端粒和端粒酶是如何保护染色体的——2009年诺贝尔生理学或医学奖工作介绍及相关研究进展
关于端粒及端粒酶的调查报告

关于端粒及端粒酶的调查报告一:引言2009年10月5日,诺贝尔生理学或医学奖颁发给了美国科学家伊丽莎白·布莱克本、卡罗尔·格雷德和杰克·绍斯塔克,以表彰他们在研究端粒和端粒酶保护染色体的机理方面的贡献。
这篇调查报告将会通过资料查询和逻辑推断等方式论述关于端粒,端粒酶以及它们与肿瘤细胞的相关内容。
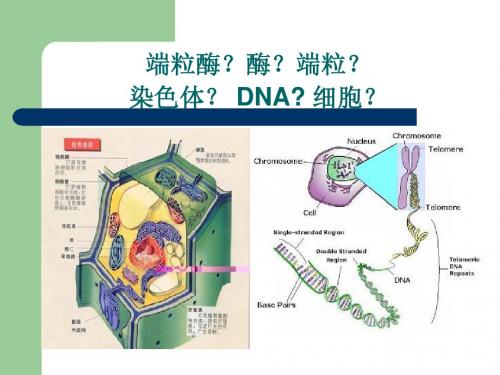
二:端粒和端粒酶2.1.1端粒端粒(Telomere)是染色体末端的重复DNA序列,在人细胞中长度约为几千到一两万碱基对,它防止细胞将天然染色体末端识别为染色体断裂,起着保护和稳定染色体的作用。
[1]随着细胞的分裂增殖,端粒会逐渐缩短。
当端粒的长度缩短到一定程度时,细胞的分裂便会停止。
因此,端粒具有调节细胞增殖的作用,是细胞分裂的“时钟”。
端粒的碱基序列具有极高的保守性,但不同物种的端粒仍有差异,例如:四膜虫重复序列为GGGGTT,草履虫为TTGGGG,人类和哺乳动物为TTAGGG.[2]2.1.2端粒的结构端粒通常由富含G的DNA重复序列,以及端粒结合蛋白和端粒相关蛋白组成。
端粒结合蛋白直接保护端粒DNA,端粒相关蛋白通过与端粒结合蛋白的相互作用间接影响端粒的功能。
端粒既可保护染色体不受核酸酶的破坏,又避免了因DNA黏性末端的裸露而发生的染色体融合。
[4][5]2.2.1端粒酶端粒酶(Telomerase),在细胞中负责端粒的延长的一种酶。
在端粒发现之后,人们便开始猜测存在这样一种酶,可以起到延长端粒的作用——因为随着细胞的分裂增殖和染色体的复制,端粒应当越来越短,但是某些细胞(如肿瘤细胞)的端粒长度却能够保持相对不变。
在1997年,Tom Cech实验室的Lingner在Euplotes aediculatus以及酿酒酵母中发现了真正的端粒酶催化亚基。
[3]2.2.2端粒酶的作用机理端粒酶主要依靠两种成分来实现其功能,一种名为端粒酶逆转录酶(TERT)的蛋白酶,另一种是作为模板的一小段RNA序列。
端粒与端粒酶的研究解读诺贝尔生理学或医学奖

端粒与端粒酶的研究解读诺贝尔生理学或医学奖一、本文概述自人类踏入科学研究的领域以来,生命的奥秘一直是科学家们孜孜不倦探索的主题。
作为生命科学的两大支柱之一,医学领域的研究进展对人类生活的影响深远而持久。
每年的诺贝尔生理学或医学奖揭晓,都会引发全球范围内的广泛关注,因为它不仅代表了该领域最前沿的科学成果,更预示了未来医学可能的突破方向。
本文将以诺贝尔生理学或医学奖为背景,深入探讨端粒与端粒酶的研究,解读这一重大科学奖项背后的科学意义和影响。
端粒和端粒酶是生物学中的两个重要概念,它们在细胞生物学、分子生物学和遗传学等领域有着广泛的应用。
端粒是真核生物染色体末端的特殊结构,具有保护染色体末端、防止染色体融合和降解的重要作用。
而端粒酶则是一种特殊的逆转录酶,能够合成端粒DNA,从而维持端粒的长度和稳定性。
近年来,随着对端粒和端粒酶研究的深入,科学家们发现它们在细胞衰老、癌症发生和发展等方面扮演着重要的角色,因此,对端粒与端粒酶的研究不仅具有重要的理论价值,也具有广阔的应用前景。
本文将从端粒与端粒酶的基本概念入手,详细介绍它们在生物学中的重要作用,回顾相关的科学发展历程,并重点解读近年来诺贝尔生理学或医学奖中涉及端粒与端粒酶的重要研究成果。
通过对这些成果的深入分析和解读,我们希望能够更好地理解端粒与端粒酶在生命科学中的地位和价值,同时也为未来的医学研究提供新的思路和方法。
二、端粒与端粒酶的基本概念端粒,也被称为端区或端粒DNA,是真核细胞线性染色体末端的特殊结构。
它们像一顶帽子,保护着染色体的末端,防止其受到损伤或与其他染色体发生融合。
端粒的存在对于维持染色体的完整性和稳定性至关重要。
端粒主要由重复的非编码DNA序列组成,这些序列在染色体末端形成特定的结构,从而起到保护作用。
端粒酶则是一种特殊的逆转录酶,能够合成端粒DNA。
其主要功能是在端粒DNA受到损伤或缩短时,通过添加重复的DNA序列来修复端粒,从而保持染色体的稳定性和长度。
染色体是如何被端粒和端粒酶保护的?——2009年诺贝尔生理学或医学奖解读

