原核与真核生物mRNA的特征比较
原核mrna结构特点

原核mrna结构特点原核mRNA(messenger RNA)是一类具有特殊结构特点的核糖核酸分子。
mRNA是基因表达的中间产物,将DNA中的遗传信息转录为蛋白质合成的模板。
与真核生物的mRNA相比,原核生物的mRNA结构相对简单。
原核mRNA的基因组结构通常是单个连续的开放阅读框(ORF,Open Reading Frame),没有内含子(intron)。
这意味着原核生物的基因组中的大部分区域都可以被转录为mRNA,而不需要经过剪接过程。
相比之下,真核生物的基因组中含有大量的内含子,需要通过剪接来选择性地将外显子连接起来。
原核mRNA的起始密码子和终止密码子相对保守。
起始密码子通常是AUG,编码甲硫氨酸(methionine),而终止密码子可以是UAA、UAG或UGA。
这些起始和终止密码子在原核生物中具有普遍性,用于识别蛋白质的起始和终止位置。
原核mRNA的5'端通常具有一个甲基鸟苷帽(m7G cap),这是一个被甲基化的鸟苷三磷酸分子。
甲基鸟苷帽可以保护mRNA免受核酸酶降解,并且在蛋白质合成中起到重要的调节作用。
真核生物的mRNA也具有甲基鸟苷帽结构,但其合成过程与原核生物有所不同。
原核mRNA的3'端通常没有聚腺苷酸尾巴(poly-A tail)。
聚腺苷酸尾巴是一段由腺苷酸组成的多聚核苷酸序列,附加在真核mRNA的3'端。
聚腺苷酸尾巴可以提高mRNA的稳定性,并参与转录后修饰和mRNA的转运。
在原核生物中,由于缺乏聚腺苷酸尾巴,mRNA的稳定性通常较低。
原核mRNA的转录和翻译过程是同时进行的,即转录和翻译可以在同一时间和同一位置进行。
这种耦合的转录-翻译机制使得原核生物的蛋白质合成速度更高,更加高效。
从这些结构特点可以看出,原核mRNA相对于真核生物的mRNA 来说更加简单和直接。
这种简单性和直接性使得原核生物能够更快地合成蛋白质,并适应其快速的生长和繁殖需求。
分子生物学-07-3-生物信息的传递-3转录后加工-小RNA-RNA拼接1
能够识别发夹 结构的内切酶
1
RNAase P 3
2 RNA酶D
4 4
4
4
2020/10/28
多种酶的处理
2020/10/28
23
tRNA的内含子去除
2020/10/28
2020/10/28
22
rRNA的转录后处理
1、原核生物和真核生物是较为相似的,分别由多种
rRNA组成它的核糖体,包括5S rRNA 、5.8S S rRNA、 18 S rRNA、 28 S rRNA
3.6原核生物RNA转录与真核生物的比较
Eukaryotes and prokaryotes produce mRNAs somewhat differently
2020/10/28
2020/10/28
1
3.6.1 原核生物mRNA的特征
1. 多顺反子,有共同的起始子和终止信号 2. 转录和翻译的时空性 3. 5′端无帽子结构, 3′端没有或只有较短的poly(A)结构 4. 起始密码子常为AUG,有时也为GUG,甚至UUG 5. 半衰期短
14
Poly(A)尾巴的功能:
(1)尾巴可能与核-质转运有关;但是无尾巴的 mRNA,如组蛋白的mRNA同样可以通过核膜 进入细胞质
(2)尾巴的长短对mRNA的翻译也无影响,因 此其功能无定论。
(3)细胞质中mRNA 的Poly(A)尾巴与蛋白质相 结合构成mRNP(一种蛋白质分子)。其功能 不清楚。
2020/10/28
帽子结构功能:
1.能被核糖体小亚基识别,促使mRNA和 核糖体的结合;
2.m7Gppp结构能有效地封闭mRNA 5’末 端,以保护mRNA免受5’核酸外切酶的降解 ,增强mRNA的稳定。
原核生物和真核生物基因表达调控特点的比较知识讲解

翻译的起始 起始tRNA是fMet-tRNA,30s小 起始tRNA是 Met-tRNA,40s小亚基首先
亚基首先与mRNA模板相结合, 与Met-tRNA相结合,再与模板mRNA结
再与fMet-tRNA结合,最后与 合,最后与60s大亚基结合生成起始复
50s大亚基结合
合物
肽链的终止
三种释放因子RF1,RF2,RF3
eRF1和eRF3
真核生物和原核生物复制的不同点:
• 真核生物DNA的合成只是在细胞周期的S期进行,而原核生物则 在整个细胞生长过程中都可进行DNA合成
• 原核生物DNA的复制是单起点的,而真核生物染色体的复制则 为多起点的。
• 真核生物中前导链的合成并不像原核生物那样是连续的,而是 以半连续的方式,由一个复制起点控制一个复制子的合成,最 后由连接酶将其连接成一条完整的新链。
原核生物
真核生物
场所
细胞核内
拟核区
mRNA
mRNA分子一般只编码一个基因
一个mRNA分子通常含多个基因RNA聚合酶一种
三种
独立转录
可以直接起始转录合成RNA 不能独立转录RNA,三种聚合酶都必须
在蛋白质转录因子的协助下才能进行
翻译
原核生物
RNA的转录
真核生物
氨基酸的活化 起始氨基酸是甲酰甲硫氨酸
从生成甲硫氨酰-tRNAi开始
原核生物转录过程示意图
真核生物转录 过程示意图
原核生物和真核生物基因表达调控特点的比较——不同点——转录
真核生物和原核生物翻译的不同点:
• 氨基酸的活化:原核起始氨基酸是甲酰甲硫氨酸,真核是从生 成甲硫氨酰-tRNAi开始的。
• 翻译的起始:原核的起始tRNA是fMet-tRNA,30s小亚基首先与 mRNA模板相结合,再与fMet-tRNA结合,最后与50s大亚基结合。 真核中起始tRNA是 Met-tRNA,40s小亚基首先与Met-tRNA相结合, 再与模板mRNA结合,最后与60s大亚基结合生成起始复合物。
三、真核生物、原核生物mRNA的区别;真核生物前mRNA的加工;tRNA二级结构与功能

