利用温敏致死性状专养雄蚕的研究进展
季铵盐∕两性壳聚糖改性真丝织物及其协同增效作用

第31卷㊀第6期2023年11月现代纺织技术AdvancedTextileTechnologyVol.31ꎬNo.6Nov.2023DOI:10.19398∕j.att.202212030季铵盐∕两性壳聚糖改性真丝织物及其协同增效作用杨㊀晟1ꎬ徐兆梅2ꎬ马廷方2ꎬ付飞亚1ꎬ刘向东1ꎬ姚菊明1(1.浙江理工大学材料科学与工程学院ꎬ杭州㊀310018ꎻ2.杭州万事利丝绸数码印花有限公司ꎬ杭州㊀310020)㊀㊀摘㊀要:化学改性是进一步拓展蚕丝用途的重要技术手段ꎬ但传统单一物质改性难以同时实现绿色和高效改性目的ꎮ使用水溶性羧化壳聚糖(CMC)和2ꎬ3 ̄环氧丙基三甲基氯化铵(GTA)作为反应原料经串联反应对真丝织物(SF)进行化学改性ꎮ期待CMC为GTA提供更多的反应位点ꎬGTA有助于稳定CMC并弥补其功能不足ꎮ对比分析了SF㊁SF∕CMC㊁SF∕GTA以及SF∕CMC∕GTA四种样品的形貌㊁结构㊁物化性能等ꎮ结果表明:串联反应在水溶剂中80ħ条件下可成功实施ꎻSF∕CMC∕GTA(1.5mV)的Zeta电位相对SF(-26.3mV)明显增加ꎬ其透气率和吸水率均最大ꎬ分别达到272g∕(m2 d)和326%ꎻSF∕CMC∕GTA抗菌效果最明显ꎬ对金黄色葡萄球菌和大肠杆菌的抗菌率均在99.9%以上ꎬ上染率相对SF提高了55倍ꎮ研究结果可为丝绸的绿色染整加工提供新的科学依据和技术路径ꎮ关键词:真丝织物ꎻ串联反应ꎻ协同增效ꎻ抗菌性能ꎻ染色性能中图分类号:TS146㊀㊀㊀文献标志码:A㊀㊀㊀文章编号:1009 ̄265X(2023)06 ̄0017 ̄11收稿日期:20221221㊀网络出版日期:20230321基金项目:浙江省重点研发计划(2121069 ̄J)ꎻ浙江省 高层次人才特殊支持计划 杰出人才项目(2021R51003)ꎻ浙江省分析测试项目(LGC22E030006)ꎻ浙江省清洁染整技术研究重点实验室开放基金项目(QJRZ2110)作者简介:杨晟(1996 )ꎬ男ꎬ山西运城人ꎬ硕士研究生ꎬ主要从事天然高分子改性方面的研究ꎮ通信作者:付飞亚ꎬE ̄mail:fufar@163.com㊀㊀蚕丝具有优良的服用性㊁生物相容性和生物可降解特性ꎬ在纺织㊁生物医用㊁食品等领域被广泛应用[1]ꎮ化学改性是拓展蚕丝用途的重要技术手段[2 ̄3]ꎬ但蚕丝的蛋白质属性对化学原料和反应路径均有较高的要求ꎬ例如高温㊁有毒试剂㊁强酸∕碱易造成蛋白质变性等[4 ̄5]ꎮ探索蚕丝纤维的绿色化学改性方法推进其高值化利用的重要技术途径ꎮ壳聚糖是自然界第二丰富的多糖生物质[6 ̄7]ꎬ具有优异生物相容性和生物活性ꎬ在纺织领域经常用作织物染色㊁印花和抗皱整理剂[8]ꎮ但壳聚糖存在大量分子间和分子内氢键ꎬ导致其在水中难以溶解ꎬ因此将壳聚糖方便㊁绿色且牢固地键连到真丝织物表面仍是挑战[9 ̄10]ꎮFerrero等[11]采用酒石酸㊁二甲基丙烯酸㊁环氧树脂等交联剂实现壳聚糖与蚕丝的化学接枝ꎬ而壳聚糖在蚕丝表面分布不均匀ꎬ大量交联剂的使用破坏了蚕丝原有良好的物化性能ꎮDavarpanah等[12]利用丁二酸酐和邻苯二甲酸酐对蚕丝进行酰化作用ꎬ后通过化学接枝壳聚糖ꎬ但反应需在有机溶剂中进行ꎬ反应条件要求高ꎬ过程复杂ꎮ水溶性羧化壳聚糖(CMC)是一种重要的水溶性壳聚糖衍生物ꎬ同样具备良好的生物相容性和生物降解性ꎬ在化工㊁环保㊁保健品方面也有广泛的应用[13]ꎮ由于CMC同时包含大量活性氨基和羧基[14]ꎬ可通过化学键共价接枝在真丝织物表面ꎬ提升稳定性ꎬ但其抗菌活性受环境pH值㊁聚合度等影响较大[15 ̄16]ꎮ张伟[17]㊁Lim等[18]尝试直接使用壳聚糖季铵盐对真丝织物进行改性ꎬ过程需要使用交联剂如柠檬酸进行固化ꎬ对真丝织物的白度㊁力学性能存在负面影响ꎮ本文提出采用水溶性的CMC和2ꎬ3 ̄环氧丙基三甲基氯化铵(GTA)对真丝织物(SF)进行串联化学改性的新方法ꎮ使用扫描电镜㊁红外光谱仪㊁X射线电光子能谱等测试手段对比分析不同真丝织物的形貌㊁结构㊁物化性能变化规律ꎮ此外ꎬ论文将研究样品的抗菌活性和染色性能ꎬ阐明CMC和GTA的协同增效作用ꎮ本方法过程简单㊁无有机溶剂㊁反应条件温和且效果明显ꎬ可为真丝织物的功能化改性提供新的技术途径ꎮ1㊀实㊀验1.1㊀实验材料真丝织物(60g∕m2(14姆米)真丝斜纹绸ꎬ杭州万事利丝绸数码印花有限公司)ꎬ羧化壳聚糖((C6H11NO4)nꎬ15kDaꎬBRꎬ水溶性ꎬ阿拉丁有限公司)ꎬ2ꎬ3 ̄环氧丙基三甲基氯化铵(C6H14ClNOꎬȡ95%ꎬ麦克林生化有限公司)ꎬ大肠杆菌(E.coliꎬATCC1555)㊁金黄色葡萄球菌(S.aureusꎬATCC547)ꎬ均购于上海鲁微科技有限公司ꎮ1.2㊀实验方法1.2.1㊀SF∕CMC和SF∕GTA的制备将脱除丝胶的真丝织物(SFꎬ5.00cmˑ5.00cmꎬ5片)以浴比1ʒ50浸入CMC水溶液(100mLꎬ质量分数为2%)中ꎬ在60ħ油浴锅中搅拌2hꎬ使CMC与SF纤维充分接触ꎬ之后将处理的SF于80ħ热处理3hꎬ再用去离子水充分清洗3次ꎬ经60ħ烘干后得到SF∕CMCꎮ另制备了仅负载GTA的真丝织物作为对比样ꎮ将SF(5.00cmˑ5.00cmꎬ5片)浸入GTA水溶液(100mLꎬ质量分数为8%)中ꎬ在80ħ油浴锅中搅拌反应10hꎬ再用去离子水充分清洗3次ꎬ经60ħ烘干后得到SF∕GTAꎮ1.2.2㊀SF∕CMC∕GTA的制备将SF∕CMC改性真丝织物(5.00cmˑ5.00cmꎬ5片)浸入GTA水溶液(100mLꎬ质量分数为8%)中ꎬ在80ħ油浴锅中搅拌反应10hꎬ再用去离子水充分清洗3次ꎬ经60ħ烘干后得到SF∕CMC∕GTAꎮ图1为串联改性法制备SF∕CMC∕GTA的过程示意图ꎮ首先CMC通过酰胺化反应接枝在真丝织物表面ꎬ富含氨基的CMC为真丝织物提供了更多的反应位点ꎬ提高真丝织物表面的反应活性ꎮGTA通过其环氧基与SF∕CMC中部分氨基发生开环反应ꎬ从而将含有阳离子基团的GTA接枝在真丝织物表面ꎮGTA基团的引入进一步提高了SF表面阳离子的数目ꎬ有望改善其抗菌活性和染色性能ꎮ同时ꎬGTA基团通过与SF表面的羧基㊁氨基㊁羟基等产生氢键相互作用ꎬ有利于进一步稳定CMC基团ꎮ整个制备过程不使用任何有机溶剂ꎬ反应条件温和(80ħ)ꎬ无有害小分子副产物生成ꎬ是绿色㊁经济㊁便捷的化学改性技术手段ꎮ图1㊀SF∕CMC∕GTA的制备过程示意Fig.1㊀SchematicdiagramofthepreparationprocessofSF∕CMC∕GTA1.3㊀基本测试采用扫描电子显微镜(SEM∕Ultra55ꎬZeissꎬ德国)对SF㊁SF∕CMC㊁SF∕GTA㊁SF∕CMC∕GTA的表面形貌进行观察分析ꎮ采用傅里叶红外光谱仪(FTIR∕TENSORⅡꎬBrochꎬ德国)再配备衰减全反射仪(ATR)分析不同样品的化学结构ꎬ扫描范围500~4000cm-1ꎮ采用二维X射线衍射仪(XRD∕D8Discoverꎬ德国)研究样品晶体结构ꎬ扫描速率2(ʎ)∕minꎬ扫描范围5ʎ~45ʎꎮ采用X射线电光子能谱(XPS∕AXSSꎬ德国)分析样品表面化学成分ꎮ采用热重分析仪(TG∕SDTA851ꎬ美国)分析热稳定性ꎬ测试温度范围30~800ħꎬ升温速率为10ħ∕minꎬ空气气氛ꎮ81 