海马结构、扣带回
在边缘系统中与记忆有关的是

在边缘系统中与记忆有关的是
边缘系统中关于记忆功能重要的部分是海马。
海马结构是边缘系统的中心结构,其位于颞叶的内侧区,左右各一,呈扇形,主要负责学习和记忆;日常生活中的短期记忆都储存在海马体中,如果一个记忆片段,比如一个电话号码或者一个人在短时间内被重复提及的话海马就会将其转存入大脑皮层,成为永久记忆。
所以海马比较发达的人,记忆力相对会比较强一些。
下丘脑,上后脑核,扣带回,海马与杏仁核一起被合称为边缘系统。
克拉弗和布西曾在实验中验证了边缘系统在情绪中的作用。
他们通过外科手术切除了猴子的颞叶。
颞叶的双侧破坏导致了情绪改变在内的一系统症状,先前野性的,具有攻击性的猴子在手术后变得非常温顺,表现很少的情绪性。
在某种情况下愤怒和恐惧永久性地消失,但在其他情况下,它们在手术后恢复,但却有现得很不兴奋——根据这一实验,曾在西文社会出现了利用手术方法切除某些罪犯或者是精神分裂症的颞叶以达到“温顺”的目的。
而下捕丘内有二个区域对饥饿,渴和性行为的调节非常重要。
这二个区域,外侧下丘脑和腹内侧下丘脑,看起来在攻击性行为中也有参与。
但是,至到现今,关于边缘系统的更加具体的功能并没有更全面或者说是有说服力的证据与研究支持。
因此,平时只是认为,这一系统不仅参与人的基本的情绪加工,同时,还对于高级情绪加工,认知,攻击,防御等方面相关联,同时,也会涉及到更多的方面。
同时,因为海马本身与记忆是有着联系的,因此,当海马
被划分入边缘系统的时候,就意味着,它不再是一个单纯的系统,而是一个复杂的功能综合模块。
或者是通道,或者是一个小中心。
海马解剖结构
AP室床通路 6齿状回分子层
PP穿通路
• 经颞叶中部做大脑半球的冠状切面,海马呈双重C环抱的外形,大C代表海马, 开口向腹内侧,小C代表齿状回,位于海马沟的背内侧,开口朝背侧
• 依据细胞形态及皮质发育的差源自,海马被分为CA1、CA2、CA3、CA4四个扇 形区
细胞类型 资料仅供参考,不当之处,请联系改正。
• 接受 – 内嗅皮质:谷氨酸、ENK – 隔区:Ach、GABA、 – 蓝斑:NA – 中缝核:5-HT – 腹侧被盖、黑质:DA
• 纤维分布 – Ach:遍布海马各区 – NA:门区、腔隙分子层 – ENK:齿状回
纤维联系资料仅供参考,不当之处,请联系改正。
• 内部神经环路
– 三突触回路:
1. 嗅区II及III层锥体细胞轴突 (形成穿通径路PP)-齿 状回分子层外2/3-颗粒细 胞树突树突棘;
资料仅供参考,不当之处,请联系改正。
位
置
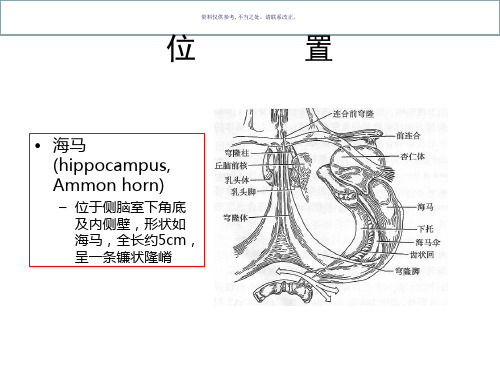
• 海马 (hippocampus, Ammon horn)
– 位于侧脑室下角底 及内侧壁,形状如 海马,全长约5cm, 呈一条镰状隆嵴
资料仅供参考,不当之处,请联系改正。
外形
• 海马前端膨大称海马足 , 被2-3个浅沟分开,沟 间隆起称海马趾
• 海马表面被室管膜上皮 覆盖,下方有一层有髓 纤维称为海马槽(室 床),室床纤维沿海马 背内侧缘集中,形成纵 行白色扁带称海马伞
– 外侧份II层-齿状回分子 层2/3和CA1、CA3腔 隙分子层
– III层穿通纤维-CA1腔隙 分子层
• 对侧海马结构:CA3锥体细 胞-对侧海马CA1、CA3
• 乳头体:乳头体-齿状回通 路(抑制性传入)
• 脑干:蓝斑核、中锋核、腹 侧被盖-齿状回(多形细胞 层)
海马结构

纤维联系
• 内部神经环路 – 三突触回路:
1. 嗅区II及III层锥体细胞轴 突(形成穿通径路PP)-齿 状回分子层外2/3-颗粒细 胞树突树突棘; 2. 颗粒细胞轴突苔藓纤维CA3透明层-锥体细胞顶树 突基部; 3. CA3锥体细胞轴突的 Schaffer侧枝-同侧CA1腔 隙层,终止于锥体细胞顶 端树突干。
传入纤维
• 隔区:隔-海马径路 • 内嗅区: – 内侧份II层-海马槽CA1腔隙分子层(室床 径路) – 外侧份II层-齿状回分 子层2/3和CA1、CA3腔 隙分子层 – III层穿通纤维-CA1腔 隙分子层 • 对侧海马结构:CA3锥体细 胞-对侧海马CA1、CA3 • 乳头体:乳头体-齿状回通 路(抑制性传入) • 脑干:蓝斑核、中锋核、腹 侧被盖-齿状回(多形细胞 层)
传出纤维
穹隆是海马传出的主要径路 • CA1区-伏核、尾壳核 • CA3区-双侧外侧隔核、同侧 CA1区、对侧CA1、CA3及齿 状回、外侧视前区、下丘脑 前份、中脑中央灰质吻部 • 部分下托-形成联合后穹隆乳头体 • 内嗅区皮质-旁嗅回、海马 旁回、扣带回、额叶、眶额 皮质、伏隔核、尾壳核
海马解剖结构
神经生物学系 施 静 2009.11.
位
• 海马(hippocampus, Ammon horn)
– 位于侧脑室下角底 及内侧壁,形状如 海马,全长约5cm, 呈一条镰状隆嵴
置
外
• 海马前端膨大称海马足 , 被2-3个浅沟分开,沟间 隆起称海马趾 • 海马表面被室管膜上皮 覆盖,下方有一层有髓 纤维称为海马槽(室 床),室床纤维沿海马 背内侧缘集中,形成纵 行白色扁带称海马伞 • 齿状回是一条窄的皮质, 由于许多血管进入而被 压成许多横沟成齿状
海马结构

