第二章 核酸的化学
生化第二章核酸的结构和功能
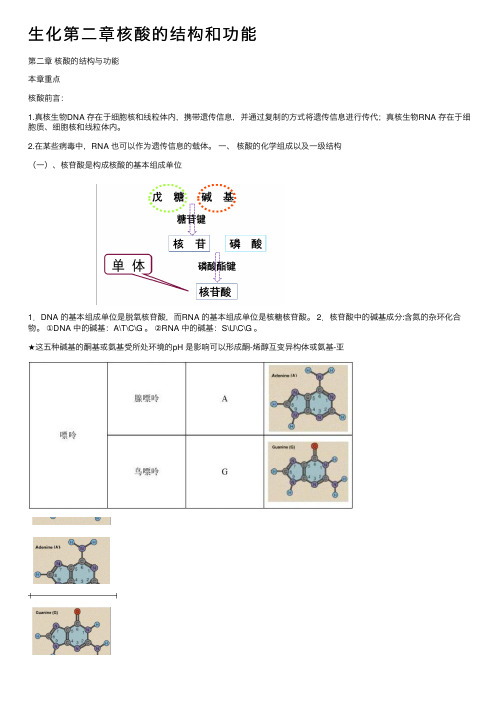
⽣化第⼆章核酸的结构和功能第⼆章核酸的结构与功能本章重点核酸前⾔:1.真核⽣物DNA 存在于细胞核和线粒体内,携带遗传信息,并通过复制的⽅式将遗传信息进⾏传代;真核⽣物RNA 存在于细胞质、细胞核和线粒体内。
2.在某些病毒中,RNA 也可以作为遗传信息的载体。
⼀、核酸的化学组成以及⼀级结构(⼀)、核苷酸是构成核酸的基本组成单位1.DNA 的基本组成单位是脱氧核苷酸,⽽RNA 的基本组成单位是核糖核苷酸。
2.核苷酸中的碱基成分:含氮的杂环化合物。
①DNA 中的碱基:A\T\C\G 。
②RNA 中的碱基:S\U\C\G 。
★这五种碱基的酮基或氨基受所处环境的pH 是影响可以形成酮-烯醇互变异构体或氨基-亚2.核糖①β-D-核糖:C-2’原⼦上有⼀个羟基。
②β-D-脱氧核糖:C-2’原⼦上没有羟基☆脱氧核糖的化学稳定性⽐核糖好,这使DNA成为了遗传信息的载体。
3.核苷①核苷②脱氧核苷③核糖的C-1’原⼦和嘌呤的N-9原⼦或者嘧啶的N-1原⼦通过缩合反应形成了β-N-糖苷键。
在天然条件下,由于空间位阻效应,核糖和碱基处在反式构象上。
3.核苷酸的结构与命名①核苷或脱氧核苷C-5’原⼦上的羟基可以与磷酸反应,脱⽔后形成磷酸键,⽣成核苷酸或脱氧核苷酸。
②根据连接的磷酸基团的数⽬不同,核苷酸可分为核苷⼀磷酸(NMP)、核苷⼆磷酸(NDP)、核苷三磷酸(NTP)。
③⽣物体内游离存在的多是5’核苷酸★细胞内⼀些参与物质代谢的酶分⼦的辅酶结构中都含有腺苷酸,如辅酶Ⅰ(NAD+),它们是⽣物氧化体系的重要成分,在传递质⼦或电⼦的过程中具有重要的作⽤。
(⼆)、DNA是脱氧核糖核苷酸通过3’,5’-磷酸⼆酯键连接形成的⼤分⼦1.脱氧核糖核苷三磷酸C-3’原⼦的羟基能够与另⼀个脱氧核糖核苷三磷酸的α-磷酸基团缩合,形成了⼀个含有3’,5’-磷酸⼆酯键的脱氧核苷酸分⼦。
2.脱氧核苷酸分⼦保留着C-5’原⼦的磷酸基团和C-3’原⼦的羟基。
《生物化学》第二章核酸化学及答案

第二章核酸化学《生物化学》一、选择题1.自然界游离核苷酸中,磷酸最常见是位于:A.戊糖的C-5′上B.戊糖的C-2′上C.戊糖的C-3′上D.戊糖的C-2′和C-5′上E.戊糖的C-2′和C-3′上2.可用于测量生物样品中核酸含量的元素是:A.碳B.氢C.氧D.磷E.氮3.下列哪种碱基只存在于RNA而不存在于DNA:A.尿嘧啶B.腺嘌呤C.胞嘧啶D.鸟嘌呤E.胸腺嘧啶4.核酸中核苷酸之间的连接方式是:A.2′,3′磷酸二酯键B.糖苷键C.2′,5′磷酸二酯键D.肽键E.3′,5′磷酸二酯键5.核酸对紫外线的最大吸收峰在哪一波长附近?A.280nm B.260nm C.200nm D.340nm E.220nm6.有关RNA的描写哪项是错误的:A.mRNA分子中含有遗传密码B.tRNA是分子量最小的一种RNAC.胞浆中只有mRNAD.RNA可分为mRNA、tRNA、rRNAE.组成核糖体的主要是rRNA7.大部分真核细胞mRNA的3′-末端都具有:A.多聚A B.多聚U C.多聚T D.多聚C E.多聚G8.DNA变性是指:A.分子中磷酸二酯键断裂B.多核苷酸链解聚C.DNA分子由超螺旋→双链双螺旋D.互补碱基之间氢键断裂E.DNA分子中碱基丢失9.DNA Tm值较高是由于下列哪组核苷酸含量较高所致?A.G+A B.C+G C.A+T D.C+T E.A+C10.某DNA分子中腺嘌呤的含量为15%,则胞嘧啶的含量应为:A.15% B.30% C.40% D.35% E.7%二、填空题1.核酸完全的水解产物是________、_________和________。
其中________又可分为________碱和__________碱。
2.体内的嘌呤主要有________和________;嘧啶碱主要有_________、________和__________。
某些RNA分子中还含有微量的其它碱基,称为_________。
生物化学第二章核酸化学