4.胸苷假尿苷胞苷臂thymidine-pseudouridine-cytidine arm :包括由5对碱
基对组成的短双螺旋区T,C茎和7个核苷酸围成的T,C环,通过T,C茎与tRNA的 其余部分相连,主要碱基对是C-G,实现tRNA与核糖体大小亚基的结合,这是
tRNA转运AA和肽键在核糖体上生成的必要条件,维系和稳定tRNA的三级结构。
5.可变臂variable:由3~21个核苷酸组成可变环,含量多者有可能形成小双螺旋, 是tRNA分类标志。
嘧啶核苷酸处的一段序列内,一定含有一个特定的腺苷酸(A),称为 分支点,内含子5 ′端、3 ′端和分支点对于剪接的发生是必需的。
tRNA的二级结构及功能(三叶草形
1. 氨基酸臂amino acid arm:包括由7个碱基对组成的局部双螺旋区 氨基酸接受茎和3‘端的CCAOH组成,CCAOH称接受端,可接受 活化的AA,是各种tRNA共同的3’端序列,CCAOH的存在对于 tRNA与核蛋白体大亚基的结合是必不可少的。 2. 二氢尿嘧啶臂DHU arm:由3~4个碱基对组成的短双螺旋区形成的 二氢尿嘧啶茎,其中第一个和最后一个碱基对通常是C-G,及由7-10
三、真核生物、原核生物mRNA的区别;真核生物 前mRNA的加工;tRNA二级结构与功能
原核生物mRNA的结构 1、典型结构:多顺反子(polycistron):即一个操纵子负责一段产物 对几条多肽链编码。 2、 5′端无帽子结构,分子中一般没有修饰核苷酸,不含有稀有碱基 3、 3 ′端无多聚A序列,或仅有很短的多聚A(少于10个)。 真核生物mRNA的结构: 1. 5′端帽子结构 真核生物普遍存在一些甲基化核苷,以特殊的方 式连接,称帽子结构。帽子结构分三种类型:m7G5’ppp5’N和 m7G5’ppp5’Nm 和m7G5’ppp5’N1mN2m。功能: (防止mRNA被核酸 酶降解、为mRNA 翻译活性所必需、与蛋白质合成的起始有关,从细 胞核向细胞质转运和增加其稳定性有关) 2. 3 ′端多聚A结构:包含200-300个残基(保护mRNA 免受核酸外切酶 的作用、与翻译有关,没有ployA翻译活性降低、与mRNA 从细胞核转 移到细胞质有关)
试比较原核和真核细胞的mRNA的异同

第一章绪论分子生物学:从分子水平研究生物大分子的结构与功能从而阐明生命现象本质的科学,主要指遗传信息的传递(复制)、保持(损伤和修复)、基因的表达(转录和翻译)与调控。
分子生物学研究内容● DNA重组技术(基因工程)1、可被用于大量生产某些在正常细胞代谢中产量很低的多肽;2、可用于定向改造某些生物的基因组结构;3、可被用来进行基础研究● 基因的表达调控1信号转导研究;2转录因子;3研究RAN剪接● 生物大分子的结构和功能研究(结构分子生物学)● 基因组、功能基因组与生物信息学研究第二章染色体和DNADNA复制时其中一条子链的合成是连续的,而另一条子链的合成是不连续的,故称半不连续复制在DNA复制时,合成方向与复制叉移动的方向一致并连续合成的链为前导链;合成方向与复制叉移动的方向从复制原点到终点,组成一个复制单位,叫复制子相反,形成许多不连续的片段,最后再连成一条完整的DNA链为滞后链。
DNA的复制过程(大肠杆菌为例)双链的解开;DNA的复制有特定的起始位点,叫做复制原点。
ori(或o)、富含A、T的区段RNA引物的合成:DnaB蛋白活化引物合成酶,引发RNA引物的合成。
引物长度约为几个至10个核苷酸,DNA链的延伸切除RNA引物,填补缺口,连接相邻的DNA片段复制时,解链酶等先将DNA的一段双链解开,形成复制点,这个复制点的形状象一个叉子,故称为复制叉在DNA复制过程中,前导链能连续合成,而滞后链只能是断续的合成5 3 的多个短片段,这些不连续的小片段称为冈崎片段。
复制的几种主要方式P421、双链环状、θ型复制、双向等速2、滚环型:单向复制的特殊方式如:ΦΧ174的双链环状DNA复制型(RF)(1)模板链和新合成的链分开;(2)不需RNA引物,在正链3‘-OH上延伸(3)只有一个复制叉;3、D环复制单向复制的特殊方式如:动物线粒体DNA真核生物中DNA的复制特点1、真核生物每条染色体上有多个复制起点,多复制子(约150bp左右);2、复制叉移动的速度较慢(约50bp/秒),仅为原核生物的1/10。
第三章mRNA特征,终止与内含子编辑

1、原核生物mRNA与真核生物mRNA的区别(1)半衰期短(转录和翻译是在同一个细胞空间里同步进行)。
真核细胞mRNA的合成和功能表达发生在不同的空间和时间范畴内。
(2)许多原核生物mRNA以多顺反子的形式存在。
真核生物单顺反子形式存在。
(3)原核生物mRNA的5’端无帽子结构,3’端没有或只有较短的多聚(A)结构。
真核生物5’端存在“帽子(7甲基-鸟苷酸)”结构,绝大多数具有多聚(A)尾巴。
(4)原核生物常以AUG(有时GUG,甚至UUG)作为起始密码子;真核生物几乎永远以AUG作为起始密码子。
2、单顺反子mRNA (monocistronicmRNA):真核生物转录的产物,只编码一个蛋白质的mRNA。
顺反子:指的是结构基因,3、多顺反子mRNA(polycistronicmRNA):原核生物转录产物,编码多个蛋白质的mRNA。
4、基因:产生一条多肽链或功能RNA所必需的全部核苷酸序列。
5、真核生物基因转录一般从嘌呤起始。
6、mRNA的帽子结构常常被甲基化,是蛋白质合成起始信号的一部分。
7、mRNA的帽子类型:零类帽子(第一个核苷酸被甲基化)、1类帽子(mRNA的主要形式,第二个核苷酸的2’-OH也被甲基化)、2类帽子(第3个核苷酸的2’-OH再被甲基化)。
8、帽子结构的功能:(1)有助于mRNA越过核膜,进入胞质;(2)保护5′不被核酶降解;(3)翻译时供IFⅢ(起始因子)和核糖体识别,是翻译所必需的。
9、Poly(A):它是在转录后加上的。
10、多聚(A)的功能(1)是mRNA由细胞核进入细胞质所必需的形式;(2)它大大提高了mRNA在细胞质中的稳定性。
(3)它可促进核糖体的有效循环。
11、在动物细胞中,mRNA的表达需转录、修饰、加工、核质转运和翻译。
12、内含子在真核基因中所占的比例很高,甚至超过99%。
13、mRNA的前体(核不均一RNA,hnRNA) 5’加“帽”和3’酶切加多聚腺苷酸 RNA的剪接连续的可译框(ORF) 进入胞质蛋白质合成的模板14、RNA加工过程及其生理功能加工过程推测的生理功能加帽子反应mRNA从细胞核向细胞质运转,翻译起始加多聚A反应转录终止,翻译起始和mRNA降解RNA的剪接从mRNA, tRNA和rRNA分子中切除内含子RNA的切割从前体RNA中释放成熟tRNA和rRNA分子15、不同生物细胞内含子的边界存在相似的核苷酸序列。
原核mRNA

