全面C3、C4、CAM植物的光合和生理生态特性的比较.doc
C3植物、C4植物与CAM植物

C3植物、C4植物与CAM植物标题: C3植物、C4植物与CAM植物摘要: [C3植物、C4植物与CAM植物]在高等植物中,光合碳同化主要有3种类型:C3途径,C4途径和景天酸代谢途径(CAM)。
C3植物中,CO2的固定主要取决于1,5-二磷酸核酮糖羧化酶(RuBPCase)的活化状态,因为该酶是光合碳循环的入口钥匙。
它催化1,5-二磷酸核酮糖(RuBP)羧化,将大气中的CO2同化,产生两分子磷酸甘油酸,可见RuBPCase在C3植物中同化CO2的重要性。
C4植物是从C3植物进化而来的一种高光效种类。
与C3植物相比,它具有在高光强……[关键词:叶绿体活性基因外稃细胞质叶片大豆]……关键词: 叶绿体活性基因外稃细胞质叶片大豆在高等植物中,光合碳同化主要有3种类型:C3途径,C4途径和景天酸代谢途径(CAM)。
C3植物中,CO2的固定主要取决于1,5-二磷酸核酮糖羧化酶(RuBPCase)的活化状态,因为该酶是光合碳循环的入口钥匙。
它催化1,5-二磷酸核酮糖(RuBP)羧化,将大气中的CO2同化,产生两分子磷酸甘油酸,可见RuBPCase在C3植物中同化CO2的重要性。
C4植物是从C3植物进化而来的一种高光效种类。
与C3植物相比,它具有在高光强,高温及低CO2浓度下,保持高光效的能力。
C4植物固定CO2的酶为磷酸烯醇式丙酮酸羧化酶(PEPCase),与C3作物中RuBPCase相比,PEPCase对CO2的亲和力高。
C4植物的细胞分化为叶肉细胞和鞘细胞,而光合酶在两类细胞中的分布不同,如PEPCase在叶肉细胞固定CO2,生成草酰乙酸(OAA),OAA进一步转化为苹果酸(Mal),Mal进入鞘细胞,脱羧,被位于鞘细胞内的RuBPCase羧化,重新进入卡尔文循环。
这种CO2的浓缩机理导致了鞘细胞内的高浓度的CO2,一方面提高RuBPCase的羧化能力,另一方面又大大抑制了RuBPCase的加氧活性,降低了光呼吸,从而使C4植物保持高的光合效率。
C4植物及C3植物的光合作用曲线比较

C4植物与C3植物的光合作用曲线比较1、光合作用强弱变化的指标光合作用强弱变化的指标通常是光合速率和光合生产率光合速率是指单位时间、单位叶面积吸收CO2的量或放出O2的量或有机物的消耗量。
一般测定光合速率的方法都没有把叶片的呼吸作用考虑在内,所以测定的结果实际是光合作用减去呼吸作用的差数,称为表观光合速率或净光合速率。
如果把表观光合速率加上呼吸速率,则得到总(真正)光合速率。
光合生产率又称净同化率,是指植物在较长时间(一昼夜或一周)内,单位叶面积生产的干物质量。
光合生产率比光合速率低,因为已去掉呼吸等消耗。
2、影响光合作用的因素内因1)叶龄:叶片的光合速率与叶龄密切相关。
从叶片发生到衰老凋萎,其光合速率呈单峰曲线变化。
新形成的嫩叶由于组织发育不健全、叶绿体片层结构不发达、光合色素含量少、光合酶含量少、活性弱、气孔开度低、细胞间隙小、呼吸细胞旺盛等原因,净光合速率很低,需要从其它功能叶片输入同化物。
