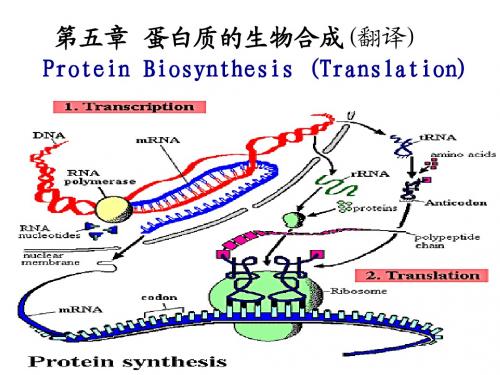
第五章protein的翻译
第3章 DNA复制(1)

Ed., 2005 吴乃虎. 基因工程原理(上、下). 1999,2001
关于学习方法
理解和掌握分子生物学基本理论
指导分子生物学实践
准确理解和运用分子生物学的基本概念 分子生物学的基本理论或结论:
知其然,知其所以然
启发! 创造性思维!!!
Central Dogma of Live
DNA
3.1.1 复制的基本过程 3.1.2 模板 3.1.3 半保留复制 3.1.4 复制的起点与方向 3.1.5 半不连续复制 3.1.6 复制的引物
3.1.7 线性DNA末端的复制
DNA复制:亲代双链DNA分子在DNA聚合酶的作
用下,分别以每条单链 DNA分子为模板,聚合与自 身碱基可以互补配对的游离的dNTP,合成出两条与 亲代DNA分子完全相同的子代DNA分子的过程
tumor antigen genes: SV40 large T Ag 端粒逆转录酶(TERT)基因 ?
Why/How to study MB?
Practical/theory questions?
4. Gene expression & its regulation
1) Developmentally/Tissue-specifically ?
Innovation
Creation
Carve-out
第3章 DNA复制
DNA Replication
遗传物质的分子机制
分子生物学的核心
Watson & Crick: 一种遗传物质必
须能行使2种功能:
自我复制 对细胞的高度特异性的影响 遗传物质的基本属性:
基因的自我复制
[分享]分子生物学名词解释1
![[分享]分子生物学名词解释1](https://img.taocdn.com/s3/m/9f30ee3359fb770bf78a6529647d27284b7337c4.png)
分子生物学名词解释第二章(主要的:核小体、半保留复制、复制子、单链结合蛋白、岗崎片段、错配修复、DNA的转座、C值矛盾、前导链与后随链。
)1. C值反常现象(C值矛盾C-value paradox):C值是一种生物的单倍体基因组DNA的总量。
真核细胞基因组的最大特点是它含有大量的重复序列,而且功能DNA序列大多被不编码蛋白质的非功能DNA所隔开,这就是著名的“C值反常现象”。
C值一般随着生物进化而增加,高等生物的C值一般大于低等生物。
某些两栖动物的C值甚至比哺乳动物还大,而在两栖动物里面,C值变化也很大。
2.DNA的半保留复制:由亲代DNA生成子代DNA时,每个新形成的子代DNA中,一条链来自亲代DNA,而另一条链则是新合成的,这种复制方式称半保留复制。
3.DNA聚合酶:●以DNA为模板的DNA合成酶●以四种脱氧核苷酸三磷酸为底物●反应需要有模板的指导●反应需要有3 -OH存在●DNA链的合成方向为5 34.DNA连接酶(1967年发现):若双链DNA中一条链有切口,一端是3’-OH,另一端是5‘-磷酸基,连接酶可催化这两端形成磷酸二酯键,而使切口连接。
但是它不能将两条游离的DNA单链连接起来DNA连接酶在DNA复制、损伤修复、重组等过程中起重要作用5.DNA 拓扑异构酶(DNA Topisomerase):拓扑异构酶І:使DNA一条链发生断裂和再连接,作用是松解负超螺旋。
主要集中在活性转录区,同转录有关。
例:大肠杆菌中的ε蛋白拓扑异构酶Π:该酶能暂时性地切断和重新连接双链DNA,作用是将负超螺旋引入DNA分子。
同复制有关。
例:大肠杆菌中的DNA旋转酶6. DNA 解螺旋酶/解链酶(DNA helicase)通过水解ATP获得能量来解开双链DNA。
E.coli中的rep蛋白就是解螺旋酶,还有解螺旋酶I、II、III。
rep蛋白沿3 ’ 5’移动,而解螺旋酶I、II、III沿5 ’ 3’移动。
7. 单链结合蛋白(SSBP-single-strand binding protein):稳定已被解开的DNA单链,阻止复性和保护单链不被核酸酶降解。
分子生物学第5章蛋白质翻译

---aa 接受臂 ;
loading aa at 3‟ end ---TΨ C loop; contact with 5s rRNA
---DHU loop;
contact with AARS ---anti-codon loop;
34
---extra loop;
分类标准
tRNACys表示转运半胱氨酸的tRNA Cys-tRNACys 表 示 携 带 了 半 胱 氨 酸 的 tRNACys
●
Paracodon ---由若干Nt组成,存在于tRNA不定位置上 ---与AARS侧链基团的分子发生特异的“契合” ---成为tRNA准确负载氨基酸的机制之一
●
tRNA的“L”三维结构与功能 “L”构型的结构力 ---二级结构中双链区的碱基堆积力和氢键 ---二级结构中非双链区在“L”结构中,形成氢键结合
成为核糖体识别第一个 AUG的信号
细菌 16S
AMEAMECCUGCGGUUGGAUGACCUCCUU
哺乳动物 18S rRNA AMEAMECCUGCGGAAGGAUGAUUA
高度相似
5.2.3. mRNA
mRNA分子5‟端的序列对于起始密码子选择 有重要作用
● In Prokaryote
poly-cistron
a) 简并现象的概念; ---一种氨基酸受2个或2个以上codon编码的遗传现象 ---编码一种aa的4个codon间, 仅3rd Nt 不同, 称为codon family 例;Ser (6 codons) 1 codon family & 2 extra codons
密码子的简并性
b) 简并现象的机理;
5.3. Genetic Code
第五章 基因表达的调节1(regulation

b. 调节蛋白识别结构模序中常含Asn,Gln, Lys或Arg,可与碱基形成氢键;
2. 调节蛋白与DNA结合motif的结构类型 a. 螺旋-转角-螺旋(Helix-Turn-Helix) 这种结构含两个α-螺旋(7-9个氨基酸) 由β-转角(20个氨基酸)连系,其中一 个螺旋为识别螺旋,通常含有许多与 DNA接触的氨基酸残基,位于大沟之中; b. 锌指(Zinc Finger) 真核生物调节蛋白常含有许多由30个氨
噬菌体λ 九. 噬菌体λ的发育调控 λ 的转录调控用级联(cascade)调控方 法,为了有序地组装噬菌体颗粒,把基 因分组分阶段表达; λ 决定溶源化或溶菌命运的调控,为了 解真核生物的发育调控提供了一种模式。
1. 溶菌(Lysis)和溶源化(Lysogeny) λ 进入细菌细胞后的两种命运: 溶菌周期(lytic cycle):新一代噬菌体 颗粒产生并导致细胞溶解; 溶源化:噬菌体DNA在寄主染色体上整 合,并随寄主DNA复制传代的过程。
第五章 基因表达的调节 (Gene regulation) regulation)
基因表达(gene expression):基因被转 录产生RNA,编码着蛋白质结构信息的 mRNA被翻译成蛋白质,总之,基因的 信息被转录,翻译成终产物的过程称为 基因表达。 一. 引论 a. 基因的多样性及表达的差异性 细胞在任何给定的时间内都表达的基因 只有一小部分;
肽链延长因子,细菌细胞中大量存在的蛋白 质(基因产物)之一; DNA损伤修复酶,每个细胞只有几个拷贝; 代谢途径的酶,因食物来源不同而变化数量; 一些影响细胞分化的蛋白只存在极短时间。 b. 基因表达的调节是细胞代谢平衡及维持发育 期间不同细胞的结构和功能差异的关键; c. 蛋白质(或RNA)在细胞内浓度可以在六个 水平上被调节;
蛋白组学翻译后修饰

先用糖苷内切酶消化,再用蛋白酶消化,通过 分析糖苷酶作用前后MS发生位移的肽段,即 可确定含糖基化的肽段 结合串联质谱,可进一步分析糖基化肽段的氨 基酸序列,从而发现糖基化位点
核糖核酸酶B中糖基化位点的分析
糖苷键酶F作用前
糖苷键酶F作用后 剩一个GlcNAc
氨基酸序列的测定
m/z=4792.23
翻译后化学修饰的生物学效应
泛素化对于细胞分化与凋亡、DNA 修复、免疫应 答和应激反应等生理过程起着重要作用; 磷酸化涉及细胞信号转导、神经活动、肌肉收缩 以及细胞的增殖、发育和分化等生理病理过程; 糖基化在许多生物过程中如免疫保护、病毒的复 制、细胞生长、炎症的产生等起着重要的作用; 脂基化对于生物体内的信号转导过程起着非常关 键的作用; 组蛋白上的甲基化和乙酰化与转录调节有关。
TSAASSSNYCNQM
RQHMDSSTSAASSSNYCNQ MMKSRNLTKDRCKPVNTF VHE
糖基化类型的分析
糖蛋白进行蛋白酶切,得到含糖肽段,对肽段直接 进行ESI-MS/MS,以及MALDI-TOF-MS的PSD, 可以直接得到单糖碎片,从而确定糖链结构。
凝集素在糖蛋白研究中的作用
真核生物的Ser,Thr,Tyr 残基. 原核生物的His,Asp,Glu
蛋白质组学在磷酸化分析中的困难
磷酸化蛋白质在细胞内的蛋白质中是相对较 低丰度的; 即使我们找到一种磷酸化蛋白质,也不能排 除有该蛋白质的其他磷酸化形式存在; 细胞内有很多磷酸酯酶,在样品处理时,这 些酶很容易将磷酸基团脱掉; 磷酸化蛋白质酶解后的磷酸化肽段,因为其 化学性质的负电性,在质谱技术中面临着难 以质子化的困难。
5第五章 植物的光合作用复习题参考答案

