第四十三章 真核生物的基因表达调控
生物竞赛-生物化学-43真核生物的基因表达调控-杨荣武《生物化学原理(第二版)(三)》

生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
DNA甲基化与印记
生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
多个启动子的选择性使用
某些真核生物的基因不止一个启动子,例如,抗肌营
养不良蛋白有8个启动子,通过使用不同的启动子可转 录出不同长度的mRNA,它们经过翻译可产生不同性 质或功能的蛋白质产物。 人谷胱甘肽还原酶的基因具有两个启动子,这两个启 动子分别指导定位于细胞质和线粒体的谷胱甘肽还原 酶的合成。指导线粒体谷胱甘肽还原酶的启动子在指 导细胞质谷胱甘肽还原酶启动子的上游。显然,上游 启动子转录出来的mRNA要比下游启动子转录出来的 mRNA要长。分析它们的核苷酸序列以后发现,长 mRNA的起始密码子位置前移,因而会多翻译一段指 导进入线粒体的信号肽序列。
生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
DNA重排
B淋巴细胞在成熟过程Ig基因经历的重排 锥体虫主要的表面抗原基因发生的重排 酿酒酵母在交配类型转换过程中发生的基因 重排
生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
抗体基因多样性产生的分子机制
1. 2.
3. 4. 5.
生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
在染色质水平上的基因调控
组蛋白的共价修饰; 染色质重塑因子对染色质的作用; 组蛋白变体取代标准组蛋白。
生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
组蛋白不同化学修饰对基因表达的影响以及其他功能
修饰形式
乙酰化 单甲基化
修饰位点
生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
生物竞赛—生物化学原理(分子生物学)—南京大学杨荣武
真核生物的基因表达调控概述

真核生物的基因表达调控概述
真核生物基因在染色质活性、DNA水平、转录水平和翻译水平的表达调控特点。
答:真核基因组结构具有基因组结构庞大、单顺反子、含有大量重复序列、基因不连续性、非编码区较多等特点。
(1)染色质结构水平对基因表达调控:①常染色质或异染色质;②染色质的状态(活性或阻遏),紧密结构会抑制基因表达,解凝集结构利于基因表达;③可以通过对组蛋白结构的修饰来实现,有组蛋白翻译后的乙酰化、甲基化、磷酸化、泛素化等;④DNA水平的调控包括基因丢失、扩增、重排和移位等方式。
(2)转录水平的调控:①RNA聚合酶、转录因子等反式作用因子和顺式作用元件(启动子强弱、增强子、沉默子)相互作用对基因转录的调控;②同一基因转录起始位点的不同,导致在不同组织细胞中的基因表达差异。
(3)转录后的加工:转录后加工的多样性,包括①加尾和剪接;②多个5′端转录起始位点或剪接位点;③多个加多聚(A)位点和不同的剪接方式;④虽无剪接,但有多个转录起始位点或加多聚(A)位点等多种方式调控基因的表达。
(4)翻译水平的调控:①翻译起始因子eIF-4F 的磷酸化激活蛋白质的合成,eIF-2α 的磷酸化引起翻译起始受阻,降低蛋白质的生物合成水平;② mRNA 结构与翻译控制:mRNA5′端m7G 帽有增强翻译水平的作用,上游AUG 密码子的存在往往抑制下游开放读框的翻译效率;③起始AUG 上游序列对翻译效率的影响,如Kozak序列;④poly(A)尾增加翻译效率;⑤poly(A)尾中富含UA 序列抑制翻译。
(5)翻译后加工水平的调控:翻译的蛋白质还需要加工、修饰、折叠和分选后才具有功能。
综上所述,真核生物基因表达调控是一个十分复杂的过程。