随着叶片的成长,光合速率不断提高。
当叶片伸展至叶面积最大和叶厚度最大时,光合速率达最大值。
通常将叶片充分展开后光合速率维持较高水平的时期,称为叶片功能期,处于功能期的叶叫功能叶。
功能期过后,随着叶片衰老,光合速率下降2)光合产物的运输:光合产物从叶片中输出的快慢影响叶片的光合速率。
例如,摘去花或果实使光合产物的输出受阻,叶片的光合速率就随之降低。
反之,摘除其他叶片,只留一个叶片和所有花果,留下叶片的光合速率就会增加。
如对苹果枝条进行环割,光合产物会积累,则叶片光合速率明显下降。
叶肉细胞中蔗糖的积累会促进叶绿体基质中的淀粉合成和淀粉粒形成,过多的淀粉粒一方面会压迫和损伤叶绿体,另一方面,由于淀粉粒对光有遮挡,从而阻碍光合膜对光的吸收。
(1)光照:光是光合作用的能量来源,是形成叶绿素的必要条件。
此外,光还调节着光合酶的活性和气孔开度,因此光是影响光合作用的重要因素1)光强:在暗中叶片无光合作用,只进行细胞呼吸释放CO2。
C3C4CAM植物光合作用之比较
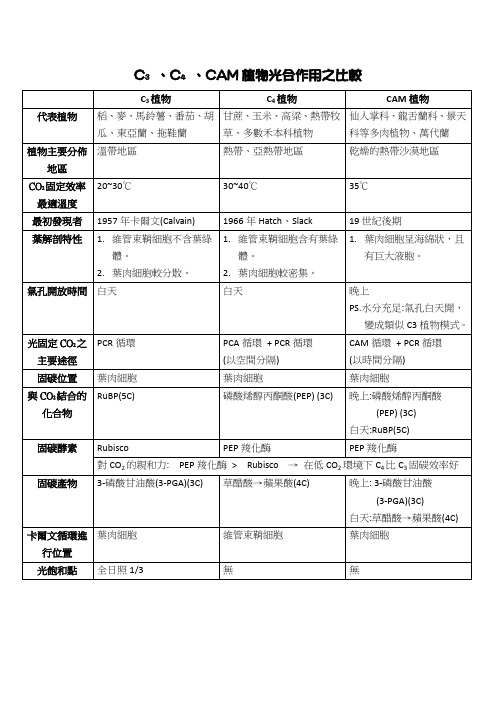
C3 植物 代表植物 瓜、東亞蘭、拖鞋蘭 植物主要分佈 溫帶地區 地區 CO2 固定效率 最適溫度 最初發現者 葉解剖特性 1957 年卡爾文(Calvain) 1. 維管束鞘細胞不含葉綠 體。 2. 葉肉細胞較分散。 氣孔開放時間 白天 1966 年 Hatch、Slack 1. 維管束鞘細胞含有葉綠 體。 2. 葉肉細胞較密集。 白天 晚上 PS.水分充足:氣孔白天開, 變成類似 C3 植物模式。 光固定 CO2 之 主要途徑 固碳位置 與 CO2 結合的 化合物 固碳酵素 PEP 羧化酶 PEP 羧化酶 > Rubisco 草醋酸→蘋果酸(4C) 葉肉細胞 RuBP(5C) PCR 循環 PCA 循環 + PCR 循環 (以空間分隔) 葉肉細胞 磷酸烯醇丙酮酸(PEP) (3C) CAM 循環 + PCR 循環 (以時間分隔) 葉肉細胞 晚上:磷酸烯醇丙酮酸 (PEP) (3C) 白天:RuBP(5C) Rubisco 對 CO2 的親和力: 固碳產物 3-磷酸甘油酸(3-PGA)(3C) PEP 羧化酶 → 在低 CO2 環境下 C4 比 C3 固碳效率好 晚上: 3-磷酸甘油酸 (3-PGA)(3C) 白天:草醋酸→蘋果酸(4C) 卡爾文循環進 葉肉細胞 行位置 光飽和點 全日照 1/3 無 無 維管束鞘細胞 葉肉細胞 19 世紀後期 1. 葉肉細胞呈海綿狀,且 有巨大液胞。 20~30℃ 30~40℃ 35℃ C4 植物 草、多數禾本科植物 熱帶、亞熱帶地區 CAM 植物 科等多肉植物、萬代蘭 乾燥的熱帶沙漠地區 稻、麥、馬鈴薯、番茄、胡 甘蔗、玉米、高粱、熱帶牧 仙人掌科、龍舌蘭科、景天
C3、C4和CAM植物的光合特性比较

C3还原 的场所 叶肉细胞叶 绿体 维管束鞘细 胞叶绿体
暗反应 途径 C3途径 C3途径 C4途径
二、生理特性
在生理上,C4植物一般比C3植物具有较强的
