及转基因烟草的耐早性[J].植物学报,2000,42(6):616-619苯丙氨酸解氨酶(Phenylalanineammonia-lyase,PAL,EC.4.3.1.5)广泛存在于各种植物和少数微生物中,催化L-苯丙氨酸(L-phenylalanine,L-Phe)脱氨生成肉桂酸和氨,是植物体内次生代谢的关键酶和限速酶。
在植物形成次生物质如木质素、植保素等中起重要作用,对植物生长发育、抵御病虫害、防紫外辐射及构成植物支撑系统等方面具有重要意义。
自Koukol首次从绿色植物中发现并分离纯化出苯丙氨酸解氨酶的研究进展贺立红1,2,张进标3*,宾金华3(1.仲恺农业技术学院生命科学学院,广州510225;2.华南农业大学园艺学院,广州510642;3.华南师范大学生命科学学院,广州510631)摘要:对苯丙氨酸解氨酶的分布、基本特性、分子结构、各种因子对其活性的影响和生理功能方面作了简要的概述,旨在为该酶的进一步研究提供参考。
植物苯丙氨酸解氨酶基因的表达调控与研究展望

果树学报2003,20(5):351~357Journal of Fruit Science植物苯丙氨酸解氨酶基因的表达调控与研究展望程水源1,2陈昆松1*刘卫红2杜何为2(1浙江大学果树研究所,浙江杭州310029;2湖北农学院园林植物与观赏园艺重点学科组,湖北荆州434025)摘要苯丙氨酸解氨酶(hhenylalanine ammonia-lyase,PAL,EC4.3.1.5)是催化苯丙烷代谢途径第一步反应的酶,也是这个途径的关键酶,对植物有非常重要的生理意义。
根据有关文献综述了植物PAL的分布与定位、酶学性质,总结了生长发育、钝化因子与调节因子、末端产物等内部因素及光、温、机械损伤与生长调节剂等外部因素对PAL的调控作用,得出外部因子是在转录水平上对酶活性实施调控的结论,并运用酶学和分子生物学方面的知识阐述了其调控机理。
还着重阐述了PAL酶在果树上的研究现状与进展,并对其今后的研究及在果树上的应用进行了展望。
关键词苯丙氨酸解氨酶;基因表达与调控;调控机理中图分类号:S501.2文献标识码:A文章编号:1009-9980(2003)05-351-07Regulation and Expression of the PAL in Plant and Its OutlookCheng Shuiyuan1,2,Chen Kunsong1*,Liu Weihong2,and Du Hewei2(1Fruit Science Institute of Zhejiang Uniuersity,Hangzhou310000hhina;2The Key Discipline Group of Garden Plant and Ornamental Horti-culture in Hubei Agricultural hollege,Jingzhou,Hubei434025hhina)Abstract Phenylalanine ammonia-lyase(PAL,EC4.3.1.5),a key enzyme of hlant secondary metabolism,catalyzed the first reaction in the biosynthesis from L-hhenylalanine to a wide variety of natural hroducts based on the hhenylhrohane skeleton.It has great influence on hlant hhysiological function.The distribution,orientation and basic characters of PAL in hlants are reviewed.The regulating effects of inner factors such as growth and develohment,and PAL-inhibitor(PAL-I)and PAL-in-hibitor-regulation(PAL-IR),and end hroducts,and outer factors such as light,temherature,and mechanical hurt,and growth regulators on PAL are also