核酸分类及命名规则
核酸可分为DNA和RNA两大类,根据来源不同可分为基因组DNA、病毒DNA、mRNA、tRNA、 rRNA等。
核酸的命名通常包括种类、来源和特定序列信息,如人类基因组DNA可命名为hgDNA,mRNA可命 名为信使RNA等。
02
DNA结构与性质
DNA双螺旋结构模型
DNA由两条反向平行的多核苷酸链 组成,形成右手螺旋结构。
长约21nt的双链RNA,可引导RISC复合物识别并切割靶mRNA,实现基因沉默。
其他小分子RNA
如piRNA、snoRNA等,在基因表达调控、RNA修饰等方面发挥作用。
04
核酸理化性质与分离纯化方法
核酸溶解度和沉淀条件
溶解度
核酸在不同溶剂中的溶解度不同,一般易溶于水,难溶于乙醇、乙醚等有机溶 剂。其溶解度受温度、pH、离子强度等因素的影响。
非同源重组
发生在非同源序列之间的重组过程。这种重 组不依赖于序列之间的相似性,而是通过一 些特殊的蛋白质和酶的作用来实现DNA片 段的连接。非同源重组可能导致基因的重排 和染色体的不稳定,进而对生物体产生遗传 影响。
07
总结与展望
核酸化学领域重要成果回顾
核酸结构与功能研
究
揭示了DNA双螺旋结构和RNA多 种功能,阐明了遗传信息存储、 传递和表达机制。
05
核酸酶及其作用机制
限制性内切酶和外切酶作用方式
限制性内切酶
识别DNA分子中的特定核苷酸序 列,并在该序列内部进行切割, 产生特定的DNA片段。
外切酶
从DNA或RNA链的末端开始,逐 个水解核苷酸,释放单个的核苷 酸或寡核苷酸。
DNA连接酶在基因工程中应用
连接DNA片段
第二章 核酸的结构与功能

核酸的结构与功能
❖ 1868年,瑞士外科医生Fridrich从外科手术绷带上的脓细胞的细 胞核中分离出一种溶于碱而不溶于酸的酸性有机化合物,其分子 中含磷2.5%、含氮14%,该物质被命名为核酸。
❖ 根据核酸分子中所含戊糖的差别: (一)脱氧核糖核酸(DNA):主要存在于细胞核中(真核细胞的 线粒体中也存在不少量的DNA),携带着决定个体基因型的遗传信 息,是遗传信息的贮存和携带者; (二)核糖核酸(RNA):主要存在于细胞核和细胞质中,参与细
比DNA复制得多,这与它的功能多样化密切相关。
一、mRNA是蛋白质合成中的模板
❖ 1960年,Jacob 和 Monod 等人用放射性核素示踪实验证实: 一类大小不同的RNA才是细胞内合成蛋白质的真正模板,于 1961年首先提出了信使RNA(mRNA)这个概念。
❖ 在各种RNA分子中,mRNA约占细胞内RNA总量的2~5%,种类 最多,分子大小相差很大;
N H
❖DN生称AN物为稀体有的D碱N基A8 N和79NH。RN45 AN36分12 子N 中NH2还含有一些65含1N4 3量2N 很O 少H的3C碱基65 1,N4 32
N
O
鸟嘌呤
RNA
胞嘧啶
胸腺嘧啶
5´
HOCH2
4´ H
OH O
H 1´
H
H
3´
2´
OH OH
β-D-核糖(构成RNA)
5´
HOCH2
遗传的相对稳定性,又可发生各种重组和突变,适应环境的 变迁,为自然选R型择细提菌供:无机毒会型。肺炎球菌
S型细菌:有毒型肺炎球菌
肺炎球菌转化实验
第三节
RNA 的结构与功能
❖ RNA和蛋白质共同担负着基因的表达和表达调控功能。 ❖ RNA通常以单链形式存在,但可通过链内的碱基配对形成
第二章 核酸的化学3

(四)核酸的变性
2.热变性和 热变性和Tm: 热变性和 加热DNA的稀盐溶液,达到一定温度后在 的稀盐溶液, 吸光度骤 加热 的稀盐溶液 达到一定温度后在260nm吸光度骤 吸光度 然增加,表明两条链开始分开,吸光度增加约40%后变化趋 然增加,表明两条链开始分开,吸光度增加约 后变化趋 于平坦,说明两条链已完全分开。这表明DNA变性是一个突 于平坦,说明两条链已完全分开。这表明 变性是一个突 变过程,类似结晶的熔解,引起DNA变性的温度称为熔点, 变性的温度称为熔点, 变过程,类似结晶的熔解,引起 变性的温度称为熔点 表示。 用Tm表示。 表示 Tm:通常把ε(p)达最高时的 : 达最高时的1/2温度叫做 温度叫做Tm。凡分子中 、 温度叫做 。凡分子中G、 C含量高的 含量高的Tm高。一般 值在70-85℃之间。 含量高的 高 一般DNA的Tm值在 的 值在 ℃之间。 3.影响 影响Tm的因素: 的因素: 影响 的因素 G-C对含量:G-C含量越高 对含量: 含量越高Tm愈高,经验公式(G+C)%= 愈高, 对含量 含量越高 愈高 经验公式( ) (Tm-69.3)×2.44 ) 溶液的离子强度:离子强度较低的介质中Tm较低。 较低。 溶液的离子强度:离子强度较低的介质中 较低 溶液的pH值 完全变性; 溶液的 值: pH>11.3时,DNA完全变性; pH﹤5. 0时, > 时 完全变性 ﹤ 时 DNA易脱嘌呤。 易脱嘌呤。 易脱嘌呤 变性剂:甲酰胺、尿素、甲醛等可破坏H键 变性剂:甲酰胺、尿素、甲醛等可破坏 键,妨碍碱基堆积 下降。 使Tm下降。 下降
核酸的光吸收值比各核苷酸光吸收值的和少30~40%,当核 酸变性或降解时光吸收值显著增加(增色效应 增色效应),但核酸 增色效应 复性后,光吸收值又回复到原有水平(减色效应 减色效应)。 减色效应
生物化学第二章 核酸化学

