内质网的结构和功能
内质网

(4) 蛋白质在粗面内质网上继续合成:
1. 处于暂停状态的蛋白质合成重新开始。 2. 信号肽反向贴附转移器,通过疏水节段插入内质网膜。 3. 肽链以袢环形式进入内质网腔,直至合成完整多肽链。 4. 内质网膜上的信号肽,被腔面的信号肽酶切掉降解。 5. 核糖体脱离内质网,重新加入新的蛋白质合成。
脂类去向: 一、嵌入到内质网脂类双层中,补充自身膜损失。 二、输送到其它膜性细胞器,补充其他膜相损失。
(六)贮存钙离子
细胞膜和ER膜上有Ca 2+ -泵,能将胞质中的Ca2+泵出胞外或 贮存在内质网(Ca2+-库)中,使细胞质中的Ca2+和细胞外及Ca2+ -库的Ca2+保持极大的浓度差。
内质网在细胞中的分布图
鸡 输 卵 管 细 胞 粗 面 内 质 网
小鼠睾丸间质细胞滑面内质网
Endoplasmic reticulum of living CHO cells displayed by green fluorescent protein(GFP)
二)内质网的分类
粗面内质网(rER) 滑面内质网(sER)
(3) 信号肽被引导到内质网膜上:
SRP受体也称停靠蛋白(docking protein): 膜整合蛋白,暴露于内质网膜表面。
过程: 1. SRP与膜上受体结合,SRP-核糖体复合体被引导到ER 膜上,与ER膜接触,核糖体结合核糖体受体。 2. 核糖体与ER膜结合成为附着核糖体,SRP从核糖体上 解离,参加下次循环。
SRP receptor binding site
Three functional binding sites
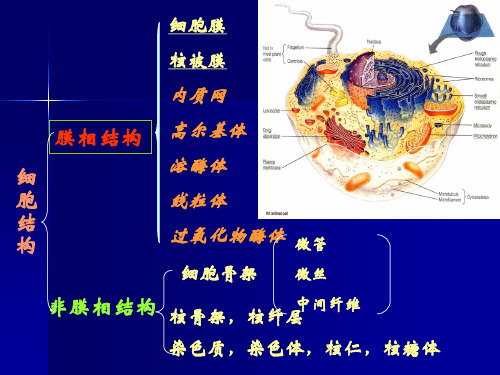
【生物知识点】内质网的结构和功能

【生物知识点】内质网的结构和功能
内质网分为粗面内质网和滑面内质网两种基本类型。
粗面内质网有核糖体颗粒附着,主要功能是外输性蛋白质及多种膜蛋白的合成、加工及转运。
滑面内质网表面光滑无核糖体附着主要参与类固醇、脂类的合成与运输,糖代谢及激素的灭活。
内质网是细胞内除核酸以外的一系列重要的生物大分子,如蛋白质、脂类(如甘油三酯)和糖类合成的基地。
滑面内质网还具有解毒功能,如肝细胞中的滑面内质网中含有一些酶,用以清除脂溶性的废物和代谢产生的有害物质。
依据内质网膜外表面是否有核糖体附着,通常将内质网分为粗面内质网和滑面内质网两种基本类型。
粗面内质网:主要形态特征为网膜胞质面有核糖体颗粒附着,并由此得名。
粗面内质网在形态上多为排列整齐的扁囊,在功能上,主要与外输性蛋白质及多种膜蛋白的合成、加工及转运有关。
因此,在具有分泌肽类激素或蛋白质功能的细胞中,粗面内质网发达,如胰腺细胞、浆细胞等。
在未分化或低分化的细胞中相对不发达,如胚胎细胞、肿瘤细胞等。
滑面内质网:电镜下呈光滑的小管、小泡样网状结构,常与粗面内质网相通。
滑面内质网是一种多功能的细胞器。
在不同细胞、同一细胞的不同发育阶段或不同生理时期,其形态结构、数量、细胞内空问分布及发达程度差异较大,而且常表现出不同的功能特性。
如睾丸间质细胞、卵巢黄体细胞及肾上腺皮质细胞中有大量的滑面内质网,是与其合成类固醇激素的功能有关;肝细胞中丰富的滑面内质网与其减毒功能有关;在平滑肌和横纹肌中的滑面内质网特化为肌质网,通过储存及释放Ca2+调节肌肉收缩。
感谢您的阅读,祝您生活愉快。
细胞内质网和高尔基体的功能