第五章植物的光合作用复习题参考答案一、名词解释1、光反应( light reaction)与暗反应(dark reaction ):光合作用中需要光的反应过程,是一系列光化学反应过程,包括水的光解、电子传递及同化力的形成;暗反应是指光合作用中不需要光的反应过程,是一系列酶促反应过程,包括CO2的固定、还原及碳水化合物的形成。
2、C3途径(C3pathway )与C4途径(C4pathway ):以RUBP为CO2受体、CO2固定后的最初产物为PGA的光合途径为C3途径;以PEP为CO2受体、CO2固定后的最初产物为四碳双羧酸的光合途径为C4途径。
3、光系统(photosystem, PS ):由不同的中心色素和一些天线色素、电子供体和电子受体组成的蛋白色素复合体,其中PSI的中心色素为叶绿素a P700,PSII的中心色素为叶绿素a P680.4、反应中心( reaction center):由中心色素、原初电子供体及原初电子受体组成的具有电荷分离功能的色素蛋白复合体结构。
5、光合午休现象(midday depression ):光合作用在中午时下降的现象。
6、原初反应(primary reaction ):包括光能的吸收、传递以及光能向电能的转变,即由光所引起的氧化还原过程。
7、磷光现象(phosphorescence phenomenon ):当去掉光源后,叶绿素溶液还能继续辐射出极微弱的红光,它是由三线态回到基态时所产生的光。
这种发光现象称为磷光现象。
8、荧光现象(fluorescence phenomenon ):叶绿素溶液在透射光下呈绿色,在反射光下呈红色,这种现象称为荧光现象。
9、红降现象(red drop ):当光波大于685nm时,虽然仍被叶绿素大量吸收,但量子效率急剧下降,这种现象被称为红降现象。
10、量子效率(quantum efficiency ):又称量子产额或光合效率。
第五章 蛋白质的翻译2

翻译后易位
post-translational translocation
◆蛋白质翻译完成后从核糖体上释放,然合
扩散至合适靶膜并与易位装置结合。
◆蛋白质合成时,其核糖体不与任何细胞器
相连,称游离核糖体(free ribosome)
细胞器官
线粒体 叶绿体 细胞核 过氧化物酶体
信号位置
类型
长度
蛋白质翻译完成后从核糖体上释放,然 合扩散至合适靶膜并与易位装置结合。
① ② ③
④
⑤
An N-terminal signal sequence is hydrophobic
牛生长素的信号肽序列 信号肽的作用是引导蛋白质的跨膜
2.2.3 信号肽与蛋白质转运的关系
(1)完整信号肽是保证蛋白质转运的必要条件; (2)仅有信号肽不足以保证蛋白质转运发生; (3)信号序列切除并不是转运所必需; (4)并非所有运转蛋白质都有可降解的信号肽。
(2)穿越过氧化物酶体膜
(peroxisome)
膜上也有类似的装置,但底物蛋白质并不直
接与其结合。需载体蛋白协助穿过膜。
蛋白质在细胞质中通过 与载体蛋白结合一起穿 越过氧化物酶体膜通道, 穿过之后释放载体蛋白。
(3)穿越细胞核膜
向核内转运需更大更复杂的装置,即核 孔(nuclear pore )。由载体蛋白与底
The signal sequence initiates membrance entry
2.3 翻译后易位
2.3.1 前导肽及其特性
2.3.2 蛋白质穿越不同器膜的过程
2.3.1 前导肽及其特性
翻译后跨膜易位的蛋白质,前体一般 含前导肽(leader peptide),前导肽在跨 膜运转中起重要作用。 过膜后,前导肽水解,蛋白质变为有 功能的蛋白质。
5蛋白质的翻译

proteins,r-proteins)组成,rRNA 组成总分子量的 60%~65%。核糖体的相对大小常常用 沉降系数单位来表示。大肠杆菌的核糖体称为 70S 核糖体,其中的小亚基称为 30S 核糖体, 大亚基称为 50S 核糖体(图 5-8) 。小亚基包含 21 种不同的蛋白质(被称为 S1 一 S21)和 16SrRNA。大亚基由 33 种蛋白质(被命名为 L1~L33)和 23S 及 5SrRNAs 组成(图 5-9)。真核 核糖体称为 80S 核糖体,其中 40S 小亚基包含 33 种蛋白质和 18SrRNA,而 60S 大亚基包含 50 种蛋白质(图 5-9)和 3 种 rRNA (28S, 5.8S 和 5S) 。 真核 5.8SrRNA 与细菌 23SrRNA 的 5SrRNAs 部分同源(表 5.1) 。古细菌核糖体类似于细菌核糖体,但有些包含与真核相同的特别亚基。
组织上,原核生物与真核生物有重要的差别(图 5.1、图 5.2 和图 5.3) 。原核生物的 mRNA 的第一个密码子 AUG 上游的一个重要特征就是 Shine-Dalgarno 序列,而真核生物 mRNA 除
第一个密码子 AUG 的上游是核糖体小亚基扫描 AUG 的信号序列(CCACC)外, 5’端非翻译区 上游为帽子结构, 3’端非翻译区内有多聚腺苷化的信号 AAUAAA 以及其下游的多聚 A 尾巴。 mRNA 是由 DNA 的模板链转录而来, 其序列与编码链相同与模板链互补。 mRNA 的 5’ →3 ’ 三联体密码子序列与蛋白质 N 端到 C 端的氨基酸序列线形相关。原核生物 mRNA 的转录和翻 译发生在时间与空间上具有相对的同一性,其 mRNA 通常不稳定,在合成后的几分钟内翻译 成蛋白质。 真核 mRNA 的合成与成熟都在核内, 成熟的 mRNA 被运往胞质, 作为模板翻译蛋白 质 , 其 稳 定 性 相 对 较 高 , 达 几 小 时 。
生物化学(人卫版)名词解释整理

生物化学名词解释整理版章节根据人民卫生出版社第7版《生物化学》划分参考教学PPT、各教辅术后习题及试卷标准答案整理录入/清水秋香第一章蛋白质结构与功能isoelectric point, pI:氨基酸的等电点:在某一pH的溶液中,氨基酸解离成阳离子和阴离子的趋势及程度相等,成为兼性离子,呈电中性。
此时溶液的pH值称为该氨基酸的等电点。
Peptide bond:肽键。
由一个氨基酸的α-羧基与另一个氨基酸的α-氨基脱水缩合而形成的酰胺键。
Glutathione,GSH:谷胱甘肽。
由谷氨酸,半胱氨酸,甘氨酸组成的三肽。
是体内重要的还原剂。
peptide unit:肽单元。
参与肽键的6个原子Cα1、C、O、N、H、Cα2位于同一平面,Cα1和Cα2在平面上所处的位置为反式(trans)构型,此同一平面上的6个原子构成了所谓的肽单元。
peptide plane:肽平面。
肽链主链的肽键C-N具有部分双键的性质,因而不能自由的旋转,使连接在肽键上的六个原子共处于一个平面上,此平面称为肽单位平面,又称酰胺平面。
通常是反式的。
α-helix:α-螺旋。
常见的蛋白质二级结构之一。
为具有最大氢键联系的右手螺旋,侧链伸向螺旋外侧,每3.6个氨基酸残基螺旋上升一圈,螺距0.54nm。
motif:模体。
在一个或几个蛋白质中出现的2个或2个以上二级结构元件的不同折叠形式,又称折叠或超二级结构。
也是在DNA中对特殊序列的描述。
zinc finger:锌指结构。
一种常见的模体。
由1个α-螺旋和2个反平行的β-折叠共3个肽段组成,形似手指, 能够结合锌离子,锌指具有结合DNA的功能。
Domain:结构域:大分子蛋白质的三级结构常可分割成一个或数个球状或纤维状的区域,折叠得较为紧密,各行使其功能,称为结构域(domain) 。
Molecular chaperon:分子伴侣。
是细胞内一类可识别肽链的非天然构象、促进各功能域和整体蛋白质正确折叠的保守蛋白质。
医学细胞生物学英译汉总结

第一章 绪论cellcell biology cell theory central dogma cytomics cytoplasmelectron microscope ,EM epigenetics gene细胞细胞生物学 细胞学说 中心法则 细胞组学 细胞质 电子显微镜 表观遗传 基因genomicshuman genome project,HGP medical cell biology model animal noncoding RNA proteomics stem cell biology translational medicine基因组学人类基因组计划医学细胞生物学 模式动物 非编码RNA 蛋白质组学 干细胞生物学 转化医学第二章细胞的概念和分子基础anticodon archaea archaebacteria bacteria cell membrane chlamydia codon cytoplasm cytosol deoxyribonucleic acid,DNAenzyme eukarya eukaryotic cell glycolipids glycoproteins message RNA , mRNAmicroRNA, miRNA mycoplasma反密码子 古菌域 古细菌 细菌 细胞膜 衣原体 密码子 细胞质 细胞质溶胶 脱氧核糖核酸 酶 真核域 真核细胞 糖脂 糖蛋白 信使核糖核酸 微小RNA 支原体 nucleoid nucleus peptide plasmid prion prokaryotic cell protoplasm ribonucleic acid ,RNA ribosomeribosome RNA , rRNA ribozyme RNA silencingsmall nuclear RNA, snRNA structural domains transfer RNA, tRNA viroid virus 拟核细胞核 肽 质粒 朊病毒 原核细胞 原生质 核糖核酸 核糖体 