滇一型杂交粳稻恢复基因的分子鉴定研究_谭亚玲
来源不同的恢复系对滇Ⅰ型不育系恢复力的探讨

来源不同的恢复系对滇Ⅰ型不育系恢复力的探讨金寿林;杨国仙;王石华;洪汝科;刘雨;谭学林【摘要】通过调查云南农业大学自育的恢复系南34和分别从上海、辽宁引进的恢复系沪R 378、C418与9个滇Ⅰ型不育系杂交共27个杂种F1的育性,以比较三个恢复系对滇Ⅰ型不育系的恢复力.结果表明:恢复系南34对滇Ⅰ型不育系恢复力强.可在生产上广泛应用;沪R378和C418对滇Ⅰ型不育系具有一定的恢复能力,在生产上可有选择的使用,组配的杂种F1应进行生态适应性和抗性鉴定后,才能应用于生产.【期刊名称】《种子》【年(卷),期】2009(028)008【总页数】3页(P79-81)【关键词】恢复系;南34;沪R378;C418;滇Ⅰ型不育系;组合;恢复力【作者】金寿林;杨国仙;王石华;洪汝科;刘雨;谭学林【作者单位】云南农业大学稻作研究所,昆明,650201;云南省大理州南涧县农业技术推广中心,云南,大理,675700;云南农业大学稻作研究所,昆明,650201;云南农业大学稻作研究所,昆明,650201;云南农业大学稻作研究所,昆明,650201;云南农业大学稻作研究所,昆明,650201【正文语种】中文【中图分类】S511滇Ⅰ型不育系,是云南农业大学李铮友教授1969年育成的我国第一个水稻雄性不育系(滇Ⅰ型红帽缨不育系),经过多年,用多个品种转育而成的一系列不育系[1],是目前生产上培育滇型杂交粳稻利用最多的不育系。
现有的绝大多数的粳稻常规品种对滇Ⅰ型不育系都是保持的,可将它们转育成不育系[2];而滇Ⅰ型恢复系或恢复材料,没有一个是直接从现在的品种中测交筛选出来,它们都是天然杂交或人工杂交得来[3]。
为了选育杂种优势强的杂交粳稻,首先必须将籼粳亚种的品种或材料进行杂交,以选育出籼粳交的恢复系[4]。
可见恢复系选育程序复杂,选育年限长,工作量大,导致恢复系单一,是制约杂交粳稻育种的关键因素之一。
在籼稻区,由于地理气候的相似性,外省选育的杂交籼稻在云南省籼稻区得到广泛的种植,显著地提高了云南省的籼稻产量。
优质杂交粳稻“滇优34”及其高产栽培技术

以 匕。
剑叶长 2— 1( 93 3 m,剑 叶 宽 1 ~ . c . 1 m。单 株 有效 穗 数 5 6 91 ~ 0穗 ,穗 长 2 ~ 3C I 0 2 1,穗 总 粒数 10 10粒 ,实 T 4~9
科 院育成 的优质 长粒 型粳稻品种滇粳优 1 号为资源 , 采 用 云南农 业 大学 滇型 杂 交水 稻育 种 技术 ,历 经 9年 时间选 育而成 的第 1 长粒 型杂 交粳 稻组 合 ,于 20 个 07 年通 过 云南 省 农 作 物 品 种 审 定 。 在 嵩 明 、大 理 、龙 陵 、鹤 庆等 地 种 植 ,表现 出抗 寒 性 强 、抗 病性 较 好 、 品质优 等特点 ,增 产 1%以上 。 0
滇优 3 4属 中早熟 组 合 ,在海 拔 16 0 20 01 的 0 ~ 0 1 1 中高海 拔 粳 稻 区种 植 ,全 生 育期 10 10d 7 ~9 。生 育期 随海 拔高 度 、栽 培节 令 、栽插 密 度 、肥 水管 理 等 条件 不 同而略有 变化 。 1 主 要农艺性 状 . 2 滇优 3 4株 形 紧 凑 ,剑 叶挺 直 ,株 高 9 ~ 0 m, 6 15c
云 南 农 业 科 技
2 1 年 第 3期 n n y
优质杂交 粳稻 “ 滇优 4,及其高 产栽培技市 3,
郭 艾 , 陶光喜 ,谭 亚玲
( 南农 业 大学稻 作研 究所 ,云 南 昆明 6 0 0 ) 云 521
滇优 3 4是 云 南农 业 大 学 稻 作 研 究 所 以 云南 省 农
滇一型杂交粳稻亲本数量性状遗传差异研究

Su y G nt Df rne o Jpnc Rc P rns Da T p td o ee c f ecs aoi i aet o i y e n i ie f a e f n