海马结构MARK(1)海马结构(hippocampal formation)包括海马(又称安蒙角cornu AmmonisCA)、下托、齿状回和围绕胼胝体形成一圈的海马残件。
齿状回至胼胝体压部,消失齿状外形,改称束状回,束状回向前上与覆盖胼胝体上面的深层灰质称灰被(又称胼胝体上回)相连续。
灰被中埋有一对纵纹,分别为内侧纵纹与外侧纵纹。
灰被与纵纹就是海马及其白质的残件。
它们向前经胼胝体膝与终板旁回连续。
海马(hippocampus)形如中药海马故名。
位于侧脑室下角底兼内侧壁,全长5 cm。
海马前端较膨大称海马足,它被2-3个浅沟分开,沟间隆起称海马趾。
海马是一条镰状隆嵴,自胼胝体压部向前到侧脑室的颞端。
海马至胼胝体压部时,从齿状回和海马旁回间翻出称Retzius回。
海马结构的位置海马表面被室管膜上皮覆盖。
室管膜上皮下面有一层有髓纤维称为海马槽(又称室床alveus)。
室床纤维沿海马背内侧缘集中,形成白色扁带称海马伞(fimbria of hippocampus),它自海马趾伸向压部,续于穹隆脚(crus of fomix)。
海马伞的游离缘直接延续于其上方的脉络丛,两者间隔以脉络裂。
海马结在下角的发育齿状回(dentate gyms)是一狭条皮质;由于血管进入被压成许多横沟呈齿状,故名。
它位于海马的内侧,介于海马沟与海马伞之间。
齿状回向前伸展至钩的切迹,在此急转弯,成光滑小束横过钩的下面,这横行段称齿状回尾。
齿状回尾将钩分成前部的前钩回,后部的边叶内回。
齿状回向后与束状回(fasciolar gyrus)相连。
在海马结构发育较好的颞中平面,作一个大脑半球的冠状切面,海马结构呈双重“C”形环抱的外形,大C锁住小C。
大C代表海马,它开口向腹内侧。
小C代表齿状回,位于海马沟的背内侧,开口朝向背侧。
海马沟的腹侧为下托(subiculum)。
海马结构的位置与安排,从发育过程来理解比较清楚。
在胚胎3个月,两个半球内侧壁上各显出一条纵行加厚部分称海马嵴(hippocampal ridge),这是海马结构的原基。
大脑结构名词中英文对照

大脑结构名词中英文对照汉英版:安蒙氏角Ammom's horn白质white matter背内侧丘脑dorsalmedial thalamus背外侧通路dorsallateral pathway背外侧膝状核dorsal lateral geniculate nucleus被盖tegmentum被盖背束dorsal tegmental bundle被盖腹区ventral tegmental area边缘皮质limbic cortex边缘系统limbic system布洛卡区Broca's area苍白球globus pallidus侧脑室lateral ventricle齿状回dentate gyrus穿质通路perforant path传出纤维efferent fibers传入纤维afferent fibers大脑半球cerebral hemisphere大脑导水管cerebral aqueduct导水管周围灰质periaqueductal grey matter第三脑室third ventricle第四脑室fourth ventricles顶盖tectum顶盖脊髓束tectospinal tract顶盖枕核系统tectopulvinar system顶叶parietal lobes豆状核lentiform nucleus端脑telecephalon额眶皮质orbital frontal cortex额叶frontal lobe额叶岛盖frontal operculum耳蜗核cochlear nucleus二级投射区sencondary projection area非特异性投射系统nonspecific projecting system缝际核raphe nucleus伏核nucleus accumbens腹内侧通路ventraomedial pathway腹内侧下丘脑ventromedial hypothalamus隔区septal area弓状核arcuate nucleus弓状束arcuate fasciculus沟sulcus钩束uncinate faciculus孤束核solitary nucleus海马hippocampus海马结构hippocampus formation海马伞fimbria黑质substantia nigra黑质纹状体束nigrotriatal bundles红核red nucleus红核束rubrospinal tract后脑metencephalon灰质gray substance基底神经节basal ganglia基底外侧核群basolateral nuclear group间脑diencephalon交叉decussation角回angular gyrus旧纹状体(苍白球)paleostriatum距状裂calcarine fissure壳核putamen扣带cingulum扣带回cingulate gyrus扣带回皮质cingulate cortex蓝斑locus coeruleus连合commissure联络皮质associative cortex裂fissure菱脑rhombencephalon漏斗infundibulum颅腔endocast脉络丛choroid plexus梅纳特基底核(无名核)basal nucleus of Meynert 末脑myelencephalon内侧前脑束medial forebrain bundle内侧膝状体medial geniculate nucleus内囊internal capsule内嗅皮质entorhinal cortex脑干brain stem脑干网状结构brainstem reticular formation脑化encephalization脑化指数encephalization index脑回gyrus脑脊膜meninges脑脊液cerebrospinal fluid脑桥pons脑室ventricles颞平台planum temporale颞叶temporal lobe旁臂核parabranchial nucleus旁室核paraventricular nucleus皮质脊髓侧束lateral corticospinal tract皮质脊髓前束ventral corticospinal tract皮质脊髓束corticospinal system皮质脊髓通路cortcospinal pathway皮质内侧核群corticomedial nuclear group皮质下subcortex皮质延髓束corticobulbar tract胼胝体corpus callosum前额皮质prefrontal cortex前脑proseucephalon前脑基底大细胞核magnocellular nucleus of the basal forebrain 前下托presubiculum穹窿fornix穹窿下器官subfornical organ丘colliculus丘脑thalamus丘脑腹后核ventral posterior nucleus of thalamus丘脑腹外侧核ventrolateral nucleus of thalamus丘脑前核anterior nucleus thalamus丘系系统lemniscal system躯体感觉皮质somatosensory cortex躯体感觉区somatosensory area乳头丘脑束mammillothalamic tract乳头体mammillary body软脑膜pia matter塞尔维氏裂Sylvian fissure三级投射区tertiary projection area上橄榄核群superior olivary complex上丘superior colliculus上丘脑epithalamus神经核nucleus视顶盖optic tectum视放核optical radiation视交叉上核suprachiasmatic nucleus视觉背侧系统dorsal system of visual function视觉腹侧系统ventral system of visual function视觉皮质(枕极)visual cortex视前内侧区medial preoptic area视前区preoptic area视上核supraoptic nucleus室周器官circumventricular organ松果体corpus pineale髓板medullary lamina髓质medullary substances or medulla特异性投射系统specific projecting system听觉皮质auditory cortex听觉皮质auditory cortex外侧丘系lateral lemniscus外侧膝状体lateral geniculate nucleus外侧膝状体纹皮质系统geniculostriate system外侧下丘脑lateral hypothalamus area外囊external capsuale网状脊髓束reticulospinal tract网状结构reticular formation维尔尼克区Wernicke area尾状核caudate nucleus纹前皮质prestriate cortex纹状皮质striate cortex纹状区striate area下颞皮质inferior temporal cortex下丘inferior colliculus下丘脑hypothalamus下托subiculum小脑cerebellum小脑上脚superior cerebella peduncle小脑小叶cerebellar folia小细胞神经分泌系统parvocellular neurosecretory system 新纹状体neostriatum杏仁核amygdala杏仁核腹侧传出通路ventral amygdalofugal pathway性二型核sexually dimorphic nucleus嗅脑rhinencephalon嗅球olfactory bulb延脑medulla oblongata眼优势柱ocular dominance columns一级感觉皮质=第一感觉区primary sensory cortex一级视觉皮质primary visual cortex一级投射皮质primary projection area一级运动皮质primary motor cortex硬脑膜dura matter运动控制系统motor control system运动皮质motor cortex枕骨大孔occipital foramen枕核pulvinar枕极occipital pole枕角occipital horns枕叶occipital lobe中缝大核nucleus raphe magnus中间块massa intermedia中间皮质mesocortex中脑mesencephalon中央被盖束central tegmental tract中央沟central sulcus中央管central canal中央后回postcentral cortex中央前回precentral gyrus终纹stria terminalis蛛网膜arachnoid蛛网膜下腔subarachnoid锥体区pyramidalis area锥体外运动系统extrapyramidal motor system锥体运动系统pyramidal motor system英汉版:afferent fibers 传入纤维Ammom's horn 安蒙氏角amygdala 杏仁核angular gyrus 角回anterior nucleus thalamus 丘脑前核arachnoid 蛛网膜arcuate fasciculus 弓状束arcuate nucleus 弓状核associative cortex 联络皮质auditory cortex 听觉皮质auditory cortex 听觉皮质basal ganglia 基底神经节basal nucleus of Meynert 梅纳特基底核(无名核)basolateral nuclear group 基底外侧核群brain stem 脑干brainstem reticular formation 脑干网状结构Broca's area 布洛卡区calcarine fissure 距状裂caudate nucleus 尾状核central canal 中央管central sulcus 中央沟central tegmental tract 中央被盖束cerebellar folia 小脑小叶cerebellum 小脑cerebral aqueduct 大脑导水管cerebral hemisphere 大脑半球cerebrospinal fluid 脑脊液choroid plexus 脉络丛cingulate cortex 扣带回皮质cingulate gyrus 扣带回cingulum 扣带circumventricular organ 室周器官cochlear nucleus 耳蜗核colliculus 丘commissure 连合corpus callosum 胼胝体corpus pineale 松果体cortcospinal pathway 皮质脊髓通路corticobulbar tract 皮质延髓束corticomedial nuclear group 皮质内侧核群corticospinal system 皮质脊髓束decussation 交叉dentate gyrus 齿状回diencephalon 间脑dorsal lateral geniculate nucleus 背外侧膝状核dorsal system of visual function 视觉背侧系统dorsal tegmental bundle 被盖背束dorsallateral pathway 背外侧通路dorsalmedial thalamus 背内侧丘脑dura matter 硬脑膜efferent fibers 传出纤维encephalization 脑化encephalization index 脑化指数endocast 颅腔entorhinal cortex 内嗅皮质epithalamus 上丘脑external capsuale 外囊extrapyramidal motor system 锥体外运动系统fimbria 海马伞fissure 裂fornix 穹窿fourth ventricles 第四脑室frontal lobe 额叶frontal operculum 额叶岛盖geniculostriate system 外侧膝状体纹皮质系统globus pallidus 苍白球gray substance 灰质gyrus 脑回hippocampus 海马hippocampus formation 海马结构hypothalamus 下丘脑inferior colliculus 下丘inferior temporal cortex 下颞皮质infundibulum 漏斗internal capsule 内囊lateral corticospinal tract 皮质脊髓侧束lateral geniculate nucleus 外侧膝状体lateral hypothalamus area 外侧下丘脑lateral lemniscus 外侧丘系lateral ventricle 侧脑室lemniscal system 丘系系统lentiform nucleus 豆状核limbic cortex 边缘皮质limbic system 边缘系统locus coeruleus 蓝斑magnocellular nucleus of the basal forebrain 前脑基底大细胞核mammillary body 乳头体mammillothalamic tract 乳头丘脑束massa intermedia 中间块medial forebrain bundle 内侧前脑束medial geniculate nucleus 内侧膝状体medial preoptic area 视前内侧区medulla oblongata 延脑medullary lamina 髓板medullary substances or medulla 髓质meninges 脑脊膜mesencephalon 中脑mesocortex 中间皮质metencephalon 后脑motor control system 运动控制系统motor cortex 运动皮质myelencephalon 末脑neostriatum 新纹状体nigrotriatal bundles 黑质纹状体束nonspecific projecting system 非特异性投射系统nucleus 神经核nucleus accumbens 伏核nucleus raphe magnus 中缝大核occipital foramen 枕骨大孔occipital horns 枕角occipital lobe 枕叶occipital pole 枕极ocular dominance columns 眼优势柱olfactory bulb 嗅球optic tectum 视顶盖optical radiation 视放核orbital frontal cortex 额眶皮质paleostriatum 旧纹状体(苍白球)parabranchial nucleus 旁臂核paraventricular nucleus 旁室核parietal lobes 顶叶parvocellular neurosecretory system 小细胞神经分泌系统perforant path 穿质通路periaqueductal grey matter 导水管周围灰质pia matter 软脑膜planum temporale 颞平台pons 脑桥postcentral cortex 中央后回precentral gyrus 中央前回prefrontal cortex 前额皮质preoptic area 视前区prestriate cortex 纹前皮质presubiculum 前下托primary motor cortex 一级运动皮质primary projection area 一级投射皮质primary sensory cortex 一级感觉皮质=第一感觉区primary visual cortex 一级视觉皮质proseucephalon 前脑pulvinar 枕核putamen 壳核pyramidal motor system 锥体运动系统pyramidalis area 锥体区raphe nucleus 缝际核red nucleus 红核reticular formation 网状结构reticulospinal tract 网状脊髓束rhinencephalon 嗅脑rhombencephalon 菱脑rubrospinal tract 红核束sencondary projection area 二级投射区septal area 隔区sexually dimorphic nucleus 性二型核solitary nucleus 孤束核somatosensory area 躯体感觉区somatosensory cortex 躯体感觉皮质specific projecting system 特异性投射系统stria terminalis 终纹striate area 纹状区striate cortex 纹状皮质subarachnoid 蛛网膜下腔subcortex 皮质下subfornical organ 穹窿下器官subiculum 下托substantia nigra 黑质sulcus 沟superior cerebella peduncle 小脑上脚superior colliculus 上丘superior olivary complex 上橄榄核群suprachiasmatic nucleus 视交叉上核supraoptic nucleus 视上核Sylvian fissure 塞尔维氏裂tectopulvinar system 顶盖枕核系统tectospinal tract 顶盖脊髓束tectum 顶盖tegmentum 被盖telecephalon 端脑temporal lobe 颞叶tertiary projection area 三级投射区thalamus 丘脑third ventricle 第三脑室uncinate faciculus 钩束ventral amygdalofugal pathway 杏仁核腹侧传出通路ventral corticospinal tract 皮质脊髓前束ventral posterior nucleus of thalamus 丘脑腹后核ventral system of visual function 视觉腹侧系统ventral tegmental area 被盖腹区ventraomedial pathway 腹内侧通路ventricles 脑室ventrolateral nucleus of thalamus 丘脑腹外侧核ventromedial hypothalamus 腹内侧下丘脑visual cortex 视觉皮质(枕极)Wernicke area 维尔尼克区white matter 白质。
海马结构