光合作用,这是与C4植物的磷酸烯醇式丙酮 酸羧化酶活性较强,光呼吸很弱有关。
1、羧化酶种类和位置 2、PEP羧化酶对CO2的亲和力强
1、羧化酶种类和位置
C3(PEP) C3 C5
多种酶参加 催化
NADP+ ATP ADP+Pi
C3ADP+Pi 4植物光合途径的比较 和C ATP (丙酮酸)
2、C3植物和C4植物光合作用途径比较
项目 种类
(CH2O)
CO2固定 CO2的受体 后产物 C5 PEP ( C3 ) 2C3 C4
C3植物 C4植物
CO2固定 的场所 叶肉细胞叶 绿体 叶肉细胞叶 绿体
C4植物
PEP 草酰乙 酸 16~18
CAM植物
光下:RuBP ; 暗中:PEP 光下:PGA;暗 中:草酰乙酸 0.2
8、CO2固定的最初产 PGA 物 0.30~0.35 9、PEP羧化酶活性 [μ mol/(mg Chl. min)]
10、光合速率 [CO2mg/(dm2.h)]
12、光饱和点
C4植物光合作用特点:
C4途径(叶肉 细胞中的叶绿 体) CO2 酶 C3(PEP) ADP+Pi C5 C4 C3途径和CO2的还 原(维管束鞘细胞 中的叶绿体)
C4
CO2
2C3 多种酶 参加催化
NADPH NADP+ ATP ADP+Pi
C3 ATP (丙酮酸) (CH2O)
C4植物光合作用特点示意图
C3植物和C4植物比较

C3植物:在光照强度较低时光合作用效率较高 C3植物:在光照强度较高时光合作用效率降低 C4植物:在光照强度较高时物:适应于湿润、肥沃的 土壤
C4植物:适应于干旱、贫瘠的 土壤
C3植物:在湿润、肥沃的土壤 中生长良好
C4植物:在干旱、贫瘠的土壤 中具有更强的生存能力
光反应:吸收 光能产生TP和
NDPH
暗反应:固定 CO2合成有机
物
光呼吸:消耗 光反应产生的 TP和NDPH释放
CO2
光合作用效率: C4植物比C3植物 更高因为C4植物 可以更有效地利 用光能提高光合
作用效率。
光反应阶段:C3植物在光反应阶段产生TP和NDPH而C4植物在光反应阶段产生TP和NDPH但 NDPH的生成量较少。
C3植物与C4植物 的应用前景比较
C3植物:广泛应用于农业生产如小麦、水稻等 C4植物:在热带地区具有较高的利用价值如甘蔗、玉米等 C3植物:在温带地区具有较高的利用价值如小麦、水稻等 C4植物:在热带地区具有较高的利用价值如甘蔗、玉米等
C3植物:在生态修复中C3 植物可以吸收大气中的二 氧化碳降低温室气体排放 减缓全球变暖。在环境保 护中C3植物可以吸收土壤 中的重金属和有毒物质降 低土壤污染。
C4植物:在生态修复中C4 植物可以快速生长提高土 壤肥力促进生态系统的恢 复。在环境保护中C4植物 可以吸收大气中的二氧化 硫等有害气体降低空气污 染。
比较:C3植物和C4植物 在生态修复和环境保护 中都有各自的优势可以 根据不同的环境和需求 选择合适的植物种类。
C3植物:适 合用于生产生 物乙醇、生物 柴油等生物能
C3植物与C4植物的比 较
汇报人:
目录
添加目录标题
C3植物与C4植物概 述
C3植物和C4植物比较

