第六章(1) 分子生物学改造
第六章:常用分子生物学技术——RNA干扰技术

识别并切割(qiēgē)与siRNA反义链互补的靶mRNA
(5)dsRNA的再生成
第十ānrǎo)
第十七页,共四十页。
RNA干扰(gānrǎo)
(RNA interference, RNAi)
siRNA(小干扰RNA):21-23nt,由dsRNA裂 解而成的小片段,可诱导mRNA降解。siRNA主要
药学 分子生物 (yào xué)
学
第一页,共四十页。
生物芯片 (biochip) (shēnɡ wù xīn piàn)
以生物、电子、机械和信息技术为基础,在 固相支持物表面建立集成、连续、微型分析系 统(xìtǒng),形成芯片,实现对生物大分子的准确、 快速、高通量、自动化检测。
第二页,共四十页。
第三十一页,共四十页。
REGγ基因敲除鼠(下图)出现脊椎(jǐ zhuī)弯曲等早衰现象
第三十二页,共四十页。
Gab1基因(jīyīn)敲除导致小鼠缺血性血管新生、侧枝循环建立出现 缺陷(图示上半部分),主要是由于血管内皮细胞管状结构形成的 信号调控通路出现障碍而引起的(图示下半部分)。
第三十三页,共四十页。
2、 RNA干扰技术缺点
“脱靶效应”(off-target effects)
第二十四页,共四十页。
RNA干扰技术(jìshù)的应用
1、基因功能研究(功能失活策略) “基因敲减(knockdown)”
2、基因治疗(基因(jīyīn)失活性治疗) (1)病毒感染性疾病
通过RNAi抑制 RNA病毒(bìngdú)复制 (2)基因过表达引起的疾病(如肿瘤)
特定功能基因的部分片段被同源的外源DNA片 段替代,从而使靶基因失活。
2024年分子生物学电子教案

2024/2/29
14
04
基因的表达与调控
Chapter
2024/2/29
15
基因表达的转录过程
1 2
转录起始
RNA聚合酶识别并结合启动子,形成转录起始复 合物。
转录延伸
RNA聚合酶沿DNA模板链移动,合成RNA链。
3
转录终止
RNA聚合酶遇到终止子,释放RNA链并停止转 录。
2024/2/29
16
2024/2/29
识别并切除受损碱基,然后通过 DNA聚合酶和连接酶进行填补和 连接。
在DNA复制发生错误时,通过姐 妹染色单体之间的同源重组进行 修复。
25
DNA复制与修复的生物学意义
遗传信息的传递
DNA复制确保了遗传信息在细胞分裂过程中的准 确传递,是生物体生长、发育和繁殖的基础。
适应环境变化
tRNA
转运RNA,负责携带氨基酸并识别 mRNA上的密码子。结构特点包括 三叶草形二级结构和倒L形三级结 构。
rRNA
核糖体RNA,是核糖体的组成成分 ,参与蛋白质合成。结构特点包括 多个大小亚基和复杂的二级、三级 结构。
13
RNA的生物学功能
遗传信息传递
RNA作为遗传信息的携带 者,在DNA的指导下合成 ,并将遗传信息传递给蛋 白质合成系统。
2024/2/29
分子生物学的发展
自20世纪50年代以来,随着DNA双螺旋结构的发 现、遗传密码的破译、基因工程技术的建立等一系 列重大科学成就的取得,分子生物学迅速崛起并渗 透到生物学的各个领域,推动了整个生命科学的飞 速发展。
4
分子生物学的研究内容
生物大分子的结构与功能
研究生物大分子如蛋白质、核酸等的 结构特点、理化性质和功能,以及结 构与功能之间的关系。
分子生物学-课件-分子生物第六章可编辑全文

• 使双链DNA解链度达到50%所需的温度称为解链温度 (Tm)、变性温 度、熔点
• DNA的解链温度一般在82-95℃ ,与DNA的分子大小和碱基组成、 溶液的pH值和离子强度(+)等有关
二、复性
• DNA复性:缓慢降温可以使热变性DNA重新 形成互补双链结构
且RNA的完整性和纯度都很高
3、氯化锂-尿素法
• 利用高浓度尿素变性蛋白质同时抑制RNA 酶,氯化锂选择性沉淀RNA
• 缺点:存在DNA污染,氯化锂沉淀RNA会 丢失一些小分子RNA,如5sRNA等
• 优点:快速、简便、产量高,尤其适用于大 量样品少量组织细胞的RNA提取
4·热酚法
• 将异硫氰酸胍、巯基乙醇和SDS等联合使用,可以快速裂 解细胞,解离核蛋白复合物,释放RNA,并有效抑制 RNase的活性
• 在某些理化因素的作用下,维 系核酸二级结构的氢键和碱基 堆积力受到破坏,DNA双螺旋 结构松散,变成单链的过程称 为核酸的变性
• 核酸双螺旋区的氢键断裂,变 成单链,但并不涉及共价键的 断裂
DNA解链曲线
DNA变性的本质是氢键的断裂
核酸的变性因素
• 变性方法
–热变性、酸碱变性、化学变性剂(乙醇、 尿素和甲酰胺 )
(3)煮沸裂解法
• 以溶菌酶、Triton裂解细菌,然后以沸水浴加热,不 仅促进细菌裂解,还可以使蛋白质和染色体DNA、质 粒DNA变性
• 当降低温度时,闭环质粒DNA复性留在上清液中,而 染色体DNA保持与细胞膜碎片结合、沉淀,可以通过 离心除去
• 在离心上清液中加入有机溶剂 (例如异丙醇)便得到质 粒DNA粗品沉淀
第六章1分子生物学改造

一、定点突变
利用分子生物学技术,在体外通过碱基取代、插入 或缺失可以使基因DNA序列中任何一个特定的碱基 发生改变。这种体外特异性改变某个碱基的技术, 称谓定点突变(site directed mutagenesis)。
定点突变具有简单易行、重复性高等优点,现已发 展成为基因操作的一种技术。这种技术不仅适用于 基因结构与功能的研究,还可通过改变基因的密码 子来改造天然蛋白质。
(%) 100 100
96 106
0 95 0
0
(℃) 41.9 41.9 46.7 48.3 52.9 57.6 58.9 65.9
wtα:野生型T4溶菌酶;pwt:假野生型酶;A-F:六种设计的半胱氨酸 变体;Tm:熔点温度
将Asn和Gln转换成其他氨基酸
当蛋白质暴露于高温时:
天冬酰胺(Asn)
而将2个Asn分别换为苏氨酸(Thr)和异亮氨酸(Ile),其半衰 期则延长。
酵母菌磷酸丙糖异构酶及其变体的热稳定性
氨基酸位点
酶
14
78
半衰期(min)
野生型 Asn
Asn
13
变体A Asn
Thr
17
变体B Asn
Ile
16
变体C Thr
Ile
25
变体D Asp
Asn
天冬氨酸(Asp) + NH3
谷氨酰胺(Gln)
谷氨酸(Glu) + NH3
导致肽链折叠的局部的改变,可能影响其活性。
如酵母的丙糖磷酸异构酶是由两个相同的亚基组成的二聚体, 每个亚基都含有两个Asn残基,均位于两个亚基相互接触的 表面上,可能与该酶的热稳定性有关。
分子生物学第5章、第6章