传入纤维
隔区:隔-海马径路 内嗅区: – 内侧份II层-海马槽CA1腔隙分子层(室床 径路) – 外侧份II层-齿状回分 子层2/3和CA1、CA3腔 隙分子层 – III层穿通纤维-CA1腔 隙分子层 对侧海马结构:CA3锥体细 胞-对侧海马CA1、CA3 乳头体:乳头体-齿状回通 路(抑制性传入) 脑干:蓝斑核、中锋核、腹 侧被盖-齿状回(多形细胞 层)
– –
(三种突触之间都是兴奋 性氨基酸-形成兴奋性前馈 通路) CA3反向投射到齿状回分子 层-解释海马腹侧惊厥易感 性问题
海马结构的外部联系:
– 皮层:海马旁回、颞上回、旁嗅回、岛叶、扣 带回、眶额皮质 – 皮层下:杏仁复合体、屏状核、内侧隔核、下 丘脑后部的乳头体上区、前丘脑、丘脑中线核 群、腹侧被盖、蓝斑
形
Hale Waihona Puke 海马分区:3锥体层 4放射层 5腔隙分子层
海马与齿状回均属于古皮质3层结构:分子层、锥体细胞层(海 马)、颗粒细胞层(齿状回)
2始层 1室床
Sch Schaffer侧枝
Mf苔藓纤维 AP室床通路 8颗粒层 7齿状回多形层 6齿状回分子层 PP穿通路
经颞叶中部做大脑半球的冠状切面,海马呈双重C环抱的外形,大C代表海马, 开口向腹内侧,小C代表齿状回,位于海马沟的背内侧,开口朝背侧 依据细胞形态及皮质发育的差异,海马被分为CA1、CA2、CA3、CA4四个扇形区
传出纤维
穹隆是海马传出的主要径路 CA1区-伏核、尾壳核 CA3区-双侧外侧隔核、同侧 CA1区、对侧CA1、CA3及齿 状回、外侧视前区、下丘脑 前份、中脑中央灰质吻部 部分下托-形成联合后穹隆乳头体 内嗅区皮质-旁嗅回、海马 旁回、扣带回、额叶、眶额 皮质、伏隔核、尾壳核
抑郁症会导致脑部结构变化吗?

