植物干细胞1
植物干细胞1

摘要:植物干细胞位于茎尖分生组织区和根尖分生组织区,是植物胚后发育中新的器官产生的源泉。
近几年,在干细胞及其周围组织区发现了一些与干细胞稳态维持有关的基因,这些基因产物与外源性信号(如生长素)一起组成复杂的调控网络控制植物的生长和发育。
表观遗传修饰作为控制基因表达的一种方式也对植物干细胞有重要的影响。
该文介绍近几年植物干细胞分化调控的最新进展。
关键词:植物干细胞;基因调控;生长素;表观遗传中图分类号:Q74动物干细胞因其在疾病治疗、组织修复和动物克隆等方面的广泛应用一直是研究人员和公众关注的焦点[1],相比之下,植物干细胞研究却门庭冷落。
由于植物细胞具有全能性,植物干细胞的概念也一直存在争议。
近几年的研究表明:在植物的茎尖分生组织(shoot apical meristem,.. SAM)和根尖分生组织(rootapical meristem,.. RAM)中,存在一群特殊的细胞,它们具有自我更新能力又能产生具有持续分裂能力的子细胞。
这些特殊细胞是植物根、茎、叶和花等器官发生的源泉,因此被认为是植物干细胞[2,3]。
与动物干细胞一样,植物干细胞的维持同样受到内源性信号和外源性信号的调控。
1. 植物干细胞的组织学特征模式植物拟南芥的SAM 是一个半球状的穹型结构,由多个功能结构域组成[图1(A)]。
干细胞位于分生组织的顶端中心区域,这个区域的细胞分裂不活跃。
中心区域中干细胞分裂后产生两部分细胞,一部分仍然保留在中心区域的叫干细胞后裔(progeny ofstem cells),.. 保持多潜能性,始终留守在原来位置,继承干细胞的衣钵;分裂出来的另一部分叫子细胞(daughter cells),.. 随着干细胞的分裂逐渐脱离中心区域到分生组织周边区域(peripheral zone,PZ),.. 在周边它们快速分裂并进行分化,融入分生组织两侧的器官原基中[4]。
由此可见,SAM是不断改变的动态结构,干细胞群的维持依赖于周围组织细胞提供的各种外源性和内源性信号分子。
植物干细胞的萃取方式

植物干细胞的萃取方式
植物干细胞的萃取方式通常涉及以下步骤,首先,选择合适的
植物组织,如茎、叶或根,这些组织通常含有丰富的干细胞。
然后,使用物理方法如超声波或高压萃取,以破碎细胞壁释放细胞内容物。
接下来,利用化学溶剂或酶解剂来提取细胞内的营养物质和活性成分。
萃取后的溶液经过过滤、离心和浓缩等步骤,最终得到植物干
细胞的提取物。
此外,一些先进的技术如超临界流体萃取和超滤技
术也被用于植物干细胞的提取,以确保提取物的纯度和活性成分的
保留。
总的来说,植物干细胞的萃取方式需要综合考虑植物的特性、萃取方法的效率以及提取物的纯度和活性,以确保最终产品的质量
和功效。
2020秋高一生物(2019)1学案:第6章 第2节 细胞的分化含解析

2020秋高一生物人教版(2019)必修1学案:第6章第2节细胞的分化含解析第2节细胞的分化学习目标核心素养1。
结合课本中实例,理解细胞分化的过程、特点及意义。
2.结合植物组织培养和非洲爪蟾的核移植过程,阐述细胞全能性的概念.3.搜集资料,了解干细胞的概念和研究意义.1.科学思维:归纳与概括细胞分化和全能性;明确其关系;阐述细胞分化实质.2.社会责任:举例说明细胞全能性,并解释其在生产实践中的应用.知识点一细胞分化及其意义1.概念:在个体发育中,由一个或一种细胞增殖产生的后代,在错误!形态、错误!结构和错误!生理功能上发生错误!稳定性差异的过程.2.特点3.实质:在个体发育过程中,不同种类的细胞中错误!遗传信息的表达情况不同(即基因的选择性表达)。
4.表现细胞水平:产生特定的细胞。
分子水平:合成特定的蛋白质。
5.实例:构成人体四种组织的细胞,如上皮细胞、骨骼肌细胞、软骨细胞、神经细胞等;植物的错误!叶肉细胞、表皮细胞、错误!储藏细胞等。
6.意义(1)是生物错误!个体发育的基础.(2)使多细胞生物体中的细胞趋向错误!专门化,有利于提高生物体各种错误!生理功能的效率。
错误!比较细胞分裂和细胞分化有什么区别和联系?提示:题型一细胞分化的实质及过程[例1]下列有关细胞分化的叙述,不正确的是()A.细胞分化是指相同细胞的后代在形态、结构和生理功能上发生稳定性差异的过程B.细胞分化是一种持久性的变化C.细胞分化使一个个体的各种细胞具有不同的遗传信息D.细胞分化是由于不同种类细胞中遗传信息的执行情况不同解题分析细胞分化没有改变细胞的遗传信息,C错误.答案C题型二细胞分化与细胞分裂的关系[例2]下列关于细胞分裂和细胞分化的叙述中,不正确的是()A.两者都是多细胞生物个体发育过程中不可缺少的过程B.无论是细胞分裂还是细胞分化,与亲代细胞相比,子细胞遗传物质都不会发生改变C.细胞分裂只发生在胚胎发育阶段,而细胞分化发生在生物体的整个生命过程中D.细胞分裂使细胞数目增加,细胞分化使细胞种类增加解题分析细胞分裂使细胞数目增加,在整个生命过程中都可能发生,细胞分裂不会使子细胞的遗传物质发生改变;细胞分化使细胞种类增加,它发生在生物体的整个生命进程中,且遗传物质也不会发生改变.答案C知识拓展分裂、分化与个体发育的关系知识点二细胞全能性1.概念:指细胞经分裂和分化后,仍具有产生完整错误!有机体或分化成其他错误!各种细胞的错误!潜能和特性。