•DNA分子内或分子间发生遗传信息的重新组合,称为 遗传重组,或基因重排。→ 重组DNA •真核生物基因组间重组多发生在减数分裂时同源染 色体之间的交换;细菌及噬菌体的基因组为单倍体, 来自不同亲代两组DNA之间可通过多种形式进行遗传 重组。 •DNA重组对生物进化起着关键的作用。 •重组分类:同源重组(homologous recombination) 、 位点特异性重组(site-specific recombination)、 转座重组(transposition recombination)和 异常重组(illegitimate recombination)。
1. 互变异构体:碱基发生烯醇式-酮式互变异构或者氨 基-亚氨基互变异构时,使碱基错配。 2. 脱氨基作用:碱基上氨基自发脱落,或在诱变剂的 作用下脱去氨基,则C→U、A →I、G →X,引起子 链错误。 3. DNA聚合酶“打滑”:DNA复制时发生碱基的环出现 象,引起一个或数个碱基的插入或缺失,易发生于 几个相同碱基串联的部位。 4. 活性氧(O3)引起的诱变:①氧化碱基与C、A配对, 造成GC → TA颠换,这种损伤可以积累;②H2O2造成 的DNA氧化损伤,此类损伤一般能被修复。
核苷酸切除修复
错配修复
错配修复对 DNA复制忠实 性的贡献力达 102-103,DNA 子链中的错配 几乎完全都被 修正,充分反 映了母链的重 要性。
大肠杆菌甲基化引 导的错配修复
重组修复
易错修复和SOS反应
•SOS反应:当DNA损伤广泛难以继续复制时,由此而
诱发出一系列复杂的反应。
•这种修复特异性低,对碱基的识别、选择能力差。
5.3.4 基因突变的后果
基因突变的后果主要是生物功能的丧失。 某一基因突变后使其所表达的蛋白质或酶失活, 有时还会引起多种酶的缺乏。 有些突变可产生功能获得性显性表现型。 典型的人体细胞突变每个基因每代发生率为107~10-5,但并非所有的突变都会导致疾病。
分子生物学习题集答案 第六章