summarized.The outer factor regulation to PAL rests on transcrihtional level.The mecha-nisms of regulation to PAL are concluded with the helh of the knowledge of enzymology and molecular biology.The hrogresses of the research on PAL in fruit trees are eshecially emhhasized in this haher,and the outlook of research and ahhlication on PAL in fruit trees is also indicated.eey Words PAL;Regulation and exhression;Mechanisms of regulation苯丙氨酸解氨酶(hhenylalanine ammonia-lyase,PAL,E.C4.3.1.5),它催化L-苯丙氨酸解氨生成反式肉桂酸,是苯丙氨酸代谢途径的关键酶。
苯丙氨酸氨解酶基因

淀纯化,溶于双蒸水。
•
按文献描述步骤用T4DNA连接酶将2.5kb rPAL-1-
cDNA与5'端脱磷酸之线性pET-28c连接重组成表达质粒,
连接时取插入片段DNA与pET-28c DNA的摩尔比为3∶1。
连接产物转化感受态大肠杆菌BL21DE3, 在含有卡那霉素
的LB平板上,筛选出阳性克隆。
获取工程菌的过程
谢谢
构必需具备与之相匹配的ATT框架。对比大肠杆菌表达质
粒pET-28a, b, c部分序列,发现在重组后,能保持正确阅
读框架,而不发生移码的只有pET-28c符合要求。若rPAL-
1-cDNA正向插入pET-28c 多克隆位点上的EcoR I位点,
预期重组后的表达质粒部分序列应如图1所示。
获取工程菌的过程
苯丙氨酸氨解酶基因
苯丙氨酸氨解酶简介
• 缩写PAL。是催化直接脱掉L-苯丙氨酸上的氨而生成反式 桂皮酸的酶。EC.4.3.1.5。是1961年J.Koukol, E.Conn在大麦中发现的。存在于高等植物、酵母、菌类 的可溶性部分。推测分子量为30万。这是一个可把苯丙氨 酸用于酚类化合物合成的酶。在很多情况下,其反应成为 酚类化合物合成的有步骤的速率阶段。在组织中的活性可 随外界因素而发生显著变化,用光照,病伤害,植物激素 处理等会使活性显著增加。另外有时还受光敏色素所支配。 在多数情况下,在组织中活性增加的同时,酶发生失活作 用,这时组织中具有活性酶的量很快就会减少,据认为, 这种失活是与类蛋白质物质作用有关。此外也已指出有非 活性的酶的存在。
获取工程菌的过程
• 目的基因的获取
获取工程菌的过程
• 二、重组子的制备及酶切分析:
•
rPAL-1-cDNA大小为2519 bp,pET-28c大小为5367
苯丙氨酸氨基转移酶基因

苯丙氨酸氨基转移酶基因
苯丙氨酸氨基转移酶基因(phenylalanine aminotransferase gene,简称Phenylalanine aminotransferase,缩写为ALT)是一种编码酶的基因,该酶在肝脏和其他组织中起着重要的代谢作用。
ALT 基因编码的酶称为苯丙氨酸氨基转移酶(phenylalanine aminotransferase,简称ALT),也称为谷丙转氨酶(glutamic-pyruvic transaminase)。
该酶主要存在于肝细胞中,是肝脏中最重要的转氨酶之一。
苯丙氨酸氨基转移酶的主要功能是催化苯丙氨酸和α-酮戊二酸之间的氨基转移反应,将苯丙氨酸转化为酪氨酸,并生成谷氨酸。
这个反应是氨基酸代谢中的一个关键步骤,对于维持蛋白质的合成和代谢平衡非常重要。
ALT 基因的表达和活性受到多种因素的调节,包括激素、药物、毒物、疾病等。
当肝细胞受到损伤或发生病变时,ALT 基因的表达会增加,导致血液中ALT 酶的水平升高。
在临床上,检测血液中ALT 酶的水平常用于诊断肝脏疾病,如肝炎、脂肪肝、肝硬化等。
ALT 酶水平的升高通常提示肝细胞的损伤或坏死。
需要注意的是,ALT 基因的表达和活性也可能受到其他因素的影响,如生理状态、饮食、药物等。
因此,在解读ALT 酶水平时,需要综合考虑多种因素,并结合临床表现和其他检查结果进行综合判断。
