蛋白质的合成
第十五章蛋白质的生物合成

第一位
(5ˊ)
U
U
C
A
G
遗传密码字典 第二位
C
A
G
第三位
(3ˊ)
U C A G
U C A G
U C A G
U C A G
6 4 组 密 码 子 中 , AUG 既 是 甲 硫氨酸的密码,又是起始密码; 有三组密码不编码任何氨基酸, 而是多肽链合成的终止密码子: UAG、UAA、UGA。
• 在原核生物和真核生物中,均存在另一 种携带蛋氨酸的tRNA,识别非起动部位 的蛋氨酸密码,AUG。
第十五章蛋白质的生物合成
核糖体的研究历史
1.早在本世纪30年代后期就发现细胞质和细胞核中都有 核酸存在,不过用1924年福尔根发明的染色法只能使细胞核 中的核酸染色。但两种核酸在260nm的吸收非常相似。
第十五章 蛋白质合成及转运
蛋白质的生物合成
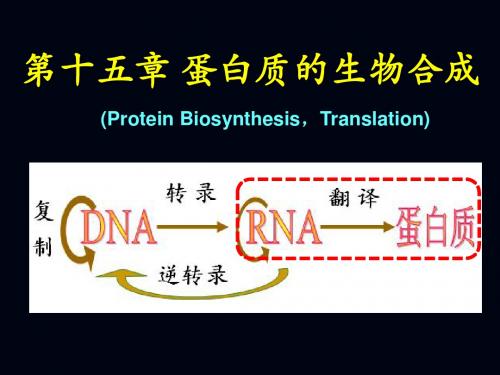
中心法则指出,遗传信息的表达最终是合成出 具有特定氨基酸顺序的蛋白质,这种以mRNA上所 携带的遗传信息,到多肽链上所携带的遗传信息的传 递,就好象以一种语言翻译成另一种语言时的情形 相似,所以称以mRNA为模板的蛋白质合成过程为 翻译(translation)。
2.Crick, F.比较了核酸和氨基酸的大小和形状后,认为不可 能在空间上互补,因此预测:(1) 存在一类分子转换器,使信 息从核酸序列转换成氨基酸序列;(2) 这种分子很可能是核酸; (3) 它不论以何种方式进入蛋白质翻译系统的模板,都必须与 模板形成氢键(即配对);(4) 有20种分子转换器,每种氨基 酸一个;(5) 每种氨基酸必定还有一个对应的酶,催化与特定 的分子转换器结合。
2.1941年,细胞学家J.Brachet和T.Caspersor注意到细 胞质中的核酸与蛋白质的合成有密切的关系。
简述蛋白质合成的过程

简述蛋白质合成的过程蛋白质是构成细胞的重要组成部分,也是细胞功能的基础。
蛋白质合成是生物体中一项重要的生化过程,通过合成蛋白质,维持细胞正常的生理功能。
蛋白质合成过程主要包括转录和翻译两个阶段。
1. 转录阶段转录是指将DNA上的基因信息转录成RNA的过程。
在转录过程中,DNA的双链解旋,RNA聚合酶与DNA的一个特定区域结合,开始合成RNA。
这个特定区域称为启动子,它位于基因的上游区域。
RNA聚合酶沿着DNA模板链向下滑动,合成RNA链。
合成RNA 的过程中,RNA聚合酶会依据DNA的碱基序列信息合成对应的RNA碱基,形成RNA单链。
合成的RNA称为信使RNA (mRNA)。
2. 翻译阶段翻译是指将mRNA上的信息转化为蛋白质的过程。
翻译过程发生在细胞质内的核糖体中。
在翻译之前,mRNA先经过剪接修饰,将其内部的非编码区域(内含子)剪除,只保留编码区域(外显子)。
然后,mRNA与核糖体中的核糖体RNA(rRNA)和转移RNA (tRNA)相互作用。
翻译的第一步是启动,mRNA的5'端与小核糖体亚基结合,tRNA 与起始密码子结合在小核糖体亚基上的P位。
然后,大核糖体亚基和小核糖体亚基结合,形成功能完整的核糖体。
核糖体沿着mRNA 滑动,tRNA的反应中心与mRNA上的对应密码子结合。
tRNA携带的氨基酸与上一个tRNA携带的氨基酸进行肽键形成,合成蛋白质的链。
这个过程称为肽链延伸。
当核糖体滑动到终止密码子时,翻译停止,蛋白质合成结束。
总结起来,蛋白质合成的过程包括转录和翻译两个阶段。
转录是将DNA上的基因信息转录成mRNA,翻译是将mRNA上的信息转化为蛋白质。
蛋白质合成是一项复杂而精细的生化过程,其中涉及到多种酶的参与和调控。
蛋白质合成的正常进行对于细胞的正常功能和生理过程具有重要意义。
分子生物学第十五章 蛋白质的生物合成

1.遗传密码种类:
• mRNA 分子有4种碱基:A、G、C、U,可组合成64个密 码子,其中61个分别代表20种不同氨基酸
• 遗传密码共有43=64种, 64: UAA、UAG、UGA 终止密码 61: AUG 起始密码 代表Met 60: 代表19种氨基酸
9
遗传密码表 第一碱基
(5/-端)
第二碱基
43
一、起始阶段
(一)原核生物翻译起始复合物的形成
参与的物质: 核糖体50S和30S大小两类亚基 mRNA 起始作用的fMet-tRNAfMet GTP供能 起始因子(IF1、IF2、IF3)
44
起始过程:
• 核糖体大小亚基解离 • 小亚基与mRNA结合:
16sRNA识别mRNA的SD序列
• fMet-tRNAfMet与mRNA起始密码子AUG结合 • 核糖体大小亚基形成起始复合物
tRNAphe Phe-tRNAphe
• 肽链起始和延长的甲硫氨酰-tRNA:
真核生物:起始: Met-tRNAiMet 延长: Met-tRNAMet
原核生物: 起始:fMet-tRNAfMet 延长:Met-tRNAMet
39
40
第三节 肽链的合成过程
41
整个翻译过程可分为 :
• 翻译的起始 • 翻译的延长 • 翻译的终止
tRNA反密码子 第1位碱基
mRNA密码子 第3位碱基
I
U
G AC
U, C, A A, G U, C U G
17
摆 动 配 对
32 1
U
123
18
• ⑤遗传密码的通用性
从原核生物到人类都共用同一套遗传密码,被称为遗传密码的通用性。 密码的通用性进一步证明各种生物进化自同一祖先。
蛋白质合成的基本过程