2.1.1 植物细胞工程的基本技术(练习)(解析版)

2.1.1 植物细胞工程的基本技术练习一、单选题1.图为植物体细胞杂交过程示意图。
利用植物细胞A 和B 培育成为杂种植株。
该过程运用的技术有:①干细胞技术②细胞融合技术③细胞核移植技术④植物组织培养技术A.①②B.②④C.②③D.③④【答案】B【解析】据图分析,植物细胞A和B培育成为杂种细胞是植物体细胞杂交过程,运用了细胞融合技术,体现了细胞膜具有一定的流动性;杂种细胞形成杂种植株是植物组织培养过程,体现了植物细胞的全能性,B正确,ACD错误。
故选B。
2.利用植物体细胞杂交获得优良新品种过程中,不需要进行的是A.去除亲本体细胞的细胞壁B.选择具有优良性状的亲本C.利用灭活的病毒诱导原生质体融合D.诱导杂种细胞愈伤组织的再分化【答案】C【解析】依题文可知,植物体细胞杂交获得优良新品种包括原生质体融合及植物组织培养两个过程,由此需要选择具有优良性状的亲本,去除亲本体细胞的细胞壁获得原生质体,然后用物理法或化学法诱导原生质体融合,不需要用灭活的病毒诱导,最后诱导杂种细胞愈伤组织的再分化。
由此C正确,A、B、D错误。
故选C。
3.植物体细胞杂交要先去除细胞壁的原因是A.植物体细胞的结构组成中不包括细胞壁B.细胞壁使原生质体失去活力C.细胞壁阻碍了原生质体的融合D.细胞壁不是原生质体的组成部分【解析】细胞壁是植物细胞结构的一部分,A项错误;细胞壁对植物细胞具有保护作用,不会使原生质体失去活力,但阻碍了原生质体的融合,因此要除去细胞壁,B项错误,C项正确;细胞壁不是原生质体的组成部分,但不是植物体细胞杂交要先去除细胞壁的原因,D项错误。
故选C。
4.驱蚊草含有香茅醛,能散发出一种特殊的柠檬型香气,从而达到驱蚊且对人体无害的效果。
驱蚊草是把天竺葵的原生质体和香茅草的原生质体进行诱导融合培育而成的。
下列关于驱蚊草培育的叙述,错误的是A.驱蚊草的培育属于细胞工程育种,优点是克服了远缘杂交不亲和的障碍B.驱蚊草培育过程要用到纤维素酶、果胶酶、PEG等试剂或电刺激等方法C.驱蚊草培育过程不同于植物组织培养,无细胞脱分化和再分化的过程D.驱蚊草培育利用了植物体细胞杂交技术,育种原理是染色体数目变异【答案】C【解析】A、驱蚊草是把天竺葵的原生质体和香茅草的原生质体进行诱导融合培育而成的,采用了植物体细胞杂交技术,属于细胞工程育种,其优点是能克服远缘杂交不亲和的障碍,A正确;B、要获得天竺葵的原生质体和香茅草的原生质体,需要采用酶解法(纤维素酶、果胶酶),诱导天竺葵的原生质体和香茅草的原生质体的融合可采用化学法(PEG等试剂)或物理法(电刺激等),B正确;C、驱蚊草培育过程不同于植物组织培养,但需要采用植物组织培养技术,因此有愈伤组织和试管苗形成,C错误;D、驱蚊草培育利用了植物体细胞杂交技术,育种原理是染色体数目变异,D正确。
人教版高二生物选修三课件:2.1.1植物细胞工程的基本技术

课后作业
据下图回答问题:
(1)过程①②是用________________处理细胞的 细胞壁,后经③过程诱导原生质体融合,实 际融合的过程中原生质体可能会形成____种 细胞团(只考虑两个细胞融合的情况)。④是 ______________的过程,与此过程密切相关的 细胞器主要_____________。
条染色体,将番茄和马铃薯采用杂交育种方法培育 (假设能成功),得到的后代应含_____________条染 色体,还必须用_______________来处理幼苗,才能 得到可育的“番茄—马铃薯”植株,此时体细胞 中含有_____ 条染色体 (4)若番茄是二倍体,马铃薯是四倍体,则杂种植 株为______倍体。
基因在特定的时间和空间条 件下 选择性 表达的结果
5、植物细胞表现出全能性的条件:
离体 一定的营养物质、植物激素 适宜的温度、光照、pH、
无菌等外界条件
二
A、脱 分化
(排列疏松而无规则,
高度液泡化的呈无定 形状态的薄壁细胞)
愈伤
组织
植物激素: 细胞分裂素、生长素
三:植物体细胞杂交
1.概念 将不同种的植物体细胞,在一定条件下融合成杂种 细胞,并把杂种细胞培育成新的植物体的技术
2、这个过程中涉及的原理是?