G O a- ag, A G iu' P Z g n2 A S o hn' W N S - a, U i g, h c hh ia - J So- ' W N n hn ' T N el' I hui , E J - eg, A X - N l n ic a u i n
@sh . m ou c o
万方数据
16 4
云南农业大学学报
第2 卷 3
பைடு நூலகம்
4 麻、 辣椒、烤烟等作物上进行了广泛研究「, 2 ] 南农业大学稻作研究所近年来选育出的3 份恢复 一。
对于滇型杂交粳稻的恢复系 、 保持系的遗传差异,
谭学林「在 1 8 5 9 年曾基于数量性状通过模糊数 〕 8
高绍 ,王石华, 长‘ ,普自 2 金寿林, 祥, ,文建成, 谭学 ’ , 林,‘
(.云南农业大学稻作研究所, 1 云南 昆明 600; 521 2 .红河州种子管理站, 云南 蒙自61 0 61 ) 0 摘要: 为了解滇型杂交粳稻亲本的遗传差异, 对云南农业大学稻作所多年来选育出的3 份恢复系和2 份保持 4 9 系的 1个农艺性状进行主成分及聚类分析。结果表明: 0 滇型杂交粳稻6 份亲本材料的 1 个农艺性状可简化 3 0 为6 个彼此互不相关的综合指标 ( 主成分) :即单穗产量、 大穗、 穗数、 株形、 穗形、 穗性状因子, 该6 个主 成分的特征根累积贡献率高达9.2 1 %。基于主成分分析在欧氏距离的水平上进行系统聚类,以35 4 . 的距离将 6 个材料划分成 7 3 大类群, 其中 7 第 类群有 4 份材料,占7.9 8 61%。第 1 2 4 5 6 , , 等5 , , 个类群的材料全 部为恢复系, 说明在滇型杂交梗稻的恢复系中具有丰富的遗传多样性。然而, 对于保持系和第 7 类恢复系而 言, 需借助分子手段进行研究以进一步了解其遗传差异。 关键词: 杂交粳稻亲本;主成分分析;聚类分析; 杂种优势 中图分类号: 51 3 S 1. 2 文献标识码: 0 A 文章编号:04 0 (08 0 - 5 1 - X 0 ) 04 - 0 3 9 2 2 1 0 5
2004年5月应结题验收的SRT计划项目一览
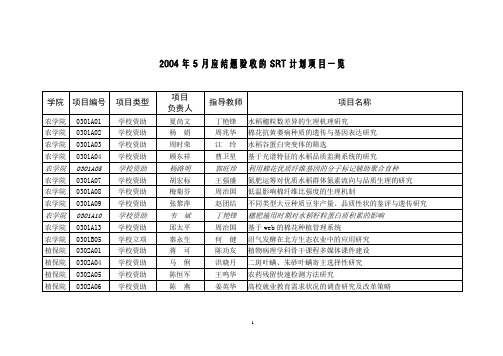
0310A01
学校资助
张明军
陈超
卫岗与光明在南京乳品市场上的比较分析
经贸院
0310A02
学校资助
刘平
应瑞瑶
盗版软件市场现状的实证分析
经贸院
0310A03
学校资助
寇成
刘荣茂
江苏城镇居民储蓄行为研究
经贸院
0310A04
学校资助
何荔薇
胡浩
南京市观光农业的现状分析及发展建议
经贸院
0310B02
学校立项
资环院
0307A01
学校资助
施春健
徐阳春
不同商品有机肥对土壤供氮特性和作物生长的影响
资环院
0307A02
学校资助
沈倩宇
宗良纲
土壤中重金属污染作物毒害图谱的建立
资环院
0307A03
学校资助
张攀伟
陈巍
增铵对蔬菜叶绿体及其内容物的影响
资环院
0307A04
学校资助
赵慧渊
刘德辉
氨基酸螯合中微量元素肥料的肥效研究
信息院
0313B01
学校立项
程冲
黄水清
基于Java平台的网络信息检索定题服务系统
信息院
0313B03
学校立项
吴亮
黄芬
基于友好人机交互的数据库导入与导出
外语院
0314A02
学校资助
麦添花
金锦珠
日本外商对江苏省投资的历史与现状分析
外语院
0314A04
学校资助
王晓妹
成春有
日语汉字吴音与吴方言的比较研究
外语院0314B01学来自立项邵小丽游衣明
日语量词研究现状调查
海拔及细胞质背景对水稻测交群体遗传分化的影响

组合 F产生 的群体 中 , 群体 的遗传 多样性 随产生群体海拔( 0m海拔 除外) 4 0 的升高而降低 。利用 N i 遗传距 离评价各群体 eS 间的遗传分化 , 正反交组合 F 产生测交群体间的遗传分化均高于 同一组合在不 同海拔下产生 的测交群体之 间的遗传 分化 ,
筛 选 出的杂 交组 合 的F群 体 中检 测 发 现一 些 S R 2 S 位 点出现不同程度 的偏分离【,推测这些偏分离可能 9 J 主要 是 由于海 拔差 异导 致 的温 度差 异选 择所 致 。但
这些 差 异是 温度 差异 对雌 配子 ,雄 配子 ,还是 对孢
