果蝇胚胎早期
果蝇心脏发育研究三十年进展

果蝇心脏发育研究三十年进展袁婺洲;谢玉莲;吴秀山;邓婷智【期刊名称】《生命科学研究》【年(卷),期】2024(28)1【摘要】果蝇(Drosophila melanogaster)作为最早用于研究心脏发育基因调控的模式生物,已经走过三十年的历程。
果蝇心脏发育过程经历了胚胎期、幼虫期和成虫期三大阶段。
在胚胎早期,Tinman、Dorsocross和Pannier等基因是关键的调控因子。
Tinman参与最早的心脏前体细胞分化和心脏细胞形成,而Dorsocross 和Pannier则影响心脏前体细胞的定向分化和心脏管腔的形成。
进入胚胎晚期和幼虫期,果蝇的心管经历进一步的发展和重塑,该过程主要受到转录因子Hand、Mef2以及Hox基因家族的调控。
在成虫期,Hox基因家族和Tinman依旧发挥重要作用。
虽然果蝇心脏与脊椎动物成熟心脏存在形态上的差异,但两者心脏的早期发育过程以及调控基因和信号通路都有保守性。
本文综述了果蝇心脏发育基因调控研究的三十年进展以及利用果蝇模型研究人类心脏相关疾病的潜在希望。
【总页数】12页(P1-11)【作者】袁婺洲;谢玉莲;吴秀山;邓婷智【作者单位】湖南师范大学生命科学学院心脏发育研究室;湖南省人民医院(湖南师范大学附属第一医院)【正文语种】中文【中图分类】Q344;Q46【相关文献】1.控制果蝇心脏早期发育的基因研究进展2.利用果蝇模型研究人类心脏早期发育的分子机理(英文)3.利用RNAi技术研究果蝇心脏发育基因的功能4.果蝇心脏发育基因调控的研究进展5.利用模式生物果蝇研究人类心脏发育的基因调控因版权原因,仅展示原文概要,查看原文内容请购买。
遗传学(第3版)第16章 发育遗传分析

(4)BICOID蛋白抑制caudal mRNA的翻译,NANOS蛋白抑 制hunchback mRNA的翻译 (5)相应地,hunchback mRNA在胚胎前部得到翻译, caudal mRNA在胚胎后部得到 翻译 (6)HUNCHBACK与BICOID蛋 白作为转录因子调控负责胚胎 前部分化的基因的表达, CAUDAL蛋白作为转录因子调 控负责胚胎后部分化的基因的 表达
16.1 遗传与发育的关系
16.1.1 遗传属性决定性状发育 发育遗传学(developmental 例如:果蝇的发育模式 genetics):是研究基因对个体发 育的调控作用的学科。 个体发育是在时间和空间尺度 上对个体的基因组进行逐步的、 程序性的解读过程。 每一个经有性生殖而来的有机 体的遗传组成都是独特的——个性 化特征(individuality);但是, 每一个个体又同时带有鲜明的物 种特征,即同一个物种内所有个 体共有的形态与躯体结构模式。
16.2 果蝇早期胚胎极性的决定与形态发生素
16.2.1 果蝇胚胎发育的起始与形态发生素
果蝇胚胎极性的形成
发育起始,沿着受精卵两条轴即前 后轴(A/p,anterior-posterior)和背
腹轴CD/V,dorsal-vontsal)形成梯度。
这是由母体基因表达的结果。
果蝇胚胎的极性是通过一系列的基因分层次表达调控的级联反应来
(2)胚轴建立与图式形成
胚胎细胞形成不同的组织、器官,构成有序空间结构的过程称为图 式形成(pattern formation)。 在动物胚胎发育中,早期的图式形成主要涉及胚轴形成(formation of embryonic axes)、体节形成(segmentation)、枝芽和器官 原基(anlage)形成及其一系列与躯体模式的建立相关的细胞分化过程 。躯体模式的建立是在一系列基因多层次、网络性的调控下完成的。
果蝇的胚胎发育

目前对果蝇胚胎早期发育机制已基本了解, 胚胎的前 – 后轴和背 – 腹轴分别独立地由 母体效应基因产物决定。这些母体效应基 因主要编码转录因子,它们的产物通常形 成一种浓度梯度并产生特异的位置信息, 以进一步激活一系列合子基因的表达。随 着这些基因的表达,胚胎被分成不同的区 域。形式。最后每一体节通过HOM-C基因的 特异性表达而确定其特征。
果蝇沿前 后轴、背 腹轴和中 侧轴建立 形体模式。
果蝇早期胚轴形成涉及一个由母体效应基 因产物构成的位置信息网络。在这个网络 中,一定浓度的特异性母源性RNA和蛋白 质沿前 – 后轴和背 – 腹轴的不同区域分布, 以激活胚胎的合子基因组的程序。 有4组母体效应基因与果蝇胚轴形成有关, 其中3组与胚胎前 – 后轴的决定有关,另一 组基因决定胚胎的背腹轴。
二、果蝇前 – 后轴的形成
1. 果蝇前后极性的产生
果蝇的胚胎,幼虫、成体的前后极性均来 源于卵子的极性。 对于调节胚胎前 – 后轴的形成有4个非常重 要的形态发生素:BICOID(BCD)和 HUNCHBACK(HB)调节胚胎前端结构的形 成,NANOS(NOS)和CAUDAL(CDL)调节胚 胎后端结构的形成。
缺口基因(gap gene)的表达区域为一些较 宽的区域,每个区域的宽度约相当于3个体 节,表达区之间可有部分重叠。当缺口基 因突变时胚胎缺失相应的区域。缺口基因 直接受母体效应基因的调控。 缺口基因最初通常在整个胚胎中都有较弱的 表达,然后随着卵裂的进行逐渐变成一些 不连续的区域。缺口基因的表达最初由母 体效应基因启动,其表达图式的维持可能 依赖于缺口基因之间的相互作用。
在动物胚胎发育中,最初的图式形成主要 涉及胚轴(embryonic axes)形成及其一系 列相关的细胞分化过程。胚轴指胚胎的前 -后轴(anterior -posterior axes)和背 – 腹 轴(dorsal -ventral axis)。 胚轴的形成是在一系列基因的多层次、网 络性调控下完成的。
果蝇体轴形成的分子机制果蝇的卵、胚胎、幼虫、成虫都有明确的前后和