细胞膜的流动性 植物细胞的全能性
三:植物体细胞杂交
1.你认为两个来自不同植物的体细胞完成融合, 5遇.到如的何第将一杂个种细 障胞 碍培 是育 什成么杂?种植株?
8.下列属于组织培养的是:( ) A.花粉培养成单倍体植株 B.芽发育成枝条 C.根尖分生区发育成成熟区 D.未受精一的定卵要发脱育离成母植体株
9.植物细胞表现出全能性的必要条件是( ) A.给予适宜的营养和外界条件 B.导入其他植物细胞的基因 C.脱离母体后,给予适宜的营养和外界条件 D.将成熟筛管的细胞核移植到去核的卵细胞内
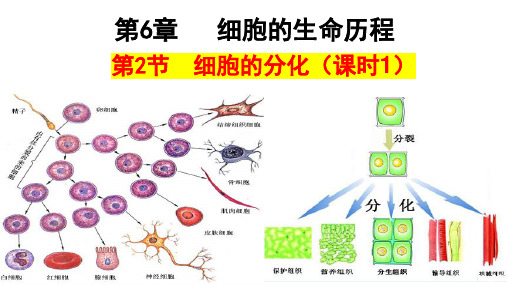
6.2细胞的分化课件高一上学期生物人教版必修1
不会
与动物一样,同一植物体中也存在形态、结构、功能不同的细胞 同一植物体的这些细胞也都来自一群彼此相似的早期胚细胞
细胞 分裂
细胞 分化
受精卵
早期胚胎细胞
叶肉细胞(含叶绿体) 光合作用 表皮细胞(角质层) 保护功能 贮藏细胞(有大液泡) 储存营养
1.实例:
细胞分化
2.概念: 在 个体发育 中,由_一__个__或__一__种__细__胞__增殖产生的后代,在_形__态__、__结__构_ 和_生__理__功__能__上发生_稳__定__性__差__异__的过程,叫做细胞分化
持久性
一般来说,分化的细胞将一直保持分化后的状态,不能恢复成早期
胚胎细胞,也不能转变成其他组织细胞。 稳定性(不可逆性)
多细胞生物体内都存在细胞分化的生命现象。
普遍性
就一个个体而言,细胞分化形成的各种细胞具有完全相同的遗传信息。
遗传物质不变性
1.为什么一个个体的各种细胞具有完全相同的遗传信息?
一个个体的各种细胞均由同一个受精卵有丝分裂形成
小块皮肤嫩肉
无菌培养基 中离体培养
成熟的男人
胎状体发育
到目前为止,人们还没有成功地将单个已分化的动 物体细胞培养成新的个体
思考·讨论
阅读资料2,完成思考1—3(讨论3+P121旁栏思考):
思考1:从资料2中可以得出什么结论? 已分化的动物体细胞的细胞核具有全能性
思考2:为什么已分化的动物体细胞的细胞 核具有全能性? 已分化的动物细胞体细胞的细胞核中含 有该动物几乎全部的遗传物质
讨论1:从资料1中可以得出什么结论? 高度分化的植物细胞仍然具有发育成完整植株的能力
讨论2:如果将胡萝卜韧皮部细胞换成其他高度分化的植物细胞, 在适宜条件下,这些细胞也能形成新的植株吗? 能
高三生物一轮复习动物细胞工程(4)

领域发挥大作用。
5.诱导多能干细胞(iPS细胞) (1)胚胎干细胞的局限性: 胚胎干细胞必须从_胚__胎__中获取 ,涉及_伦_理__问__题__,因而限制了在医学上的应用。
(2)诱导多能干细胞概念: 通过体外诱导小鼠成纤维细胞,获得了类似胚胎干细胞的一种细胞,称为诱导
细胞株 从原代培养细胞群中筛选出进行传代培养的细胞,能传到40-50代,其遗传物质
没有发生变化。
细胞系 传到40-50代后有部分细胞遗传物质发生了变化,具有癌细胞的特点,失去
接触抑制,可能在培养条件下无限制的传代下去。
区别: 细胞系的遗传物质改变,具有癌细胞的特点,失去接触抑制, 容易传代培养。
三、干细胞培养及其应用
动物细胞培养
动物细胞 动物细胞融合技术与单克隆抗体
工程
动物细胞核移植技术
克隆动物
一、动物细胞培养
1.动物细胞培养概念:指从动物体中取出相关的组织,将它分散成单个细胞,然后在适 宜的培养条件下,让这些细胞生长和增殖的技术。2.动物细胞培
养的原理:细胞增殖(有丝分裂)。3.动物细胞培养的地位:动物细胞工程的基础。
上皮细胞
肌细胞
神经细胞
4.干细胞的应用 (1)应用原理:有着自我更新能力及分化潜能的干细胞,与组织、 器官的发育、再生和修复等密切相关。 (2)应用实例: 造血干细胞:治疗白血病及一些恶性肿瘤放疗或化疗后引起的 造血系统、免疫系统功能障碍等疾病
神经干细胞:治疗神经组织损伤和神经系统退行性疾病(如帕 金森病、阿尔茨海默病等)
多能干细胞,简称iPS细胞。
诱导方法:①将特定基因或特定蛋白导入细胞(成纤维细胞以及已分化的T细胞、 B细胞均可);
6.1 细胞的增殖(第1课时)-2024-2025学年高一生物爱上课堂优质课件(人教版必修1)

后期
染色体:4条→8条 染色单体:8条→0条 DNA:8条
三、有丝分裂(高等植物细胞为例) 分裂期(末期) 末:两消两现重建壁
染色体变成染色质;纺 锤体消失,核膜、核仁重现 在赤道板位置出现细胞板 细胞板扩展成细胞壁,形 成两个子细胞
后期
染色体:8条→4条 染色单体:0条
细 胞 板
末期
DNA:8条-4条
三、有丝分裂(高等植物细胞为例) 复习提问:下图所示序号的结构名称是什么?