原核生物转录的mRNA是一个多顺反子。
• 几乎所有mRNA都可以被分成3部分:编码 区和位于AUG之前的5'端上游非编码区以及 位于终止密码子之后不翻译的3'端下游非编 码区。编码区从起始密码子AUG开始经一 连串编码氨基酸的密码子直至终止密码子。 对于第一个顺反子来说,一旦mRNA的5'端 被合成,翻译起始位点即可与核糖体结合, 而后面几个顺反子翻译的起始就会受其上 游顺反子结构的调控。
真核生物
主要在核质 hnRNA,需加工 单顺反子,5’ 端有cap,3’ 端有poly A 较稳定,半衰 期为24h
名词解释
1. 编码区:从起始密码子AUG开始,经过一连串 编码氨基酸的密码子直至终止密码子的碱基 序列 2. 5’端上游非编码区(5’UTR):位于ATG之 前不编码的碱基序列 3. 3’端下游非编码区(3’UTR):位于终止密 码子之后不翻译的区域 4. 单顺反子mRNA:只编码一个蛋白的mRNA 5. 多顺反子mRNA:编码多个蛋白质的mRNA
2
在原核生物中(包括病毒),一个mRNA分子能够编码 一种或多种蛋白质,所有蛋白质都能翻译出来。
单顺反子mRNA(monocistronic mRNA)把只编码一个 蛋白质的mRNA称为单顺反子mRNA。 多顺反子mRNA(polycistronic mRNA):把编码多个 蛋白质的mRNA称为多顺反子mRNA。
3.帽子结构的功能
① 有助于mRNA越过核膜,进入胞质; ② 保护5’端不被核膜降解; ③ 翻译时供IFiii和核糖体识别,是翻译所必须的; 而原核生物是通过起始密码子AUG上游的一段 SD序列的保守区与核糖体结合
4.多聚A尾的功能
① 是mRNA由核内进入胞质所必须的 ② 增加了mRNA在细胞质内的稳定性,mRNA刚进 入胞质时,A尾较长,随着mRNA在胞质内时间 的延长,A尾逐渐变短,mRNA进入降解的过程 ③ 可以促进核糖体的有效循环
真核生物和原核生物mRNA的区别
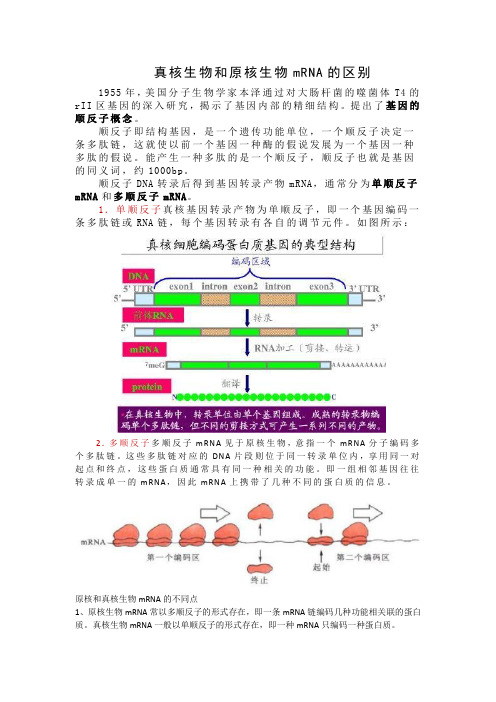
真核生物和原核生物mRNA的区别1955年,美国分子生物学家本泽通过对大肠杆菌的噬菌体T4的rII区基因的深入研究,揭示了基因内部的精细结构。
提出了基因的顺反子概念。
顺反子即结构基因,是一个遗传功能单位,一个顺反子决定一条多肽链,这就使以前一个基因一种酶的假说发展为一个基因一种多肽的假说。
能产生一种多肽的是一个顺反子,顺反子也就是基因的同义词,约1000bp。
顺反子DNA转录后得到基因转录产物mRNA,通常分为单顺反子mRNA和多顺反子mRNA。
1.单顺反子真核基因转录产物为单顺反子,即一个基因编码一条多肽链或RNA链,每个基因转录有各自的调节元件。
如图所示:2.多顺反子多顺反子mRNA见于原核生物,意指一个mRNA分子编码多个多肽链。
这些多肽链对应的DNA片段则位于同一转录单位内,享用同一对起点和终点,这些蛋白质通常具有同一种相关的功能。
即一组相邻基因往往转录成单一的mRNA,因此mRNA上携带了几种不同的蛋白质的信息。
原核和真核生物mRNA的不同点1、原核生物mRNA常以多顺反子的形式存在,即一条mRNA链编码几种功能相关联的蛋白质。
真核生物mRNA一般以单顺反子的形式存在,即一种mRNA只编码一种蛋白质。
2、原核生物mRNA的转录与翻译一般是偶联的,即转录尚未完毕,蛋白质的转译合成就已开始。
真核生物转录的mRNA前体则需经过加工,加工为成熟的mRNA与蛋白质结合生成信息体后才开始,工作信息体中蛋白质与RNA之比约为3。
3、原核生物mRNA半寿期很短,一般为几分钟,最长只有数小时。
真核生物mRNA的半寿期较长,如胚胎中的mRNA可达数日。
4、原核与真核生物mRNA的结构特点也不同。
真核生物mRNA由5′端帽子结构、5′端不翻译区、翻译区、3′端不翻译区和3′端聚腺苷酸尾巴组成,原核生物mRNA无5′端帽子结构和3′端聚腺苷酸尾巴。
分子生物技术问答题-农大期末考试复习题