果蝇体轴形成的分子机制果蝇的卵、胚胎、幼虫、成虫都有明确的前后和背腹轴,在果蝇最初的发育中,由母源效应基因及其编码蛋白构成位置信息的基本网络,激活合子基因的表达,控制果蝇躯体模式的建立。
1、果蝇胚胎的极性果蝇早期胚轴形成设计由母源效应产物构成的位置信息网络,其中有3组与前-后轴形成有关,为前端系统;1组决定胚胎的背腹轴,即后端系统;还有一组决定背腹轴形成。
2、果蝇前-后轴的形成在果蝇前-后轴的形成中,有四个非常重要的形态发生素:Biociod(BCD)和Hunchback(HB)调节胚胎前端结构的形成;Nanos(NOS)和Candal(CDL)调节胚胎后端结构的形成。
1)前端组织中心: BCD蛋白浓度梯度前端系统至少包括4个主要的基因,其中起关键作用的是BCD,bcd是一种母源效应基因,在卵子发生时,bcd mRNA于滋养细胞中转录,再转运至卵子中并定位于卵子前极。
受精后迅速翻译,BCD具有决定胚胎极性和组织空间图示的功能。
受精后BCD蛋白在前端积累并向后端弥散,形成从前向后稳定的浓度梯度,主要覆盖胚胎前2/3区域。
bcd 基因也是同源异型框基因,BCD蛋白是一种转录调节因子,可与DNA特异性结合并激活合子靶基因的表达。
BCD蛋白浓度梯度可以同时特意新启动不同基因的表达,从而将胚胎划分为不同的区域。
2)后端组织中心:Nanos蛋白和Candal蛋白浓度梯度后端系统在控制图式形成中起到作用与前端系统相似。
决定胚胎后端的最初信息也是母源效应基因转录产物,在卵子发生过程中,后端决定子Nanos(NOS)的mRNA在卵室前端的滋养细胞中转录,通过转运定位到卵子后极,在成熟卵中定位于生殖质。
后端系统是通过抑制转录因子HB的翻译起作用,该系统包括约10个基因,这些基因都是腹部图示形成所必须的基因。
NOS活性从后端向前弥散形成浓度梯度,在胚胎后部抑制hb mRNA的翻译。
HB蛋白的分布区域主要位于胚胎前半部分。
5第五章 果蝇胚轴形成

果蝇形体模式的分子机制:发育生物学研究的重大成
果之一
2
胚轴指胚胎的前后轴(anterior - posterior axis) 和背腹轴(dorsal- ventral axis)。所有多细胞机体至 少具有一种主要的胚轴。两侧对称的动物还具有中 侧轴(mediolateral axis)或左右轴(left-right axis)。
具有正常形态特征的一类关系密切的基因。这些基因 都含有一段高度保守的180 bp 的DNA片段,该片段称 为同源异型框 (homebox)。
功能:它将决定产生哪一类体节(前胸部、中胸部或 后胸部)。在每一个体节确定后,调控进一步的结构 特化。
35
在果蝇中,大多数同源异型基因是在第3号染色 体上,排成两簇。一簇叫触角足复合体(Antp-
22
表达特点:
(1). 都编码转录因子;
(2). 都在合胞胚期开始 表达; (3). 其产物的半衰期一 般较短,仅数分钟,因 而它们的扩散距离较短; (4). 其表达局限在一定 的区域,其突变会导致 胚胎在该区域及附近区 域的缺失。
lose of Kruppel
23
功能:
1. 彼此调节确定表达界限;
末端系统:Torso信号途径,决定胚胎两端不分节的原头区和尾 节
背腹轴
TOLL降解CS :Dorsal(dl)-DL在细胞核中
腹部
DL和CS结合,Toll无活性: Dorsal(dl)-DL在细胞质中
背部:
二、分节基因和胚胎体节的形成(研究 缺失突变体):
缺口基因
成对控制基因
体节极性基因
同源异型基因
13
果蝇的发育过程及其调控机制