第六章基因表达调控自测题(一)选择题A型题1. 关于基因表达调控的说法错误..的是A. 转录起始是调控基因表达的关键B. 环境因素影响管家基因的表达C. 在发育分化和适应环境上有重要意义D. 表现为基因表达的时间特异性和空间特异性E. 真核生物的基因表达调控较原核生物复杂的多2. 下列哪项属于可调节基因A. 组蛋白编码基因B. 5S rRNA编码基因C. 异柠檬酸脱氢酶编码基因D. 肌动蛋白编码基因E. 血红蛋白编码基因3. 与α-酮戊二酸脱氢酶系协调表达的是A. 肉毒碱脂酰转移酶IB. 柠檬酸合成酶C. 丙酮酸羧化酶D. 葡萄糖-6-磷酸酶E. HMG-CoA合成酶4. 乳糖操纵子中,能结合别位乳糖(诱导剂)的物质是A. AraCB. cAMPC. 阻遏蛋白D. 转录因子E. CAP5. 乳糖操纵子模型是在哪个环节上调节基因表达A. 复制水平B. 转录水平C. 转录后水平D. 翻译水平E. 翻译后水平6. 乳糖操纵子的调控方式是A. CAP的正调控B. 阻遏蛋白的负调控C. 正、负调控机制不可能同时发挥作用D. CAP拮抗阻遏蛋白的转录封闭作用E. 阻遏作用解除时,仍需CAP加强转录活性7. 与分解代谢相关的操纵子模型中,存在分解代谢物阻遏现象,参与这一调控的主要作用因子是A. 阻遏蛋白B. AraCC. 衰减子D. cAMP-CAP复合物E. 诱导剂8. 原核细胞中,识别基因转录起始点的是A. 阻遏蛋白B. 转录激活蛋白C. 基础转录因子D. 特异转录因子E. σ因子9. 使乳糖操纵子实现高表达的条件是A. 乳糖存在,葡萄糖缺乏B. 乳糖缺乏,葡萄糖存在C. 乳糖和葡萄糖均存在D. 乳糖存在E. 葡萄糖存在10. 大肠杆菌可以采用哪种方式调控转录终止A. 阻遏作用B. 去阻遏作用C. 反义控制D. 衰减作用E. 降低转录产物的稳定性11. 关于色氨酸操纵子错误..的描述是A. 核蛋白体参与转录终止B. 衰减子是关键的调控元件C. 色氨酸不足时,转录提前终止D. 转录与翻译偶联是其转录调控的分子基础E. 色氨酸存在与否不影响先导mRNA的转录12. 下列哪种因素对原核生物的翻译没有..影响A. microRNAB. 稀有密码子所占的比例C. mRNA的稳定性D. 反义RNAE. 调节蛋白结合mRNA13. 真核生物基因表达调控的关键环节是①染色质活化②转录起始③转录后加工④翻译起始⑤翻译后加工A. ①+②+③B. ①+②+④C. ①+②D. ②+③E. ②14. 下列哪种染色质结构的变化不.利于基因表达A. 组蛋白乙酰化B. 核小体解聚C. CpG岛甲基化D. 基因扩增E. 染色质结构松散,对DNA酶I敏感15. 下列哪项不.属于真核生物基因的顺式作用元件A. 激素反应元件B. 衰减子C. 启动子D. 沉默子E. 增强子16. 与RNA聚合酶相识别和结合的DNA片段是A. 增强子B. 衰减子C. 沉默子D. 操纵子E. 启动子17. 下列哪项不.参与调控真核细胞基因的特异性表达A. 反应元件B. 特异转录因子C. 增强子D. 基础转录因子E. 沉默子18. 与原核生物相比较,真核生物的基因表达调控包括多个层次,下列哪项不.是其调控复杂性特有的分子基础A. 含有重复序列B. 断裂基因C. 转录与翻译分离D. 细胞内被膜性结构分隔形成多个区域E. 染色质结构19. 能够与基础转录因子结合的是A.上游启动子元件B.TATA boxC.增强子D.反应元件E. Pribnow box20. 有关基础转录因子的叙述,正确的是A. 与非转录核心序列相结合B. 决定基因表达的特异性C. 其种类和数量在不同组织中差别很大D. 辅助RNA聚合酶结合启动子E. 在原核生物中的种类比真核生物少21. 不.属于特异转录因子的是A. TF II DB. HSFC. AP1D. 类固醇激素受体E. NF-κB22. 关于特异转录因子的说法,正确的是A. 调控管家基因的表达B. 仅通过蛋白质-蛋白质相互作用进行调控C. 仅通过DNA-蛋白质相互作用进行调控D. 仅通过RNA-蛋白质相互作用进行调控E. 起转录激活或者转录抑制作用23. 锌指结构可能存在于下列哪种物质中A. 阻遏蛋白B. RNA聚合酶C. 转录因子D. 端粒酶E. 核酶24. 下列哪种氨基酸在转录因子的转录激活结构域中含量丰富A. LysB. ArgC. AspD. HisE. Trp25. 下列哪种因素不.会影响真核细胞中mRNA的稳定性A. 5' 端帽子B. siRNAC. poly A尾D. 去稳定元件E. miRNA26. 小干扰RNA调节基因表达的机制是A. 封闭mRNA上的核蛋白体结合位点B. 特异性降解靶mRNAC. 形成局部双链,抑制靶mRNA的模板活性D. 使翻译出的蛋白质进入泛素化降解途径E. 使翻译提早终止27. eIF-2对翻译起始具有重要的调控作用,下列哪项是它的活性形式A. 磷酸化B. 脱乙酰化C. 乙酰化D. 脱磷酸化E. ADP-核糖基化28. 不.影响真核生物翻译起始的因素是A. eIFB. 帽子结合蛋白C. RNA编辑D. mRNA非翻译区的二级结构E. miRNA29. 原核生物中,某种代谢途径相关的几种酶类往往通过何种机制进行协调表达A. 顺反子B. 操纵子C. 转录因子D. 衰减子E. RNAi30. 生物体在不同发育阶段,蛋白质的表达谱也相应变化,这主要取决于A. 转录调控元件的差异B. 翻译调控元件的差异C. 基础转录因子的差异D. 特异转录因子的差异E. 翻译起始因子的差异31. 原癌基因通常是A. 管家基因B. 可调节基因C. 突变的基因D. 表达增强的基因E. 促进凋亡的基因32. 管家基因编码的产物不.包括A. 细胞外生长因子B. 电子传递链的成员C. 细胞的主要结构蛋白D. 转录因子E. 血红蛋白33. 下列哪项不.是可调节基因的特点A. 组织特异性B. 阶段特异性C. 时间特异性D. 空间特异性E. 组成性表达34. 关于操纵子的说法,正确的是A. 几个串联的结构基因由一个启动子控制B. 几个串联的结构基因分别由不同的启动子控制C. 一个结构基因由不同的启动子控制D. 转录生成单顺反子RNAE. 以正性调控为主35. 核蛋白体调控转录终止的典型例子是A. 乳糖操纵子B. 半乳糖操纵子C. 阿拉伯糖操纵子D. 色氨酸操纵子E. 以上都不对36. 色氨酸操纵子的显著特点是A. 阻遏作用B. 诱导作用C. 衰减作用D. 分解物阻遏作用E. 抗终止作用37. 关于色氨酸操纵子的调控,正确的说法是A. 色氨酸存在时,仅生成前导mRNAB. 色氨酸不足时,转录提前终止C. 是翻译水平的调控D. 具有抗终止作用E. 依赖ρ因子进行转录调控38. 细菌优先利用葡萄糖作为碳源,葡萄糖耗尽后才会诱导产生代谢其他糖的酶类,这种现象称为A. 衰减作用B. 阻遏作用C. 诱导作用D. 协调调节作用E. 分解物阻遏作用39. 关于分解物阻遏的作用机制,说法正确的是A. 葡萄糖缺乏时,cAMP浓度低B. 葡萄糖缺乏时,CAP浓度低C. 葡萄糖缺乏时,cAMP不能与CAP形成复合物D. 葡萄糖缺乏时,cAMP-CAP复合物浓度高E. 葡萄糖缺乏时,cAMP-CAP复合物失去DNA结合能力40. 大肠杆菌的乳糖操纵子模型中,与操纵基因结合而调控转录的是A. 阻遏蛋白B. RNA聚合酶C. 调节基因D. cAMP-CAPE. 启动子41. 翻译终止阶段,新生多肽链的释放涉及哪种化学键的断裂A. 肽键B. 磷酸二酯键C. 氢键D. 疏水键E. 酯键42. IPTG诱导乳糖操纵子表达的机制是A. 使乳糖-阻遏蛋白复合物解离B. 与阻遏蛋白结合,使之丧失DNA结合能力C. 与乳糖竞争结合阻遏蛋白D. 与RNA聚合酶结合,使之通过操纵序列E. 变构修饰RNA聚合酶,提高其活性43. 下列哪一项是真核生物可调节基因的表达调控特有的机制A. 基础转录因子B. 衰减子C. RNA聚合酶D. 增强子E. 阻遏蛋白44. 基础转录因子属于DNA结合蛋白,它们能够A. 结合转录核心元件B. 结合增强子C. 结合5' 端非翻译区D. 结合3' 端非翻译区E. 结合内含子45. 特异转录因子不.能够A. 结合RNA聚合酶B. 结合基础转录因子C. 结合其他特异转录因子D. 结合转录非核心元件E. 结合沉默子46. 基因特异性表达的根本机制是A. 顺式作用元件的种类不同B. RNA聚合酶活性的差异C. 基础转录因子的质和量的差异D. 特异转录因子的质和量的差异E. 表达产物后加工过程的差异47. 下列哪一类分子常具有亮氨酸拉链的结构特征A. 生长因子B. 酪氨酸蛋白激酶受体C. G蛋白D. 转录因子E. 丝/苏氨酸蛋白激酶B型题A. 阻遏蛋白B. AraCC. 衰减子D. CAPE. ρ因子48. 葡萄糖缺乏时,细菌中cAMP浓度升高,可以结合49. 实验室常使用IPTG作为诱导剂,其作用是结合50. 阿拉伯糖操纵子的主要调节因子是51. 色氨酸操纵子的调节作用依赖于A. 转录起始B. 转录终止C. 翻译起始D. 翻译终止E. mRNA稳定性52. 原核基因表达调控的最基本环节是53. 真核基因表达调控的最基本环节是A. eIF-2B. 特异转录因子C. EF-2D. 基础转录因子E. eRF54. 真核细胞中管家基因的转录需要55. 真核细胞中可调节基因的转录主要取决于56. 真核细胞中参与翻译起始复合物形成的是57. 绿脓杆菌外毒素抑制真核细胞蛋白质合成的靶点是58. 真核细胞中识别终止密码子的是A. 阻遏蛋白B. 转录因子C. cAMP-CAPD. 终止因子E. 核蛋白体59. 色氨酸操纵子的转录调控依赖于60. 乳糖缺乏时,乳糖操纵子不转录,这主要取决于61. 葡萄糖缺乏时,乳糖操纵子转录,这主要取决于A. 基础转录因子B. 特异转录因子C. 起始因子D. 阻遏蛋白E. ρ因子62. 人血红蛋白表达特异性的决定因素是63. 小鼠异柠檬酸合成酶的表达需要哪一类蛋白质因子辅助64. 大肠杆菌β-半乳糖苷酶表达的关键调控因素是X型题65. 管家基因的转录受哪些因素控制A. 基础转录因子B. 增强子C. 特异转录因子D. 启动子E. 反应元件66. 大肠杆菌乳糖操纵子中,属于调控元件的是A. 操纵基因B. 调节基因C. 启动子D. 阻遏蛋白E. CAP结合位点67. 色氨酸操纵子模型中,哪些因素参与调控作用A. 释放因子B. 核蛋白体C. CAP复合物D. 阻遏蛋白E. 衰减子68. 原核生物转录终止的调控机制涉及A. RNA干扰B. 分解物阻遏C. ρ因子D. 衰减E. 抗终止69. 真核生物的基因表达调控表现在A. 转录水平B. 翻译水平C. 染色质水平D. 转录后加工E. 翻译后加工70. 真核生物中,影响RNA聚合酶转录活性的因素包括A. 启动子B. 增强子C. 基础转录因子D. 衰减子E. 特异转录因子71. 真核生物基因表达的空间特异性的机制包括A. 特异转录因子的种类不同B. 同种特异转录因子的浓度不同C. 特定组织的基因中存在组织特异性启动子D. 特异转录因子的排列组合不同E. 增强子等调控元件在不同组织的基因中分布不同72. 转录因子的DNA结合结构域包含哪些结构类型A. 螺旋-片层-螺旋B. 锌指C. 螺旋-转角-螺旋D. 亮氨酸拉链E. 螺旋-环-螺旋73. 与siRNA相比较,miRNA的显著特点是A. 单链B. 在转录后水平发挥作用C. 与靶mRNA碱基互补D. 不降解靶mRNAE. 个别碱基与靶mRNA序列不完全匹配74. 转录因子的作用机制包括A. DNA-DNA相互作用B. DNA-RNA相互作用C. DNA-蛋白质相互作用D. RNA-蛋白质相互作用E. 蛋白质-蛋白质相互作用75. 真核生物的基因转录涉及哪些物质的相互作用A. operatorB. cis-acting elementC. polysomeD. trans-acting factorE. RNA polymerase76. 在同一个体的不同组织中A. 基因的表达谱不同B. 基因组结构不同C. 特异转录因子的种类不同D. 存在的蛋白质的种类不同E. 特异性启动子的种类不同77. 真核生物独有的转录调控机制涉及A. 启动子B. 增强子C. 转录因子D. 组蛋白E. SD序列78. 生物对环境的适应性表现在A. 基因变异B. 合成不同种类的mRNAC. 合成不同种类的蛋白质D. 产物的反馈抑制E. 蛋白质活性的快速调节79. 下列哪些情况对于真核生物的基因转录具有调控作用A. 反式作用因子的磷酸化B. 类固醇激素与胞内受体结合C. 特定DNA序列的甲基化D. 组蛋白的乙酰化E. 蛋白质因子的羟基化80. 管家基因的含义是A. 在各组织细胞中都表达B. 在特定的组织细胞中表达C. 在不同发育阶段都表达D. 在特定的发育阶段表达E. 表达程度在不同时空条件下差异显著81. 关于特异转录因子的描述,正确的是A. 在所有组织细胞中组成性表达B. 在不同组织细胞中存在的种类不同C. 在不同组织细胞中的浓度不同D. 调控管家基因的转录E. 是真核生物基因表达特异性的根源所在(二)名词解释1. 管家基因(housekeeping gene)2. 可调节基因(regulated gene)3. 顺式作用元件(cis-acting element)4. 反式作用因子(trans-acting factors)/转录因子(transcription factor, TF)5. 基础转录因子(basal/general transcription factor)6. 特异转录因子(special transcription factor)7. 操纵子(operon)8. 衰减(attenuation)9. 锌指(zinc finger)结构10. 亮氨酸拉链(leucine zipper)结构(三)简答题1. 简述顺式作用元件与反式作用因子对基因表达调控的影响。
《分子生物学导论》笔记_学习笔记