细胞内质网和高尔基体的功能细胞内有许多重要的细胞器,其中包括质体、线粒体、溶酶体、核糖体和核糖体等。
质网与高尔基体是质体中非常重要的细胞器,它们对于细胞内物质的合成、转运和分解等起着重要的作用。
一、细胞内质网细胞内质网(ER)是一种有网状结构的膜系统,又称内质网,主要包括粗面内质网和平滑内质网两种类型。
1. 粗面内质网粗面内质网的表面布满有许多小突起,其中富含有许多核糖体,主要参与细胞蛋白质的合成和转运。
当细胞内需要大量合成蛋白质时,粗面内质网的数量会不断增加,并且在蛋白质的合成过程中起到了重要的作用。
2. 平滑内质网平滑内质网的表面没有粗面内质网那么颗粒状,而是长成一片平滑的薄膜状。
平滑内质网包含一系列重要物质的合成如脂肪酸和甾体激素等,也是胆固醇合成和代谢的重要场所。
此外,平滑内质网还参与钙离子的调节和质膜的代谢。
二、高尔基体高尔基体与内质网一样,都是一种膜状结构,且其中含有类似于内质网小泡的中心内腔和许多层叠的薄膜结构。
高尔基体是细胞内分泌系统中的重要器官,主要负责对细胞内物质的合成、后加工处理、聚合和分解等过程。
高尔基体可以分为Golgi近浅面、中央层和深浅液泡依次组成。
其中,近浅面主要负责将前体蛋白和小泡从内质网向高尔基体运输;中央层则参与前体蛋白的后加工;深浅液泡负责将经过后加工的成品物或嵌合了异物抵抗某种疾病的核糖体运出细胞,或进入某个胞吞作用的列表或其他细胞器内部。
三、质网与高尔基体的协作质网和高尔基体在细胞内起着密不可分的作用。
在细胞分泌过程中,由内质网合成并转运至粗面内质网中的前体蛋白,再到高尔基体中进行所需的后加工处理,之后再通过高尔基体中的深浅液泡运输至目的地,以完成细胞分泌的各项职能。
此外,质网还能够与高尔基体共同参与对细胞外文化环境的调节。
当细胞内部受到外界压力的影响后,质网和高尔基体会协同作用调节细胞内物质的合成和外排,保证细胞处于一个最佳的状态下。
总之,质网和高尔基体是细胞内非常重要的细胞器。
内质网

UPR:蛋白质在 ER 内的正确折叠需要许多 分子伴侣的协助,当 ER 中未折叠或错误折叠 的蛋白增多时,应激信号通过内质网膜传递 到细胞核,继而引起一系列特定的靶基因转 录和蛋白质翻译水平下调,以使细胞继续存 活,这种反应称为未折叠蛋白反应(UPR)。
(2)内质网超负荷反应(EOR)
ER 的钙稳态
ER是细胞内的钙储存库。 钙在内质网内以钙离子或钙结合蛋白的形式存 在。而钙结合蛋白与蛋白质折叠及修饰有关, ER的分子伴侣和折叠酶大多数是强力钙离子结 合蛋白。钙离子作为信号转录因子,发挥两种 作用:一是由内质网腔释放入胞质作为第二信 使;二是在内质网腔内调节钙依赖蛋白酶的活 性。所以内质网腔内钙离子的稳态对维持细胞 的功能至关重要。
EOR 是指正确折叠蛋白在内质网上过度积聚 时引起的内质网超负荷,从而导致一系列信 号物质的激活,EOR 也使机体自我保护性反
应之一。
(3)固醇调节级联反应
• • 内质网膜上含有固醇调节元件结合蛋白(SREBP)
,其无活性的前体大分子与内质网膜和核膜相连,在 内质网应激时内质网膜上的固醇耗竭,导致 SREBP 与 内质网膜和核膜结合的裂解,从而激活引导固醇生物 合成的起动子,即固醇调节因子,引起脂肪酸和胆固 醇的合成增加。通过恢复蛋白质正确构象,减少蛋白 质在内质网腔中的堆积,从而对细胞起保护作用,但 内质网应激反应时间过长或强烈可活化继而引起级联 反应,导致细胞凋亡。
Thank you !
三、内质网在细胞凋亡中的作用
据研究表明:内质网是细胞凋亡调节中的重要环 节。与内质网应激有关。
内质网应激:当细胞受到某些打击(如缺氧、药 物毒性等)后,内质网腔内氧化环境被破坏,钙代 谢失调,ER功能发生紊乱,突变蛋白质产生或者蛋 白质二硫键不能形成,引起未折叠蛋白或错误折叠
第十一章内质网
(四)膜脂的合成
大部分膜脂在糙面内质网膜组装。膜脂的合成与蛋 白质的合成、运输过程密切相关。是细胞内唯一合成膜 脂的细胞器,作为细胞膜成分的“源泉”。
脂类去向:
一、嵌入到内质网脂类双层中,补充自身膜损失。
二、输送到其它膜性细胞器,补充其他膜相损失。
①胞液面磷脂合成:
新合成的脂类分子最初只嵌入内质网脂双层的细胞质 基质面。
分类: ①粗面内质网(rough endoplasimic reticulum,rER) ②光面内质网(smooth endoplasimic reticulum,sER)
粗面内质网(rER) 滑面内质网(sER)
RER has ribosomes on the cytosolic side of continuous, flattened sacs(cisternae); SER is an interconnecting network of tubular membrane elements.
真核细胞膜结合区室的主要功能
细胞器
胞质溶胶 细胞核 内质网 高尔基 溶酶体 内体 过氧化物酶体 线粒体
主要功能
代谢的主要场所,蛋白质合成部位 基因组存在场所,DNA,RNA的合成地 脂的合成场所,蛋白质合成和集散地 蛋白质和脂的修饰,分选和包装 细胞内的降解作用 内吞物质的分选 毒性分子的氧化 通过氧化磷酸化合成ATP
②一个位于内部的内信号肽疏水序列
(2)多次跨膜蛋白整合到ER膜的方式:
具有两个以上起始转移序列和停止转移序列,多 肽链前后来回重复的通过脂双层。
(二).蛋白质的修饰与加工 包括糖基化、羟基化、酰基化、二硫键形成等,其中最主 要的是糖基化。 糖基化的作用: ①使蛋白质能够抵抗消化酶的作用; ②赋予蛋白质传导信号的功能; ③某些蛋白只有在糖基化之后才能正确折叠。
内质网和高尔基体的结构和功能