简述真核生物基因表达调控过程

简述真核生物基因表达调控过程真核生物基因表达调控过程是指在真核生物细胞中,如何通过一系列的调控机制,将基因中的遗传信息转化为蛋白质,以实现细胞功能的正常发挥。
基因表达调控过程可以分为转录调控和转录后调控两个阶段。
在转录调控阶段,首先是在细胞核中进行转录。
细胞核中的DNA被RNA聚合酶酶识别并解链,形成单链mRNA。
但并不是所有基因都会被转录,细胞会根据需要选择性地进行转录。
这是通过转录因子的作用来实现的。
转录因子是一类能够与DNA特定序列结合的蛋白质,它们能够促进或抑制转录的进行。
转录因子的结合位点位于启动子区域,当转录因子结合到启动子区域时,会引发一系列的反应,包括启动RNA聚合酶的活性和引导其结合到合适位置上,从而促使转录的进行。
转录因子的表达受到多种因素的调控,如细胞内的信号分子、细胞周期等。
转录后调控是指在mRNA合成后,通过一系列的调控机制来决定其在细胞中的命运。
mRNA在合成后需要经过剪接、修饰和运输等过程。
剪接是指将mRNA中的内含子去除,将外显子进行连接的过程。
通过剪接的不同方式,可以生成不同的mRNA亚型,从而在翻译过程中产生不同的蛋白质。
修饰是指在mRNA上加上帽子和尾巴等化学修饰,这些修饰可以保护mRNA不被降解,并帮助mRNA与翻译机器结合。
运输是指mRNA离开细胞核,进入到细胞质中,进一步参与翻译过程。
这个过程受到RNA结合蛋白的调控。
在翻译过程中,mRNA被核糖体识别并翻译成蛋白质。
这个过程也受到多种调控机制的影响。
一方面,mRNA上的启动子序列会影响翻译的起始位置,从而决定蛋白质的翻译起始位点。
另一方面,mRNA的稳定性也会影响翻译的效率和蛋白质的表达水平。
mRNA 的稳定性受到RNA结合蛋白和非编码RNA的调控。
总的来说,真核生物基因表达调控过程是一个复杂而精细的调控网络。
通过转录调控和转录后调控的相互作用,细胞可以根据内外环境的需要,在不同的时空位置上产生不同类型的蛋白质,以实现细胞功能的正常发挥。
真核生物基因表达的调控

第10章真核生物基因表达的调控本章教学要求1.熟悉真核基因组的结构特点、真核生物在DNA水平、转录水平和翻译水平上基因表达调控的特点。
2.掌握以下概念:顺式作用元件、反式作用因子、启动子、增强子,熟悉沉默子、基本转录因子、特异转录因子。
3.了解转录因子的结构特点。
本章教学重点和难点1、真核生物在DNA水平和转录水平基因表达调控的特点。
2、转录因子的结构特点。
教学方法与手段讲授与交流互动相结合,采用多媒体教学。
授课内容10.1 真核生物基因表达调控的特点和种类一、真核生物基因表达调控的特点原核生物的调控系统就是要在一个特定的环境中为细胞创造高速生长的条件,或使细胞在受到损伤时,尽快得到修复,所以,原核生物基因表达的开关经常是通过控制转录的起始来调节的。
真核基因表达调控的最显著特征是能在特定时间和特定的细胞中激活特定的基因,从而实现"预定"的、有序的、不可逆转的分化、发育过程,并使生物的组织和器官在一定的环境条件范围内保持正常功能。
真核生物基因表达调控与原核的共同点:•基因表达都有转录水平和转录后的调控,且以转录水平调控为最重要;•在结构基因上游和下游、甚至内部存在多种调控成分,并依靠特异蛋白因子与这些调控成分的结合与否调控基因的转录。
真核生物基因表达调控与原核的不同点:1、真核基因表达调控的环节更多:转录与翻译间隔进行,具有多种原核生物没有的调控机制;个体发育复杂,具有调控基因特异性表达的机制。
2、真核生物活性染色体结构的变化对基因表达具有调控作用:DNA拓扑结构变化、DNA 碱基修饰变化、组蛋白变化;3、正性调节占主导,且一个真核基因通常有多个调控序列,需要有多个激活物。
二、真核生物基因表达调控的种类根据其性质可分为两大类:一是瞬时调控或称为可逆性调控,它相当于原核细胞对环境条件变化所做出的反应。
瞬时调控包括某种底物或激素水平的升降,及细胞周期不同阶段中酶活性和浓度的调节。