染色体是如何被端粒和端粒酶保护的?——2009年诺贝尔生
理学或医学奖解读
李大可
【期刊名称】《生命科学研究》
【年(卷),期】2010(014)003
【摘要】端粒对维持染色体的稳定和延长细胞寿命至关重要,其长度的维持有赖于端粒酶的存在.布莱克本和绍斯塔克发现端粒中的一种独特DNA序列能保护染色体免于退化,格雷德和布莱克本发现了端粒酶及其作用.这些发现揭示了端粒形成和端粒酶保护染色体的机理,3位美国科学家因此荣获2009年诺贝尔生理学或医学奖.【总页数】5页(P278-282)
【作者】李大可
【作者单位】复旦大学,生命科学学院,生理学和生物物理学系,立人实验室,中国上海,200433
【正文语种】中文
【中图分类】Q291
【相关文献】
1.端粒和端粒酶与衰老、癌症的潜在关系——2009年诺贝尔生理学或医学奖简介[J], 孔令平;汪华侨
2.端粒和端粒酶是如何保护染色体的——2009年诺贝尔生理学或医学奖工作介绍及相关研究进展 [J], 宋伟;宋德懋
3.端粒、端粒酶的发现和意义——2009年诺贝尔生理学或医学奖简介 [J], 张婷;王晓民
4.三名美国科学家因发现染色体端粒酶及其功能分享2009年诺贝尔医学奖 [J],
5.端粒和端粒酶的发现——2009年度诺贝尔生理学或医学奖成果介绍 [J], 郗鹏因版权原因,仅展示原文概要,查看原文内容请购买。
2009年诺贝尔生理学或医学奖

2009年诺贝尔生理学或医学奖引言2009年,诺贝尔生理学或医学奖揭晓了由三位科学家共同获得的荣誉。
他们通过对细胞生物学和遗传调控的研究,做出了重要的贡献,为人类健康和医学领域的发展带来了突破性的进展。
本文将对这三位诺贝尔奖获得者及其研究成果进行介绍和分析。
诺贝尔奖获得者2009年诺贝尔生理学或医学奖由伊丽莎白·布莱克本、卡罗尔·格雷德尔和杰克·沙泌尔共同获得。
他们的研究突破了细胞生物学和分子遗传学的重要难题,为后续研究和治疗疾病提供了重要的理论基础。
研究成果端粒酶逆转录酶的发现和功能伊丽莎白·布莱克本和卡罗尔·格雷德尔的工作主要集中在细胞端粒酶逆转录酶(telomerase)的研究上。
端粒酶逆转录酶是一种能够延长染色体末端的酶,它在细胞分裂过程中起着关键的作用。
在布莱克本和格雷德尔的研究中,他们发现了端粒酶逆转录酶的存在,并揭示了它与细胞衰老和癌症发展之间的关系。
通过对细胞中端粒酶逆转录酶的活性进行研究,布莱克本和格雷德尔发现了一种叫做“端粒”的结构。
端粒位于染色体末端,能够保护染色体免受损伤和衰老。
他们的发现为后续研究提供了重要的线索,帮助科学家们更好地理解染色体的稳定性和细胞衰老的机制。
RNA干扰的发现与应用杰克·沙泌尔的工作则集中在RNA干扰(RNA interference)的研究上。
RNA干扰是一种基因调控的机制,通过介导特定RNA分子的降解或抑制,来控制靶基因的表达。
沙泌尔的研究发现了一种叫做“小干扰RNA”的分子,它们能够干扰靶基因的转录或翻译过程。
这项发现不仅揭示了RNA干扰机制的存在,还为科学家们开辟了一条新的基因治疗途径。
利用小干扰RNA可以有效地靶向控制基因表达,为治疗疾病提供了新的思路和方法。
科学意义和应用前景这三位诺贝尔奖获得者的研究成果为细胞生物学和遗传调控领域带来了重大的突破,对生命科学的发展产生了深远影响。
2009年诺贝尔生理学或医学奖研究成果介绍

2009年诺贝尔生理学或医学奖研究成果介绍摘要:2009年10月5日瑞典卡罗林斯卡医学院诺贝尔生理学或医学奖评审委员会宣布将本年度诺贝尔生理学或医学奖授予三位美国科学家伊丽莎白·布莱克本(Elizabeth H Blackburn)、卡罗尔·格雷德(Carol W.Greider)和杰克·绍斯塔克(Jack W.Szostak),以表彰他们在上世纪80年代发现了“端粒和端粒酶是如何保护染色体的”。
本文主要介绍端粒和端粒酶是如何保护染色体的。
关键词:端粒,端粒酶,染色体Elizabeth Blackburn教授1948年出生在澳大利亚塔斯马尼亚州霍巴特市,毕业于墨尔本大学,1975年在剑桥大学获博士学位,而后在耶鲁大学做博士后,1990年至今在加州大学任教。
Jack Szostak教授1952年出生在英国伦敦,毕业于加拿大麦吉尔大学,1977年在美国康奈尔大学获博士学位,现供职于哈佛医学院、麻省总医院和霍华休斯医学研究所。
这两位科学家合作证实了真核生物的端粒具有保护染色体末端的作用。
Carol Greider教授1961年出生在美国加州的圣地亚哥,1987年在加州大学Black—burn教授的指导下获博士学位,而后在冷泉港实验室做博士后,1997年至今任教于约翰·霍普金斯大学医学院。
Greider教授与Blackbum教授合作发现了催化延伸端粒结构的端粒酶。
1端粒能保护染色体末端以下仅以哺乳动物的端粒为例加以介绍。
哺乳动物端粒的重复序列为(TTAGGG/AATCCC),其中G链3’端是一段单链的悬突(overhang),C链5’端以序列(ATC)结束。
电镜观察发现,端粒结构是一个双环结构,称为T环(T—loop),3’端的悬突替代G链的一段序列与C链配对,形成D环(D-loop),T环的形成使得染色体的末端被包裹保护起来而免遭破坏。
哺乳动物的端粒与一个6种蛋白构成的复合物shelterin结合,这6种蛋白分别为TRFl、TRF2、POTl、TIN2、Rapl和TPPl。
2009诺贝尔 端粒酶文章