内质网和高尔基体的结构和功能内质网和高尔基体是细胞内最重要的器官之一。
它们分别位于细胞质中,并且彼此之间紧密相连通。
在细胞的代谢和生长过程中,这两个器官发挥着重要的生物学功能。
本文将就它们的结构和功能进行探讨。
内质网,又称为内质膜系统,是细胞质中最重要的细胞器之一。
它由许多同心环状的膜片构成,这些膜片可以从细胞膜、核膜和高尔基体膜等地方形成。
内质网主要分为粗面内质网和平滑内质网两种。
粗面内质网上有大量的核糖体附属,因此也被称为囊泡样内质网。
而平滑内质网则没有核糖体,外形较为扁平。
两种结构的功能也并不相同。
粗面内质网的作用主要与蛋白质的合成有关。
刚开始的时候,氨基酸通过核孔进入细胞质,然后与核糖体以及粗面内质网上的肽链合并。
合并过程中需要包括蛋白质组装以及折叠等过程。
随后,成型的蛋白质通过囊泡的运输进入高尔基体。
在这个过程中,粗面内质网还会为肝细胞合成胆固醇,和肾细胞合成荷尔蒙等。
所以说,粗面内质网在蛋白质的合成及附加多项代谢活动方面都发挥着重要的生物学功能。
平滑内质网的作用则相对单一些。
它负责合成细胞中许多脂质物质,包括各种脂肪和糖类等。
此外,平滑内质网在肝组织内还负责解毒和代谢过度酒精等化学物质。
这些功能都是和细胞内代谢过程直接相关的。
高尔基体是一个通过囊泡运输与内质网联系,并且与腹泡颗粒一同参与表面组织的主要细胞器之一。
高尔基体位于细胞质中,其中含有大量酵素,同时也是蛋白质和糖的合成和加工中心。
高尔基体上的许多囊泡是供运输细胞膜和蛋白质使用的,这些囊泡是由内质网上的蛋白质分解产生的。
高尔基体主要分为两个部分:囊泡样和网状结构。
囊泡样高尔基体主要负责细胞内的物质运输。
其中包括运输组织、荷尔蒙和一些卟啉物质等。
而网状高尔基体则负责蛋白质和糖的平面合成和附加代谢。
总体来说,内质网和高尔基体结构简单、功能强大,是细胞最主要的两个器官之一。
在细胞中,这两个器官并不是互相孤立的,而是相互联系、互相配合的。
内质网的结构域功能

第三节ER的化学组成
1.与氧化反应电子传递相关的酶系 NADPH-细胞色素c还原酶、Ctyb5、细胞色素P-450 2.与脂类物质代谢功能反应相关的酶类 脂肪酸CoA连接酶 3.与碳水化合物代谢功能反应相关的酶类 葡萄糖-6-磷酸酶 4.参与蛋白质加工转运的多种酶类
核糖体结合糖蛋白(ribophorin)只分布在RER,P450酶系只分布在SER。
4、环孔板(annulate lamellae)
一、AL 的形态结构及分布特征
•
AL 在形态上与内质网十分相似,其 基本结构单位类似内质网的潴泡,是 由双层膜围成的扁平囊状结构。 • 但 AL 与内质网也存在明显区别,即 双层膜上具有规律性分布的、与核孔 十分相似的膜孔结构。膜孔上常有弥 散的非膜性无定形物质分布。 •因此在形态结构上 AL 兼有内质网和 核膜的特征。
微粒体仍具 有内质网的 基本特征; 粗面微粒体 仍保留RER 所具备的功 能
常用大鼠肝细胞 ER 的微粒体 来分析其化学组成。
φ100nm
第二节内质网的种类与结构特点
3、微粒体
A B
A: 粗面微粒体(来自RER)
B: 滑面微粒体(多来自SER,部分来 自细胞膜和高尔基体或其他膜性碎片
第二节内质网的种类与结构特点
酶蛋白含量多是内质网膜区别于高尔基体膜囊和 质膜的主要标志
第四节ER的功能
一、RER的功能
1.进行蛋白质合成
2.合成蛋白的修饰与加工
3.膜的生成
4.物质运输
第四节ER的功能
一、RER的功能
(一)进行蛋白质合成
蛋白质合成两个特定区域
1、游离核糖体(合成结构蛋白)
2、RER表面的核糖体(非膜蛋白、镶嵌蛋白)
内质网的结构和功能