子体的选择所致还缺乏实验资料证明 ,因此对于环 境条件对水稻选择所致 的遗传变异还需作进一步 的研 究 。 为 明确海 拔及 细胞 质 背景 对水 稻 正反交 F在 不 一 同产生的测交群体遗传分化的影响, 本研究利用适 于 高 海 拔 冷 凉 环 境 的粳 稻 地 方 品种 和 适 于温 和 气 候具 籼稻 细胞 质 背景 的 品系杂交 ,在海 拔 高低相 差 达 1 0 0m的4 海 拔点 的种植 正反 交F并与 育性 稳 8 个 1 定 的滇 一 型 细 胞 质 不 育 系 杂 交 产 生 的8 测 交 群 个 体 ,用 s R 记检 测 发现 海拔 因素 和细胞 质 背景 都 s标 对测交群体的遗传分化产生了影响。
同一组合 在 4 个海 拔产生的测交群体间的遗传分化总体趋势是 随海拔差异 的增 加而增大。 研究证 实了环境温度差异及细胞质 背景 导致 F 产生的雄 配子基因型发生选择 ,导致雄配子基 因型 的遗传 比例偏离孟德 尔规律 ,影 响其后代群 体的遗传多样性 和遗传分化 。
关键词 : 水稻测交群体 ; 海拔 ;细胞 质 ;遗传多样性 ;遗传分化
SSR标记及在水稻遗传育种中的应用

SSR标记及在水稻遗传育种中的应用张桂莲,陈立云,张顺堂,雷东阳(湖南农业大学水稻科学研究所,长沙,410128)摘 要:SSR标记是在PCR基础上发展起来的新型分子标记,已开始应用于基因组研究的各个领域。
概述了SSR类型、特点、原理,总结了其在水稻遗传育种方面的应用,并讨论了其所存在的问题。
关键词:SSR;分子标记;水稻;遗传育种中图分类号:S511.03503 文献标识码:A 文章编号:100125280(2004)0520325204 简单重复序列(Si m p le sequence repeat,SSR)亦称微卫星DNA(m icro satellite DNA),是一类以少数几个核苷酸(一般1~6个)串联重复的DNA序列。
SSR广泛分布于真核生物基因组中。
对SSR的研究始于动物基因组。
1974年Sk inner等在寄居蟹DNA中发现了一类(TA GG)n[1]。
1982年H a m ada等在人心肌肌动蛋白(H a225)的内含子中发现了一个重复25次(T G)序列[2]。
随后在其它生物的研究中也发现了相似的串联重复序列。
由于SSR在不同基因组型间表现高度的多态性,使其很快成为一种新的分子标记[3],在多种作物中广泛应用。
本文拟就SSR标记类型、特点、原理及其在水稻遗传育种中的应用作一简要综述。
1 SSR类型根据SSR核心序列排列方式的不同可将SSR分为完全型(perfect)、不完全型(i m perfect)和复合型(compound)。
完全型SSR是指核心序列以不间断的重复方式首尾相连构成的DNA;不完全型SSR是指在SSR的核心序列之间有3个以下的非重复碱基,但两端的连续重复数大于3;复合型SSR是指2个或2个以上的串联核心序列有3个或3个以上的连续的非重复碱基分隔开,但这种连续性的核心序列重复数不少于5。
W eber 等[4]研究指出,这3种类型SSR在人类基因组中所占的比例差异很大,分别为64%,25%和11%,3种类型内完全型是SSR标记中应用较多的一种类型。
滇型杂交粳稻新组合滇杂35

225 kg /hm , 培育移栽时单 株带 2~ 3 个分蘖 的壮 秧。适龄早栽, 浅插稀植 , 叶龄为 4 . 5~ 6 . 0 叶时移 栽较好。在海拔 1 800 m 左右地区栽插密度为 37 . 5 万 ~ 45 . 0 万穴 / hm , 海拔 1 500 m 左右地区栽插密 度为 30 万 ~ 45 万穴 /hm 。施足底肥, 增施 P、 K 肥, P、 K 肥全部作中层底肥施用。海拔 1 500 m 以下地 区 N 肥作底肥一次清 , 海拔 1 600 m 以上地区 N 肥 的 50 % 作底肥施用, 50 % 插后 7 d 作追肥施用。抽 穗期补施钾肥以提高粒重。总施肥量为纯 N 207 ~ 360 kg /hm , 普通 过磷酸钙 750 kg /hm , 硫酸钾 75 kg /hm 。按常规稻栽培管理技术防治病虫害。