李纳斯·鲍林(Linus Pauling)
DNA的二级结构是三螺旋?
1962年,三人分享诺贝尔生理医学奖
DNA的二级结构是双螺旋
(1)DNA分子由两条多聚 脱氧核糖核苷酸链(简称 DNA单链)组成。两条链 沿着同一根轴平行盘绕, 形成右手双螺旋结构。 螺旋中的两条链方向相 反,即其中一条链的方 向为5′→3′,而另一条链 的方向为3′→5′,螺旋结 构上有大沟和小沟。
两类 核酸在分子组成上的异同点
组分 磷酸 戊糖 碱 嘌呤 基 嘧啶
RNA
DNA
磷酸Βιβλιοθήκη 核糖脱氧核糖AG
U
C
T
核苷酸的衍生物
ⅰ ATP (腺嘌呤核糖核苷三磷酸)
d d d
核苷酸及其多磷酸化合物
ⅱ 环腺苷酸(cAMP)和环鸟苷酸 (cGMP)
•这两种环核苷酸在细胞代谢调节中具有重要作 用,是传递激素作用的媒介物。称为二级信使。
(2)两条链上的碱基通 过氢键相结合,形成碱 基对。碱基的相互结合 具有严格的配对规律, 即A与T结合,G与C结 合,碱基之间的这种一 一对应关系,称为碱基 互补配对原则。A和T之 间形成两个氢键,G与C 之间形成三个氢键。
碱基互补配对
A
T
C
G
(3)嘌呤和嘧啶碱基位于螺旋 的内侧,磷酸和脱氧核糖基 位于螺旋外侧,彼此以3 ’-5’ 磷酸二酯键连接,形成DNA 分子的骨架。碱基环平面与 螺旋轴垂直,糖基环平面与 碱基环平面成90°角。
级结构的可能性较小。
* mRNA的功能 把DNA所携带的遗传信息,按碱基互补
配对原则,抄录并传送至核糖体,用以决定 其合成蛋白质的氨基酸排列顺序。
原核细胞
真核细胞
细胞质
第二章-核酸化学

2021/4/9
42
2021/4/9
酵母tRNA Ala 的二级结构
氨基酸臂 D臂(二氢尿嘧啶臂) D环 TC臂 TC环 额外环 AC臂(反密码臂) AC环
43
三级结构 —— 倒L型
2021/4/9
44
2021/4/9
45
二、rRNA的分子结构
原核生物rRNA有3类:5S、16S、23S 真核生物rRNA有4类: 5S、5.8S、18S、28S
核苷酸
Phosphates
戊糖(核糖、脱氧核糖)
Nucleotides
核苷 Pentoses
Nucleosides
碱基(嘌呤、嘧啶)
2021/4/9
Nitrogenous Base
4
㈠ 碱 基 Nitrogenous Base
主要包括嘌呤碱和嘧啶碱。
2021/4/9
5
嘌呤(purine)
N 7
许多rRNA的一级结构及由一级结构推导出来的 二级结构都已阐明,但是对许多rRNA的功能迄今 仍不十分清楚。 已有一些rRNA具有酶的活性,称为核酶 (ribozyme)。
2021/4/9
46
2021/4/9
5sRNA的二级结构
47
三、mRNA的分子结构
顺反子(cistron):一个基因就是一个顺反子。 原核生物的mRNA一般是多顺反子。 真核生物的mRNA一般是单顺反子。
1953年,Watson和Click提出DNA双螺旋模型。
1958 Crick提出遗传信息传递的中心法则。
2021/4/9
2
核酸的种类和分布
1. 脱氧核糖核酸(deoxyribonucleic acid, DNA) 原核: 裸露的DNA分子集中于核区 真核: 细胞核DNA:与组蛋白、非组蛋白形成染色体
第二章核酸化学下ppt课件

3'
3' U attacks 3' end of IVS
19-IVS 作为催化剂
GOH-3'
GpGpGpApGpGOH-5'
CpCpCpCpCOH "C5"
CpCpCpCpCpCOH "C6"
GOH-3'
5'-CpCpCpCpCOH-3' GpGpGpApGpGOH-5'
CpCpCpCOH "C4"
• tRNA主要作用是将氨基酸转运到核糖体mRNA复合物的相应位置用于蛋白质合成
• 20种基本氨基酸每一种都至少有一种tRNA • tRNA分子较小,平均沉降系数为4S • 大多数tRNA分子具有类似的三叶草二级结
构
tRNA三叶草二级结构
3’-
受体端
76 75
(acceptor
stem)
74 73
1
1
ddTTP
CAddT CATddT CATTACGddT
8
T
7
G
6
C
5
A
4
T
3
T
2
A
1
C
DNA测序技术的应用
• RNA序列的测定,将RNA反转录成互补 DNA(cDNA) ,测定cDNA序列后即可推 断出RNA的序列
• 蛋白质的氨基酸序列的测定,也可以通过 测定DNA序列,然后用遗传密码来推断。
核酸的性质—结构稳定性
• 碱基对间的氢键
• 碱基堆积—碱基堆积力对维持核酸的空间 结构起主要作用
• 环境中的正离子—环境中的Na+、K+、 Mg2+、Mn2+等离子,可消除核酸中磷酸基 间的静电斥力,对核酸结构的稳定有重要 作用
第2章 核酸的结构与功能

第二章核酸的结构和功能核酸是以核苷酸为基本组成单位的线性多聚生物信息分子。
分为DNA和RNA两大类。
其化学组成见下表:DNA RNA碱基①嘌呤碱 A、G A、G②嘧啶碱 C、T C、U戊糖β-D-2 脱氧核糖β-D-核糖磷酸磷酸磷酸碱基与戊糖通过糖苷键相连,形成核苷。
核苷的磷酸酯为核苷酸。
根据核苷酸分子的戊糖种类不同,核苷酸分为核糖核苷酸与脱氧核糖核苷酸,前者是RNA的基本组成单位,后者为DNA的基本组成单位,核酸分子中核苷酸以3’,5’-磷酸二酯键相连,形成多核苷酸链,是核酸的基本结构。
多核苷酸链中碱基的排列顺序为核酸的一级结构。
多核苷酸链的两端分别称为3’-末端与5’-末端。
DNA的二级结构即双螺旋结构的特点:⑴两条链走向相反,反向平行,为右手螺旋结构;⑵脱氧核糖和磷酸在双螺旋外侧,碱基在内侧;⑶两链通过氢键相连,必须A与T、G与C配对形成氢键,称为碱基互补规律。
⑷大(深)沟,小(浅)沟。
⑸螺旋一周包含10个bp,碱基平面间的距离为0.34nm,螺旋为3.4nm,螺旋直径2nm;⑹疏水作用。
氢键及碱基平面间的疏水性堆积力维持其稳定性。
DNA的基本功能是作为遗传信息的载体,并作为基因复制转录的模板。
mRNA分子中有密码,是蛋白质合成的直接模板。
真核生物的mRNA一级结构特点:5’-末端“帽”,3’-末端“尾”。
tRNA在蛋白质合成中作为转运氨基酸的载体,其一级结构特点:含有较多的稀有碱基;3’-CCA-OH,二级结构为三叶草形结构。
rRNA与蛋白质结合构成核蛋白体,作为蛋白质合成的“装配机”。
细胞的不同部位还存在着许多其他种类小分子RNA,统称为非mRNA小RNA(snmRNAs),对细胞中snmRNA 种类、结构和功能的研究称为RNA组学。
具有催化作用的某些小RNA称为核酶。
碱基、核苷、核苷酸及核酸在260nm处有最大吸收峰。
加热可使DNA双链间氢键断裂,变为单链称为DNA变性。
DNA变性时,OD260增高。
生物化学试题 核酸化学