内质网的结构和功能
1. 结构:Endoplasmic reticulum(内质网)是由一系列的膜蛋白组成的有机结构,可以形成交错的平台,连接到细胞核和胞浆中的其他机制。
可能包括多孔充气体系统、纤维结构和膜循环链,以及纤维微管等结构。
2. 功能:内质网有多种功能,但其主要功能是负责蛋白质的合成和细胞内代谢的调节。
它也参与建立和维护细胞形态,同时会参与膜组分的分泌和转运。
此外,内质网还可以参与脂质的生物合成、调节激素信号转导、参与核酸合成和代谢,以及细胞凋亡过程中重要蛋白质的注射和递送过程等。
细胞质内主要的细胞器及功能

细胞质内主要的细胞器及功能1. 内质网(endoplasmic reticulum):内质网是一个复杂的膜系统,分为粗面内质网和平滑内质网。
粗面内质网上有许多附着的核糖体,负责合成蛋白质并将其包装成囊泡。
平滑内质网则参与脂质代谢、糖代谢和钙离子的贮存等。
内质网在细胞内运输、折叠和修复蛋白质等方面起着重要作用。
2. 核糖体(ribosome):核糖体是细胞内的蛋白质合成工厂,由RNA和蛋白质组成。
核糖体通过翻译mRNA上的遗传密码来合成蛋白质。
核糖体可以存在于细胞质中,也可以附着在内质网上。
3. 高尔基体(Golgi apparatus):高尔基体由一系列扁平的膜囊泡组成,主要负责蛋白质的加工、修饰和分拣。
高尔基体接收从内质网运输过来的蛋白质,经过一系列的修饰,如糖基化、磷酸化等,然后分拣并运输到细胞的不同位置或分泌出细胞外。
4. 溶酶体(lysosome):溶酶体是一种含有多种水解酶的囊泡,主要负责细胞内外废物的降解和消化。
溶酶体可以分解细胞内的蛋白质、核酸、糖类等有机物质,同时也可以降解细胞外的细菌和病毒等。
5. 线粒体(mitochondria):线粒体是细胞内的能量合成器,主要参与细胞的呼吸作用和能量代谢。
线粒体内有许多内膜,形成了许多被称为氧化磷酸化系统的结构,通过这些结构,线粒体可以将有机物质氧化成二氧化碳和水,并转化为细胞所需的能量。
6. 中心体(centrosome):中心体是一对圆柱形的微管结构,位于细胞质中,起着细胞分裂和形成纺锤体的重要作用。
中心体内含有许多微管,可以形成纺锤体,参与细胞的有丝分裂过程。
7. 高尔基体体(peroxisome):高尔基体体是一种具有氧化功能的细胞器,主要参与脂肪酸的代谢和氧化反应。
高尔基体体内含有多种氧化酶,可以氧化有机物质,并产生氢过氧化物等物质。
8. 核仁(nucleolus):核仁是位于细胞核内的一个小体,主要参与核糖体的组装和合成。
核仁内含有DNA、RNA和蛋白质等物质,核仁可以合成和组装核糖体的RNA和蛋白质。
内质网