核糖体RNA 核酶RNA 沉默 小核RNA 结构域 转运RNA 类病毒 病毒第三章细胞生物学研究方法Abbe limitagaroseatomic force microscope,AFM autoradiographybiochipcell culturecell free systemcell linecell strainChIPAbbe 限度 琼脂糖 原子力显微镜放射性自显影技术 生物芯片 细胞培养 非细胞体系 细胞系 细胞株 染色质免疫沉淀技术 CLIPcolumn chromatography cytochemistry technique differential centrifugation diffraction pattern DNA denaturation dNTPelectrospray ionization mass spectrometry,ESI-MS embedding紫外交联免疫沉淀技术 柱层析细胞化学技术 差速离心 X-射线衍射图 DNA 变性4种脱氧核苷酸 电喷雾电离质谱 包埋emission lightemzyme cytochemistryequilibrium sedimentation excitation lightfixationflow cytometerfluorescence microscope fluorescence resonanceemergy transfer,FRETfreeze-etchfreeze-fracturegene chipgreen fluorescentprotein,GFPhigh performance liquidchromatography,HPLCimmunocytochemistry,ICC immunomagnetic microsphere immuno-precipitation,IPin situ hybridization,ISHisoelectric focusinglaser capturemicrodissection,LCMmatrix-assisted laserdesorptionionization/time-of-flight , MALDI-TOF-MSmetal shadowingNorthern blotnuclear magnetic resonance spectroscopy,NMR over-expression phage display发射光 酶细胞化学技术 平衡沉降激发光 固定 流式细胞仪 荧光显微镜荧光共振能量转移 冰冻蚀刻 冰冻断裂 基因芯片 绿色荧光蛋白 高效液相层析 免疫细胞化学技术 免疫磁珠免疫沉淀 原位杂交技术 等点聚焦 激光俘获显微切割电泳 基质辅助激光解吸/电离飞行时间质谱金属投影法 Northern 印迹杂交 核磁共振光谱过表达 噬菌体展示polymerase chainreaction,PCR primary cultureprobeprotein chipproteomeproteomicsquantitative PCR ,qPCR radioisotoperenaturationresolutionrestriction nucleasescanning electronmicroscope,SEM scanning tunnelingmicroscope,STM SDS polyacrylamid gel electrophoresissecondary culturesectionsingle molecular fluorescence imaging southern blot stainingtandem massspectrometry,MS/MStransfectiontransmission electron microscope,TEM velocity sedimentation yeast two-hybridzation phase contrast microscope聚合酶链式反应原代培养 探针 蛋白质芯片 蛋白质组 蛋白质组学 荧光实时定量PCR 反应放射性同位素 复性 分辨率 限制性内切酶 扫描电子显微镜扫描隧道显微镜SDS-聚丙烯酰胺凝胶电泳 传代培养 切片 单分子荧光成像 Southern 印迹杂交染色 串联质谱 转染透射电子显微镜速度沉降 酵母双杂交 相差显微镜第四章细胞膜与物质的穿膜运输active transport adaptinantiportaquaporin ,AQPbilayerbiomembranecarrier proteincell coatcell membrane主动运输衔接蛋白 反向运输 双分子层 水孔蛋白 生物膜 载体蛋白 细胞外被 细胞膜 channel proteincholesterol clathrin coated pits coated vesicle constitutive secretion endocytosis exocytosis extrinsic protein通道蛋白 网格蛋白 胆固醇 有被小窝 有被小泡 连续性分泌 胞吞作用 胞吐作用 膜外在蛋白facilitated diffusion fluid mosaic model glycolipid intrinsic proteinligand-gated channel lipid raftslipid anchored protein lipid-linked protein lamella structure model membrane lipid membrane proteinmembrane transport protein micellepassive diffusion passive transport 易化扩散 流动镶嵌模型 糖脂膜内在蛋白 配体门控通道 脂筏脂锚定蛋白 脂连接蛋白 片层结构模型 膜脂 膜蛋白 膜运输蛋白 球状分子团 被动扩散 被动运输 peripheral protein phagocytosis phospholipid pinocytosis plasma membrane regulated secretion simple diffusionstress-activated channel symporttransmembrane protein unit membraneunit membrane model vesicular transport voltage-gated channel外周蛋白 吞噬作用 磷脂 胞饮作用 质膜 受调分泌 简单扩散 应力激活通道 同向运输 穿膜蛋白 单位膜 单位膜模型 小泡运输 电压门控通道第五章内膜系统annulate lamellaeautophagic lysosomecalreticulincis Golgi networkcisternaeclathrin-coated vesiclecontinuous secretioncotranslation insertiondiscontinuous secretionendoplasmic reticulumgated transportglucose regulated protein94,GRP94 glycosylationGolgi complexheavy-chain bindingprotein,BiP heterophagic lysosome internal signal peptideinternal start-transfer peptidelysosomemedial Golgi stackmolecular chaperonemyeloid bodyN-linked glycosylationphagolysosome孔环状片层 自噬溶酶体 钙网素 顺面高尔基网 扁平囊泡 网格蛋白有被囊泡 连续分泌 共翻译插入 非连续分泌 内质网 门孔运输 葡萄糖调节蛋白94糖基化 高尔基复合体 重链结合蛋白 异噬溶酶体内信号肽 内开始转移肽 溶酶体 高尔基中间膜囊 分子伴侣 髓样体 N-连接糖基化 异噬溶酶体 phospholipid exchangeproteins,PEP primary lysosome protein disulfide isomerase,PDI retention protein reticulo-plasmin rough endoplasmic reticulum ,RER sarcoplasmic reticulum secondary lysosome signal hypothesis signal patch signal peptide signal recognition particle,SRPSRP-receptor,SRP-R smooth endoplasmic reticulum ,SERtarget-SNAREs,t-SNAREs tertiary lysosome trans Golgi network translocontransmembrane transport unfolded protein response, UPR磷脂交换蛋白初级溶酶体蛋白二硫键异构酶驻留蛋白 网质蛋白 糙面内质网肌质网 次级溶酶体 信号肽假说 信号斑 信号肽信号识别颗粒信号识别颗粒受体 光面内质网靶SNAREs 三级溶酶体 反面高尔基网 转运体 穿膜运输未折叠蛋白反应vacuoles vesicle vesicles 大囊泡囊泡小囊泡vesicle-SNAREs,v- SNAREsvesicular