镉超富集植物球果蔊菜对镉_砷复合污染的反应及其吸收积累特征
复合污染土壤上几种叶类蔬菜对Cd和As的富集效应

复合污染土壤上几种叶类蔬菜对Cd和As的富集效应MENG Yuan;ZHANG Liang;WANG Lin-quan;SHANGGUAN Yu-xian;YANG Yang;LI Xue-fang;LI Na【摘要】[目的]不同蔬菜镉、砷富集系数各异,对镉和砷污染土壤的响应也不同.研究复合污染土壤上不同叶类蔬菜对Cd和As的积累效应,为轻度-中度Cd和As污染土壤的合理与安全利用提供适宜的蔬菜种类.[方法]采集了西安市12个污染程度不同的菜地耕层土壤,于2015年3月6日—5月26日在西北农林科技大学资源环境学院遮雨大棚内进行了盆栽试验.供试7种叶菜,包括菠菜、油菜、生菜、油麦菜、苋菜、空心菜和茼蒿.蔬菜收获后,测量了蔬菜产量、Cd和As含量与吸收累积量,计算了蔬菜对Cd和As的富集系数等,并用线性回归模型研究了不同蔬菜栽培的土壤Cd和As安全临界值.[结果]镉污染土壤(0.6~2.4 mg/kg)对大多数蔬菜生物量有抑制效应,中、低浓度镉砷复合污染(Cd 1.0~2.4 mg/kg,As 24.9~26.8 mg/kg)对供试蔬菜生长没有叠加效应.镉污染土壤上,菠菜、油菜、苋菜叶、生菜可食部Cd含量均超出食品安全限量标准(0.2 mg/kg),其中菠菜和油菜Cd最高超标4倍以上;而茼蒿和空心菜茎秆Cd未超标.虽然供试蔬菜砷含量随着土壤砷含量增加有升高趋势,但叶菜As含量没有超标.7种蔬菜Cd富集系数为0.083~0.491,高低顺序为油菜、菠菜、生菜和苋菜叶>油麦菜、苋菜茎和空心菜叶>空心菜茎和茼蒿.菠菜、油菜、生菜、油麦菜、苋菜、空心菜和茼蒿土壤Cd安全临界值分别为0.33、0.38、0.46、1.15、0.59~1.79、1.49~8.16和8.98~17.11 mg/kg,其中菠菜、油菜和生菜阈值与现行标准(0.3~0.6 mg/kg)相当,而油麦菜、苋菜、空心菜和茼蒿均大于土壤重金属污染限量值.As富集系数为0.002~0.006,空心菜叶和茼蒿叶片As富集系数显著高于其他蔬菜.7种蔬菜的土壤As临界阈值分别为62.31、70.35、70.21、67.41、67.86~90.43、57.21~75.70和72.43~105.06 mg/kg,均高于现行标准(25 mg/kg).[结论]中等程度的Cd和As复合污染土壤上,Cd对蔬菜的生长有显著的抑制,As与Cd没有叠加作用.不同蔬菜的产量、污染程度和安全阈值等有显著差异,因此选择低富集、抗污染蔬菜品种是利用中低重金属污染土壤的一条可行途径.空心菜和茼蒿对Cd富集系数低,可推荐在中、低污染土壤上种植.【期刊名称】《植物营养与肥料学报》【年(卷),期】2019(025)006【总页数】10页(P972-981)【关键词】镉;砷;富集系数;土壤阈值;菠菜;茼蒿【作者】MENG Yuan;ZHANG Liang;WANG Lin-quan;SHANGGUAN Yu-xian;YANG Yang;LI Xue-fang;LI Na【作者单位】;;;;;;【正文语种】中文土壤污染状况调查公报 (2014) 的发表和土壤污染防治行动计划 (2016) 的出台表明土壤污染治理刻不容缓。
植物激素、螯合剂复合处理对镉胁迫下籽粒苋生长和富集镉的影响

㊀山东农业科学㊀2023ꎬ55(11):151~156ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2023.11.022收稿日期:2023-01-11基金项目:山东省自然科学基金项目(ZR2022MC203)ꎻ山东省高校科技计划项目(J14LE54)ꎻ菏泽学院植物生物学重点实验室资助项目作者简介:郭梦露(1986 )ꎬ女ꎬ山东菏泽人ꎬ讲师ꎬ主要从事植物修复和植物抗逆研究ꎮE-mail:549802453@qq.com植物激素、螯合剂复合处理对镉胁迫下籽粒苋生长和富集镉的影响郭梦露ꎬ沈国明ꎬ王宜磊(菏泽学院农业与生物工程学院/植物生物学重点实验室ꎬ山东菏泽㊀274015)㊀㊀摘要:为探讨植物激素和螯合剂(EDTA)复合处理对镉胁迫下籽粒苋生长与修复率的影响ꎬ本试验以生物量较大的籽粒苋为材料ꎬ采用盆栽方法ꎬ研究不同复合处理对镉胁迫下籽粒苋生长以及镉富集量的影响ꎮ结果表明:与单一Cd2+胁迫处理相比ꎬ植物激素和螯合剂的复合处理缓解Cd2+胁迫对籽粒苋生长的影响ꎬ使其各部分的生物量增加ꎬ增强根部和地上部对Cd2+的吸收和富集ꎬ进一步提高了籽粒苋对Cd2+的植物修复效率ꎬ且复合处理以根部浇灌方式的修复效率更高ꎮ关键词:镉胁迫ꎻ籽粒苋ꎻ植物修复ꎻ植物激素ꎻ螯合剂中图分类号:S519:X503.231㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2023)11-0151-06EffectsofPhytohormonesandChelatingAgentCombinedTreatmentsonGrowthandCadmiumAccumulationofGrainAmaranthunderCadmiumStressGuoMengluꎬShenGuomingꎬWangYilei(CollegeofAgriculturalandBiologicalEngineering/KeyLaboratoryofPlantBiologyꎬHezeUniversityꎬHeze274015ꎬChina)Abstract㊀Inordertoinvestigatetheeffectsofdifferentphytohormonescombinedwithchelatingagent(EDTA)onthegrowthandremediationefficiencyofgrainamaranthundercadmiumstressꎬthisexperimentwasconductedtakinggrainamaranthwithlargebiomassasmaterialandbypotmethodtostudytheeffectsofdifferentcombinedtreatmentsonthegrowthandcadmiumaccumulationamountofgrainamaranthundercad ̄miumstress.TheresultsshowedthatꎬcomparedwiththesingleCd2+treatmentꎬthecombinedtreatmentsofphytohormonesandchelatingagentalleviatedtheinfluenceofCd2+stresstothegrowthofgrainamaranthꎬin ̄creasedthebiomassofeachpartꎬenhancedtheabsorptionandenrichmentofCd2+inrootandshootꎬandfur ̄therimprovedthephytoremediationefficiency.Thecombinedtreatmentshadhigherremediationefficiencybyrootwatering.Keywords㊀CadmiumstressꎻGrainamaranthꎻPhytoremediationꎻPhytohormoneꎻChelatingagent㊀㊀随着工业和经济的快速发展ꎬ土壤重金属污染问题日益严重ꎬ我国受Cd㊁As㊁Pb㊁Hg㊁Zn等重金属污染的耕地面积约占总耕地面积的五分之一[1]ꎮ重金属排放到环境中ꎬ严重污染水体和土壤ꎬ其中有高毒性和潜在致癌性的Cd是污染超标率最高的重金属ꎻ土壤Cd点位超标率最高达7%ꎬ其通过植物吸收㊁积累㊁富集后经食物链进入人体ꎬ从而对人类健康造成威胁[2-3]ꎮCd胁迫对植物生长发育有明显的抑制和毒害作用[4]ꎬ可通过抑制种子中的淀粉酶㊁蛋白酶活性影响淀粉和蛋白质的分解ꎬ从而对种子萌发产生影响[5]ꎮ因而ꎬ有必要采取不同技术措施对Cd污染进行有效修复ꎮ植物修复技术因具有成本低㊁不引入二次污染等优势而前景广阔ꎬ其修复效率主要取决于植物生物量和其累积重金属的能力ꎮ选择生物量大㊁富集量大的植物以及添加外源激素可以大大提高植物的修复效率[6-12]ꎮ众多研究结果表明ꎬ植物激素能缓解重金属的植物毒性ꎬ提高植株的抗逆性ꎬ促进其生长发育[13-15]ꎻ螯合剂对重金属离子具有较强的螯合力ꎬ施加螯合剂能提高重金属的迁移能力和生物有效性[16-21]ꎮ如Wang等[22]研究EDTA(乙二胺四乙酸)强化盐生植物景天三七对土壤中Pb㊁Cd的去除效果发现ꎬEDTA可以强化景天三七修复土壤的能力ꎬ使Pb㊁Cd的去除率分别达到37.87%和41.61%ꎮ综上ꎬ很多研究表明植物激素和螯合剂的添加可提高植物修复效率ꎬ但关于植物激素和螯合剂复合处理的研究较少ꎬ两种添加剂复合处理是否会大大增加植物修复效率仍未可知ꎻ外源激素的添加方式有多种ꎬ具体何种添加方式能最大程度增加修复效率也鲜有研究ꎮ龙玉梅[23]等通过温室盆栽试验比较籽粒苋㊁龙葵㊁商陆㊁青葙4种植物对镉污染土壤的修复效果ꎬ结果表明ꎬ4种植物可收集的镉总量排序为籽粒苋>青葙>商陆>龙葵ꎮ籽粒苋对镉胁迫的耐受性及镉富集能力均较强ꎬ是一种良好的Cd污染土壤修复材料[24-27]ꎮ本试验以一年生籽粒苋(AmaranthushybridusL.)为材料ꎬ选取叶面喷施和根部浇灌两种添加方式ꎬ研究植物激素和螯合剂复合处理对镉胁迫条件下籽粒苋的植株生长状况和富集镉情况的影响ꎬ旨在寻求提高污染土壤植物修复效率的方法ꎬ为研究植物激素与螯合剂效应提供一定的理论与实践依据ꎮ1㊀材料与方法1.1㊀试验材料籽粒苋是苋科苋属一年生草本C4植物ꎬ叶宽大㊁柔软㊁直立ꎬ主茎粗壮ꎬ适应性强ꎬ具有耐旱㊁耐盐碱㊁耐贫瘠土壤的特点ꎮ籽粒苋在国内北方地区种植较多ꎬ生育期内可进行多次收割ꎬ鲜草产量可达130t/hm2ꎮ本试验选用美国籽粒苋K472ꎬ种子购于山东省种子有限公司ꎮ1.2㊀试验试剂Hoagland全培养液配方为:①微量元素:每升水中加H3BO32.86g㊁MnCl2 4H2O1.81g㊁ZnSO4 7H2O0.22g㊁CuSO4 5H2O0.08g㊁H2MoO4 H2O0.02gꎻ②大量元素:每升培养液中加1mol/LKH2PO41mL㊁1mol/LKNO35mL㊁1mol/LCa(NO3)25mL㊁1mol/LMgSO42mLꎻ③Fe-EDTA溶液:每升水中加入Na2-EDTA7.45g㊁FeSO4 7H2O5.57gꎮ镉溶液为20mg/kgCd(NO3)2 4H2OꎻEDTA添加浓度为1.5mg/Lꎻ三种激素(IAA㊁GA㊁SA)的添加浓度选取前期试验得到的最佳浓度ꎬ分别为100㊁500㊁100mg/Lꎬ与EDTA分别配成不同浓度的水溶液ꎮEDTA㊁Cd(NO3)2 4H2O和植物激素均为化学分析纯试剂ꎬ购于山东星伟化学试剂有限公司ꎮ1.3㊀试验设计及方法试验于2022年3月20日开始ꎮ对籽粒苋种子进行表面消毒后浸种12hꎬ暗中催芽24h后于温室中培养幼苗ꎬ每天定时更换Hoagland培养液ꎬ培养22dꎬ于4月12日将幼苗移至镉溶液和Hoagland培养液的复合处理液中让植株适应稳定ꎮ5月29日进行植物激素与螯合剂的复合添加处理ꎬ施用方式分为叶面喷施(P)和根部浇灌(J)两种ꎬ每一种添加方式分别对应三种植物激素(IAA㊁GA㊁SA)与螯合剂EDTA的复合处理ꎮ以浇施蒸馏水为空白对照ꎬ共设7个处理ꎬ每处理重复3次ꎬ共21组ꎮ不同复合处理设计见表1ꎮ㊀㊀表1㊀㊀㊀㊀试验设计处理配方CK蒸馏水叶面喷施PIAA100mg/LIAA+1.5mg/LEDTAPGA500mg/LGA+1.5mg/LEDTAPSA100mg/LSA+1.5mg/LEDTA根部浇灌JIAA100mg/LIAA+1.5mg/LEDTAJGA500mg/LGA+1.5mg/LEDTAJSA100mg/LSA+1.5mg/LEDTA1.4㊀测定指标及方法收获的籽粒苋植株用去离子水洗净后沥干水分ꎬ将根㊁茎㊁叶分开ꎬ105ħ杀青30minꎬ70ħ下烘至恒重ꎬ分别称其干重ꎮ采用火焰原子吸收光谱法测定植物样品Cd含量ꎮ转运系数=地上部Cd含量/根系Cd含量ꎻ251山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀富集系数=茎㊁叶㊁根中Cd含量/培养液中Cd含量ꎮ1.5㊀数据处理与分析所有数据均为3次重复平均值ꎮ利用Mi ̄crosoftExcel2010和DPS7.5统计软件进行数据分析及差异显著性检验ꎬ采用OriginPro9.1制图ꎮ2㊀结果与分析2.1㊀不同植物激素㊁螯合剂复合处理对籽粒苋生长的影响不同植物激素与螯合剂复合处理下ꎬ籽粒苋各器官生物量的方差分析和多重比较见表2ꎮ与对照相比ꎬ复合处理显著增加籽粒苋根㊁茎㊁叶的生物量(P<0.05)ꎬ且浇灌较喷施处理的效果更明显ꎮ叶部的生物量明显高于茎部㊁根部ꎮJSA复合处理籽粒苋根部生物量最大ꎬ为对照的3.28倍ꎻJGA复合处理下ꎬ茎㊁叶部的生物量最大ꎬ较对照分别高110.21%㊁42.59%ꎮ三种激素与螯合剂复合处理对籽粒苋根部生物量的影响表现为(SA+EDTA)>(GA+EDTA)>(IAA+EDTA)ꎬ对茎㊁叶部生物量的影响表现为(GA+EDTA)>(SA+EDTA)>(IAA+EDTA)ꎮ㊀㊀表2㊀不同复合处理下籽粒苋各部位生物量处理根部生物量/g茎部生物量/g叶部生物量/gCK0.103ʃ0.014g0.284ʃ0.018f0.587ʃ0.033gPIAA0.235ʃ0.020f0.467ʃ0.026e0.722ʃ0.031ePGA0.246ʃ0.027e0.588ʃ0.025b0.813ʃ0.030bPSA0.328ʃ0.018b0.484ʃ0.029d0.738ʃ0.024dJIAA0.257ʃ0.026d0.495ʃ0.030c0.713ʃ0.026fJGA0.269ʃ0.025c0.597ʃ0.029a0.837ʃ0.023aJSA0.338ʃ0.035a0.499ʃ0.031c0.746ʃ0.027c㊀㊀注:同列数据后不同小写字母表示处理间差异显著(P<0.05)ꎮ2.2㊀不同植物激素㊁螯合剂复合处理对籽粒苋吸收Cd的影响不同复合处理下籽粒苋根㊁茎㊁叶中的Cd含量见图1ꎮ与对照相比ꎬ复合处理显著增加籽粒苋根㊁茎㊁叶对Cd的吸收(P<0.05)ꎮ三种激素与螯合剂复合处理对根㊁茎㊁叶中Cd含量影响的整体表现为(GA+EDTA)>(SA+EDTA)>(IAA+ED ̄TA)ꎬ且根部浇灌的影响显著高于叶面喷施ꎮ叶部的Cd含量显著高于茎部㊁根部ꎮ不同复合处理中ꎬPIAA处理下根㊁茎㊁叶部的Cd含量最小ꎬ分别为对照的1.28㊁1.54㊁1.18倍ꎻJGA复合处理下ꎬ根㊁茎㊁叶部的Cd含量最大ꎬ分别为对照的1.60㊁2.73㊁1.23倍ꎮ表明浇灌GA和螯合剂使超富集植物籽粒苋幼苗各部对Cd的吸收进一步加强ꎬ修复效率进一步提高ꎮ同一部位柱上不同小写字母表示处理间差异显著(P<0.05)ꎬ下同ꎮ图1㊀不同复合处理下籽粒苋各部位Cd含量2.3㊀不同植物激素㊁螯合剂复合处理对籽粒苋转运Cd的影响不同植物激素与螯合剂复合处理下籽粒苋地上部对Cd的转运系数见图2ꎮ各复合处理下籽粒苋地上部的转运系数均大于20ꎬ显著高于对照ꎬ且浇灌方式较喷施效果更佳ꎮ不同复合处理对籽粒苋地上部转运Cd能力的影响表现为(GA+ED ̄TA)>(SA+EDTA)>(IAA+EDTA)ꎮ不同复合处理中ꎬPIAA处理下籽粒苋地上部对Cd的转运系数最低ꎬ为对照的1.31倍ꎻJGA处理下籽粒苋地上部对Cd的转运系数最高ꎬ为对照的1.68倍ꎮ说明浇灌GA和螯合剂显著提高了Cd从根部向地上部的转运能力ꎬ进而提高籽粒苋对Cd污染的修复效率ꎮ图2㊀不同复合处理下籽粒苋地上部对Cd的转运系数351㊀第11期㊀㊀㊀㊀郭梦露ꎬ等:植物激素㊁螯合剂复合处理对镉胁迫下籽粒苋生长和富集镉的影响2.4㊀不同植物激素、螯合剂复合处理对籽粒苋富集Cd的影响不同复合处理下籽粒苋根部和地上部对Cd的富集系数如图3所示ꎮ各复合处理下籽粒苋地上部的富集系数均大于1ꎬ显著高于对照ꎮ不同复合处理均增大了籽粒苋根部和地上部对Cd的富集系数ꎬ且浇灌处理相比于喷施效果更明显ꎮ不同复合处理对籽粒苋Cd富集系数的影响总体趋势表现为(GA+EDTA)>(SA+EDTA)>(IAA+EDTA)ꎮ籽粒苋地上部对Cd的富集系数显著高于根部ꎮ不同复合处理中ꎬPIAA处理的根部和地上部的Cd富集系数最小ꎬ分别为对照的1.42㊁1.57倍ꎻJGA处理下根部和地上部的Cd富集系数均最大ꎬ分别为对照的1.89㊁2.02倍ꎮ表明浇灌GA和螯合剂显著增强籽粒苋对Cd的富集能力ꎬ提高Cd从培养液向根及根向地上部的迁移ꎬ进而强化籽粒苋对Cd污染的修复效率ꎮ图3㊀不同复合处理下籽粒苋对Cd的富集系数3㊀讨论与结论众多学者开展关于采用绿色环保植物修复技术缓解重金属镉等对土壤和环境危害的研究时ꎬ多选用高生物量超富集植物及添加外源植物激素等措施增加修复效率[28-29]ꎮ王凯[30]在研究螯合剂及组合用于强化籽粒苋修复镉污染土壤效果中发现ꎬ复合螯合剂具有最高的镉移除量ꎬ并且对土壤中的一些酶如过氧化氢酶具有显著的促进作用ꎬ强化了籽粒苋的修复效率ꎮ肖艳辉等[31]采用盆栽试验ꎬ比较不同浓度胺鲜酯对籽粒苋生长及富集镉锌的影响ꎬ发现10mg/L胺鲜酯处理不仅能显著提高籽粒苋根际土壤有效态镉和锌含量ꎬ还能显著增加籽粒苋的生物量ꎻ施用胺鲜酯处理的籽粒苋叶㊁茎㊁根中镉含量均增加ꎮ已有研究表明赤霉素除具有调控植物生长发育的功能外ꎬ还可增强植物对重金属的耐受性并提高其对重金属的吸收量ꎬ赤霉素介导下植物对重金属的耐性机理主要包括植物生物量维持㊁抗氧化作用增强㊁光合系统修复㊁重金属区隔化和信号传递等[32]ꎮ1ˑ10-6mol/LGA使银胶菊(Partheniumhysteropho ̄rus)的Cd积累量和生物富集系数分别提高289%和128%[33]ꎻPb污染下施加1~10μmol/LGA可有效增加黑麦草(Loliumperenne)对Pb的富集[34]ꎻ质量浓度为10~1000mg/LGA可提高龙葵(Solanumnigrum)地上部对Cd的富集量[35]ꎮ本研究得出ꎬ植物激素和螯合剂复合处理显著增加籽粒苋的生物量ꎬ显著提高其对镉的吸收㊁转运和富集水平ꎮ这可能是植物激素和螯合剂共同作用的结果ꎬ既能提高植株的抗逆性ꎬ促进其生长发育ꎬ又能加强其对重金属离子的螯合力ꎬ提高重金属生物修复的有效性ꎮ从籽粒苋生物量来看ꎬ不同植物激素㊁螯合剂复合处理显著增加籽粒苋根㊁茎㊁叶部的生物量ꎬ根部浇灌方式效果明显优于叶面喷施ꎮJGA处理的籽粒苋茎㊁叶部生物量最大ꎻJSA处理的根部生物量最大ꎮ从籽粒苋对Cd的吸收量㊁转运系数㊁富集系数来看ꎬ不同植物激素㊁螯合剂复合处理对3个指标的影响均表现为(GA+EDTA)>(SA+EDTA)>(IAA+ED ̄TA)ꎬ显著高于对照ꎬ且根部浇灌处理效果显著高于叶面喷施处理ꎬ即浇灌GA与EDTA复合处理(JGA)下籽粒苋地上部对Cd的吸收量㊁转运系数㊁富集系数均最大ꎮ原因可能是由于EDTA对重金属离子具有较强的螯合力ꎬ在土壤中施用EDTA可促进土壤中的重金属成为可溶态ꎬ从而有效增加其生物利用率ꎻGA可以诱导细胞膜超级化ꎬ参与调节介导Cd转运的膜蛋白ꎬ促进Cd通过细胞膜进入植物体ꎬ增加植物细胞对Cd的富集ꎬ但相关机制有待进一步研究ꎮ本试验为提高植物对重金属污染土壤的修复效率提供了方法ꎬ为研究植物激素与螯合剂效应提供了一定的理论和实践依据ꎮ但外源施加激素㊁螯合剂的种类㊁浓度㊁持续时间㊁喷施方式等众多因素对植物的生理活动均会造成不同影响ꎬ这451山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀种影响程度仍需进一步研究ꎻ植物激素㊁螯合剂提高植物耐受重金属的相关机制需要更深入的研究ꎻ重金属胁迫下多种植物激素㊁螯合剂的调控功能和机制研究需要进一步完善ꎮ参㊀考㊀文㊀献:[1]㊀环境保护部ꎬ国土资源部.全国土壤污染状况调查公报[R].北京:环境保护部ꎬ2014:1-5.[2]㊀HaoQJꎬJiangCS.HeavymetalconcentrationsinsoilsandplantsinRongxiManganeseMineofChongqingꎬSouthwestofChina[J].ActaEcologicaSinicaꎬ2015ꎬ35(1):46-51. [3]㊀徐剑锋ꎬ王雷ꎬ熊瑛ꎬ等.土壤重金属污染强化植物修复技术研究进展[J].环境工程技术学报ꎬ2017ꎬ7(3):366-373. [4]㊀游来勇ꎬ王昌全ꎬ罗娟ꎬ等.Cd胁迫对水稻生长及内源激素含量的影响[J].生态环境学报ꎬ2015ꎬ24(5):860-865. [5]㊀张娜.褪黑素处理对渗透胁迫下黄瓜种子萌发及幼苗生长的影响及其分子机制[D].北京:中国农业大学ꎬ2014. [6]㊀ChagasJKMꎬdeFigueiredoCCꎬdaSilvaJꎬetal.Theresid ̄ualeffectofsewagesludgebiocharonsoilavailabilityandbio ̄accumulationofheavymetals:evidencefromathree ̄yearfieldexperiment[J].JournalofEnvironmentalManagementꎬ2021ꎬ279(24):111-118.[7]㊀DiaoZHꎬQianWꎬZhangZWꎬetal.RemovalsofCr(VI)andCd(II)byanovelnanoscalezerovalentiron/peroxydisul ̄fateprocessanditsFenton ̄likeoxidationofpesticideatrazine:coexistingeffectꎬproductsandmechanism[J].ChemicalEngi ̄neeringJournalꎬ2020ꎬ397(8):125-129.[8]㊀DiaoZHꎬYanLꎬDongFXꎬetal.Degradationof2ꎬ4 ̄di ̄chlorophenolbyanovelironbasedsystemanditssynergismwithCd(II)immobilizationinacontaminatedsoil[J].Chemi ̄calEngineeringJournalꎬ2020ꎬ37(9):122-127. [9]㊀DiaoZHꎬDongFXꎬYanLꎬetal.SynergisticoxidationofBi ̄sphenolAinaheterogeneousultrasound ̄enhancedsludgebio ̄charcatalyst/persulfateprocess:reactivityandmechanism[J].JournalofHazardousMaterialsꎬ2020ꎬ38(4):121-126. [10]DiaoZHꎬDuJJꎬJiangDꎬetal.Insightsintothesimultane ̄ousremovalofCr6+andPb2+byanovelsewagesludge ̄derivedbiocharimmobilizednanoscalezerovalentiron:coexistenceeffectsandmechanism[J].ScienceoftheTotalEnvironmentꎬ2018ꎬ64(2):505-515.[11]DiaoZHꎬYanLꎬDongFXꎬetal.Ultrasound ̄assistedcata ̄lyticreductionofCr(VI)byanacidminedrainagebasedoncouplingwithFeS2systemfromaqueoussolutions:performanceandmechanism[J].JournalofEnvironmentalManagementꎬ2021ꎬ27(8):111-120.[12]DongFXꎬYanLꎬZhouXHꎬetal.SimultaneousadsorptionofCr(Ⅵ)andphenolbybiochar ̄basedironoxidecompositesinwater:performanceꎬkineticsandmechanism[J].JournalofHazardousMaterialsꎬ2021ꎬ41(6):125-130.[13]李韦瑶ꎬ肖汇川ꎬ张碧莹ꎬ等.不同激素对山韭种子萌发特性的影响[J].种子ꎬ2022ꎬ41(6):76-83.[14]戴陶宇ꎬ王前程ꎬ张迎迎ꎬ等.外源赤霉素对盐胁迫下番茄种子萌发和幼苗生长的影响[J].种子ꎬ2022ꎬ41(3):74-80.[15]张盛楠ꎬ黄益宗ꎬ李颜ꎬ等.Cd胁迫下不同外源植物激素对水稻幼苗抗氧化系统及Cd吸收积累的影响[J].环境科学2021ꎬ42(4):2040-2046.[16]WangHLꎬTianCYꎬJiangLꎬetal.Remediationofheavymetalscontaminatedsalinesoils:ahalophytechoice?[J].EnvironmentalScience&Technologyꎬ2014ꎬ48(1):21-22. [17]BabaeianEꎬHomaeeMꎬRahnemaieR.Chelate ̄enhancedphy ̄toextractionandphytostabilizationoflead ̄contaminatedsoilsbycarrot(Daucuscarota)[J].ArchivesofAgronomy&SoilSci ̄enceꎬ2016ꎬ62(3):339-358.[18]HuYHꎬWeiSHꎬZhouQXꎬetal.Applicationofchelatorinphytoremediationofheavymetalscontaminatedsoils:areview[J].JournalofAgro ̄EnvironmentScienceꎬ2010ꎬ29(11):2055-2063.[19]YanLꎬLiCLꎬZhangJJꎬetal.EnhancedphytoextractionofleadfromartificiallycontaminatedsoilbyMirabilisjalapawithchelatingagents[J].BulletinofEnvironmentalContaminationandToxicologyꎬ2017ꎬ99(2):208-212.[20]陈柯罕ꎬ张科ꎬ李取生ꎬ等.四种盐生植物对CdPb复合污染提取修复效果比较研究[J].农业环境科学学报ꎬ2017ꎬ36(3):458-465.[21]WangHꎬSunLNꎬLiHBꎬetal.EffectofdifferentchelatorsapplicationonCdaccumulationinmetalpollutedsoilsbyBetavulgarisvar.ciclaL.[J].EcologyandEnvironmentꎬ2008ꎬ17(6):2249-2252.[22]WangYHꎬChenDYꎬJiangZYꎬetal.PhytoremediationofthesoilcontaminatedbyPbꎬCdandsecondarysalinizationwiththeenhancementofEDTA[J].JournalofAgro ̄Environ ̄mentScienceꎬ2018ꎬ37(9):1866-1874.[23]龙玉梅ꎬ刘杰ꎬ傅校锋ꎬ等.4种Cd超富集/富集植物修复性能的比较[J].