第二章核酸化学.三、典型试题分析1. 下列几种DNA分子的碱基组成比例不同,哪一种DNA的Tm值最低(1999年生化试题)A. DNA中A-T占15%B.DNA中G-C占25%C. DNA中G-C占40%D.DNA中A-T占80%E. DNA中G-C占55%[答案] D2. 核酸的各基本单位之间的主要连接键是(2000年生化试题)A.二硫键B.糖苷键C.磷酸二酯键D.肽键E,氢键[答案) C3.DNA的二级结构是:A.α—螺旋B.β-片层C.β—转角D.超螺旋结构E,双螺旋结构[答案) E4. DNA的热变性特征是A. 碱基间的磷酸二酯键断裂B.一种三股螺旋的形成C.黏度增高D.融解温度因G-C对的含量而异E.在260nm处的光吸收降低[答案] D5. 下列关于DNA碱基组成的叙述,正确的是A. 不同生物来源的DNA碱基组成不同B,同一生物不同组织的DNA碱基组成不同C. 生物体碱基组成随着年龄变化而改变D.A和C含量相等E.A+T=G+C[答案] A6,DNA受热变性时(士998年硕士研究生入学考试题)A. 在260nm波长处的吸光度下降B,多核苷酸链断裂成寡核苷酸链 C. 碱基对可形成氢键D,加入互补RNA链,再冷却,可形成DNA/RNA杂交分子E. 溶液黏度增加[答案] D7,在核酸中占9%"-11%,且可用之计算核酸含量的元素是(1997年硕士研究生入学考试题)A. 碳B,氧C.氮D.氢E.磷[答案] E8,下列关于B-DNA双螺旋结构模型的叙述中哪一项是错误的?A. 两条链方向相反B,两股链通过碱基之间的氢键相连C.为右手螺旋,每个螺旋为10个碱基对D.嘌呤碱和嘧啶碱位于螺旋外侧E.螺旋的直径为20A[答案) D四、测试题(一)A型题1.从小鼠的一种有荚膜的致病性肺炎球菌中提取出的DNA,可使另种无荚膜、不具有致病性的肺炎球菌转变为有荚膜并具致病性的肺炎球菌A.DNA是遗传物质,蛋白质是遗传信息的体现者B,DNA是遗传信息的体现者,蛋白质是遗传物质C.DNA与蛋白质均是遗传物质D.RNA是遗传物质,DNA和蛋白质是遗传信息的体现者E.DNA和蛋白质是遗传物质,RNA是遗传信息的体现者2.核酸中一般不含有的元素是A.C B.H C.O D.P E,S3,在核酸中占9%'--11%且可用之计算核酸含量的元素是A.C B.H C.O D.P E.S4.A(腺嘌呤)与G(鸟嘌呤)在结构上的差别是A,A的C6上布羟基,G的C6上有氨基B.A的C6上有羟基,G的C2上有甲基C,A的C6上有甲基,G的C6上有羰基D.A的C2上有氨基,G的C2上有羟基E.A的C6上有氨基,G的C2上有氨基5,T(胸腺嘧啶)与U(尿嘧啶)在结构上的差别是A.T的C2上有氨基,U的C2上有O ·B.T的C5上有甲基,U的C5上无甲基C.T的C4上有氨基,U的C4上有OD.T的Cl上有羟基,U的C1上无羟基E.T的C5上有羟甲基,U的C5上无羟甲基6,通常既不见于DNA又不见于RNA的碱基是A. 腺嘌呤B.黄嘌呤巳鸟嘌呤D,胸腺嘧啶E,尿嘧啶7,自然界游离核苷酸中的磷酸最常位于A. 戊糖的C-2’上B.戊糖的C-3’上C.戊糖的C-5’上D.戊糖的C-2’及C-3’ E. 戊糖的C-2’及C-5’上8,假尿嘧啶核苷的糖苷键是·A.C-C键B.C-N键C.N-N键D.C-H键E.N-H键9,核苷酸中碱基(N)、戊糖(R)和磷酸(P)之间的连接关系是A. N-R-PB. N-P-RC. R-N-PD.P-N-R E.R-P-P-N10,脱氧胸苷的英文简写符号为A.AdR B,CdR C.UdR D. TdR E. CdR11,CR表示A. 脱氧胞苷B.胞苷 C. 腺苷D.脱氧腺苷E,脱氧核糖12,含有稀有碱基较多的核酸是A.rRNA B.tRNA C.mRNA D.hnRNA E。
生科第二章-核酸化学

组 蛋 白 与 DNA的 结 合
➢组蛋白核心(héxīn): H2B, H2A, H3, H4。H1 组蛋白在核小体之间。
第三十四页,共74页。
DNA的存在(cúnzài)形式
第三十五页,共74页。
染色体包装------多级螺旋(luóxuán)模型
压缩倍数 7
6
40
DNA → 核小体 → 螺线管 →
第十九页,共74页。
3.2 核酸(hé suān)的一级结构(primary structure)
一级结构-----核酸(hé suān)分子中核苷酸的排列 顺序及连接方式。核苷酸的排列顺序代表了遗传信 息。
➢碱基序列(base sequence)即为DNA的一级结构。通 常碱基序列由DNA链的5 →3 方向(fāngxiàng)写。 ➢n个核苷酸组成的DNA链中可能有的不同序列总数为 4n。
DNA double helix类型
bp/turn
D vertical rise/bp
11
2.3
0.255
10
2.0
0.34
12
1.8
0.37
direction 右 右 左
第二十八页,)结构(tertiary structure)
1. 环状DNA的超螺旋结构 DNA双螺旋进一步扭曲(niǔ qǔ)成超螺旋构成三级结 构。 一段双螺旋(luóxuán)在螺旋(luóxuán)均已形成的 情况下,双链环不发生进一步扭曲,称松弛环形 DNA。 若将线形DNA的螺旋(luóxuán)先拧松两周再连接 成环时,解链部分形成突环称解链环型DNA。
第二十页,共74页。
核苷酸的连接方式
1、核酸的基本结构(jiégòu)单位:核苷酸
核酸的化学组成