1.试比较DNA复制与转录的异同点。
相同点:都以DNA链作为模板,合成的方向均为5’→3’,聚合反应均遵从碱基配对原则,核苷酸之间形成磷酸二酯键使核苷酸链延伸。
不同点:复制转录模板两股链均复制,全部信息模板链,部分信息原料dNTP NTP酶DNA聚合酶RNA聚合酶产物子代双链DNA(半保留复制)mRNA,tRNA,rRNA引物RNA引物无配对G-C; T-A A-U; T-A; C-G2.简述真核与原核基因转录方面存在的差异。
原核生物真核生物RNA聚合酶只有1种。
3种,分别转录不同的RNAmRNA结构通常5‘帽子和3’尾巴顺反子功能相近的基因通常形成一个操纵子,由共同的调控区进行转录调控。
转录起始不同的聚合酶有不同的启动子调控转录终止在RNA水平发生作用:不依赖ρ因子的终止子在柄部富含GC碱基对,而且连接一串富含U的茎环结构;依赖于ρ因子的终止子通过ρ因子与β亚基的作用促使转录终止。
3类RNA聚合酶的转录因子都需要富含AT的序列。
3.简述RNA的种类及其生物学作用。
4.比较真核生物和原核生物mRNA的异同。
原核生物和真核生物mRNA的差别在于可翻译顺反子的数目,真核生物的mRNA是单顺反子。
而且真核生物的mRNA在其3’末端有多聚腺苷酸尾巴(polyA),而5’末端有7-甲基鸟嘌呤帽子。
真核生物的mRNA尾部区域有时会携带特定的去稳定因子。
5.简述真核生物mRNA基因的转录过程。
1) 模板识别:RNA聚合酶与双链DNA的启动子开始结合;DNA双链被分开;形成转录泡。
2) 转录的起始:RNA中第一个核苷酸键合成;合成前9 个核苷酸键时,酶仍旧位于启动子上;聚合酶成功地完成了RNA链的延伸并离开启动子。
3) 延伸阶段:延伸阶段包括DNA 结构的改变带来的转录泡前移。
在转录泡中,模板链的瞬间解旋区与新生的RNA链在延伸点发生配对。
RNA聚合酶沿DNA移动,RNA链逐渐延长,酶一边移动一边将DNA解螺旋,使一段新的模板以单链形式暴露出来。
原核生物和真核生物基因表达调控复制、转录、翻译特点的比较

原核生物和真核生物基因表达调控、复制、转录、翻译特点的比较1.相同点:转录起始是基因表达调控的关键环节①结构基因均有调控序列;②表达过程都具有复杂性,表现为多环节;③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性;2.不同点:①原核基因的表达调控主要包括转录和翻译水平。
真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。
②原核基因表达调控主要为负调控,真核主要为正调控。
③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。
④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。
⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。
原核生物基因以操纵子的形式存在。
转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。
翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。
真核生物基因表达的调控环节较多:在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。
在转录水平主要通过反式作用因子调控转录因子与TA TA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。
在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。
在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。
真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。
分子生物学考试复习题及答案

分⼦⽣物学考试复习题及答案第⼀章绪论⼀.简述分⼦⽣物学的主要内容。
1.DNA重组技术(⼜称基因⼯程)2.基因表达调控研究3.⽣物⼤分⼦的结构功能的研究——结构分⼦⽣物学4.基因组、功能基因组与⽣物信息学研究⼆.什么是遗传学的中⼼法则和反中⼼法则?遗传学中⼼法则:描述从⼀个基因到相应蛋⽩质的信息流的途径。
遗传信息贮存在DNA中,DNA被复制传给⼦代细胞,信息被拷贝或由DNA转录成RNA,然后RNA翻译成多肽。
反中⼼法则:由RNA逆转录到DNA,再由DNA转录到RNA,然后RNA翻译成多肽、蛋⽩质。
第⼆章染⾊体与DNA⼀、名词解释半保留复制:DNA复制时,以亲代DNA的每⼀条链作为模版,合成完全相同的两个双链⾃带DNA分⼦,每个⼦代DNA分⼦中都含有⼀条亲代DNA链的复制⽅式。
冈崎⽚段:在DNA复制过程中,前导链能连续合成,⽽滞后链只能是断续的合成5 3 的多个短⽚段,这些不连续的⼩⽚段称为冈崎⽚段。
复制⼦:从复制原点到终点,组成⼀个复制单位,叫复制⼦。
插⼊序列:最简单的转座⼦,不含有任何宿主基因,它们是细菌染⾊体或质粒DNA的正常组成部分。
DNA的变性和复性:变性:指DNA双链的氢键断裂,完全变成单链。
复性:指热变性的DNA缓慢冷却,单链恢复成双链。
双向复制:复制从⼀个固定的起始点开始,同时向两个⽅向等速进⾏。
引物酶:⼀种特殊的RNA聚合酶,在DNA模版上合成⼀段RNA链,这段RNA链是DNA复制起始时必需的引物。
转座⼦:存在与染⾊体DNA上可⾃主复制和位移的基本单位。
碱基切除修复:⾸先由糖苷⽔解酶识别特定受损核苷酸上的N-β糖苷键,在DNA上形成AP位点,由AP核酸内切酶把受损的核苷酸的糖苷-磷酸键切开,移去包括AP位点在内的⼩⽚段DNA,由聚合酶Ⅰ合成新⽚段,DNA连接酶最终把两者连接成新的DNA链。
DNA重组修复:即“复制后修复”。
机体细胞对在复制起始时尚未修复的DNA部位可先复制再修复。
在复制时,先跳过损伤部位,在和成链中留下⼀个缺⼝。
原核生物与真核生物mRNA比较