收稿日期 : 2007- 06 - 29 基金项目 : 国家科技支撑 计划 ! 高产优 质多抗 水稻育 种技术 研究及 新品种培育 ∀ ( 2006BAD 01A 01- 7) ; 云南省 ! 十一五 ∀科技 攻关项目 ( 2006NG 05) 作者简介 : 金寿林 ( 1969 - ), 男 , 云南南涧人 , 实验师。电话 : 08715227810; E- m ai: l jsl w hz @ 163. com。
( 云南农业大学 稻作研究所 , 云南 昆明
摘 要 : 滇杂 35 是云南农业大学稻作研究所用育性稳定的滇
樨 , 陈丽娟
650201)
型不育系合系 42- 5A 与优质恢复系南 34配组育 成的三系杂交粳
稻新组合。该组合具有产量较高、 抗稻瘟病、 耐瘠薄、 适应性广等特点 , 于 2006年通过云南省农作物品种审定。 关键词 : 滇型杂交粳稻 ; 滇杂 35 ; 选育 中图分类号 : S334; S338 文献标识码 : B 文章编号 : 1005- 3956 ( 2008) 01 - 0087- 02
特色香紫糯新品种滇香紫1号

特色香紫糯新品种滇香紫1号李 娟1 董陈文华1 郭应忠2 普世皇1 王 翔2黄大军1 金寿林1 洪汝科1 陈丽娟1 谭学林1 文建成1(1云南农业大学稻作研究所,昆明650201;2云南省普洱市墨江县农业技术推广中心,普洱654800)摘要:滇香紫1号是利用广南紫米与鄂香1号进行杂交组配选育的有色稻新品种,由云南农业大学稻作研究所和墨江县农业技术推广中心自主选育而成,于2019年通过云南省农作物品种审定委员会审定(滇审稻2019001号),具有株型好、抗倒伏,花青素含量高,米粒紫黑色,香味浓,食味品质高,综合抗性好,推广应用前景好等特点。
对其选育过程、特征特性及栽培技术措施等进行了介绍。
关键词:有色稻;香味;滇香紫1号;选育;应用有色稻作为一种特殊的水稻种质资源,主要由不同色素沉积在水稻种子的种皮内而形成。
其中紫米因种皮富含大量花青素而呈紫黑色,花青素是一种天然的色素,还具有抗氧化药用功效[1-2]。
紫米中蛋白质、氨基酸、维生素、植物脂肪和铁、锌、磷等微量元素种类均比白米丰富[3],因其丰富的营养和药用价值,紫米在民间也被称之为“药谷”“补血米”“长寿米”[4]。
随着生活水平的提高,人们对稻米的需要越来越多样化,紫米等有色米也越来越受到人们的青睐。
滇香紫1号花青素含量高、米质好、香味浓,深受消费者喜爱,具有较好的发展前景。
1 品种来源利用来源于云南文山地方老品种广南紫米为母本,来源于中国水稻研究所选育的常规籼型香稻品种鄂香1号为父本,2009年在云南农业大学稻作研究所昆明温室进行常规杂交,得到杂种F1;2010年和2011年在云南元江早季和晚季进行系谱法选择,得到F2、F3和F4世代;2012年在云南弥勒市竹园镇种植,并选择出F5世代,种植编号为G5-144,综合农艺性状好并趋于稳定;2013年在云南弥勒市竹园镇扩大种植,该品系群体整齐,米粒黑色,蒸煮食味性好,于当年定名为滇香紫1号。
2015年和2016年在云南优质米生产区域普洱市墨江县、西双版纳州勐海县、红河州元阳县、楚雄州彩云镇不同生态环境下进行品种比较试验评价,该品系农艺性状稳定,群体整齐,株叶型好,株高适中,抗倒性好,田间产量和抗性表现较突出,室内分析外观紫黑色,长粒型,具浓郁香味,米饭外观油亮,口感润滑,食味性好。
云南稻微核心种质籼粳回交后代功能成分含量差异及评价

云南稻微核心种质籼粳回交后代功能成分含量差异及评价杜娟;杨加珍;杨晓梦;杨涛;李霞;普晓英;刘建伟;曾亚文【摘要】[目的]了解云南稻微核心种质籼粳回交后代功能成分含量差异,为有效利用云南稻微核心种质奠定基础.[方法]通过方差分析和显著性检验,对108份云南稻微核心种质及121份籼粳回交后代4种功能成分含量进行测定和比较分析,筛选功能稻米选育的桥梁材料.[结果]微核心种质及其回交后代糙米黄酮、γ-氨基丁酸、抗性淀粉、生物碱含量的变幅和变异系数均较大,微核心种质供体功能成分变异系数从高到低依次是:黄酮>抗性淀粉>γ-氨基丁酸>生物碱;回交后代变异系数从高到低依次是:抗性淀粉>生物碱>黄酮>γ-氨基丁酸.