蛋白质合成的基本过程蛋白质是构成生物体细胞的重要组成部分,参与了生物体内的几乎所有生化过程。
蛋白质的合成是细胞内最为重要的生物化学过程之一,也是维持生命活动正常进行的基础。
蛋白质的合成过程包括转录和翻译两个阶段,通过这两个阶段,细胞可以根据遗传信息合成出具有特定功能的蛋白质。
下面将详细介绍蛋白质合成的基本过程。
一、转录阶段转录是指在细胞核内DNA模板上合成RNA的过程。
在蛋白质合成中,首先需要将DNA上的遗传信息转录成RNA,形成mRNA(信使RNA),mRNA携带着DNA上的遗传信息,将其带到细胞质中进行翻译合成蛋白质。
1.1 RNA聚合酶的结合转录的第一步是RNA聚合酶与DNA模板的结合。
RNA聚合酶是一种酶类蛋白质,它能够识别DNA上的启动子区域,并在该区域结合,开始合成RNA链。
1.2 RNA链的合成RNA聚合酶在DNA模板上沿着3'→5'方向移动,合成RNA链时是在5'→3'方向进行的。
RNA链的合成过程与DNA复制有所不同,RNA链的合成速度较快,而且只合成一条链。
1.3 终止转录在DNA上的终止子区域,会有一些特定的序列,当RNA聚合酶合成到这些序列时,转录过程会终止,RNA链会从DNA模板上脱离,形成成熟的mRNA。
二、翻译阶段翻译是指在细胞质中mRNA的遗传信息被翻译成氨基酸序列的过程。
翻译过程中涉及到多种RNA和蛋白质,包括tRNA(转运RNA)、rRNA (核糖体RNA)和核糖体等。
2.1 核糖体的结合在翻译的起始阶段,mRNA会与核糖体结合,核糖体是一种由rRNA和蛋白质组成的细胞器,能够将mRNA上的遗传信息翻译成氨基酸序列。
2.2 tRNA的运载tRNA是一种带有特定氨基酸的RNA分子,它能够将氨基酸运载到核糖体上,与mRNA上的密码子配对,完成氨基酸的添加。
2.3 氨基酸的连接在核糖体上,tRNA将氨基酸按照mRNA上的密码子顺序连接起来,形成氨基酸链。
蛋白质的合成与运输

)
【解析】 内质网是脂质合成的场所,所以合成固醇类激素的分泌细胞内质网 应该比较发达。分泌蛋白的修饰和加工是由内质网和高尔基体共同完成。生物膜之 间可通过具膜小泡的转移进行相互转化。
【答案】 B
5.如图表示用含 3H 标记的氨基酸培养液培养某细胞过程中蛋白质的合成和分泌 示意图,则该细胞中出现 3H 的部位依次为( )
【答案】 B
2.为了研究酵母菌细胞内蛋白质的合成, 研究人员在其培养基中添加 3H 标记的 亮氨酸后,测得与合成和分泌乳蛋白相关的一些细胞器上放射性强度的变化曲线如 下图中甲,有关的生物膜面积变化如图乙,其相关结构关系如图丙,则下列有关说 法不正确的是( )
A.图丙中首先可观察到 3H 标记的细胞器是③ B.能在图丙中④上观察到 3H 标记表明可能有分泌蛋白合成 C.图甲中 c 曲线所指的细胞结构是高尔基体 D.图乙中 d 曲线表示的细胞结构是内质网
提示:原核细胞除细胞膜外无其他膜结构,因此没有内膜系统。
[思维升华] 生物膜的种类、结构和功能联系
细胞膜:单层膜,细胞的外界膜 核膜:双层膜,具核孔,细胞核的外界膜 生物膜 双层膜结构:叶绿体、线粒体 (1) 的种类 细胞器膜单层膜结构:内质网、高尔基 体、液泡等
1.关于蛋白质的分选和运输,下列说法不正确的是( A.蛋白质合成后,一般在其氨基酸序列中含有分选信号 B.游离的核糖体合成的蛋白质中无分选信号
)
C.核糖体在细胞中的存在部位不同,也影响蛋白质的去向 D.蛋白质只有被准确地运输到相应的部位才能执行特定的功能
【解析】 无论是在内质网的核糖体上还是游离的核糖体上合成的蛋白质,一 般其氨基酸序列中均含分选信号,决定着蛋白质的去向和最终定位,而核糖体在细 胞中的存在部位不同,也影响蛋白质的去向。
细胞内各种蛋白质的合成和转运途径

细胞内各种蛋白质的合成和转运途径引言:细胞是生物体的基本单位,其中蛋白质是构成细胞的重要组成部分。
细胞内的蛋白质合成和转运途径是维持细胞正常功能的关键过程。
本文将介绍细胞内蛋白质合成的主要途径,包括转录、翻译和后转录修饰,以及蛋白质的转运途径,包括核糖体、内质网和高尔基体等。
一、蛋白质合成的途径1. 转录蛋白质合成的第一步是转录,即将DNA中的基因信息转录成RNA。
在细胞核中,DNA的双链解旋,RNA聚合酶结合到DNA上,根据DNA模板合成mRNA。
mRNA是一条单链RNA,它携带着从DNA中转录得到的基因信息。
2. 翻译翻译是蛋白质合成的第二步,即将mRNA上的基因信息翻译成蛋白质。
翻译发生在细胞质中的核糖体中。
核糖体由rRNA和蛋白质组成,它能够识别mRNA上的密码子,并将相应的氨基酸连接起来,形成多肽链。
翻译的过程包括起始、延伸和终止三个阶段,通过tRNA和蛋白因子的参与完成。
3. 后转录修饰蛋白质合成的最后一步是后转录修饰,即对新合成的蛋白质进行修饰和折叠。
这一过程发生在内质网和高尔基体中。
内质网是一个复杂的膜系统,它能够将新合成的蛋白质进行折叠和修饰,如糖基化、磷酸化等。
高尔基体则进一步对蛋白质进行修饰,并将其定位到细胞的不同位置。
二、蛋白质的转运途径1. 核糖体核糖体是蛋白质合成的场所,它位于细胞质中。
在核糖体中,mRNA上的密码子与tRNA上的反密码子互补配对,通过蛋白因子的辅助,将氨基酸连接成多肽链。
核糖体能够识别起始密码子和终止密码子,从而控制蛋白质的合成过程。
2. 内质网内质网是一个复杂的膜系统,它位于细胞质中。
内质网上的核糖体能够合成蛋白质,并将其进行折叠和修饰。
折叠不正确的蛋白质将被内质网上的分解酶降解,而正确折叠的蛋白质则会进一步转运到高尔基体或其他细胞器。
3. 高尔基体高尔基体是一个复杂的膜系统,它位于细胞质中。
高尔基体接收来自内质网的蛋白质,并对其进行进一步修饰和定位。
蛋白质合成PPTPPT课件