大多数的真核mRNA转录后在5'-端加一个7-甲基鸟苷,同 时第一个核苷酸的C'2也是甲基化的,这种m7GpppNm结构被称 为帽子结构(cap sequence)。帽子结构具有促进核蛋白体与 mRNA的结合、加速翻译起始速度的作用,同时可以增强mRNA 的稳定性。
原核生物与真核生物mRNA比较
5.多聚(A)结构
除组蛋白基因外,在真核mRNA的3'末端,有一多聚腺苷酸 (poly A)结构,通常称为多聚A尾。一般由数十个至一百几十 个腺苷酸连接而成。poly A是RNA生成后加上去的。
原核生物3’端没有或只有较短的多聚(A)结构。 多聚(A)是mRNA由细胞核进入细胞质所必需的形式,它大 大提高了mRNA在细胞质中的稳定性。mRNA刚从细胞核进入细 胞质时,其多聚(A)尾巴一般比较长,随着mRNA在细胞质内逗 留时间延长,多聚(A)逐渐变短消失,mRNA进入降解过程。
内含子在真核基因中所占的比例很高,甚至超过99%
原核生物与真核生物mRNA比较
几乎所有mRNA都可以被分成3部分:
1、编码区:从起始密码子AUG开始经一连串编码 氨基酸的密码子直至终止密码子。
2、位于AUG之前的5’端上游非编码区 3、位于终止密码子之后不翻译的3’端下游非编
码区。
原核生物与真核生物mRNA比较 原核生物mRNA转录物与DNA模板链序列结构之间 的对应关系
真核生物mRNA的合 成和加工:细胞核
原核生物与真核生物mRNA比较
1.原核生物mRNA的半衰期短
mRNA的转录和翻译是在同一个细胞空间里同步 进行的,蛋白质合成往往在mRNA刚开始转录时就 被引发了。真核生物中有特殊的机理来增加或减少 特定分子的半衰期。
第八章 真核生物的转录

● 真 核 生 物 转 录 起 始 复 合 物
转录因子 TBP
转录复合体
TAFs
TFIIA TFIIB TFIIF Pol II
TFIIE
RNA pol Ⅱ的转录起始
结构基因
DNA
P
O
Z
Y
A
Z: β-半乳糖苷酶 Y: 透过酶
A:乙酰基转移酶
● 5’ 端无“帽子”结构, 3’ 端没有或只有较短
的poly(A )结构。
SD序列:mRNA中用于结合原核生物核糖体的序列。
2、真核生物mRNA的特征 “基因”的分子生物学定义:产生一条多肽链或 功能RNA所必需的全部核甘酸序列。 ● 5’ 端存在“帽子”结构 ●多数mRNA 3’ 端具有poly(A )尾巴(组蛋白除外) ●以单顺反子的形式存在
原核生物和真核生物mRNA结构的比较
五、RNA合成与DNA合成异同点
相同点:
1、都以DNA链作为模板
2、合成的方向均为5’→3’
3、聚合反应均是通过核苷酸之间形成的3’,5’-磷 酸 二酯键,使核苷酸链延长。
不同点:
模板 原料
酶 产物
复制 两条链均复制 dNTP
DNA聚合酶 子代双链DNA (半保留复制) A-T;G-C RNA引物
增强子的作用方式
2. 通用转录因子
能直接、间接辨认和结合转录上游区段
DNA的蛋白质,统称为反式作用因子(transacting factors) ,现已发现数百种 。 反式作用因子中,直接或间接结合RNA
原核生物mrna的特征

原核生物mrna的特征原核生物的mRNA具有以下特征:1.单一转录本:原核生物的基因组通常是圆形或线性的,基因数量较少。
每个基因一般只有一个转录本(mRNA),不会剪接产生多种不同形式的mRNA。
2. 没有内含子:原核生物的mRNA通常是连续的,没有内含子(introns)。
它们的DNA序列中不存在内含子,因此转录过程中不需要剪接(splicing)。
3. 短寿命:原核生物的mRNA在细胞内的寿命相对较短。
这是因为它们通常不含稳定的3'非翻译区(3' untranslated region, 3' UTR)和5'非翻译区(5' untranslated region, 5' UTR)等稳定性序列,容易被核酸酶降解。
4.具有起始密码子和终止密码子:原核生物的mRNA具有起始密码子(通常为AUG)和终止密码子(如UAA,UAG和UGA)。
起始密码子标志着翻译的起始点,而终止密码子指示翻译的终止点。
5.翻译前没有修饰:原核生物的mRNA在翻译之前一般没有进行修饰。
与真核生物不同,原核生物的mRNA不经历RNA剪接、mRNA甲基化和3'端加帽修饰等过程。
6.能够进行同时转录和翻译:在原核生物中,mRNA的合成和翻译可以同时进行。
由于原核生物的细胞没有明显的亚细胞器,如核和线粒体,mRNA的合成(转录)和翻译(蛋白质合成)可以在细胞质内同时进行。
7.具有短3'UTR:原核生物的mRNA的3'非翻译区相对较短,一般不超过50个碱基。
3'UTR在真核生物中常参与调控转录后的mRNA稳定性和翻译效率,但在原核生物中,这个区域的作用较小。
8. 具有SD序列:原核生物的mRNA在起始密码子的上游通常含有Shine-Dalgarno序列(SD序列)。
SD序列与原核生物中的核糖体结合,帮助确定起始点,促进翻译的起始。
9.基因的排列相对简单:原核生物的基因组相对较小,基因数量有限,并且基因组的排列较为紧凑。
分子生物学复习资料11111111111111111111111111111111

名词解释1. DNA拓扑异构酶:能在闭环DNA 分子中改变两条链的环绕次数的酶,它的作用机制是首先切断DNA,让DNA绕过断裂点以后再封闭形成双螺旋或超螺旋DNA.2. 信号肽:在起始密码子后,有一段编码疏水性氨基酸序列的RNA区域,被称为信号肽序列,它负责把蛋白质引导到细胞内膜结构的亚细胞器内。
3. 启动子:与基因表达启动相关的顺式作用元件,是结构基因的重要成分。
它是一段位于转录起始位点5’端上游区大约100~200bp以内的具有独立功能的DNA序列,能活化RNA聚合酶,使之与模板DNA准确地结合并具有转录起始的特异性。
4. 冈崎片段:是在DNA半不连续复制中产生的长度为1000~2000个碱基的短的DNA片段,能被连接成一条完整的DNA链。
5. 密码的简并性:由一种以上密码子编码同一个氨基酸的现象称为简并,对应于同一氨基酸的密码子称为同义密码子。
6. DNA重组体:根据人们的意愿利用限制性内切酶和DNA连接酶对不同生物的遗传基因进行切割、拼接或者重新组合,形成具有新的遗传性状的DNA 。
7. 信号转导:在细胞通讯系统中,细胞识别与之相接触的细胞,或者识别周围环境中存在的各种化学和物理信号,并将其转变为细胞内各种分子活性的变化,从而改变细胞的某些代谢过程,影响细胞的生长速度,甚至诱导细胞凋亡,这种针对外源信息所发生的细胞应答反应全过程称为信号转导。
8. 摆动假说:Crick为解释反密码子中某些稀有成分的配对以及许多氨基酸有2个以上密码子的问题而提出的假说。
9. C值反常现象:指C值往往与种系的进化复杂性不一致的现象。
10. 半不连续复制:DNA复制过程中前导链的复制时连续的,而另外一条链,即后随链的复制是中断的、不连续的。
11. 基因组:生物有机体的单倍体细胞中所有DNA,包括核中的染色体DNA和线粒体、叶绿体等亚细胞器中的DNA.12. 转录:是指拷贝出一条与DNA链序列完全相同(除了T——》U之外)的RNA单链的过程,是基因表达的核心步骤。
分子生物学3生物信息的传递(上)——从DNA到RNA