荸荠苯丙氨酸解氨酶基因的克隆和表达

荸荠苯丙氨酸解氨酶基因的克隆和表达SONG Mubo;SHUAI Liang;DUAN Zhenhua;QIN Li;LI Lin;CHENZhenlin;FANG Fang【摘要】[目的]克隆荸荠Eleocharis tuberosa苯丙氨酸解氨酶基因(PAL),分析其序列特征及其在荸荠不同组织中和鲜切荸荠贮藏过程中的表达情况,为揭示鲜切荸荠黄化机理提供理论依据.[方法]通过RT-PCR和RACE技术从荸荠中克隆PAL基因的cDNA全长,采用生物信息学方法对其序列和所编码的蛋白进行预测分析,利用荧光定量PCR技术分析PAL基因在荸荠不同组织和鲜切荸荠贮藏过程中的表达情况.[结果]克隆得到荸荠PAL基因全长cDNA,将其命名为CwPAL,该序列长度为2485 bp,含有1个2142 bp的完整开放阅读框,共编码713个氨基酸.CwPAL蛋白分子式为C3437H5514N944O1058S34,相对分子质量为78079,等电点为5.97,原子总数为10987个.CwPAL包含PAL-HAL和PLN02457结构域及典型的PAL 酶活性中心序列(GTITASGDLVPLSYIAG).CwPAL的二级结构以α-螺旋为主,其三维结构模型呈典型的\"海马状\"结构.系统进化分析表明,CwPAL与菠萝Ananas comosus和海枣Phoenix dactylifera的PAL蛋白亲缘关系较近.荧光定量PCR 分析表明,CwPAL基因在荸荠皮中的表达量最高,鲜切荸荠贮藏过程中CwPAL基因表达量快速上升,水杨酸处理显著抑制了CwPAL基因的表达.[结论]CwPAL属于典型的苯丙氨酸解氨酶家族,该基因可能通过调控苯丙烷代谢从而影响鲜切荸荠的黄化.【期刊名称】《华南农业大学学报》【年(卷),期】2019(040)001【总页数】7页(P77-83)【关键词】荸荠;苯丙氨酸解氨酶;基因克隆;表达分析;黄化;水杨酸【作者】SONG Mubo;SHUAI Liang;DUAN Zhenhua;QIN Li;LI Lin;CHEN Zhenlin;FANG Fang【作者单位】;;;;;;【正文语种】中文【中图分类】S645.3;TS255.36荸荠Eleocharis tuberosa别名马蹄、地栗、乌芋等,是非常受欢迎的莎草科水生蔬菜[1]。
从基因工程乳酸菌中高效提取苯丙氨酸解氨酶的方法研究

生物技术通报BIOTECHNOLOGYBULLETIN・研究报告・2008年第3期收稿日期:2007-11-28作者简介:陈星(1973-),实习研究员,研究方向:基因诊断与治疗,E-mail:xing18785@yahoo.com.cn苯丙氨酸解氨酶(PhenylalanineAmmoniaLyase,PAlE.C.4.3.1.5)是高等植物苯丙酸类化合物代谢过程中的一个关键酶,可催化苯丙氨酸脱氨生成肉桂酸和氨,已成为当前生物化工领域研究与开发的热点[1]。
有研究表明,该酶在体内外可有效降低苯丙氨酸浓度,对苯丙酮尿症及某些底物依赖性肿瘤有治疗作用,在临床治疗领域亦有良好的应用前景[2 ̄5]。
目前,PAL酶虽可从水稻、小麦等植物中分离纯化,但工艺复杂、费用高,且天然来源的PAL性质不稳定,生物半衰期短,难以适应大规模工业化生产的需求。
乳酸菌是一种重要的食品级微生物,它具有生长速度适中,易于培养,抗原性弱等特点,工业中可作为用于生产肽、酶或代谢物的宿主菌。
乳酸菌基因表达系统构建已经成为许多人的研究课题,尤其是食品级的高效表达系统的构建和应用更是近年来的研究热点[6]。
利用国际上最先进的乳酸乳球菌NICE表达系统,并使用偏爱密码子成功获得了高效表达PAL的食品级基因工程菌NZ3900/pNZ8149-palart[7]。
乳酸乳球菌菌壁较厚且结构致密,传统的超声破碎和强度较大的细胞裂解液法难以在充分破碎菌体的同时保持外源蛋白的功能和结构完整。
因此,基因工程乳酸菌外源蛋白的提取、纯化成为研究和应用中的难点。
结合物理、化学和生物等方法,探索出一条从基因工程乳酸菌中简便、有效的提纯苯丙氨酸脱氨酶(PAL)的技术路线。
实验证明完全能够适应体外细胞学试验的要求,为基因工程乳酸菌的应用和PAL酶的来源等相关研究开辟了新途径。