蛋白质合成的细胞定位
总结词
蛋白质合成主要发生在细胞内的核糖体上,核糖体是 细胞内蛋白质合成的场所。
详细描述
核糖体是细胞内一种由RNA和蛋白质组成的颗粒状结 构,主要存在于细胞质中。核糖体在蛋白质合成过程中 起着至关重要的作用,它能够读取mRNA上的遗传信 息,将一个个氨基酸按照特定的顺序连接起来形成多肽 链。同时,核糖体还具有催化肽键形成的酶活性,促进 蛋白质合成的进行。除了核糖体外,细胞内还有其他一 些细胞器也参与了蛋白质的合成过程,如内质网、高尔 基体等。这些细胞器在蛋白质的修饰、加工和运输等方 面起着重要作用。
蛋白质合成PPT课件
目录
• 蛋白质合成简介 • 蛋白质合成的过程 • 蛋白质合成的调控 • 蛋白质合成与疾病的关系 • 研究展望
01
蛋白质合成简介
蛋白质合成的基本概念
总结词
蛋白质合成是指细胞内利用已有的小分子物质作为原料,通过一系列酶促反应将氨基酸 按照特定的顺序连接起来形成多肽链,进而形成具有特定结构和功能的蛋白质的过程。
翻译后加工与修饰
总结词
翻译后加工与修饰是蛋白质合成的重要环节,涉及多 种酶促反应和化学修饰。
详细描述
翻译后加工与修饰是蛋白质合成的最后阶段,涉及到 多种酶促反应和化学修饰。这些加工和修饰包括剪切 、磷酸化、糖基化、乙酰化等,有助于完善蛋白质的 结构和功能。这些加工和修饰过程通常在特定的细胞 器或细胞部位进行,需要特定的酶和化学环境的支持 。通过翻译后加工与修饰,蛋白质的结构和性质得以 最终确定,从而发挥其在细胞生命活动中的重要功能 。
04
蛋白质合成与疾病的关系
蛋白质合成异常与疾病的发生
癌症
蛋白质合成异常可能导 致细胞增殖失控,引发
第15章 蛋白质的生物合成
进 位
成肽 转 位
成肽
成肽是由转肽酶催化形成肽键的过程。
Tu GTP
Tu GDP
5'
AUG
3'
fMet fMet
Tu GTP
5'
AUG
3'
(三) 原核生物翻译终止
当mRNA上的终止密码子出现后,多肽链合成 停止,肽链从肽酰-tRNA中脱落,mRNA、核糖体 等解体,此时翻译过程终止。 终止密码子不被任何氨基酰-tRNA识别,只有 释放因子(RF)能够识别它并进入核糖体的A位。 1. 释放因子可引起转肽酶活性的改变,从而使 肽链从tRNA上脱落。 2. 释放因子可引起核糖体结构的改变,从而使 复合体趋于解体。
解码
反密码子(anti-codon) 反密码子是位于tRNA、可与mRNA的三联体密 码子配对的三个相邻核苷酸。在蛋白质的合成中, 起解读密码、辅助将特异的氨基酸引入合成位点。 tRNA
5'
AUG
反密码子
3'
mRNA
密码子
遗传密码的特点
翻译时遗传密码的阅读方向是 5’→3’,即读 码从mRNA的起始密码子AUG开始,按5’→3’ 的方向逐一阅读,直至终止密码子。
COORF
5'
UAG
3'
第四节
蛋白质翻译后加工和靶向输送
Posttranslational Modification and Targeting Transfer of Protein
蛋白质翻译后加工
《蛋白质的合成》课件

翻译水平的调控
翻译起始
翻译后修饰
翻译起始是蛋白质合成的关键步骤之 一,涉及核糖体与mRNA的结合以及 起始密码子的识别。
翻译后的蛋白质还需要经过一系列的 修饰才能成为有功能的分子,如磷酸 化、乙酰化、糖基化等。
翻译延长与终止
在翻译过程中,核糖体的移位速度和 肽酰-tRNA的形成受到多种因素的调 节,如mRNA的结构、tRNA的浓度 和种类、核糖体的构象等。
04
CATALOGUE
蛋白质合成的应用
蛋白质工程
蛋白质工程是通过修改或设计蛋白质 的氨基酸序列,以达到改善蛋白质的 某些功能或创建新功能的技术。
蛋白质工程在医药、农业和工业领域 有广泛应用,例如设计新的药物、改 良酶的催化效率和稳定性、优化蛋白 质的表达和纯化等。
药物设计与开发
01
蛋白质合成在药物设计与开发中 发挥着关键作用,因为许多药物 是直接与蛋白质相互作用来发挥 其治疗作用的。
蛋白质合成的分子机制
mRNA在蛋白质合成中的作用
信息传递者
mRNA是DNA遗传信息的传递者 ,将遗传信息从DNA传递到核糖 体,用于指导蛋白质的合成。
遗传信息的解码
mRNA上的密码子与tRNA上的反 密码子互补配对,实现遗传信息 的解码,确保氨基酸按照正确的 顺序连接起来。
tRNA在蛋白质合成中的作用
氨基酸转运
tRNA作为氨基酸的转运载体,将氨基酸按照mRNA上的指令准确运送到核糖 体的A位点,参与蛋白质的合成。
反密码子识别
tRNA的反密码子能够与mRNA上的密码子进行互补配对,确保氨基酸按照正确 的顺序加入到多肽链中。
rRNA在蛋白质合成中的作用
核糖体的组成
rRNA是核糖体的主要组成部分,核 糖体是蛋白质合成的场所。
第八章:蛋白质的合成与加工