transport囊泡SNAREs囊泡转运第六章线粒体与细胞的能量转换ATP synthase complex biological oxidation cellular oxidation cellular respiration chemiosmotic coupling hypothesiscristaeelementary particle glycolysisinner membrane intercristae space intermembrane space intracristae spacematrixmatrix spacematrix-targeting sequence,MTS ATP合酶复合体生物氧化细胞氧化细胞呼吸化学渗透假说嵴基粒糖酵解内膜嵴间腔膜间腔嵴内空间基质基质腔基质导入序列mitochondrial disordersmitochondrial phaseouter membraneoxidative phosphorylationpostmitochondrial phasepremitochondrial phasereactive oxygen species, ROSsubstrate-levelphosphorylationtranslocation contact sitetranslocon of the innermembrane,Timtranslocon of the outermembrane,Tomtricarboxylic acid cycle,TCAcycle线粒体疾病线粒体期外膜氧化磷酸化线粒体后期线粒体前期活性氧底物水平磷酸化转位接触点内膜转位子外膜转位子三羧酸循环第七章细胞骨架与细胞的运动actincell cortexcochicine contractile ring cytochalasin B cytoskeletondynamic instability model dyneinfilopodiaintegrin intermediate filaments, IF kinesin lamellipodia microfilaments , MF microfilament associated protein,MAP 肌动蛋白细胞皮层秋水仙素收缩环细胞松弛素B细胞骨架非稳态动力学模型动力蛋白丝状伪足整合蛋白中间纤维驱动蛋白片状伪足微丝微丝结合蛋白microtubule organizing center,MTOCmicrotubules, MTmicrotubule-associadedprotein ,MAPmyosinnucleation phasephalloidinpolymerization phasesliding filament modelsteady state phasetaxoltreadmilling modeltubulinvinblastine微管组织中心微管微管结合蛋白肌球蛋白成核期鬼笔环肽聚合期滑动丝模型稳定期紫杉醇踏车模型管蛋白长春新碱第八章细胞核acrocentric chromosome annular subunitbandcentral domaincentral plug centromerechromatidchromatin chromosomecolumn subunit constitutive heterochromatin cytoplasmic ringdense fibrillar component,DFCeuchromatinexportin facultative heterochromatinfibrillar center,FCfish-trapgenomegranular component,GC heterochromatin nucleosomenucleusouter nuclear membrane pairing domain perinuclear space primary constriction replication origin satellitescaffold radial loop structure model 近端着丝粒染色体环状亚单位带型中央结构域中央栓着丝粒染色单体染色质染色体柱状亚单位组成性异染色质胞质环致密纤维组分常染色质输出蛋白兼性异染色质纤维中心捕鱼笼基因组颗粒中心异染色质核小体细胞核外核膜配对结构域核周间隙主溢痕复制源随体染色体骨架-放射环模型histonekaryophilic proteinsecondary constrictionsolenoidkaryotypekinetochorekinetochore domainluminal subunitmetacentric chromosomeminibandmultiple coiling modelnuclear localization signal,NLSnuclear matrixnuclear matrix associatedproteinnuclear matrix proteinnuclear porenuclear pore complex,NPCnuclear ringnucleolar cyclenucleolar matrixnucleolar organizernucleolar organizing regionnucleolusnucleosomal histonespokesubmetacentric chromosomesupersolenoidtelocentric chromosometelomere组蛋白亲核蛋白次溢痕螺线管核型动粒动粒结构域腔内亚单位中着丝粒染色体微带多级螺旋化模型核内定位信号核基质核基质结合蛋白核基质蛋白核孔核孔复合体核质环核仁周期核仁基质核仁组织者核仁组织区核仁核小体组蛋白辐亚中着丝粒染色体超螺线管端着丝粒染色体端粒第九章基因信息的传递与蛋白质合成activating domain adaptive expression alternative splicing aminoacyl site aminoacyl-tRNA antibiotics anticodon antisense strand 转录激活域适应性表达可变剪接A位点氨酰-tRNA抗生素反密码子反义链CAAT boxcentral dogmachaperoninschromatin remoldingcis-acting elementcoding regioncoding strandcodonCAAT盒中心法则伴侣素染色质重塑顺式作用元件编码区编码链密码子commalessconstitutive expressionconstitutive splicingdegeneracydirectionDNA binding domainenhancerexonGC boxgenegene clustergene familygeneral transcription factor genetic codegenomehelix-loop-helix ,HLHhelix-turn-helix ,HTHheterogeneous nuclear RNA ,hnRNAhighly repetitive sequence inducible geneintronleucine zippermiddle repetitive sequence negative regulationnon-coding regiontranslocationtranspeptidaseubiquitin-proteasomeunique sequenceuniversalwobblezinc finger连续性 组成性表达 常规剪接 简并性 方向性 DNA 结合域 增强子 外显子 GC 盒 基因 基因簇 基因家族 通用转录因子遗传密码 基因组 螺旋-环-螺旋 螺旋-转角-螺旋 不均一核RNA高度重复序列可诱导基因 内含子 亮氨酸拉链 中等重复序列负性调控 非编码区 转位 转肽酶 泛素- 蛋白酶体 单一序列 通用性 摆动性 锌指 operonpeptide formation peptidyl-tRNA site polyribosome positive regulation pribnow box Promoter Registerregulatory gene repetitive sequence repressible gene repressor remodelerribosome circulation sense strandShine-Dalgarno sequence silencer small nuclearribonucleoprotein particle ,snRNPspatial specificity spliceosome split gene structural gene sumoylation TATA box template strand temporal specificity terminatortrans-acting factor transcriptiontranscription factor translation操纵子 成肽 P 位点多聚核糖体 正性调控 Pribnow 盒 启动子 进位 调控基因 重复序列 可阻遏基因 阻遏蛋白 染色质重建子 核糖体循环 有意义链 SD 序列 沉默子核糖核蛋白颗粒空间特异性 剪接体 断裂基因 结构基因 sumo 化 TATA 盒 模板链 时间特异性 终止子反式作用因子 