果蝇的发育过程及其调控机制果蝇作为模式生物,其发育过程及调控机制已经被广泛研究。
果蝇的发育过程主要包括卵母形成、卵的受精、胚胎发育、幼虫期和蛹期等阶段。
这些阶段都受到不同的调控机制影响。
一、卵母形成和受精卵母形成过程在果蝇体内发生,一般从卵巢的端部开始,向基部发展。
在卵母形成早期,睾丸激素和卵泡刺激素对生殖细胞的发育起着重要作用。
随着卵母不断生长发育,卵母细胞核的复制和分裂也不断进行,最终形成完整的卵母。
成熟的卵母受到雄性精子的受精作用,形成受精卵。
在受精卵中,父本和母本细胞贡献的基因发挥不同的作用,影响胚胎的发育命运和表型表达。
二、胚胎发育受精卵受到一系列的调控因素作用,经过不同的胚胎发育阶段,最终形成成熟的幼虫体。
胚胎发育过程受到多个信号通路和基因网络的调控。
在果蝇的胚胎发育中,Wnt信号通路通过不同的基因调节细胞分裂和定向移动,对胚胎的头尾轴和背腹轴形成起着重要作用。
Hedgehog信号通路则调节胚胎前期的背腹轴形成。
同时,一个非常重要的因子是胰岛素样生长因子信号通路,其对幼虫的大小和发育有关键调节作用。
胚胎发育过程中也涉及到了一些胚胎基因调控系统,如TGFβ/Activin和Notch信号通路,调节着胚胎的细胞命运和分化。
三、幼虫期和蛹期在幼虫期和蛹期,果蝇表现出不同的形态和功能,同时也涉及到不同的代谢途径和调控机制。
在幼虫期,果蝇需要做出重要的生长和分化决策。
通过对食物和营养的感知和代谢途径的调控,果蝇可以根据生长的需要积累能量和物质,维持其生命活动的正常进行。
例如调节能量代谢的AMPK和Sirt1,以及调节食欲和代谢途径的Insulin/IGF1和TOR信号通路,都对幼虫的生长分化起着重要作用。
蛹期是果蝇生命周期中的一个转折点,幼虫期积累的营养和物质被重新配置为成虫身体的各种组织和器官。
在成虫器官分化和重构的过程中,也涉及到不同的调控机制。
例如调节上皮细胞形成和器官增殖的Notch信号通路,调节生殖腺发育的JAK/STAT信号通路,以及影响翅膀和大脑发育的Wnt和Hedgehog信号通路,都对蛹期的发育起着至关重要的作用。
果蝇胚胎发育与前-后轴

• 在受精卵的中央,细胞核进行多次有丝分裂。果蝇受精卵
平均每8分钟一次核分裂,形成多大256个细胞核。之后细 胞核移动到受精卵的外围,继续进行有丝分裂。
• 在第九个分裂周期中,大约五个细胞核到达胚胎的后极表
面,开始被细胞膜包围,这些极细胞将来形成成虫的配子。
• 大多数其他的细胞核在第十分裂周期到达胚胎的外围,之 后经历减慢的四个分裂周期。细胞核经过这些分裂阶段,
• 尽管所有的核公用一个细胞质,并不是意味着每个细胞核
周围的细胞质是一样的。每一个核周围都包含细胞骨架蛋
白。在第十个周期,各个细胞核到达卵的外围时,每一个
受精卵周围都包围着微管和微丝。细胞核和它们周围的细
胞质被称作活质体(energids)
• Although the nuclei divide within a common cytoplasm, this does not mean that the cytoplasm is itself uniform. Karr and Alberts (1986) have shown that each nucleus within the syncytial blastoderm is contained within its own little territory of cytoskeletal proteins. When the nuclei reach the periphery of the egg during the tenth cleavage cycle, each nucleus becomes surrounded by microtubules and microfilaments. The nuclei and their associated cytoplasmic islands are called energids
果蝇细胞周期蛋白Cyclin D在胚胎发育不同时期的表达

激 活 G1 时期 特 有 的周 期 蛋 白依 赖性 激 酶 C K4 磷 D , 酸 化 Rb 磷 酸化 的 Rb蛋 白从 其 所 结 合 的 E F转 录 , 2
A u . 2 07 g 0
20 0 7年 8月
果 蝇 细 胞 周 期 蛋 白 C ci 在 胚 胎 yl D n 发 育 不 同 时 期 的 表 达
石 松 婷
( 华 大 学 生 物 科 学 与 技 术 系 , 京 10 8 ) 清 北 0 0 4
摘要 : 细胞周期蛋 白 C cn ( yD 是 发育中重要的蛋白, yl C c ) iD 作者利用 原位杂交 法和免疫 组化法 , 别对野生 型的果蝇 分
C AF, 亦可 以结合 转 录 因子 T FⅡD 5等 , 过和 这些 [ 通
转 录因子 的结合 , y D1 到促 进基 因转 录的功 能. Cc 起
研 究 中最重要 的模式 生物 之一 [ . 期 经 典 遗传 学 理 1早 ]
论 的建 立就 得益于对 果蝇 的观 察研 究 . 进 入后 基 因 在 组时代 的今 天 , 随着 果 蝇 全 基 因组 序 列 的 测 定 完 成 ,
导致 翅原基 中产 生过 量 的细胞 [ . 今为止 , 7迄 ] 关于 C — y
以及 日渐成 熟 的果 蝇 实 验 技术 , 蝇 在 发 育 生 物 学 , 果
神经 生物学 等 当今诸 多 热点 生 命 科 学 领 域 的研 究 中
起着 相 当重要 的作用 .
细胞周 期 (el y l) 细 胞 生命 活 动 的 基 本 过 cl c c 是 e
关 键 词 : yD 原位杂交 ; cc ; 免疫组化 ; 果蝇
中 图分 类号 : 4 Q32
果蝇胚胎发育和体轴形成的分子机制