O HO P OH2C OH OH OH 核 核 核三 三 O B O HO P OH2C OH OH 脱脱 核核 核三 三 O B
B=三腺腺,鸟腺腺,胞胞胞,尿胞胞尿尿三尿胞 三
2.核苷酸的衍生物
(1) ATP (腺嘌呤核糖核苷三磷酸) (腺嘌呤核糖核苷三磷酸 腺嘌呤核糖核苷三磷酸)
ATP是生物体内分布最广和最重要的一种核苷酸衍生物。它的 是生物体内分布最广和最重要的一种核苷酸衍生物。 是生物体内分布最广和最重要的一种核苷酸衍生物 结构如下: 结构如下:
HOCH2 H H OH O H H OH H OH H OH HOCH2 H O H H OH
D-核核 核
D-2-脱脱核核 脱
1.核苷酸
(3)核苷 nucleoside
核苷由戊糖和碱基缩合而成,嘌呤的N9或嘧啶的 核苷由戊糖和碱基缩合而成,嘌呤的N9或嘧啶的 N9 O N1与戊糖 与戊糖C -OH以 糖苷键相连接。 N1与戊糖C-1’-OH以C-N糖苷键相连接。
二、核酸的生物学功能 (一)DNA是主要的遗传物质 一 DNA是主要的遗传物质
S
1944年,O.T.Avery(美) 1944年 O.T.Avery( 肺炎链球菌的转化实验, 转化实验 肺炎链球菌的转化实验,首次 证明DNA DNA是细菌遗传性状的转化 证明DNA是细菌遗传性状的转化 因子。 因子。 十年后证明DNA是遗传物质 十年后证明DNA是遗传物质 DNA
R
S+R S菌体的DNA + R 菌体的DNA
S
(一)DNA是主要的遗传物质 一 DNA是主要的遗传物质
1952年,美国冷泉港 年 Hershey-Chase 噬菌体浸染细菌的实验。 噬菌体浸染细菌的实验。
生物化学重点_第二章核酸化学

生物化学要点 _第二章核酸化学第二章核酸化学一、核酸的化学构成 :1、含氮碱 : 参加核酸与核苷酸构成的含氮碱主要分为嘌呤碱与嘧啶碱两大类。
构成核苷酸的嘧啶碱主要有三种——尿嘧啶 (U) 、胞嘧啶 (C)与胸腺嘧啶 (T),它们都就是嘧啶的衍生物。
构成核苷酸的嘌呤碱主要有两种——腺嘌呤 (A) 与鸟嘌呤 (G),它们都就是嘌呤的衍生物。
2、戊糖 :核苷酸中的戊糖主要有两种,即β-D- 核糖与β-D-2- 脱氧核糖 ,由此构成的核苷酸也分为核糖核苷酸与脱氧核糖核酸两大类。
3、核苷 :核苷就是由戊糖与含氮碱基经脱水缩合而生成的化合物。
由“罕有碱基”所生成的核苷称为“罕有核苷”。
如 :假尿苷 (ψ)二、核苷酸的构造与命名:核苷酸就是由核苷与磷酸经脱水缩合后生成的磷酸酯类化合物,包含核糖核苷酸与脱氧核糖核酸两大类。
核苷酸又可按其在 5’位缩合的磷酸基的多少 ,分为一磷酸核苷 (核苷酸 )、二磷酸核苷与三磷酸核苷。
别的 ,生物体内还存在一些特别的环核苷酸 ,常有的为环一磷酸腺苷 (cAMP) 与环一磷酸鸟苷 (cGMP),它们往常就是作为激素作用的第二信使。
核苷酸往常使用缩写符号进行命名。
第一位符号用小写字母 d 代表脱氧 ,第二位用大写字母代表碱基 ,第三位用大写字母代表磷酸基的数量 ,第四位用大写字母 P 代表磷酸。
三、核酸的一级构造 :核苷酸经过 3’ ,5-磷’酸二酯键连结起来形成的不含侧链的多核苷酸长链化合物就称为核酸。
核酸拥有方向性,5’-位上拥有自由磷酸基的尾端称为5’-端,3’-位上拥有自由羟基的尾端称为3’-端。
DNA 由 dAMP 、dGMP、dCMP 与 dTMP 四种脱氧核糖核苷酸所构成。
DNA 的一级构造就就是指 DNA 分子中脱氧核糖核苷酸的摆列次序及连结方式。
RNA由AMP,GMP,CMP,UMP 四种核糖核苷酸构成。
四、 DNA 的二级构造 :DNA 双螺旋构造就是 DNA 二级构造的一种重要形式 ,它就是 Watson与 Crick 两位科学家于 1953 年提出来的一种构造模型 ,其主要实验依照就是 Chargaff 研究小组对 DNA 的化学构成进行的剖析研究,即 DNA 分子中四种碱基的摩尔百分比为 A=T 、 G=C、 A+G=T+C(Chargaff 原则 ),以及由 Wilkins 研究小组达成的 DNA晶体 X 线衍射图谱剖析。
第二章(2)核酸的化学(性质)