内质网素被作为内质网的标志性分
子伴侣。
SRP-核蛋白体 复合结构
分泌性 蛋白
1、分泌性蛋白质转入内质网合成的过程:
2、跨膜驻留蛋白的插入与转移
单次跨膜蛋白插入内质网膜的2种机制: 新生肽链协同翻译插入机制 起始转移信号肽——引导肽链转移 停止转移肽——使移位子由活性状态转为钝化 状态而终止肽链的转移。 内信号肽介导的内开始转移肽插入转移机制 内信号肽——位于多肽链中的信号肽序列 多次跨膜蛋白质的转移插入 基本机制与单次跨膜蛋白质插入差不多,一般 认为以内信号肽作为开始转移信号的。
2.下列关于内质网蛋白的叙述,哪项错误( ) A插入内质网膜成为跨膜蛋白 B留在内质网 C运送到线粒体 D运送到溶酶体 3.下列蛋白质合成后的加工、修饰的叙述,下 列哪项不正确( ) A正确的折叠 B二硫键的修饰 C糖基化 D蛋白原的水解 E分泌 4.由信号肽介导进入内质网的蛋白有( ) A溶酶体酶 B核蛋白 C分泌性蛋白 D内质网膜蛋白 E过氧化物酶体酶
发现过程 组成与形态 基本类型 功能 病理变化
内质网的发现
一.1945年,Porter等人在培养的小鼠 成纤维细胞中初次观察到细胞质内部 有网状结构,将之名为内质网(ER)。
二.1954年,研究证实:内质网是一 类由大小、形态各异的膜性囊泡所构 成的细胞器。
在电镜下观察,内质网(ER)由一层单位 膜构成的形状大小不同的扁平囊、小囊及 小管组成,并连成一个连续网状管道系统。
1内质网的结构和功能

拓展资料第一单元第二章第一节济南三中邱晨(整理)1.内质网的结构和功能内质网是由Porter等人在1945年发现的。
他们利用电镜在成纤维细胞中观察到一些形态和大小略有不同的网状结构,并集中在内质中,因此将这些结构称为内质网。
内质网是由一层膜形成的囊状、泡状和管状结构,并形成一个连续的网膜系统。
内质网通常占细胞的生物膜系统的一半左右,占细胞体积的10%以上。
根据内质网上是否附有核糖体,将内质网分为粗面内质网和光面内质网两类。
粗面内质网多呈大的扁平膜泡,排列整齐。
它是核糖体和内质网共同组成的复合结构,普遍存在于细胞中,特别是合成分泌蛋白的细胞。
在结构上,粗面内质网与细胞核的外层膜相连。
无核糖体附着的内质网称为光面内质网,通常为小的管状和小的泡状,广泛存在于各种类型的细胞中。
光面内质网是脂质合成的重要场所。
内质网可通过出芽方式,将合成的蛋白质或脂质转运到高尔基体。
2.高尔基体的结构和功能高尔基体是意大利科学家高尔基(C.Golgi)在1898年发现的,是普遍存在于真核细胞中的一种细胞器。
在电镜下观察到,由一些排列较为整齐的扁平膜囊堆叠在一起,构成了高尔基体的立体结构。
扁平膜囊多呈弓形,也有的呈半球形,均由光滑的膜围绕而成。
在扁平膜囊外还包括一些小的膜泡。
整个高尔基体结构分为形成面和成熟面,来自内质网的蛋白质和脂质从形成面逐渐向成熟面转运。
高尔基体与细胞的分泌功能有关,能够收集和排出内质网所合成的物质,它也是聚集某些酶原的场所,参与糖蛋白和黏多糖的合成。
高尔基体还与溶酶体的形成有关,并参与细胞的胞吞和胞吐作用。
3.溶酶体的结构和功能溶酶体是动物细胞中一种由膜构成的细胞器,呈小球状,外面由一层非渗透性单位膜包被。
溶酶体是一种动态结构,它不仅在不同类型细胞中形态大小不同,而且在同一类细胞的不同发育阶段也不相同。
溶酶体的主要功能是消化作用,其消化底物的来源有三种途径:一是自体吞噬,吞噬的是细胞内原有物质;二是吞噬体吞噬的有害物质;三是内吞作用吞入的营养物质。
研究内质网应激相关蛋白质的结构和功能

研究内质网应激相关蛋白质的结构和功能内质网是细胞内一种重要的膜系统,它负责合成、折叠、修饰和运输蛋白质。
通过完成这些任务,内质网有助于保持细胞的内外环境平衡,并发挥多个细胞生理学功能。
然而,内质网也面临着各种压力和挑战,例如病原体感染、药物毒性和代谢异常等因素,这些因素都可能导致内质网的应激反应,促使内质网产生一系列的结构和功能改变,从而影响细胞和个体的健康状态。
本文将介绍内质网应激相关蛋白质的结构和功能,探讨它们对细胞应激的响应机制及其在多个疾病中的作用。
一、内质网应激反应机制内质网应激反应是一种由多个信号通路和蛋白质调节器所调控的生物学过程。
在正常情况下,内质网中的蛋白质会经过正确的折叠和修饰,并在内质网内被运输到它们要去的地方。
但是,当内质网处于应激状态时,细胞会启动多个应激反应通路,以增强或恢复内质网的正常功能。
其中,IRO1(inhibitor of RNA polymerase I)和PERK(PKR-like ER kinase)通路是比较典型的内质网应激反应通路。
内质网应激反应的信号传导机制包括如下几个步骤:①由IRE1、PERK或ATF6等膜蛋白感受内质网应激信号,②感受到应激信号的膜蛋白自我磷酸化,激活相应的内生性激酶活性,③磷酸化的IRE1和PERK可以分别调节XBP1和ATF4等转录因子,促使它们在核内启动基因表达,并促进细胞存活。
这些通路的成熟度和细胞内信号转导机制,是内质网应激机制的重要组成部分。
二、内质网应激相关蛋白质的结构和功能为了更好地适应压力环境,内质网在多个不同的性质下都会形成一些新的蛋白质,这些蛋白质被称为内质网应激相关蛋白质。
内质网应激相关蛋白质大多是转录因子、分子伴侣、酶和调节蛋白等,这些蛋白质在应激反应机制中具有十分重要的功能。
1. XBP1XBP1是内质网应激反应中最为著名的转录因子之一,它的发现和研究也为内质网应激反应的理解提供了很多的帮助。
XBP1基因编码的蛋白质可以分为XBP1u 和s两种类型,在进行应激反应后,XBP1的切割酶IRE1会切割s和XBP1u两种转录因子中的一个,切割后的XBP1s能够转录大量抗应激蛋白,并在细胞内调节直接参与应激反应的一系列靶基因。
内质网