粳稻回交后代具有选育富含黄酮、γ-氨基丁酸等成分材料的优势,而籼稻回交后代具有选育高生物碱含量材料的优势.在籼粳各回交后代中,筛选出糙米总黄酮、γ-氨基丁酸和抗性淀粉含量分别都较高的共10个优良株系,这些株系可以作为功能稻米选育的桥梁材料及功能成分基因定位的近等基因系材料.[结论]云南微核心种质籼粳回交后代糙米的4种功能成分均存在广泛的变异,以微核心种质及滇屯502、合系35培育具有高功能成分近等基因系的方案是可行的.【期刊名称】《西南农业学报》【年(卷),期】2018(031)010【总页数】5页(P1993-1997)【关键词】微核心;籼粳;功能【作者】杜娟;杨加珍;杨晓梦;杨涛;李霞;普晓英;刘建伟;曾亚文【作者单位】云南省农业科学院生物技术与种质资源研究所,云南昆明650223;云南省农业生物技术重点实验室,云南昆明650223;云南省农业科学院生物技术与种质资源研究所,云南昆明650223;云南省农业生物技术重点实验室,云南昆明650223;云南省农业科学院生物技术与种质资源研究所,云南昆明650223;云南省农业生物技术重点实验室,云南昆明650223;云南省农业科学院生物技术与种质资源研究所,云南昆明650223;云南省农业生物技术重点实验室,云南昆明650223;云南省农业科学院生物技术与种质资源研究所,云南昆明650223;云南省农业生物技术重点实验室,云南昆明650223;昆明田康科技有限公司,云南昆明650205;云南农业大学农学与生物技术学院,云南昆明650201;云南省农业科学院生物技术与种质资源研究所,云南昆明650223;云南省农业生物技术重点实验室,云南昆明650223【正文语种】中文【中图分类】S511【研究意义】稻米是全球一半以上人口的主食。
云南地方老品种水稻中恢复基因位点遗传差异分析

云南地方老品种水稻中恢复基因位点遗传差异分析寇姝燕;邹茜;刘慰华;谭亚铃;文建成;金寿林;洪汝科;谭学林;袁平荣【期刊名称】《中国农学通报》【年(卷),期】2012(28)21【摘要】目前生产上使用的水稻恢复系之间的遗传基础过于狭窄,应积极开发水稻新恢复系的选育工作。
云南地理气候条件特殊,地方老品种资源丰富,是筛选新恢复系的理想材料。
以61份云南地方老品种水稻资源为父本,通过测交筛选到6份可以恢复野败型不育系的品种。
再利用5对特异引物,对这6份恢复材料和7份恢复系的恢复基因位点遗传差异进行分析。
结果表明:5对特异引物对6份恢复材料的扩增结果各不一样,而与对照恢复系仅有2.8kb的一条扩增带型一致。
说明云南地方老品种水稻中蕴藏的恢复基因与目前生产上应用的恢复基因的亲缘关系较远。
从云南地方老品种水稻中发掘利用新型恢复基因和资源,是突破水稻恢复基因遗传基础狭窄、筛选新质源恢复系的一种有效策略。
【总页数】6页(P27-32)【关键词】云南地方稻种;测交;育性恢复;遗传差异【作者】寇姝燕;邹茜;刘慰华;谭亚铃;文建成;金寿林;洪汝科;谭学林;袁平荣【作者单位】云南省农业科学院粮食作物研究所;云南农业大学稻作研究所【正文语种】中文【中图分类】S-3【相关文献】1.云南省水稻品种稻瘟病抗性差异与遗传多样性相关研究 [J], 涂敏;王云月;卢宝荣;杨学辉2.云南元阳哈尼梯田水稻地方品种月亮谷的遗传变异分析 [J], 董超;徐福荣;杨文毅;汤翠凤;张恩来;杨雅云;阿新祥;张斐斐;卢光德3.中国西双版纳水稻老品种在育性恢复基因Rf-1位点的遗传多样性 [J], 寇姝燕;谭亚玲;李俊;周英;陈丽娟;谭学林4.云南地方水稻品种遗传多样性分析及其保护意义 [J], 朱明雨;王云月;朱有勇;卢宝荣5.元阳白脚老粳水稻地方品种内遗传异质性及意义 [J], 高东;王云月;何霞红;李成云;朱有勇因版权原因,仅展示原文概要,查看原文内容请购买。
中国西双版纳水稻老品种在育性恢复基因Rf-1位点的遗传多样性

中国西双版纳水稻老品种在育性恢复基因Rf-1位点的遗传多样性寇姝燕;谭亚玲;李俊;周英;陈丽娟;谭学林【期刊名称】《分子植物育种》【年(卷),期】2009(7)4【摘要】云南西双版纳素有"物种基因库"的美称,是稻种资源的多样性中心区和优异种质富集区之一。