绿体
C3
C4途径 C3途径
3. C4植物比C3植物光合作用强的原因?
• 结构原因:
光合在叶肉细胞
光合在维管束鞘细胞
C3 中进行,淀粉积 C4
中进行。有利于光合
累影响光合。
产物的就近运输,防
止淀粉积累影响光合。
维管束鞘细胞 发育不好,无花环 型,叶绿体无或少
发育良好,花环 型,叶绿体较大
• 生理原因: a. PEPC(磷酸烯醇式丙酮酸羧化酶)对CO2的Km(米氏常数)远小于Rubisco(1,
比较C3植物和C4植物
2013.10.17
光合C同化途径
C3途径
RuBP(1, 5 - 二磷酸核酮糖羧化 酶)
C4途径
CO2 浓度降低驱使 C4 植物扩张。
CAM(景天酸代谢)途径
• C3植物 光合作用时CO2中的C直接转移到C3(3-
磷酸甘油酸)里的植物,叫做C3植物。 例如:小麦、水稻、大麦、大豆、马铃
冷草
翦股颖
高羊茅 早熟禾
结缕草
热草
地毯草
狗牙根
雀稗
5-二磷酸核酮糖羧化酶),所以C4对CO2的亲合力大,低CO2浓度(干旱) 下,光合速率更高。
b. C4植物将CO2泵入维管束鞘细胞,改变了CO2/O2比率,改变了Rubisico的
作用方向,降低了光呼吸。
c. 高光强又可推动电子传递与光合磷酸化,产生更多的同化力,以满足C4植物
PCA循环对ATP的额外需求。
C4植物这种独特的作用,被形象的比 喻成“二氧化碳泵”。
C3途径和C4途径
• CO2固定的途径
C4植物有两条:C4途径和C3途径 C3植物有一条:C3途径 • 上述途径分别发生的场所
C3、C4、CAM植物的某些光合特征和生理特征比较
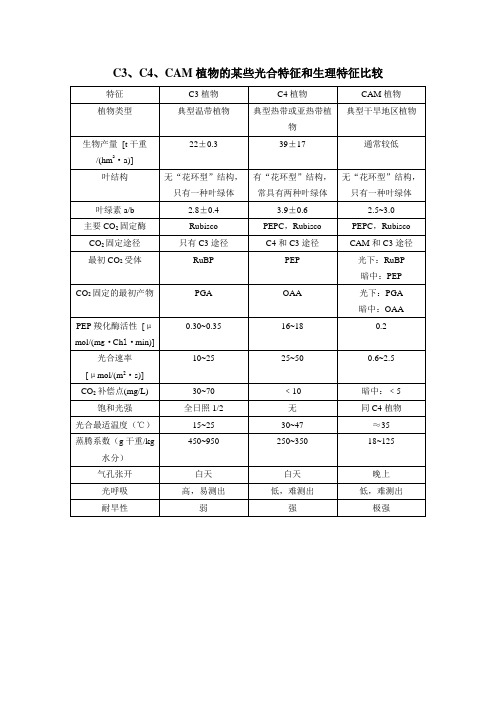
白天
晚上
光呼吸
高,易测出
低,难测出
低,难测出
耐旱性
弱
强
极强
0.30~0.35
16~18
0.2
光合速率
[μmol/(m2·s)]
10~25
25~50
0.6~2.5
CO2补偿点(mg/L)
30~70
﹤10
暗中:﹤5
饱和光强
全日照1/2
无
同C4植物
光合最适温度(℃)
15~25
30~47
≈35
蒸腾系数(g干重/kg水分)
450~950
250~350
18~125
气孔Байду номын сангаас开
叶绿素a/b
2.8±0.4
3.9±0.6
2.5~3.0
主要CO2固定酶
Rubisco
PEPC,Rubisco
PEPC,Rubisco
CO2固定途径
只有C3途径
C4和C3途径
CAM和C3途径
最初CO2受体
RuBP
PEP
光下:RuBP
暗中:PEP
CO2固定的最初产物
PGA
OAA
光下:PGA
暗中:OAA
PEP羧化酶活性[μmol/(mg·Chl·min)]
C3、C4、CAM植物的某些光合特征和生理特征比较
特征
C3植物
C4植物
CAM植物
植物类型
典型温带植物
典型热带或亚热带植物
典型干旱地区植物
生物产量[t干重/(hm2·a)]
22±0.3
39±17
通常较低
叶结构
无“花环型”结构,只有一种叶绿体
C3植物与C4植物光合作用机理的差别与意义

C4植物光合作用特点示意图
C3、C4植物的特性比较
1. C3途径在叶肉细胞和叶绿体内进行. 2. C3途径利用两个同化力.把CO2还原 为有机物,同时把活跃的化学能→稳定 的化学能,贮存在有机物中去. 3. 所有绿色植物 ,对CO2还原都有这条 途径,并非只有这一条途径.把只有这一 条途径的植物叫C 3植物. • 例: 木本植物 蔬菜作物都是C3植物.