二是发育调控或称不可逆调控,是真核基因调控的精髓部分,它决定了真核细胞生长、分化、发育的全部进程。
分子遗传学章真核生物基因的表达调控
第22页/共74页
DNA Structure – Major and Minor Grooves
第23页/共74页
反式作用因子根据作用方式分为三类:
①通用转录因子。普遍存在的转录因子。如TATA box结合 因子 TFⅡD、GC box结合因子SP1 等。 ②组织特异性转录因子。与基因表达的组织特异性有很大 关系。 ③诱导性反式作用因子。活性能被特异的诱导因子所诱导。
位于转录起始位点附近,具有相对固定位置,且为转录起始 所必需的序列元件。
启动子一般模式:
核心启动子,TATA box 上游启动子成分(UPE),CAAT box等 TATA框控制转录的精确性,而UPE则控制转录的起始频率,启动子的 强度决定于UPE的数目和种类。
第13页/共74页
第14页/共74页
构型出现的结构单元,每间隔6个氨基酸出现一个亮 氨酸残基,能形成两性α-螺旋。两个具有这种结构的 因子接触后可借助侧链疏水性交错对插,形成稳定卷 曲螺旋结构的二聚体。
功能:可使碱性区与DNA的亲和力明显增加。
第32页/共74页
Leucine Zipper
第33页/共74页
④螺旋-环-螺旋 (helix-loop-helix,HLH)结构:
同源盒基因家族各基因间具有一相同的保守序 列,称为同源结构域。所含的至少二个α螺旋中形 成“转折”,第三个α螺旋与DNA大沟相互作用 是同源盒蛋白与DNA结合的主要力量 。
功能:调节与机体各部分正常发育或正常分化的 有关基因的表达。
第30页/共74页
同源结构域及其作用
第31页/共74页
③碱性亮氨酸拉链(basic 1eucine zipper,bLZ) 有些肽链C末端有一段30个氨基酸序列以α-螺旋
《分子生物学》真核生物基因表达的调控

4.具有细胞特异性或组织特异性
②富含谷氨酰胺结构域
SPl 是启动子 GC 盒的结合蛋白,除结合 DNA 的锌指结构以 外, SP1 共有 4 个参与转录活化的区域,其中最强的转录激活 域很少有极性氨基酸而谷氨酰胺含量却达 25 %左右。 酵母的 HAP1、HAP2和哺乳动物细胞中的Oct-1、Oct-2、Jun、AP2以及 血清应答因子(SRF)等都有同样的结构域。
• 真核生物细胞内绝大部分DNA是用于储存调控信息 的,用于合成蛋白质的信息仅占很小一部分。 • 真核生物能在特定时间和特定的细胞中激活特定的 基因,从而实现“预定”的、有序的、不可逆转的 分化发育过程,并使组织器官在一定环境条件下保 持正常功能。 • 真核生物基因表达的调控更加复杂、多层次、涉及 更多的蛋白质因子参与。
亮氨酸拉链
转录激活结构域
(transcription activating domain)
转录因子的 DNA 结合结构域本身并不具有调控转录活性的功 能,其转录活化功能是由另一种结构域,即转录激活结构域 所作用的。 转录激活域通常是由30~100氨基酸残基组成,根据氨基酸组 成的特点主要包括以下三种类型:
(3)染色体组蛋白的修饰
组蛋白磷酸化、乙酰化、泛素化以及H3组蛋白巯基化等现 象,会改变染色质的结构状态,调控基因转录的开关。 组蛋白的N端含有数个赖氨酸,它们的侧链基团在细胞正常 的pH范围内带正电荷,DNA的磷酸根带负电荷,能与之紧密 结合,使转录因子或RNA聚合酶很难与DNA结合进行转录。 组蛋白的赖氨酸残基被乙酰化后就不带电荷,与DNA的结合 力降低,有利于转录调控因子的结合。 组蛋白乙酰转移酶、组蛋白去乙酰酶
真核基因表达调控的特点

真核基因表达调控的特点尽管我们现在对真核基因表达调控知道还不多,但与原核生物比较它具有一些明显的特点。
真核基因表达调控的环节更多如前所述:基因表达是基因经过转录、翻译、产生有生物活性的蛋白质的整个过程。
同原核生物一样,转录依然是真核生物基因表达调控的主要环节。