转录 转录因子 翻译第十章细胞连接与细胞黏附adhering junctionadhesion beltanchoring junctioncadherincell adhesioncell adhesion molecule ,CAMcell junctionchemical synapse黏着连接 黏着带 锚定连接 钙黏着蛋白 细胞黏附 细胞黏附分子 细胞连接 化学突触 claudincommunicating junction connexon desmosomedesmosome junction electric coupling electronic synapse filamin密封蛋白 连接连接子 桥粒 桥粒连接 电耦联 电突触 细丝蛋白focal adhesiongap junction hemidesmosome heterophilic binding homophilic binding immunoglobin-superfamily, Ig-SF inside outintegrinintracellular anchor protein linker-dependent binding metabolic coupling epithelial-mesenchymal 黏着斑间隙连接半桥粒异亲型结合同亲型结合免疫球蛋白超家族由内向外整联蛋白细胞内锚定蛋白连接分子依赖性结合代谢耦联上皮-间质转型通讯neural cell adhesionmolecule,N-CAMoccludinoccluding junctionoutside inplectinselectinsynapsetalintransmembrane adhesionprotein神经细胞黏附分子闭合蛋白封闭连接由外向内网蛋白选择素突触踝蛋白穿膜黏连蛋白transition,EMT第十一章细胞外基质及其与细胞的相互作用anchorage dependent growth anoikisbasal lamina,basement membrane,BM chondroitin sulfate,CS collagencollagen disease collagenasecore proteindermatan sulfate,DS elastaseelastinextracellular matrix, ECM fibronectin, FN glycosaminoglycan, GAG 锚定依赖性生长失巢凋亡基膜硫酸软骨素胶原胶原病胶原酶核心蛋白硫酸皮肤素弹性蛋白酶弹性蛋白细胞外基质纤连蛋白糖胺聚糖glycosyltransferasesheparan sulfate,HSheparinhyaluronic acid,HAkeratan sulfate,KSlaminin,LNmatrix metalloproteinases,MMPnidogen,entactinperlecanproteoglycan,PGsyndecantriple helixtropoelastin糖基转移酶硫酸乙酰肝素肝素透明质酸硫酸角质素层粘连蛋白基质金属蛋白酶巢蛋白渗滤素蛋白聚糖连接素三股螺旋可溶性弹性蛋白原第十二章细胞的信号转导adenylate cyclase, AC calmodulin,CaM cAMP-dependent protein kinase A,PKA cascadecGMP depedent protein kinase G,PKG cyclic AMP,cAMP cyclic GMP,cGMP diacylglycerol,DAG effector protein 腺苷酸环化酶钙调蛋白cAMP依赖蛋白激酶A级联反应cGMP依赖蛋白激酶G环磷酸腺苷环磷酸鸟苷二酰基甘油效应蛋白first messengerG proteinG protein linked receptorguanylate cyclase,GCinositol trisphosphate,IP3insulinion channel receptorligandprotein tyrosine kinase,PTKreceptorsecond messenger第一信使G蛋白G蛋白偶联受体鸟苷酸环化酶三磷酸肌醇胰岛素离子通道受体配体酪氨酸蛋白激酶受体第二信使serine/threonine kinases ,STKsignal transduction丝氨酸/苏氨酸蛋白激酶 信号转导 signaling networktyrosine-specific protein kinase receptor ,TPKR信号网络酪氨酸蛋白激酶型受体第十三章细胞分裂与细胞周期amitosis anaphase anti oncogene aster bivalentCdk inhibitor ,CKI cell cycle cell divisioncellular oncogene ,C-onc centrosome chalone checkpoint chiasmachiasma terminalization contractile ring cyclincyclin-dependent kinase ,CDKcytostatic factor,CSF direct division growth factor无丝分裂 后期 抑癌基因 星体 二价体Cdk 激酶抑制物 细胞周期 细胞分裂 细胞癌基因 中心体 抑素 检测点 交叉 交叉端化 收缩环 细胞周期蛋白细胞周期蛋白依赖性激酶细胞静止因子 直接分裂 生长因子indirect divisioninterphasekinetochore microtubule maturation promotingfactor ,MPF meiosismetaphasemitosismitosis,Mmitotic apparatuspolar microtubuleprophaseproto-oncogenerecombination nodulespindlesynapsissynaptonemal complex ,SC telophasetetradV-oncogene ,V-onc间接分裂 分裂间期 动粒微管成熟促进因子减数分裂 中期 有丝分裂 分裂期 有丝分裂器 极微管 前期 原癌基因 重组小结 纺锤体 联会 联会复合体 末期 四分体 癌基因 第十五章细胞分化apical ectodermal cap cell determination cell differentiation cellular reprogramming cleavagecombinatory control compensatory regeneration dedifferentiation differential expression ectodermembryonic induction endoderm顶端外胚层帽 细胞决定 细胞分化 细胞重编程 卵裂 组合调控补偿性再生 去分化 差异表达 外胚层 胚胎诱导 内胚层epimorphosis regeneration gastrulation genomic imprinting grafting experiment histone code homeobox homeobox gene homeodomain homeodomain protein homeosis homeotic gene housekeeping gene微变态再生 原肠形成 基因组印记 胚胎移植实验 组蛋白密码 同源异形框 同源异形框基因 同源异形结构域 同源异形域蛋白 同源异形转变 同源异形基因 管家基因induced pluripotent stem cells juxtacrine interaction lateral inhibitionlocus control region,LCR long non-coding RNA,lncRNAluxury geneluxury proteinmaster control gene maternal effect gene,MEG mesoderm metamorphosis microRNA, miRNA morphallaxis regeneration non-coding RNA诱导多能干细胞,iPS细胞近分泌相互作用侧向抑制基因座控制区长链非编码RNA奢侈基因奢侈蛋白细胞分化主导基因母体效应基因中胚层变态微小RNA变形再生非编码RNAorganogenesisparacrine factorpluripotent cellregenerationregeneration blastemasmall interfering RNA, siRNAsomatic recombinationspatial specificitystage specificitytemporal specificitytotipotent celltotipotent nucleustransdeterminationtransdifferentiationunipotency器官发生旁分泌因子多能(干)细胞再生再生胚芽小干扰RNA体细胞重组阶段特异性空间特异性时间特异性全能(干)细胞全能性细胞核转决定转分化单能第十六章细胞衰老与细胞死亡anoikisanti-apoptosis gene apoptosis apoptotic bodies autophagosome autophagycell deathcell senescencecell shrinkage