果蝇胚胎发育和体轴形成的分子机制果蝇是一种十分常见的昆虫,它的胚胎期相当短暂,只需要不到两天的时间就能完成发育。
在这个过程中,许多重要的分子和信号通路都在起作用,对于了解发育和形态建成的分子机制具有重要意义。
本文将着重讨论果蝇胚胎发育和体轴形成的分子机制。
一、胚胎发育的基本过程果蝇的生命周期很短,在一般的实验室条件下,一只果蝇的寿命约为两个月。
雌性果蝇通常会在食物和水的混合物中产卵,大约在24小时内就会孵化。
在这之后的几天中,果蝇的发育非常迅速,逐渐从卵到幼虫,然后到成虫。
而在这个发育过程中,果蝇胚胎发育是非常关键的一步。
胚胎发育的过程可以分为五个连续的阶段。
第一阶段为卵细胞核形成,包括以交配卵产生的核和父母亲体细胞内孤雌生殖卵产生的核。
第二阶段是卵剖面形成,包括老二系统等。
通过定期锯切进行紧密环绕,则表明胚胎发展正常。
长时间僵滞不进则可能会在胚胎发展的后期出现显著的异常。
第三阶段是胚胎心脏形成,包括神经脑、背板神经织物、翼的最初生长和原胚内的胚芽形成。
第四阶段胚胎形成,则大半是身体器官、肌肉、皮肤等的形成。
卵子在此时分化为头部,胸部和腹部。
第五阶段是成虫形成,也就是最后的阶段。
它涉及更广泛的组织分化,包括瓢虫和蜜蜂等其他昆虫几乎都会经历的阶段。
以上这些阶段综合起来,构成了果蝇胚胎发育过程中的基本过程。
接下来我们将要探究果蝇胚胎形态的建立和维持所涉及的分子机制。
二、体轴形成的分子机制在果蝇发育的过程当中,体轴形成似乎是整个过程中最显著的阶段之一。
而体轴形成涉及到许多基因和信号通路的调控。
1. Wnt信号通路Wnt信号通路是胚胎发育中非常重要的一个通路。
Wnt分子在体轴形成中具有特殊作用,因为它们能够影响Dorsal基因的表达。
Dorsal基因在果蝇胚胎发育中发挥着严格的调控作用,在形成体胚层时非常重要。
Wnt拮抗剂能够使得胚胎中Dorsal基因表达出现异常,从而导致体轴形成不良。
2. Hedgehog信号通路Hedgehog信号通路也是在胚胎发育中重要的信号通路。
果蝇的发育和成虫盘解剖
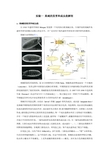
实验一果蝇的发育和成虫盘解剖1、果蝇胚胎发育和成虫盘自1910年遗传学鼻祖Morgan发现第一个突变体白眼果蝇以来,生物学家将果蝇作为遗传学研究的模式动物已经近百年,并广泛应用于现代遗传学和发育生物学研究的模型。
果蝇生命周期如图1。
果蝇胚胎发育速度快,前13次卵裂每次只间隔9min,细胞核成倍增加成为一个合胞体(syncytim),发育过程中的胚胎为观察分析卵裂、早期胚胎发生和躯体模式形成等发育调控机制的提供了很好的材料。
果蝇胚胎发育的梯度假说被证实,在1997年和1998年连续两年被《Science》杂志评为当年十大突破成就之一。
现已鉴定出在一些卵子中形成梯度、调节细胞定位和分化并决定胚胎发育方式的形态发生素(morphogen)。
果蝇存在变态过程,由幼虫(larval)经蛹(pupal)转变为成虫。
成虫盘(imaginal discs)是果蝇早期胚胎发育期间保留下来的而在幼虫期开始分化的。
变态期间,幼虫表皮转成蛹壳而成虫盘则发育成为成虫外层。
成虫盘首先出现在20~40细胞群的胚胎中,由胚胎上皮内陷形成,最初通过各种标记基因的表达而被确认。
每一个成虫盘都由单层上皮构成片状或囊状,并有一个细茎与胚胎和幼虫的上皮连接。
最终每一个液囊展平,液囊的两端具有不同的特征,并执行不同的发育任务。
一端形成较厚而高度折叠的成虫盘上皮,另一端形成较薄而展开的周膜。
大部分成虫外胚层结构由成虫盘上皮演化而来。
成虫盘在一、二、三龄幼虫期都有不同程度的细胞增殖,在晚期三龄幼虫后,即变态之前,每个成虫盘形成了数万个细胞。
在变态之前,大约产卵后108h(AEL),25℃放置,三龄幼虫即蠕动——“爬”出培养基,生活在培养瓶的瓶壁上,这个阶段约12h,在这个阶段末期,果蝇幼虫前端的呼吸孔外翻,幼虫停止蠕动并开始蛹化。
上皮形成蛹的桶状体壁——蛹壳,此时为白色的蛹前期阶段(WPP)。
大约1h后,蛹壳变为深褐色。
蛹化开始的第一个5-6h后,果蝇仍然处于蛹前期阶段,成虫细胞仍然分泌形成蛹的表皮。
果蝇发育基因