但真核基因转录发生在细胞核(线粒体基因的转录在线粒体内),翻译则多在胞浆,两个过程是分开的,因此其调控增加了更多的环节和复杂性,转录后的调控占有了更多的分量。
图中标出了真核细胞在分化过程中会发生基因重排(gene rearrangement),即胚原性基因组中某些基因会再组合变化形成第二级基因。
例如编码完整抗体蛋白的基因是在淋巴细胞分化发育过程中,由原来分开的几百个不同的可变区基因经选择、组合、变化、与恒定区基因一起构成稳定的、为特定的完整抗体蛋白编码的可表达的基因。
这种基因重排使细胞可能利用几百个抗体基因的片段,组合变化而产生能编码达108种不同抗体的基因,其中就有复杂的基因表达调控机理。
此外,真核细胞中还会发生基因扩增(gene amplification),即基因组中的特定段落在某些情况下会复制产生许多拷贝。
最早发现的是蛙的成熟卵细胞在受精后的发育程中其rRNA 基因(可称为rDNA)可扩增2000倍,以后发现其他动物的卵细胞也有同样的情况,这很显然适合了受精卵其后迅速发育分裂要合成大量蛋白质要求有大量核糖体的需要。
又如MTX (methotrexate)是叶酸的结构类似物,能竞争性抑制细胞对叶酸的还原利用,因而对细胞有毒性,但当缓慢提高MTX浓度时,一些哺乳类细胞会对含有利用叶酸所必需的二氢叶酸还原酶(dihydrofolate reductase,DHFR)基因的DNA区段扩增40-400倍,使DHFR的表达量显著增加,从而提高对MTX的抗性。
基因的扩增无疑能够大幅度提高基因表达产物的量,但这种调控机理至今还不清楚。
真核基因的转录与染色质的结构变化相关真核基因组DNA绝大部分都在细胞核内与组蛋白等结合成染色质,染色质的结构、染色质中DNA和组蛋白的结构状态都影响转录,至少有以下现象:染色质结构影响基因转录细胞分裂时染色体的大部分到间期时松开分散在核内,称为常染色质(euchromatin),松散的染色质中的基因可以转录。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
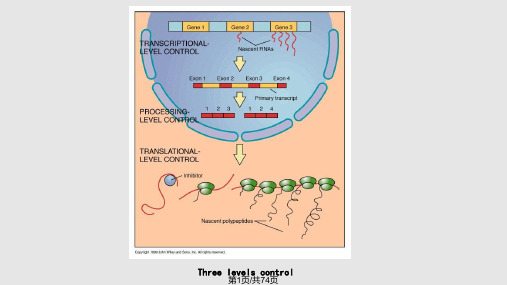
五. 在翻译及翻译后加工水平上的调控
真核生物与原核生物在 调控机制上的主要差别
调控的原因:原核生物基因表达调节的目的是为了更 有效和更经济地对环境的变化做出反应,而多细胞真 核生物基因表达调节的主要目的是细胞分化,它需要 在不同的生长时期和不同的发育阶段具有不同的基因 表达样式; 调控的层次:原核生物基因表达调控主要集中在转录 水平,但真核生物基因表达的转录后水平调节与其在 转录水平上的调节各占“半壁江山”,而某些调控层 次是真核生物特有的,比如染色质水平、RNA后加 工水平和mRNA运输等; 调控的手段:原核生物绝大多数的基因组织成操纵子, 但真核生物一般无操纵子结构。
不是所有的转录因子都能够与DNA结合,也
不是所有的转录因子都是激活基因的转录。
转录因子的结构
1. DNA结合结构域,直接与顺式作用元件结合的转录因 子都具有此结构域。转录因子通常使用此结构域之中 的特殊α-螺旋与顺式作用元件内的大沟接触,通过螺 旋上的特殊氨基酸残基的侧链与大沟中的特殊碱基之 间的次级健(主要是氢键)相互识别而产生特异性。 许多转录因子在此结构域上富含碱性氨基酸,这可能 有利于和DNA骨架上带负电荷的磷酸根发生作用; 2. 效应器结构域,这是转录因子调节转录效率(激活或 阻遏)、产生效应的结构域; 3. 多聚化结构域,此结构域的存在使得转录因子之间能 够组装成二聚体或多聚体。下面将集中介绍前两种结 构域,特别是DNA结合结构域。
第四十三章 真核生物 的基因表达调控
提纲
一. 在染色质水平上的调控 二. 在DNA水平上的调控 三. 在转录水平上的调控
1. 顺式作用元件 2. 转录因子 3. 转录水平调控的实例
四. 在转录后加工及运输水平上的调控
1. 2. 3. 4.