江苏农业科学ꎬ2019ꎬ47(8):296-300. [24]JinHJꎬXuMJꎬChenHꎬetal.Comparativeproteomicanaly ̄sisofdifferentiallyexpressedproteinsinAmaranthushybridusL.rootsundercadmiumstress[J].WaterꎬAirandSoilPollu ̄tionꎬ2016ꎬ227(7):220-225.[25]谢运河ꎬ纪雄辉ꎬ吴家梅ꎬ等.镉砷污染土壤 三高 富集植物筛选与修复成本分析[J].环境科学与技术ꎬ2020ꎬ43(1):116-121.[26]何明倩ꎬ卢欣ꎬ唐研ꎬ等.冬氨酸二丁二酸醚和酒石酸配施对籽粒苋生长和镉富集特征的影响[J].四川农业大学学报ꎬ2021ꎬ39(6):827-834.[27]张燕ꎬ葛燚ꎬ刘晓月ꎬ等.施用螯合剂对重金属污染土壤中籽粒苋镉吸收转运的影响[J].湖南农业科学ꎬ2021ꎬ21(11):46-50.[28]张欣ꎬ范仲学ꎬ郭笃发ꎬ等.外源物质对农作物镉毒害的缓551㊀第11期㊀㊀㊀㊀郭梦露ꎬ等:植物激素㊁螯合剂复合处理对镉胁迫下籽粒苋生长和富集镉的影响解效应[J].山东农业科学ꎬ2011(1):68-72.[29]周凯ꎬ张苏明ꎬ孙梦媛ꎬ等.东南景天对胁迫条件下重金属锌和镉吸收积累特性的研究[J].山东农业科学ꎬ2021ꎬ53(8):90-93.[30]王凯.复合螯合剂强化籽粒苋修复Cd污染土壤效果研究[D].武汉:华中农业大学ꎬ2019.[31]肖艳辉ꎬ李应文ꎬ邹碧ꎬ等.不同浓度胺鲜酯对籽粒苋富集重金属镉锌的影响[J].生态环境学报ꎬ2019ꎬ28(12):2433-2437.[32]CamaraMCꎬVandenbergheLPSꎬRodriguesCꎬetal.Cur ̄rentadvancesingibberellicacid(GA3)productionꎬpatentedtechnologiesandpotentialapplications[J].Plantaꎬ2018ꎬ248(5):1049-1062.[33]AliNꎬHadiF.Phytoremediationofcadmiumimprovedwiththehighproductionofendogenousphenolicsandfreeprolinecon ̄tentsinPartheniumhysterophorusplanttreatedexogenouslywithplantgrowthregulatorandchelatingagent[J].EnvironmentalScienceandPollutionResearchꎬ2015ꎬ22(17):13305-13318. [34]HeSYꎬHeZLꎬWuQLꎬetal.EffectsofGA3onplantphys ̄iologicalpropertiesꎬextractionꎬsubcellulardistributionandchemicalformsofPbinLoliumperenne[J].InternationalJour ̄nalofPhytoremediationꎬ2015ꎬ17(12):1153-1159. [35]JiPHꎬTangXWꎬJiangYJꎬetal.Potentialofgibberellicacid3(GA3)forenhancingthephytoremediationefficiencyofSolanumnigrumL.[J].BulletinofEnvironmentalContamina ̄tionandToxicologyꎬ2015ꎬ95(6):810-814.651山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀。
镉对植物的毒害及植物解毒机制研究进展

杨红霞ꎬ陈俊良ꎬ刘㊀崴.镉对植物的毒害及植物解毒机制研究进展[J].江苏农业科学ꎬ2019ꎬ47(2):1-8.doi:10.15889/j.issn.1002-1302.2019.02.001镉对植物的毒害及植物解毒机制研究进展杨红霞ꎬ陈俊良ꎬ刘崴(国家地质实验测试中心ꎬ北京100037)㊀㊀摘要:综述近年来镉对植物的毒害及植物解毒机制的相关研究成果ꎮ主要从植物生长㊁光合作用㊁植物酶活性㊁植物细胞分裂等方面阐述镉的毒害机制ꎬ并从植物对镉的吸收和转运㊁耐受机制㊁影响因素等方面讨论植物的解毒机制ꎮ㊀㊀关键词:镉ꎻ毒害ꎻ解毒ꎻ机制㊀㊀中图分类号:Q948.12ꎻX173㊀㊀文献标志码:A㊀㊀文章编号:1002-1302(2019)02-0001-08收稿日期:2017-09-22基金项目:国家自然科学基金(编号:41301566)ꎻ中国地质科学院基本科研业务费项目(编号:YYWF201620)ꎻ国土资源部公益性行业科研专项(编号:200911043-26)ꎻ中国地质大调查项目(编号:12120113015400)ꎮ作者简介:杨红霞(1979 )ꎬ女ꎬ山东济南人ꎬ博士ꎬ教授级高级工程师ꎬ硕士生导师ꎬ主要从事环境地球化学及元素形态分析研究ꎮE-mail:yanghongxia1@sina.comꎮ㊀㊀镉(Cd)是一种生物毒性很强的重金属ꎬ是植物生长㊁发育的非必需元素ꎬ当植物体中Cd积累到一定程度时ꎬ就会表现出各种毒害症状ꎬ如叶片枯黄㊁茎秆缩短㊁侧根减少㊁叶绿素含量降低㊁抗氧化酶活性降低㊁细胞膜通透性发生改变ꎬ导致细胞损伤ꎬ从而对植物的生长产生抑制作用[1-2]ꎮ当Cd达到一定的浓度时ꎬ不仅影响植物的正常生长ꎬ而且影响植物的生理活动ꎬ主要表现为影响植物的光合作用㊁植物酶活性㊁植物细胞分裂及膜活性㊁呼吸作用㊁蒸腾作用等ꎮ通常植株表现为发育缓慢㊁植株矮小㊁叶片失绿等症状ꎮ某些植物对镉产生超累积作用ꎬ在重金属镉胁迫下ꎬ有一系列的耐性和解毒机制[3-6]ꎮ目前研究表明ꎬ植物的耐Cd机制主要包括螯合作用㊁区隔化作用㊁㊁抗氧化作用㊁排外作用㊁固定钝化作用㊁应激作用ꎮ本文着重讨论了镉对植物产生的毒害及植物对镉吸收和耐性机制的研究进展ꎬ从而为植物抗镉性机制㊁防治镉污染提供理论依据ꎬ其超积累的生理生化机制对于Cd污染环境的植物修复研究具有重要意义ꎮ1 植物受镉的毒害镉对植物的毒害表现为2个方面:其一ꎬ镉对植物生长的影响ꎻ其二ꎬ镉对植物生理活动的影响ꎮ一般表现为植物细胞及整个植株的生长受到抑制㊁根系活力降低㊁幼苗生长缓慢㊁叶片缩卷甚至枯黄等ꎮ镉影响植物的生理特性主要表现为影响植物的光合作用㊁抑制植物保护酶的活性㊁细胞膜通透性改变㊁硝酸还原酶活性下降等ꎮ1.1㊀镉对植物生长的影响㊀㊀Cd对植物生长的影响主要表现出 低促高抑 现象ꎮ而Cd对植物生长毒害的临界浓度随着不同土壤类型ꎬ不同植物种类而改变ꎮ何俊渝等研究结果表明ꎬ当Cd2+浓度为5~15mg/L时ꎬCd2+可以促进玉米种子的萌发ꎬ提高玉米的发芽势和发芽率ꎬ促进玉米芽与幼根的生长ꎻ当Cd2+的浓度高于50mg/L时ꎬ明显抑制了种子萌发和幼苗生长[7]ꎮ王明新等通过试验发现ꎬ随着营养液Cd浓度的增加ꎬ叶片光合色素含量呈先上升后下降趋势ꎬ丙二醛含量则呈线性递增趋势ꎬ高浓度Cd处理(ȡ0.1mmol/L)对孔雀草生长产生了显著的胁迫响应[8]ꎮ郭智等研究发现ꎬ镉胁迫下龙葵幼苗地上部和根系生长均受到严重抑制ꎬ其中根系受抑制程度更严重[9]ꎮ宋阿琳等研究表明ꎬCd胁迫下ꎬ小白菜体内Cd含量和累积量随处理浓度的上升而显著增加ꎬ而且根系中的Cd含量显著高于地上部[10]ꎮCd对植物根系的毒害主要通过加速根系细胞的老化ꎬ影响细胞分裂ꎬ从而改变RNA合成ꎬ抑制硝酸还原酶活性ꎬ减少根部对硝酸盐和其他养分的吸收[11]ꎮCd对植物叶片发育的影响ꎬ主要表现为叶片光合作用降低ꎬ叶绿体含量减少ꎬ叶片缩卷ꎬ严重时甚至发黄ꎮ当Cd2+浓度高时ꎬ叶绿体内基粒片层发生肿胀或结构紊乱ꎬ线粒体内膜脊肿胀或消失ꎬ细胞核核膜消失ꎬ从而导致植物生长受到严重影响ꎮ张利红等通过试验发现ꎬ在Cd浓度为0.05㊁0.10㊁0.25㊁0.50mmol/L的水培处理中ꎬ根系活力表现为先升高后降低的变化趋势ꎬ小麦生长受到一定程度的抑制ꎬ并随溶液中Cd浓度的增加ꎬ抑制加重[12]ꎮ张丽等通过盆栽试验发现ꎬ随着土壤中重金属镉溶液处理浓度的增大ꎬ润草1号叶绿素和游离脯氨酸的含量均表现为先升高后降低ꎬ而根系活力则逐渐降低ꎬ细胞膜透性逐渐升高[13]ꎮ毕君等研究表明ꎬ虽然羽衣甘蓝㊁华北景天㊁木槿地上部分的镉含量大于10mg/kgꎬ但其生长却未出现异常ꎬ故此3种植物为镉富集能力较强的植物ꎻ夹竹桃㊁木槿㊁紫穗槐根系的含镉量均大于50mg/kgꎬ这对于污染土壤中Cd离子的固定和清除能起到一定的作用[14]ꎮ1.2㊀镉对植物生理活动的影响重金属对植物生长产生的不利影响可通过植物的一些生理活动及其生理生化特征反映出来ꎮ1.2.1㊀对植物光合作用的影响㊀植物的光合作用不仅能合成有机物ꎬ而且能释放大量氧气净化空气ꎮ而Cd对光合作用影响的机制目前还没有一致的看法ꎮ有报道ꎬCd是通过影响叶绿素的生物合成ꎬ破坏叶绿体的结构来影响植物的光合作用ꎮ张利红等通过水培试验发现ꎬ叶绿素含量随Cd浓度的增加而降低[12]ꎮ在过量Cd处理下ꎬ会影响植物体内叶绿素的生物合成ꎬ使叶片叶绿素含量下降[15]ꎮ陈良等通过水培1 江苏农业科学㊀2019年第47卷第2期试验表明ꎬ在Cd胁迫下ꎬ南芋5号(NY5)和南芋2号2种菊芋幼苗净光合速率(Pn)㊁气孔导度(Gs)㊁蒸腾速率(Tr)均明显下降[16]ꎮ滕振宁等研究结果表明ꎬ镉处理后ꎬ敏感的水稻品种YZX和相对耐受的品种XY-12的净光合速率均降低ꎻ镉对2种水稻品种光响应曲线参数的影响存在差异ꎬ其内禀量子效率㊁最大净光合速率㊁光饱和点与不添加镉的对照相比均有所减小ꎬ而镉处理后水稻剑叶光补偿点却大大增加[17]ꎮ张媛华研究表明ꎬCd显著降低了绿豆幼苗的根长㊁根数㊁株高及生物量ꎬ不仅降低了植物的光合速率ꎬ还降低了幼苗茎叶中微量矿质元素铁(Fe)㊁锰(Mn)㊁锌(Zn)㊁铜(Cu)的含量ꎬ促进根中Zn㊁Fe㊁Cu的积累ꎬ这说明Cd可以通过改变植物对微量元素的积累影响植物的生长发育和光合作用[18]ꎮ张永平等试验发现ꎬ镉胁迫下ꎬ哈密绿和秀绿这2个甜瓜品种幼苗生长量㊁净光合速率㊁气孔导度㊁蒸腾速率和气孔限制值(Ls)均降低ꎬ且镉浓度越高ꎬ下降幅度越大ꎬ胞间CO2浓度(Ci)随着镉浓度的增加而升高ꎻ哈密绿幼苗生长和光合作用受镉胁迫的影响小于秀绿[19]ꎮ1.2.2㊀对植物保护酶系统的影响㊀Cd对植物的毒害首先是作用于细胞内的氧化还原系统ꎬ进而引起一系列氧化还原反应ꎬ使抗氧化系统发生改变ꎬ且Cd易与细胞膜上蛋白体结合进而破坏酶系统[20]ꎮ植物体内各种抗氧化酶活性变化比较复杂ꎬ同时保护酶系统也用来保护植物免受氧化伤害ꎮ植物保护酶系统的组成主要有超氧化物歧化酶(SOD)㊁过氧化物酶(POD)㊁过氧化氢酶(CAT)ꎮ张利红等通过试验表明ꎬ超氧化物歧化酶㊁过氧化物酶㊁丙二醛含量和细胞膜透性随Cd浓度的增加而增加[12]ꎮ黄辉等进行了水平玉米幼苗试验ꎬ结果显示在高浓度镉胁迫下幼苗生长几乎停止ꎻ根部受到氧化损伤ꎬSOD㊁CAT活性升高ꎬPOD㊁抗坏血酸过氧化物酶(ASA-POD)被明显抑制ꎬ叶片中CAT㊁POD及ASA-POD活性显著升高[21]ꎮ当土壤镉含量小于22.3mg/kg时ꎬ桑树POD㊁SOD活性无明显变化ꎻ当其含量超过22.3mg/kg时ꎬSOD活性降低ꎻ当Cd含量超过145mg/kg时ꎬPOD活性才表现出明显的降低趋势[22]ꎮ此外ꎬCd还抑制固氮酶㊁脱氧核酸酶㊁核糖核酸酶㊁根系脱氢酶㊁淀粉酶㊁硝酸还原酶㊁多酚氧化酶㊁蛋白酶㊁抗坏血酸过氧化酶㊁乳酸脱氢酶等多种酶的活性[23]ꎮCd通过影响这些酶的活性ꎬ进而影响植物的碳水化合物代谢㊁氮代谢㊁核酸代谢㊁呼吸作用等多种代谢活动ꎮ1.2.3㊀对植物细胞的影响㊀杨居荣发现在Cd胁迫条件下ꎬ烟草㊁莴苣㊁甜菜㊁胡萝卜㊁紫花苜蓿5种植物细胞的生长受到了明显的抑制[24]ꎮ任安芝等用Cd处理青菜ꎬ随着Cd浓度的提高ꎬ细胞膜的透性增强ꎬ膜脂过氧化程度提高[25]ꎮ植物叶片细胞膜受到伤害后ꎬ细胞内的大量离子和有机物外渗ꎬ同时外界有害有毒的物质也容易进入植物细胞体内ꎬ从而导致植物体生理活动失调[26]ꎮ方继宇等研究表明ꎬCd在马缨丹根和叶细胞可溶性组分中的分配比例最高ꎬ分别占62.1%~54.2%和59.8%~52.6%ꎬ其次在细胞壁中ꎬ分别占23.8%~34.4%和28.7%~39.5%ꎬ在细胞器和细胞膜中的分配比例较低ꎬ表明镉在马缨丹根和叶细胞中的主要分布位点是可溶性组分和细胞壁ꎬ细胞壁固持和液泡区隔化可能是马缨丹对镉的重要解毒和耐性机制之一ꎬ对保持马缨丹的细胞完整性具有重要意义[27]ꎮ随着生物技术的发展ꎬ对细胞的超微结构的研究越来越多ꎮ在低Cd浓度处理下ꎬ叶绿体基粒和线粒体内腔峭突减少ꎻ在高Cd浓度胁迫下ꎬ叶绿体垛叠无规律ꎬ线粒体肿胀或消失[28-29]ꎮ1.2.4㊀对其他生理特征的影响㊀另外ꎬCd胁迫还会造成植物其他生理特征的一些反应ꎬ如通过降低氮素吸收和硝酸还原酶活性来改变氨基酸组成㊁阻碍蛋白质合成以及加速蛋白质的分解ꎮVallee等研究表明ꎬCd使硝酸还原酶活性下降[30]ꎮ杨金凤等研究表明ꎬ随着Cd浓度的增加ꎬ油菜体内硝酸还原酶活性及根系活力降低ꎬ油菜根内游离脯氨酸含量逐渐增加[31]ꎮ陈愚等研究结果表明ꎬ一定浓度的镉能够改变沉水植物的硝酸还原酶活性ꎬ抑制超氧化物歧化酶活性ꎬ从而破坏其抗氧化防御系统[32]ꎮ赵天宏等试验结果表明ꎬ与硝态氮相比ꎬCd胁迫下ꎬ铵态氮能够显著提高小麦地上部干质量㊁氮素积累量和小麦叶片的叶绿素含量ꎻ而当施氮量在150~300kg/hm2范围内ꎬ硝态氮对小麦叶片硝酸还原酶活性和可溶性蛋白含量的促进作用高于铵态氮[33]ꎮ2 