抑郁症会导致脑部结构变化吗?
抑郁症有时候被看作心理问题,有时候又被看作神经问题。
二者之间有个很重要的区分,即抑郁症是否已经存在脑神经化学层面甚至是脑部结构层面的改变;如果有,那就更倾向于神经问题。
脑神经化学层面是指大脑神经递质和神经递质浓度是否超出正常的范围。
抑郁症与血清素水平偏低有关几乎是共识性的理论,这就是说,严重的抑郁症患者的脑部很可能出现神经递质浓度的异常。
有研究表明,抑郁症患者的脑部结构会发生一些变化,这主要集中在边缘系统,包括杏仁核、海马体、前扣带回和左侧前额叶皮质。
具体来说:抑郁症患者的杏仁核体积增大,而且呈正相关;重度抑郁患者,尤其是病情反复的患者,海马体的体积会变小,因此记忆力会下降;前扣带回也会发生变化,这个部分的变化导致患者的情绪反应更多是负向消极的;左侧前额叶皮质发生变化,影响患者的计划决策、神经活动调控及理性思维功能。
通常我们把左侧前额叶皮质称为“理性脑”,把杏仁核称为“情绪脑”。
有研究表明,左侧前额叶皮层与抑郁的想法和行为有关。
尽管尚未完全了解左侧前额叶皮质的病理生
理学,但专家已经发现大脑前额叶区域的结构异常,特别是内侧和左侧前额叶皮质异常会导致抑郁症。
这一研究带来的影响就是经颅磁刺激这样的新技术被用于治疗抑郁症,其工作原理是刺激大脑的目标区域,帮助该区域形成更多具有更好传导性的连接,这一过程被称为“神经可塑性”,是较为前沿的治疗方法。
除此之外,通过训练可以一定程度上缓解杏仁核带来的不良情绪反应。
例如,已经有研究指出,杏仁核与纹状体影响情绪,而睡眠不好,就会对二者产生负面影响。
由此可见,睡眠不好会影响人的生活和生命质量,对抑郁症患者而言,更应该高度关注睡眠问题。
海马的两个记忆回路

海马的两个记忆回路海马→穹窿→乳头体→乳头丘脑束→丘脑前核→扣带回→海马,这条环路是30年代就认识到的边缘系统的主要回路,称为帕帕兹环。
在这条环路中,海马结构是中心环节。
所以,在40-50年代曾认为海马结构与情绪体验有关。
近些年发现,内侧嗅回与海马结构之间存在着三突触回路,它与记忆功能有关。
三突触回路始于内嗅区皮层,这里神经元轴突形成穿通回路,止于齿状回颗粒细胞树突,形成第一个突触联系。
齿状回颗粒细胞的轴突形成苔状纤维(Mossy fibers)与海马CA3区和锥体细胞的树突形成第二个突触联系。
CA3区锥体细胞轴突发出侧支与CA,区的锥体细胞发生第3个突触联系,再由CAi锥体细胞发出向内侧嗅区的联系。
这种3突触回路是海马齿状回内嗅区与海马之间的联系,具有特殊的机能特性,成为支持长时记忆机制的证据。
1966年,罗莫(T. Lomo)首先报道了他称之谓长时程增强(Long-term potentiation,LTP)的现象,即电刺内嗅区皮层向海马结构发出的穿通回路时,在海马齿状回可记录出细胞外的诱发反应。
如果电刺激由约100个电脉冲组成,在1-10秒内给出,则齿状回诱发性细胞外电活动在5-25分钟之后增强了2.5倍,说明电刺激穿通回路引起齿状回神经元突触后兴奋电位的LTP,因而这些神经元单位发放的频率增加。
后来他们又报道,海马齿状回神经元突触电活动的LTP现象可持续散月的时间。
他们认为,由短暂电刺激穿通回路所引起的三突触神经回路持续性变化,可能是记忆的重要基础。
每侧的海马齿状回都接受两侧内侧嗅区来的穿通纤维,但以同侧性联系为主,对侧性联系较少。
如果在单侧刺激内嗅区,则发现在同侧海马齿状回内很容易引起LTP现象,而在对侧海马齿状回内,则很难引起这种现象。
如果用建立经典条件反射的程序对两侧内嗅区施以刺激时,就会发现LTP效应的呈现也符合经典条件反射建立的基本规律,从而证明LTP 现象可能是一种学习的脑机制。
海马解剖及MR诊断

2020/4/4
48
仁 爱 |精 诚 |融 汇 |卓 越
病理及病理生理特点
• 主要病理特征为海马结构内神经元数目减少,神经元树突棘丧失及星形胶质细 胞的反应性增生,其中以CA1区神经元细胞减少最为显著,CA3、CA4区呈中 等程度减少,CA2区相对减少(CA2区对神经元凋亡有抵抗作用)随着病情发 展,病变区海马组织出现萎缩,可同时伴有颞叶萎缩
2020/4/4
16
•内侧颞叶实际 为一组相互联 系的结构 •包括海马和海 马附近的3个皮 层区:内嗅皮 层、嗅周皮层 (二者合称嗅 皮层)和旁海 马皮层
海马足 下托 内嗅区
2020/4/4
仁 爱 |精 诚 |融 汇 |卓 越
17
仁 爱 |精 诚 |融 汇 |卓 越
边缘叶(Limbic lobe)
海马硬化的病理改变及影像表现
2020/4/4
46
仁 爱 |精 诚 |融 汇 |卓 越
海马内部分区cornu Ammonis
• 分CA1, CA2, CA3, CA4区, 含有椎体细 胞。
• CA1细胞最丰富,是下托的直接延续 • CA2 位于comu Ammonis弯向齿状回
前部分头侧。 • CA3 区是comu Ammonis的过渡部分 • CA4 区由齿状回包绕。 • 海马槽 (a)是白质纤维,将海马与颞角分
• 海马硬化与癫痫之间可能互为因果关系 ✓ 海马硬化神经细胞丢失可刺激剩余神经元代偿生长和神经突触异常重组,引起
局部神经纤维异常放电,诱发癫痫 ✓ 癫痫异常放电反复刺激会引起并加剧局部脑组织神经元细胞的代谢改变,进而
导致细胞膜通透性增加,细胞水肿、萎缩、细胞外间隙增宽等病理改变。
2020/4/4
海马的结构及功能