1.核酸的两性性质及等电点
分子中既含有酸性基团(磷酸基),也含有碱性基 团(氨基),因而核酸具有两性性质. 因而核酸具有两性性质. 因而核酸具有两性性质 由于核酸分子中的磷酸是一个中等强度的酸,而碱性 (氨基)是一个弱碱,所以核酸的等电点比较低。 核酸的等电点比较低。 核酸的等电点比较低 如DNA的等电点为4~4.5,RNA的等电点为2~2.5。 RNA的等电点比DNA低的原因:是RNA分子中2′-OH 通过氢键促进了磷酸基上质子的解离,而DNA没有这种 作用。
影响复性速度的因素:
1、分子量越大,复性越难。 分子量越大,复性越难。 单链片段浓度越大,随机碰撞的频率越高,复性越容易。 2、单链片段浓度越大,随机碰撞的频率越高,复性越容易。 片断内的重复序列多,则容易形成互补区,复性较快。 3、片断内的重复序列多,则容易形成互补区,复性较快。 维持溶液一定的离子强度,可加快复性速度。 4、维持溶液一定的离子强度,可加快复性速度。 此外,DNA的复性也与它本身的组成和结构有关 的复性也与它本身的组成和结构有关。 此外,DNA的复性也与它本身的组成和结构有关。
HOCH
2
O H
OH H
HOCH
2
O H
OH H
H H OH D-核核 核
H H OH
OH
H
D-2-脱脱核核 脱
2.核酸的水解
(1)酸或碱水解
核酸分子中的磷酸二酯键可在 酸或碱性条件下水解切断。 DNA和RNA对酸或碱的耐受程度 有很大差别。例如,在0.1 mol/L NaOH溶液中,RNA几乎可以完全水解, DNA在同样条件下则不受影响。
核酸外切酶的作用方式是从多聚核苷酸链的一端(3′-端或5′核酸外切酶的作用方式是从多聚核苷酸链的一端(3′-端或5′的作用方式是从多聚核苷酸链的一端 5′ 端)开始,逐个水解切除核苷酸;核酸内切酶的作用方式刚好和外切 开始,逐个水解切除核苷酸;核酸内切酶的作用方式刚好和外切 酶相反,它从多聚核苷酸链中间开始,在某个位点切断磷酸二酯键。 酶相反,它从多聚核苷酸链中间开始,在某个位点切断磷酸二酯键。
第二章3-核酸化学PPT课件

▪
戊糖在外,双螺旋每转一
小 沟
周 为10碱基对(bp)
▪
A型结构
▪
碱基平面倾斜20º,螺旋
变粗变短,螺距2~3nm。
2.0 nm
大 沟
DNA的三级结构
➢DNA的三级结构:指双螺旋进一步扭曲 形成的超螺旋。 ➢包括:线状DNA形成的纽结、超螺旋和 多重螺旋、环状DNA形成的结、超螺旋 和连环等
线状DNA形成的超螺旋
▪ 多核苷酸链均有5’-末端和3’-末端 ▪ 核酸的碱基顺序本身就是遗传信息存储的分子形
式。生物界物种的多样性即寓于DNA分子中四种 核苷酸千变万化的不同排列组合之中。
二、DNA的二级结构
DNA的双螺旋模型
▪ 1953年,J. Watson和F. Crick 在前人研究工作的基础上, 根据DNA结晶的X-衍射图谱和 分子模型,提出了著名的
➢DNA分子中具有特定生物学功能的片 段称为基因(gene)。
➢一个生物体的全部DNA序列称为基因 组(genome)。
RNA的结构与功能
▪ 一、结构特点
1. 碱基组成 A、G、C、U (A= U/G=C)
稀有碱基较多,稳定性较差,易水解 2. 多为单链结构,少数局部形成螺旋 3. 分子较小 4. 分类 ➢mRNA ➢tRNA ➢rRNA
三、核酸的变性
▪ 稳定核酸双螺旋次级键断裂,空间结构破坏,变成单链结 构的过程。核酸的的一级结构(碱基顺序)保持不变。
▪ 变性表征 生物活性部分丧失、粘度下降、浮力密度升高、紫外
吸收增加(增色效应) ▪ 变性因素
pH(>11.3或<5.0) 变性剂(脲、甲酰胺、甲醛) 低离子强度 加热
➢核酸是存在于细胞中的一类大分子酸 性物质,包括核糖核酸(RNA)和脱氧 核糖核酸(DNA)两大类。
生物化学讲义第二章核酸化学

核酸的结构与功能【目的和要求】1. 熟悉核酸的种类、分布和主要的生物学功能。
2.掌握核酸的化学组成、核苷酸的连接方式。
3.归纳区分两类核酸在化学组分上的异同点。
4.说出DNA二级结构的模型及其主要特点。
5.简述RNA分子组成和结构的特点。
6.简述三种RNA结构特点和主要功能。
7.了解核酸重要的理化特性及其在医学上的应用。
8.能说出生物体内重要的单核苷酸及其生化功能。
【本章重难点】1.核酸的种类、分布和生物学功能。
2.核酸的化学组成。
3.DNA和RNA的分子结构与功能。
4.核酸的变性、复性及杂交。
5.生物体内重要的单核苷酸。
学习内容第一节核酸的化学组成第二节 DNA的分子结构第三节 RNA的分子结构第四节核酸的理化性质第一节核酸的化学组成一、核酸(nucleic acid)的分类、分布与生物学功能分类分布生物学功能核糖核酸(RNA)细胞质参与蛋白质的生物合成5 % 蛋白质合成的直接模板tRNA 15 % 活化与转运AArRNA 80 % 充当装配机,提供场所脱氧核糖核酸(DNA ) 核内、染色质遗传的物质基础** 基因 —— DNA 分子中的功能片段(决定遗传特性的碱基序列)。
二、核酸的分子组成1.核酸的元素组成:C.H.O.N.和P ;代表元素P ,平均含量9~10%。
2.核酸的基本组成单位:核苷酸(nucleotide )1)核苷酸的组成戊糖、碱基:核苷、核苷酸:核苷酸链:3/,5/-磷酸二酯键;3/-羟基端,5/-磷酸基端水解 水解 磷酸 戊糖(戊糖、脱氧戊糖)核酸 核苷酸核苷 嘧啶(C.T.U )碱基嘌呤(A.G)2)核苷酸的结构与命名3)核苷酸的功用3.两类核酸在分子组成上的异同点第二节 DNA 的分子结构一、DNA 的一级结构组成DNA 分子的基本单位是四种脱氧核苷酸:dAMP 、dCMP 、dGMP 和dTMP1.DNA 的碱基组成规律:Chargaff 规则:①同一生物不同组织的DNA 样品,其碱基成分含量相同。
核酸的化学