3. fMet-tRNAfMet被起始因子IF2和一分子的GTP一同带到小亚基 的P位
密码虽有简并性,但在特定的物种中,基因并非平均地使用 每个密码子,这种现象称为密码子偏倚(codon bias)。 如亮氨酸可由6个密码子编码(TTA、TTG、CTT、CTC、 CTA和CTG)。但是在人类基因中,大多被CTG编码,而且 几乎不被TTA或CTA编码。密码子偏倚的生物学原因并不清 楚,但不同种属的生物有不同的偏好。
(3)密码子具有普遍性
高等生物和低等生物在很大程度上共用一套密码子,说明核 酸中的核苷酸序列与蛋白质中的氨基酸序列之间对应关系的 遗传密码在进化的早期就已确定下来了。但是,也存在一些 例外的情况。
例外1:在支原体中,终止密码子UGA被用来编码Trp,而 UGG仍是Trp的密码子,但用得很少。
ቤተ መጻሕፍቲ ባይዱ
例外2:在一些纤毛虫(单细胞原生动物)种,终止密码子 UAA、UAG被用来编码谷氨酰胺。
2. 核糖体小亚基与IF3附着于mRNA上的核糖体结合位点上
30S亚基在mRNA上的结合位置,正好使起始密码子置于30S 亚基P位上。而在30S亚基的A位上结合有IF1,其作用是防止 tRNA在翻译的起始阶段结合到A位上
IF3在上一轮蛋白质合成即将结束时开始与小亚基结合,并 协助70S核糖体解离成大、小亚基。由此可看出,IF3具有双 重功能:一方面它与30S亚基结合后,阻止30S亚基与50S亚 基重新结合成70S核糖体,使30S亚基处于游离状态;另一 方面它还能辅助30S亚基与mRNA结合,30S亚基也必须有 IF3的参与才可能与mRNA形成复合体。
7.2.2 tRNA的三级结构
通过X-衍射来研究几种 酵母的tRNA晶体,发现 各种tRNA 存在共同的三 维结构。每一tRNA的三 叶草型二级结构都要折叠 成致密的倒L型三级结构。
蛋白质合成的基本过程

蛋白质合成的基本过程蛋白质是构成生物体细胞的重要组成部分,参与了生物体内的各种生命活动。
蛋白质的合成是一个复杂而精密的过程,需要多种生物分子和酶的协同作用。
本文将介绍蛋白质合成的基本过程,包括转录和翻译两个主要阶段。
一、转录转录是蛋白质合成的第一步,发生在细胞核内。
转录的过程是将DNA上的遗传信息转录成RNA的过程。
具体步骤如下:1.1 RNA聚合酶的结合:在转录开始前,RNA聚合酶会与DNA上的启动子序列结合,形成转录起始复合物。
1.2 DNA的解旋和开放:RNA聚合酶在启动子序列的作用下,使DNA解旋并开放,暴露出一段DNA模板链。
1.3 RNA合成:RNA聚合酶沿着DNA模板链逐一将核苷酸加入RNA链中,根据DNA模板的碱基序列合成RNA链。
RNA链的合成是以5'→3'方向进行的。
1.4 终止:当RNA聚合酶到达终止子序列时,转录终止,RNA链与DNA分子分离,形成初级转录产物。
二、翻译翻译是蛋白质合成的第二步,发生在细胞质中的核糖体上。
翻译的过程是将RNA上的密码子翻译成氨基酸序列的过程。
具体步骤如下:2.1 tRNA的递送:tRNA携带特定的氨基酸,通过抗密码子与mRNA 上的密码子互补配对,将氨基酸递送到核糖体上。
2.2 核糖体的组装:核糖体由大亚基、小亚基和mRNA组成,tRNA 带来的氨基酸在核糖体上进行连接。
2.3 蛋白质合成:核糖体沿着mRNA上的密码子逐个读取,根据密码子对应的氨基酸将氨基酸连接成多肽链。
2.4 终止:当核糖体读取到终止密码子时,翻译终止,核糖体释放合成的多肽链,蛋白质合成完成。
总结:蛋白质合成是一个精细的生物学过程,包括转录和翻译两个主要阶段。
在转录过程中,RNA聚合酶将DNA上的遗传信息转录成RNA;在翻译过程中,核糖体将RNA上的密码子翻译成氨基酸序列,合成蛋白质。
这两个过程密切配合,确保蛋白质的准确合成,从而维持生物体内正常的生命活动。
第二章 蛋白质的合成、转运、加工与修饰

顺反子: 顺反子 : 编码一种多肽链并连同起始信号和终止 信号在内的DNA区段。 区段。 信号在内的 区段 单顺反子mRNA:编码一种多肽链的mRNA分子。 :编码一种多肽链的 分子。 单顺反子 分子 多顺反子mRNA: 编码数种不同多肽链的同一条 : 多顺反子 mRNA分子。多见于原核生物。 分子。 分子 多见于原核生物。 反义链/有意义链 ( ) 模板链 双链DNA分子中 模板链: 反义链 有意义链/(-)链/模板链:双链 有意义链 分子中 被转录成RNA转录本的链。 转录本的链。 被转录成 转录本的链 正义链/无意义链 ( ) 正义链 无意义链/(+)链 无意义链
(S) )
SD 序 列 / 核 糖 体 结 合 位 点 ( ribosomal binding site , RBS) : 原核细胞 的翻译起始密码子AUG的上游 ) 原核细胞mRNA的翻译起始密码子 的翻译起始密码子 的上游 相距8~ 个核苷酸处有一段由 个核苷酸处有一段由4~ 个核苷酸组成的富含 相距 ~13个核苷酸处有一段由 ~6个核苷酸组成的富含 嘌呤的序列, 为核心, 嘌呤的序列 , 以 5’-AGGA-3’为核心,它与核糖体小亚基 为核心 上的16S-rRNA 的近 末端处的一段短序列互补。 的近3’末端处的一段短序列互补 末端处的一段短序列互补。 上的 Kozak序列 Kozak序列:a favorable context for efficient eukaryotic 序列: translation initiation(PuNNATGPu)。(S) ( ) ) 典型的Poly(A)加尾信号:AATAAA。(S) 加尾信号: 典型的 加尾信号 。 ) cDNA 末 端 快 速 扩 增 法 ( rapid amplification of cDNA ends, RACE)(S) , ) )
蛋白质生物合成的过程