植物对镉的吸收及运输植物的正常生长除需要氮(N)㊁磷(P)㊁钾(K)㊁硫(S)㊁钙(Ca)㊁镁(Mg)这些元素外ꎬ还需要铁(Fe)㊁锌(Zn)㊁锰(Mn)㊁镍(Ni)㊁铜(Cu)和钼(Mo)等微量元素ꎮ植物对金属离子的吸收㊁运输㊁储存有其对应的机制ꎬ金属的吸收和运输通过细胞膜上的转运蛋白来进行ꎬ并且植物有着敏感的机制ꎬ使细胞内金属离子浓度在植物的生理活动范围内[34-35]ꎮ2.1㊀吸收机制土壤中重金属的有效态与植物生长过程密切相关[36]ꎮ植物对金属的吸收方式有以下2种:(1)通过根系分泌物螯合重金属ꎮ例如ꎬ根系分泌物的羧基具有酸性ꎬ与重金属结合是一种潜在的增加重金属有效性的途径[37-38]ꎮ(2)利用金属的流动性ꎮ金属离子先通过植物根毛细胞膜上的通道进入ꎬ再通过细胞膜的选择通透性以共质体途径进入植物根的表皮细胞[39-40]ꎮ植物根系吸收重金属后向木质部运输ꎮ大多数的金属离子都是进入到木质部ꎬ少数金属离子进入到韧皮部ꎮZhao等研究表明ꎬCd超累积植物天蓝遏蓝菜(Thlaspicaerulescens)根细胞质膜上有高选择性的Cd转运系统ꎬ对Cd吸收性较强[41]ꎮ植物体对重金属转运还会受外部环境的影响ꎮ如Salt等报道ꎬ有机酸参与印度芥菜木质部中Cd的运输ꎬ根系分泌的有机酸也参与了部分植物对Cd的吸收㊁运输[42]ꎮClemens研究表明ꎬ镉作为植物的非必需元素ꎬ可通过钙㊁铁㊁锰㊁锌等必需元素的吸收通道进入植物体内[43]ꎮ杨菲等研究了水稻对镉的吸收情况ꎬ结果表明ꎬ镉既可以以Cd-谷胱甘肽(GSH)㊁Cd-植物螯合肽(PC)等螯合形式ꎬ也可以以自由离子Cd2+的形式进入根细胞内[44]ꎮ2.2㊀影响因素㊀㊀影响植物对重金属Cd的吸收有以下6个因素:(1)植物的种类ꎮ植物对重金属的选择性吸收与植物种类的特性有关ꎬ根据不同物种对重金属的选择吸收和累积生物量的不同ꎬ来判断是否可以作为该种重金属的超累积植物ꎮ(2)植物生长条件ꎮ不同生长环境和种植条件会影响植物对重金属的吸收ꎬ比如pH值㊁施用肥料品种㊁添加螯合剂的不同ꎬ都会影响到植物对重金属的吸收ꎮ(3)植物根际ꎮ植物根系能吸收各2 江苏农业科学㊀2019年第47卷第2期种重金属元素ꎬ并能进行存储和代谢ꎬ还能通过改变根系粗细和长短来适应重金属的胁迫[45]ꎮ植物根际分泌物能够活化土壤中的某些重金属ꎬ从而增加植物的吸收ꎮ(4)环境因素ꎮ植物对重金属的吸收会随着土壤温度㊁水分㊁光照等的变化而改变ꎮ(5)重金属的有效性ꎮ不同土壤的pH值㊁Eh(土壤氧化还原电位)㊁植物根系分泌物㊁土壤的有机质含量等因素都会影响重金属的有效性ꎬ从而使植物可以选择性地吸收不同形态的重金属ꎮ(6)重金属离子间的相互影响ꎮ不同种重金属离子间存在相互竞争的附着位点ꎬ这会改变植物对重金属离子的吸收能力ꎬ当某一离子浓度过高时ꎬ植物对其他离子的选择性吸收将产生影响[46]ꎮ2.3㊀累积及运输能够累积和运输Cd的植物主要具备以下几个特点:(l)植物根系具有较强的耐Cd能力和吸收能力ꎻ(2)根系吸收Cd后通过蒸腾作用能够有效地运输到植物的地上部ꎻ(3)叶片能对Cd区隔化ꎬ将大多数的Cd离子储存在液泡中[6]ꎮ细胞膜上有运输重金属离子的转运蛋白及高亲和力结合位点[47-48]ꎮ为了让植物更多地吸收重金属ꎬ达到植物修复效果ꎬ研究者须清楚地理解重金属的运输受哪些基因调控ꎮ张玉秀等研究表明ꎬ土壤微环境会使植物对Cd2+的吸收产生影响ꎻ植物根细胞壁会选择性吸收㊁吸附和固定土壤中的Cd2+ꎬ其中大部分Cd2+被截留在细胞壁中ꎬ其余的则通过主动运输或协助扩散等方式透过细胞膜进入根细胞中ꎻ部分Cd在根细胞液泡中积累ꎬ部分通过木质部运输到地上部分ꎻ茎叶部的大部分Cd2+则通过络合作用被固定在液泡中ꎬ仅少量在细胞壁和细胞质中被截留[35]ꎮ与植物累积运输Cd相关的基因有以下几类: (1)锌铁调控蛋白(ZRTꎬIRT-likeproteinꎬ简称ZIP)家族的基因ꎮ当植物对Fe离子的吸收供不应求时ꎬ植物就会吸收更多的Cd离子[49-50]ꎮEide等对拟南芥的研究发现ꎬ当缺少Fe时ꎬ植物转运蛋白Zip家族有关运输的基因AtIRT1会优先吸收Fe2+到根表皮ꎬ同时运输Mn2+㊁Zn2+㊁Cd2+等二价态金属离子[51]ꎮ另有报道称ꎬZip家族的许多成员都参与了上述4种离子的运输[52]ꎮ(2)阳离子扩散促进剂(cationdiffusionfacilitatorꎬ简称CDF)家族蛋白的基因ꎮCDF是另一类转运运输蛋白的催化剂ꎬ其主要作用是促进Zn2+的运输ꎻ另外ꎬ对Co2+㊁Fe2+㊁Cd2+㊁Ni2+㊁Mn2+的运输也起到了一定的作用[53-54]ꎮ(3)天然抗性相关巨噬细胞蛋白(naturalresistance-associatedmacrophageprotein1ꎬ简称NRAMP1) (一种膜内在蛋白)的基因ꎮNRAMP1基因在拟南芥中与Fe2+㊁Mn2+㊁Cd2+的运输有关[55-56]ꎮ(4)CPX型重金属ATP酶(Cpx-typeheavymetalATPases)的基因ꎮ已证实大量有机体内有该基因ꎬ且该基因也是Cu㊁Zn㊁Cd㊁Pb等重金属横跨细胞膜运输必不可少的调控基因[57]ꎮ㊀㊀重金属Cd被植物吸收后ꎬ大部分富集在根部ꎬ少量迁移到地上部ꎬ通常Cd在植物体内积累的含量从大到小的顺序是根>茎>叶>子实ꎮ有报道ꎬ印度芥菜在Cd浓度为0.1mg/L的溶液中时ꎬ在地上部植株干质量中Cd的浓度达到110mg/kgꎬ而在根浓度干质量中Cd的浓度达到670mg/kgꎮ这主要是因为Cd在印度芥菜根系中能快速地螯合ꎬ形成Cd-S4的混合物ꎬ达到Cd的累积效果ꎮ在印度芥菜根系中ꎬCd的运输主要靠木质部ꎬ而在植株地上部Cd的运输则是靠植物的蒸腾作用[58]ꎮ这表明印度芥菜具有在根部截留重金属的能力ꎬ从而避免较高浓度的重金属进入地上部分ꎬ产生更为严重的生理毒害作用[59]ꎮ苏徳纯等研究发现ꎬ印度芥菜能够活化㊁吸收石灰性土壤中的难溶态Cdꎬ随着土壤中CdCO3加入量的增加ꎬ印度芥菜根系和地上部中的Cd含量显著增加ꎬ印度芥菜吸收的Cd有71%~82%在地上部累积[60]ꎮ孙涛等研究发现ꎬ在土壤重金属含量较低的情况下ꎬ大多数重金属离子先在印度芥菜根部富集ꎬ而只有少数运输到地上部分ꎬ这可能表明印度芥菜在根部具有截留重金属的能力ꎬ从而避免高浓度重金属进入地上部分产生更严重的生理毒害作用[59]ꎮ3 植物耐镉机制在高浓度Cd的胁迫下ꎬ有些植物的生长发育基本不受影响ꎻ而在轻微的重金属污染条件下ꎬ有些植物就停止生长发育ꎬ这表明植物对重金属具有某种抗性机制ꎮ植物在重金属胁迫下ꎬ有一系列的耐性和解毒机制[4-5ꎬ61]ꎮ有的植物是通过阻止重金属进入根系来耐Cdꎬ还有些植物通过地上部对Cd的富集来耐Cd[62]ꎮ目前研究表明ꎬ植物的耐Cd机制主要包括以下几个方面:螯合作用㊁区隔化作用㊁抗氧化作用㊁排外作用㊁固定钝化作用㊁应急作用ꎮ其中ꎬ植物的区隔化作用和螯合作用占主导地位ꎬ虽然大多数学者对其在重金属的耐受机制中的作用进行了广泛的研究ꎬ但关于多变复杂的螯合肽的研究至今未达到共识ꎮ3.1㊀螯合作用螯合作用主要是指重金属离子进入植物体内后ꎬ植物体内的金属硫蛋白(MTs)㊁植物螯合肽(PCs)㊁谷胱甘肽(GSH)㊁柠檬酸㊁草酸㊁苹果酸等能和重金属离子发生螯合作用ꎬ形成稳定的螯合物ꎬ从而降低重金属毒性或解毒作用[63]ꎮ重金属的解毒机制一般是配合基通过螯合作用与重金属离子形成配合物ꎬ然后储存于植物液泡中[64]ꎮ不同的螯合肽物质在重金属解毒机制中的作用不同ꎮ重金属通过细胞膜一般须要被配体螯合ꎬ以此减少与细胞复合物不必要的联系ꎮ与重金属非超积累物种相比ꎬ重金属超积累物种中具有更高的植物螯合肽积累的能力ꎬ这表明植物螯合肽在重金属隔离㊁重金属解毒和重金属排除等方面起着重要作用[65]ꎮ3.1.1㊀谷胱甘肽㊀谷胱甘肽是一种非巯基蛋白ꎬ在细胞(线粒体㊁叶绿体㊁液泡㊁细胞质)中大量存在ꎻ由于结构特殊ꎬ谷胱甘肽在有机体中能发挥较大的功能ꎬ在细胞的新陈代谢过程中起主要作用[66]ꎮ谷胱甘肽的高稳定性和亲水性在螯合重金属㊁解毒外来污染物的过程中起防御作用[67-69]ꎮ在重金属胁迫下ꎬ谷胱甘肽能结合重金属离子形成复合物ꎬ而结合重金属的离子主要受谷胱甘肽转移酶支配ꎬ形成的这种复合物会被运输到植物叶片的液泡中ꎬ从而保护植物细胞ꎬ防止其受害[70]ꎮ单长卷等研究表明ꎬ冰草能够通过增强叶片中参与抗坏血酸㊁谷胱甘肽的合成和循环代谢的酶的活性ꎬ维持植物体内谷胱甘肽㊁抗坏血酸水平及其氧化还原状态ꎬ从而抵御干旱造成的氧化胁迫[71]ꎮ谷胱甘肽生物合成物质的增加可增强植物对Cd的耐性ꎬ并促进Cd在一些植物地上部的积累ꎬ从而达到植物修复的目的[72]ꎮ与前述观点不同的是ꎬCobbett3江苏农业科学㊀2019年第47卷第2期等对拟南芥进行试验发现ꎬGSH的含量与植物耐重金属有较强的关系ꎬ但并不是意味着GHS含量越高ꎬ对重金属的耐性越强[73]ꎮ植物可能在耐重金属胁迫时ꎬ并非仅GSH单独作用ꎬ而是有复杂的机制[74]ꎮ由于谷胱甘肽金属配合物运输机制的研究鲜有报道ꎬ因此该方面有待于进一步了解ꎮ3.1.2㊀金属硫蛋白㊀金属硫蛋白分子量低ꎬ富含半胱氨酸的小分子蛋白质ꎬ能通过一连串的巯基来络合重金属ꎬ存在于绝大多数真核生物中[75]ꎮ在重金属胁迫下ꎬ和其他螯合物相比ꎬ金属硫蛋白在生物解毒方面发挥着重要作用ꎬ并且许多研究已证实其为重金属胁迫的生物指示者[76-78]ꎮ植物金属硫蛋白中半胱氨酸(Cys)存在形式有Cys-x-CysꎬCys-x-x-CysꎬCys-Cys(串)ꎬx代表Cys以外任何1种氨基酸ꎻ植物MTs分为6个亚家族:P1㊁P2㊁p2vꎬp3ꎬpec㊁p21[79]ꎮ金属硫蛋白最早在动物组织内被发现ꎬ动物接触Cd后在肝脏合成Cd的金属硫蛋白ꎬ汇集于肾脏ꎬ形成尿镉排出[80-81]ꎮ张海燕等将大蒜的AsMT2b基因转到拟南芥中表达ꎬ使拟南芥对重金属Cd的耐性明显增强ꎬ对Cd2+的累积量也随即增加[82]ꎮ同样的结果在Grispen等的研究[83]中也得到体现ꎬ其在烟草中转入AtMT2b基因ꎬ提高了烟草对砷(As)的耐性ꎬ增加了As向植株地上部的转移ꎮ前面的研究很好地解释了植物MTs能提高植物对重金属的耐性ꎬ但其对重金属的解毒机制尚不清楚ꎬ可能是单一作用ꎬ也可能有其他转运蛋白和螯合肽的协同作用ꎬ包括运输转移等ꎮGuo等研究发现ꎬ在拟南芥中ꎬ有6种MTs:MT1aꎬMT2aꎬMT2bꎬMT3ꎬMT4a㊁MT4bꎮ其中MT1a对Cd的耐性和累积在拟南芥中是必需的ꎬPCs和MTs可以共同保护拟南芥免受镉的毒害[84]ꎮ3.1.3㊀植物螯合肽㊀PCs是一种由半胱氨酸㊁谷氨酸(Glu)和甘氨酸(Gly)组成的含巯基螯合多肽ꎬ其能和金属离子结合形成重金属PC螯合物[85]ꎮ植物螯合肽广泛地存在于单子叶㊁双子叶㊁裸子和藻类植物的细胞中ꎬ只要有微量的重金属ꎬ就能在植物细胞中检测到植物螯合肽ꎮPCs是重金属离子代谢ꎬ维持细胞体内平衡不可缺少的[86-87]ꎮPCs主要通过植物螯合态酶的作用ꎬ由谷胱甘肽合成ꎮ一些研究者报道ꎬ一定浓度的重金属胁迫下ꎬPCs能增加植物螯合肽的形成ꎬ其主要以(γ-Glu-Cys)n-Gly(n=2~11)的结构存在ꎬ植物螯合肽将和重金属Cd形成分子量为2500~3600的复合物[88]ꎮ虽然PCs在对重金属Cd的解毒过程中发挥着重要的作用ꎬ但其在生理或生态联系上的机制还不是很清楚ꎮWagner认为ꎬ在低浓度Cd污染的土壤中ꎬCd2+会与液泡中的柠檬酸发生螯合反应ꎻ而在高浓度Cd污染的土壤中ꎬPCs扮演着相当重要的角色[89]ꎮHowden等认为ꎬ即使在0.6μmol/L的Cd浓度下ꎬ甚至无污染的土壤中ꎬPCs含量也能被检测到ꎬ其在金属螯合体系中起到不可替代的作用[90]ꎮ张妍茹等研究发现ꎬ植物螯合肽合成酶(phytochelatinsynthaseꎬ简称PCs)可在重金属离子激活下ꎬ以谷胱甘肽及其巯醇盐为双底物催化合成植物螯合肽ꎬ从而解除生物体内重金属的毒性[91]ꎮ研究表明ꎬ在高等植物中分离最多的重金属螯合肽是PCsꎬPCs具有解除重金属对植物细胞的毒害以及维持细胞内必需金属离子浓度相对稳定的作用[92]ꎮ3.2㊀区隔化作用㊀㊀区隔化作用在植物耐重金属胁迫以及对重金属解毒的过程中发挥着重要的作用ꎬ其能阻止Cd2+在细胞质中自由流通ꎬ将Cd2+限制在一定的区域内ꎮSalt等研究发现ꎬ液泡内积累的Cd浓度是液泡外浓度的38倍[93]ꎮ细胞质和液泡对植物耐Cd的作用机制主要有以下3点:(1)在镉胁迫下ꎬ胁迫细胞质溶液中形成植物螯合肽(主要是PC3)和谷胱甘肽ꎬ这些物质或络合Cd2+形成低分子量复合物ꎬ从而限制Cd2+在细胞质特定区域活动[94]ꎮ(2)Kneer等研究发现ꎬ在液泡膜上存在着大量的S2-ꎬ能与大量的Cd2+结合[95]ꎮ(3)液泡内部ꎬ存在着各种有机酸(柠檬酸㊁草酸㊁苹果酸)和氨基酸ꎬ能与Cd2+结合形成各种高分子的复合物ꎬ使Cd2+的毒性降低并限制其流动[96]ꎮ杨红霞等研究发现ꎬ印度芥菜幼苗分别用0.5㊁1.0㊁2.0㊁3.0㊁5.0mg/L镉胁迫1㊁5㊁7㊁10㊁14d后收获ꎬ采用电感耦合等离子体质谱法测定印度芥菜根系和叶片亚细胞中的镉含量[97]ꎮ结果表明ꎬCd含量在细胞壁中的比例为50%~64%ꎬ细胞液中为22%~38%ꎬ细胞器中为7%~17%ꎬCd在细胞壁中所占比例显著高于在细胞液和细胞器中ꎬ该结果证明了细胞壁区隔化在Cd的解毒机制中具有重要的作用ꎮ㊀㊀另外ꎬ植物根系分泌物与根际形成的植物-微生物微系统能防御Cd2+等进入植物根系ꎬ这与植物种类㊁分泌物组成及根系表皮的细胞壁结构密切相关[98]ꎮ因此ꎬ不仅是液泡起到了区隔化作用ꎬ细胞壁在区隔化作用中也发挥了很好的作用ꎮ在细胞水平上ꎬ重金属超富集植物可使富集的重金属暂时储存于液泡中ꎬ缓解重金属对细胞质及细胞器的各种生理代谢活动的伤害[99]ꎮ首先ꎬ可以直接结合Cd2+ꎬ该反应短时间内就可以完成ꎻ其次ꎬ细胞壁中的纤维素㊁果胶质和糖蛋白极易形成网状结构ꎬ一方面阻止了Cd2+的流动ꎬ另一方面在一定程度上能固定Cd2+[77]ꎮ3.3㊀抗氧化作用重金属污染能导致植物体内产生大量的超氧阴离子自由基(O-2 )㊁羟自由基( OH)㊁一氧化氮自由基(NO )㊁氢过氧自由基(HOO )㊁烷氧基(RO )㊁烷过氧基(ROO )等活性氧自由基(ROS)ꎬ使蛋白质和核酸等生物大分子变性㊁膜脂过氧化ꎬ导致植物受伤害ꎮ据报道ꎬ植物的抗氧化系统在植物耐重金属胁迫中起非常重要的作用ꎬ并能通过其被氧化的程度来指示重金属的污染程度[100]ꎮ每个植物细胞都有合成抗氧化物质的能力ꎬ为避免在胁迫条件下ꎬ自由基的产生被潜在地破坏ꎬ植物在重金属胁迫下能合成各种抗氧化物质ꎬ以发挥防御作用[101]ꎮ为了应对胁迫ꎬ植物会产生抗氧化剂ꎬ包括抗氧化酶[SOD㊁CAT㊁POD㊁抗坏血酸过氧化物酶(APX)]及非酶抗氧化剂(GSH㊁AsA等)ꎬ以其来清除自由基ꎬ从而抵抗ROS对细胞的损伤[102]ꎮ这些抗氧化物质通过不同的机制来实现如活性氧物质的清除㊁重金属的螯合㊁抗氧化性物质的更新㊁抑制膜脂被伤害和修复被氧化所损失的DNA分子等功能[103]ꎮIannelli等研究发现ꎬ在镉胁迫下芦苇的各个器官中过氧化氢酶㊁超氧化物歧化酶等酶活性上升ꎬGSH含量会增加ꎬ谷胱甘肽硫转移酶(GST)活性也相应增强[104]ꎮAllen研究表明ꎬ在叶绿体中如果谷胱甘肽的合成增多ꎬ将会导致细胞的氧化性损伤ꎬ这可能是因为叶绿体所有的氧化还原状态被改变[105]ꎮ也有不同学者认为ꎬ不同抗氧化物质的比例减少ꎬ可以作为细胞内活性氧清除的1个信号[90ꎬ106]ꎮIannelli等的研究结果表明ꎬ以Cd2+处理的芦苇与对照相比ꎬ根㊁匍匐茎㊁4 江苏农业科学㊀2019年第47卷第2期。