1 核膜 2 核仁 3 染色质
4细胞壁
三、有丝分裂(高等植物细胞为例)
染色质(体)
着丝点
高度螺旋
染色质
复制
染色体
1.着丝点可能在中间, 可能靠 近某一端。 2.一个着丝点,就意味着一条染 色体。
染色体
姐妹染色单体
三、有丝分裂(高等植物细胞为例)
三、有丝分裂(高等植物细胞为例) 分裂期(前期) 前:膜仁消失显两体
核膜
核仁 着丝点 染色质
①出现染色体 ②出现纺锤体
③核膜核仁消失
间期(末)
染色体: 4条
染色单体:8条
纺锤丝 染色体 着丝点
前期
DNA:8条
三、有丝分裂(高等植物细胞为例) 分裂期(中期) 中:形定数晰赤道齐 辨认的最好时期
①纺锤丝牵引染色体运动,使每条染 色体的着丝粒整齐地排列在赤道板上
4.出现细胞板,形成细胞壁
下图是高等植物细胞增殖各时期的示意图,其中排列顺序是紊乱的,据图回答
问题:
1
4
2
3一二来自三四五
六
(1)图中所标号码所表示的为:
1.______核__仁______,2.___纺___锤__体__(___丝__)____,3.__染__色___体_____,4.__细___胞__板__(___壁__)_____.
植物基因与茎尖分生组织的形成与维持

• WUS通过和这些转录共阻遏物结合共同抑 制参与干细胞分化过程中的基因表达,因 而干细胞能保持其多能性的特征。
• 此外,WUS在转录过程中还受SYD的调控, SYD是一种染色质重构ATP酶,这类酶能够 促进基因的顺式调控区域和转录因子的结 合。
• 因此,SYD对WUS在干细胞组织中心的表 达起正调控作用,从而引起WUS的高水平 表达。这种作用是维持茎端分生组织中干 细胞特征所必需的。
3. CLV基因
• 拟南芥CLV丧失功能的突变株clv1、 clv2 clv3
在枝条和花的分生组织的干细胞区积累未分化的 干细胞,导致SAM庞大,其中细胞的分裂速度并没 有发生变化;
• 因此,从表型性状推测,在野生型植株中CLV的 功能是限制分生区干细胞增殖,CLV基因的作用 与STM和WUS相反,对SAM的中央区域细胞的未 分化状态这一特征起负调控作用。
• CLV3 is expressed in L1 and L2 layer.
• CLV3 and CLV1 temporal expression patterns are very similar.
• CLV3 is a soluble protein
• contains a signal peptide to direct the protein into the secretory pathway.