H
HOH2C 5′ O
4′ 3′
HOH2C 5′ O
4′ 3′ 2′
OH
1′
OH
OH
OH
OH
核 糖 尿苷
核 糖 假尿苷 (ψ)
(四)核苷酸
N H O
-
H
N
9
H
N
O
-
-
O —
P ‖ O
~O
—
P ‖ O
~O
—
-
O
-
N H 腺嘌呤
P HOH2C 5′ O OH ‖ 4′ 1′ O 3′ 2′ OH OH AMP ADP
O
5′
O=P—O—CH2 O O
1′ 3′
G
G
T
A
OH OH O
P
3′ 5′
P
P
OH
O=P—O—CH2 O
5′
T
O
-
3′
pGpTpAOH
pG-T-A pGTA
OH OH O
-
O=P—O—CH2 O
5′
A
O
3′
OH OH
(二)DNA的双螺旋结构
1953年,Watson 和Crick 提出。
1.双螺旋结构的主要依据 (1)Wilkins和Franklin发现不同来源的DNA纤维具有相 似的X射线衍射图谱。 (2)Chargaff发现DNA中A与T、C与G的数目相等。后 Pauling 和Corey发现A与T生成2个氢键、C与G生成3个 氢键。 (3)电位滴定证明,嘌呤与嘧啶的可解离基团由氢键连 接。
0.2 0.1 0 220
240
260 ¨³ ² ¤ /nm
生物化学第二章 核酸的结构