《分子生物学导论》笔记第一章:分子生物学概述1.1分子生物学的定义与发展1.2分子生物学的研究对象1.3分子生物学与其他学科的关系1.4分子生物学的重要性第二章:DNA的结构与功能2.1DNA的双螺旋结构2.2DNA的复制机制2.3DNA的修复与重组2.4DNA的功能与基因表达第三章:RNA的类型与作用3.1信使RNA(mRNA)3.2转运RNA(tRNA)3.3核糖体RNA(rRNA)3.4小RNA及其功能第四章:蛋白质的合成与功能4.1转录与翻译过程4.2蛋白质的结构层次4.3蛋白质的折叠与修饰4.4蛋白质的功能与作用机制第五章:基因调控机制5.1基因表达调控的基本概念5.2转录因子与增强子5.3表观遗传学与基因表达5.4RNA干扰与基因沉默第六章:分子生物学的应用6.1分子生物学在医学中的应用6.2分子生物学在农业中的应用6.3分子生物学在生物技术中的应用6.4未来发展与挑战第1章:分子生物学概述分子生物学的定义与发展分子生物学是研究生命现象的分子基础的科学,主要关注生物大分子的结构、功能及其相互作用。
其核心内容包括DNA、RNA和蛋白质的相互关系。
分子生物学的起源可以追溯到20世纪初,随着显微镜技术的发展,科学家们对细胞组成的认识逐渐深入。
1940年代,随着DNA的双螺旋结构被发现,分子生物学开始正式形成。
关键概念包括:DNA(脱氧核糖核酸):遗传信息的载体,结构为双螺旋。
RNA(核糖核酸):在基因表达中起到中介作用,主要类型有信使RNA(mRNA)、转运RNA(tRNA)和核糖体RNA(rRNA)。
蛋白质:由氨基酸构成,承担细胞内外的多种功能。
重要发展里程碑:1953年,沃森和克里克提出DNA双螺旋结构。
1961年,霍普金斯等人发现RNA的转译机制。
1970年代,基因工程技术的引入,推动了分子生物学的应用。
考点:分子生物学定义的准确描述DNA、RNA和蛋白质的基本功能和相互关系重要历史事件及其影响分子生物学的研究对象分子生物学的研究对象主要包括核酸(DNA和RNA)、蛋白质、酶及其相互作用。
分子生物学第六章:DNA损伤与修复

48
4.直接插入嘌呤
DNA链上嘌呤的脱落造成无嘌呤位点,能被
DNA嘌呤插入酶(insertase)识别结合,并在K+
存在下催化游离的嘌呤碱基或脱氧核苷与DNA无
嘌呤部位形成糖苷键。且催化插入的碱基有高
度专一性、与另一条链上的碱基严格配对,使
DNA完全恢复。
49
三、碱基切除修复(Base
Excision Repair,BER)
35
第二节
错配修复
DNA修复
DNA的修复主要类型:
直接修复
切除修复 重组修复 跨损伤修复 (SOS修复)
36
一、错配修复
在DNA复制过程中, DNA聚合酶能够利用
其3ˊ一5ˊ外切核酸酶活性去除错配核苷酸,但
是这种校正作用并不十分可靠, 某些错配核苷酸
可能逃避检测, 出现于新合成的DNA链中。 错
胞嘧啶
O6-乙基鸟嘌呤 胸腺嘧啶
25
(一)烷化剂对DNA的损伤 2.碱基脱落 烷化鸟嘌呤的糖苷键不稳定,容易脱 落形成DNA上的无碱基位点,复制时可以 插入任何核苷酸,造成序列的改变。
26
(一)烷化剂对DNA的损伤
3.断链
DNA链的磷酸二酯键上的氧也容易被 烷基化,结果形成不稳定的磷酸三酯键, 易在糖与磷酸间发生水解,使DNA链断裂。
不识别任何特殊的碱基损失,而是识 别双螺旋形状的改变;修复时切除含有损 伤碱基的那一段 DNA。
54
55
56
核苷酸切除修复 (大肠杆菌)
紫外线诱导uvrA、 uvrB、uvrC和uvrD 四种基因表达
UvrA:识别损伤 部位 UvrB:解旋双链
57
UvrC:
5ˊ末端内切
分子生物学总结