植物对重金属镉超富集及耐受机理的研究

密级:中国科学院大学UniversityofChineseAcademySciences博士学位论文2013年5月专业综述:调控植物镉吸收转运及耐受性的分子遗传机制研究进展专业综述调控植物镉吸收转运及耐受性的分子遗传机制研究进展摘要镉(cadmium,Cd)对生物有很大毒害作用,因此对抗镉作用机制的研究,增强植物对镉的抗性,调控植物体内不同组织和器官的镉积累水平均具有重要意义。
Cd2+主要通过同为二价阳离子的Fe2+、Ca2+或zn”的低特异性转运蛋白或通道蛋白进入植物细胞内,主要被贮存在根细胞液泡中,但有相当部分通过木质部导管长途转运地上部分,再通过韧皮部进行再分配,储存在不同组织的液泡或者细胞壁等部位。
在植物和真菌的重金属解毒机制中,植物螯合肽(phytochelatins,PCs)起着核心的作用,能与重金属螫合为复合物,再通过跨液泡膜转运区室化到液泡,降低细胞质中重金属成分的含量。
本文对近年来调控植物镉吸收转运及耐受性的分子遗传机制研究新进展进行了概述。
关键词镉;螫合;区室化。
在生命的演化进程中,植物形成了多样化的机制来维持体内与周围环境存在的可利用重金属离子之间的平衡关系。
植物面临着两个重要任务,一是从生长环境中选择吸收生长所必需的重金属离子并拒绝吸收非必需的重金属离子,二是在细胞内维持这些金属离子保持最适宜的生理浓度(Cobbetteta1.,2002)。
对金属离子的吸收和累积机制随着植物的种类的不同而不同,即使是在同一个属,不同种植物的吸收机制也不尽相同(Singha1.,2003)。
植物可通过根部直接吸收水溶性重金属,植物根组织不但可以通过根细胞膜上的质子泵使根际(rhizosphere)酸化,而且能够分泌具有金属螫合功’-能的低分子量复合物,使得土壤的金属离子更易被根部吸收。
现在并不清楚是否植物铁载体(phytosiderophore)或有机酸(例如柠檬酸盐)参与了非必需的毒性金属离子(例如Cd”)的吸收过程。
超富集植物遏蓝菜对重金属吸收_运输和累积的机制

生物工程学报Chin J Biotech2010, May 25; 26(5): 561–568 Chinese Journal of Biotechnology ISSN 1000-3061 cjb@©2010 CJB, All rights reserved.超富集植物遏蓝菜对重金属吸收、运输和累积的机制刘戈宇,柴团耀,孙涛中国科学院研究生院生命科学学院,北京 100049摘要:遏蓝菜Thlaspi caerulescens可以在其地上部累积大量重金属如锌、镉等,是公认的超富集植物。
由于该植物生物量小,不宜直接用于重金属污染的土壤植物修复,而被广泛作为一种模式植物来进行重金属富集机制研究。
遏蓝菜对重金属离子的累积大致经过螯合剂解毒、地上部长距离运输以及在液泡中的储存等生理过程。
已经发现的植物体内的金属螯合剂——有机酸、氨基酸、植物络合素(PCs)、金属硫蛋白(MT) 和尼克烟酰胺NA等,区室化以及长距离运输相关的转运蛋白——ZIP (ZRT/IRT like protein)、CDF (Cation diffusion facilitator)、Nramp (Natural resistance and macrophage protein) 和HMA (Heavy metal ATPase) 等家族,以上各种基因、多肽与蛋白等共同参与了植物对金属累积与耐受过程并发挥各自重要的作用。
以下主要介绍了遏蓝菜重金属超富集相关的基因、多肽和蛋白,以及它们在重金属螯合作用和运输过程中的功能。
关键词:遏蓝菜,重金属,富集,螯合,转运Heavy metal absorption, transportation and accumulation mechanisms in hyperaccumulator Thlaspi caerulescensGeyu Liu, Tuanyao Chai, and Tao SunCollege of Life Science, Graduate University of Chinese Academy of Sciences, Beijing 100049, ChinaAbstract:Thlaspi caerulescens, the famous model plant of heavy-metal hyperaccumulator, can uptake and accumulate large amount of heavy metals in its above-ground part of the plants. However, the very low biomass in Thlaspi caerulescens makes this plant unfit for direct application in phytoremediation. In recent years, there are many reports about the physiological and molecular characterization of Thlaspi caerulescens under heavy metals stresses, including absorption, transport and intracellular detoxification processes (e.g., chelation and compartmentation). Research teams have conducted many studies of chelators in plants, such as organ acid, amino acid, phytochelatins, metallothioneins and nicotianamine, and so on. Several transport protein families, such as Zinc Regulated Protein, Cation Diffusion Facilitator, Natural Resistance and Macrophage Protein and Heavy Metal ATPase, play important role in short/long distance transport in the plant. In this review, we summarize the current knowledge of the physiological and molecular mechanisms of heavy metals accumulation in Thlaspi caerulescens, with particular emphasis on the roles of transporters and chelatins in modulating plant heave-metal-stress responses.Keywords:Thlaspi caerulescens, heavy metal, accumulation, chelation, transportationReceived:November 17, 2009; Accepted: March 8, 2010Supported by: National High Technology Research and Development Program of China (863 Program) (No. 2006AA10Z407), Genetically Modified Organisms Breeding Major Projects (No. 2009ZX08009-130B).Corresponding author: Tuanyao Chai. Tel/Fax: +86-10-88256343; E-mail: tychai@国家高技术研究发展计划 (863计划) (No. 2006AA10Z407),转基因生物新品种培育科技重大专项 (No. 2009ZX08009-130B) 资助。
镉对植物的毒害及植物解毒机制研究进展

镉对植物的毒害及植物解毒机制研究进展2. 镉对植物的毒害镉是一种重金属元素,其在土壤中的积累会严重影响植物的生长和发育。
镉会通过植物的根部被吸收进入到植物体内,随着土壤中镉的积累,镉会影响植物的营养吸收和水分利用,从而导致植物的生长受阻和光合作用受抑制。
镉还会影响植物的生理代谢过程,导致植物对抗逆境的能力下降,出现生理障碍和形态异常。
最终,植物的生长受限,产量减少,甚至无法正常生长,甚至可能导致植物死亡。
3. 植物解毒机制在受到镉污染的环境中,植物为了适应环境并减轻镉的毒害,发展了一系列的解毒机制。
植物可以通过根系排除来减少镉在植物体内的积累。
根系排除是植物对抗镉污染的一种重要途径,它通过增加细胞膜的通透性和特异性转运蛋白的表达来减少镉的内部富集。
植物可以通过螯合剂和酶解毒来减轻镉的毒害。
螯合剂可以与镉离子结合形成不活跃的物质,减少镉对植物的伤害。
酶解毒则可以通过酶的催化作用将有害的物质转化为无害的物质,以减轻镉对植物的损害。
植物还可以通过抗氧化系统减轻镉的毒害。
抗氧化系统能够清除植物体内镉引起的氧化应激,减轻镉对植物的氧化损害,保护植物的生理代谢过程,从而提高植物对镉的抵抗能力。
4. 镉对植物解毒机制研究进展近年来,伴随着生物技术和分子生物学的发展,关于镉对植物解毒机制的研究也得到了较大的进展。
一方面,研究者通过利用研究模型植物(如拟南芥、水稻等)进行功能基因组学和转录组学的研究,揭示了一系列参与植物抗镉的关键基因和信号通路,为深入了解植物解毒机制提供了重要的参考。
通过基因敲除和过表达等技术手段,研究者发现了一些与植物对抗镉毒害相关的基因,如金属运输蛋白(如HMA、NRAMP等)、螯合剂合成酶(如PCS、MTs等)、抗氧化酶(如SOD、CAT等)等。
一些研究者还通过比较不同植物对镉反应的差异,发现了一些对镉高效积累和耐受的植物种质资源,为植物资源的筛选和开发提供了重要的参考。
通过不断深入的研究,植物对镉的解毒机制逐渐被揭示,为植物的镉污染防治提供了新的途径和手段。
农作物土壤中镉含量分析及作物镉富集系数评价研究

农作物土壤中镉含量分析及作物镉富集系数评价研究
黄娇;郑智溢
【期刊名称】《农业与技术》
【年(卷),期】2015(000)003
【摘要】基于农产地土壤、农作物中的镉含量分析,进行二者的相关性研究,以及不同作物对土壤镉富集吸收能力的表征与比较分析。
结果表明,土壤中镉含量平均值为0.21mg/kg ,农作物含镉量在0.004~0.73mg/kg ;在蔬菜中镉含量高低的次序为:薯芋类>叶菜类>茄果类>豆类>瓜类>水生类。
作物镉富集系数的对数值概率分布服从正态性,富集系数对数值的平均数可用来表征比较作物对镉的富集能力。
【总页数】3页(P20-22)
【作者】黄娇;郑智溢
【作者单位】乐清市食品质量安全检测中心,浙江乐清 325600;乐清市食品质量安全检测中心,浙江乐清 325600
【正文语种】中文
【中图分类】X825
【相关文献】
1.浙江省主要优势农产品产地土壤-农作物镉含量空间分布及相关性研究
2.恩施富硒土壤区主要农作物硒镉特征以及和根系土硒镉关系研究
3.土壤与水稻籽粒镉含量相关性分析及水稻产地土壤镉临界值的研究
4.浙江省主要优势农产品产地土壤-
农作物镉含量空间分布及相关性研究5.不同修复技术对土壤有效态镉含量及作物镉吸收的影响
因版权原因,仅展示原文概要,查看原文内容请购买。
植物对重金属镉的响应及其耐受机理

植物对重金属镉的响应及其耐受机理
宋瑜;金棵;曹宗英;王晓娟
【期刊名称】《草业学报》
【年(卷),期】2008(017)005
【摘要】重金属Cd作为非必需微量元素,经根系吸收并累积时对植物具有很强的毒性,因而开展植物对Cd的响应途径及其调控机理研究,对改良植物对Cd的耐受性以及开发超累积植物均具有重要意义.植物硫代谢、抗氧化系统和Cdz2+跨膜运输是植物对重金属镉响应的主要途径,本研究综述了以上3种耐受机制的研究进展,包括Cd2+诱导植物硫转运蛋白、硫还原相关酶类以及半胱氮酸、谷胱廿肽和植物螯合肽合成及其基因表达调控,Cd2+诱发的植物抗氧化反应及其基因表达,质膜和液泡转运蛋白促进Cd2+运输和隔离的基因调控.