果蝇发育基因一、引言果蝇(Drosophila melanogaster)是一种广泛应用于生物学研究的模式生物。
其简单的遗传学和发育学特性使得其成为了研究基因功能和发育过程的理想模型。
果蝇发育过程中涉及到许多基因,其中一些被称为果蝇发育基因。
二、什么是果蝇发育基因果蝇发育基因是指在果蝇胚胎发育过程中起关键作用的基因。
这些基因控制着胚胎从受精卵到成虫的不同阶段的发育过程,包括细胞分裂、细胞分化、器官形成等。
三、果蝇发育基因分类1.早期启动子元件(maternal effect genes)早期启动子元件是由母体提供给受精卵的mRNA和蛋白质。
这些基因控制着受精卵最初几个小时内的细胞分裂和形态变化。
早期启动子元件包括bicoid(bcd)、nanos(nos)等。
2.轴形成基因(zygotic genes)轴形成基因是在受精卵中形成的,控制着胚胎的轴向。
轴形成基因包括dpp、hedgehog(hh)等。
3.分割基因(segment polarity genes)分割基因控制着胚胎体节间的分割。
分割基因包括wingless(wg)、engrailed(en)等。
4.家族基因(homeotic genes)家族基因控制着器官和组织的发育。
家族基因包括Antennapedia、Ultrabithorax等。
四、果蝇发育基因举例1.bicoidbicoid是早期启动子元件中最为重要的一个,它在受精卵前端高浓度表达,控制着头部和前体的形成。
当bicoid缺失时,受精卵无法正常发育,最终导致死亡。
2.hedgehoghedgehog是轴形成基因中的一个,它在受精卵中表达,并控制着腹侧和后部结构的形成。
当hedgehog缺失时,会导致一系列畸形,如腹侧结构缺失等。
3.winglesswingless是分割基因中的一个,它在胚胎体节间表达,并控制着体节间边界的形成。
当wingless缺失时,会导致体节间边界消失,最终导致胚胎死亡。
果蝇的胚胎发育(PPT文档)

3. 后端组织中心:
NANOS蛋白和CAUDAL蛋白浓度梯度
后端系统包括约10个基因,这些基因的突 变都会导致胚胎腹部的缺失。在这一系统
缺口基因、成对控制基因以及体节极性基 因共同调节同源异型基因(homeotic gene) 的表达,决定每个体节的发育命运。
2. 前端组织中心
BICOID(BCD)蛋白浓度梯度
前端系统至少包括4个主要基因,其中bicoid
(bcd)基因对于前端结构的决定起关键的作
用。BCD具有组织和决定胚胎极性与空间图 式的功能。
如果前端和后端系统都失活,果蝇胚胎仍 可产生某些前后图式,形成具有两个尾节 的胚胎。
Torso系统基因的失活会导致胚胎不分节的部分,即前 端原头区和后端尾节,缺失
三、果蝇背 – 腹轴的形成
与果蝇胚轴形成有关的4组母体效应基因中, 背–腹系统最为复杂,涉及约20个基因。其
中dorsal(dl)等基因的突变会导致胚胎背
现已筛选到与胚胎前后轴和背腹轴形成有 关的约50个母体效应基因(maternal effect gene)和120个合子基因(zygotic gene)。 目前,对果蝇胚轴形成的调控机制已有了 一个较为清晰的认识。
在果蝇最初的发育中,由母体效应基因构 建位置信息的基本网络,激活合子基因的 表达,控制果蝇形体模式的建立。
在动物胚胎发育中,最初的图式形成主要 涉及胚轴(embryonic axes)形成及其一系 列相关的细胞分化过程。胚轴指胚胎的前 -后轴(anterior -posterior axes)和背 – 腹 轴(dorsal -ventral axis)。
果蝇发育基因