2009诺贝尔端粒酶文章2009年诺贝尔生理学或医学奖颁发给了三位科学家——伊丽莎白·H·布莱克本、卡罗尔·W·格雷德尔和杰克·W·斯托斯,以表彰他们在端粒酶研究上所做出的突出贡献。
他们的研究不仅深化了我们对人类健康和长寿的理解,更为未来的医学和抗衰老研究带来了新的指导意义。
做为生命的时钟,端粒酶是一种可以在人类染色体末端发挥关键作用的酶。
它能够维持染色体末端的稳定性和完整性,以免发生不正常的DNA损伤和降低细胞的功能。
随着细胞的分裂和老化,端粒酶的活性逐渐降低,导致端粒缩短,最终导致细胞进入老化状态或死亡。
这对于人类衰老和疾病的发生有着重要的影响。
为了更好地理解端粒酶的作用和机制,布莱克本等科学家们开展了一系列的实验和研究。
他们的工作不仅揭示了端粒酶在维持染色体完整性中的重要作用,同时还发现了一种由RNA和蛋白质组成的复合物——端粒酶。
该复合物能够延长端粒的长度,保护染色体免受DNA 损伤和降低细胞功能。
这项研究对于理解细胞衰老和肿瘤发展的过程具有重要意义。
这项突破性的研究为未来的医学和抗衰老研究提供了新的指导意义。
首先,我们能够更深入地了解细胞老化和疾病发生的机制,从而寻找新的治疗方法。
其次,通过调节端粒酶的活性,我们能够延长端粒长度,保护染色体和细胞免受损伤,延缓细胞衰老和死亡。
这将有助于延长人类的寿命和改善人类健康。
此外,端粒酶的研究还可以为肿瘤治疗提供新的思路。
由于肿瘤细胞具有无限增殖能力,它们通常会表现出较高的端粒酶活性。
研究人员通过抑制或调节端粒酶的活性,可以阻止肿瘤细胞的增殖和生长,从而有望开发出更高效的肿瘤治疗方法。
总之,2009年诺贝尔生理学或医学奖的颁发对于推动细胞老化、疾病和肿瘤研究有着重要的意义。
通过深入研究端粒酶和端粒的作用机制,我们能够更好地理解人类的衰老过程,为延缓衰老、改善人类健康和开发更有效的肿瘤治疗方法奠定了坚实的基础。
端粒酶
端粒=生命时钟
端粒就像一顶高帽子置于染色体头上,被科学 家称作“生命时钟”。在新细胞中,细胞每分 裂一次,端粒就缩短一次,当端粒不能再缩短 时,细胞就无法继续分裂而死亡。 卡萝尔・格雷德和伊丽莎白・布莱克本则确定 了端粒酶,端粒酶是形成端粒DNA的成分。这 些发现解释了染色体的末端是如何受到端粒的 保护的,而且端粒是由端粒酶形成的。
步加强。在这个过程中产生了一群“超级老
鼠”,它们的寿命更长,而且更有抗癌能力。
?
病 毒 会 程 序 性 死 亡 不
端粒酶 “一半是魔鬼,一半是天使”
因为端粒酶的活跃,癌细胞不停
增殖; 但是,如果能够调控正常细胞的 端粒酶,使之具备相当的活性, 那么正常细胞的寿命就可能延长, 起到抗衰老的作用。
一个实验
2008年,西班牙国立癌症研究中心的科学家 将端粒酶植入小白鼠的干细胞中,这些小白鼠 的寿命比正常情况下延长了50%。这种改良老 鼠经过继续喂养,它们的新DNA形态会进一Βιβλιοθήκη 端粒=生命时钟端粒酶
2009年诺贝尔奖
2009年诺贝尔生理学或医学奖授予美国科学 家伊丽莎白・布莱克本、卡萝尔・格雷德和杰 克・绍斯塔克,以表彰他们“发现端粒和端粒 酶是如何保护染色体的”。
这三位科学家解决了生物学上的一个重大问题, 即在细胞分裂时染色体如何进行完整复制,如 何免于退化,其中奥秘蕴藏在端粒和端粒酶上。
染色体的保护者——端粒与端粒酶

研究论文H A I X I A K E X U E年第3期(总第3期)3海峡科学染色体的保护者—端粒与端粒酶福建教育学院理科研修部郑俊敏[摘要]端粒是位于染色体末端、能保护染色体不被降解的特殊结构。
端粒酶则是能合成端粒D N A 的酶,使得端粒的长度和结构得以稳定。
端粒和端粒酶的发现推动了科学家对人类衰老和肿瘤发生机制的研究,有助于相关疾病的预防和治疗。
文章对端粒和端粒酶的发现过程、其主要结构和功能、以及与相关疾病的预防和治疗作了简要介绍。
[关键词]端粒端粒酶结构功能2009年10月5日,瑞典皇家科学院将诺贝尔生理学或医学奖授予了3位美国科学家:伊丽莎白布莱克本(Elizabeth H.Blackburn ),卡萝尔格雷德(Carol W.Greider )和杰克绍斯塔克(Jack W.Sz ostak )。
诺贝尔奖主页上介绍他们获奖的原因是揭示了“染色体是如何被端粒和端粒酶保护的”。
端粒是一种在染色体末端部分的一个帽子样的特殊结构(图1),其主要作用是使染色体不被降解。
端粒酶则是帮助端粒合成的分子,它维持着端粒的长度和结构的稳定,从而保护染色体。
绝大多数成体细胞缺乏端粒酶,导致端粒保护染色体的功能逐渐丧失,这被认为是决定细胞寿命的一个重要因素〔1〕。
而在一些细胞中,如胚胎干细胞和癌细胞,端粒酶的活性高度表达,使得这些细胞不断分裂却不会遭受染色体损伤,延迟了衰老〔2〕。
同时,一些遗传病也是由于端粒酶活性的缺失而导致的。
正是由于这三位科学家开创性的工作,向人们揭开了端粒和端粒的神秘面纱,并了解了它们与人体衰老和许多疾病的关系。
而整个端粒和端粒酶的研究进程就像相继解开一个个智力谜团一样有趣,充满了思想的光辉。
重现这个思路对广大教师和学生都是有启发意义的。
本文也提供了一个很好的科学问题推演的教学案例。
图1端粒及其基本结构1端粒和端粒酶的发现染色体末端对于染色体稳定性的重要作用,最早是在上世纪30年代,Barbara M clintock (因为发现玉米的转座子获得诺奖)和Hermann M uller (因为发明用X 射线突变基因而获得诺奖)在对玉米和果蝇的研究中提出来的〔3,4〕。
端粒及端粒酶的研究进展