3、参与糖原代谢:在肝细胞中,糖原颗粒常 与光面内质网伴存。当糖原丰富时光面内质 网常被遮盖而不易分辨,当饥饿使糖原减少 时光面内质网就明显可见。对肝细胞光面微 粒体分析表明,其膜上有葡萄糖-6-磷酸酶, 这种酶与糖原代谢有关。
4、参与药物代谢和细胞解毒:一些脂溶性代 谢产物和药物,有一定毒性,不易直接被排 出,需在肝细胞中经过氧化、还原、水解和 结合等生物转化过程,降低其毒性,且成为 易溶于水的物质利于排出体外。光面内质网 上有混合功能氧化酶系统,如NADPH细胞色 素C还原酶、细胞色素P450等。当给大鼠大 量注射脂溶性苯巴比妥时,可见肝细胞的光 面内质网显著增生,其内参与药物代谢的酶 活性也增强,表明肝细胞的光面内质网与药 物代谢有密切关系。
2、蛋白质的修饰与加工:包括糖基化、酰基化、 氨基酸羟基化以及肽链间的二硫键形成等。糖基化 是内质网中最主要的蛋白修饰。糖基化在糖基转移 酶作用下发生,包括:(1)N-连接的糖基化( Nlinked glycosylation)。即N-乙酰葡萄糖胺与天冬 酰胺残基的—NH2连接,在粗面内质网中进行。(2) O-连接的糖基化( O- linked glycosylation)。即半 乳糖或N-乙酰半乳糖胺与丝氨酸、苏氨酸、羟脯氨 酸、羟赖氨酸残基—OH连接,主要在高尔基体内进 行。酰基化主要指软脂酸与半胱氨酸共价结合,发 生在内质网的细胞质基质侧。
内质网的形态结构 内质网的主要功能 内质网与基因表达的调控
内质网
1945年Porter首先在电子显微镜下观察成纤维细胞 时发现一种网状结构位于细胞质内,故名为内质网 (endoplasmic reticulum,ER)。后来发现内质网 可存在于细胞质的任何部位,内质网广泛分布于各 种细胞中,它是一个由单位膜包围的封闭结构,这 些互相连续的封闭结构在形态上呈现为囊状、管状 和泡状,在细胞质内形成一个三维网状结构。内质 网膜所包围的空间称内质网腔,内质网腔与胞质溶 胶之间物质交换必须通过内质网膜进行。内质网与 高尔基体之间的物质交换是通过运输小泡进行的。
细胞内质网的形态结构和功能研究