•
海马结构的构筑: 2. 海马结构的构筑 : 海马和齿状回均 属古皮质archipallium, archipallium,都是由三层细胞 属古皮质archipallium,都是由三层细胞 组成: 分子层、 锥体细胞层( 海马) 组成 : 分子层 、 锥体细胞层 ( 海马 ) 或颗 粒细胞层(齿状回)和多形层。 粒细胞层(齿状回)和多形层。
•
齿状回dentate gyrus:是一狭条皮质, 齿状回dentate gyrus:是一狭条皮质,由于 血管进入被压成许多横沟呈齿状, 血管进入被压成许多横沟呈齿状, • 故名。它位于海马的内侧。 故名。它位于海马的内侧。 • 在海马结构较好发育的颞中平面, 在海马结构较好发育的颞中平面,作一 个大脑半球的冠状切面,海马结构呈双重C 个大脑半球的冠状切面,海马结构呈双重C环抱 的外形, 锁住小c 代表海马, 的外形,大C锁住小c。大C代表海马,它开口向 腹内侧。 代表齿状回,位于海马沟的背内侧, 腹内侧。小c代表齿状回,位于海马沟的背内侧, 开口朝向背侧。海马沟的腹侧为下托subculum subculum。 开口朝向背侧。海马沟的腹侧为下托subculum。
• 依据细胞形态,不同皮质区发育的差异以 依据细胞形态, 及 各 种 纤 维 通 路 的 不 同 , Lorente do No{1934 把海马又分成CA 1934} CA1 CA2 CA3 No{1934}把海马又分成CA1、CA2、CA3、 CA4 CA4 CA4 四 个 扇 形 区 。 CA4 位 于 齿 状 回 门 ,CA3 区内有最大的锥体细胞,CA ,CA3 内 ,CA3 区内有最大的锥体细胞 ,CA3 区由 大锥体细胞组成, CA1 大锥体细胞组成, CA1区是邻近下托的部 由小锥体细胞组成,CA 区是移行区, ,CA2 分,由小锥体细胞组成,CA2区是移行区, 由大和小锥体细胞组成。 由大和小锥体细胞组成。
海马的结构及功能

• ( 4 ) 杏 仁 皮 质 投 射 amygdalo-cortical projection:投射至眶额回、颞上、中、下回。
整理版ppt
9
• 4.功能
• 电刺激或切除杏仁体,可产生行为、
整理版ppt
18
• (三)海马结构
•
海 马 结 构 hippocampal formation
包括海马(安蒙氏角)、下托、齿状回等。
•
1.外形与位置
•
海马hippocampus :形如中药海
马故名。位于侧脑室下角底兼内侧壁。
全长5cm 。
整理版ppt
19
整理版ppt
20
• 齿状回dentate gyrus:是一狭条皮质,由于 血管进入被压成许多横沟呈齿状,
整理版ppt
14
• 2.传出纤维
• 〈1〉内侧隔核发纤维经穹窿返回海马。
• 〈2〉自隔核发纤维经内侧前脑束分布 至下丘脑外侧区,有些纤维向尾侧伸展终 于中脑网状结构。
• 〈3〉自隔核发纤维至丘脑前腹核与 背内侧核,又经髓纹至缰核。
整理版ppt
15
• 3.功能
• (1)情绪反应:对隔区的剌激,有时 引起病人高兴和愉缺的反应,有的病人咯 咯发笑,言语增多,显得比平时更为随和。 损毁隔区的动物,立即出现怒反应增加 和 感 情 超 常 的 “ " 隔 综 合 征 ” septal syndrome。
扣带回发纤维经扣带束直接终止于海马和或经内嗅区中继后发纤维至海马和下托从隔核发纤维胆碱能纤维经穹窿海马伞终止于海马和齿状回一侧海马发纤维经同侧海马伞穹窿脚通过海马连合至对侧穹窿脚与海马伞终止于对侧海马和齿状回
海马结构

–
–
(三种突触之间都是兴奋 性氨基酸-形成兴奋性前馈 通路) CA3反向投射到齿状回分子 层-解释海马腹侧惊厥易感 性问题
• 海马结构的外部联系:
– 皮层:海马旁回、颞上回、旁嗅回、岛叶、扣 带回、眶额皮质 – 皮层下:杏仁复合体、屏状核、内侧隔核、下 丘脑后部的乳头体上区、前丘脑、丘脑中线核 群、腹侧被盖、蓝斑
海马的化解剖
• 固有 – 锥体细胞:谷氨酸、Ach – 颗粒细胞:谷氨酸、强啡肽 – 中间神经元:GABA、CCK、nNOS、VIP、SS • 接受 – 内嗅皮质:谷氨酸、ENK – 隔区:Ach、GABA、 – 蓝斑:NA – 中缝核:5-HT – 腹侧被盖、黑质:DA • 纤维分布 – Ach:遍布海马各区 – NA:门区、腔隙分子层 – ENK:齿状回
形
海马分区:
3锥体层 4放射层 5腔隙分子层
海马与齿状回均属于古皮质3层结构:分子层、锥体细胞层(海 马)、颗粒细胞层(齿状回)
2始层 1室床
Sch Schaffer侧枝
Mf苔藓纤维 AP室床通路 PP穿通路
8颗粒层
7齿状回多形层 6齿状回分子层
• 经颞叶中部做大脑半球的冠状切面,海马呈双重C环抱的外形,大C代表海马, 开口向腹内侧,小C代表齿状回,位于海马沟的背内侧,开口朝背侧 • 依据细胞形态及皮质发育的差异,海马被分为CA1、CA2、CA3、CA4四个扇形区
细胞类型
• 锥体细胞或颗 粒细胞 • 中间神经元 (5%-8%):篮 细胞、腔隙分 子层中间神经 元、吊灯样细 胞 • 门区细胞:苔 藓细胞
锥体细胞示意图
• 胞体长径20-30uM。短径1020uM。CA1区排列2-3层,CA3区 的细胞数是CA1区的1.5-2.0倍, 排列疏松,最多可达10层。 • 树突与苔藓纤维形成突触,构 成透明层 • CA2区细胞排列最紧密,但不与 苔藓纤维形成突触,只接受下 丘脑乳头上区的传入纤维 • 轴突由海马伞经穹窿进入隔区, 并经穹窿连合投射到对侧CA1、 CA3区
边缘系统病变

短暂性全面遗忘症是指突发的顺行性和逆行性遗忘,持续 24 小时左 右。病因不明,通常累及 50-70 岁的中老年人,可由精神紧张诱发。 发病时,由于病人出现短暂性获得新信息的功能障碍,此时病人会出 现重复提问等,但意识和远期记忆存在,无神经功能缺损症状。该病 可自行缓解,发作过后病人完全恢复记忆能力,唯独发作期的经历完
IAE 是流感病毒感染的罕见并发症,主要累及 5 岁以下儿童,其变异
型 ANE 定义为脑部对称性出血病灶,最为常见的临床特征是全面癫 痫发作和精神状态改变,包括意识水平、言语异常和谵妄。影像学可 发现对称性分布的脑部病灶,累及大脑白质、丘脑和脑干深部结构, 特征为双侧坏死或出血性丘脑病灶。
神经梅毒
三期梅毒是由闭塞的小动脉内膜炎引起的,通常累及中枢神经系统的滋养血管。 三期梅毒有以下三种类型:
神经梅毒
心血管梅毒
晚发性良性(如树胶肿的)梅毒
神经梅毒可分为两大类:
(1)早期中枢神经系统的损害仅限于脑膜, (2)随后有脑实质的侵犯。
脑膜血管梅毒的病理改变是伴有血管周围炎症的动脉内膜炎。其主要表现为:动
惠普尔病是一种少见的系统性疾病。男女发病之比为(4~8):1, 以30~50岁发病者为多,平均发病年龄为39岁。其临床特征为小肠 吸收不良、发热、皮肤色素沉着、贫血、淋巴结肿大、关节炎、关节 痛、胸膜炎、瓣膜性心内膜炎和中枢神经系统症状。 1907 年惠普尔 (Whipple)首次报道本病,并描述了其临床特征及小肠、淋巴结的 病理改变。
蛋白含量增高,淋巴细胞计数增多,葡萄糖在正常范围内。
边缘叶脑炎的患者通常表现为急性或亚急性起病,伴有行为或心境障
碍;短期记忆或其它认知障碍(如失语);局灶性癫痫;炎性脑脊液。
脑区包含的电极