【总页数】8页(P84-91)
【作者】宋瑜;金棵;曹宗英;王晓娟
【作者单位】兰州大学草地农业科技学院,甘肃,兰州,730020;兰州大学草地农业科技学院,甘肃,兰州,730020;兰州大学草地农业科技学院,甘肃,兰州,730020;兰州大学草地农业科技学院,甘肃,兰州,730020
【正文语种】中文
【中图分类】Q946
【相关文献】
1.几种湿地植物对重金属镉胁迫的生理生化响应 [J], 张超兰;陈文慧;韦必帽;刘小珍;吕沛峰
2.植物对重金属镉的耐受机制 [J], 张军;束文圣
3.植物对重金属镉抗性机理及利用的研究综述 [J], 申强;徐金益;张亮;王程亮;薛永;
4.植物重金属镉(Cd^(2+))吸收、运输、积累及耐性机理研究进展 [J], 仇硕;张敏;孙延东;黄苏珍
5.3种蕨类植物对锰污染土壤的耐受性及生理响应 [J], 汪结明;王良桂;樊亚珍;吴净因版权原因,仅展示原文概要,查看原文内容请购买。
土施超富集植物秸秆对荠菜生长及镉积累的影响

土施超富集植物秸秆对荠菜生长及镉积累的影响胡容平;石军;黄廷友;林立金【期刊名称】《水土保持通报》【年(卷),期】2015(35)5【摘要】[目的]研究超富集植物秸秆对富集植物重金属积累的化感作用,为镉污染土壤的植物修复研究提供参考。
[方法]通过在镉污染土壤中施用镉超富集植物〔(红果黄鹌菜Youngia erythrocarpa)、三叶鬼针草(Bidens pilosa)、少花龙葵(Solanum americanum)和豨莶(Siegesbeckia orientalis)〕秸秆,研究了4种镉超富集植物秸秆施入土壤对镉富集植物荠菜生长及镉积累的影响。
[结果]4种镉超富集植物秸秆施入土壤均提高了荠菜地上部分生物量、总生物量和抗性系数,同时提高了荠菜叶片SPAD值(绿色度)。
土施红果黄鹌菜秸秆提高了荠菜根系、茎、叶片及地上部分的镉含量,分别比各自对照高11.81%,102.07%,12.00%和54.95%,同时也提高了土样有效态镉含量,其余3种处理均低于各自对照。
土施红果黄鹌菜秸秆的荠菜地上部分和整株的镉积累量均高于对照,分别为42.11和54.74μg/株,比各自对照高83.09%和54.11%,而其余3种处理的镉积累量均低于对照。
[结论]土施红果黄鹌菜能够提高荠菜对土壤镉的积累,有利于提高荠菜对镉污染土壤的修复效果。
【总页数】5页(P217-221)【关键词】超富集植物秸秆;荠菜;镉;植物化感【作者】胡容平;石军;黄廷友;林立金【作者单位】四川省农业科学院植物保护研究所/农业部西南作物有害生物综合治理重点实验室;绵阳市农业科学研究院;四川农业大学果蔬研究所【正文语种】中文【中图分类】X53;S154【相关文献】1.施用富集植物秸秆对水田芥生长及镉积累的影响 [J], 王进;陈发波;林立金;吕秀兰;廖明安;蒋伟;任纬2.土施富集植物秸秆对牛膝菊生长及镉积累的影响 [J], 汤福义;林立金;廖进秋;廖明安;何静;杨代宇;张潇3.土施镉超富集植物秸秆对镉胁迫牛膝菊光合特性的影响 [J], 胡容平;石军;黄廷友;林立金;廖明安4.土施超富集植物秸秆对镉污染条件下葡萄生长及镉含量的影响 [J], 李欣欣;钟莉莎;马倩倩;张潇;林立金;廖明安5.耐性植物秸秆覆盖对荠菜生长及镉积累的影响 [J], 王恒;石军;林立金;杨代宇;黄科文;张潇因版权原因,仅展示原文概要,查看原文内容请购买。
超富集植物吸收镉的原理

超富集植物吸收镉的原理
一、超富集植物的特征
超富集植物是可以大量吸收和蓄积重金属元素的植物,如蓄积镉的虎刺梅和蓄积砷的膽砂藻等。
二、对镉吸收量大
超富集植物对镉元素的吸收量非常大,可以在植物体内蓄积高浓度的镉而不会中毒死亡。
三、根系分泌有机酸
根系会分泌柠檬酸、苹果酸等有机酸,与镉结合成可溶性盐,利于镉被吸收。
四、镉离子运输机制
植物细胞膜上的转运蛋白会主动运送镉离子进入根细胞。
还可能依靠综合作用被动吸收。
五、允许镉进入尖端部位
超富集植物的细胞壁结构允许镉离子进入茎尖、叶尖等部分的细胞堆积。
六、细胞内镉结合
植物可合成螯合剂如蛋白、阻酸等与镉结合,形成结合态固定在细胞液泡中。
所以超富集植物通过一系列生理机制,能大量吸收和蓄积镉元素,这可以用于土壤镉的去除修复。
植物激素、螯合剂复合处理对镉胁迫下籽粒苋生长和富集镉的影响
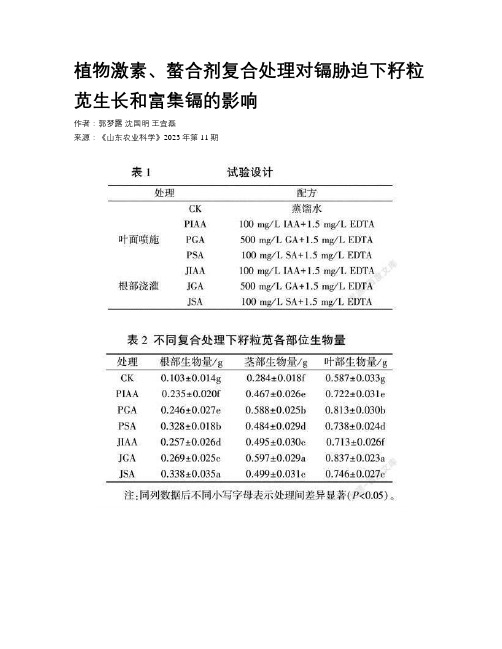
植物激素、螯合剂复合处理对镉胁迫下籽粒苋生长和富集镉的影响作者:郭梦露沈国明王宜磊来源:《山东农业科学》2023年第11期关键词:镉胁迫;籽粒苋;植物修复;植物激素;螯合剂随着工业和经济的快速发展,土壤重金属污染问题日益严重,我国受Cd、As、Pb、Hg、Zn等重金属污染的耕地面积约占总耕地面积的五分之一。
重金属排放到环境中,严重污染水体和土壤,其中有高毒性和潜在致癌性的Cd是污染超标率最高的重金属:土壤Cd点位超标率最高达7%,其通过植物吸收、积累、富集后经食物链进人人体,从而对人类健康造成威胁。
Cd胁迫对植物生长发育有明显的抑制和毒害作用,可通过抑制种子中的淀粉酶、蛋白酶活性影响淀粉和蛋白质的分解,从而对种子萌发产生影响。
因而,有必要采取不同技术措施对Cd污染进行有效修复。
植物修复技术因具有成本低、不引入二次污染等优势而前景广阔,其修复效率主要取决于植物生物量和其累积重金属的能力。
选择生物量大、富集量大的植物以及添加外源激素可以大大提高植物的修复效率。
众多研究结果表明,植物激素能缓解重金属的植物毒性,提高植株的抗逆性,促进其生长发育;螯合剂对重金属离子具有较强的螯合力,施加螯合剂能提高重金属的迁移能力和生物有效性。
如Wang等研究EDTA(乙二胺四乙酸)强化盐生植物景天三七对土壤中Pb、Cd的去除效果发现,EDTA可以强化景天三七修复土壤的能力,使Pb、Cd的去除率分别达到37.87%和41.61%。
综上,很多研究表明植物激素和螯合剂的添加可提高植物修复效率,但关于植物激素和螯合剂复合处理的研究较少,两种添加剂复合处理是否会大大增加植物修复效率仍未可知:外源激素的添加方式有多种,具体何种添加方式能最大程度增加修复效率也鲜有研究。
龙玉梅等通过温室盆栽试验比较籽粒苋、龙葵、商陆、青葙4种植物对镉污染土壤的修复效果,结果表明,4种植物可收集的镉总量排序为籽粒苋>青葙>商陆>龙葵。
七种本土植物对土壤中镉的富集效果比较研究
七种本土植物对土壤中镉的富集效果比较研究作者:王振艳丁军孙向辉赵京陈婉秋来源:《湖北农业科学》2020年第12期摘要:针对河南省新乡市王村镇及其周边地区镉污染现状,选取7种长势较好的本土植物,对其镉富集特征和修复效果进行了对比分析。
结果表明,草本植物的镉耐受能力整体高于灌木或乔木,但紫薇(Lagerstroemia indicaL)表现出较高的富集能力和转移系数;狗尾草[Setaria viridis(L)Beauv.]根中镉含量达82.5 mg/kg,根部富集系数为3.149;苋菜(Amaranthus tricolor L)各部分镉含量、转移系数和富集系数在7种植物中均处于优势地位。
综合分析认为,苋菜、狗尾草和紫薇可作为该区域Cd高富集植物来净化土壤。
关键词:镉污染;土壤修复;本土植物;富集特征中图分类号:X53文献标识码:A文章编号:0439-8114( 2020)12-0061-03DOl:10.1408 8/j .cnki.issn043 9- 8114.2020.12.012开放科学(资源服务)标识码(OSID):2014年环保部与国土资源部发布《全国土壤污染状况调查公报》,结果显示,全国土壤总的点位超标率为16.1%,污染类型以无机型为主,无机污染物超标点位数占全部超标点位的82.8%,重金属污染物镉点位超标率为7.0%[1]。
镉是毒性较强的重金属元素,在环境中具有生物遷移性强、毒性持久、难降解等特点[2],不仅会引起土壤功能失调、土质下降,还会通过食物链富集、转移,长期摄人受到镉污染的食品,会造成镉在体内蓄积,从而危害人类的生命和健康。
因此对镉污染农田土壤的治理已经引起国内外的广泛重视。
河南省新乡市被中国轻工业联合会、中国电池工业协会联合授予“中国电池工业之都”荣誉称号。
王村镇位于新乡市西北部,地理坐标东经113°50'10”-113°52'17”,北纬35°20'57”-35.21 '50”。
蔬菜对土壤高浓度镉的吸收与积累特点及耐性

关键词 : 蔬菜 ;土壤 ; d 吸收 与积 累 ; C ; 耐性
中图 分 类 号 : 6 1 S0 文献标 志码 : A 文 章 编 号 :0 2—10 (0 1 0 0 9 0 10 3 2 2 1 )4— 15— 2 7 ℃烘 干 至恒 重 , 烘 干 至 恒 重 的蔬 菜 地 上 部 和 根 部 用 玻 璃 5 将 研钵 分 别 磨 成 粉 末 , 分 混 匀 。 C 充 d含 量 采 用 万 拓 原 子 荧 光 A S一 3 a 定 。 F 20 测
耐 性 指 数 : ( 理 生 物 鲜 重/ 照 生 物 鲜 重 一1 × 处 对 )
1 0% [ ] 0
究。一般认 为 , 同等 条 件 下 C d在 蔬 菜 中的 含 量 要 大 于 瓜
果 , 蔬 菜 中又 以 叶 菜 类 对 C 而 d的 富 集 量 较 大 。 这 些 研究 主要 集 中在 C d浓 度 较 低 的 土 壤 环 境 , 关 于 蔬 菜 在 土 而 壤高 c d浓 度 条 件 下 的 研 究 报 道 较 少 。 本 试 验 通 过 盆 栽 试
为 C 鲜 重 8 9g的 1 . % 。有 研 究 表 明 , 性 指 数 越 大 表 K . 24 耐
供试蔬菜为菠菜 ( 日本华 波 l号 ) 萝 卜( 力高精 品今 、 杰
春红 ) 芹菜( 、 京都实芹王 ) 大 白菜 ( 高 8 、 德 7—14 、 l ) 白菜 ( 寒
山正 京 矮 帮 苏 州 青 ) 处 理 所 用 镉 试 剂 为 分 析 纯 氯 化 镉 ;
c d吸收系数 = 菜中 c 蔬 d含量/ 土壤 中 c d含量“
镉胁迫对两种蔊菜生长的影响及其在植物体内的积累和转运特性

http : //D01:10.3969/j.issn.1000-1700.2020.06.010沈阳农业大学学报,2020,51 (6):714-720 Journal of Shenyang Agricultural University关 萍,金 恺,袁蕴宁,等.镉胁迫对两种蔊菜生长的影响及其在植物体内的积累和转运特性[J].沈阳农业大学学报,2020,51(6):714-720.镉胁迫对两种蔊菜生长的影响及其 在植物体内的积累和转运特性关 萍",金 恺2,袁蕴宁",曲 波",邵美妮1笃许玉凤1a(1.沈阳农业大学a.生物科学技术学院,b.林学院,沈阳110161;2.辽宁省高速公路运营管理有限责任公司,沈阳110000)摘 要:为探究植物对镉胁迫的响应机制和富集特性,以欧亚蔊菜(Rorippa sylvestris 冤和两栖蔊菜(Rorippa amphilia)为试验材料, 采用盆栽方式,镉处理浓度分别为0,50,100,200,400mg-kg -1,通过测定株高、生物量、光合参数以及植物体内镉的含量等指标,研究 镉胁迫对两种蔊菜生长的影响,分析镉的吸收和转运特性,探讨两种蔊菜对镉的耐受性和富集能力。
结果表明:当镉浓度为50mg-kg -1时,两种蔊菜生长受到的影响较小;当镉浓度高于50mg-kg -1时,两种蔊菜生长受到抑制。
随着镉浓度增加,叶绿素含量逐渐减少,最大净光合速率P ”逐渐下降,最大光合效率(Fv/Fm )逐渐降低,表现岀光抑制;光合电子传递速率(ETR 冤逐渐降低,而非光化学淬 灭系数(NPQ)逐渐升高,说明叶片的光合系统受到破坏。
随着镉处理浓度增加,两种蔊菜的根、茎、叶中镉含量逐渐增大,各处理间差异显著,除对照外,各处理地上部分镉含量都超过100mg - kg -1 ;地上富集系数和地下富集系数呈现增加的趋势,且均大于1.0;欧亚蔊菜的转运系数都大于1.0,各处理间差异不显著;两栖蔊菜在镉浓度为50mg ・kg -1时,地上转运系数大于1.0;欧亚蔊菜的转运 系数均高于两栖蔊菜。
一种强化超富集植物修复镉污染土壤的强化剂及其应用[发明专利]
![一种强化超富集植物修复镉污染土壤的强化剂及其应用[发明专利]](https://img.taocdn.com/s3/m/ead904a4f7ec4afe05a1df03.png)
专利名称:一种强化超富集植物修复镉污染土壤的强化剂及其应用
专利类型:发明专利
发明人:代惠萍,魏树和
申请号:CN201711129748.1
申请日:20171115
公开号:CN107889840A
公开日:
20180410
专利内容由知识产权出版社提供
摘要:本发明公开一种强化超富集植物修复镉污染土壤的强化剂及其应用,所述强化剂由大白菜提取物、球果蔊菜提取物和水按照质量比(0.01‑1):(1‑20):(200‑2000)混合,其中,球果蔊菜提取物是通过以下方法提取得到的:(1)将球果蔊菜风干至水分含量为8%‑10%,粉碎,得到球果蔊菜粉末;(2)按照料液比1g:100mL,取球果蔊菜粉末与超纯水混合,在25‑110℃
下,3000‑10000rpm的转速下浸提10‑200h,过滤,得到滤渣;(3)将滤渣烘干,即可。
本发明提供的强化超富集植物修复镉污染土壤的强化剂,环保安全,不会对土壤造成二次污染,能增强超富集植物对土壤中镉的吸附和富集效果。
申请人:陕西理工大学
地址:723000 陕西省汉中市东关小关子
国籍:CN
代理机构:西安亿诺专利代理有限公司
代理人:张建
更多信息请下载全文后查看。
球果蔊菜对重金属的超富集特征

球果蔊菜对重金属的超富集特征魏树和;周启星;任丽萍【期刊名称】《自然科学进展》【年(卷),期】2008(018)004【摘要】利用室外盆栽试验、小区模拟试验和污染区采样试验的方法,从22科65种农田杂草植物中筛选重金属超富集植物.盆栽试验结果表明:在Cd污染水平为25.0和50.0mg·kg-1时,球果蔊菜(Rotrippa globosa)茎和叶中Cd含量均超过100 mg·kg-1这一Cd超富集植物应达到的临界含量标准,地上部富集系数大于1,地上部Cd含量大于根部Cd含量,而且植物的生长未受抑制,这些特征完全符合Cd 超富集植物的基本特征.小区试验和污染区采样试验中,球果蔊菜也表现出Cd超富集植物的基本特征,可以基本认为是Cd超富集植物.【总页数】7页(P406-412)【作者】魏树和;周启星;任丽萍【作者单位】中国科学院沈阳应用生态研究所陆地生态过程重点实验室,沈阳110016;中国科学院沈阳应用生态研究所陆地生态过程重点实验室,沈阳110016;南开大学环境科学与工程学院,天津300071;中国科学院沈阳应用生态研究所陆地生态过程重点实验室,沈阳110016【正文语种】中文【中图分类】S6【相关文献】1.当归植株对重金属的富集特征研究 [J], 仲启祥;朱锦福;谢慧春2.铁尾矿库区白茅对重金属的吸收与富集特征 [J], 宋凤敏;张兴昌;刘瑾;蒲东浩;赵亚峰;乔权3.地质高背景农田土壤中水稻对重金属的富集特征及风险预测 [J], 唐豆豆;袁旭音;汪宜敏;季峻峰;文宇博;赵万伏4.三叶鬼针草等7种常见菊科杂草植物对重金属的超富集特征 [J], 魏树和;杨传杰;周启星5.滨州湿地植物对重金属的富集特征 [J], 张再旺;靳建超;李赫;张吉强;许骥坤;田园;马春茂;周同娜因版权原因,仅展示原文概要,查看原文内容请购买。
镉超积累植物及植物镉积累特性转基因改良研究进展

镉超积累植物及植物镉积累特性转基因改良研究进展蔡利娟;范仲学;全先庆;张延新;杜瑞雪【期刊名称】《广西植物》【年(卷),期】2009(029)005【摘要】植物提取修复技术是一项既经济又环保的土壤镉(Cd)污染修复技术,该技术的关键是筛选Cd超积累植物或利用基因工程手段改良植物以提高其Cd积累能力.人们已发现遏兰菜等7种Cd超积累植物及美人蕉等潜在的Cd超积累植物.还发现了许多与Cd耐受和积累能力有关的基因:(1)编码与Cd积累、耐受有关酶的基因,如细菌中的ACC(1-aminocyclopropane-1-carboxylic acid),植物中的PCS(Phytochelatin Synthase)基因;(2)编码金属结合蛋白的基因:MT(Metallothionein)、转运蛋白(P-type ATPase、ABC型转运器)基因;(3)其它相关基因:Hvhsp17、PvSR2(Phaseolus vulgaris stress-related gene number 2)等.并将其中的一些基因转入到其它生物中,提高了其对Cd的耐受性和积累量,为实现Cd污染土壤修复的目标奠定基础.【总页数】7页(P658-663,702)【作者】蔡利娟;范仲学;全先庆;张延新;杜瑞雪【作者单位】山东省农业科学院,高新技术研究中心,山东省作物与畜禽品种改良生物技术重点实验室,济南,250100;青岛农业大学,生命科学学院,山东,青岛,266109;山东省农业科学院,高新技术研究中心,山东省作物与畜禽品种改良生物技术重点实验室,济南,250100;山东省农业科学院,高新技术研究中心,山东省作物与畜禽品种改良生物技术重点实验室,济南,250100;山东省农业科学院,高新技术研究中心,山东省作物与畜禽品种改良生物技术重点实验室,济南,250100;山东省农业科学院,高新技术研究中心,山东省作物与畜禽品种改良生物技术重点实验室,济南,250100【正文语种】中文【中图分类】Q945.78【相关文献】1.氮素营养对重金属超积累植物东南景天吸收积累锌和镉的影响 [J], 张圆圆;窦春英;姚芳;叶正钱2.超积累植物和化学改良剂联合修复锌镉污染土壤后的微生物特征 [J], 彭桂香;蔡婧;林初夏3.间作条件下超积累和非超积累植物对重金属镉的积累研究 [J], 霍文敏;邹茸;王丽;范洪黎4.氮对超积累植物东南景天生长和镉积累的影响 [J], 李继光;李廷强;朱恩;杨肖娥;林国林;柳丹;韩晓日;张玉龙5.镉超积累植物龙葵叶片中镉的积累与有机酸含量的关系 [J], 孙瑞莲;周启星;王新因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