卡尔文循环图
Calvin-Benson 循环(光合碳还原循 环)
过程分为 3个阶段
(1)羧化阶段
指进入叶绿体的CO2与受 体RuBP结合,并水解产生PGA的反 应过程(图4-17中的反应1)。 (2)还原阶段 指利用同化力将3-磷酸甘 油酸还原为甘油醛-3-磷酸的反应过程 (图4-17中的反应2、3):
(3)再生阶段 指由甘油醛-3-磷酸重新形 成核酮糖-1,-5-二磷酸的过程(图4-17中 的反应4~14)。
CO 2的固定 即:羧化阶段. CO2+RUBP.(1.5二磷酸核酮糖)→2个 三磷酸甘油质.(3C) • 磷酸甘油质被还原 : 即还原阶段 ATP,NADPH 3-PGA------------→ 3—GAP.(磷酸甘 油醛) 还原 • 1.5二磷酸核酮糖再生阶段 淀粉.蔗糖 酶 ↑ 磷酸甘油醛---------------→ 果糖—磷 酸→葡萄糖—6—磷酸 醛缩酶 ↓ 1. 5二磷酸核酮糖
C3植物与C4植物光合作用机 理的差别与意义
冯亚丽
C3植物和C4植物
1.概念 2.光合作用机理 3.差别与意义
什么是C3植物和C4植物?
• C3植物也叫三碳植物。光合作用中同化二 氧化碳的最初产物是三碳化合物3-磷酸甘 油酸的植物。碳三植物的光呼吸高,二氧 化碳补偿点高,而光合效率低。如小麦、 水稻大豆、棉花等大多数作物。 • C4植物,CO2同化的最初产物不是光合碳 循环中的三碳化合物3-磷酸甘油酸,而是四 碳化合物苹果酸或天门冬氨酸的植物。如 玉米、甘蔗、高粱、苋菜等。
C3植物、C4植物和CAM植物的比较

1、C3植物、C4植物和CAM植物 的定义和基本特点
C3植物是指通过卡尔文循环进行光合作用的植物,它们在二氧化碳固定时使 用酶 Rubisco。C4植物是指通过C4途径进行光合作用的植物,它们在二氧化碳固 定时使用酶 PEPC。而CAM植物则是指通过CAM途径进行光合作用的植物,它们在 二氧化碳固定时使用酶 PEP羧化酶。
C4植物的进化
C4植物是指通过C4途径进行二氧化碳固定的植物。这类植物的光合作用效率 较C3植物更高,且受环境因素影响更小。其进化历程大致如下:
C4植物可能最初出现在约6千万年前,当时地球大气中的二氧化碳浓度开始 逐渐降低,导致全球气候逐渐变得干旱。为了适应这种环境,一部分C3植物开始 尝试进行C4途径固定二氧化碳。成功适应这一转变的植物逐渐繁衍开来,形成了 现在约20%的全球植被。典型的C4植物包括玉米、甘蔗等。
分利用效率适应性最强。此外这三种类型的植物在基因组方面也存在差异具 有各自的独特性和进化发展历程。
未来发展方向:针对三种不同类型的植物的特点和优劣我们可以得出以下发 展方向:首先,针对C3植物的光合作用过程可以通过基因工程技术手段来提高它 们的二氧化碳固定能力和光能利用率从而提高其产量;其次针对C4植物在恶劣环 境下的适应能力可以通过深入探究它们的生理机制和基因表达模式来提高它们的 环境适应性并培育出耐旱、耐盐等新品种;最后针对CAM植物的
C3植物的进化
C3植物是指通过卡尔文循环进行二氧化碳固定的植物。这类植物的光合作用 效率较高,但受环境因素影响较大。其进化历程大致如下:
最初的C3植物可能出现在约2.5亿年前,当时地球大气中的二氧化碳浓度较 高,导致全球气候温暖湿润,适合这种植物生长。随着时间的推移,地球气候逐 渐变得干燥,C3植物为了适应这种环境,开始在自身生理代谢上做出调整,例如 增加气孔开度、加强水分吸收等。这些调整使得C3植物能够在干旱条件下生存并 逐渐繁衍开来。现在,C3植物已经成为了地球上最为普遍的一类植物,包括了如 水稻、小麦、大豆等重要的农作物。
C3植物和C4植物的光合特征