细胞内质网的形态结构和功能研究细胞内质网(Endoplasmic Reticulum)是一种重要的细胞质结构,扮演着细胞代谢、物质转运和分泌等多种生理功能的关键角色。
这一质网结构在细胞表现出极为多样化的形态和功能,迄今为止研究尚未完全深入。
本文将从光学显微镜下的细胞质结构描述到EM级别下的结构细节,并介绍质网发生的各种生理反应,综合阐述对细胞内质网的新一代结构和功能研究。
一、细胞内质网的形态结构细胞内质网是一种延伸分支的薄膜状结构,呈现出时空多样的变化和极其复杂的拓扑关系。
从形态上分,细胞内质网可以分为光滑内质网(Smooth Endoplasmic Reticulum,SER)和粗面内质网(Rough Endoplasmic Reticulum,RER)。
SER相对平整,质膜上缺乏与核糖体的结合系,涉及到乳酸脱氢酶、细胞色素P450酶等的储藏与水解功能,与一些在脂类代谢方面有着密切联系。
RER表面则有着众多的细胞核糖体附着着,参与到蛋白质的翻译、合成等诸多生物学任务和分泌等生理过程。
二、细胞内质网的细节结构TEM技术是研究细胞内膜体系最有效的手段之一,通过TEM可以更加清晰地描绘出细胞内质网的细节结构。
一部分ER膜质被腔室分成了大小不一、形状参差不齐的颗粒(cisternae),像陡峭谷壑般被逐层悬空或重叠在一起,颗粒之间的尺延伸衔接呈曲线状,形成了一个细胞内的巨型网状结构。
三、细胞内质网的功能特性细胞内质网功能多样,与蛋白质的翻译、配体结合、各种细膜蛋白的产生、分泌有着密切的关系。
同时质网也与细胞的代谢有关,如对钙离子、磷脂分子、甘油磷酸二酯等的参与和分解。
四、细胞内质网的分子调节近年来,随着单细胞测序的技术不断发展,人们开始深入了解到生物体在分子水平上调节内质网机制的分子调控过程,如成簇计算机学习技术的应用,对内质网网络的谱系分析等,都帮助学界了解细胞内质网的复杂调控机制,对于开发高效的分子治疗手段和生物医学技术具有极其重要的应用价值。
内质网

蛋白质转入内质网合成涉及的4种成分:
• 信号肽:被合成肽链N端的一段特殊氨基酸 序列,是引导肽链进入内质网腔的一段序 列,又称开始转移序列。
• 信号识别颗粒(SRP):由6种结构不同的 多肽组成,SRP与信号肽结合,导致蛋白 质合成暂停。
• 信号识别颗粒受体(SRP-R):内质网上的膜 整合蛋白,可与SRP特异结合。
网,完成于高尔基体。
不一定完成于高尔基体
3.细胞内蛋白质质量监控的重要因子为
。
4.在多次跨膜蛋白肽链上,常常有 个或者 个
以上的疏水性起始转移肽和停止转移肽。
5.滑面内质网合成脂质通常在 侧
6论述内质网和线粒体电子传递链的不同。
见书P106页
3.下列蛋白质合成后的加工、修饰的叙述,下
列哪项不正确( )
A正确的折叠
B二硫键的修饰
C糖基化
D蛋白原的水解
E分泌
4.由信号肽介导进入内质网的蛋白有( )
A溶酶体酶
B核蛋白 C分泌性蛋白
D内质网膜蛋白
E过氧化物酶体酶
1.由内质网合成并运输到高尔基体、溶酶体、
细胞膜以外的蛋白质大多是糖蛋白
2.N-连接和O-连接的寡糖修饰都是起始于内质
滑面内质网的功能
参与脂质的合成和转运
ER合成的膜脂以出芽小泡的方式转运至高尔基体 ,溶酶体和质膜上,或以水溶性磷脂转换蛋白为载 体,与之结合成复合体进入细胞质,通过自由扩散 转至其他膜上。
参与糖原的代谢:糖元的分解
是否参与糖元的合成,目前还不确定。
细胞解毒的主要场所
基本机制:催化多种化合物的氧化或羟化
糖基化:单糖或者寡糖与蛋白质之间通过共 价键的结合形成糖蛋白的过程。
催化以上过程的糖基转移酶,是存在于粗面 内质网网膜腔面的一种膜整合蛋白。
内质网