chromatin condensation ClassIIIPI3Kcytochrome C, cyt C death receptor, DR DNA ladders 失巢凋亡抗凋亡基因细胞凋亡凋亡小体自噬泡细胞自噬细胞死亡细胞衰老细胞皱缩染色质凝聚III型磷脂酰肌醇三磷酸激酶细胞色素C死亡受体DNA梯状条带DNA stainabilityendonucleaseHayflick life spaninterleukin-1β convertingenzyme, ICEnecrosisnerve growth factor receptor,NGFRpermeability transition pores,PT poresphosphatidylserine, PSprogrammed cell death, PCDreactive oxygen species, ROStumor necrosis factor receptor,TNFRDNA可染性核酸内切酶Hayflick 界限白细胞介素-1β转换酶细胞坏死神经生长因子受体渗透转变孔磷脂酰丝氨酸程序性细胞死亡活性氧类物质肿瘤坏死因子受体第十七章干细胞与组织的维持与再生adult stem cell asymmetry division cancer stem cell, CSC chimeracorneal stem cell directly induced differentiation 成体干细胞不对称分裂癌干细胞嵌合体角膜干细胞直接诱导分化dedifferentiationembryoid body, EBembryonic carcinoma cell,ECembryonic germ cell,EGembryonic stem cell,ESepidermal stem cell去分化类胚体胚胎癌细胞胚胎生殖细胞胚胎干细胞皮肤干细胞hematopoiesis stem cell ,HSCinner cell mass ,ICMintegral membrane protein induced pluripotent stem cellintegrinintestinal stem cellliver stem celllung stem cellmarrow cavitymesenchymal stem cell, MSCmultipotent stem cellmyogenic stem cellneural stem cells, NSCpancreatic stem cellplasticitypluripotent stem cell造血干细胞 内细胞团 整合膜蛋白诱导多能干细胞,iPS细胞 整联蛋白 小肠干细胞 肝干细胞 肺脏干细胞 骨髓腔 间充质干细胞 专能干细胞 肌肉干细胞 神经干细胞 胰腺干细胞 可塑性 多能干细胞 pluripotenyprogenitor cell renal stem cell self-maintenance somatic stem cellspermatogonial stem cell stage-specific embryonic antigen,SSEA stem cell stem cell niche symmetry division tissue specific stem cell totipotencytotipotent stem cell transdetermination transdifferentiation unipotency多能 前体细胞 肾脏干细胞 自稳定性 成体干细胞 精原干细胞胚胎阶段特异性抗原干细胞 干细胞巢 对称分裂组织特异性干细胞 全 能 全能干细胞 转决定 转分化 专 能。
5蛋白质翻译
AAAAAAA-OH
mRNA上存在遗传密码 mRNA分子上从5至3方向,由AUG开始,
每3个核苷酸为一组,决定肽链上某一个氨 基酸或蛋白质合成的起始、终止信号,称 为三联体密码(triplet codon)。
从mRNA 5端起始密码子AUG到3端终止
密码子之间的核苷酸序列,各个三联体密码 连续排列编码一条多肽链,称为开放阅读框 架(open
5' 7 m Gppp
reading frame, ORF)。
编码区 ORF AUG UAA 3' AAA……An
5'非翻译区
3'非翻译区
2、遗传密码的破译
• 基因密码的破译是六十年代分子生物学最辉煌的成就; • 1954年,物理学家George Gamow的数学推理(43=64 ); • 1961年,Brenner和Crick首先肯定了三个核苷酸的推理 ;
U
One tRNA anticodon may pair with as many as three codons.
3、遗传密码的特点
(1)连续性(commaless) :两个密码子之间无 任何核苷酸加以隔开和重叠,如插入/删除 碱基,可发生移码突变或框移
5′…. UACGGACAUCUG….3′
酪 插入 甘 组 蛋
5′….UACCGGACAUCUG….3′
酪 精 苏 半胱
缺失
5′…. UACGACAUCUG….3′
核苷酸结合技术
Nirenberg和Leder,1964
核苷酸结合技术
蛋白质的生物合成(翻译)

已发现少数例外,如动物细胞的线粒体、植 物细胞的叶绿体等。 密码的通用性进一步证明各种生物进化自同 一祖先。
16
4. 方向性(direction): 指阅读mRNA模板上的三联体密码时,只能沿 5’→3’方向进行。
17
5. 摆动性(wobble) 转运氨基酸的tRNA的反密码需要通过碱基互 补与mRNA上的遗传密码反平行配对结合,但 反密码与密码间不严格遵守常见的碱基配对规 律,称为摆动配对。
4
一、mRNA模板及遗传密码
mRNA是翻译的直接模板
遗传学将编码一个多肽的遗传单位称为顺反 子(cistron)。 原核细胞中数个结构基因常串联为一个转录 单位,转录生成的 mRNA 可编码几种功能相 关的蛋白质,为多顺反子(polycistron) 。 真核 mRNA只编码一种蛋白质,为单顺反子 (single cistron) 。
47
1. 核蛋白体大、小亚基分离: IF-1和IF-3与小亚基结合,促进核蛋白体大、小 亚基拆离,为新一轮合成作准备。
IF-3
IF-1
48
2. mRNA在小亚基的精确定位结合:
5' IF-3
AUG
3' IF-1
49
S-D序列
原核mRNA的起始部位由一段富含嘌呤的特殊
核苷酸顺序组成,称为S-D序列(核蛋白体结合
39
五、延长因子(EF)
与多肽链合成的延伸过程有关的蛋白因子称为 延长因子(elongation factor,EF)。 原核生物中存在3种延长因子(EFTU,EFTS, EFG),真核生物中存在2种(EF1,EF2)。 EF的作用主要促使氨基酰tRNA进入核蛋白的 受体,并可促进移位过程。
40
翻译蛋白质

翻译蛋白质Protein TranslationProtein translation, also known as protein biosynthesis, is the process by which cellular ribosomes synthesize proteins. This process occurs in all living organisms and is crucial for maintaining various cellular functions.The process of protein translation involves several steps. Firstly, the DNA sequence of a gene is transcribed into a complementary RNA sequence through a process called transcription. This RNA molecule, known as mRNA (messenger RNA), carries the genetic information from the DNA to the ribosome for protein synthesis.Next, the mRNA molecule is transported out of the nucleus and into the cytoplasm, where the ribosomes reside. The ribosome binds to the mRNA at a specific location called the start codon, which marks the beginning of a protein-coding sequence.Once the ribosome has recognized the start codon, it begins the process of protein synthesis. The ribosome moves along the mRNA molecule, reading the genetic code in the form of codons - three nucleotide bases that specify a particular amino acid or signal the termination of protein synthesis.At each codon, the ribosome recruits a specific