顶面观 侧面观
高一生物必修一第六章知识点总结

高一生物必修一第六章知识点总结生物学作为自然科学中的一门重要学科,在高中阶段占据了非常重要的地位。
高一生物必修一第六章主要讲解了细胞的结构和功能,为我们理解生物体的组成和基本生命活动提供了基础知识。
本文将对该章节进行总结,帮助学生们更好地掌握相关知识点。
一、细胞的结构细胞是生物体的基本结构和功能单位,根据细胞的形态、功能以及缺核与否的区别,可将其分为原核细胞和真核细胞。
原核细胞不含细胞核,细菌是其代表;真核细胞则含有细胞核,包括动植物细胞。
1.细胞膜:细胞膜是细胞的外包膜,由磷脂双分子层组成,具有选择性通透性,可控制物质进出细胞。
2.细胞质:细胞质是细胞膜与细胞核之间的物质,包含细胞器和细胞质基质。
3.细胞核:细胞核是细胞的控制中心,包含着遗传信息和调控生物体基因表达的指令。
二、细胞的功能细胞作为生物体的基本结构单位,具有许多重要的功能,主要包括代谢、繁殖、生长和调控等。
1.代谢:细胞通过代谢反应将营养物质转化为能量,维持生物体的正常生命活动。
2.繁殖:细胞通过细胞分裂实现繁殖,将遗传信息传递给下一代。
3.生长和发育:细胞通过物质的吸收和合成实现生长和发育,保持细胞和生物体的正常功能。
4.调控:细胞能够对内外环境进行感知并作出相应的调节反应,以维持内稳态。
三、细胞器的功能细胞器是细胞内的亚细胞结构,各有不同的功能。
主要的细胞器包括溶酶体、高尔基体、内质网、核糖体和线粒体等。
1.溶酶体:溶酶体参与细胞对外界物质的吞噬和降解。
2.高尔基体:高尔基体参与蛋白质的合成、包装和分泌。
3.内质网:内质网用于蛋白质的合成、修饰和折叠。
4.核糖体:核糖体是蛋白质合成的场所。
5.线粒体:线粒体是细胞内的能量制造器,参与细胞的呼吸作用。
四、细胞的分化和特异性细胞分化是指在发育过程中细胞通过表达不同基因而产生差异,从而形成不同种类和功能的细胞。
细胞的特异性使得人体的各个组织和器官能够担负不同的任务。
1.干细胞:干细胞是具有自我更新和分化潜能的细胞,具有重要的医学应用前景。
植物生长因子与干细胞功能关系的研究

植物生长因子与干细胞功能关系的研究植物是能够自我更新和再生的生命体,这一过程与干细胞的功能密切相关。
干细胞是一种特殊的细胞类型,能够自我复制并产生多种类型的细胞。
然而,植物干细胞比动物干细胞更加丰富和多样化,其中一些细胞甚至可以扮演多种角色。
植物营养和激素调节是干细胞发育和功能维持的关键,而植物生长因子则在这一过程中发挥着重要作用。
植物中的生长因子包括激素、小分子信号和环境因素等。
其中,植物激素是一组复杂的天然化合物,可以影响植物的各种生长和发育过程。
激素作为生长调节因子,广泛存在于植物体内,并通过信号传导途径发挥作用。
在细胞水平上,植物激素通过受体介导的信号传导通路作用于基因表达和蛋白质合成等分子水平上,从而影响植物生长和干细胞发育功能。
植物激素对干细胞发育和功能维持的影响是复杂和多样的。
例如,植物激素生长素是最初被发现的植物激素之一,它在植物生长中起着很重要的作用。
生长素对植物干细胞的分化和扩增具有正向调节作用,但另一方面,生长素还可以对植物干细胞造成抑制作用和发育限制。
其他几种植物激素如赤霉素、细胞分裂素和脱落酸等也对植物干细胞功能产生着影响,但具体的作用机制仍需要进一步研究。
近年来,植物生长因子的研究逐渐扩展到了干细胞分化和调控等多个方面。
许多研究表明,植物生长因子可以通过调节植物干细胞命运和抗逆能力等机制,促进植物生长发育和抵抗环境胁迫等生物学过程。
例如,植物干细胞中某些内源性因子可以影响干细胞分化程度和细胞命运,在复杂的环境中表现出不同的表型和品质。
同时,一些外源性因素如其他激素、气体信号和胁迫等环境因素也会影响植物干细胞的功能发挥。
总的来说,植物生长因子与干细胞功能之间的关系是非常复杂和多样化的。
通过研究各种生长因子和激素的作用机制,能够帮助我们更深入地理解植物干细胞的发育和功能维持。
同时,这些研究也可以为植物干细胞应用于生产和保护环境等领域提供有价值的指导和指导意义。
未来,我们有望通过深入探究植物生长因子和干细胞功能的相互作用研究,为植物生长发育和未来生产提供更加有效和可行的方案和策略。
6.2细胞的分化课件-高一上学期生物人教版(2019)必修1

巩固练习
2.科学家用甲母羊乳腺细胞的细胞核与乙母羊卵细胞的细胞质重组 成一个细胞,成功培养了“多莉”羊。下列关于此实验的说法错误的
是( D)
A.高度分化的细胞仍然保持全能性 B.重组细胞的细胞核内含有保持物种遗传性所需要的全部遗传物质 C.在理论上,可以运用上述方法开拯救濒危物种 D.运用上述方法,科学家已经可以用单个细胞来培养新的动物体
实验证明:已分化的动物体 细胞的细胞核仍具有全能性。
精讲点拨
二、细胞的全能性
1.概念
细胞的全能性是指细胞经 分裂和分化 后,仍具有产生 完整有机体 或分 化成其他各种细胞 的潜能和特性。
注:1. 起点:已分裂、分化的细胞; 终点:产生完整有机体或分化成其他各种细胞
2.未分化的细胞也具有全能性。如受精卵、动物和人的早期胚胎细胞、植物体分 生组织细胞。 且未分化的细胞全能性 > 已分化细胞的全能性
2.如果将胡萝卜韧皮部细胞换成其他已高度 分化的植物细胞,在适宜的条件下,这些细 胞也能形成新的植株吗?