C3植物和C4植物的光合特征人们根据光合作用碳素同化的最初光合产物的不同,把高等植物分成两类:(1)C3植物。
这类植物的最初产物是3-磷酸甘油酸(三碳化合物),这种反应途径称C3途径,如水稻、小麦、棉花、大豆等大多数植物。
(2)C4植物。
这类植物以草酰乙酸(四碳化合物)为最初产物,所以称这种途径为C4途径,如甘蔗、玉米、高粱等。
一般来说,C4植物比C3植物具有较强的光合作用,其原因可从结构和生理两方面来探讨。
结构与功能是有密切关系的,是统一的。
C4植物叶片的维管束薄壁细胞较大,其中含有许多较大的叶绿体,叶绿体没有基粒或基粒发育不良;维管束鞘的外侧密接一层成环状或近于环状排列的叶肉细胞,组成了“花环型”(Kranz type)结构。
这种结构是C4植物的特征。
叶肉细胞内的叶绿体数目少,个体小,有基粒(图3-28)。
维管束鞘薄壁细胞与其邻近的叶肉细胞之间有大量的胞间连丝相连。
C3植物的维管束鞘薄壁细胞较小,不含或很少叶绿体,没有“花环型”结构,维管束鞘周围的叶肉细胞排列松散(图3-29)。
前面说过,C4植物通过磷酸烯醇式丙酮酸固定二氧化碳的反应是在叶肉细胞的细胞质中进行的,生成的四碳双羧酸转移到维管束鞘薄壁细胞中,放出二氧化碳,参与卡尔文循环,形成糖类,所以甘蔗、玉米等C4植物进行光合作用时,只有维管束鞘薄壁细胞形成淀粉,在叶肉细胞中没有淀粉。
而水稻等C3植物由于仅有叶肉细胞含有叶绿体,整个光合过程都是在叶肉细胞里进行,淀粉亦只是积累在叶肉细胞中,维管束鞘薄壁细胞不积存淀粉。
在生理上,C4植物一般比C3植物具有较强的光合作用,这是与C4植物的磷酸烯醇式丙酮酸羧化酶活性较强,光呼吸很弱有关。
前面已经提过,卡尔文循环的CO2固定是通过核酮糖二磷酸羧化酶的作用来实现的,C4途径的CO2固定是由磷酸烯醇式丙酮酸羧化酶催化来完成的。
两种酶都可使CO2固定。
但它们对CO2的亲和力却差异很大。
磷酸烯醇式丙酮酸羧化酶对CO2的Km值(米氏常数)是7μmol,核酮糖二磷酸羧化酶的Km值是450μmol。
c4植物比c3植物光合速率高的原因

C4植物相对于C3植物具有更高的光合速率,主要原因有以下几点:
1. C4植物具有更高的光合效率:C4植物的光合作用通过C4途径和C3途径两个步骤进行。
在C4途径中,C4植物通过将CO2固定为C4有机酸(如草酸)来增加CO2的浓度,并将其传输到叶绿体内。
这一过程可以克服光合作用初级产物CO2在叶绿体中的低浓度限制,并增加CO2利用的效率。
2. C4植物具有更高的光合作用整体效率:C4植物将光合作用的关键反应分离在不同的细胞类型中。
在外部细胞(称为束鞘细胞)中,C4植物在高CO2浓度下进行光合作用的第一步,形成C4有机酸。
然后,C4有机酸在内部细胞(称为甲酸细胞)中解离为CO2,并在这些细胞中进行C3光合作用。
这种分离可以减少C4植物在高温和干旱条件下的光合作用损失,并提高整体光合作用的效率。
3. C4植物具有更高的水分利用效率:C4植物的光合速率高,同时水分蒸腾速率相对较低。
这是因为C4植物在CO2获取过程中,CO2浓度较高,不需要开放气孔来获取足够的CO2。
相比之下,C3植物需要开放气孔来吸收CO2,但这会导致较高的水分蒸腾速率。
因此,C4植物相对C3植物具有更高的水分利用效率。
总的来说,C4植物比C3植物具有更高的光合速率主要是因为它们通过C4途径增加了CO2浓度,分离了光合作用的关键反应,并且具有更高的水分利用效率。
这些特性使得C4植物在热带和亚热带地区具有更好的适应性和竞争优势。
光合作用—C3、C4、CAM区别

光合作用光合作用可分为:光反应和碳反应(旧称暗反应)两个阶段。