蛋白质生物合成的过程蛋白质是构成生命体的重要组成部分,其生物合成过程也是生命活动的重要环节之一。
蛋白质生物合成包含了两个主要的过程:转录和翻译。
在这两个过程中,多种分子和酶的参与,共同完成了蛋白质的合成。
转录是蛋白质生物合成的第一步,它发生在细胞核内。
在这一过程中,DNA的信息被转录成RNA分子,这个过程由RNA聚合酶完成。
RNA聚合酶可以识别DNA链上的启动子区域,并沿着DNA链逐渐合成RNA分子。
RNA分子的合成是由核苷酸单元的连接而成的,这些核苷酸单元包括腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)和尿嘧啶(U)。
RNA分子的合成是由DNA模板的编码信息决定的,这也就是RNA分子与DNA分子之间的信息转换。
在翻译过程中,RNA分子将信息转化为蛋白质的氨基酸序列。
这个过程发生在细胞质中,由核糖体完成。
核糖体是由RNA和蛋白质组成的复合物,其中RNA分子起到了信息传递的作用,而蛋白质则提供了支持和催化的功能。
在翻译过程中,RNA分子的信息被翻译成一系列的氨基酸,这些氨基酸按照特定的顺序连接在一起,形成了蛋白质分子。
蛋白质的生物合成过程是一个高度协调的过程,多种分子和酶在其中发挥了重要的作用。
在转录过程中,RNA聚合酶需要与其他蛋白质组成复合物,才能识别启动子区域并完成RNA分子的合成。
在翻译过程中,核糖体需要与多种因子协同作用,才能完成氨基酸的连接和蛋白质的合成。
此外,蛋白酶和蛋白质折叠酶等分子也参与了蛋白质的后续加工过程,保证了蛋白质的正确折叠和功能发挥。
总的来说,蛋白质生物合成是一个复杂而精细的过程,其中涉及到多种分子和酶的协同作用。
这个过程不仅是生命活动的基础,也具有重要的生物学意义。
通过对蛋白质生物合成过程的研究,人们可以更好地理解生命的本质和机制,同时也可以为生物医学研究和药物研发提供有力的支持。
蛋白质合成基本步骤

蛋白质合成基本步骤蛋白质是生物体内重要的基础物质,参与了许多生命活动的调控和实施。
蛋白质的合成是生物体内的一项重要过程,其基本步骤包括转录和翻译两个阶段。
1. 转录阶段转录是指在DNA模板上合成RNA的过程。
首先,DNA的双链解旋,使得DNA的编码链作为模板进行转录。
随后,核酸酶将RNA 的核苷酸与DNA模板上的互补碱基配对,形成RNA链。
这个过程中,A-T和G-C的碱基配对规则得以保持。
转录的终止是由终止密码子信号序列引发的。
2. 翻译阶段翻译是指通过核糖体将RNA的信息转换为蛋白质的过程。
此阶段分为三个步骤:起始、延伸和终止。
起始阶段:在起始阶段,mRNA的起始密码子与tRNA的抗密码子完全互补配对,导致核糖体与mRNA的起始区域结合。
同时,氨基酸的甲基化tRNA与核糖体的P位结合,形成一个功能齐全的起始复合体。
延伸阶段:在延伸阶段,核糖体依次从mRNA上读取下一个密码子,使tRNA与mRNA进行互补配对。
通过蛋白质合成酶的催化作用,将氨基酸与前一个氨基酸连接起来,形成多肽链。
这个过程将一直进行,直到遇到停止密码子。
终止阶段:终止阶段是指当核糖体读取到停止密码子时,翻译过程停止。
停止密码子并不对应具体的氨基酸,而是信号终止翻译的标志。
当核糖体读取到停止密码子时,特定的释放因子结合到核糖体上,导致核糖体、mRNA和新合成的蛋白质分离。
除了这两个基本步骤外,蛋白质合成还受到其他因素的调控。
1. 转录的调控转录的调控是指通过调节DNA转录的速率和位置来控制蛋白质合成的过程。
转录调控是生物体内基因表达的重要方式之一。
在转录过程中,转录因子结合到启动子区域,以调节特定基因的转录水平。
2. 翻译的调控翻译的调控是指通过调节翻译的速率和位置来控制蛋白质合成的过程。
在翻译过程中,一些调控因子可以结合到mRNA和核糖体上,以影响翻译的效率和准确性。
蛋白质的合成是一个复杂的过程,包括转录和翻译两个阶段。
在这个过程中,DNA的信息被转录成RNA,然后通过翻译过程转化为蛋白质。
蛋白质合成、加工和转运的过程

一、蛋白质的合成1、核糖体是合成蛋白质的机器,其功能是按照mRNA的指令由氨基酸合成蛋白质。
2、游离核糖体游离于胞质中,合成细胞内的基础蛋白质;附着核糖体,附着在内质网表面,构成粗面内质网的核糖体,合成分泌蛋白和膜蛋白。
3、蛋白质合成的一般过程:1)氨基酸的活化。
氨基酸和tRNA在氨酰—tRNA合成酶作用下合成活化的氨酰—tRNA。
2)起始、延伸和终止。
3)蛋白质合成后的加工。
肽链N端Met的去除;氨基酸残基的化学修饰,乙酰化、甲基化、磷酸化等;肽链的折叠;二硫键的形成。
二、蛋白质的分泌合成、加工修饰和转运1、信号肽介导分泌性蛋白在粗面内质网的合成。
1)信号肽是蛋白质合成中最先被翻译出来的一段氨基酸序列,通常由18-30个疏水氨基酸组成,能指引核糖体与内质网结合,并引导合成的多肽链进入内质网腔。
2)新生分泌性蛋白质多肽链在胞质中的游离核糖体上起始合成。
当新生肽链N端的信号肽被翻译后,可立即被细胞质基质中的信号识别颗粒(SRP)识别、结合。
3)与信号肽识别结合的SRP,识别结合内质网膜上的SRP-R,并介导核糖体锚泊附着于内质网膜的通道蛋白移位子上。
而SRP则从信号肽—核糖体复合体上解离,返回细胞质基质中重复上述过程。
4)在信号肽的引导下,合成中的肽链,通过由核糖体大亚基的中央管和移位子蛋白共同形成的通道,穿膜进入内质网网腔。
随之,信号肽序列被内质网膜戗面的信号肽酶且除,新生肽链继续延伸,直至完成而终止。
最后完成肽链合成的核糖体大、小亚基解聚,并从内质网上解离。
2、跨膜驻留蛋白的插入和转移决定了蛋白质的两种去处:1)穿过膜进腔,为可溶性蛋白质,包括分泌蛋白和内质网驻留蛋白。
2)嵌入内质网膜中,形成膜蛋白。
3、粗面内质网与外输性蛋白质的分泌合成、加工修饰和转运过程密切相关。
1)新生多肽链的折叠与装配,与合成同时发生。
内质网为新生多肽链正确的折叠和装配提供了有利的环境。
分子伴侣通过对多肽链的识别结合来协助它们的折叠组装和转运。
蛋白质合成、修饰及分泌