分⼦⽣物学总结分⼦⽣物学总结第⼀章绪论⼀. DNA重组技术和基因⼯程技术.DNA重组技术⼜称基因⼯程,⽬的是将不同的DNA⽚段按照⼈们的设计定向连接起来,在特定的受体细胞中与载体同时复制并得到表达.产⽣影响受体细胞的新的遗传性状.基因⼯程技术还包括其他可能使⽣物细胞基因组结构得到改造的体系.第⼆章染⾊体与DNA⼀. DNA的⼀、⼆、三级结构特征.DNA⼀级结构特征1. 双链反向平⾏配对⽽成2. 脱氧核糖和磷酸交替连接,构成DNA⾻架,碱基排在内侧3. 内侧碱基通过氢键互补形成碱基对DNA⼆级结构特征绕DNA双螺旋表⾯上出现的螺旋沟,宽的沟称为⼤沟,窄沟称为⼩沟。
⼤沟,⼩沟都、是由于碱基对堆积和糖-磷酸⾻架扭转造成的。
DNA三级结构特征拓扑异构酶拓扑异构酶负超螺旋松弛DNA 正超螺旋溴已啶溴已啶⼆. 原核⽣物DNA具有哪些不同于真核⽣物DNA的特征.1. 结构简练2. 存在转录单元3. 有重叠基因三. DNA复制通常采取哪些⽅式.1. 线性DNA双链的复制.2. 环状DNA双链的复制分为θ型、滚环型和D-环型等.四. 原核⽣物DNA的复制特点.1. DNA双螺旋的解旋2. DNA复制的引发3. 冈崎⽚段与半不连续复制4. 复制的终⽌5. DNA聚合酶五. 细胞通过哪⼏种修复系统对DNA损伤进⾏修复?1. 错配修复2. 碱基切除修复3. 核苷酸切除修复4. DNA直接修复六. 什么是转座⼦?可分为哪些种类?转座⼦是存在与染⾊体DNA上可⾃主复制和位移的基本单位原核⽣物转座⼦的类型: 1. 插⼊序列 2. 复合转座⼦ 3. TnA家族第三章⽣物信息的传递(上)⼀. 什么是编码链?什么是模板链?与mRNA序列相同的那条DNA链称为编码链;将另⼀条根据碱基互补原则指导mRNA合成的DNA链称为模板链。
三. 简述σ因⼦的作⽤.σ因⼦的作⽤是负责模板链的选择和转录的起始,它是酶的别构效应物,使酶专⼀性识别模板上的启动⼦.四. 什么是Pribnow box?它的保守序列是什么?RNA聚合酶全酶与模板DNA结合后,⽤DNase I⽔解DNA,然后⽤酚抽提,沉淀纯化DNA后得到⼀个被RNA聚合酶保护的DNA⽚段,约有41-44个核苷酸对.在被保护区内有⼀个由5个核苷酸组成的共同序列,是RNA聚合酶的紧密结合点,称为Pribnow box. Pribnow区的保守序列是: TTGACA五. 简述原核⽣物和真核⽣物mRNA的区别.(⼀)原核⽣物mRNA的特征1、半衰期短2、多以多顺反⼦的形式存在3、5’ 端⽆“帽⼦”结构, 3’ 端没有或只有较短的polyA 结构。
医学分子生物学第六章_信号转导
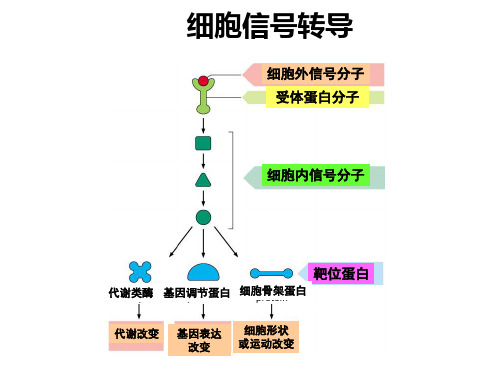
调节蛋白质功能 水平,调节细胞分化和增
和表达水平
殖
受体的结构特点
• 结合结构域-----识别外源信号分子并与之结 合
• 效应结构域-----转换配体信号,使之成为细 胞内分子可识别的信号
3、信号转导分子和分子开关
• 信号转导分子(signaling molecule):细 胞内执行信号转导的成分的一些蛋白质分 子和小分子活性物质。
• 信号转导分子组织在支架蛋白上的意义:
① 保证相关信号转导分子容于一个隔离而稳定的信号转导 通路内,避免与其他不需要的信号转导通路发生交叉反 应,以维持信号转导通路的特异性;
② 增加调控复杂性和多样性。
信号转导通路中的一些环节是由多种分子聚集形成的 信号转导复合物(signaling complex)来完成信号 传递的。
激酶
磷酸基团的受体
蛋白丝氨酸/苏氨酸激酶 蛋白酪氨酸激酶 蛋白组/赖/精氨酸激酶 蛋白半胱氨酸激酶 蛋白天冬氨酸/谷氨酸激酶
丝氨酸/苏氨酸羟基 酪氨酸的酚羟基 咪唑环,胍基,ε-氨基 巯基 酰基
蛋白磷酸酶衰减或终止蛋白激酶诱导的效应
• 蛋白质磷酸酶(phosphatidase)使磷酸化的 蛋白分子发生去磷酸化,与蛋白激酶共同 构成了蛋白质活性的调控系统。
及信息传递,是指一个细胞发出的信息通过介 质传递到另一个细胞并与靶细胞相应的受体相 互作用,然后通过信号转导产生胞内一系列生 理生化反应,最终表现为细胞整体的生物学效 应的过程。
T淋巴细胞
(一)细胞通讯的方式
靶细胞
细胞间隙连接
细胞表面分子接触通讯 可溶型信号分子
化学信号介导通讯
❖分泌化学信号
根据体内化学信号分子作用距离,可以将 其分为三类:
分子生物学 第六章

摆动性
• 反密码子与密码子之间的配对并不完全遵照 碱基互补规律,称为摆动配对。
二、tRNA
(一)结构特点 1.二级结构:三叶草结构
四环: 二氢尿嘧啶环 反密码子环 额外环 胸腺嘧啶假尿嘧啶胞嘧啶环 一臂: 氨基酸接受臂
2.三级结构——“倒L型”
(二)起始tRNA
密码子 氨基酸 表示方法
(二)延伸
1.进位 • 氨酰-tRNA 按照mRNA 分子的编码 信息进入并 结合到核糖 体A位。
(二)延伸
2.成肽
• 转肽酶催化 肽酰-tRNA 上的肽酰基 转移到A位 氨酰-tRNA 上的氨基酸 α-氨基上。
(二)延伸
3.转位
• 转位酶催化核 糖体沿mRNA 的3‘方向移动 一个密码子的 距离,使 mRNA上的下 一个密码子进 入A位,肽酰tRNA由A位移 入P位。
三、修饰
(一)磷酸化 是指在蛋白激酶的催化作用下,ATP的γ-磷酸 基被转移到蛋白质特定位点上的过程。 通常蛋白质的丝氨酸、苏氨酸和在糖基转移酶的作用下,蛋白质的特定 氨基酸残基被共价连接上寡糖链的过程。 • 糖链与氨基酸的连接主要有O型连接和N型 连接两种方式。
终止密码子: 琥珀石(UAG) 赭石(UAA) 卵白石(UGA)
起始密码子: AUG(甲硫氨酸)
2.特性
(1)完整性:有始有终 (2)方向性:5’到3’ (3)连续性:不中断、无重叠 (4)简并性:多对一 (5)统一性:万物统一 (6)摆动性::3’位可变 (7)偏爱性:使用频率各异
简并性
• 一种氨基酸具有 两个或两个以上 的密码子为其编 码,这一特性称 为遗传密码的简 并性。
一、mRNA (一)结构特点
原核 生物
真核 生物
第六章 基因调控1:原核生物基因的表达调控 分子生物学习题