transfer RNA (tRNA) molecule that carries the corresponding amino acid. The tRNA molecule recognizes the codon through its anticodon, a complementary sequence of nucleotides. This ensures that the appropriate amino acid is added to the growing protein chain.As the ribosome continues to move along the mRNA, it joins the amino acid carried by each tRNA to the growing protein chain through a peptide bond. This process repeats until the ribosome encounters a stop codon, indicating the completion of protein synthesis.After protein translation, the newly synthesized protein undergoes further modifications to become functional. These modifications may include folding, post-translational modifications such as phosphorylation or glycosylation, and subcellular localization.The accuracy and efficiency of protein translation are essential for cellular health. Errors in translation can lead to the production of faulty proteins, which can have detrimental effects on cellular functions and contribute to the development of various diseases.In conclusion, protein translation is a complex and highly regulated process that is vital for the synthesis of functional proteins. Understanding the mechanisms of protein translation can provide valuable insights into cellular processes and disease mechanisms.。
细胞生物学进展名词解释

第一章 subculture
缩写
中文 冻存 低温保护剂
传代培养
passage subcultivation ratio
第二章
Archaebacteria Eubacteria Eucaryotes Procaryotes Metazoa (Metazoan) Homo sapiens Mus musculus Xanopus leavis Drosophila malanogaster Caenorhabditis elegans vertical gene transfer horizontal gene transfer
oncogene addiction
癌蛰伏 原纤毛
杂合丢失 Hayflick界限 染色体碎接 DNA永生化链假说
癌基因成瘾性
解释 细胞冻存是细胞保存的主要方法之一。利用冻存技术将细胞置于-196℃液氮中低温 保存,可以使细胞暂时脱离生长状态而将其细胞特性保存起来,这样在需要的时候 当原代培养成功以后,随着培养时间的延长和细胞不断分裂,一则细胞之间相互接 触而发生接触性抑制,生长速度减慢甚至停止;另一方面也会因营养物不足和代谢 物积累而不利于生长或发生中毒。此时就需要将培养物分割成小的部分,重新接种 到另外的培养器皿(瓶)内,再进行培养。这个过程就称为传代(passage)或者再
是指物种内部的个体在正常繁殖过程中,发生于垂直世代之间的基因转移。 发生于不同物种之间的基因转移。
The DNA damages caused by genotoxic agents that can alter the coding information of DInNheAri.table changes in phenotype that do not result from changes in DNA sequence. 泛指 各种非经DNA序列改变所产生的可遗传的变化(包括个体和细胞水平的遗传变化) 。已知Gene - RNA - protein -求证phenotype 维持细胞在机体中最基本的功能所必需的基因,因而也是在所有或大部分有核细胞 内都能保持高水平表达的基因,亦即其表达与否与细胞分化状态无关。 是参与多细胞生物细胞社会性相互作用的基因,其表达具有细胞类型特异性,亦即 与细胞分化密切相关,故又称tissue specific genes 在物种形成事件(speciation event)之后歧化(diverge)的一对基因,它们分别存 在于两个具有共同祖先的物种中,彼此互为正源基因。 在基因倍增事件(duplication event)之后歧化的一对基因,它们存在于同一个物种 中,彼此互为旁源基因。 different variants of the same gene. 基因座上的任何突变形式、表观遗传修饰形式、 以及突变前的形式(野生型)彼此之间均称为等位基因。广义alleles也包括不含基 因的任何locus上的变异
第5章 蛋白质protein-PPT课件

NATIVE
Slow under physiological conditions
DENATURED
AGGREGATED or other ingredient interactions
Unfolding forces some hydrophobic AA to surface
Consequences of Denaturation
羟基氨基酸
丝氨酸Ser 苏氨酸Thr
含硫氨基酸
半胱氨酸Cys 胱氨酸Cys-Cys 蛋氨酸Met
芳香族氨基酸
苯丙氨酸Phe 酪氨酸Tyr
杂环氨基酸
组氨酸His 色氨酸Trp
脯氨酸Pro 羟脯氨酸Hyp
Hydroxyproline(Hyp) -OH
必需氨基酸
赖氨酸 亮氨酸 异亮氨酸 缬氨酸 苏氨酸 蛋氨酸 苯丙氨酸 色氨酸
半必需氨基酸 胱氨酸〔蛋氨酸〕 酪氨酸〔苯丙氨酸〕
氨基酸的性质 两性离子
酸碱性 等电性 疏水性 与茚三酮反响生成蓝紫色物质
1.0 NaOH 0.5
0
0.5 HCl
1.0
等离子点 pK2 pK1
pH
5.1.3 氨基酸的呈味电性酸性质
苦味 鲜味
5.2 蛋白质的结构Protein Structure
G D N G H bo G n e d l G eH G V D T S W con
One native state
S=k ln W
Increased chain entropy
蛋白质天然
结构是各种吸引和 推斥相互作用的净 结果,在生理条件 下是最稳定的状态
Many denatured states
5.2.1 蛋白质的一级结构
第五章 蛋白质的翻译2
能特异切割X-Y 2肽。其中X是Ala,Asp和Glu; Y是Ala或Pro。
2 蛋白质易位
2.1 2.2 2.3 2.4 概 述 共翻译易位 翻译后易位
细菌蛋白质易位 2.5 分子伴侣
Back
2.1 概 述
细胞中蛋白质合成:绝大多数在细胞质中合成; 小一部分在细胞器(叶绿体和线粒体)中合成。 定位于细胞器内的大部分蛋白质在细胞质中合 成,细胞器内合成的留在细胞器内。 蛋白质插入或穿过生物膜的过程称为蛋白质易 位(protein translocation)。
Back
1 蛋白质的加工
1.1 N端fMet或Met切除
1.2 二硫键形成
1.3 特定氨基酸化学修饰
1.4 新生肽链中非功能片段切除
Back
1.1 N端fMet或Met的切除
细菌蛋白质N端甲酰基能被甲酰化酶水解。
不管原核生物或真核生物,N端Met常在肽 链合成完前被切除。 有些病毒mRNA可翻译成很长的多肽链, 经蛋白酶水解后得到几个功能蛋白质分子。
1.2 二硫键形成
mRNA中没有胱氨酸密码子,而不少 蛋白质含有二硫键.这是蛋白质合成后
通过两个半胱氨酸的氧化作用生成的。
1.3 特定氨基酸的化学修饰
氨基酸侧链修饰作用包括磷酸化(如核糖体 蛋白质)、糖基化(如各种糖蛋白)、甲基化 (如组蛋白、肌肉蛋白质)、乙基化(如组蛋 白)、羟基化(如胶原蛋白)和羧基化等。
2.5.2 生物学功能
(1)帮助新生蛋白质正确折叠
(2)纠正错误折叠或介导其降解
暴露的疏水部分相互作用
Proteins emerge from the ribosome or from passage through a membrane in an unfolded state that attracts chaperones to bind and protect them from misfolding.