脑区包含的电极导言:脑区是指脑组织中的某一特定区域,而电极是用于记录和刺激神经活动的装置。
本文将介绍几个常见的脑区,并说明与之相关的电极。
一、额叶电极额叶是大脑皮层的一个重要区域,包括额上回、额中回、额下回等结构。
额叶电极是植入在额叶区域的电极,用于记录和刺激额叶神经活动。
额叶电极可用于治疗癫痫、抑郁症等疾病,也可以用于研究认知功能和情绪调节等相关领域。
二、颞叶电极颞叶是大脑皮层的一个重要区域,位于太阳穴附近,包括颞上回、颞中回、颞下回等结构。
颞叶电极是植入在颞叶区域的电极,用于记录和刺激颞叶神经活动。
颞叶电极可用于治疗癫痫、失语症等疾病,也可以用于研究听觉、语言等相关领域。
三、顶叶电极顶叶是大脑皮层的一个重要区域,位于头顶部位,包括顶上回、顶中回、顶下回等结构。
顶叶电极是植入在顶叶区域的电极,用于记录和刺激顶叶神经活动。
顶叶电极可用于治疗癫痫、运动障碍等疾病,也可以用于研究空间认知、运动控制等相关领域。
四、枕叶电极枕叶是大脑皮层的一个重要区域,位于后脑勺部位,包括枕上回、枕中回、枕下回等结构。
枕叶电极是植入在枕叶区域的电极,用于记录和刺激枕叶神经活动。
枕叶电极可用于治疗癫痫、睡眠障碍等疾病,也可以用于研究视觉、记忆等相关领域。
五、扣带回电极扣带回是大脑皮层的一个重要区域,位于大脑的中央部位,包括上扣带回、下扣带回等结构。
扣带回电极是植入在扣带回区域的电极,用于记录和刺激扣带回神经活动。
扣带回电极可用于治疗帕金森病、运动障碍等疾病,也可以用于研究运动控制、意识等相关领域。
六、海马电极海马是大脑皮层的一个重要区域,位于颞叶内侧,是记忆和空间导航的重要中枢。
海马电极是植入在海马区域的电极,用于记录和刺激海马神经活动。
海马电极可用于治疗癫痫、阿尔茨海默病等疾病,也可以用于研究记忆、学习等相关领域。
七、杏仁核电极杏仁核是大脑皮层的一个重要区域,位于颞叶内侧,参与情绪和记忆的调节。
杏仁核电极是植入在杏仁核区域的电极,用于记录和刺激杏仁核神经活动。
大脑边缘系统

大脑边缘系统边缘系统limbic system由与边缘叶有关的皮质及皮质下结构(如杏仁体、下丘脑、上丘脑、背侧丘脑前核和中脑被盖等)组成边缘叶limbic lobe在半球内侧面,位于胼胝体周围和侧脑室下角底壁的一圈弧形结构:隔区(包括胼胝体下区和终板旁回)、扣带回、海马旁回、海马和齿状回等,它们属于原皮质和旧皮质。
边缘系统是指高等脊椎动物中枢神经系统中由古皮层、旧皮层演化成的大脑组织以及和这些组织有密切联系的神经结构和核团的总称。
古皮层和旧皮层是被新皮层分隔开的基础结构。
边缘系统的重要组成包括,海马结构、海马旁回及内嗅区、齿状回、扣带回、乳头体以及杏仁核。
上述结构通过帕帕兹环[Papez 环路]相互联系,并与其他脑结构(新皮层、丘脑、脑干)有广泛联系,所以边缘系统的作用是使中脑、间脑和新皮层结构之间发生信息交换。
通过与下丘脑及植物神经系统的联系,边缘系统参与调解本能和情感行为,其作用是自身生存和物种延续。
此外,海马结构还对学习过程和记忆发挥着突出的作用。
因此如果海马结构或与之功能联系的结构受损,则导致遗忘综合征。
其病变部位不同,产生的记忆障碍形式也不同。
边缘系统所包括的大脑部位相当广泛,如梨状皮层、内嗅区、眶回、扣带回、胼胝体下回、海马回、脑岛、颞极、杏仁核群、隔区、视前区、下丘脑、海马以及乳头体都属于边缘系统。
边缘系统的主要部分环绕大脑两半球内侧形成一个闭合的环,故此得名。
边缘系统内部互相连接与神经系统其他部分也有广泛的联系。
它参与感觉、内脏活动的调节并与情绪、行为、学习和记忆等心理活动密切相关。
边缘系统的功能:调节内脏活动刺激边缘系统的后眶回、扣带回、岛叶、颞极、梨状皮层、旁杏仁皮层、后海马皮层等部位,可以引起人及动物的呼吸、血管以及其他内脏反应。
刺激下丘脑不同部分所引起的内脏反应最为明显,在出现竖毛、瞳孔扩大的同时,血压急剧升高,心率加快以及出现饮水、摄食、排尿、排粪、流涎和呕吐等反应。
刺激眶回皮层,可引起血压下降,心率变慢。
解剖-海马

胼胝体上回 向两侧进入胼胝体沟,于
沟底移行于扣带回;向前绕过胼胝体膝, 移行于两侧的胼胝体下回;向后绕过胼胝 体压部,移行于两侧的束状回。
束状回 fasciolar gyrus 是条灰质带,向
前下外方移行于齿状回和海马。
海马结构的外形
2.齿状回 dentate gyrus 是狭长
海马结构的皮质构造
锥体细胞的树突在各层接受不同的轴突
终末。1)来自对侧海马的连合纤维,终 于底树突;2)来自内嗅区的轴突和返回 侧支,在分子层终于尖树突;3)来自齿 状回的苔状纤维,在辐射层包绕尖树突; 4)篮状细胞的终末形成许多轴-体突触。 传入纤维和锥体细胞具有兴奋作用,有 些中间神经元,特别是篮状细胞有抑制 作用。
海马结构的皮质构造
海马伞可以认为是穹隆的起点,沿侧脑 室下角底壁后进,而后弯向上,到胼胝 体压部的下方,大部分纤维转向前,构 成穹隆脚。左、右穹隆脚在胼胝体压部 的下方逐渐靠近,其中有部分纤维越至 对侧,经对侧海马脚、海马伞,至对侧 海马结构,于是在两侧海马脚之间形成 一个薄的三角形白质板,叫穹隆连合 commissure of fornix,也叫海马连合。
锥体细胞的轴突发自细胞的基底部或底 树突,轴突进入海马槽,后进构成海马 伞。轴突在海马槽内还发出侧支,侧支 大部分返回分子层,终于邻近锥体细胞 的尖树突,部分终于多形细胞层。
1.4辐射层 stratum radiatum 由锥体
细胞尖树突和轴突的返回侧支组成,因 纤维排列规则呈放射状而得名。
海马结构的皮质构造
纤维数量很大,它们经由两条通路到达海 马和齿状回。其一是来自内嗅区内侧部和 下托的纤维,到达海马皮质的多形层,主 要止于锥体细胞;其二是来自内嗅区外侧 部的纤维,到达海马皮质的分子层,止于 齿状回和海马之全长。
海马结构(HF)