从基因工程乳酸菌中高效提取苯丙氨酸解氨酶的方法研究陈星1王璞2姚滢秋2肖白1(1首都医科大学附属北京朝阳医院基础医学研究中心,北京100020;2首都师范大学生命科学学院,北京100037)摘要:为从高表达苯丙氨酸解氨酶(PAL)的基因工程乳酸乳球菌NZ3900/pNZ8149-palart中有效提取所表达的PAL酶,针对乳酸乳球菌的特点,结合各种物理、化学、生物学等方法,探索出一条简便、高效的技术路线。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
苯丙氨酸氨解酶基因 在大肠杆菌 BL21DE3中的表达
苯丙氨酸氨解酶简介
• 缩写PAL。是催化直接脱掉L-苯丙氨酸上的氨而生成反式 桂皮酸的酶。EC.4.3.1.5。是1961年J.Koukol, E.Conn在大麦中发现的。存在于高等植物、酵母、菌类 的可溶性部分。推测分子量为30万。这是一个可把苯丙氨 酸用于酚类化合物合成的酶。在很多情况下,其反应成为 酚类化合物合成的有步骤的速率阶段。在组织中的活性可 随外界因素而发生显著变化,用光照,病伤害,植物激素 处理等会使活性显著增加。另外有时还受光敏色素所支配。 在多数情况下,在组织中活性增加的同时,酶发生失活作 用,这时组织中具有活性酶的量很快就会减少,据认为, 这种失活是与类蛋白质物质作用有关。此外也已指出有非 活性的酶的存在。
获取工程菌的过程
结果分析
• 在BL21DE3宿主菌中,它既能应用异丙基硫代-β-D-半乳 糖苷(IPTG)使lac阻遏蛋白失活,从而使T7启动子脱阻遏 并诱导T7聚合酶,提高表达目的基因水平,同时,由于宿 主菌不表达OmpT和Lon蛋白酶,因此表达产物又比较稳 定,不会被宿主菌蛋白水解酶所降解。我们建株的工程菌, 经超声破碎后的粗提物,按Zucker建立的苯丙氨酸氨解酶 活性测定方法,测得的酶比活性为461.7 nmol.g-1.min-1。 该质粒还具有表达His6-Tag结构,便于应用Ni柱亲和层析, 快速纯化,经凝血酶酶切,有可能获得具有活性的天然 PAL。为今后PAL的基础研究(如高等植物中的UV反应, 创伤防御,氧化应激,凋亡等)及临床治疗(如苯丙酮尿 症)奠定了基础。
获取工程菌的过程
• 根据rPAL-1-cDNA及pET-28c酶切图谱,选用BamH I酶切 重组质粒,鉴定插入方向。并用测序引物对重组质粒 PET-28c-rPAL-1-cDNA进行测序,进一步确定其插入方 向及阅读框架。
获取工程菌的过程
• 工程菌的诱导表达及SDS-聚丙酰胺凝胶电泳分析: • 挑取正向插入克隆,接种于含50 μg/mL卡那霉素的 LB培养基中,37℃振荡培养16 h,以1∶200比例接种于新 鲜的含抗生素的培养基中, 37℃振荡培养2.5 h进行扩增,取 100 mL菌液作为诱导前对照。然后加入终浓度为1 mmol/L的异丙基硫代-β-D-半乳糖苷(IPTG),在37℃振荡 培养中诱导工程菌表达,分别于诱导1,3,5,7 h四时间点取样, 以BL21DE3及经pET-28c空载体转化的BL21DE3为对照, 在超声裂解液中将大肠杆菌超声破碎,200 W 10 s,停10 s,重复10次。4 ℃ 15 000 r/min离心15 min,上清液和 沉淀分别进行10% SDS-聚丙酰胺凝胶电泳(SDS-PAGE), 200 V,30 min,考马斯亮兰染色,结果经Kodak digital scienceTM电泳扫描仪分析确定其分子量及蛋白表达量。
获取工程菌的过程
• 筛选与鉴定 • 用EcoR I酶将pET-28c从多克隆位点处切开,在其5' 端用牛小肠碱性磷酸酶(CIP)脱去磷酸,纯化后溶于双 蒸水中备用。 • 用EcoR I酶切质粒pUC19-rPAL-1-cDNA,通过1.5% 琼脂糖凝胶电泳分离(80V 4 h),切取2.5kb的rPAL-1cDNA条带,用透析袋法回收,经酚、氯仿抽提,乙醇沉 淀纯化,溶于双蒸水。 • 按文献描述步骤用T4DNA连接酶将2.5kb rPAL-1cDNA与5'端脱磷酸之线性pET-28c连接重组成表达质粒, 连接时取插入片段DNA与pET-28c DNA的摩尔比为3∶1。 连接产物转化感受态大肠杆菌BL21DE3, 在含有卡那霉素 的LB平板上,筛选出阳性克隆。
获取工程菌的过程
• 目的基因的获取
获取工程菌的过程