果蝇发育基因引言果蝇(Drosophila melanogaster)是一种被广泛用于生物研究中的模式生物。
其快速繁殖和易于繁育的特点使得科学家们可以对果蝇进行大规模的遗传实验。
通过研究果蝇的发育基因,我们可以深入了解发育过程中的分子机制,并对生物体的发育和变异有更全面的理解。
果蝇的发育果蝇的发育过程包括胚胎发育、幼虫发育和成虫发育三个阶段。
这些不同的发育阶段由一系列基因调控。
在整个发育过程中,果蝇的外部形态和内部器官都会发生显著的变化。
胚胎发育果蝇的胚胎发育从受精之后开始,持续约24小时。
在胚胎发育过程中,一系列发育基因被启动,并在特定的时期和特定的胚胎区域发挥作用。
这些基因包括转录因子和信号分子,它们相互作用,形成一个复杂的发育基因网络。
该网络调控胚胎细胞的分化、移动和器官形成。
幼虫发育在胚胎发育结束后,果蝇进入幼虫阶段。
幼虫发育分为三个期间,分别是一仲、二仲和三仲。
在这些期间里,果蝇经历吃食、蜕皮和体型增长等过程。
这些过程受到一系列发育基因的调控。
其中一些基因控制幼虫的体型,包括体长和体宽的增加,并在幼虫的节段和器官的形成中发挥重要作用。
成虫发育当幼虫发育到一定阶段,就会进入蛹化阶段,最终成为成虫。
在这个过程中,果蝇的身体组织会完全分化,外部形态和器官逐渐形成。
蛹期持续约10天,成虫在蛹的内部逐渐形成。
成虫发育过程中的发育基因主要调控外部形态、器官的形成和性别的分化。
发育基因的研究方法果蝇发育基因的研究主要依赖于基因突变和遗传分析的方法。
通过对突变体的研究,科学家们可以发现与发育相关的基因,并研究其功能。
同时,利用遗传分析的方法,可以通过交叉杂交和后代分析推断和鉴定新的发育基因。
此外,近年来基因编辑技术的快速发展,如CRISPR-Cas9技术,使得科学家们能够直接修改果蝇基因组,研究特定基因对发育的影响。
这些技术的应用扩展了我们对发育基因的认识,并深入了解发育过程中的细节。
发育基因的功能与调控果蝇发育基因具有多种功能和调控方式。
果蝇的性别决定

果蝇的性别决定 2005-7-7 11:19:37 来源:生命经纬果蝇的性别分化需要很多调控环节,我们先看一下总的途径,然后再介绍细节。
果蝇早期胚胎中性指数(sex index)(X:A)决定了性别的分化。
与性指数相关的性别决定途径是由Thomas Cline Baker(1984,1993)及其同事们建立的的。
在些途径中有主调节基因和系列性别特异基因,它们涉及到RNA的不同剪接,产生不同的雌性和雄性的转录因子。
在此调节途经的第一步是由性指数(X染色体的条数和常染色体组数之比) X:A的比例决定的。
性指数使转录因子具有特殊的浓度。
而这种特殊的浓度充当了主调节基因的开关。
当开关打开时RNA能正常剪切而产生下一步调节的活性转录因子,从而激活雌性特异基因,使胚胎发育为雌体;若开关是关闭的,那么因剪接的方式不同选择性地产生了另一种转录因子,它能激活性特异基因,使胚胎发育为雄体。
在讨论性别决定调节途径的细节之前,我们先来了解一下性别特异突变。
性别转换基因tra(transformer)发生突变使得X:A比率为雌性(X:A=1)的果体发育成雄性表型,但核型并不改变。
双重性别基因dsx(doublesex,)突变使个体变成为间性,而不受性染色体的控制。
还有一种突变会导致一种性别致死,这就是性别致死Sxl(Sex-lethal)基因,其突变有的等位基因是隐性的,可特异导致雌性致死;而另一些等位基因是是显性的,可导致雄性致死。
无女儿da (daughterless,)基因是母体基因,对性别决定具有母体效应,da纯合时母体不产生“女儿”,而“儿子”发育却正常,这是由于其产物对Sx1的早期表达起着正调控作用。
在卵产生时da若无活性,那么在受精后无论X染色体数量如何,都不能激活Sx1,结果所有的子代都向雄性方向发育,但XX个体因X-连锁基因的剂量比XY多了一倍,因剂量不适而死亡。
特大刚毛基因emc (extramacrochaetae)也是母体效应基因,其作用与da相反,是Sx1的负调因子,emc的突变将抑制雄性方向的发育。
4 body plan

Nanos控制hunchback mRNA翻译的机制
A-P轴线形成模式
Hunchback:母体mRNA在卵中均匀分布,受精后前区高浓度的Bicoid蛋白激活合子 hunchback基因的表达,从而帮助形成Hunchback蛋白浓度梯度。
Caudal: 母体mRNA在卵中均匀分布,受精后Bicoid蛋白抑制其在前区的表达,而 Caudal蛋白形成类似于nanos的浓度梯度。
2. Bicoid基因提供A-P轴线形态素梯度
Bicoid编码一 种转录因子。 其突变体缺失 头胸结构,原 头区由尾区取 代。
Bicoid mRNA和蛋白质的分布
在未受精卵中,bicoid mRNA定位在胞质前端;其受 精后翻译出的蛋白质沿AP轴 扩散,形成浓度梯度,为胚胎 的后续分化提供位置信息。
eve
ftz
Pair-rule基因的突变导致胚胎缺失相应的区域
2. Pair-rule
由
不
同 的 调 控 区 控 制
基 因 的 不 同 表 达
横
纹
Pair-rule基因的每个表达横纹由一组Gap转录因子控制
Pair-rule基因表达 的间隔性重复,无 法通过单一浓度梯 度来控制,而是由 多个转录因子来控 制。例如, Evenskipped在第三类体 节中的表达受 bicoid和hunchback的激活,而受 giant和kuppel的抑 制。
哺 乳 动 物 的 基 因
Hox
Engrailed的表达受高浓度fushi tarazu和even-skipped的激活
体节边界的维持
机制:pair-rule基
因的表达时间较短, 不足以维持体节极 性基因的长期表达, 后者必须依赖于另 外的机制。
发育生物学1