分子生物学3生物信息的传递(上)——从DNA到RNA第三章生物信息的传递(上)——从DNA到RNA重点:1.启动子与转录起始2. 原核生物和真核生物mRNA的特征比较3. 内含子的剪接、编辑及化学修饰难点:1.启动子与转录起始2. 终止和抗终止3. 内含子的剪接、编辑及化学修饰第四节启动子与转录起始大肠杆菌RNA聚合酶与启动子的相互作用主要包括启动子区的识别、酶与启动子的结合及因子的结合与解离等。
1. 原核启动子的基本结构(1)启动子:是一段位于结构基因5 ′端上游区的DNA序列,能活化RNA聚合酶,使之与模板DNA准确的相结合并具有转录起始的特异性。
基因的特异性转录取决于酶与启动子能否有效地形成二元复合物,所以,RNA聚合酶如何有效地找到启动子并与之结合是转录起始过程中首先要解决的问题。
我们知道,转录的起始是基因表达的关键阶段,而这一阶段的重要问题是RNA聚合酶与启动子的相互作用。
启动子的结构影响了它与RNA聚合酶的亲和力,从而影响了基因表达水平。
(2)转录单元:是一段从启动子开始至终止子结束的DNA序列。
RNA聚合酶从转录起点开始沿着模板前进,直到终止子为止,转录出一条RNA链。
在细菌中,一个转录单元可以是一个基因,也可以是几个基因(3)转录起点:是指与新生RNA链地一个核苷酸相对应DNA链上的碱基。
常常把起点前面,即5′末端的序列称为上游,而把其后面即3′末端的序列称为下游。
在描述碱基的位置时,一般用数字表示,起点为+1,下游方向依次为+2、+3等,上游方向依次为-1、-2、-3等。
启动子区是RNA聚合酶的结合区,其结构直接影响到转录的效率。
那么,启动子区有什么结构特点呢?(4)绝大部分原核启动子都存在-10区和-35区-10区:在-6~-13bp之间,共同序列为TATAAT,又称pribnow 框,酶在此处与DNA结合成稳定的复合物,在转录方向上解开双链形成开放型起始结构。
-35区:共同序列为TTGACA,是RNA聚合酶起始识别区,这一识别过程与σ因子有关。
原核生物mRNA

原核生物mRNA一般5′端有一段不翻译区,称前导顺序,3′端有一段不翻译区,中间是蛋白质的编码区,一般编码几种蛋白质。真核生物mRNA(细胞质中的)一般由5′端帽子结构、5′端不翻译区、翻译区(编码区)、3′端不翻译区和3′端聚腺苷酸尾巴构成分子中除m7G构成帽子外,常含有其他修饰核苷酸,如m6A等。真核生物mRNA通常都有相应的前体。从DNA转录产生的原始转录产物可称作 原始前体(或mRNA前体)。一般认为原始前体要经过hnRNA核不均-RNA的阶段,最终才被加工为成熟的mRNA。
1956年Elliot Volkin和 Lawrence Astrachan作了一项很有意思的观察:当E.coli被T2感染,迅速停止了RNA的合成,但噬菌的RNA却开始迅速合成。用同位素脉冲一追踪标记表明噬菌的RNA在很短的时间内就进行合成,但很快又消失了,表明RNA的半衰期是很短的。由于这种新合成的RNA的碱基比和T2的DNA碱基比相似,而和细菌的碱基比不同,所以可以确定新合成的RNA是T2的RNA。由于T2感染细菌时注入的是DNA,而在细胞里合成的是RNA,可见DNA是合成RNA的模板。最令人信服的证据来自DNA-RNA的杂交实验。Hall.B.D和Spiegeman,S,将T2噬菌体感染E.coli后立即产生的RNA分离出来,分别与T2和E.coli的DNA进行分子杂交,结果发现这种RNA只能和T2的DNA杂交形成“杂种”链,而不能和E.coli的DNA进行杂交。表明T2产生的这种RNA(即mRNA)至少和T2的DNA中的一条链是互补的。
[编辑本段]信使RNA的合成与加工
第一节 DNA转录生成RNA
一、定义
(一)转录单位
(二)启动子(promoter)
第三章 转录(2)

剪接的基本过程: 剪接的基本过程:
以碱基互补的方式识别 前体5 ①由Ul snRNA以碱基互补的方式识别 以碱基互补的方式识别mRNA前体 ' 前体 剪接点; 剪接点;
LacY LacA 操纵基因 LacZ 启动子 (P) ) 调节基因( ) 调节基因(R)TGTTGACATTT…ATTTGTTATAATG CAT… 半乳糖苷酶) 半乳糖苷通透酶)半乳糖乙酰酶) (β半乳糖苷酶) β半乳糖苷通透酶)半乳糖乙酰酶) ( (β … … … … 1040bp (O) ) 3510bp 780bp 825bp 6~9bp 4~7bp
(三)内含子的剪接、编辑及化学修饰 内含子的剪接、
真核基因大多是断裂的,即一个基因可由多 真核基因大多是断裂的, 断裂的 个内含子和外显子间隔排列而成。 个内含子和外显子间隔排列而成。内含子在真 核基因中所占的比例很高,可超过99%。 核基因中所占的比例很高,可超过 。
1、RNA中的内含子 、 中的内含子 真核基因表达往往伴随着RNA的剪 真核基因表达往往伴随着 的剪 前体分子中切除被 接过程,从 前体 接过程 从mRNA前体分子中切除被 称为内含子 内含子(intron)的非编码序列, 的非编码序列, 称为内含子 的非编码序列 并使基因中被称为外显子 并使基因中被称为外显子(exon)的编 的编 外显子 码序列拼接形成成熟mRNA。 码序列拼接形成成熟 。
CPSF
CstF
poly(A)尾的功能 尾的功能
poly(A)可能是 可能是mRNA由细胞核进入细胞质 可能是 由细胞核进入细胞质 所必需的形式。 所必需的形式。 提高了mRNA在细胞质中的稳定性。当 在细胞质中的稳定性。 提高了 在细胞质中的稳定性 mRNA刚从细胞核细进入胞质时,其 刚从细胞核细进入胞质时, 刚从细胞核细进入胞质时 poly(A)较长,随着 较长, 较长 随着mRNA在细胞质内逗留 在细胞质内逗留 时间延长, 逐渐变短消失, 时间延长,poly(A)逐渐变短消失,mRNA 逐渐变短消失 开始降解。 开始降解。
原核生物真核生物基因表达比较