03
蛋白质分泌
囊泡运
01
02
03
04
囊泡运输是指蛋白质在细胞内 合成后,通过一系列的囊泡转 运过程,将其从内质网等合成 场所转运至高尔基体进行加工 和修饰,再通过胞吐作用分泌 到细胞外。
囊泡运输是指蛋白质在细胞内 合成后,通过一系列的囊泡转 运过程,将其从内质网等合成 场所转运至高尔基体进行加工 和修饰,再通过胞吐作用分泌 到细胞外。
蛋白质合成、修饰及分泌
目
CONTENCT
录
• 蛋白质合成 • 蛋白质修饰 • 蛋白质分泌 • 蛋白质合成、修饰及分泌的相互关
系
01
蛋白质合成
氨基酸的合成
氨基酸是蛋白质的基本组成单 位,通过生物合成或从食物中 摄取来获得。
生物合成氨基酸主要通过转氨 基作用、脱羧基作用和氨基转 移作用等途径来完成。
不同的生物合成氨基酸的方式 不同,例如植物和微生物可以 通过光合作用将二氧化碳转化 为有机物,而动物则必须从食 物中摄取氨基酸。
核糖体合成
02
01
03
核糖体是细胞内合成蛋白质的场所,由大、小两个亚 基组成。
核糖体合成蛋白质的过程包括氨基酸的活化、肽链合 成的起始、肽链的延长和终止等步骤。
在核糖体合成过程中,mRNA作为模板指导氨基酸按 照特定的顺序排列,形成具有一定功能的蛋白质。
胞吐作用的异常可以导 致多种疾病的发生,如 神经退行性疾病、免疫 系统疾病等。
04
蛋白质合成、修饰及分泌的相互关系
合成与修饰的关系
合成是修饰的前提
蛋白质合成首先需要氨基酸按照特定 的序列连接成多肽链,然后经过翻译 后修饰,如磷酸化、乙酰化等,才能 形成具有生物活性的蛋白质。
蛋白质的合成及生物学意义

蛋白质的合成及生物学意义1.引言1.1 概述蛋白质是生命体中最重要的一类有机物,广泛存在于细胞内的各个结构和器官中。
它们不仅是构成细胞的基本组成元素,还承担着许多生物学功能,如酶的催化作用、细胞信号传导、免疫系统的功能等。
蛋白质的合成是细胞内的一个复杂过程,它涉及到DNA的转录和翻译过程。
蛋白质的合成始于DNA的转录过程。
在细胞核中,DNA的双链被解旋,然后通过转录酶将其中的一条链作为模板,合成RNA分子。
所合成的RNA分子被称为信使RNA(mRNA),它与DNA模板链上的碱基互补配对。
转录过程完成后,mRNA会通过核孔离开细胞核,进入细胞质。
接下来,mRNA进入细胞质后会与核糖体结合,进而开始翻译过程。
翻译是将RNA分子的信息转化为氨基酸序列的过程。
核糖体会按照mRNA上的密码子信息,从细胞质中寻找适配的转移RNA(tRNA),并将tRNA携带的氨基酸添加到正在合成的蛋白质链上。
这个过程一直进行到达到终止密码子,蛋白质的合成才会停止。
蛋白质的合成过程受到多种调控机制的影响。
调控机制既包括转录水平上的调控,也包括翻译水平上的调控。
在转录水平上,细胞会根据环境的需要,调控特定基因的表达,从而合成特定的蛋白质。
在翻译水平上,细胞可以通过调节核糖体的结合、tRNA的可用性以及蛋白质的翻译后修饰等方式,对蛋白质的合成进行调控。
蛋白质的合成与生物学意义密不可分。
蛋白质是细胞内生命活动的主要参与者,它们可以通过酶的作用促进代谢反应的进行,调节细胞内的信号传导通路,参与细胞的增殖和分化等过程。
此外,蛋白质还可以作为结构蛋白质维持细胞的形态稳定性,构成细胞骨架和细胞器的组成部分。
因此,了解蛋白质的合成过程以及其在生物学中的意义,对于深入理解细胞的功能和调控机制具有重要意义。
总而言之,蛋白质的合成是一个复杂的过程,通过转录和翻译将DNA 中的遗传信息转化为蛋白质的氨基酸序列。
蛋白质在细胞内扮演着重要的角色,参与了多个生物学过程的调控和执行。
蛋白质合成的过程中脱去的水分子