镉超富集植物球果 菜对镉2砷复合污染的反应及其吸收积累特征孙约兵1,2,周启星1,33,任丽萍1(11中国科学院沈阳应用生态研究所陆地生态过程重点实验室,沈阳 110016;21安徽师范大学重要资源保护与利用研究安徽省重点实验室,安徽芜湖 241000;31南开大学环境科学与工程学院,天津 300071)摘要:采用室外盆栽模拟方法,比较和分析了镉(Cd )2砷(As )复合污染处理对球果 菜(Rorippa globosa ,十字花科焊菜属,1种从农田杂草中新发现的镉超富集植物)的生长及其对Cd 、As 吸收和积累特征的影响.结果表明,在低浓度Cd 2As 复合污染条件下,由于Cd 、As 的拮抗作用,从而促进植物的生长发育,同时也促进地上部对Cd 的吸收和积累.在Cd 2As 复合污染处理浓度为10mg Πkg 和50mg Πkg 时,株高和地上部干重达到最大(分别为3519cm 和212g Π盆),叶片中Cd 积累量高于同浓度单一Cd 处理.Cd 2As 复合污染高浓度处理表现出对球果 菜生长以及Cd 的吸收和积累有协同的抑制作用.同时,球果 菜根部对As 的吸收能力大于其地上部,相应的富集系数≤013,转移系数≤016,说明球果 菜对As 有一定的排斥作用.这些结果表明,球果 菜有很强的忍耐Cd 2As 复合污染的能力,对修复Cd 2As 复合污染土壤具有一定的潜力.关键词:镉超富集植物;球果 菜;复合污染;植物修复中图分类号:X17114 文献标识码:A 文章编号:025023301(2007)0621355206收稿日期:2006207210;修订日期:2006208221基金项目:海外青年学者合作研究基金项目(20428707);国家自然科学基金重点项目(20337010);中俄自然资源与生态环境联合研究中心项目作者简介:孙约兵(1978~),男,硕士,主要研究方向为污染土壤的植物修复.3通讯联系人,E 2mail :Zhouqx @G row th R esponses of Rorippa globosa and Its Accumulation Characteristics of Cd and As under the Cd 2As Combined PollutionS UN Y ue 2bing1,2,ZH OU Qi 2xing1,3,RE N Li 2ping1(11K ey Laboratory of T errestrial Ecological Process ,Institute of Applied Ecology ,Chinese Academy of Sciences ,Shenyang 110016,China ;21Provincial K ey Laboratory of C onservation and Exploitation of Biological Res ources ,C ollege of Life Sciences ,Anhui N ormal University ,Anhui Wuhu 241000,China ;31C ollege of Environmental Science and Engineering ,Nankai University ,T ianjin 300071,China )Abstract :Rorippa globosa has been identified as a new Cd 2hyperaccumulating plant species.In the present study ,growth responses of Rorippa globosa and its accumulation characteristics of Cd and As were examined under the condition of Cd 2As combined pollution.The results showed that Cd and As had an antag onistic effect on enhancing the growth of the plants and Cd uptake and accumulation under the low concentration Cd and As treatments.When Cd and As concentrations were 10mg Πkg and 50mg Πkg ,the highest height of the plants and the dry weight of shoots were up to 3519cm and 212g Πpot ,respectively ;and the accumulation of Cd in the leaves under the combined pollution was higher than that at the same level under single Cd pollution.H owever ,there were synergic adverse effects on plant growth and Cd uptake under the high concentration Cd and As combined pollution.Meanwhile the accumulation of As in the roots was higher than that in the shoots ,the translocation factor ≤013and the bioaccumulation factor ≤016,which showed that Rorippa globosa had an excluding effect on As uptake.These results con firmed that Rorippa globosa had the strong tolerance ability to the Cd 2As combined pollution ,and the potential applied to phytoremediation of contaminated s oil by Cd and As.K ey w ords :Cd 2hyperaccumulator ;Rorippa globosa ;combined pollution ;phytormediation 由于工业废物的排放和不合理的农业管理措施,致使农田土壤中镉(Cd )、砷(As )污染日趋严重,并不同程度地导致了人和动物体急性、亚急性和慢性中毒,还可诱发引起致癌、致畸[1~3].由于Cd 、As 污染在土壤环境中表现为具有隐蔽性、长期性和不可逆转性的特点,使得这2种金属污染土壤的治理和修复成为亟需解决的现实问题.对于具有经济高效、绿色净化和易于后续处理等优点的植物修复技术[4~6],其关键是选育出对污染元素有较强吸收能力的超富集植物(hyperaccumulator ).这种植物对重金属的吸收积累量超过一般植物的100倍而不影响其正常生长[7].目前,有关超富集植物的衡量标准有3个:一是临界含量标准,即植物茎或叶中重金属达到其临界含量,其中Cd 为100mg Πkg ,As 为1000mg Πkg[7];二是富集系数标准,即富集系数大于110,但有时甚至达50~100[8];三是转移系数标准,即重金属在植物地上部积累量大于其根部积累量[9],同第28卷第6期2007年6月环 境 科 学E NVIRONME NT A L SCIE NCEV ol.28,N o.6Jun.,2007时还需要对它们有一定的耐受能力.但是,这些已知的超富集植物都不同程度存在着不能同时超量积累多种重金属、生物量较小、生长缓慢,且在基因工程培育理想的超富集植物方面进展也十分缓慢等缺点,从而影响植物修复技术的有效性和广泛应用[10].与作物相比,杂草抗逆境能力强,同时还具有较强的争光、争水和争肥的能力,是修复污染土壤较理想的种质资源.球果 菜(Rorippa globosa)是我国境内利用农田杂草发现的为数不多的1种镉超富集植物(专利号:C N20041002098112).已有研究表明,当土壤Cd达到25mgΠkg时,其开花期和成熟期叶片中Cd含量分别为13116mgΠkg和15011mgΠkg[11],地上部Cd积累量大于其根部积累量,且地上部富集系数大于110,符合超富集植物的基本特征[7].鉴于镉超富集植物对Cd2As复合污染土壤的修复以及同时对Cd、As具有超富集作用的植物鲜见报道,本试验旨在研究Cd2As 复合污染条件下球果 菜的生长反应及其对Cd、As的吸收和积累特征,探讨它们互作的内在机制,以期为Cd2As复合污染土壤的修复和治理提供科学依据.1 材料与方法111 试验地点概况试验地点设在中国科学院沈阳生态实验站内,地理位置为东经123°41′、北纬41°31′,海拔约50m,属温带半湿润大陆性气候,年平均温度5~9℃,无霜期为127~164d,年降水量650~700mm.盆栽试验土壤采自该站表土(0~20cm),土壤类型为草甸棕壤,其理化性质为pH值6156,有机质12126gΠkg,全N0191gΠkg,全P014gΠkg,全K183gΠkg,总Cd0117 mgΠkg,总Cu3219mgΠkg,总Zn2811mgΠkg,总Pb 1111mgΠkg,总As1014mgΠkg.112 试验方法将供试土壤风干后过4mm筛后,每盆装土215kg,与一定量的污染物充分混匀后装入塑料盆(<=20cm,H=15cm),Cd2As复合污染处理,设3个Cd水平(mgΠkg):10、25、50;2个As水平(mgΠkg):50、250;另外设1个对照,共7个处理,即为:①CK;②Cd10mgΠkg+As50mgΠkg;③Cd10mgΠkg+As250 mgΠkg;④Cd25mgΠkg+As50mgΠkg;⑤Cd25mgΠkg+ As250mgΠkg;⑥Cd50mgΠkg+As50mgΠkg;⑦Cd50 mgΠkg+As250mgΠkg.以CdCl2・215H2O和Na2H AsO4・7H2O的形式加入.平衡3周后,选择生长一致的球果 菜幼苗分别移栽入各盆中,每盆3棵苗,重复3次.为了使其在自然状况下生长,不施底肥且露天栽培,根据盆中土壤缺水情况,不定期浇水(水中未检出Cd和As),使土壤含水量经常保持在田间持水量的80%左右.为防止污染物淋溶渗漏损失,在盆下放置塑料托盘并将渗漏液倒回盆中.植物生长时间为68d.113 样品分析植物样品分为根、茎、叶和籽实4部分,分别用自来水充分冲洗以去除粘附于植物样品上的泥土和污物,然后再用去离子水冲洗,沥去水分,在105℃杀青10min.之后,在70℃下烘干至衡重,将植物样品粉碎备用.植物样品采用H NO32HClO4法消化(二者体积比为3∶1),原子吸收分光光度计测定样品中的Cd含量.As的测定方法是在酸性条件下,加入20%硫脲(体积分数)将As5+还原成As3+,利用HG2AFS法测定[12].114 数据分析所有检测的数据都重复3次,在计算机上用Micros oft Excel2003进行平均值和标准差的运算,以Mean±S D形式表示.并利用最小显著性差异测验(LS D测验)进行植物样品差异显著性测验.2 结果与讨论211 球果 菜对Cd2As复合污染的生长反应从外观上看,Cd2As 复合污染条件下球果 菜生长正常,叶片没有出现如萎黄、缺绿等受胁迫症状.通过盆栽梯度实验发现,植株在株高和地上部干重对Cd2As复合污染的反应,表现为相一致的规律(图1和图2).在低浓度Cd2As复合污染条件下,球果 菜地上部的生长表现出一定促进效应.当Cd 、As复合浓度为10mgΠkg和50mgΠkg时,植物平均株高达到最大值,为3519cm;地上部干重也达到最大,是对照的111倍,达到212gΠ盆.随着土壤中施加Cd、As含量逐步提高,对球果 菜生长发育逐渐具有协同的抑制作用,表现在平均株高和地上部生物量均有所降低.然而,当Cd2As复合浓度水平增加到50mgΠkg和250mgΠkg时,株高和地上部生物量则显著降低(p<0105),尤其与对照相比,其株高和地上部干重分别下降了35%和43%.当Cd投加浓度相同时,与As浓度为50mgΠkg相比较,在As浓度为250mgΠkg条件下植物株高和生物量地上部干重总体有所降低,但不太明显(只有在Cd为50mgΠkg时,株高显著下降);同样,在同一As投加浓度条件下,随6531环 境 科 学28卷着Cd 投加浓度的上升,球果 菜生长发育受到抑制程度也基本上愈加明显.作为超富集植物,其在成熟期的生物量通常是一个重要的评价指标[13],尤其是地上部生物量,在土壤中重金属污染程度较高,但只要没有达到能够抑制植物生长的临界浓度条件下,植物地上部生物量一般不会下降;如果超过这一临界浓度,植物生长就会受到抑制,其叶色、株高等生长特性就会发生不同程度地变化,但最终集中反映在植物地上部生物量会显著降低[14,15].由图1和图2可知,在低浓度Cd 2As 复合污染条件下,由于Cd 、As 之间表现出拮抗作用,促进球果 菜的生长和发育,株高和生物量与对照相比均有所提高;当Cd 2As 复合浓度增加到25mg Πkg 和50mg Πkg 时,株高和生物量虽然减少但并不显著,同时在Cd 2As 复合污染浓度梯度处理中植物根部干重受影响的程度不太明显(p >0105)(如图2),说明球果 菜对Cd 2As 复合污染有很强的耐性 图1 Cd 2As 复合作用对球果 菜株高的影响Fig.1 E ffects of Cd 2As combined pollution onthe height of Rorippaglobosa图2 Cd 2As 复合作用对球果 菜生物量的影响Fig.2 E ffects of Cd 2As combined pollutionon the biomass of Rorippa globosa能力.对于Cd 2As 复合污染土壤的植物修复有一定的潜力,因为对重金属的强耐性能力是超富集植物修复复合污染土壤的一个重要特征[16].212 Cd 2As 复合污染对球果 菜体内Cd 积累的影响表1表示的是Cd 2As 复合污染处理下球果 菜根、茎、叶和籽实4个部分Cd 含量.由表1可看出,随着土壤中Cd 投加浓度的提高,植物体各部分Cd 含量均有所增加.植物体中Cd 含量表现为:叶片>茎>根>籽实,植物地上部含量大于地下部的浓度.当Cd 投加浓度≥25mg Πkg 时,叶片中Cd 含量高于镉超富集植物临界含量标准[7],尤其当Cd 2As 复合污染浓度为50mg Πkg 和50mg Πkg ,叶片中Cd 含量达到24118mg Πkg.可见,在较高的复合污染浓度条件下,球果 菜仍然有很强的Cd 吸收积累能力,对于Cd 2As 复合污染土壤的治理有一定的修复潜力.当Cd 为同一浓度处理水平情况下,高As (250mg Πkg )条件下抑制了植物对Cd 的吸收,植物体各部分中Cd 含量基本上都有所减少,但在Cd 为10mg Πkg 和25mg Πkg 浓度下,高As (250mg Πkg )浓度对Cd 的吸收抑制程度不太明显(p >0105);而当土壤中Cd 浓度达到50mg Πkg 时,与低As (50mg Πkg )浓度相比,高As (250mg Πkg )浓度下植株根、茎和叶中Cd 含量显著下降(p <0105).在Cd 单一处理浓度为25mg Πkg 时,与成熟期的球果 菜叶片中Cd 含量为15011mg Πkg [11]相比,在Cd 2As 复合污染处理水平为25mg Πkg 和50mg Πkg 时,叶片中Cd 的积累量却达到18010mg Πkg.说明了投加一定含量的As 能够表现出球果 菜对Cd的吸收机能的促进作用.周启星研究发现[17],当土壤环境中同时存在Cd 和As 时,As 有促进苜蓿吸收Cd 的功能;同时,As 也可促进水稻对Cd 的吸收,当土壤投加As 浓度为30mg Πkg 时,水稻根、茎叶、籽实Cd 含量水平随着土壤中As 含量的增加而增加[18].植物对Cd 的吸收能力,直接受土壤中投加的Cd 、As 浓度及其交互作用制约,这或许是Cd 2As 复合污染的实质[17].Cd 、As 之间交互作用表现为协同效应,Cd 、As 在协同作用时,Cd 3(AsO 4)2可透过植物细胞膜,因此在复合污染条件下,一定浓度的As 投加能够促进植物对Cd 的吸收和积累[18].转移系数(translocation factor ,TF )是指地上部元素的含量与地下部同种元素含量的比值[19],用来评价植物将重金属从地下向地上的运输和富集能力.转移系数越大,则重金属从根系向地上部器官转运能力越强.从表2可看出,Cd 2As 复合污染条件下球75316期孙约兵等:镉超富集植物球果 菜对镉2砷复合污染的反应及其吸收积累特征 表1 Cd2As复合污染对球果 菜体内Cd 积累量的影响T able1 E ffects of Cd2As combined pollution on Cd accumulation in Rorippa globosa处理Πmg・kg-1Cd积累量Πmg・kg-1根茎叶籽实Cd10+As501213±12.2bc3411±17.1cd6410±32.0cd1716±7.3a Cd10+As250712±2.1c1214±7.8d2713±16.2d217±0.1a Cd25+As504018±6.5b9214±27.8b18010±40.0ab3616±16.5a Cd25+As2503410±23.0bc5612±15.2bc12016±46.5bc4818±10.4a Cd50+As509116±12.9a17614±35.5a24118±7.1a7016±39.1a Cd50+As2504010±33.6b8013±21.5b14116±51.2b3010±5.2a 显著性水平(p)<0101<0101<0101>0105 F71123131615119果 菜对Cd仍然具有较强的转运和富集能力,各复合污染处理梯度水平下其TF>1.