本研究利用8个位于Rf-1位点的PCR分子标记对53份西双版纳地方籼稻、粳稻和光壳稻种质资源的遗传多样性和遗传分化进行了分析,共检测到15种多态性扩增片段。
Shannon's信息指数和Nei's遗传多样性指数分析表明西双版纳水稻地方品种在Rf-1位点具有较高的遗传多样性,这些遗传变异主要来自于不同品种之间的遗传差异。
分析表明西双版纳籼稻的遗传多样性要显著高于粳稻和光壳稻。
结果还表明籼稻与粳稻和光壳稻之间的分化要显著大于粳稻与光壳稻之间的分化,光壳稻与粳稻的亲缘关系更近。
这一结果可能表明水稻Rf-1位点的遗传分化与水稻的籼粳分化紧密相关,还暗示水稻Rf-1位点遗传分化可能是与籼粳分化同时进行的。
【总页数】8页(P653-660)【关键词】水稻;Rf-1位点;遗传多样性;西双版纳【作者】寇姝燕;谭亚玲;李俊;周英;陈丽娟;谭学林【作者单位】云南农业大学稻作研究所;云南西双版纳州农业科学研究所【正文语种】中文【中图分类】S831.8;Q943【相关文献】1.利用PCR标记对恢复水稻BT型胞质雄不育性的核基因Rf-1进行遗传制图 [J], ToshiyukiKomori;雷波2.宁夏主要水稻品种(系)Rf-1基因位点多态性分析 [J], 王坚;刘炜3.水稻多基因型和单基因型品种抗病性及群体内稻瘟菌遗传多样性初析 [J], 唐志明;金晓春;李晓方;刘彦卓;刘志霞;卢东柏;毛兴学;高云;卢德城4.云南地方老品种水稻中恢复基因位点遗传差异分析 [J], 寇姝燕;邹茜;刘慰华;谭亚铃;文建成;金寿林;洪汝科;谭学林;袁平荣5.Rf-1位点CAPS标记对水稻不同胞质雄性不育育性恢复系的关系分析 [J], 谭亚玲;王石华;洪汝科;文建成;郭艾;寇淑燕;金寿林;谭学林因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
研究报告RESEARCH REPORT滇一型杂交粳稻恢复基因的分子鉴定研究谭亚玲*谭学林云南农业大学稻作研究所,昆明,650201*通讯作者,cmsong@1631com摘要用位于水稻第10染色体的微卫星标记OSR33、RM228对285份滇型材料进行恢复基因的分子鉴定研究,结果表明:¹OSR33、RM228标记的基因型依材料的不同而出现分子量不同的带型,且带型与材料有关;ºOSR33和RM228鉴定材料的准确率分别为96%和90%;OSR33、RM228标记的基因型值与黑染花粉率极显著正相关,可用于鉴别恢复系;»用M apm aker/QTL软件分析供试材料的表现型,在OSR33、RM228之间,探测到1个与育性恢复有关的主效QT L,可解释8312%的表型变异,与OSR33和RM228的遗传距离分别为313cM和710cM。
关键词滇一型杂交粳稻,恢复系,分子鉴定Molecular Identific ation of the Fertility-restorer-gene of Dian-type1Hybrid Rice Tan Yaling*T an XuelinRice Research Institute of Yunnan Agricultural University,Kunmi ng,650201*Corresponding author,cmsong@1631comABST RACTThe identification of fertility-restoring-gene of Dian-type1hybrid rice w ith two simple sequence repeat (SSR)markers,OSR33and RM228locating in the middle of the long arm of chromosome10,were studied. The correct frequency of the identification w as95%w ith OSR33,and90%with RM228.Statistics analysis show ed very