绪论发育的核心问题是细胞分化,而导致细胞分化的则是基因的相互作用。
发育是从基因型实现表现型的过程,是基因组内的基因选择性表达的结果。
发育生物学:是应用现代生物学的技术研究生物发育机制的科学。
它主要研究多细胞生物体的从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
胚胎诱导:是指在胚胎发育过程中,相邻细胞或组织间通过相互作用,决定其中一方或双方细胞的分化方向。
诱导作用可使细胞互为不同,胞质决定成分的区域化及细胞的不对称分裂使细胞具有不同的特性。
调整发育:胚胎为保证正常的发育,可以产生胚胎细胞位置的移动和重排,这样的发育为调整发育。
发育生物学的发展基础及过程如何?研究哪些问题?答:发展基础:胚胎学、遗传学、细胞生物学。
发展过程:形态→机理;组织器官→细胞→分子它主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
同时,也研究生物种群系统发生的机制。
生物发育的主要过程和基本规律是什么?答:主要过程:A)细胞分裂:细胞分裂快、没有细胞生长的间歇期,因而新生细胞的体积比母细胞小。
B)图式形成:(1) 躯体轴线的制定;(2) 胚层的形成C)原肠作用:最突出的形态变化发生在原肠作用开始之后。
D)细胞分化:人类胚胎可最后发育出至少250种不同细胞类型,分化通常是不可逆的。
E)细胞生长:胚胎在基本的pattern形成之后,其体积会显著增长,原因在于细胞数量增加、细胞体积增加、胞外物质的积累。
不同组织器官的生长速度也各异。
基本规律:受精→卵裂→原肠胚形成→神经胚形成→器官形成(organogenesis)→幼体发育→生长为成体幼体→成体经历变态发育基因控制细胞行为是通过控制细胞中的蛋白质的产生而实现的, 管家蛋白几乎存在于所有类型细胞中,通常用于产生能量、在代谢途径中生成或降解产物,如组蛋白及转录或翻译中所必需的蛋白因子。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Developmental Biology
Homeotic selector genes 的表达调控
7 Antp Ubx AbdA
Homeotic selector genes 的表达受Gap和Pair-rule 基因的控制。如abdA和 abdB受到Hunchback和 Kuppel的抑制,使腹部 基因不能在头部和胸部表 达。Ubx的激活需要一定 浓度的Hunchback, Antp 由Kuppel激活。
Developmental Biology
体节边界的维持 机制:pair-rule基
因的表达时间较短, 不足以维持体节极 性基因的长期表达, 后者必须依赖于另 外的机制。 高浓度eve或 ftz激活eng基因, 不表达eve和ftz的区 域表达wingless, 其后Eng和Wingless 间的互作使二者的 表达都得以维持, 从而使体节边界得 到巩固。
Bicoid是控制头胸发育的一个关键母体基因,其 不同浓度开启不同合子基因的表达。其它的前 区母体基因主要涉及bicoid mRNA在未受精卵中 的定位及控制其翻译。
Developmental Biology
3. Nanos和Caudal蛋白梯度控制后区结构
nanos
Nanos决定后部区的发育,它在受精后形成P-A浓度梯度,其作用是与 hunchback mRNA结合,阻止后者在后区的翻译,帮助形成Hunchback蛋白梯度。
五、体节极性基因(segment polarity genes)与细胞谱系的建立
体节极性基因是指在pair-rule基因表达之后立即表达的 基因,它们决定了体节的边界和体节内细胞的命运。这些 基因的产物包括扩散分子、受体、转录因子等多种类型。 类体节之间或体节之间均没有细胞的相互迁移。
Developmental Biology Engrailed是确定类体节和体节边界的关键基因,它在每个类体
Developmental Biology
1. A-P轴线由三类母体基因控制:突变鉴定 anterior class, posterior class, terminal class.
Developmental Biology
Developmental Biology
2. Bicoid基因提供A-P轴线形态素梯度
Developmental Biology
Developmental Biology
Homeotic selector genes 突变将 改变其表达区的结构
Ultrabithorax缺失 后,第三胸节(T3) 的平衡棒会象T2 一样长出一对翅膀
Developmental Biology
如果使原来只在T1表达的Antennapedia也在头部表达,则 头窝中长出的不是触角而是腿。
Pair-rule基因表达 的间隔性重复,无 法通过单一浓度梯 度来控制,而是由 多个转录因子来控 制。例如, Evenskipped在第三类 体节中的表达受 bicoid和hunchback的激活,而受 giant和kuppel的抑 制。
Developmental Biology
Developmental Biology
Developmental Biology
卵母细胞A-P极性的确定
Developmental Biology
卵母细胞D-V极性的确定
在受精前腹部的卵泡细 胞合成 Pipe、Windbeutel、 Nudel,定位在腹部卵外膜 上,受精后经一系列反应, 最终使卵周质腹部区的 Dorsal蛋白释放进入核中, 启动腹部特异性基因的表 达。
Developmental Biology
4. 卵膜表面受体的激活决定胚胎AP轴的两个端点