当Cd投加浓度相同时,球果 菜体内的转移系数随着As 投加浓度的增加而减少,说明高As抑制了Cd向地上部转移;但在Cd为50mgΠkg时,高As处理下植物的富集系数则大于低As条件下,这可能与高As显著抑制根部对Cd的吸收积累有关;与低As(50mgΠkg)相比,其Cd积累量减少了56%(见表1).当投加As浓度相同时,植物体Cd的转移系数随着土壤中Cd含量的增加而减少.表2 Cd2As 复合污染条件下球果 菜Cd、As的富集系数和转移系数T able2 Bioaccumulation and translocation factors of Cd and As inRorippa globosa under the Cd2As combined pollution处理Πmg・kg-1Cd As富集系数(BF)转移系数(TF)富集系数(BF)转移系数(TF)Cd10+As50 Cd10+As250 Cd25+As50 Cd25+As250 Cd50+As50 Cd50+As250614217712418418218512318414316216315012011012011013011015013016013014013 富集系数(bioaccumulation factor,BF)也称吸收系数,是指植物中某元素含量与土壤中元素含量之比[10].富集系数表征土壤2植物体系中元素迁移的难易程度,这是反映植物将重金属吸收转移到体内能力大小的评价指标.富集系数越高,表明植物地上部重金属富集质量分数大.从表2可看出,当Cd投加浓度相同时,高As(250mgΠkg)处理下表现出抑制球果 菜地上部对Cd的吸收效率;与低As(50mgΠkg)处理相比,球果 菜叶中Cd的积累量分别减少了58%、33%和42%;当投加As浓度相同时,植物体的富集系数则在Cd为25mgΠkg达到最大值,这可能与植物地上部的富集系数与其土壤中污染物投加含量有关,在适宜浓度下,能够促进植物对Cd的吸收积累,富集系数达到最大.总之,由于液泡的区室化作用和植物体内某些有机物(如MTs、PCs、有机酸等)对重金属的螯合作用,降低了重金属的毒性,从而能够促进植物体对重金属的吸收积累[20~22].在复合污染处理中,球果 菜体内都有较高的转移系数和富集系数,显示出对Cd 有很强的耐性和富集特征. 213 Cd2As 复合污染对球果 菜体内As积累的影响从图3可看出,Cd2As复合污染作用下球果 菜吸收的As主要集中在根部.在土壤Cd2As复合浓度为25mgΠkg和250mgΠkg时,根部As积累量达到11516mgΠkg,是地上部As 积累量的318倍.在Cd2As 复合污染处理条件下球果 菜对As吸收和转移能力很低,富集系数为011~013,转移系数为013~016 (表2),说明球果 菜对As有一定的排斥作用.在Cd投加浓度为10mgΠkg和25mgΠkg时,与低As(50 mgΠkg)处理相比,在高As(250mgΠkg)处理下植物根部和地上部As积累量显著增加(p<0105);在Cd处理浓度为50mgΠkg时,根部和地上部As积累量增加不太明显,可能在高Cd浓度下抑制了植物对As的吸收和积累.在As处理浓度为50mgΠkg条件下,根部和地上部As的积累量随着Cd处理浓度增加而增加;而在As处理浓度为250mgΠkg条件下,表现出低Cd处理浓度促进根部和地上部对As的吸收和积累,高Cd处理浓度抑制根部和地上部对As的吸收和积累.植物对重金属的排斥机制通常包括2个方面,一是减少根部对重金属的吸收,二是重金属在根部通过区室化保存,从而限制向地上部转移[20].排异植物最重要的特征就是植物体尤其是地上部重金属含量较低[23],排异植物是重金属污染土壤稳定修复较理想的修复植物;与超富集植物相反,以体外抗体为主导机制,排异植物减少其向地上部转移[24].球果 菜对As的排斥性虽然影响其对As的提取修8531环 境 科 学28卷复,但却是其对As 耐性能力的表现,根部吸收的As 只有少量转移到地上部,从而降低了As 对植物体的毒害.图3 Cd 2As 复合污染作用对球果 菜体内As 积累量的影响Fig.3 E ffects of Cd 2As combined pollution on Asaccumulation in Rorippa globosa214 Cd 2As 复合污染对球果 菜地上部和根系中Cd 、As 吸收量的影响超富集植物地上部积累大量的重金属有利于将其收获并妥善处理后,即可将该土壤中重金属移出土体,从而达到重金属污染治理与生态修复的目的[25],同时地上部高含量重金属还有一定的经济效益,如通过灰化冶炼提取重金属[26,27].球果 菜从土壤中吸收的Cd 主要积累在地上部,占整个植株的86%~99%(图4).当As 投加浓度相同时,地上部Cd 积累量随着土壤中投加Cd 含量增加而增加;当Cd 处理为同一浓度时,高As (250mg Πkg )处理能显著抑制地上部Cd 的积累,与低As (50mg Πkg )处理相比,球果 菜地上部Cd 积累量分别减少了69%、49%和59%.虽然球果 菜地上部As 含量小于根部,但由于地上部生物量大,其As 积累量则是根部的112~315倍.图4 Cd 2As 复合污染作用下球果 菜体内Cd 的积累量Fig.4 Accumulation of Cd in Rorippa globosa under theCd 2As combined pollution 植物有效数(plant effective number ,PE N )和重金属提取率(metal extraction ratio ,MER )被用来评价超富集植物修复重金属污染能力.其中,植物有效数是指植物体内积累1g 重金属所需要植物地上部或整个植株的数目[28].在Cd 2As 复合污染水平为50mg Πkg 和50mg Πkg 时,球果 菜体内积累1g 镉需要植物地上部或整个植株数分别为>12126和>10836,球果 菜体内积累1g 砷需要植物地上部或整个植株数分别为>33638和>240633.重金属提取率是指植物地上部积累重金属的量与土壤中同种重金属量的比值,即[29]:MER =c 地上部重金属×m 地上部干重c 土壤中重金属×m 土壤重量×100%(1) 在Cd 2As 复合污染不同浓度处理条件下球果 菜地上部对Cd 的提取率分别为0111%、0103%、0109%、0105%、0107%和0103%,可见由于高As 处理抑制了植物地上部对Cd 的吸收和积累,植物对Cd 的提取率也相应减少;在同样复合污染处理中,As 的提取率分别为01007%、01003%、01008%、01002%、01007%和01002%.总之,Cd 超富集植物球果 菜在Cd 2As 复合污染条件下对Cd 仍然有很强的吸收富集能力,对As 的吸收富集能力则较弱,这可能是植物体对不同污染物具有不同的耐性机制[10],从而最大程度地适应污染的胁迫以利于植物的生存.3 结论(1)在Cd 2As 复合污染条件下,两者低浓度处理由于Cd 、As 的拮抗作用,能够促进球果 菜的生长发育,株高和生物量都有所增加;同时,与Cd 单一污染相比,低As 浓度处理能够促进植物对Cd 的吸收和积累.高浓度处理,Cd 、As 两者能够表现出对球果菜有协同的抑制作用,不仅植株生长发育减缓,而且地上部Cd 的积累量也降低.作为镉超富集植物,在复合污染条件下,地上部能够积累大量的Cd ,其富集系数和转移系数都大于1.在Cd 2As 复合污染条件下,球果 菜对As 有一定的排斥作用,表现为植株地上部As 的积累量明显低于根部,其富集系数和转移系数都小于1.(2)在本实验中,镉超富集植物球果 菜对Cd 有超富集能力,同时对As 有一定的排斥作用,说明球果 菜对Cd 2As 复合污染有很强的耐性机制,对于修复Cd 2As 复合污染土壤有一定的潜力.作为农田杂草型超富集植物的球果 菜既具有野生植物的95316期孙约兵等:镉超富集植物球果 菜对镉2砷复合污染的反应及其吸收积累特征性状,也具有作物的某些栽培特征,有很强的生态适应性和竞争能力,生长迅猛,光合作用效率高,能够在较短时间内完成其生长史,抗逆性和抗虫害能力强,具有一般超富集植物无法比拟的优势,是Cd2As 复合污染土壤植物修复比较理想的材料.参考文献:[1]M cG rath S P,Zhao F J,Lombi E1Plant and rhiz osphere processesinv olved in phytoremediation of metal2contam inated s oils[J]1Plantand S oil,2001,232(1~2):207~2141[2]Reeves R D,Baker J M1M etal2accumulating plants[A].In:Phytoremediation of T oxic M etals:Using Plants to Clean Up theEnvironment[C].Eds.,Raskin H and Ensley B D.London:JohnW iley&S ons,Inc.,2000.193~230.[3]周启星,黄国宏.环境生物地球化学及全球环境变化[M].北京:科学出版社,2000.[4]孙铁珩,李培军,周启星.土壤污染形成机理与修复技术[M].北京:科学出版社,2005.[5]Chaney R L,M alik M,Li Y M,et al.Phytoremediation of s oilmetals[J].Environ Biotechnology,1997,8(2):279~284. [6]Salt D E,Sm ith R D,Raskin I.Phytoremediation[J].Annu RevPlant Physiol Plant M ol Biol,1998,49(5):643~668.[7]Baker A M J,Brooks R R.T errestrial higher plants whichhyperaccumulate metallic elements—a review of their distribution,ecology and phytochem istry[J].Biorecovery,1989,1(4):811~826.[8]Brooks R R.Plants that hyperaccumulate heavy metals:their role inphytoremediation,m icrobiology,archaeology,m ineral explorationand phytom ining[M].Ox ford,UK:CAB International,1998. [9]魏树和,周启星,王新,等.一种新发现的镉超积累植物龙葵(Solanum nigrum L.)[J].科学通报,2004,49(24):2568~2573.[10]周启星,宋玉芳.污染土壤修复原理与方法[M].北京:科学出版社,2004.[11]W ei S H,Zhou Q X.Phytoremdiation of cadm ium2contam inateds oils by Rorippa globosa using tw o2phase planting[J].Environ SciP ollut Res,2006,13(3):151~155.[12]W ei C Y,Chen T B.Arsenic accumulation by tw o brake fernsgrowing on an arsenic m ine and their potential in phytoremediation[J].Chem osphere,2006,63(6):1048~1053.[13]Ernst W H O,Nelissen H J M.Life2cycle phases of a zinc2andcadm ium2resistant ecotype of Silene vulgaris in risk assessment ofpolymetallic m ine s oils[J].Eviron P ollution,2000,107(3):329~338.[14]W ei S H,Zhou Q X,W ang X,et al.A newly2discovered Cd2hyperaccumulator Solanum nigrum L.[J].Chin Sci Bulletin,2005,50(1):33~38.[15]W ei S H,Zhou Q X.Identification of weed species withhyperaccumulative characteristics of heavy metals[J].Progress inNatural Science,2004,14(6):495~503.[16]Y ang X E,Long X X,Y e H B,et al.Cadm ium tolerance andhyperaccumulation in a new Zn2hyperaccumulating plant species(Sedum alfredii Hance)[J].Plant and S oil,2004,259(122):181~189.[17]周启星.镉2砷污染苜蓿的联合效应及机理的研究[J].应用基础与工程科学学报,1994,2(1):81~87.[18]周启星.复合污染生态学[M].北京:中国环境科学出版社,1995.[19]Fayiga A O,M a L Q,Cao X D,et al.E ffects of heavy metals ongrowth and arsenic accumulation in the arsenic hyperaccumulatorPteris vittata L.[J].Environmental Pllution,2004,132(2):289~296.[20]BarcelóJ,P oschenrieder C.Phytoremediation:principles andperspectives[J].C ontributions to Science,2003,2(3):333~344.[21]孙瑞莲,周启星.高等植物重金属耐性与超积累特性及其分子机理研究[J].植物生态学报,2005,29(3):497~504. [22]Alkorta J,Hernández2Allica J M,Becerril I,et al.Chelate2enhanced phytoremediation of s oils polluted with heavy metals[J].Environmental Science and BioΠT echnology,2004,3(1):55~70.[23]Baker A J M.Accumulators and excluders2strategies in the responseof plants to heavy metals[J].Journal of Plant Nutrient,1981,3(4):643~654.[24]顾继光,周启星,王新.土壤重金属污染的治理途径及其研究进展[J].应用基础与工程学学报,2003,11(2):143~151.[25]Anders on T A,G uthie E A,W alton B T.Bio2remediation in therhiz ophere[J].Envion Sci T echnol,1993,27(4):673~676. [26]Brooks R R.Plants that hyperaccumulate heavy metals:their role inphytoremediation,m icrobiology,archaeology,m ineral explorationand phytom ining[M].Ox ford,UK:CAB International,1998. [27]Anders on C W N,Brooks R R,Chiarucci A,et al.Phytom ining fornickel,thallium and g old[J].Journal of G eochem ical Exploration,1999,67(123):407~415.[28]G arcía G,Faz#,Cunha M.Performance of Piptatherum miliaceum(Sm ilo grass)in edaphic Pb and Zn phytoremediation over a shortgrowth period[J].International Biodeterioration&Biodegradation,2004,54(223):245~250.[29]M ertens J,Luyssaert S,Verheyen e and abuse of trace metalconcentrations in plants tissue for biom onitoring and phytoextraction[J].Environmental P ollution,2005,138(1):1~4.0631环 境 科 学28卷。