基因表达: 基因表达(gene expression)是指细胞在生命过 程中,把储存在DNA顺序中遗传信息经过转录和翻 译,转变成具有生物活性的蛋白质分子。
122210101436熊雪薇
.
1
转录: 原核生物
真核生物
原料: 模板: 酶: 其他因子:
NTP (ATP UTP CTP GTP)
14
蛋白质生物合成需要酶类、蛋白质因子:
15
16
氨基酸的活化:
氨基酸与特异的tRNA结合形成氨基酰-tRNA的过程称为 氨基酸的活化,氨基酸活化形成氨基酰-tRNA。
原核、真核生物----都有两种Met-tRNA:
原核生物起始氨基酰-tRNA:
fMet-tRNAfMet
真核生物起始氨基酰-tRNA:
原核细胞中数个结构基因常串联为一个转录单位转录生成的mrna可编码几种功能相关的蛋白质为多顺反子polycistron真核mrna只编码一种蛋白质为单顺反子singlecistron许多真核生物基因转录后有一个对mrna外显子加工的过程使mrna序列中出现移码错义无义突变导致同一前体mrna翻译出序列功能不同的蛋白质
6
7
转录起始:
原核生物:RNA聚合酶和DNA
的特殊序列——启动子 (promoter)结合后,就能启动 RNA合成。
RNA聚合酶全酶( 2 )与模 板结合,形成闭合转录复合体。
DNA双链局部解开,形成开放转 录复合体。
真核生物:转录起始需要启动
子 、RNA聚合酶和转录因子的 参与。
少数几个反式作用因子的搭配启 动特定基因的转录
DNA ?
RNA-聚合酶
RNA-聚合酶 I II III
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
原核与真核生物mRNA的特征比较原核生物中:•mRNA的转录和翻译发生在同一个细胞空间,•这两个过程几乎是同步进行的。
真核细胞中:真核细胞mRNA的合成和功能表达发生在不同的空间和时间范畴内。
•mRNA以较大分子量的前体RNA出现在核内,•只有成熟的、相对分子质量明显变小并经化学修饰的mRNA才能进入细胞质,参与蛋白质的合成。
mRNA的组成:•编码区(coding region):从起始密码子AUG开始经一连串编码氨基酸的密码子直至终止密码子。
•5’端上游非编码区(5’UTR):位于AUG之前不翻译的区域。
•3’端下游非编码区(3’UTR):位于终止密码子之后不翻译的区域。
原核生物mRNA的特征•半衰期短。
•许多原核生物mRNA以多顺反子的形式存在。
•原核生物mRNA的5’端无帽子结构,3’端没有或只有较短的多聚(A)结构。
原核生物mRNA的特征1.半衰期短•原核生物中,mRNA的转录和翻译是在同一个细胞空间里同步进行的,蛋白质合成往往在mRNA刚开始转录时就被引发了。
•大多数细菌mRNA在转录开始1分钟后就开始降解。
mRNA降解的速度大概只有转录或翻译速度的一半。
原核生物mRNA的特征2. 许多以多顺反子的形式存在:原核细胞的mRNA(包括病毒)有时可以同时编码几个多肽。
Prokaryotic mRNA (polycistrionic)单顺反子mRNA (monocistronic mRNA):只编码一个蛋白质的mRNA。
多顺反子mRNA(polycistronic mRNA):编码多个蛋白质的mRNA。
原核生物mRNA的特征3. 原核生物mRNA的5’端无帽子结构,3’端没有或只有较短的多聚(A)结构。
原核生物起始密码子AUG上游有一被称为Ribosome Binding Site (RBS)或SD序列(Shine –Dalgarno sequence)的保守区,因为该序列与16S-rRNA 3’端反向互补,所以被认为在核糖体-mRNA的结合过程中起作用。
原核生物mRNA的特征4. 原核生物常以AUG(有时GUG,甚至UUG)作为起始密码子;真核生物几乎永远以AUG作为起始密码子。
真核生物mRNA的特征•单顺反子形式存在。
•5’端存在“帽子”结构。
•绝大多数具有多聚(A)尾巴。
真核生物mRNA的结构模式Eukaryotic mRNA (monocistrionic)“基因”的分子生物学定义:产生一条多肽链或功能RNA所必需的全部核苷酸序列!A gene can be defined as following: The entire nucleic acid sequence that is necessary for the synthesis of a functional polypeptide or RNA molecule.真核生物mRNA的特征1.真核生物mRNA的5’端存在“帽子”结构。
真核生物基因转录一般从嘌呤起始,其5’端大都经过修饰。
5’Capping•通过5′→5′磷酸二酯键在原初mRNA的5’端倒扣一个“G”。
•包括三个连续的酶促反应。
•5′末端加上鸟苷是由鸟苷转移酶催化的。
•帽子结构是GTP和原5’三磷酸腺苷(或鸟苷)缩合反应的产物。
•mRNA的帽子结构常常被甲基化,是蛋白质合成起始信号的一部分。
•加帽反应发生的很早。
真核生物mRNA的“帽子”结构鸟苷酸-7甲基转移酶2’-O-甲基转移酶mRNA的帽子结构常常被甲基化零类帽子(cap0):第一个甲基出现在所有真核细胞的mRNA中(单细胞真核生物mRNA主要是这个结构),由鸟苷酸-7甲基转移酶催化,称为零类帽子。
1类帽子(cap1):如在第二个核苷酸(原mRNA 5’第一位)的2’-OH位上加另一个甲基,这步反应由2’-O-甲基转移酶完成。
一般把有这两个甲基的结构称为1类帽子。
真核生物中以这类帽子结构为主。
2类帽子(cap2): 在某些生物细胞内,mRNA链上的第三个核苷酸的2’-OH位也可能被甲基化,因为这个反应只以带有1类帽子的mRNA为底物,所以被称为2类帽子。
只占有帽mRNA总量的10%-15%以下。
帽子结构的功能(1)有助于mRNA越过核膜,进入胞质;(2)保护5′不被核酶降解;(3)翻译时供IFⅢ(起始因子)和核糖体识别,是翻译所必需的。
真核生物mRNA的特征2. 绝大多数真核生物mRNA具有多聚(A)尾巴。