与内质网有关的疾病
相关疾病
• 研究发现,内质网应激是慢性代谢疾病的 重要标志,也是连接免疫系统与代谢系统 的桥梁。代谢系统的细胞作为合成代谢活 跃的细胞,具有高度发达的内质网。内质 网被视为“代谢感受器”,与内分泌网络 建立了广泛而密切的联系。在营养过剩状 态下,内质网发生应激,成为触发代谢疾 病的重要因素。下面主要从炎症和细胞凋 亡的角度阐述内质网应激的致病机制。
介绍: 内质网是具有重要生理功 能的细胞器,是真核细胞蛋白质 合成、钙离子贮存以及脂质生成 的主要场所。内质网的功能状态 对蛋白质的折叠、成熟及转运至 关重要。细胞稳态改变可以干扰 内质网功能而引起内质网应激 (endoplasmic reticulum stress, ERS)。
内质网应激
• 定义:内质网应激是内质网功能紊乱时, 蛋白质出现错误折叠并与未折叠蛋白在腔 内聚集,以及钙平衡紊乱的状态。 • 信号通路:内质网应激主要包括三条信号 通路:(1)非折叠蛋白反应;(2)内质网超负 荷反应;(3)固醇级联反应。而非折叠蛋白 反应是其中研究较多的信号通路,其最终 目的是通过减少蛋白质合成,促使蛋白质 降解和增加分子伴侣合成帮起细胞凋亡。
相关疾病
• 内质网应激与2型糖尿病 • 2型糖尿病介绍:胰岛素抵抗(insulin resistance, IR)即胰岛素靶组织对胰岛素的敏感性降低,通常 指胰岛素介导的葡萄糖利用率下降,是2型糖尿病 的重要特征之一。在正常情况下,胰岛素能够激 活肌肉、肝脏、脂肪组织中的胰岛素信号通路, 使其摄取葡萄糖并转化为糖原或脂肪进行储存, 从而达到降低血糖的功能。但在2型糖尿病患者中, 胰岛素的这一重要功能受损,外周组织(如肌肉、 肝脏、脂肪组织等) 对胰岛素敏感性下降,从而直 接导致2型糖尿病的发生。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
内质网的结构与功能
一、结构
内质网膜约占细胞总膜面积的一半,就是真核细胞中最多的膜。
内质网(endoplasmic reticulum,ER)就是内膜构成的封闭的网状管道系统。
具有高度的多型性。
可分为粗面型内质网(rough endoplasmic reticulum,RER,图6-20)与光面型内质网(smooth endoplasmic reticulum,SER,图6-21)两类。
二、RER的功能
(一)蛋白质合成
蛋白质都就是在核糖体上合成的,并且起始于细胞质基质,但就是有些蛋白质在合成开
始不久后便转在内质网上合成,这些蛋白质主要有:①向细胞外分泌的蛋白、如抗体、激素;
②跨膜蛋白,并且决定膜蛋白在膜中的排列方式;③需要与其它细胞组合严格分开的酶,如溶
酶体的各种水解酶;④需要进行修饰的蛋白,如糖蛋白。
(二)蛋白质的修饰与加工
包括糖基化、羟基化、酰基化、二硫键形成等,其中最主要的就是糖基化,几乎所有内质网上合成的蛋白质最终被糖基化。
糖基化的作用就是: ①使蛋白质能够抵抗消化酶的作用;
②赋予蛋白质传导信号的功能;③某些蛋白只有在糖基化之后才能正确折叠。
(三)新生肽链的折叠、组装与运输
COP II介导由内质网输出的膜泡运输,这种膜泡由内质网的排出位点(exit sites)以出芽的方式排出,内质网的排出位点没有结合核糖体,随机分布在内质网上。
不同的蛋白质在内质网腔中停留的时间不同,主要取决于蛋白质完成正确折叠与组装的时间,这一过程就是在属
于hsp70家族的ATP酶的作用下完成的,需要消耗能量。
有些无法完成正确折叠的蛋白质被输出内质网,转入溶酶体中降解掉,大约90%的新合成的T细胞受体亚单位与乙酰胆碱受体都被降解掉,而从未到达靶细胞膜。
三、ER的其它功能
合成膜脂:大多数膜只就是完全在内质网中合成的,例外的情况包括:①鞘磷脂就是在内质网上开始合成的,但完成于高尔基体;②某些线粒体与叶绿体独有的膜脂就是驻留在这些
细胞器中的酶催化合成的。
ER合成的膜脂以膜跑运输的方式转运至高尔基体,溶酶体与质膜上,或借磷脂转移蛋白(phospholipid transfer protein,PTP)形成水溶性复合物,转至其她膜上。
解毒作用:SER中的P450酶系属于单加氧酶(monooxygenase),又称为多功能氧化酶(mixed function oxidase)、羟化酶(hydroxylase),因其还原态的吸收峰在450nm处,故名。
主要分布在SER中,但也存在于质膜、线粒体、高尔基体、过氧化物酶体、核膜等细胞器的膜中,具有解毒作用,通常可将脂溶性有毒物质,代谢为水溶性物质,使有毒物质排出体外。
有时也会将致癌物代谢为活性致癌物。
P450种类繁多,但都就是与其她辅助成分组成一个呼吸链来实现其功能,呼吸链中的P450还原酶实际就就是一种黄素蛋白。
P450催化O2分子中的一个原子加到底物分子上使之羟化,另一个氧原子被NADH或NADPH提供的氢还原生成水,在此氧化过程中无高能磷酸化合物生成。
甾体类激素的合成:在生殖腺与肾上腺的内分泌细胞中,SER、线粒体,可能还有高尔基体上的一些酶共同参与甾体类激素的合成。
调节血糖浓度:使葡糖6-磷酸水解为磷酸与葡萄糖,释放糖至血液中。
细胞中的糖元可被酶转化为葡糖1-磷酸,再转变为葡糖6-磷酸,但由于膜对磷酸化的糖就是高度不通透的,葡糖6-磷酸只有在去磷酸化以后才能通过质膜,进入血液。
形成一些特殊结构:如肌细胞中的SER特化成的肌质网可储存钙离子,作为细胞内信号物质。
支撑作用:内质网就是细胞内最丰富的膜,形成了一种网络结构,提供机械支撑作用,并成为细胞质中酶附着的支架。