第六章基因调控1:原核生物基因的表达调控分子生物学习题第六章基因调控1:原核生物基因的表达调控分子生物学习题第六章基因表达调控1:原核生物基因表达调控名词解释:操纵子基因表达持家基因正调控和负调控安慰诱导物衰减子(弱化子)魔斑结构基因和调节基因本底水平表达填空1操纵子的基因表达调节系统属水平的调节,乳糖操纵子模型由和1961年明确提出的。
色氨酸操纵子包含和两方面的调控。
2、能够诱导操纵子但不是代谢底物的化合物称为诱导物。
能够诱导乳糖操纵子的化合物就是其中一例。
这种化合物同蛋白质结合,并使之与分离。
乳糖操纵子的体内功能性诱导物是。
3、色氨酸就是一种调节分子,被视作。
它与一种蛋白质融合构成。
通过掌控起至促进作用。
色氨酸操纵子受到另一种系统------的调控,它牵涉至第一个结构基因被mRNA前的mRNA。
4、大肠杆菌乳糖操纵子调节基因编码的与结合,对lac结合,对lac表达实施负调控;与复合物结合于上游部分,对lac表达实施正调控。
5、操纵子中没有基因产物的是和选择题1、下面哪些真正就是乳糖操纵子的诱导物?()a.乳糖b.蜜二糖c.o-硝基苯酚-β-半乳糖苷(onpg)d.异丙基-β-半乳糖苷e.异乳糖2、色氨酸操纵子的调控促进作用就是受到两个相互单一制的系统控制的,其中一个须要前导肽的译者,下面哪一种调控这个系统?()a.色氨酸b.色氨酰-trnatrpc.色氨酰-trnad.campe.以上都不正确3、Dozul蛋白(阻抑蛋白)辨识操纵子中的()a逼舳基因b苯峁够因c辈僮莼因d蹦诤子e蓖庀宰4、乳糖、阿拉伯糖、色氨酸等小分子物质在基因表达调控中作用的共同特点是a庇肫舳子结合b庇dna结合影响模板活性c庇rna聚合酶结合影响其活性d庇氲鞍字式岷疑跋旄玫鞍字式岷dnae庇氩僮莼因融合5.下面那项不属于原核生物压低元的结构a:启动子b:终止子c:操纵子d:内含子6、以下有关操纵子的阐释哪个就是错误的()a操纵子是由启动基因、操纵基因与其所控制的一组功能上相关的结构基因组成的基因表达调控单位b操纵子不包含调节基因c代谢底物往往是该途径的可诱导酶的诱导物,代谢终产物往往是可阻遏酶的辅阻遏物d真核细胞的酶合成也存在诱导和阻遏现象,因此也是由操纵子进行调控的7、操纵子调节系统属于哪一种水平的调节?a复制水平的调控b转录水平调控c转录后加工的调控d翻译水平的调控8、对调节基因下述哪些论述哪些是对的()a是编码阻遏蛋白的结构基因b各种操纵子的调节基因都与启动基因二者紧邻c调节基因就是操纵子的组成部分d 调节基因的抒发Seiches迁移的调控区9、以下有关Dozul蛋白的哪些就是对的()aDozul蛋白就是调节基因表的的产物b可诱导操纵子的阻遏蛋白具有直接与操纵子基因结合的活性,与诱导物相互作用后丧失活性c可以Dozul操纵子的Dozul蛋白没轻易与操纵子基因融合的活性,与辅阻遏物融合后才有此活性d阻遏蛋白可与rna聚合酶竞争同一结合部位10、关于启动基因的下述论点哪些是错误的()a启动基因就是rna聚合酶辨识并最县融合的一段dna序列b启动基因就是最先被rna聚合酶mRNA的dna序列c启动基因就是dna上含有a-t碱基对的部分d启动基因就是引起dna激活的特定序列11、下列有关降解物基因活化蛋白(cap)的哪些论点是正确的()acap-camp可专一地与启动基因结合,促进结构基因的转录bcap可单独与启动子相互作用,促进转录ccap-camp可以与调节基因融合,掌控Dozul蛋白的制备dcap-camp可与rna聚合酶竞争地结合于启动基因,从而阻碍结构基因的转录12、与乳糖操纵子操纵基因结合的物质是()arna聚合酶bdna聚合酶cDozul蛋白d反华密码子是非题1、葡萄糖和乳糖并存时,细菌优先利用乳糖并启动乳糖操纵子()2、小分子物质如itpg诱导乳糖操纵子抒发时起负调控促进作用与操纵基因结合阻抑结构基因的抒发()3、色氨酸操纵子中含有衰减子区,其调控作用主要受trp浓度高低影响()4、色氨酸操纵子(trpoperon)中含有衰减子序列()5、camp在laz操纵子中起正调控作用,其浓度受环境中的葡萄糖影响,与其浓度成正比()6、大肠杆菌乳糖操纵子真正的诱导物不是乳糖,而是它的异构体别乳糖()7、操纵基因又称操纵子,如同启动基因又称启动子一样()8、可诱导操纵子是负责调节糖分解代谢的,可阻遏操纵子是负责调节氨基酸代谢的()问答题:1、试述乳糖操纵子的结构及负控诱导的调控机理2、色氨酸操纵子的结构特点?其弱化子在调控中如何起至促进作用?3、详述色氨酸操纵子中启动子调控促进作用特点。
分子生物学全套课件(2024)

2024/1/26
17
蛋白质在细胞中的作用
蛋白质可以作为酶催化生物体内 的化学反应,维持生命活动的正 常进行。
蛋白质可以作为载体运输物质, 如血红蛋白运输氧气和二氧化碳 。
蛋白质可以作为抗体参与免疫反 应,保护机体免受病原体的侵害 。
蛋白质是细胞结构和功能的基础 ,参与细胞的各种生命活动,如 催化、运输、免疫、调节等。
2024/1/26
21
基因表达调控的分子机制
DNA结合蛋白的作用
识别并结合特定DNA序列,影响基因转录。
染色质结构与基因表达
染色质结构的变化可影响基因的可及性和转 录活性。
2024/1/26
信号转导与基因表达调控
细胞外信号通过信号转导途径影响基因表达 。
转录后调控机制
包括mRNA剪接、转运、定位和降解等过程 对基因表达的调控。
比较基因组学分析
通过比较不同物种或不同个体之间的基因组差异,揭示物种进化、基 因功能等生物学问题。
生物信息学在基因组学中的应用
利用生物信息学方法对基因组数据进行挖掘和分析,发现新的基因、 突变位点以及与疾病相关的遗传变异等。
27
THANK YOU
感谢观看
2024/1/26
28
2024/1/26
8
DNA的复制与修复
01
02
03
DNA复制的过程
起始、延伸和终止三个阶 段,涉及多种蛋白质和酶 的参与。
2024/1/26
DNA复制的特点
半保留复制、半不连续复 制等。
DNA修复的机制
直接修复、切除修复、重 组修复和SOS修复等,用 于纠正复制过程中产生的 错误。
9
DNA的转录与表达
第六章_酶分子修饰
一分子RNA酶 + 6.5分子右旋糖苷——酶活提高为 原来的2.25倍。
一分子胰凝乳蛋白酶 + 11分子右旋糖苷——酶活 提高原来的 5.1倍。
一分子胰蛋白酶 + 11分子右旋糖苷——酶活提高 0.30倍
(2)增加酶的稳定性
酶的保存或使用一段时间后,由于各种因素影响,原 来完整的空间结构受到破坏,酶活力降低,甚至完全 丧失催化能力。
聚乙二醇琥珀酰亚胺衍生物:
是MPEG的羟基与琥珀酰亚胺类物质反应。在pH7-10时, 对酶的氨基进行修饰。
聚乙二醇胺类衍生物:
是MPEG的羟基与胺类化合物反应生成的。可以对酶的羰 基进行修饰。
聚乙二醇马来酸酐衍生物:
MPEG的羟基与马来酸酐反应生成共聚物(PM),其中 马来酸酐通过酰胺键对酶分子的氨基进行修饰。
3)修饰:
活化后的大分子修饰剂与经分离纯化的酶液,以一 定的比例混合,在一定的温度、pH值等条件下反 应,使两者以共价键结合。 4)分离:
通过凝胶层析等方法进行分离,将具有不同修饰度 的酶分子分开,从中获得具有较好修饰效果的修饰 酶。
右旋糖苷————(高碘酸HIO4)活化右旋糖苷
三、大分子修饰的作用
酶对人体是一种外源蛋白,当经注射进入人体后 会成为抗原,刺激体内产生抗体。当这种酶再次进 入体内,抗体就会与作为抗原的酶特异结合,而使 酶失去催化能力。
抗体与抗原间特异结合是由于它们之间特定分子结构引 起的。用酶分子修饰法使酶的结构产生改变,抗体、抗 原就不再特异结合,可能降低或消除其抗原性。
利用水溶性大分子对酶进行修饰可降低或消除酶的抗原 性。
2-羟基-5-硝基苄溴(HNBB)和4-硝基苯硫 绿可以比较专一地对吲哚基进行修饰,但也可以 与巯基反应。
细胞分子生物学 第六章 内质网和蛋白质合成