torso突变体缺少原头区和尾区, torso蛋白为酪氨酸激酶类受体。 未受精前,torso已均匀地分布 在卵的质膜上。但其腺体torsolike 定位在两端的卵外膜 (vitelline membrane)上,不能与torso结合。 受精时,torsolike得以释放, torsolike与torso结合, torso活化, 启动信号传导。 Torsolike蛋白的存在量很低, 受精后其扩散距离有限。其突变体 类似torso突变体。
Dorsal
Developmental Biology
2. 背部命运决定于Decapentaplegic (dpp)
dpp编码信号蛋白,给胚胎大量注射dpp mRNA导致外胚 层全部发育为羊浆膜,一定的高浓度dpp使腹部外胚层命运改 变为背部外胚层命运。Dpp浓度梯度形成机制如下图:
Developmental Biology
eve
ftz
Developmental Biology
Pair-rule基因的突变导致胚胎缺失相应的区域
Developmental Biology
2 横 纹 由 不 同 的基 调因 控的 区不 控同 制表 达 . Pair-rule
Developmental Biology
Pair-rule基因的每个表达横纹由一组Gap转录因子控制
Developmental Biology
类 体 节 与 体 节 的 关 系 及 其 发 育 命 运
Developmental Biology
类体节的边界由pair-rule基因活性决定。它们的表达特点包括:
(1). 每个基因只在半数类体节中表达,如even-skipped在奇数类体节(1、3、 5、7、9、11、13)中表达,而fushi tarazu在偶数类体节中表达。表达横 纹一般只有3个细胞宽。 (2). 绝大多数编码转录因子,表达开始于胚胎细胞化前夕。
Developmental Biology
Gap/Pair-rule proteins (repressor)
Developmental Biology
哺 乳 动 物 的 基 因
Hox
Developmental Biology
体节内细胞命运的确定
同一体节内不同区 域的表皮角质结构 有所不同,这可能 是由Hedgehog和 Wingless的梯度分 布控制,或者由它 们作用logy
六、不同体节的发育命运决定于 homeotic selector genes
转基因实验证明hunchback合子基因的表达区域
Developmental Biology
kuppel hunchback
。
则 对 后 者 起 抑 制 作 用
一 定 浓 基度 因的 的 表 达 , 而 高蛋 浓白 度激 的活 hunchback
Developmental Biology
四、Pair-rule基因的表达界定胚胎的类体节
1.类体节(parasegment)由pair-rule基因活性界定
在原肠作用开始后,胚胎表面沿AP轴线出现一些过渡 性的浅沟,将胚胎分为14个区域,这些区域即为类体节。 每个类体节受一套特定的基因的控制,做为独立的发育 单位,将逐渐获得自身特有的特性。 原肠期后,胚胎沿AP轴线出现有规则的节段,即体节 (segments),每个体节有不同的特性及发育命运。 体节是在类体节的基础上形成的,即一个体节是由前一 个类体节的后半部和下一个类体节的前半部组成。
Developmental Biology
三、受精后合子基因的表达
Developmental Biology 1. Dorsal激活腹部合子基因、抑制背部基因
预置中胚层twist和snail 中胚层特异性基因 预置腹部外胚层rhomboid Ventralized embryos: snail和twist基因在所有细胞中都表达,而dpp、 tolloid、zerknullt在所有细胞中都不表达。 Dorsalized embryos: 基因表达状态与ventralized embryos中相反。
节的前部表达,占居一行细胞,从而确定了类体节的前部边界。 体节出现后,它在每个体节的后部表达,将一个体节分成前后两 个区域,两个区域内的细胞不发生交换,各自有不同的发育命运。
Developmental Biology
Developmental Biology
Engrailed的表达受高浓度fushi tarazu和even-skipped的激活
3. Gap基因的表达使胚胎沿AP轴线区域化
Gap基因是指那些在受精后最早沿AP轴线呈区域 性表达的合子基因。 Gap基因的表达特定: (1). 都编码转录因子; (2). 都在多核胚期开始表达; (3). 其产物的半衰期一般较短,仅数分钟,因而它们的 扩散距离较短; (4). 其表达局限在一定的区域,其突变会导致胚胎在该 区域及附近区域的缺失。
Bicoid编码一 种转录因子。 其突变体缺失 头胸结构,原 头区由尾区取 代。
Developmental Biology
Bicoid mRNA和蛋白质的分布
在未受精卵中, bicoid mRNA定 位在胞质前端; 其受精后翻译 出的蛋白质沿 AP轴扩散,形 成浓度梯度, 为胚胎的后续 分化提供位置 信息。
Developmental Biology
5. 卵膜中的母体蛋白决定胚胎D-V极性
Developmental Biology
Dorsal蛋白的V-D浓度梯度的形成
Developmental Biology
6. 卵发生过程中母体基因产物的合成及分布
Developmental Biology
卵子发生过程中某些基因特异性表达
Developmental Biology
Developmental Biology
GAP 基 因 突 变 体
Developmental Biology
母体bicoid蛋白激活合子hunchback的表达,hunchback 蛋白再为其它Gap基因的表达提供位置信息