Your site here
LOGO
端粒酶应用的展望
肿瘤的诊断 :研究结果表明,85%的
恶性肿瘤组织端粒酶活性均呈阳性,而癌旁 组织的阳性率仅有4%。因此端粒酶活性可 以作为肿瘤诊断的指标之一。可以用许多 材料来检测端粒酶的活性,不必采用手术取 样,因此简单易行,操作方便。且端粒酶活性 水平还可以区分肿瘤的恶性程度,从而选择 恰当的治疗方法。
端粒及端粒酶 研究进展
——王振
宇
LOGO
端粒和端粒酶的概述
2009年,美国的三位科学家Elizabeth H· Blackburn、Carol W· Greider和Jack W· Szostak发表了题为“端粒和端粒酶是 如何保护染色体的”而共同获得诺贝尔生 理学或医学奖。也是从这一重大研究成果 开始,端粒和端粒酶的研究为人类衰老和 肿瘤带来了福音。
Your site here
LOGO
端粒和端粒酶
端粒酶的结构: 一般认为端粒酶由端粒 酶RNA(TR)、端粒酶相关蛋白和端粒酶催 化亚基(TERT)3部分组成。许多物种的TR 已经被成功克隆,如纤毛虫、酵母、人和鼠 等。
Your site here
LOGO
端粒的功能:
1.保护染色体末端 真核生物的端粒DNA-蛋白复合物,如帽 子一般,保护染色体末端免于被化学修饰或 被核酶降解,同时可能还有防止端粒酶对端 粒进行进一步延伸的作用。改变端粒酶的模 板序列将导致端粒的改变,从而诱导细胞衰 老和死亡。
Your site here
LOGO
端粒的功能:
3.决定细胞的寿命
染色体复制的上述特点决定了细胞分裂 的次数是有限的,端粒的长度决定了细胞的 寿命,故而被称为“生命的时钟”。
端粒与端粒酶的发现历程——记诺贝尔生理学或医学奖

端粒与端粒酶的发现历程——记诺贝尔生理学或医学奖本文是基于个人理解来整理的端粒和端粒酶的发现历史,因为知识时间有限,其中必有偏差和谬误的地方,关键之处还是以原始文献为主。
本人之所以赶这趟诺贝尔奖热,花大量的时间进行文献阅读和整理,是因为它提供了一次极好的向公众传播科学思想的机会。
由于端粒和端粒酶领域的一系列发现贯穿着"发现现象/问题"-"提出概念/模型"-"实验验证"的思路,重现这个思路对科学工作者是有启发意义的。
本文也提供了一个很好的教学案例。
引言-到底是"谁"得诺奖了?2009年诺贝尔生理学或医学奖授予了UCSF(加州大学旧金山分校)的Elizabeth Blackburn(简称Liz),Johns Hopkins University(约翰霍普金斯大学)的Carol Greider(简称Carol),以及Howard Medical School (哈佛医学院)的Jack Szostak。
诺贝尔奖主页上介绍她/他们获奖的原因是揭示了"how chromosomes are protected by telomeres and the enzyme telomerase"(染色体是如何被端粒和端粒酶保护的),这样描述是非常专业的。
当然更多的公众媒体为了吸引眼球,会用"Aging Research Wins Nobel Prize"(衰老研究摘取诺贝尔奖)的标题,这颇有误导之嫌。
"揭开衰老与癌症的奥秘",这样的标题更是耸人听闻,偏离这个诺贝尔奖的用意了。
不可否认端粒和端粒酶的发现能获得诺贝尔奖,是因为它跟衰老和癌症的潜在关系获得了更多公众的关注。
但是迄今为止它只是衰老和癌症的correlator(相关者),勉强算得上indicator(指示者),还远不是causer (引起者)。
端粒和端粒酶与衰老_癌症的潜在关系_2009年诺贝尔生理学或医学奖简介

端粒和端粒酶与衰老、癌症的潜在关系———2009年诺贝尔生理学或医学奖简介孔令平① 汪华侨②①副教授,广州医学院从化学院,广州510182;②教授,中山大学中山医学院人体解剖学与脑研究室,广州510080关键词 端粒 端粒酶 细胞 衰老 癌症 美国科学家伊丽莎白・布莱克本、卡萝尔・格雷德和杰克・绍斯塔克三人同时获得2009年诺贝尔生理学或医学奖,这是由于他们发现“染色体是如何被端粒和端粒酶保护的”,这一研究成果揭开了人类衰老和肿瘤发生等生理病理现象的奥秘。
本文将就端粒和端粒酶的发现、结构和功能及其与人类衰老、癌症的潜在关系等方面做一简要介绍。
人的生老病死,这或许是生命最为简洁的概括,但其中却蕴藏了无穷无尽的奥秘。
2009年10月5日,瑞典卡罗林斯卡医学院宣布将2009年诺贝尔生理学或医学奖授予美国科学家伊丽莎白・布莱克本(Elizabet h H.Blackburn)、卡萝尔・格雷德(Carol W.Greider)和杰克・绍斯塔克(J ack W.Szostak),以表彰他们发现“染色体是如何被端粒和端粒酶保护的”。
这3位科学家的发现“解决了一个生物学重要课题,即染色体在细胞分裂过程中是怎样实现完全复制,同时还能受到保护且不发生降解”。
由此可能揭开了人类衰老和罹患肿瘤等严重疾病的奥秘。
染色体是生物细胞核中的一种易被碱性染料染色的线状物质。
大家都知道,正常人的体细胞有23对染色体,这对人类生命具有重要意义,其中的X和Y染色体是决定男女性别的性染色体。
在染色体的末端,有一个像帽子一样的特殊结构,这就是端粒。
作为染色体末端的“保护帽”,端粒具有维持染色体的相对稳固、防止DNA互相融合及重组的功能,犹如卫兵那样守护染色体不受损害。
而端粒酶的作用则是帮助合成端粒,使得端粒的长度等结构得以稳定。
“染色体携有遗传信息。
端粒是细胞内染色体末端的‘保护帽’,它能够保护染色体,而端粒酶在端粒受损时能够恢复其长度。
2009年分子生物学诺贝尔奖感想启发评价