•除组蛋白基因外,真核生物mRNA的3’末端都有多聚(A)序列,其长度因mRNA种类不同而变化,一般为40-200个左右。
•由多聚(A)聚合酶催化的;•它是在转录后加上的;•Poly(A)被特异的蛋白质PABP结合。
•在高等生物中(酵母除外)在poly(A)上游11-30nt 处有一特殊序列AAUAAA ,这一序列是高度保守的。
•对于初级转录产物的准确切割及加多聚(A)是必需的。
1. CPSF (cleavage and polyadenylation specificity factor) & CstF (cleavage stimulation factor) bind to the poly-A signal, leading to the RNA cleavage.2. Poly-A polymerase (PAP ) adds ~ 200 As at the 3’end of the RNA, using ATP as a substrate.Polyadenylation and termination多聚腺苷化(polyadenylation)反应要经过2个阶段(1)首先将一个短的寡聚A序列(10nt)加到3’端,此反应绝对依赖于AAUAAA序列,这是由poly(A)聚合酶在特殊因子指导下完成的。
(2)寡聚A尾巴延伸到240nt的长度。
此反应并不需要AAUAAA序列,但需要一个识别寡聚A并指导poly(A)聚合酶延伸的刺激因子。
多聚(A)的功能•是mRNA由细胞核进入细胞质所必需的形式;•它大大提高了mRNA在细胞质中的稳定性。
mRNA刚从细胞核进入细胞质时,其多聚(A)尾巴一般比较长,随着mRNA在细胞质内逗留时间延长,多聚(A)逐渐变短消失,mRNA进入降解过程。
•它可促进核糖体的有效循环。
Poly-A in the 3’end promotes the efficient recycling of ribosomesPoly (A) -•尽管大部分真核mRNA有poly (A)尾巴,细胞中仍有多大1/3没有poly (A)的mRNA,将其称为Poly (A) –•约1/3的Poly (A) –mRNA编码了不同形式的组蛋白。
Poly(A)+ RNA can be separated from other RNAs by fractionation on Sepharose-oligo(dT)在动物细胞中,mRNA的表达需转录、修饰、加工、核质转运和翻译。
内含子的剪接、编辑及化学修饰RNA 中的内含子非编码区编码区真核基因大多是断裂的:•一个基因可由多个内含子和外显子间隔排列而成。
•内含子在真核基因中所占的比例很高,甚至超过99%。
Exons (外显子): the coding sequencesIntrons (内含子) : the intervening sequences部分人类基因中内含子序列所占的比重分析基因长度(kb)内含子数量内含子所占比重(%)胰岛素 1.4267ß-球蛋白 1.4269血清蛋白181389胶原蛋白组分VII3111771 VIII因子1862595萎缩性肌强直因子240078>99剪接(Splicing)RNA splicing: removal of introns and joining of exons.•It takes place in the nucleus before the mature mRNA can be exported to the cytoplasm.•Catalyzed by spliceosome(RNA +protein)mRNA的前体(核不均一RNA,hnRNA)5’加“帽”和3’酶切加多聚腺苷酸RNA的剪接连续的可译框(ORF) (open reading frame)通过核孔进入细胞质蛋白质合成的模板RNA splicing可通过体外凝胶电泳进行分析RNA加工过程及其生理功能加工过程推测的生理功能加帽子反应mRNA从细胞核向细胞质运转,翻译起始加多聚A反应转录终止,翻译起始和mRNA降解RNA的剪接从mRNA, tRNA和rRNA分子中切除内含子RNA的切割从前体RNA中释放成熟tRNA和rRNA分子•真核基因平均含有8-10个内含子,前体分子一般比成熟mRNA大4-10倍。
•不同生物细胞内含子的边界存在相似的核苷酸序列。
•内含子的“功能”及其在生物进化中的地位是一个引人注目的问题。
内含子剪接异常引起疾病例如:地中海贫血病人的珠蛋白基因中,大约有1/4的核苷酸突变发生在内含子的5’或3’边界保守序列上,或者直接干扰了前体mRNA的正常剪接。
生物体内的各种内含子内含子类型细胞内定位GU-AG细胞核,前体mRNA (真核)AU-AC细胞核,前体mRNA(真核)I类内含子细胞核,前体rRNA(真核),细胞器RNA,少数细菌RNA II类内含子细胞器RNA,部分细菌RNAIII类内含子细胞器RNA双内含子细胞器RNA tRNA前体中的内含子细胞核,tRNA前体(真核)GU-AG或AU-AC分别代表了不同内含子的5’和3’边界序列。
断裂基因发现不久,Crick(1978年)就提出了一系列发人深思的问题:(1)剪切作用若是通过酶来进行的,那么这种剪接酶是怎样识别RNA上特定的位点?(2)剪切酶有没有特异性?对不同的RNA是否需要不同的酶?(3)切除的RNA是以线状还是环状存在的?切下的RNA的命运将如何?Chambon等分析比较了大量结构基因的内含子切割位点,发现有2个特点:(1)内含子的两个末端并不存在同源或互补。
(2)连接点具有很短的保守序列,亦称边界序列。
100种内含子的5′端都是GU;3′端都是AG,因此称为GU-AG法则(GU-AGrule),又称为Chambon法则。
A脊椎动物前体mRNA中常见内含子剪接所必需的保守序列两个剪接位点的序列是不同的:•左边的剪接位点称供体(donor)位点,•右边的剪接位点称受体(acceptor)位点。
GU-AG法则(GU-AG rule)不适用于线粒体、叶绿体的内含子,也不适用于酵母的tRNA基因。