蛋白质合成的过程中脱去的水分子
蛋白质合成是生物体内重要的生物化学过程,其过程中蛋白质的合成是通过多肽键的形成来完成的。
在蛋白质合成过程中,脱去的水分子是由氨基酸之间的羟基和羧基所释放的。
具体来说,当两个氨基酸通过肽键连接时,羧基(-COOH)和氨基(-NH2)之间的羟基(-OH)和氢原子(-H)释放出一个水分子,这个过程称为脱水缩合反应。
在这个反应过程中,羧基中的羟基氧原子和氨基中的氢原子结合形成水分子,同时两个氨基酸分子通过肽键连接在一起,形成了一个新的多肽链。
蛋白质合成过程中脱去的水分子是非常重要的,因为它代表着氨基酸之间的连接过程,这种连接是蛋白质结构形成的基础。
蛋白质的空间结构和功能都受到氨基酸连接方式的影响,而脱水缩合反应正是这种连接的关键步骤之一。
除了蛋白质合成过程中的脱水缩合反应外,水还在其他生物化学反应中发挥着重要作用,例如水解反应中水分子参与了酶促反应的催化过程。
总的来说,水在生物体内的化学反应中起着至关重要的作用,蛋白质合成过程中脱去的水分子只是其中之一。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第一章糖一、糖的概念糖类物质是多羟基(2个或以上)的醛类(aldehyde)或酮类(Ketone)化合物,以及它们的衍生物或聚合物。
据此可分为醛糖(aldose)和酮糖(ketose)。
还可根据碳层子数分为丙糖(triose),丁糖(terose),戊糖(pentose)、己糖(hexose)。
最简单的糖类就是丙糖(甘油醛和二羟丙酮)由于绝大多数的糖类化合物都可以用通式Cn (H2O)n表示,所以过去人们一直认为糖类是碳与水的化合物,称为碳水化合物。
现在已经这种称呼并恰当,只是沿用已久,仍有许多人称之为碳水化合物。
二、糖的种类根据糖的结构单元数目多少分为:(1)单糖:不能被水解称更小分子的糖。
(2)寡糖:2-6个单糖分子脱水缩合而成,以双糖最为普遍,意义也较大。
(3)多糖:均一性多糖:淀粉、糖原、纤维素、半纤维素、几丁质(壳多糖)不均一性多糖:糖胺多糖类(透明质酸、硫酸软骨素、硫酸皮肤素等) (4)结合糖(复合糖,糖缀合物,glycoconjugate):糖脂、糖蛋白(蛋白聚糖)、糖-核苷酸等(5)糖的衍生物:糖醇、糖酸、糖胺、糖苷三、糖类的生物学功能(1) 提供能量。
植物的淀粉和动物的糖原都是能量的储存形式。
(2) 物质代谢的碳骨架,为蛋白质、核酸、脂类的合成提供碳骨架。
(3) 细胞的骨架。
纤维素、半纤维素、木质素是植物细胞壁的主要成分,肽聚糖是细胞壁的主要成分。
(4) 细胞间识别和生物分子间的识别。
细胞膜表面糖蛋白的寡糖链参与细胞间的识别。
一些细胞的细胞膜表面含有糖分子或寡糖链,构成细胞的天线,参与细胞通信。
2红细胞表面ABO血型决定簇就含有岩藻糖。
第一节单糖一、单糖的结构1、单糖的链状结构确定链状结构的方法(葡萄糖):a. 与Fehling试剂或其它醛试剂反应,含有醛基。
b. 与乙酸酐反应,产生具有五个乙酰基的衍生物。
c. 用钠、汞剂作用,生成山梨醇。
图2最简单的单糖之一是甘油醛(glyceraldehydes),它有两种立体异构形式(Stereoismeric form),图7.3。
这两种立体异构体在旋光性上刚好相反,一种异构体使平面偏振光(Plane polarized liyot)的偏振面沿顺时针方向偏转,称为右旋型异构体(dextrorotary),或D型异构体。
另一种异构体则使平面偏振不的编振机逆时针编转,称左旋异构体(levorotary,L)或L型异构体。
像甘油醛这样具有旋光性差异的立体异构体又称为光学异构体(Cptical lsmer),常用D,L表示。
以甘油醛的两种光学异构体作对照,其他单糖的光学异构构与之比较而规定为D型或L 型。
差向异构体(epimer):又称表异构体,只有一个不对称碳原子上的基因排列方式不同的非对映异构体,如D-等等糖与D-半乳糖。
链状结构一般用Fisher投影式表示:碳骨架、竖直写;氧化程度最高的碳原子在上方,2、单糖的环状结构在溶液中,含有4个以上碳原子的单糖主要以环状结构。
单糖分子中的羟基能与醛基或酮基可逆缩合成环状的半缩醛(emiacetal)。
环化后,羰基C 就成为一个手性C原子称为端异构性碳原子(anomeric carbon atom),环化后形成的两种非对映异构体称为端基异构体,或头异构体(anomer),分别称为α-型及β-型头异构体。
环状结构一般用Havorth结构式表示:3 用FisCher投影式表示环状结构很不方便。
Haworth结构式比Fischer投影式更能正确反映糖分子中的键角和键长度。
转化方法:①画一个五员或六员环②从氧原子右侧的端基碳(anomerio carbon)开始,画上半缩醛羟基,在Fischer投影式中右侧的居环下,左侧居环上。
构象式:Haworth结构式虽能正确反映糖的环状结构,但还是过于简单,构象式最能正确地反映糖的环状结构,它反映出了糖环的折叠形结构。
3、几种重要的单糖的链状结构和环状结构(1) 丙糖:D-甘油醛二羟丙酮(2) 丁糖:D-赤鲜糖D-赤鲜酮糖(3) 戊糖:D-核糖D-脱氧核糖D-核酮糖D-木糖D-木酮糖(4) 己糖:D-葡萄糖(α-型及β型) D-果糖(5) 庚糖:D-景天庚酮糖4、变旋现象在溶液中,糖的链状结构和环状结构(α、β)之间可以相互转变,最后达到一个动态平衡,称为变旋现象。
从乙醇水溶液中结晶出的D—glucose称为α-D-(+)Glucose([α]20D=+113°),从吡啶溶液中结晶出的D—glucose称为β-D-(+)glucose([α]20D=+18.7°)。
将α-D-(+)葡萄糖与β-D-(+)葡萄糖分别溶于水中,放置一段时间后,其旋光率都逐渐转变为+52.7︒C。