2009年分子生物学诺贝尔奖感想启发评价端粒酶的研究思路值得学习曾武威、章静波(中国医学科学院基础医学研究所):10月5日,瑞典卡罗琳斯卡医学院宣布,将2009年诺贝尔生理学或医学奖授予3位美国科学家,以表彰他们“发现端粒和端粒酶是如何保护染色体的”。
这一发现解答了生物学上的一个重大问题,即染色体在细胞分裂时是如何以完整的形式自我复制的,以及它们是如何受到保护而免于降解的。
3位科学家揭示了其答案就在染色体的末端——端粒,以及形成端粒的酶——端粒酶上。
我们身体中的每个细胞都含有完整的基因组,携带基因的线状dna分子被包裹在染色体中。
在染色体的末端部分有一个像帽子一样的特殊结构,这就是端粒。
它是细胞内染色体末端的保护帽,能够保护染色体。
而端粒酶的作用则是形成端粒,使得端粒的长度等结构得以稳定。
1982年,伊丽莎白·布莱克本应用dna测序技术揭示了端粒由短的重复dna片段组成。
随后,她和杰克·邵斯塔克发现,单细胞生物四膜虫的端粒可以保护其自身dna分子,也可以保护种属相隔很远的酵母微染色体。
端粒的保护作用在进化上具有如此巨大跨度的保守性,提示这是一种非常基本的生物机制在起作用。
于是,布莱克本与当时是其研究生的卡萝尔·格雷德开始探索端粒的维护机制。
1984年,他们发现了端粒酶。
它是由rna和蛋白质组成的,其rna成分中包含端粒dna的特征性序列ccccaa。
端粒合成时,该序列作为模板,而其蛋白质成分中含有酶活性。
端粒酶通过其自身rna提供的模板将端粒dna延伸,从而以此分子末端为平台,使dna聚合酶能够复制完整长度的染色体,而不会遗漏其最末端部分。
在此之前,分子生物学家们一直无法解释dna在细胞分裂时如何加到复制的dna双链的末端,这项发现解释了这一长时间令人困惑的“末端复制问题”。
端粒和端粒酶的发现历程教学提纲

端粒和端粒酶的发现历程端粒和端粒酶的发现历程廖新化引言2009年诺贝尔生理学或医学奖授予了UCSF(加州大学旧金山分校)的Elizabeth Blackburn (简称Liz),Johns Hopkins University(约翰霍普金斯大学)的Carol Greider(简称Carol),以及Howard Medical School(哈佛医学院)的Jack Szostak。
诺贝尔奖主页上介绍她/他们获奖的原因是揭示了“how chromosomes are protected by telomeres and the enzyme telomerase”(染色体是如何被端粒和端粒酶保护的)。
端粒和端粒酶的研究进程中贯穿着“发现现象/问题”-“提出概念/模型”-“实验验证”的思路,整个过程就像相继解开一个个puzzle(智力谜团)一样有趣,充满了思想的光辉。
重现这个思路对科学工作者是有启发意义的。
本文也提供了一个很好的科学问题推演的教学案例。
染色体末端的两个难题以及端粒的概念20世纪70年代初,对DNA聚合酶特性的深入了解引申出了一个染色体的复制问题。
DNA聚合酶在复制DNA的时候必须要有引物来起始,而且它的酶活性具有方向性,只能沿着DNA5’到3’的方向合成。
染色体复制之初可以由小RNA作为引物起始合成,之后细胞的修复机器启动,DNA聚合酶能够以反链DNA为模板,以之前合成的DNA为引物,合成新的DNA取代染色体中间的RNA引物。
但是线性染色体最末端的RNA引物因为没有另外的引物起始,没有办法被DNA 取代。
所以线性染色体DNA每复制一轮,RNA引物降解后末端都将缩短一个RNA引物的长度(图1,简化的示意图,实际上染色体的DNA双链末端不会是平的)。
尽管这个引物不长,但是细胞千千万万代地不断复制,如果不进行补偿,染色体不断缩短,最终就会消失。
James Watson(因为发现DNA双螺旋结构获得诺奖)最早就明确指出了这个“末端隐缩问题”,并猜想染色体也许可以通过在复制前联体(染色体末端跟末端连起来)的方式来解决末端复制的问题[1]。
2009诺贝尔生理及医学奖

生老病死,这或许是人类生命最为简洁的概括,但其中却蕴藏了无数的奥秘。获得2009年诺贝尔生理学或医学奖的三位美国科学家,凭借“发现端粒和端粒酶是如何保护染色体的”这一成果,揭开了人类衰老和罹患癌症等严重疾病的奥秘。 在生物的细胞核中,有一种易被碱性染料染色的线状物质,它们被称为“染色体”。正常人的体细胞有23对染色体,它们对人类生命具有重要意义,例如众所周知,决定男女性别的就是一对染色体。在染色体的末端部分有一个像帽子一样的特殊结构,这就是端粒。而端粒酶的作用则是帮助合成端粒,使得端粒的长度等结构得以稳定。 “染色体携有遗传信息。端粒是细胞内染色体末端的‘保护帽’,它能够保护染色体,而端粒酶在端粒受损时能够恢复其长度。”获奖者之一的伊丽莎白·布莱克本介绍说:“伴随着人的成长,端粒逐渐受到‘磨损’。于是我们会问,这是否很重要?而我们逐渐发现,这对人类而言确实很重要。” 卡罗林斯卡医学院发布的新闻公报说,这三位科学家的发现“解释了端粒如何保护染色体的末端以及端粒酶如何合成端粒”。借助他们的开创性工作,人们知道,端粒不仅与染,端粒变短,细胞就老化。相反,如果端粒酶活性很高,端粒的长度就能得到保持,细胞的老化就被延缓。 不过需要指出的是,近年来陆续有研究发现,端粒和染色体等虽然与细胞老化有关,进而影响衰老,但并非唯一的因素,“生命衰老是一个非常复杂的进程,它有许多不同的影响因素,端粒仅仅是其中之一”。 “这是有关人类衰老、癌症和干细胞等研究的谜题拼图中重要的一片,”新闻公报说,“他们的发现使我们对细胞的理解增加了新的维度,清楚地显示了疾病的机理,并将促使我们开发出潜在的新疗法。”
诺贝尔生理学或医学奖