光反应条件:光照、光合色素、光反应酶。
场所:叶绿体的类囊体薄膜。
(蓝细菌等微生物的反应场所在细胞膜:色素所在地)过程:①水的光解:2H2O→4[H]+O2(在光和叶绿体中的色素的催化下)。
②ATP的合成:ADP+Pi+能量→ATP(在酶的催化下)。
影响因素:光照强度、CO2浓度、水分供给、温度、酸碱度、矿质元素等。
意义:①光解水,产生氧气。
②将光能转变成化学能,产生ATP,为碳反应提供能量。
③利用水光解的产物氢离子,合成NADPH(还原型辅酶Ⅱ),为碳反应提供还原剂NADPH(还原型辅酶Ⅱ)。
碳反应条件:多种酶。
场所:叶绿体基质。
过程:①碳的固定:C5+CO2→2C3(在酶的催化下)②C3+[H]→(CH2O)+C5(在ATP供能和酶的催化下)影响因素:温度、CO2浓度㊣卡尔文循环(Calvin Cycle)可分为三个阶段: 羧化(碳的固定)、还原(3-磷酸甘油醛(G3P(PGAL))的合成)、二磷酸核酮糖的再生。
卡尔文循环,又称还原磷酸戊糖循环(以对应呼吸作用中的氧化磷酸戊糖途径)、C3循环(CO2固定的第一产物是三碳化合物)、光合碳还原,还是光合作用的暗反应的一部分。
反应场所为叶绿体内的基质。
㊣固碳作用:※碳三植物(小麦、水稻、大豆、棉花等)二氧化碳经气孔进入叶片后,直接进入叶肉进行卡尔文循环。
而C3植物的维管束鞘细胞很小,不含或含很少叶绿体,卡尔文循环不在这里发生。
※碳四植物(玉米、甘蔗、高粱、苋菜等)碳四植物维管束周围有两种不同类型的细胞:靠近维管束的内层细胞称为鞘细胞,围绕着鞘细胞的外层细胞是叶肉细胞。
①叶肉细胞里的磷酸烯醇式丙酮酸(PEP)经PEP羧化酶的作用,与CO2结合,形成苹果酸或天门冬氨酸。
②这些四碳双羧酸转移到鞘细胞里,通过脱羧酶的作用释放CO2,后者在鞘细胞叶绿体内经核酮糖二磷酸(RuBP)羧化酶作用,进入光合碳循环(卡尔文循环。
C4植物与C3植物的光合作用曲线比较【范本模板】

C4植物与C3植物的光合作用曲线比较1、光合作用强弱变化的指标光合作用强弱变化的指标通常是光合速率和光合生产率光合速率是指单位时间、单位叶面积吸收CO2的量或放出O2的量或有机物的消耗量。
一般测定光合速率的方法都没有把叶片的呼吸作用考虑在内,所以测定的结果实际是光合作用减去呼吸作用的差数,称为表观光合速率或净光合速率.如果把表观光合速率加上呼吸速率,则得到总(真正)光合速率。
光合生产率又称净同化率,是指植物在较长时间(一昼夜或一周)内,单位叶面积生产的干物质量。
光合生产率比光合速率低,因为已去掉呼吸等消耗.2、影响光合作用的因素内因1)叶龄:叶片的光合速率与叶龄密切相关。
从叶片发生到衰老凋萎,其光合速率呈单峰曲线变化。
新形成的嫩叶由于组织发育不健全、叶绿体片层结构不发达、光合色素含量少、光合酶含量少、活性弱、气孔开度低、细胞间隙小、呼吸细胞旺盛等原因,净光合速率很低,需要从其它功能叶片输入同化物。
随着叶片的成长,光合速率不断提高。
当叶片伸展至叶面积最大和叶厚度最大时,光合速率达最大值。
通常将叶片充分展开后光合速率维持较高水平的时期,称为叶片功能期,处于功能期的叶叫功能叶。
功能期过后,随着叶片衰老,光合速率下降2)光合产物的运输:光合产物从叶片中输出的快慢影响叶片的光合速率。
例如,摘去花或果实使光合产物的输出受阻,叶片的光合速率就随之降低。
反之,摘除其他叶片,只留一个叶片和所有花果,留下叶片的光合速率就会增加.如对苹果枝条进行环割,光合产物会积累,则叶片光合速率明显下降。
叶肉细胞中蔗糖的积累会促进叶绿体基质中的淀粉合成和淀粉粒形成,过多的淀粉粒一方面会压迫和损伤叶绿体,另一方面,由于淀粉粒对光有遮挡,从而阻碍光合膜对光的吸收。