• 二、重组子的制备及酶切分析: • rPAL-1-cDNA大小为2519 bp,pET-28c大小为5367 bp,通过T4 DNA连接酶连接后的重组表达质粒应为7886 bp。挑取四个重组阳性克隆,经EcoR I酶切图谱鉴定,所 得片段符合预计大小(见图2,lane2, 4, 6, 8)。由于应 用单酶切插入目的基因所获得的重组质粒,存在着正反两 种插入方向。为了获得所需的正向插入重组克隆,经查阅 GenBank, 在对rPAL-1-cDNA及pET-28c质粒的酶切图谱 分析后,选用各自都只有一个酶切位点并能显著区分正反 插入方向的BamH I进行酶切鉴定。根据理论分析,经 BamH I酶切后可能出现的二种情况如下:正向插入生成 的二个片段大小预计应为337bp和7549 bp,反向插入分 别为2194 bp和5692 bp(见图3)。四个重组克隆的 BamH I酶切图谱见图2,lane 1, 3, 5, 7。由此可知,获得 了3个正向插入重组克隆和1个反向插入重组克隆。
• 苯丙氨酸氨解酶(L-phenylalanine ammonia lyase, PAL)广 泛分布于植物和某些真菌中,它可使苯丙氨酸降解为无毒 的苯丙烯酸。在动物实验中证明,口服该酶可降低实验性 苯丙酮尿症动物血清中异常增高的苯丙氨酸浓度,已有学 者研究将该酶固化后,口服用于治疗苯丙酮尿症。尽管 PAL在临床治疗应用中具有良好的前景,但无法从真菌和 高等植物细胞中大量提取PAL,为此,试图应用基因工程 技术探索从大肠杆菌中获取大量廉价的PAL,藉以通过口 服在肠道内消除苯丙氨酸,或用于生产廉价的无苯丙氨酸 食物,达到治疗苯丙酮尿症的目的。
获取工程菌的过程
• • 三、重组质粒测序: 重组表达质粒测序结果如图4所示,与设计时预期的 完全相同。说明该重组质粒已正向插入目的基因,对阅读 框架分析,完全肯定了该质粒能正确表达rPAL-1-cDNA。
获取工程菌的过程
• 重组子的转化 • 按文献,将质粒pUC19-rPAL-1-cDNA转化大肠杆菌DH5α, 并加以扩增。根据文献在水稻苯丙氨酸氨解酶基因 (GenBank/EMBL, Accession number: x16099)ATG下 游131-112碱基段设计测序引物5' TGGATCTTGACCAGCGGCT 3',对pUC19-rPAL-1cDNA质粒进行反向测序。测序按Perkin Elmer公司推荐 的BigDye标记终止碱基双脱氧法在ABI PrismTM310自动 序列分析仪上进行。根据测序结果,在表达质粒pET-28 系列中,选择与之相匹配的表达载体。
结果
• 表达质粒pET-28系列的选择: • 应用测序引物对质粒pUC19-rPAL-1-cDNA进行部分 测序,结果见图1。从rPAL-1-cDNA序列中首先找出翻译 起始密码ATG,根据3个碱基编码1个氨基酸的原则,由此 往上推算可知,要求表达载体在EcoR I接头处的三联体结 构必需具备与之相匹配的ATT框架。对比大肠杆菌表达质 粒pET-28a, b, c部分序列,发现在重组后,能保持正确阅 读框架,而不发生移码的只有pET-28c符合要求。若 rPAL-1-cDNA正向插入pET-28c 多克隆位点上的EcoR I位 点,预期重组后的表达质粒部分序列应如图1所示。
预期目标
• 遗传性缺乏苯丙氨酸羟化酶所致的苯丙酮尿症是 人类常见的先天性苯丙氨酸代谢缺陷病,是引起 先天性智力降低的主要原因之一。为防止痴呆的 发生,在出生后至发育期必需服用不含或含极少 量苯丙氨酸的合成食物,费用十பைடு நூலகம்昂贵。用酶在 肠道内/外将消化后的苯丙氨酸降解,是一个有希 望的取代途径。
预期目标
获取工程菌的过程
• 10% SDS-聚丙烯酰胺凝胶电泳及扫描分析: • 以大肠杆菌BL21DE3和经pET-28c质粒转化的 BL21DE3为对照,与经pET-28c-rPAL-1-cDNA 转化的 BL21DE3在同等条件下按照方法六项中描述的方法进行 扩增、诱导表达、裂解和SDS-聚丙烯酰胺凝胶电泳分析, 结果如图5。工程菌在分子量66.2 kD和97.4 kD间,出现 明显的表达条带,表达的目的蛋白主要存在于沉淀中(以 包涵体形式存在),而对照组BL21DE3及经pET-28c质粒 转化的BL21DE3都没有出现。IPTG诱导5和7 h时蛋白表 达量最高,表达产物His-rPAL的分子量为78.6 kD,与文 献报导一致。IPTG诱导1,3,5,7 h蛋白表达量占菌体总蛋白 量的百分数分别为21.40%, 30.60%, 35.40%, 35.43%。