美国科学家罗伯特·霍维茨。他们为研究器官发育和程序性细胞死亡过程中的基因调节作用做出了重大贡献。
2001年,美国科学家利兰·哈特韦尔、英国科学家保罗·纳斯和蒂莫西·亨特。他们发现了导骤锔胞分裂的关键性调节机制,这一发现为研究治疗癌症的新方法开辟了途径。
k型的毒杀型T细胞杀死。这与已知免疫系统的功用玩全吻合,免疫细胞只攻击病毒感染的细胞,而不会影响正常细胞。而实验出乎他们意料的部分是,被病毒感染的k型标的细胞,并不会被b型老鼠的毒杀型T细胞所杀死。T细胞在辨认外来抗原时,很明显地受限于自己的组织抗原。这样的结果,在他们以多种不同纯种老鼠及多种不同的细胞实验中得到玩全确认。因此他们提出一个模型,指出T细胞的辨识病毒需要双重承认,一个是自己细胞上的组织抗原,另一个是外来病毒抗原。单独的组织抗原,或是单独的病毒抗原,不会引起T细胞反应。如果病毒与不是自身的组织抗原同时出现,也不会被T细胞辨认。这也就是说T细胞辨识的是自己(self)加上异己(nonself)。免疫系统一直被认为主要是区分自己和异己,而对异己予以摧毁。因此杜赫堤和辛克纳吉的「自己加异己」辨认,引起极大的回响。尤其又带了几分哲理推敲,令人印象鲜明。他们的T细胞辨认模式,在几年内得到玩全的证实。杜赫堤利辛克纳吉的发现,也是第一个展示组织抗原免疫机能的报告。我们身体细胞上的组织抗原,原来是引发T细胞免疫反应的关键分子。这个发现可以说是以后二十年来T细胞免疫研究的基础。时至今日,我们已经知道所谓的双重承认事实上是病毒其中一段胜肽与组织抗原结合的复合体被辨识,T细胞则是以其表面的T细胞受体来辨认搭宕合体(图四)。如果T细胞受体与组织抗原及病毒胜 的复合体可以吻合,则其结合力会使毒杀型T细胞活化,进而杀死被病毒感染的细胞。这也是辅助型T细胞用以辅助B细胞产生抗体的辨认方式。辅助型T细胞借着表面抗体辨识含有特定抗原胜 的组织抗原,而与该特定B细胞结合。因此T细胞受体、病原胜肽与组织抗原三者之作用,可以说是对抗外来感染的免疫反应(包括抗体和毒杀性T细胞)之根本。我们也可以理解,为什么杜赫堤和辛克纳吉的实验会有这么大的影响。
端粒与端粒酶

染色体是真核生物遗传物质的载体, 染色体是真核生物遗传物质的载体,维持其稳定性对于 高等生物至关重要.然而,作为生命蓝图的承载者, 高等生物至关重要.然而,作为生命蓝图的承载者,染 色体却处在一个危机四伏的环境中, 色体却处在一个危机四伏的环境中,经常遭到细胞内多 种不利因素的干扰和破坏.可以想象, 种不利因素的干扰和破坏.可以想象,如果得不到有效 保护, 稳定性难以维持, 保护,DNA稳定性难以维持,后果将不堪设想.染色体 稳定性难以维持 后果将不堪设想.
探索一步步深入
1984年,Carol与Liz精心讨论设计实验,用四膜虫的核抽提液与体外的 1984 端粒DNA进行温育,试图在体外检测到这个"酶"活性,看到端粒的延伸 。经过不断优化条件,尤其是把底物换成体外合成的高浓度的端粒DNA 后,同年的圣诞节终于清楚地看到了"酶"活性。在测序胶的同位素曝光 片上,端粒底物明显被从新加上了DNA碱基,而且每六个碱基形成一条 很深的带,与四膜虫端粒重复基本单位为六个碱基正好吻合。这种酶活 性不依赖于DNA模板,只对四膜虫和酵母的端粒DNA进行延伸,而对随机 序列的DNA底物不延伸;并且该活性不依赖于DNA聚合酶。由于同源重组 对序列没有特异性的要求并且依赖于DNA聚合酶的活性,至此,她们澄 清了这两种假说,证明了有一种"酶"来延伸端粒DNA。这种酶后来被命 名为"端粒酶"(telomerase)。
探索一步步深入
20世纪五六十年代 20世纪五六十年代,当科学家们尝试解析真核生物如何实现染色体DNA的精 世纪五六十年代 确复制时,又一个端粒相关的难题产生了。DNA聚合酶进行每一轮线性DNA的 复制都会导致少量末端核苷酸的丢失,其结果是,真核生物线性染色体,作 为基因的载体,会在细胞分裂过程中逐渐缩短。1972年,James Watson提出 了"末端复制问题",他同时推测,真核生物需要一种特殊机制来确保线性染 色体末端的完整复制。同时,Alexey Olovnikov也推测,染色体末端的逐渐 缩短将导致细胞的衰老。 20世纪七十年代 20世纪七十年代,Hayflick首次提出将体外培养的正常人成纤维细胞的“ 世纪七十年代 有限复制力”作为细胞衰老的表征。在此过程中,细胞群中的大部分细胞 经历了一定次数的分裂后便停止了,但它们并没有死亡,仍保持着代谢活 性,只是在基因表达方式上有一定的改变。于是Hayflick猜测细胞内有一 个限制细胞分裂次数的“钟”,后来通过细胞核移植实验发现,这种“钟 ”在细胞核的染色体末端——端粒。
09诺贝尔——端粒酶机理的 发现及意义

2009年诺贝尔生理学或医学奖得主主要成果及对人类社会的影响生命科学学院 2010级生物学试验班马福林辅导教师:丁显平教授内容摘要端粒是真核生物染色体末端的一种特殊结构, 对于维持染色体稳定性具有十分重要的意义。
北京时间10月5日下午17时30分,2009年度诺贝尔生理学或医学奖在瑞典卡罗林斯卡医学院揭晓,三名美国科学家Elizabeth.Blackburn、Carol W. Greider以及Jack W. Szostak共同获得该奖项。
他们发现了由染色体冠制造的端粒酶,这种染色体的自然脱落物将引发衰老和癌症。
关键词端粒 端粒酶 染色体 诺贝尔生理学与医学奖 延缓衰老 治疗癌症1. 简述获奖者成果2009年度诺贝尔生理学或医学奖于北京时间10月5日下午17时30分,在瑞典卡罗林斯卡医学院揭晓,三位美国科学家伊丽莎白•布兰克波恩(Elizabeth H. Blackburn)、卡罗尔•格雷德(Carol W. Greider)以及杰克•绍斯塔克(Jack W. Szostak)共同因发现了端粒和端粒酶保护染色体的机理而获得该奖项卡罗林斯卡医学院方面称,这三人“解决了生物学上的一个重大问题”,即在细胞分裂时染色体如何进行完整复制,如何免于退化。
其中奥秘全部蕴藏在端粒和端粒酶上。
由染色体根冠制造的端粒酶是染色体的自然脱落物,能引发衰老和癌症。
端粒也被科学家称作“生命时钟”,在新细胞中,细胞每分裂一次,端粒就缩短一次,当端粒不能再缩短时,细胞就无法继续分裂而死亡。
Elizabeth H. Blackburn他们发现的端粒酶,在一些失控的恶性细胞的生长中扮演重要角色。
大约90%的癌细胞都有着不断增长的端粒及相对来说数量较多的端粒酶。
换句话说,正常的细胞,每次复制时都会发生端粒的缩短,因此,细胞复制的次数是有限的,一般50次左右。
但是这三人发现癌细胞发现了一种酶,因为这种酶的关系,癌细胞的端粒不会发生正常细胞中的缩短现象。
2009年诺贝尔奖生理学或医学奖端粒与端粒酶