Na+,K+, Cs+ 窄深
宽深
Na+低盐
Li+
Na+, Mg++高 盐
宽中等 深 宽中等 深 平浅
窄中 等深 窄中 等深 窄深
• 真核生物染色体DNA是线型双链DNA。
• • 细胞器DNA、原核生物的染色体DNA、 质粒DNA都是环状双链
补充知识:
(一)三链DNA(triple helix DNA)
脱氧鸟嘌呤核苷酸(dGMP) 脱氧胞嘧啶核苷酸(dCMP) 脱氧胸腺嘧啶核苷酸(dTMP)
多磷酸核苷酸:NMP,NDP,NTP
N O
HO P O CH2O N
OH
AMP OH OH
NH2 N
N
N
O
O
HO P O P O CH2O N OH OH
ADP
OH OH
N
O
O
O
HO P O P O P O CH2O N OH OH OH
7)碱基环氮原子的烷基化反应
8)环外氨基的反应
9)环外氧的烷基化反应
(二)戊 糖
(三) 磷酸:
(四)核苷酸
1. 核苷(ribonucleoside)
碱基和核糖(脱氧核糖) 糖苷键
N-9 C-1'
N-1
NH 2
N
N
N
N
HOCH 2
9
O
1
HO
OH
NH2
N
O N1 HOCH2
O
1
OH H
• 核苷:腺苷,鸟苷 • 脱氧核苷:
定义: 核酸中核苷酸的排列顺序, 也称为碱基序列。
核苷酸的连接 以3',5'–磷酸二酯键连接
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
在生物体内,绝大多数超螺旋DNA以负超螺旋的形式存在,也就是说,一旦超螺旋解开,则会形成解链环形DNA,有利于DNA复制或转录。
螺旋具有相同的结构,但L值不同的分子称为拓扑异构体。DNA拓扑异构酶切断一条链或两条链,拓扑异构体可以相互转变。W的正表示双链闭环的螺旋圈在增加,W的负表示减少。L和T的正负表示螺旋方向,右手为正,左手螺旋为负;L值必定是整数。
一、转运RNA(transfer RNA,tRNA)
1.分子量最小的RNA,约占总RNA的15%。主要功能是在蛋白质生物合成过程中,起着转运氨基酸的作用。
2.1965年Holley等测定了酵母丙氨酸tRNA的一级结构,并提出二级结构模型。一级结构特点:核苷酸残基数在73~95;含有较多的稀有碱基(如mG、DHU等);5'-末端多为pG,3'- 末端都是-CCA。
核苷酸是两性电解质,有等电点。核苷酸有互变异构和紫外吸收。(含氧的碱基有酮式和烯醇式两种互变异构体,在生理pH条件下主要以酮式存在)
二、核苷酸的连接方式
RNA和DNA链都有方向性,从5'→ 3'。前一位核苷酸的3'- OH与下一位核苷酸的5'位磷酸基之间形成3',5'-磷酸二酯键,从而形成一个没有分支的线性大分子,两个末端分别称为5'末端和3'末端。大分子的主链由相间排列的戊糖和磷酸构成,而碱基可看作主链上的侧链基团,主链上的磷酸基是酸性的,在细胞pH下带负电荷;而碱基有疏水性。
第二章 核酸的化学
教学目标:
1.掌握DNA和RNA在化学组分、分子结构和生物功能上的特点。
2.掌握DNA双螺旋结构模型和t-RNA二级结构的要点,了解核酸的三级结构。
3.熟悉核酸的性质(一般性质、DNA热变性、复性与分子杂交)。
4.掌握基因组的概念,原核生物和真核生物基因组的特点。了解DNA测序物体内游离存在的多是5'- 核苷酸(如pA、pdG等)。常见的核苷酸为AMP、GMA、CMP、UMP。常见的脱氧核苷酸有dAMP、dGMA、dCMP、dTMP。AMP是一些重要辅酶的结构成分(如NAD+、NADP+、FAD等);环化核苷酸(cAMP/cGMP)是细胞功能的调节分子和信号分子。ATP在能量代谢中起重要作用。
1.环形DNA可形成超螺旋
当将线性过旋或欠旋的双螺旋DNA连接形成一个环时,都会自动形成额外的超螺旋来抵消过旋或欠旋造成的应力,目的是维持B构象。过旋DNA会自动形成额外的左手螺旋(正超螺旋),而欠旋形成额外的右手螺旋(负超螺旋)。
一段双螺旋圈数为10的B-DNA连接成环形时,不发生进一步扭曲,称松弛环形DNA(双螺旋的圈数=链绕数,即T=L,超螺旋数W=0;L=T+W),但将这一线形DNA的螺旋先拧松一圈再连接成环时,解链环形DNA存在的扭曲张力,可导致双链环向右手方向扭曲形成负超螺旋(T=10,L=9,W = -1)。
在DNA一级结构中,有一种回文结构的特殊序列,所谓回文结构即DNA互补链上一段反向重复顺序,正读和反读意义相同,经反折可形成“十字形”结构,在转录成RNA后可形成“发夹”样结构,有调控意义。
→ GCTA GTTCA CTC TGAAC AATT →
← CGAT CAAGT GAG ACTTG TTAA ←
tRNA中含有较多的稀有碱基(修饰碱基),多为甲基化的。
(三)核苷
是碱基和戊糖生成的糖苷。通过C1'- N9或C1'- N1糖苷键连接,用单字符表示,脱氧核苷则在单字符前加d。常见的修饰核苷有:次黄苷或肌苷为I、黄嘌呤核苷X、二氢尿嘧啶核苷D、假尿苷Ψ等。注意符号的意义,如m5dC。
RNA的C2'位羟基是游离的,是一个易发生不良反应的位置,它使RNA的化学性质不如DNA稳定,能较DNA产生更多的修饰组分。RNA的种类、大小、结构都比DNA多样化,按照功能的不同和结构的特点,RNA主要分为tRNA、rRNA和mRNA三类。此外,细胞的不同部位还存在着另一些小分子RNA,如核内小RNA(snRNA)、核仁小RNA(snoRNA)、胞质小RNA(scRNA)等,分别参与mRNA的前体(hnRNA)和rRNA的转运和加工过程。
2.真核细胞染色体
真核细胞DNA是线形分子,与组蛋白结合,其两端固定也形成超螺旋结构。DNA被紧密地包装成染色体来自三个水平的折叠:核小体、30nm纤丝和放射环。
核小体是染色体的基本结构单位,是DNA包装的第一步,它由DNA结合到组蛋白上形成复合物,在电镜下显示为成串的“念珠”状。组蛋白是富含精氨酸和赖氨酸的碱性蛋白质,其氨基酸序列在进化中是高度保守的。组蛋白有5种,H2A、H2B、H3和H4各两分子组成的八聚体是核小体核心颗粒,DNA缠绕其上,相邻核小体间的DNA称为连接DNA且结合H1。200 bpDNA的长度约为68nm,被压缩在10nm的核小体中。压缩比约为7。30nm纤丝是第二级压缩,每圈含6个核小体,压缩比是6。30nm螺旋管再缠绕成超螺旋圆筒,压缩比是40。再进一步形成染色单体,总压缩近一万倍。典型人体细胞的DNA理论长度应是180 cm,被包装在46个5μm的染色体中。
DNA分子很大,最小的病毒DNA约含5000b。1965年Holley用片段重叠法完成酵母tRNAala 76nt 序列测定;1977年Sanger利用双脱氧法(酶法)测定了φX174单链DNA5386b的全序列。1990年实施的人类基因组计划(HGP),用15年,投资30亿美元,完成人类单倍体基因组DNA3×109bp全序列的测定。该计划由美、英、日、法、德、中六国科学家合作,于2003年提前完成,生命科学进入后基因组时代,研究重点从测序转向对基因组功能的研究。
讨论:列表说明DNA和RNA在化学组成、分子结构和生物功能方面的主要特点。
第二节 DNA的分子结构
一、 DNA的一级结构 (primary stucture)
DNA的一级结构是指分子中脱氧核苷酸的排列顺序,常被简单认为是碱基序列(base sequence)。碱基序有严格的方向性和多样性。一般将5'- 磷酸端作 为多核苷酸链的“头”,写在左侧,如pACUGA( 5'→ 3') 。
氢键维持双螺旋的横向稳定。碱基对平面几乎垂直螺旋轴,碱基对平面间的疏水堆积力维持螺旋的纵向稳定。
4.碱基在一条链上的排列顺序不受限制。遗传信息由碱基序所携带。
5.DNA构象有多态性。
Watson和Crick根据Wilkins 和Franklin拍摄的DNA X-射线照片是相对湿度92%的DNA钠盐所得的衍射图,因此Watson-Crick 双螺旋结构称B-DNA。细胞内的DNA与它非常相似。另外还有A-DNA、C-DNA、D-DNA。
1.两条反向平行的多核苷酸链形成右手螺旋。一条链为5'→ 3',另一条为3'→ 5'。(某些病毒的DNA是单链分子ssDNA)
2.碱基在双螺旋内侧,A与T,G与C配对,A与T形成两个氢键,G与C形成三个氢键。糖基-磷酸基骨架在外侧。表面有一条大沟和一小沟。
3.螺距为3.4 nm,含10个碱基对(bp),相邻碱基对平面间的距离为0.34 nm。螺旋直径为2 nm。
四、DNA和基因组
1.DNA分子中的最小功能单位称作基因,为RNA或蛋白质编码的基因称结构基因,DNA中具调节功能而不转录生成RNA的片段称调节基因。基因组(genome)是某生物体所含的全部基因,即全部DNA或完整的单套遗传物质(配子中的整套基因)。
2.细菌、噬菌体、大多数动植物病毒的基因组即指单个DNA分子。最小病毒如SV40的基因组仅有5226b,含5个基因。大肠杆菌含4.6×106 bp,有3000~4000个基因,DNA完全伸展总长约1.3mm。
导入:核酸是生物遗传的物质基础。它的发现和研究进展如何?
1868年瑞士青年医生Miescher从脓细胞核中分离出一种含磷量很高的酸性化合物,称为核素。其继任者Altman发展了从酵母和动物组织中制备不含蛋白质的核酸的方法,于1889年提出核酸(nucleic acid)这一名称。早期核酸研究因“四核苷酸假说”的错误进展缓慢。
原核生物基因组的特点是:结构简炼,绝大部分为蛋白质编码(结构基因);有转录单元,即功能相关的基因常串联一起,并转录在同一mRNA(多顺反子mRNA)中;有基因重叠现象,即同一段DNA携带两种不同蛋白质的信息。
3.真核生物基因一般分布在若干条染色体上,其特点是:有重复序列(按重复次数分单拷贝序、中度重复序和高度重复序);有断裂基因(由不编码的内含子和编码的外显子组成)。酵母基因组有1.35×107bp,含6374个基因。人类基因组有3×109 bp,含4万个基因。
一、核酸的基本组成单位
核酸是一种多聚核苷酸,用不同的降解法得到其组成单位——核苷酸。而核苷酸又由碱基、戊糖和磷酸组成。
(一) 戊糖
DNA含β-D-2-脱氧核糖,RNA含β-D-核糖。这是核酸分类的依据。核糖中的C记为1'……5'。
(二)碱基(base)
核酸中的碱基有两类:嘌呤碱和嘧啶碱。有5种基本的碱基外,还有一些含量甚少的稀有碱基。DNA和RNA中常见的两种嘌呤碱是腺嘌呤(adenine,A)、鸟嘌呤(guanine,G)。而嘧啶碱有所不同:RNA主要含胞嘧啶(cytosine,C)、尿嘧啶(uracil,U),DNA主要含胞嘧啶、胸腺嘧啶(thymine,T)。