(1)光照:光是光合作用的能量来源,是形成叶绿素的必要条件。
此外,光还调节着光合酶的活性和气孔开度,因此光是影响光合作用的重要因素1)光强:在暗中叶片无光合作用,只进行细胞呼吸释放CO2.随着光强的增高,光合速率相应提高,当达到某一光强时,叶片的光合速率与呼吸速率相等,净光合速率为零,这时的光强称为光补偿点在一定范围内,光合速率随着光强的增加而呈直线增加;但超过一定光强后,光合速率增加转慢;当达到某一光强时,光合速率就不再随光强增加而增加,这种现象称为光饱和现象。
C4植物与C3植物的比较 甄选

C4植物与C3植物的比较 (优选.)rd植物与植物的比较人们根据光合作用碳素同化的最初光合产物的不同,把高等植物分成两类:(1)C 3植物。
这类植物的最初产物是3-磷酸甘油酸(三碳化合物),这种反应途径称C 3途径,如水稻、小麦、棉花、大豆等大多数植物。
(2)C 4植物。
这类植物以草酰乙酸(四碳化合物)为最初产物,所以称这种途径为C 4途径,如甘蔗、玉米、高粱等。
一般来说,C 4植物比C 3植物具有较强的光合作用,原因有:一、叶片的显微结构——重点比较维管束鞘细胞结构 C 4植物叶片的维管束薄壁细胞较大,其中含有许多较大的叶绿体,叶绿体没有基粒或基粒发育不良;维管束鞘的外侧密接一层成环状或近于环状排列的叶肉细胞,组成了“花环型”结构。
这种结构是C 4植物叶片所特有的特征。
叶肉细胞内的叶绿体数目少,个体小,有基粒。
C 3植物的维管束鞘薄壁细胞较小,不含或很少叶绿体,没有“花环型”结构,维管束鞘周围的叶肉细胞排列松散。
含有“花环状”环绕在维管束鞘细胞外含有大含有排列疏松不含小维管束C 4 植物C 3 植物叶绿体排列叶绿体细胞大小叶肉细胞维管束鞘细胞比较类型维管束C 4 植物的叶片结构海绵组织维管束鞘细胞栅栏组织维管束鞘细胞一部分叶肉细胞植物类型 植物 植物叶片的解剖结构 无“花环型”结构 维管束鞘细胞及周围的一部分叶肉细胞构成“花环型”结构叶绿体的类型 有一种类型的叶绿体,主要位于叶肉细有两种类型的叶绿体,叶肉细胞的叶绿体正常,维管束鞘细胞中 胞的叶绿体没有基粒二、淀粉粒形成的场所 C 4植物通过磷酸烯醇式丙酮酸固定二氧化碳的反应是在叶肉细胞中进行的,生成的四碳双羧酸转移到维管束鞘薄壁细胞中,放出二氧化碳,参与卡尔文循环,形成糖类,所以甘蔗、玉米等C 4植物进行光合作用时,只有维管束鞘薄壁细胞形成淀粉,在叶肉细胞中没有淀粉。
而水稻等C 3植物由于仅有叶肉细胞含有叶绿体,整个光合过程都是在叶肉细胞里进行,淀粉亦只是积累在叶肉细胞中,维管束鞘薄壁细胞不积存淀粉。
C_3植物、C_4植物和CAM植物的比较

C_3植物、C_4植物和CAM植物的比较
罗红艺
【期刊名称】《高等函授学报:自然科学版》
【年(卷),期】2001(14)5
【摘要】根据光合作用中碳同化途径的不同 ,把植物分为C3 植物、C4 植物和CAM植物。
【总页数】4页(P35-38)
【关键词】C3植物;C4植物;CAM植物;光合特征;光合作用;碳同化;光合形态【作者】罗红艺
【作者单位】华中师范大学生命科学学院
【正文语种】中文
【中图分类】Q945.11
【相关文献】
1.C_3植物与C_4植物的比较 [J], 张晓丽;魏俊杰
2.C_4,C_3,CAM植物叶磷酸烯醇式丙酮酸羧化酶分子聚体的比较 [J], 焦德茂
3.C_3植物和C_4植物的区别 [J], 何世顺
4.C_3C_4C_3—C_4中间植物的光合特征及C_3—C_4中间植物在进化上的地位[J], 唐萍
5.浅论C_3植物和C_4植物 [J], 杨胜利
因版权原因,仅展示原文概要,查看原文内容请购买。