能
合作解疑
二、细胞的全能性
条件: ①离体 ②植物激素 ③一定的营养条件 ④适宜的环境(pH、温度等) ⑤无菌环境
合作解疑
二、细胞的全能性
讨论 1.将肠上皮细胞单独培养能获得新的个体吗?
不能 2.与资料1中的实验相比,你能从资料2中的 实验得出什么结论?
精讲点拨
二、细胞的全能性
2.原因 细胞内含有本生物物种全套的遗传信息。
3.细胞全能性大小的比较
(1)胚胎干细胞> 造血干细胞 (2)受精卵 > 生殖细胞(精子、卵细胞)>体细胞 (3)植物细胞 > 动物细胞
细胞全能性大小主要受细胞分化程度影响。细胞分化程度越高 ,全能性越_低___; 分化程度越 低 ,全能性越_高__。
组培形成层分生组织细胞作为一种植物天然产物的来源

知识点分生组织(Meristerm): 植物体内能连续或周期性地进行细胞分裂的组织。
由这种组织衍生的细胞,经生长和分化形成其他各种组织,而其本身始终保持着分裂能力。
细胞接触紫杉醇后会在细胞内积累大量的微管,这些微管的积累干扰了细胞的各种功能,特别是使细胞分裂停止于有丝分裂期,阻断了细胞的正常分裂。
红豆杉醇:别名紫杉醇,紫素,特素,紫烷素,路泰,泰素。
非常重要的一个问题是,天然产物中化学成分的不同是由于植物的不同造成的。
原则上讲,植物细胞培养为天然产物的生产提供了一个很具有吸引力的选择。
但是,因为组培经常会需要在工业化范围的脱分化,而形成脱分化的细胞(DDCs)在商业上不具有操作可行性(不经济or容易发生基因突变)。
为了避免这一脱分化过程,我们分离、培养得到了天然的未分化的形成层分生组织细胞(CMCs)。
将各种深度测序技术结合起来,明确了与干细胞一致的标记基因和转录程序。
这种想法被近一步的CMCs的形态、他们对γ-射线、类放射药物的敏感程度及它们具有高分化能力支持(由于形成层分生组织的这些优点,使得将CMCs作为一种天然产物的想法得到进一步的支持)。
本文主要研究通过东北红豆杉CMCs 悬浮培养,产生抗癌药物――紫杉醇。
此过程避开具有商业障碍的脱分化这一步,这些细胞能够为可持续生产品种繁多的植物天然产物提供一种成本效益高,环境友好的平台。
只有植物干细胞,也就是生长在茎尖、根尖及维管内部的分生组织细胞,才能够进一步分化或再生出干细胞。
由于这些分生组织细胞在理论上能够无限次的分裂,因此它们被认为是永远不会死亡的一类细胞。
所以,自从19世纪40年代植物组培开始,细胞悬浮培养一般都是先经历一个脱分化过程,然后对脱分化的细胞进行悬浮培养。
当前的一些研究表明,这个过程可能不是必须的。
不考虑这个过程中会涉及什么机理性问题,这个过程的结果是在一个给定的器官中发生有丝分裂,产生多细胞混合的扩繁的细胞群的过程。
由此得到的细胞进行悬浮培养获得细胞产物,可能由于在脱分化过程中会产生有害的或是后成说的基因突变,而致使产生天然产物量小及产品性质很不一致等问题。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
摘要:植物干细胞位于茎尖分生组织区和根尖分生组织区,是植物胚后发育中新的器官产生的源泉。
近几年,在干细胞及其周围组织区发现了一些与干细胞稳态维持有关的基因,这些基因产物与外源性信号(如生长素)一起组成复杂的调控网络控制植物的生长和发育。
表观遗传修饰作为控制基因表达的一种方式也对植物干细胞有重要的影响。
该文介绍近几年植物干细胞分化调控的最新进展。
关键词:植物干细胞;基因调控;生长素;表观遗传中图分类号:Q74动物干细胞因其在疾病治疗、组织修复和动物克隆等方面的广泛应用一直是研究人员和公众关注的焦点[1],相比之下,植物干细胞研究却门庭冷落。
由于植物细胞具有全能性,植物干细胞的概念也一直存在争议。
近几年的研究表明:在植物的茎尖分生组织(shoot apical meristem,.. SAM)和根尖分生组织(rootapical meristem,.. RAM)中,存在一群特殊的细胞,它们具有自我更新能力又能产生具有持续分裂能力的子细胞。
这些特殊细胞是植物根、茎、叶和花等器官发生的源泉,因此被认为是植物干细胞[2,3]。
与动物干细胞一样,植物干细胞的维持同样受到内源性信号和外源性信号的调控。
1. 植物干细胞的组织学特征模式植物拟南芥的SAM 是一个半球状的穹型结构,由多个功能结构域组成[图1(A)]。
干细胞位于分生组织的顶端中心区域,这个区域的细胞分裂不活跃。