可变剪接 可变加尾 组织特异性RNA编辑 在mRNA运输水平上的调控
真核生物在转录水平进行基因表达调控的主要方式
激活蛋白激活基因表达的可能机制
绝缘子作用的分子模型
ICR绝缘子对小鼠Igf2和H19基因表达的控制
转录因子
转录因子是泛指除RNA聚合酶以外的一系列 参与DNA转录和转录调节的蛋白质因子,分 为基础转录因子和调节转录因子,其中前者 专指从核心启动子开始进行精确转录所必需 的一组最低数量的蛋白质的总称,其它转录 因子参与转录的调节,激活或阻遏基因的转 录,因此属于调控转录因子。
阻遏蛋白与沉默子结合以后抑制基因表达的三种方式
转录水平调控的实例
1. 酵母细胞半乳糖代谢相关基因的表达调控
2. 热休克蛋白的基因表达调控 3. 激素诱导的基因表达调控 4. 金属硫蛋白的基因表达调控 5. 生物发育过程中的组织特异性基因表达
酵母半乳糖代谢相关基因的表达调控
酵母参与半乳糖代谢的3个基因Gal1、Gal7和Gal10的
组蛋白的修饰与染色质构象变化的关系
酵母HO基因的转录激活
组蛋白变体对基因表达的影响
所有真核细胞内的组蛋白都具有五种标准的形式。这
五种标准的形式由多拷贝基因编码,在进化上高度保 守。但细胞内还存在与这五种标准形式相对应的不同 变体。这些变体的氨基酸序列与相应的标准组蛋白在 一级结构上差别可能很小,也可能很大,表达的方式 可能是组成型,也可能具有组织特异性。编码这些变 体的基因与相应的标准组蛋白的基因并不呈等位基因 的关系,且通常是单拷贝。 如果一种变体代替了相应的标准组蛋白参入到一个或 多个核小体结构之中,就等于在染色质上创造了一些 “特区”,或者在染色质上打上了特殊的标记。这些 特化的区域或者特殊的标记可能具有多种不同的功能, 但最重要的功能可能是调节特定的基因表达
激活蛋白的效应器结构域即是激活基因转录的激活 结构域。转录因子上的DNA结合结构域只能让转 录因子与特定的顺式作用元件结合,以“锁定”被 调节的目标基因,激活基因表达的功能由转录因子 上专门的激活结构域承担。 已发现三种常见的激活结构域:(1)酸性结构 域——富含酸性氨基酸残基,但常有1个疏水的氨 基酸残基镶嵌在其中。(2)富含Gln结构域;(3) 富含Pro结构域。
抗体轻链基因的重排机制
抗体重链基因重排、转录、后加工和翻译
D基因和J基因连接的多样性
锥体虫主要表面抗原基因的重排
锥体虫是由一种叫采采蝇的吸血蝇传播的血液寄生
性原生动物,它是非洲昏睡病的病原体。在应付宿 主细胞的免疫系统方面,锥体虫会不断而有序地改 变其表面抗原,以逃避免疫系统对它的攻击。 锥体虫的表面抗原性质与其可变的表面糖蛋白 (VSG),由它形成一种保护性的外被。锥体虫基 因组约有1000拷贝的VSG基因,但并不是所有的 VSG基因都能表达。只有处于表达偶联位点的拷贝 (ELC)才有可能被转录,其它位点上的VSG被称 为基本拷贝(BC),无转录活性。
DNA重排
B淋巴细胞在成熟过程Ig基因经历的重排 锥体虫主要的表面抗原基因发生的重排 酿酒酵母在交配类型转换过程中发生的基因 重排
抗体基因多样性产生的分子机制
1. 2.
3. 4. 5.