1977年在康奈尔大学 获得博士学位。师从中 国工程学院外籍院士吴 瑞教授
目前是马萨诸塞综合
医院遗传学教授,并同
时任职于美国霍华德·休
斯医学研究所
2021/10/10
8
2.获奖者
卡萝尔·格雷德 (Carol reider)
➢1961年出生在美国加利 福尼亚州,
➢1987年获得博士学位, 其导师正是伊丽莎白·布 莱克本,之后曾在美国科 尔德斯普林实验室从事博 士后研究
➢1997年起开始担任约翰 斯·霍普金斯大学医学院 教授
2021/10/10
9
3.获奖成果— 端粒的发现
赫尔曼·马勒和芭芭拉·麦克林托克发现染色体
1930 末端的一个特殊结构他们认为该结构可以阻止
染色体彼此黏附,并设想其可能具有保护性作用。
赫尔布莱克波恩在研究一种简单的池生生物
1978 四膜虫的染色体时,获得了一个突破性的发现,在该动
Ӂ 1990年开始担任美国加利福 尼亚大学旧金山分校生物学和 生理学教授。
Ӂ 2007年伊丽莎白·布莱克本因 学术成就卓著曾被美国《时代 》周刊评为年度全球最具影响 力的100个人物之一。
2021/10/10
7
2.获奖者
杰克·绍斯塔克(Jack
Szostak)
美国人。1952年生于 伦敦,在加拿大长大。
2021/10/10
16
4.获奖成果— 端粒酶
概念
端粒酶是一种RNA与蛋白的复合体,它以 自身RNA上的一个片段为模板通过逆转录合 成端粒重复序列,并通过一种RNA依赖性聚 合酶(如逆转录酶)机制加到染色体3’末 端以延伸端粒。
组成
RNA(作为模板) 蛋白质(反转录酶)
作用
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
虽然Blackbum和Szostak的实验证实了端粒在 物种进化中的保守性以及其保护染色体稳定性的重 要功能,但是端粒的复制问题始终未能解决。科学 家们试图用多种模型来解释这个问题,包括同源重 组或转座作用、回文结构或发卡结构等,也有科学家 设想存在一种全新的酶催化合成端粒。
万方数据
色体DNA发生同源重组,或很快被降解而不能长时 间稳定存在。因此,如何使线性质粒分子在酵母细 胞核内稳定存在并进行复制,便成为Szostak实验室 亟待解决的一个难题。
1980年,Blackburn在戈登会议(Gordon Confer- ence)上关于嗜热四膜虫染色体端粒结构的报告引 起了Szostak的极大兴趣,他们决定合作设计一个方 案来提高酵母线性DNA分子的稳定性。他们将嗜 热四膜虫rDNA一个长1.5kb、含有端粒的末端片 段,连接到酵母环状质粒酶切后形成的线性分子两 端,构成了一个全新的线性DNA分子。令人惊讶的 是这种分子转入酵母后,它们可以稳定存在并且进 行复制,子代DNA保持了嗜热四膜虫端粒的结构特 点(图3)。如果将这种分子一端换成酵母自身染色 体的末端片段,这种稳定性同样存在一J。Blackbum 和Szostak的这一研究成果的重要性在于发现了嗜 热四膜虫rDNA的端粒能在亲缘关系较远的酵母中 发挥功能这一现象,说明端粒在进化上具有很强的 保守性。在后续的研究中,Blackburn和Szostak测 定了酵母的端粒是由序列C1—3A不规则重复构 成,并且发现酵母中特有的端粒重复序列同样可以 添加到嗜热四膜虫线性质粒分子的末端发挥同样的 功能㈨。
圈4端粒合成模式图 在G链上端粒酶催化端粒沿3 7—巧7延长,再以此
为模板合成互补c链
随后,她们发现这种特殊的转移酶是一种由 RNA和蛋白质构成的核蛋白(ribonucleoprotein, RNP)复合体,两种成分均是其发挥功能所必需的, 她们将这种酶命名为端粒酶(telomerase)¨J。接着, 两人又克隆了嗜热四膜虫端粒酶上一段159bp长的 RNA片段(159 RNA),该片段上含有(CAAC— CCCAA)序列,如果设法干扰这一段序列,那么端粒 酶将无法正常工作,这说明端粒酶可能以此为模板 来延长端粒¨1。1990年,Blackburn实验室的Gou— Liang Yu将这段序列突变,结果合成的端粒序列随 之变化,且与之互补,这充分说明了端粒酶是以其上
四、端粒维持染色体末端的稳定性 Elizabeth Blackburn和Jack Szostak两位科学家 在上世纪80年代初发现了端粒能够保护染色体末 端,并且这种保护作用具有进化保守性,但具体机制 当时并不清楚。经过20多年的研究,科学家已经对 端粒的结构和功能有了更为深刻的理解。以下仅以 哺乳动物的端粒为例加以介绍。 哺乳动物端粒的重复序列为(TFAGGG/AATC. CC),其中G链3 7端是一段单链的悬突(overhang) (图6),C链5 7端以序列(ATC)结束。电镜观察发 现,端粒结构是一个双环结构,称为T环(T—loop),3’ 端的悬突替代G链的一段序列与C链配对,形成D 环(D-loop)(图6),T环的形成使得染色体的末端 被包裹保护起来而免遭破坏u 0|。哺乳动物的端粒 与一个6种蛋白构成的复合物sheherin结合,这6 种蛋白分别为TRFl、TRF2、POTl、TIN2、Rapl和 TPPl。TRFl和TRF2(telomeric repeat binding factor 1 and 2)能够特异性识别并结合双链重复序列上的 5 7一YTAGGG,丌R-3 7序列,POTl(protection of telo— mel七¥1)则能识别并结合3 7端悬突和D环上单链重 复序列上的5 7一(T)TAGGGTYAG-3’序列,上述三种
3’硝’方向先将G链延长,再以此为模板合成互补