原因就是葡萄糖的不同结构形式相互转变,最后,各种结构形式达到一定的平衡,其中α型占36%,β型占63%,链式占1%。
图5 葡萄糖的变旋5、构型与构象构型:分子中由于各原子或基团间特有的固定的空间排列方式不同而使它呈现出不同的较定的立体结构,如D-甘油醛与L-甘油醛,D-葡萄糖和L葡萄糖是链状葡萄糖的两种构型,α-D-葡萄糖和β-D-葡萄糖是环状葡萄糖的两种构型。
一般情况下,构型都比较稳定,一种构型转变另一种构型则要求共价键的断裂、原子(基4团)间的重排和新共价键的重新形成。
图3甘油醛的构型:构象:由于分子中的某个原子(基团)绕C-C单键自由旋转而形成的不同的暂时性的易变的空间结构形式,不同的构象之间可以相互转变,在各种构象形式中,势能最低、最稳定的构象是优势对象。
图1-3 吡喃型己糖构象6、构型与旋光性旋光性是分子中具有不对称结构的物质的一种物理性质。
显然,构型不同旋光性就不同。
构型是人为规定的,旋光性是实验测出的。
因此,构型与旋光性之间没有必然的对应规律,每一种物质的旋光性只能通过实验来确定。
二、单糖的物理化学性质(一)物理性质旋光性:是鉴定糖的一个重要指标甜度:以蔗糖的甜度为标准溶解性:易溶于水而难溶于乙醚、丙酮等有面溶剂(二)化学性质1、变旋图7-11在溶液中,糖的链状结构和环状结构(α、β)之间可以相互转变,最后达到一个动态平衡,称为变旋现象。
三者间的比例因糖种类而异。
只有链状结构才具有下述的氧化还原反应。
2、糖醛反应(与酸的反应)(1) Molish反应5Molish反应可以鉴定单糖的存在。
(2) Seliwannoff反应据此区分酮糖与醛糖。
还可利用溴水区分醛糖与酮糖。
3、氧化反应氧化只发生在开链形式上。
在氧化剂、金属离子如Cu2+、酶的作用下,单糖可以发生几种类型的氧化:图7、12醛基氧化:糖酸(aldonic acid)伯醇基氧化:醛酸(uronic acid)醛基、伯醇基同时氧化:二酸(alduric acid)能被弱氧化剂(如Fehhing试剂、Benedict试剂)氧化的糖称为还原性糖,所有的单糖都是还原性糖。
单糖氧化形成的羟基可以进一步形成环状内酯(Lactone)。
内酯在自然界中很普遍,如L-抗坏血酸(L-ascorbio acid),又称VC (Vitamcn c),就是D-葡萄糖酸的内酯衍生物。
分子量176.1,它在体内是一种强还原剂。
豚鼠(guinea pig)、猿(ape)和人不能合成Vc,从能合成Vc的肝脏微粒体中分离到合成Vc的三种酶,人和猿缺乏gulonolactone oxidase)。
缺乏抗坏血酸将导致坏血病(scurvy),龄龈(gum)、腿部等开始出血,肿胀,逐渐扩展到全身,柑橘类果实(citrus frait)中含有丰富的Vc。
4、还原反应单糖可以被还原成相应的糖醇(Sugar alcohol)。
D-葡萄糖被还原成D-葡萄糖醇,又称山犁醇(D-Sorbitol)。
糖醇主要用于食品加工业和医药,山犁醇添加到糖果中能延长糖果的货架期,因为它能防止糖果失水。
用糖精处理的果汁中一般都有后味,添加山犁醇后能去除后味。
人体食用后,山犁醇在肝中又会转化为果糖。
5、异构化在弱碱性溶液中,D-葡萄糖、D-甘露糖和D-果糖,可以通过烯醇式相互转化(enediol6intermediate)图7.15D-葡萄糖异构化为D-甘露糖后,由于其中的一个手性碳原子的构型发生变化,又称差向异构化(epimerization)。
6、酯化生物体中最常见也是最重要的糖酯是磷酸糖酯和硫酸糖酯。
磷酸糖酯及其衍生物是糖的代谢活性形式(糖代谢的中间产物)。
硫酸糖酯主要发现于结缔组织的蛋白聚糖中(Proteo glycan),由于硫酸糖酯带电荷,因此它能结合大量的水和阳离子。
葡萄糖的核苷二磷酸酯,如UDPG参与多糖的生物合成。
7、糖苷化单糖环状结构上的半缩醛羟基与醇或酚的羟基缩合失水成为缩醛式衍生物,通称为糖苷(glycosides)。
8、糖脎反应(亲核加成)糖脎反应发生在醛糖和酮糖的链状结构上。
糖脎易结晶,可以根据结晶的形状,判断单糖的种类。
三、重要的单糖四、重要的单糖衍生物1、糖醇2、糖醛酸单糖的伯醇基被氧化成-COOH。
动物体内有两种很重要的糖醛酸:α-D-葡萄醛酸和差向异构物β-L-艾杜糖醛酸,它们在结缔组织中含量很高。
glucuronic acid β-L-iduronate7 葡萄糖醛酸是肝脏内的一种解毒剂,它与类固醇、一些药物、胆红素(血红蛋白的降解物)结合增强其水溶性,使之更易排出体外。
3、氨基糖(糖胺,amino sugar, glycosamine)单糖的一个羟基(通常是C2位)被氨基取代。
常见的氨基糖有D-葡萄糖胺(D-glucosamine)和D-半乳糖胺(D-galactosamine)。
氨基糖的氨基还经常被乙酰化形成N-乙酰糖胺。
4、糖苷单糖的半缩醛羟基与其它分子的醇、酚等羟基缩合,脱水生成缩醛式衍生物,称糖苷Glycoside。
半缩醛部分是Glc,称Glc糖苷。
半缩醛部分是Gal,称Gal糖苷。
O糖苷、N糖苷、S糖苷。
糖苷物质与糖类的区别:糖是半缩醛,不稳定,有变旋;苷是缩醛,较稳定,无变旋。
糖苷大多数有毒。
5、脱氧糖重要的有6-脱氧D-甘露糖,L-岩藻糖(L-fucose)和2-脱氧D-核糖。
岩藻糖常见于一些糖蛋白中,如红细胞表面ABO血型决定簇。
第二节双糖和三糖双糖在自然界中含量也很丰富,它是人类饮食中主要的热源之一。
在小肠中,双糖必须在酶的作用下水解成单糖才能被人体吸收。
如果这些酶有缺陷的话,那么人体摄入双糖后由于不能消化它就会出现消化病。
未消化的双糖进入大肠,在渗透压的作用下从周围组织夺取水分(腹泻,diarrhea),结肠中的细菌消化双糖(发酵)产生气体(气胀和绞痛或痉孪)。
最常见的双糖消化缺陷是乳糖过敏,就是由于缺乏乳糖酶(Lactose),解决办法就是乳糖酶处理食物或避免摄入乳糖。