第5章 翻译
一、 mRNA是蛋白质生物合成的直接模板 (一)mRNA结构
• 不同生物mRNA序列,都具有5-端非翻译 区、开放阅读框架区和3-端非翻译区。 • 真核生物mRNA的5-端有帽子结构, 3-端 有多聚腺苷酸(polyA)尾。 1961年,Nirenberg 证明 了mRNA的模板作用。
5'
一、原核生物的肽链合成过程
(一)肽链合成的起始
• 指 mRNA 和起始氨基酰-tRNA分别与核糖体结合 而形成翻译起始复合物 (translational initiation complex)的过程。 • 参与起始物质:30S小亚基、50S大亚基、mRNA、 fMet-tRNAfMet、起始因子、GTP和Mg2+。
摆动配对
密码子、反密码子摆动配对
tRNA反密码 子第1位碱基
U
I
U
G
A C
mRNA密码 子第3位碱基
C,A,U A,G C,U U G
二、核糖体是蛋白质生物合成的场所
核糖 体的 组成
核糖体的组成
核蛋 白体 原核生物 蛋白质 S值 真核生物
rRNA 蛋白质
S值
rRNA
小亚基
大亚基
核蛋白体
21种
36种
同义密码子
为同一种氨基酸编码的各密码子,亦称简并性密码子。 • 前两个碱基均相同,只是第三个碱基不同。 • 若头两个碱基发生点突变,可译出不同氨基酸,而第 三个碱基的突变,不会影响氨基酸的翻译。 •遗传密码的特异性主要取决于前两位碱基。 密码子简并性的生物学意义:减少基因突变对蛋白质功 能的影响。 GCU ACU
ORF 编码区
AUG UAA
3'
m7Gppp
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• type I: Val,Arg,Gln,Glu,Ile,Leu,Met,Trp,Tyr,paracodon 大多位于反密码子臂
• type II: paracodon 大多位于氨基酸接受臂,个别还同时 在附加臂上有相应碱基
两类酶与tRNA反应时--接近模式不同
• AUG前有 S – D 序列
• S – D序列(Shine-Dalgarno sequence): 位于AUG 上游 4 ~13 个NT处的富含嘌呤的 3 ~9个NT的共同序列 ,其一致序列为 AGGAGG;与核糖体小亚基内16S rRNA 的 3’端富含嘧啶的序列3’------UCCUCC-----5’互 补,使 tRNA正确定位于起始codon。
• 需要三种底物: AA、tRNA 、ATP • 因此有三个位点:aa binding site、tRNA binding site、
ATP site • Prok中AARS有20种,对AA专一 • 按反应机制AARS分为两类
• Type Ⅰ类酶 先将氨酰基转移到 tRNA 3’ 端A的 2’-OH 然后通过转酯反应转移到 3’ –OH上
5.1.2 mRNA
• 单顺反子mRNA(monocistronic mRNA)
• 编码一个蛋白质的….. • 部分Prok 所有Euk的mRNA • 多顺反子mRNA(polycistronic mRNA ): • 部分原核mRNA • mRNA 分子的组成: • 编码区 起始AUG到终止密码子 • 前导序列 AUG之前的 5’ 端非编码区(5’UTR) • 尾巴 终止密码子之后,不翻译的 3’ 端
Prok.mRNA转录、翻译和降解周期
真核生物mRNA的特征
• 一般为单顺反子 • 5’ 端的帽子对翻译有增强作用,且 5’ 帽子和3’polyA尾
对翻译效率的调节有协同作用 • “第一AUG规律” 即----- 绝大部分以AUG作为起始
codon • 起始AUG有如下特点
• C C A C C AUG G;-3位的A和+4位的G….. 对于准确翻译 至关重要
当…………,有S-D 序列的特征)
• Prok中多顺反子mRNA中各基因的产物数量不一定相 等
• 比如乳糖操纵元中,Z:Y:A=1:0.5:0.2
Spacer 较长 Spacer 较短
• 其他特征
• 5’ 端有300个左右NT的leader(A/G----AUG)
• AUG作为起始密码子(行的场所 ,含大小亚基 • Prok. protein 约占细胞的10%,rRNA占总RNA的80%
,Euk中相对比例小些; • 细菌中常以多聚核糖体的形式; • Euk中大多与细胞骨架和内质网膜结合在一起
核糖体的结构
• 同工tRNA
• 携带AA相同而反密码子不同的一组tRNA • 结构上能被AA– tRNA合成酶识别的共性
副密码子与氨酰基tRNA的合成
• 氨酰基tRNA的合成执行着双重功能 • 为肽建的合成提供能量 • 执行遗传信息的解读 • 模板mRNA只能识别特异的tRNA而不是AA
• AA– tRNA合成酶(AARS)
生命的中心法则
5.1 基本元件
• tRNA • mRNA • rRNA与核糖体
5.1.1 tRNA
• 最小的 RNA,4S,70 ~ 80个base; • 1964年,Holly. R. 鉴定出 tRNAphe 的二级结构为三
叶草形(77个NT); • 三叶草结构包括5个臂、4个环
• 氨基酸接受臂 • tRNA 的5’与3’-末端碱基配对形成 • 3’ 端永远为不配对的CCA序列 • 最后的A的 3’ 或 2’-OH可以被氨酰化
“L”结构的稳定; • “L”结构中碱基堆积力大,使其拓扑结构趋于稳定; • wobble base位于“L”结构末端,堆积力小、自由度大
、使碱基配对摇摆。
tRNA 的种类
• 附加臂的大小
• Ⅰ类tRNA:3-5 bp (75%) • Ⅱ tRNA: 13-21 bp (25%)
• 起始tRNA
• Prok中, 携带甲酰甲硫氨酸(fMet) • Euk中, 携带甲硫氨酸(Met)
• TypeⅡ类酶 直接将氨酰基转移至 3’ –OH 上
• Paracodon (副密码子)
• tRNA中决定负载特定氨基酸的空间密码,为同一种AARS 所 识别的一组同功受体具有相同的副密码子
tRNA
Ala (GGC)
tRNA
Ala (UGC)
具有G3 :U70 paracodon
• AARS 对paracodon 的识别与结合是通过氨基酸与碱基之间的 连接实现的
• D臂、反密码子臂、TψC臂、附加臂 ; • DHU环、反密码子环、 TψC环、附加环
tRNA真实的空间构型
“ L”形三级结构
“L”结构域的功能
• aa accept arm 位于“L”一端,契合于核糖体的肽基; • anti-codon arm 位于”L”另一端,与结合在核糖体小
亚基上的 codon of mRNA配对; • TΨC loop & DHU loop位于“L”两臂的交界处,利于
Chapter 5
Protein 翻译 Protein translation
从DNA转录到RNA可以利用碱基互补配对,但mRNA 与的碱基序列与蛋白质的氨基酸序列却没有直接的对 应关系,他们之间也没有直接的作用力,那么在这两 种完全不同的遗传语言之间是如何沟通、如何传递信 息的呢?
信息的传递者就是tRNA,准确的来说,应该是氨酰基 tRNA,它利用反密码子与mRNA配对,通过氨酰 tRNA 合成酶、利用副密码子来选择特异的氨基酸。
原核生物mRNA的特征
• 大部分为多顺反子
• spacer: 各顺反子之间(基因之间)的非编码区长度变化很大 ,从重叠到100个碱基不等;
• 因此,多顺反子的mRNA翻译有两种情况:
• spacer较长时,下一个基因的产物合成起始是独立的 • spacer长度较短时,两个顺反子使用相同的核糖体或小亚基( 相