VJ和VDJ的重组连接 连接的多样性,重排过程中的连接是不精确 的,这种“故意”的不精确连接可导致氨基 酸的变化或缺失,从而影响到抗原结合位点 的结构,实际上它是抗体上出现高变区的原 因 插入的多样性,TdT作用导致DJ连接或V-DJ 连接中随机插入若干个核苷酸到V基因、D基 因或J基因的末端 体细胞超突变,主要发生在V基因 选择性剪接和选择性加尾
在转录水平上的基因表达调控
真核生物的蛋白质基因的转录除了启动子、RNA聚
合酶II和基础转录因子以外,还需要其它顺式作用元 件和反式作用因子的参与。 参与基因表达调控的主要顺式作用元件有:增强子、 沉默子、绝缘子和各种反应元件;参与基因表达调控 的反式作用因子也称为转录因子,它们包括激活蛋白、 辅激活蛋白、阻遏蛋白和辅阻遏蛋白。 激活蛋白与增强子结合激活基因的表达,而阻遏蛋白 与沉默子结合,抑制基因的表达。辅激活蛋白缺乏 DNA结合位点,但它们能够通过蛋白质与蛋白质的 相互作用而行使功能,作用方式包括:招募其它转录 因子和携带修饰酶(如激酶或HAT)到转录复合物而 刺激激活蛋白的活性;辅阻遏蛋白也缺乏DNA结合 位点,但同样通过蛋白质与蛋白质的相互作用而起作 用,作用机理包括:掩盖激活蛋白的激活位点、作为 负别构效应物和携带去修饰酶去中和修饰酶(如磷酸 酶或HDAC)的活性。
DNA甲基化导致基因转录活性丧失的三种可能机制
DNA印记
就许多真核生物的某些基因而言,在一个发育的个体之中,两个
等位基因中只有一个才表达,而哪一个被表达是由亲代决定的: 有的是来自父本的基因才能表达,如IGF-2基因,有的是来自母本 的基因才表达。这种由亲代决定的等位基因的选择性表达的机制 被称为印记。印记的手段是甲基化,不表达的等位基因的CpG岛 上的C被甲基化,表达的等位基因的CpG岛上的C没有被甲基化。 在配子形成时期,许多基因就开始以性别特异性的方式进行甲基 化反应,性别特异性的甲基化导致胚胎内来自不同亲本的等位基 因的区别表达。 尽管在胚胎发育的早期,其胚性细胞内的甲基化样式重新设定, 需要经历一波又一波的去甲基化和新甲基化反应,但这并不影响 到印记基因的甲基化。 在个体发育的整个阶段,由于组织特异性甲基化酶的作用,不同 类型的细胞其甲基化样式会发生改变,但被印记的基因始终得到 维持,这要归功甲基化与印记
多个启动子的选择性使用
某些真核生物的基因不止一个启动子,例如,抗肌营
养不良蛋白有8个启动子,通过使用不同的启动子可转 录出不同长度的mRNA,它们经过翻译可产生不同性 质或功能的蛋白质产物。 人谷胱甘肽还原酶的基因具有两个启动子,这两个启 动子分别指导定位于细胞质和线粒体的谷胱甘肽还原 酶的合成。指导线粒体谷胱甘肽还原酶的启动子在指 导细胞质谷胱甘肽还原酶启动子的上游。显然,上游 启动子转录出来的mRNA要比下游启动子转录出来的 mRNA要长。分析它们的核苷酸序列以后发现,长 mRNA的起始密码子位置前移,因而会多翻译一段指 导进入线粒体的信号肽序列。
锥体虫主要表面抗原基因的重排
DNA甲基化、羟甲基化与基因表达
DNA甲基化是一种复制后加工反应。真核生物DNA的 甲基化位点主要是CpG 二核苷酸之中的C,甲基供体 为SAM,由DNA甲基化酶催化,C甲基化后成为5-甲 基胞嘧啶。 CpG序列在基因组中的分布并不均一,它们通常成簇 存在,形成所谓的CpG岛。每一个CpG岛长度在1~2kb 左右,通常位于基因的启动子附近或内部,并有可能 延伸到基因的第一个外显子。 甲基化样式与基因表达有关:活性基因的CpG岛处于 去甲基化状态,非活性基因的CpG岛处于甲基化状态。 管家基因的CpG岛在所有的细胞都呈去甲基化状态, 而组织特异性基因的CpG岛只是在表达它的细胞才处 于去甲基化状态。 羟甲基化修饰也是发生在DNA 的C 上也是调节基因的 表达,特别是在哺乳动物的脑细胞和胚胎干细胞中。
表达受到半乳糖的诱导,这3个基因虽然靠得很近,但 并不象原核生物那样组成操纵子。 在Gal1和Gal10之间有一段上游激活子序列(UAS), 其一致序列是CGG-N11-CCG,为转录因子Gal4蛋白的 结合位点。 在没有半乳糖时,Gal4蛋白二聚体与Gal80蛋白组成的 复合物与UAS结合,这时Gal80与Gal4的激活结构域结 合,而阻止Gal4去激活Gal基因的转录;在有半乳糖时, 半乳糖的代谢物与Gal80上的结合位点结合,诱使 Gal80与Gal4解离并与细胞质中的Gal3结合,Gal4的激 活结构域因此暴露出来,于是,它能将一系列促进和 参与转录的蛋白质,包括SAGA(具有HAT活性)、 介导蛋白、基础转录因子和RNA聚合酶II招募到Gal基 因的启动子周围,激活Gal基因的表达
泛酰化 酵母H2BK123 哺乳动物H2B120
转录阻遏
转录激活 减数分裂
组蛋白乙酰化或去乙酰化与染色质转录活性的关系