一、化学组成
1、水和无机离子等一些小分子。离子主要是Na+、K+、Ca2+、Mg2+、
Cl-等。 2、脂类、糖类、氨基酸、核苷酸及其衍生物等中等分子。
3、蛋白质、多糖和RNA等大分子。
4、含有糖原等一些处于贮存状态的重要化合物。
二、细胞质溶质的基本属性
1、酸度稳定
主要内容
第一节 细胞质溶质
第二节 内质网 第三节 核糖体 第四节 蛋白质合成的命运
1 、胞质溶质:是指细胞质中可溶性 的液体。含有大量的各种可溶性蛋白 和代谢物质。在细胞的物质代谢、维 持细胞内环境稳定性等方面具有非常 重要的作用。 2.内膜系统:细胞内的许多膜性细胞 器在结构和功能上有一定的连续性, 构成了膜体系,故称为内膜系统,主 要包括内质网、高尔基体、溶酶体等, 但不包括半自主性的线粒体和叶绿体。
SRP为GTP结合蛋白,可结合GTP。当SRP与信号序列和核糖体结合
后,翻译过程暂时停止。 糙面内质网上有信号识别颗粒的受体,SRP引导核糖体与内质网膜 上的的SRP受体结合。内质网膜上存在转运体,转运体可形成通道。当 SRP与SRP受体结合后,新生肽链随即从核糖体的大单位进入转运体的中
央通道。
肽链信号序列进入内质网腔后,随即被膜上与转运体结合的信号肽 酶切除。切除信号肽的新生肽链则继续穿过转运体通道进入内质网腔。
动有条不紊地进行。细胞骨架对细胞质溶质中的各种小泡运输也具有 导向作用。
三、细胞质溶质的功能
1、维持一些代谢反应途径
细胞质溶质中含有很多酶,大部分中间代谢是在细胞质溶质中进 行的。例如:糖酵解的起始步骤
2、保持细胞内环境的稳定
缓冲pH值,使酸度保持在 pH7.2 ,为各种中间代谢反应的进行
第六章:分子生物学研究方法(下全文编辑修改

人类染色体端粒DNA的荧光原位杂交
4. 基因定点突变技术
• 通过改变基因特定位点核苷酸序列来改变所编码 的氨基酸序列,用于研究某个(些)氨 基酸残基 对蛋白质结构功能的影响。
二、基因敲除技术
• 基因敲除(gene knock-out)又称基因打靶,通 过外源DNA与染色体DNA之间的同源重组,进行 精确的定点修饰和基因改造。
方法: • 以9~11bp作为标签(tag)=49=262144组合 • 串联tag并通过两端接头PCR扩增 • 扩增产物进行测序 • 每个tag通过Genebank或EST数据库进行比对 • 确认tag代表的基因表达情况
2. RNA的选择性剪接从一个 mRNA前体产生不同的mRNA剪接异构体的过程。
2.凝胶阻滞试验 ➢ 是用于研究DNA与蛋白质相互作用的一种
特殊的凝胶电泳技术。
➢ 当DNA与蛋白质结合时,在电泳中会受到阻滞, 说明可能与某种特殊蛋白结合了。
3. DNAase足迹试验 ➢ 蛋白质结合在DNA片段上,能保护结合部位不被
DNAase水解,电泳中对应于结合部位没有条带。
• 常用RT-PCR法研究某个基因是否存在选择性剪切。
3.原位杂交技术( In Situ Hybridization,ISH)
• 用标记的核酸探针,在组织、细胞上对核酸进行 定位和相对定量研究的一种手 段。
• 探针标记用同位素或地高辛、生物素荧光标记等。
地
同
高
位
辛
素
标
标
记
记
染色体原位杂交
三、蛋白质与RNA、DNA相互作用
1. 酵母单杂交系统 • 将待测转录因子cDNA与表达载体导入酵母细胞,
该基因产物如 果能够与顺式作用元件相结合,就 能激活启动子,使报告基因得到表达。
分子生物学(杨洋)第六章 rna剪接-rna splicing