significant correlation betw een pollen fertility and marker genotypes.T he results indicated that the tw o m arkers could be used for identifying restorer lines of Dian type rice.A major-fertility-restoring-gene w as detection between OSR33and RM228by QT L mapping approach.T he locus w as closed to markers OSR33and RM228w ithin313cM and710cM respectively,w hich ex plained8312%phenotypic variance.KEY WORDSDian-type1hybrid rice,Fertility-restoring-g ene,M olecular identification水稻是我国重要的粮食作物之一,杂交水稻的育成为我国粮食生产和温饱问题的解决做出了巨大贡献。
有效利用水稻杂种优势的基础是雄性不育性。
研究利用最多的水稻雄性不育有两种类型:一类是遗传因素所致,包括细胞核基因控制的雄性不育、细胞质基因控制的雄性不育、细胞分子植物育种,2004年,第2卷,第2期,第209)214页M olecular P lant Br eeding,2004,V ol12,No12,209)214核基因与细胞质基因互作控制的雄性不育;另一类是由遗传和环境互作控制的雄性不育,包括光敏感及温敏感雄性不育(谭学林和师常俊,1999)。
目前,水稻生产上大面积使用的是由质核基因互作控制的雄性不育系。
在籼稻上,主要是利用野败型雄性不育,累计推广的2亿hm2杂交稻中,野败型雄性不育系占50%(曾千春等,2000)。
粳稻上则是包台型和滇一型雄性不育,虽然杂交粳稻与杂交籼稻同在1973年实现三系配套,但其未能象杂交籼稻那样得到广泛应用,主要的一个原因是粳型杂交稻的恢复系选育难,其根本原因是粳型不育系不像籼型那样,有现成的品系可作恢复系,粳型(包括包台型和滇一型)的恢复基因只存在于籼稻中,不存在于粳稻品种中,而且不是所有的籼稻都具有恢复基因,大约只有30%左右的籼稻品种有恢复基因。
所以,选育滇型恢复系和其它粳稻恢复系一样必须通过籼粳交,从籼稻中引入恢复基因(李铮友和纳信真,1980)。
此外,在选育恢复系时,由于很少有形态特征可用于鉴别恢复系,其鉴别必须通过测交,这一方法需经历两个生长季节,而且结果的准确性常受环境条件的影响。
因此,恢复基因的鉴别及恢复系的选育一直是粳型杂交水稻研究的瓶颈。
近20年来,分子标记技术的迅猛发展,加快了水稻育性恢复基因的定位和作图的研究步伐。
不同学者采用不同的分子标记技术对不同质源的育性恢复基因进行了定位研究,其中以野败型、BT型、滇一型研究较多。
尽管不同学者将野败型的恢复基因定位于第1、7、10染色体上(Bharaj et al1, 1995;Akagi et al1,1996;Zhang et al1,1997;Yao et al1,1997;Tan et al1,1998a;Jing et al1,2001;景润春等,2000),但许多结果均表明第10染色体长臂中部有一个与育性恢复有关的主效基因,距OSR33 3158cM;包台型的育性恢复基因也位于第10染色体,与OSR33紧密连锁(约317cM)(Fukuta et al1,1992;Ichikaw a et al1,1997;Liang,2000)。
滇型细胞质雄性不育系育性恢复基因的定位研究相对较少,陶大云(2002,私人通讯)发现滇一型细胞质雄性不育系育性恢复基因位于第10染色体长臂中部,OSR33、RM228之间,其连锁距分别为316cM 和815cM。
Tan等用分离群体混合法,采用微卫星分子标记技术,研究滇一型细胞质雄性不育系育性恢复基因,在第10染色体探测到一个与育性恢复有关的主效基因,其与OSR33、RM228紧密连锁,距OSR33314cM,距RM228510c M。