海马结构(HF)来源:内蒙古医学影像论坛作者:李木子商务合作:*****************版主微信号:fsslong2海马结构(HF)是大脑边缘系统的重要组成部分,属大脑古皮质,位于端脑颞叶的内侧深部,左右各一。
海马结构的组成:海马(Ammon’s角)、海马伞、齿状回、束状回、下托、围绕胼胝体的海马残体(胼胝体上回/灰背)、海马旁回、钩(深面为杏仁体)。
深红色 GD 齿状回浅红色 CA 海马白色 Sub 下托灰色 GPh 海马旁回在海马结构发育较好的颞中平面,作一个大脑半球的冠状切面,海马结构呈双重“C”形环抱的外形,大C锁住小C。
大C代表海马,它开口向腹内侧。
小C代表齿状回,位于海马沟的背内侧,开口朝向背侧。
海马沟的腹侧为下托(subiculum)。
灰质部分:海马--齿状回--束状回--灰被白质部分:海马伞--穹窿脚--穹窿联合--穹窿海马(hippocampus)=Ammon角Ammon角,因形似海马而得名,弓形隆起,5cm,古皮质海马由海马沟被挤到侧脑室下角底壁(内侧壁),由海马沟内陷卷曲而成,前段较宽,有时有2-3个浅沟将其分割成若干个隆起,称海马足(头、脚),沟间隆起为海马趾。
海马位于侧脑室颞角后方、内侧,与灰质等信号;海马趾间脑脊液海马(Ammon 角)分为头、体、尾三部分:头部因其上缘有海马趾,而呈波浪形外观体位于海马沟与脉络膜裂之间尾的特征是海马槽形成海马伞海马常规扫描方式●常规扫描方位:轴位,冠状位。
●轴位:以冠状位和矢状位作为参考定位。
在冠状位上定位线平行于两侧颞叶底部的连线;矢状面上平行于前后联合的连线或者与胼胝体的前后连线平行。
扫描范围至少包括整颞叶范围。
●冠状位:以矢状位和轴位作为参考定位。
在轴位上与大脑纵裂垂直;在矢状位上定位线与海马长轴垂直。
扫描范围包括整个颞叶及海马。
层厚3mm● T2需加上下饱和带。
【今日推荐】《中华医学影像技术学》丛书国内该领域专家理论和实践的全面展现中华医学会影像技术分会倾心之作!。
阿尔兹海默症诊断标准