Step 1: The OH of the conserved A at the branch site attacks the phosphoryl group of the conserved G in the 5’ splice site. As a result, the 5’ exon is released and the 5’-end of the intron forms a three-way junction structure.
Trans-splicing:原本位于两个不同RNA分子 上的外显子可以连接到同一条mRNA中
标准剪接:套索结构
反式剪接:Y型分支结构
Figure 13-5
Not a lariat
Topic 2 THE SPLICESOME MACHINERY (剪接体)
2.1 RNA splicing is carried out by a large complex called spliceosome (RNA剪接是由一个称为剪接体的大复合体 执行的) The above described splicing of introns from pre-mRNA are mediated by the spliceosome. The spliceosome comprises about 150 proteins and 5 snRNAs(5种RNA,核内小RNA). Many functions of the spliceosome are carried out by its RNA components(剪接体的多数 功能是由其RNA而不是蛋白质执行的)
分支点
RNA-RNA interactions between different snRNPs, and between snRNPs and pre-mRNA
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
❖ 为了使目的基因的特定位点发生突变,所设计的寡 核苷酸引物的序列除了所需的突变碱基外,其余的 则与目的基因编码链的特定区段完全互补。
2.突变过程
1)合成含有目的 基因的正链DNA; 2)合成含有特殊 突变碱基的引物; 3)制备异源双链 DNA; 4)富集和转化双 链DNA分子; 5)筛选突变体并鉴定
❖ B Ile Cys Thr Thr Ala Thr Cys 1
❖ C Ile Ile Cys Thr Ala Cys Leu 1
❖ D Cys Cys Thr Thr Cys Thr Cys 2
❖ E Ile Cys Cys Thr Ala Cys Cys 2
❖ F Cys Cys Cys Thr Cys Cys Cys 3
❖ 将β-IFN分子中的游离Cys残基用Ser来取代,因为 Cys与Ser分子结构相似,前者有一个S原子,而后 者是一个O原子,减少了二聚体的形成,提高了活 性蛋白质的得率。
增加酶的活性
❖ 用定点突变技术不仅可以提高蛋白质的稳定 性,还可以提高酶的活性。要改变酶的活性, 需要一个详细描述了酶的活性位点的图谱。 有了这样的资料,研究者们即可推测酶与底 物的亲和程度,并利用定点突变技术对蛋白 质进行改造,提高酶的活性。
(二)Kunkel定点突变法
❖ 体外DNA合成往往是不完全的,所以部分合 成的DNA分子必须通过蔗糖密度梯度离心除 去,获得纯化的突变DNA。
❖ 理论上来说,DNA是半保留复制的,应用寡 核苷酸定点突变时,所形成的噬菌体中携带 突变基因的应为一半。但实际上,由于技术 上的原因通常只有1-5%的噬菌斑含有突变基 因的噬菌体。
❖ 当感染型单链M13噬菌体感染大肠杆菌后,在菌体内借助宿 主的酶系统先把ssDNA复制为双链(dsDNA),称为复制 型(replication form,RF)M13(RF- M13)。
❖ 广泛用于DNA序列分析和噬菌体表面展示系统(phage display)和单链核酸的制备。
1.基本原理
❖ 再使用化学合成的含有突变碱基的寡核苷酸短片段 作引物,启动M13单链DNA分子进行复制,随后这 段寡核苷酸引物便成为新合成的DNA子链的一个组 成部分。
天冬氨酸(Asp) + NH3
❖ 谷氨酰胺(Gln)
谷氨酸(Glu) + NH3
❖ 导致肽链折叠的局部的改变,可能影响其活性。
❖ 如酵母的丙糖磷酸异构酶是由两个相同的亚基组成的二聚体, 每个亚基都含有两个Asn残基,均位于两个亚基相互接触的 表面上,可能与该酶的热稳定性有关。
❖ 若将两个Asn全部换成Asp,则变体酶即使在常温下也不稳 定,而且酶活性也大为降低;
❖ 如枯草杆菌蛋白酶之所以易被氧化失活,是 由于催化部位的丝氨酸(Ser221)邻近的甲硫 氨酸(Met222)易被氧化成硫氢化物,若以其 他氨基酸取代Met222,则可提高酶的氧化稳 定性而又不影响其催化活性。这是结构分析 和定点突变改造蛋白质的成功范例。
❖ 枯草杆菌蛋白酶可作为洗涤剂的添加剂,但由于其 只能水解苯丙氨酸(Phe)羧基所形成的肽键,底物 作用范围过窄而限制了洗涤剂的高效性,若用带正 电荷的赖氨酸(Lys)取代位于活性中心166位的甘氨 酸(Gly),所获得的突变酶不仅能水解苯丙氨酸(Phe) 羧基所形成的肽键,而且可以水解酸性氨基酸谷氨 酸(Glu)所形成的肽键,使其底物作用范围拓宽,因 而可能成为最高效的洗涤剂添加酶,这是定点突变 改变蛋白质生物学活性的成功例子。
③ 双链DNA导入正常的大肠杆菌 中,其尿嘧啶N-糖基化酶除去 DNA链上的尿嘧啶碱基。
④ 结果原来的M13模板链被降解, 只有突变链因不含U,被保留下 来。这种方法产生的M13噬菌体 中含有突变DNA的比例大大增 加。
(三) PCR定点突变
❖ Polymerase Chain Reaction (PCR)是一 种体外酶促合成特定DNA片断的技术,是根 据人类的需要对复杂生命过程的一种简单化 的模拟。PCR技术的原理是DNA半保留复制。 Kari.B.Mullis首创。
基因,即可表达出适合于商业用途的蛋白质。
❖ 在体外,通过碱基取代、插入或缺失的方法, 使基因DNA序列中的某个特定碱基发生改变, 从而改变蛋白质的结构,称为蛋白质工程。
❖ 通过蛋白质工程,人们可以随心所欲地改变 蛋白质的结构及其理化性质和生物学功能。 例如,我们可以利用蛋白质工程改变酶的Km、 Vmax,酶促反应的最适温度、最适pH值、 酶促反应的特异性以及酶蛋白的稳定性等。
先设置一个PCR反应产生一个含突变的 DNA片段,然后再以此DNA片段作为引物 与原模板退火进行PCR扩增得到含突变的完 整的基因。因为作为引物使用的DNA片段较 通常的引物要大许多甚至有上百碱基,所以 命名为大引物PCR法。
大引物PCR 定点突变技术路线
(圆点指突变位点)
3、环状突变 PCR 法
定义:以整个带有目的基因的质粒为模板,用突变剂 进行扩增,最后产生环状的带有目的突变的质粒。
方法: ①两个引物反向、紧邻但没有重叠区,扩增产物是平
末端的线性DNA,需用T4连接酶环化处理。 ②以Stratagene公司开发的定点突变试剂盒为代表,
两个引物也是反向的并且其5’端有l5个碱基以上的 重叠区,扩增产物为带黏性末端的线性DNA,可自 行环化。
❖ 例如:当人β-干扰素基因(β-IFN)在大肠杆菌中表 达时,尽管其表达量很高,但其比活性只及天然蛋 白质的10%。
❖ DNA序列分析显示, β-IFN所编码的蛋白质有三个 Cys残基,其中2个形成分子内二硫键,而另一个游 离的Cys可能涉及到β-IFN分子间二硫键的形成,导 致二聚体的产生,使单聚体含量减少,活性降底。
蛋白质分子的生物学改造
❖ 蛋白质在生命现象中具有重要作用,在医药、轻工 等行业也具有重要作用,如胰岛素、枯草杆菌蛋白 酶等;
❖ 天然生物材料中蛋白质含量较少; ❖ 基因克隆技术克隆基因,在宿主细胞中可表达特定
蛋白质(重组蛋白); ❖ 多数天然蛋白质的理化特性不能适应于工业用途; ❖ 利用现代分子生物学技术,改变克隆基因中的特定
环状突变 PCR 法
环状突变 PCR 法
通过特殊氨基酸的改变提高蛋白的稳定性
例:葡萄糖异构酶(Glucose isomerase) 用双引物法对葡萄糖异构酶基因进行了体外定点 诱变,以Pro138替代Gly138,在酶比活相近的情况下, 突变型葡萄糖异构酶的热半衰期比野生型长一倍,最 适反应温度提高10~12C。
具体步骤:
① 将待突变的基因克隆入M13 DNA载体上,导入具有dUTP酶 (dut)和N-尿嘧啶脱糖苷酶 (ung)双缺陷的大肠杆菌 (dut-,ung-)菌株中。
② Dut缺陷导致细胞内dUTP水平 上升,并在DNA复制时,部分 取代dTTP进入DNA新生链中。 又由于ung缺陷使掺入DNA的 dUTP残基不能除去。由这种大 肠杆菌菌株产生的M13单链 DNA大约有1%的T被U所取代, 然后进行DNA定点突变。
通过改变活性中心附近的氨基酸提高酶的催化活性
应用举例:大肠杆菌碱性磷酸酶
Ser101
Asp101
Wild Type
D101S
E.coli Alkaline phosphatase: Mutation of D101S
二、编码基因随机突变和重组
易错PCR(Error prone PCR) DNA改组(DNA shuffling)
11
❖ 酶的稳定性以在100℃时半衰期或者失活率来表示。 半衰期越长酶越稳定
减少游离Cys残基的数目
❖ 在利用DNA重组技术表达外源基因时,常常 会遇到这种情况:外源蛋白的表达量很大, 但其生物活性却很低,即外源蛋白的比活性 很低。
❖ 利用蛋白质工程技术减少游离的半胱氨酸 (Cys)残基数目,以减少蛋白错误折叠的 可能性,从而可以提高蛋白质的生物活性。
三、定点突变原理
四、定点突变的常用方法
(一)M13DNA寡核苷酸突变
❖ 基因工程中,噬菌体是一种常用的基因载体,其中又以M13 和 λ为最常用。
❖ M13噬菌体是一种环形DNA,基因组大小为6.4kb,在颗粒 中包装的仅是正链的DNA,有时也称为感染型单链 DNA(single stranded DNA, ssDNA) 。
(%) 100 100
96 106
0 95 0
0
(℃) 41.9 41.9 46.7 48.3 52.9 57.6 58.9 65.9
❖ wtα:野生型T4溶菌酶;pwt:假野生型酶;A-F:六种设计的半胱氨酸 变体;Tm:熔点温度
将Asn和Gln转换成其他氨基酸
❖ 当蛋白质暴露于高温时:
❖ 天冬酰胺(Asn)
❖ 蛋白质工程的主要技术之一是定点突变技术.
一、定点突变
❖ 利用分子生物学技术,在体外通过碱基取代、插入 或缺失可以使基因DNA序列中任何一个特定的碱基 发生改变。这种体外特异性改变某个碱基的技术, 称谓定点突变(site directed mutagenesis)。
❖ 定点突变具有简单易行、重复性高等优点,现已发 展成为基因操作的一种技术。这种技术不仅适用于 基因结构与功能的研究,还可通过改变基因的密码 子来改造天然蛋白质。
PCR进行寡核苷酸 定点突变的示意图
1、重叠延伸PCR (OE-PCR)
首先设计两个PCR反应以产生两个含突 变位点的DNA片段,随后将上述两个 PCR产物混合,二者通过末端互补区结 合并在适宜的温度下互为引物延伸得到 完整的含突变的基因,对第一次PCR产 物进行纯化处理可显著提高突变效率
2、大引物PCR法(megaprimer PCR )
据分析,可能由于Pro138替代Gly138引入的一个 吡咯环刚好能够填充Gly138附近的空洞,使突变蛋白 的空间结构更具刚性,从而提高了酶的热稳定性。