中心区域中干细胞分裂后产生两部分细胞,一部分仍然保留在中心区域的叫干细胞后裔(progeny ofstem cells),.. 保持多潜能性,始终留守在原来位置,继承干细胞的衣钵;分裂出来的另一部分叫子细胞(daughter cells),.. 随着干细胞的分裂逐渐脱离中心区域到分生组织周边区域(peripheral zone,PZ),.. 在周边它们快速分裂并进行分化,融入分生组织两侧的器官原基中[4]。
由此可见,SAM是不断改变的动态结构,干细胞群的维持依赖于周围组织细胞提供的各种外源性和内源性信号分子。
在拟南芥的根尖分生组织中心,也有一群分裂缓慢的细胞,称为静止中心(quiescent center,QC)[图1(B)]。
在胚胎发生中,QC的建立不是来自胚体,而是来自胚柄最上部的细胞。
QC 细胞通过不对称分裂产生子细胞,子细胞或者保留QC细胞功能,或者取代邻近细胞。
实质上直接从QC 衍生出来的细胞就是干细胞,它们能够产生根部特定的组织类型。
研究拟南芥根的发育表明,RAM 中干细胞的维持受位置信息的影响[5]。
当QC被切去后,邻近的细胞就发育成一个新的有功能的QC。
QC释放信号分子以维持根尖干细胞局部微环境。
2. 植物干细胞调控的分子机制2.1 WUS-CLV反馈调节环控制茎尖干细胞稳态1996 年Laux等[6]利用诱变技术发现,WUSCHEL(WUS)基因的编码产物是维持干细胞数量的内源性信号分子。
WUS mRNA在干细胞组织中心区域表达,wus突变体花的数量明显减少且提前脱落,由此可见,在野生型植株中WUS 基因的功能是通过维持中心干细胞库以促进茎和花分生组织的活性。
WUS 基因的异位表达诱导异位干细胞的形成,表明WUS表达区域必须受到严格的调控以维持干细胞的正确位置和数量。
与WUS基因功能相反,CLAVATA(CLV)基因突变则产生过多的SAM细胞,形成膨大的分生组织,茎尖呈环型而不是尖型[7]。
在突变植株的一生中,由于产生过多的干细胞,其结果是枝条粗大,产生较多的花分生组织。
因此,从表型性状推测,在野生型植株中CLV的功能是限制分生区干细胞增殖。
研究表明,CLV表达区域位于WUS之上[8]。
CLV3是干细胞分泌的小分子多肽,作为配体与CLV1-CLV2(在干细胞之下表达)组成的受体复合体相互作用。
激活CLV 复合体启动下游信号事件,导致WUS 表达区域受到限制。
在野生型拟南芥中,干细胞稳态受到正负信号的反馈调节。
CLV3是一个负调控信号,控制WUS在分生组织中心很窄范围内表达。
在CLV3 突变体中,WUS表达区域向两侧及向上扩展。
转基因CLV3过表达的植株类似WUS 突变体,说明WUS基因的活性受到CLV信号通路的下行调节。
WUS既是CLV负控信号的重要靶基因,又是促进干细胞数量的正调控基因。
然而,诱导WUS 基因异位表达却能促进CLV3 基因转录,说明WUS 和CLV 组成了一个反馈调节环[4,8]。
当干细胞后裔脱离干细胞区域,成为器官原基的一部分后,干细胞数量减少引起CLV3信号水平降低。
负控信号水平降低引起WUS表达区域扩展,通过这个正调控途径使干细胞数量增加。
干细胞数量增加到一定程度时反馈激活CLV3信号表达,限制WUS表达区域进一步扩大,保持干细胞数量的恒定。
2.2 SHR-SCR 信号通路对根尖干细胞的调控作用SCARECROW(SCR)和SHORTROOT(SHR)属于GRAS 家族的转录因子,对根尖分生区的维持起关键作用[9]。
SCR在QC、内皮层/皮层初始区和已分化的内皮层表达,其功能缺失导致初始区及QC干细胞的连续性丧失。
SHR在根尖中柱组织表达,其蛋白质转运到邻近细胞层(包括QC)。
SHR功能缺失导致QC的结构不规则,缺乏QC特异性标志分子和根部停止生长等表型。
在内皮层/皮层初始区,SHR激活SCR,促进干细胞的不对称分裂。
SHR 和SCR 都是维持QC 功能必需的,因为在shr突变体中,SCR在QC区域表达并不能挽救QC 缺失[10]。
最近研究发现,PLETHORA (PLT1/2)基因对胚胎发生中干细胞群的形成和胚后发育中干细胞数量的维持亦至关重要[11]。
PLT基因在QC和周围干细胞区域表达,与SCR分布区域重叠,说明它们共同为干细胞微环境提供信号。
PLT基因异位表达诱导茎尖分生区形成异位的根尖干细胞微环境,因为SHR和SCR基因在此茎尖组织表达。
上述研究说明,根尖干细胞微环境的形成依靠SHR 和SCR 提供信号分子。
2.3 生长素对植物干细胞的调控作用生长素(auxin)对植物的生长、发育具有重要的调控作用,是熟知的影响干细胞微环境的外源性信号[12]。