育性恢复基因分子定位的日趋精确,为恢复基因的分子鉴别提供了可能。
但分子标记辅助选择时,首先应考虑前景选择的可靠性。
而前景选择的可靠性主要取决于标记与目标基因间的连锁程度,只有连锁非常紧密,才能够达到较高的准确率。
选择的准确率随重组率的增加迅速下降,在分子辅助选择育种中,若要求选择的准确率达到90%以上,则标记与目标基因的重组率必须小于5%,当重组率达到10%时,选择的准确率已降到80%(方宣钧等,2001)。
根据图距小于10cM时,图距近似等于重组率,微卫星标记OSR33、RM228与滇一型恢复基因的重组率分别近似为4%、5%。
从理论上讲,如用这2个微卫星标记对滇一型杂交粳稻恢复基因选择的准确率均达到90%以上,可用于滇一型杂交粳稻恢复基因的分子标记辅助选择研究,能对现有材料早期进行选择,大大缩短育种周期。
因此,本试验采用微卫星标记OSR33和RM228筛选了滇型水稻的一些稳定品系,进行恢复基因的分子鉴别及恢复系的分子标记辅助选择研究。
1材料与方法111供试材料供试材料共285份,其中滇一型杂交水稻不育系及相应的保持系106对(编号为/不0字号);测交组合179对(不育系与稻作所籼粳交自育品系的测交一代,及相应测交父本179份;编号为/测0字号);于2001年种植于云南农业大学稻作所试验田,栽培管理同一般大田栽培。
112方法11211供试材料的育性检测开花期取不育系及测交一代的成熟颖花,用1% I2-KI溶液染色镜检。
圆形、黑染者为可育花粉;形状不规则、不染色者(典败),圆形但浅染(染败)或圆形不染者(圆败)均为不育花粉。
统计花粉可育率,成熟时考查不育系及测交一代结实情况。
11212供试材料的分子鉴定分蘖末期取保持系、测交组合父本的叶片,采用脲-酚变性法提取水稻总DNA(Tan et al1,210分子植物育种M olecular Plant Breeding1998a),溴化乙锭荧光比色测定DNA 浓度,聚丙烯酰胺凝胶电泳,银染检测多态性的方法。
PCR 扩增反应液组成、扩增程序及引物序列如下:反应液组成:ddH 2O 8L l 、10@buffer 2L l 、MgCl 2(25mol/L )2L l,dNTP (215umol/L )2L l 、引物(2umol/L)各2L l,模板DNA 2L l 、DNA 聚合酶1u 扩增程序:94e 4min;94e 1min 、55e 1min 、72e 1min,35个循环;72e 4minOSR33:5c -AACGCGAGGACACGTACTTAC -3c ;5c -ACGAGA TACGT ACGCCCTTG -3cRM228:5c -CTGGCC ATTAGTCCTTGG -3c ;5c -GC TTGCGGCTCTGCTTAC-3c 11213分子鉴定材料的可靠性分析及育性恢复基因位点的确定对106对不育系及179对测交组合进行花粉育性与标记基因型的相关分析;以花粉育性为对照,统计106对不育系及179对测交组合分子鉴定的出错率,其中测交组合黑染花粉率大于30%(计为育性恢复),及黑染花粉率小于10%(计为育性不恢复)。
并用Mapmaker/QTL 软件分析供试材料的表现型,确定恢复基因的位点。
分析时黑染花粉率大于30%的材料为育性恢复/20,小于10%的材料为育性不恢复/10,将标记基因型值为/1,20、/1,30、及黑染花粉率为10%~30%作为缺失,参数(LOD)为013,最大距离5010cM 。
2结果与分析211恢复基因的分子标记带型OSR33有分子量不同的3种带型,分别记为/10、/20、/30,其中以/30型分子量最大,/20型的分子量最小(图1)。
与OSR33不同的是,微卫星标记RM228的带型以/组0的形式出现,依据主要带型位置的不同,将其分为两类,分别记为/10、/20,以/10型分子量最大(图2)。
带型出现类型与材料类型有关,/10型主要出现于保持系及测交一代不育的测交父本;/20、/30带型主要出现在测交一代的花粉育性为半不育,结实表现/恢0的测交父本及30份自育品系。