的c链,这样就保证了子代DNA链5 7端的完整性 (图4)。她们用一段合成的端粒作为底物,加入嗜 热四膜虫的无细胞浸出物(cell free extract)中,并混 合了Ot一32P—dGTP与dATP、drrI'P、dCTP,孵育一段时 间后将产物进行凝胶电泳分析。结果发现,DNA寡 聚体的延长是以6碱基序列重复相加来完成的,上 述结果说明了确实存在一种此前未知的酶来催化端 粒特有的重复序列延伸。为了排除这种延伸是由已 知的DNA聚合酶以四膜虫自身染色体上的 (CCCCAA)序列为模板完成的,她们用微球菌核酸 酶(micrococcal nuclease)破坏无细胞浸出物中四膜 虫自身染色体上的(CCCCAA)n序列,但上述过程 依然存在,证实了上述结论。另外,嗜热四膜虫中的 这种转移酶还能催化(rlTGGGG)叠加到酵母端粒重 复序列构成的引物上,这说明此种转移酶同样具有 进化上的保守性。但其对随机序列的DNA底物不 延伸;并且该活性不依赖于DNA聚合酶旧J。
一、端粒问题研究的由来 “端粒”(telomere)的概念最早由果蝇遗传学家 Hermann J.Muller(1946年诺贝尔生理学或医学奖
获得者)提出。Muller于上世纪20年代用x射线对 果蝇进行了人工诱变,在对突变基因的分析中,他发 现很多突变与染色体片段的重排——倒转、易位或 缺失有关。重排是通过片段的重新连接(reattach— ment)来实现的,但是这种连接不会发生在染色体游 离端(free end)与游离端、或者游离端与断端之间。 1938年,Muller在当时的科学杂志The Collecting Net上发表了一篇题为“The remarking of Chromo一 ¥omes”的演讲稿,文中他将染色体的游离端命名为 端粒。端粒的字面意思是末端部分(end part),由两 个希腊词根“telos”(末端)和“mel'o¥“(部分)合成。 Muller设想端粒是一种末端基因,指出“这种末端基 因具有某种特殊的功能,即可以对染色体的末端起 到封闭(sealing)的作用,从某种意义上讲,如果染色 体的末端不被封闭,染色体就不会持续存在”。 Muller的结论由Barbara McClintock(1983年诺贝尔 生理学或医学奖获得者)在玉米中得到证实。1941 年McClintock在分析X射线诱导玉米发生基因突 变时,同样发现了染色体存在易位、缺失等现象,这 些都是DNA断端融合(fusion)的结果,但是“染色 体的末端不会与任何染色体片段发生融合”,也就 是说,DNA断端具有“黏”性,易于与其他断端融合, 而正常的染色体末端则非常稳定¨J。产生这一现 象的原因是什么?合理的推测是,染色体末端不同 于DNA断端,它应该有一个特殊的结构来避免它们 之间发生融合。这是有关端粒的第一个问题。
图3带有端粒的人工微染色体可以在酵母菌中稳定存在 嗜热四膜虫rDNA的一个长1.5kb、含有端粒的末端片段连接到 酵母线性质粒分子的两端,构成了一个全新的线性质粒分子。 将其转入酵母细胞后,可以稳定存在(http://nobelprize.org/no- bel_prizes/medicine/laureates/2009/)
上世纪50年代以后,随着DNA双螺旋结构的 发现以及DNA聚合酶(DNA polymerase)催化DNA 复制机制的阐明,染色体末端的复制问题随之出现。 1972年,James Watson(1962年诺贝尔生理学或医 学奖获得者)在探讨噬菌体r17的DNA复制时【2 o指 出,真核生物线性DNA的复制始于内部起始点,以 短链RNA分子作引物,沿503方向延伸互补链。
DNA链3 7—7方向的延伸,子代DNA链5 7端的引
物被切去后会留下一段空隙,导致子代DNA分子上 有一段不完整的57末端,DNA链也会随着不断的复 制而逐渐缩短(图2)。但实际上真核生物线性 DNA末端的复制是可以完整的,这个机制是什么? 这是有关端粒的第二个问题。
围2其核生物线状DNA在复制过程中导致末端部分缺失
上述两个有关端粒的问题引起了一些科学家的 兴趣,其中就有Blackburn教授和Szostak教授。
二、Blackburn与Szostak合作发现端粒能保护 染色体末端
1975年,Blackburn在耶鲁大学Joseph Gall的 实验室里检测了嗜热四膜虫(Tetrahymena ther- mophila)编码核糖体RNA的线性rDNA分子的端粒 序列,结果表明是由核苷酸六聚体(CCCCAA/ GGGGTI')连续重复构成,重复的次数在20—70之 间;进一步研究还发现端粒重复序列具有方向性,即 在端粒的3 7端由(TYGGGG)重复序列构成(称为G 链),5 7端由(CCCCAA)重复序列构成(称为C链)。 除此之外,Blackburn还揭示了四膜虫端粒的重复序 列具有不连续性,这种不连续性的存在有助于对不 同的端粒进行标记,但是其意义还不完全清楚[3】。
’北京大学医学部八年制临床医学专业2004级学生 △通讯作者
万方数据
生堡型堂进屋!QBiblioteka Q笙筮堡!鲞筮!塑其中,前导链(1eading strand)连续合成,随从链(1ag- sing strand)先分段沿5 7_+3’方向合成冈崎片段 (Okazaki fragment),然后切去各段引物,再由DNA 连接酶将冈崎片段连接成完整的DNA链。分析上 述过程时不难发现,由于DNA聚合酶不能催化
的RNA片段为模板来合成端粒的‘91。至此,嗜热四 膜虫端粒合成的机制已经非常清楚了(图5)。
I
2
,
‘,}酣^儿粕E鲋E
图5端粒酶催化端粒延伸模型 嗜热四膜虫端粒G链上带有一个13个碱基的悬突(0velllng) (1)端粒酶识别端粒底物末端的1TGGGG重复序列,并与端粒 酶RNA上的5"-CAACCCCAA-3 7模板序列配对;(2)合成TrG序 列;(3)端粒酶移位使末端的TrGGGGTIV,重新与模板序列配 对;(4)继续延伸端粒,复制模板序列完成TTGGGGTrG序列的 合成。(引自参考文献Is J)
与此同时,上世纪70年代Szostak在哈佛大学 建立了自己的实验室,试图在酵母中构建一种人工 线性DNA分子,来研究酵母(Yeast)染色体同源重 组机制(homologous recombination)。但当他用限制 性内切酶酶切酵母的环形质粒时,由于存在“黏”性 末端,酶切后形成的线性DNA分子很快与酵母的染