生长素在PIN蛋白(生长素运输辅助因子家族蛋白)协助下长距离极性运输,在这个过程中建立生长素梯度效应。
在胚后的根尖分生区,最高的生长素梯度位于QC 下的远端干细胞。
生长素最高梯度效应参与根尖干细胞微环境的位置特化,干扰生长素效应或者极性运输诱导分生区组织结构紊乱,形成异位的QC和干细胞[13]。
Aida等[11]报道,PLT基因表达受生长素效应调节,但PLT基因突变不影响生长素在根尖分布和初级生长素效应,说明PLT在生长素下游发挥作用。
Pin基因突变影响QC 在胚后根尖中的位置及PLT基因表达模式,进一步说明生长素对干细胞区域化的影响是通过PLT基因介导的[13]。
此外,3个PIN基因的表达受到PLT基因调节。
上述研究表明,生长素启动胚根发育,调节PLT 基因表达使干细胞位置特化;PLT 基因又反馈调节PIN基因表达,通过稳定生长素最高梯度效应进一步使根尖干细胞微环境区域化[9]。
此外,生长素的另外一个重要作用是控制茎尖干细胞分化、促进器官原基形成[14]。
生长素向顶端分生组织运输,控制该区域的细胞参与两侧器官发生。
器官原基总是在分生组织的高生长素浓度区域形成,一旦形成器官原基,它像海绵一样吸取周围细胞中的生长素。
新的原基器官的出现除效应于高生长素浓度外,还是KNOX 基因下调表达的结果。
在玉米中,通过抑制生长素运输实验表明:生长素积累抑制Ⅰ型KNOX基因,如SHOOT MERISTEMLESS(STM)基因,在PZ区域表达[15]; 反之,亦有研究表明,KNOX基因的表达调节生长素的极性运输。
生长素和KNOX基因是否通过相互作用建立一个自动调节环,这个问题尚需继续探索。
2.4 表观遗传机制对植物干细胞的调节与动物干细胞一样,植物干细胞也受到染色质活性变化的调节。
研究表明,染色质装配因子FAS1 和FAS2 限制WUS 和SCR 基因的活性,且在FAS1 或FAS2 突变植株中观察到WUS 基因异位表达现象,说明WUS 是FAS1和FAS2的靶基因[16]。
BRU1,一个稳定复制后染色质结构的蛋白质,与WUS基因的正调控有关[5]。
最近研究发现,WUS 是染色质重塑因子SYD 的直接靶基因。
SYD 属于ATP 酶家族蛋白,使DNA 模板易于形成转录复合体从而促进转录[17]。
Syd突变植株SAM提前进入终端分化与WUS表达水平降低的表型一致。
染色质免疫沉淀证明SYD 结合到WUS 启动子区域,说明SYD是WUS基因的上游调控因子[17]。
在动物中,PcG蛋白家族通过组蛋白甲基化和组蛋白去乙酰基化关闭与干细胞分化有关的基因,使干细胞保持未分化状态[3]。
在植物中,研究表明干细胞全能性丧失与PcG介导的基因沉默有关。
Katz等[18]研究表明,FIE和CLF蛋白(分别与PcG家族蛋白Eed和Ezh2同源)抑制STM基因表达。
STM 基因的主要功能是抑制分生组织中细胞的分化,保证分生组织内细胞的扩增,从而能有足够数量的细胞成为器官原基。
上述研究表明,染色质修饰对植物干细胞和动物干细胞的调控具有相似的分子机制。
在动物中,AGO(ARGONAUTE)蛋白与干细胞自我更新有关[3]。
在拟南芥胚胎发生中,AGO家族蛋白ZLL对建立稳定的茎尖干细胞群非常关键。
在zll突变体中,种子萌发后不久茎尖分生区就进行终端分化,只产生一个或少数几个花器官[19]。
ZLL异位表达诱导细胞过度分裂和形成异位分生组织,说明该蛋白质在建立干细胞群中发挥作用。
研究已证实,AGO家族蛋白的功能是通过siRNA 和miRNA 控制mRNA 稳定性或者抑制mRNA 翻译[20]。
除控制mRNA 翻译外,在裂殖酵母和拟南芥中还发现,AGO 蛋白参与miRNA 介导的组蛋白甲基化过程[3]。
尽管miRNA和染色质失活之间机制尚待阐明,但众多的研究表明它们对干细胞的自我更新和分化有重要影响。
3. 总结在过去几年里,植物干细胞的研究取得了很大进展,研究人员发现了一些与干细胞数量维持和分化有关的基因,这些基因与外源性信号一起组成复杂的网络控制植物的生长和分化。
尤其重要的是,人们逐渐发现植物干细胞和动物干细胞尽管形态和功能各异,干细胞稳态的维持却具有相似分子机制。
调控染色质活性变化的PcG 蛋白家族和通过miRNA 发挥作用的AGO蛋白家族在动、植物干细胞共同存在,这一现象进一步激发研究人员的兴趣。
因此,这些领域的研究必将促进人们对干细胞生物学特性的了解。
研究幹細胞的應用叫再生醫學,再生醫學是未來人類持久保持健康長壽的支柱。
甚麼是幹細胞:幹細胞是一群尚未完全分化的原始母細胞,它具有自我更新及分化的潛能。