阿尔兹海默症诊断标准
阿尔茨海默症的诊断标准:
1.临床表现:年龄在40-90岁,表现为认知能力下降,尤其是近期记忆力障碍,并有明显精神和行为异常,后期生活不能自理。
2.影像学检查:头颅磁共振可发现大脑半球萎缩,海马结构及后扣带回萎缩。
脑电图为弥漫性的慢波,可明确诊断。
3. 精神量表:可进行相关认知量表检查,如阿尔茨海默病行为病理评定量表、蒙特利尔认知评估、简易精神量表等相关认知量表,以判断患者是否存在进行性认知功能障碍。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
海马结构一.形态海马结构(hippocampal formation)包括海马(hippocampus)又称安蒙角(Ammon’s born),齿状回(dentate gyrus)和围绕胼胝体形成一圈的海马残件(灰被indusium grisem).齿状回随海马伞向后,至胼胝体压部,它与海马伞分开,改为束状回,束状回向前上与覆盖在胼胝体上面的灰质称胼胝体上回(ssupracallosal gyrus)(灰被)相连续,灰被中埋有一对纵纹,分别为内侧纵纹和外侧纵纹。
灰被与纵纹就是海马及其白质的残件,它们向前经胼胝体膝与胼胝体下回连续。
(一)海马海马形似中药海马,故得名。
其位于侧脑室下角的底和内侧壁,全长约5cm,前段较膨大,称海马角,他被2-3个浅沟分开,沟间隆起,称海马趾;海马表面被室管膜上皮覆盖,室管膜上皮下面一层有髓鞘纤维称室床,室床纤维沿海马背内侧缘集中,形成白色扁带称海马伞,构成穹窿系统的起始步,它自海马趾伸向压部,续于穹窿角。
海马内的细胞构筑分为三层,从海马裂到脑室依次为①分子层;②椎体细胞层;③多形层。
根据细胞形态和皮质区发育差异等特点,在横断面上海马又可分为CA1、CA2、CA3和CA4四个区。
CA4位于齿状回门内,内有大的椎体细胞;CA3有来自齿状回颗粒细胞的轴突(即苔状纤维);CA2内有少量轴突;CA1内含有小的椎体细胞。
(二)齿状回齿状回是一条灰皮质,由于血管进入形成沟而成齿状,故名。
它位于海马的内侧,海马裂与海马伞之间,齿状回向后与束装回相连,其前端抵海马回钩和海马回之间。
海马接受扣带回来的纤维经扣带直接或间接地终止于海马,从隔核发出的纤维经穹窿,海马伞终止于海马CA3、CA4区和齿状回。
一侧的海马也可经同侧海马伞,穹窿脚,通过海马联合投射至对侧的海马和齿状回,海马还可经室床通路接受内嗅区外侧份的传出纤维,这些纤维主要分布于CA1区和下托的深层,内侧份纤维则经穿通道、下托进入海马CA1-CA3斜角带核,在穹窿的行程中发出纤维至丘脑前核和板内核的吻部,部分纤维可向尾侧进入中脑被盖和中央灰质。
二.海马的功能海马具有多方面的生理功能,50年代不少实验已经证明海马可接受来自外周的视觉、听觉、触觉、痛觉、本体感受性和内感受性刺激,感受性冲动经过脑干网状结构传递至海马,引起海马电活动的变化。
近年来,由于神经科学的迅速发展,不少学者认为海马是情感和学习记忆等高级神经活动的重要部位。
(一)行为反应毁损海马后,动物的行为发生一系列变化,最重要的是更乐于从事新的活动,在恐惧或应激的情况下,动物好动反应灵活,热衷于进行新的活动,有时也会出现“幻觉”,但很少出现攻击性动作。
这些反应对动物具有一定的生理意义,如毁损海马的鼠,当遇到猫时,其表现为饥饿情绪反应增强,食欲亢进,性活动异常以及性情温顺,但辨认能力和记忆能力减退。
毁损海马不同区域,对动物的行为有着不同的影响,如毁损全部海马或仅毁损海马腹部的动物较毁损背部者活动增加更明显;毁损海马腹部,动物的允水频率增加,而毁损背部则不出现此现象。
说明在行为变化中,海马内存在着区域性差异。
(二)植物性功能早期实验电刺激猫、狗和猴的海马,没有发现呼吸和血压的变化,因此称海马为“内脏活动静止区(autonomically silent regions)”.70年代有报道电刺激海马可引起一系列的内脏反应,如动物鸣叫、呼吸减慢或停止、心率和血压也有变化,同时尚可出现流延、恶心、瞳孔扩大和竖毛等反应;但刺激海马所引起的植物性功能变化一般是轻而短暂的。
(三)海马的Q节律从大鼠、兔、猫等哺乳类动物海马上记录的海马电活动,可分为三种类型:①慢正弦节律(rhythmical slow activity ,RSA),是一种特殊的同步化慢波,频率4-7Hz,又称Q节律(Q rhythmus);②高振幅不规则慢波(large irregular activity, LIA),频率比Q节律少,发放持续时间50-100ms;③高频低振幅去同步化波(small irregular activity,SIA),发放持续时间在大鼠不超过1-2s,在兔和猫持续时间则较长。
海马电活动不仅是海马神经元活动的表现,而且与动物的行为有关,特别是Q节律,可作为指标来研究动物行为或精神活动与海马神经元活动之间的关系。
1.Q节律发生的机制清醒动物,当各种感觉刺激(如嗅、视、听或触觉等)引起动物注意反应时或电刺激脑干网状结构,均可在海马记录到Q节律,当毁损隔区或切断穹窿后,同样刺则不出现Q节律。
因而认为Q节律的发生是由于脑干网状结构被激活,上行经隔核或穹窿至海马,激活了海马的CA1锥体细胞和齿状回颗粒细胞活动的结果。
也有实验证明,海马Q节律的活动是依赖于海马与背内测隔核的联系,当该核被毁损时,则感觉刺激以及电刺激脑干网状结构所引起的海马Q节律被消除,同时海马自发活动的振幅也降低;然而毁损外侧隔核则不影响Q节律的发放。
实验中又观察到,隔核神经元的活动不受海马活动的影响,如应用5-HT受体拮抗剂麦角酰二乙胺(D-lysergic acid diethylamide,LSD)可以抑制依色林(eserine)诱发的海马Q节律,但不抑制隔核神经元节律性发放;此外,应用低电频电刺激内侧隔核产生的海马Q节律与自然存在的Q节律与自然存在的Q节律相似。
而目前较多学者认为,隔核细胞的作用如海马Q节律的起搏点,隔核内胆碱能细胞在调制海马Q节律中有着重要的作用。
2.Q节律的生理意义一般人认为海马与脑干网状结构相联系,参与维持觉醒状态。
当动物接受各种传入刺激引起皮质出现去同步化快波时,海马则出现同步化的Q节律;当动物接受新异动因刺激出现朝向反应和注意的行为活动时,海马的Q节律也出现。
但当动物处于瞌睡或入睡状态在皮质出现慢波时,海马则出现快的去同步电活动。
因此,不少学者认为Q节律是动物处于“注意”、“警觉”状态或探究反射时海马出现的电活动,是一种特殊形式的“警觉反应”。
然而在大白鼠实验中发现,Q节律往往和躯体运动同时出现,在静止时虽表现为警觉状态,但不出现Q节律,故认为Q节律和躯体活动有关。
近年来又有人认为Q波节律与某些动物的典型行为有关,如鼠的探索与鼻嗅和兔的奔跑等,当这些具有生物生物学意义的行为占优势时,在海马上记录到的主要是Q节律。
可以认为当中枢正在组织及调节行为活动时,海马却受到抑制性调节。
因此,海马Q 节律的意义有人认为是代表激活反应,是中枢“觉醒反应”的一部分;而另一种意见则相反,认为Q节律是海马被抑制的表现。
(四)学习与记忆多年来人们与记忆在脑内的定位问题进行了大量的研究,从脑损伤及脑电图的资料表明,学习和记忆是皮质和皮质下大量神经元以一定的时空模式高度配合完成的。
越是复杂的学习记忆,越需要较高水平的神经结构的作用也越重要。
目前认为,颞叶→海马回→海马→穹窿→下丘脑乳头体→丘脑前核→扣带回→海马结构所构成的海马环路,可能与学习记忆有关。
在动物实验中,毁损海马能促进操作性逃避工作的学习,但可减弱在“T”迷宫工作逃避足部电击的视觉辨别问题的学习。
这种逃避条件反应的差异可能是由于毁损海马的动物,不但影响空间的位置信息的识别能力,同时也降低了动物对亮度信息的识别能力。
临床资料也证明海马参与学习记忆过程。
各种精神病特别是记忆丧失症的患者,在其海马内可找到病理性变化。
为了治疗颞叶癫痫而进行海马切除,会导致严重的并发症并牵涉到病人的心理改变以及新近记忆的丧失,丧失程度决定于切除部位的大小。
在手术切除第三脑室囊肿而损伤穹窿时,也可使患者丧失近期记忆。
双侧海马损伤的患者,当他们在一段时间内集中注意力,则可以记住一个短句或一个短位的数字,但当患者把注意力转向某些事物时,即使是片刻,也将完全忘记这个短句或短位数字。
由于这种障碍,使患者不能学习新事物,也不能记忆新近的经历,但对于发病前工作中获得的技能以及生活中曾发生的事情,仍有良好的记忆。
这说明海马与近期记忆有密切关系,可能与第一级记忆向第二级记忆转移过程有关。
看来记忆痕迹主要并非存储于海马,海马可能参与选择应予存储的信息以及从存储记忆中找寻和提取有关的信息,即参与储存记忆痕迹的神经过程。
与记忆有关的海马结构神经元主要是锥体细胞和齿状回颗粒细胞。
海马环路活动可促进学习和记忆,因环路活动不仅使神经元的活动时间延长,而且可使神经元的活动发生总和而不断增强,从而使神经元间的突触联系容易化,有助于促进学习和记忆过程。
中枢胆碱能系统的活动与学习记忆有密切关系。
有学者提出,胆碱能神经通路参与构成记忆痕迹,胆碱能突触即为记忆突触,它是学习记忆的结构和生理基础。
据报道,用药物阻滞胆碱能系统所诱发的学习记忆功能障碍与海马结构损伤者极为相似,如学习能力下降和严重的顺行性遗忘等,但远期记忆一般不受影响。
有学者认为,可能是在学习记忆过程中记忆突触对Ach的敏感性增加,神经元的兴奋性增高,而利于记忆痕迹的储存。
隔-海马-边缘叶胆碱能通路在此过程中起着重要的作用。
虽然有不少事实说明海马环路与学习记忆有关,但也存在着矛盾的结果和相反的意见。
有实验观察到毁损动物的海马或穹窿并不明显影响学习记忆。
临床上有1例病例,经检查两侧海马小而异常,并且不存在有穹窿和海马伞,而病人生前的学习记忆成绩名列前茅,无行为或腺癌破坏两侧穹窿和乳头体的病人也没有记忆缺损。
因而认为,内侧颞叶切除引起的记忆障碍主要是由于破坏了颞干和颞叶新皮质。
颞干是指颞叶内侧,颞叶皮质和杏仁传入与传出纤维集中的白质,并不还有海马纤维。
切断猴的颞干后,影响动物的学习记忆能力;破坏海马和海马回而不损及颞干时并不引起记忆障碍。
因此,有关记忆的神经基础还有待进一步(五)与内分泌的关系激素的昼夜节律性释放与穹窿系统有关,切除穹窿的动物皮质类固醇分泌的昼夜节律降低。
海马内存有特异的类固醇受体,对促进肾上腺皮质激素分泌具有反馈调控的作用。
刺激海马可影响肾上腺皮质对各种应激情况的反应。
此反应可根据动物所处的状态而起抑制或异化作用,当动物在应激情况下刺激海马,可降低皮质类固醇水平;反之则促进分泌。
促肾上腺皮质激素和肾上腺皮质激素能影响海马细胞的电活动。
切除肾上腺的大鼠,注射皮质类固醇可抑制海马锥体细胞神经元的单位放电,注射肾上腺皮质激素则放电增加。
海马对促性腺激素的分泌也具有抑制作用。
电刺激海马,可抑制排卵以及黄体生成激素或卵泡雌激素的分泌;在海马内埋置雌激素可降低血清中卵泡激素的含量;切断海马伞可加速雌鼠的成熟。
上述资料说明海马与内分泌之间有着密切的关系。
(六)对痛觉的调制切除狒狒和猴的海马,动物对痛的反应降低;为恶痛病人施行海马切除,能使疼痛缓解。
刺激猫海马背侧,可出现较长时间的镇痛作用。
提示海马可能参与痛觉的调制。
在记录猕猴或猫海马自发电活动的实验中,给予躯体或内脏的伤害性刺激,发现海马的Q节律发生明显改变,说明伤害性刺激可到达海马,并且影响海马的电活动。