现代纺织技术第31卷采用紫外可见分光光度计(UH4150ꎬ日本)测量染料吸收率ꎬ扫描范围420~700nmꎮ采用通用材料试验机(Instron5943ꎬ美国)测试样品的机械拉伸性能ꎮ采用固体表面ZETA电位仪(DLS∕SURPASSꎬ奥地利)分析SF㊁SF∕CMC㊁SF∕GTA及SF∕CMC∕GTA表面Zeta电位ꎬ测试pH范围为3~10ꎮ1.4㊀抗菌性能测试按照改进的AATCC100-1999«纺织材料中抗菌整理剂的鉴定»方法来测试真丝织物样品对大肠杆菌(E.coli)和金黄色葡萄球菌(S.aureus)的抗菌率ꎮ抗菌实验前ꎬ将待测真丝织物样品在紫外灯下灭菌30minꎮ抗菌测试中ꎬ首先将样品(1.5cmˑ1 5cm)上滴入20μL的标准菌液ꎬ培养1hꎮ取出样品放入到含有5mLPBS缓冲溶液(4ˑ106CFU∕mL)的试管中ꎬ恒温培养箱中震荡10minꎮ最后最后ꎬ将100μL的PBS溶液涂在LB琼脂平板上ꎬ在37ħ下培养24hꎮ抗菌率(BR)按照式(1)计算:BR∕%=B-ABˑ100(1)式中:A和B分别是待测真丝织物样品和原始真丝织物做完抗菌率实验后固体培养基中菌落的个数ꎮ每组样品均重复测试3次取平均值ꎮ1.5㊀水气透过率测试根据ASTME ̄96方法ꎬ进行了水气透过率实验ꎮ在直径为15mm的试管中装有蒸馏水ꎬ使得水面离试管口约3mmꎬ再用待测真丝织物封住试管口ꎬ用橡皮筋栓牢ꎮ记录24h前后试管中水质量的变化ꎮ用式(2)计算真丝织物的透气性:T=m0-maπ r2(2)式中:T是透气率为每天每平方米的真丝织物表面透出去水的质量ꎬg∕(m2 d)ꎻm0是测试前试管内水的质量ꎬgꎻma是测试后试管内水的质量ꎬgꎻr是试管的内壁半径ꎬmꎮ每组样品重复测试3次取平均值ꎮ1.6㊀吸水性测试将待测真丝织物样品浸没在蒸馏水中10minꎬ然后悬挂10minꎬ直到样品不滴水ꎮ记录待测真丝织物样品吸收水前后质量的变化ꎬ按式(3)计算真丝织物样品的吸水性:w∕%=wa-w0w0ˑ100(3)式中:w是真丝织物的吸水率ꎬ%ꎻw0是吸水前真丝织物的质量ꎬgꎻwa是吸水后真丝织物的质量ꎬgꎮ每组样品均重复测3次取平均值[19]ꎮ1.7㊀染色性能测试活性染料是一种分子结构上带有活性基团的水溶性染料ꎬ能与蛋白质纤维上的氨基发生共价键结合ꎮ本实验通过浸渍法染色ꎬ首先用去离子水为参照液ꎬ取活性红3BF染料配制成样品注入比色皿中ꎬ使用可见光分光光度计在波长为420~700nm范围内扫描得到相对应的吸光度ꎮ经过吸光度与扫描波长ꎬ找出染料的最大吸收波长λmaxꎮ然后分别将不同真丝织物在25ħ下浸入染料中搅拌10min完成上染ꎬ对不同真丝织物染色前染液㊁染色后残液取样ꎬ测其吸光度ꎬ按式(4)计算真丝织物样品的上染率:R∕%=1-A1A0æèçöø÷ˑ100(4)式中:R为真丝织物样品的上染率ꎬ%ꎻA0为染色原液的吸光度ꎻA1为染色后残液的吸光度ꎮ每组样品均重复测3次取平均值ꎮ2㊀结果与讨论2.1㊀改性真丝织物的形貌分析图2为真丝织物经CMC㊁GTA处理前后的SEM照片ꎮ如图2所示ꎬ改性前的SF纤维具有光滑㊁均匀的表面ꎮ经过化学改性后ꎬ通过观察样品的低倍SEM图像ꎬ发现SF∕CMCꎬSF∕GTA和SF∕CMC∕GTA形貌没有明显变化ꎬ表明化学改性对真丝织物整体结构影响较小ꎮ高倍SEM图像显示ꎬ从SF∕CMCꎬSF∕GTA到SF∕CMC∕GTAꎬ纤维表面呈现出越来越粗糙的趋势ꎬ造成这一现象的原因为:化学反应破坏了组成蚕丝的蛋白微纤之间的氢键[20]ꎬ进而导致蚕丝表面有部分纤丝出现ꎮ2.2㊀改性真丝织物的结构分析图3(a)㊁图3(b)为SF和不同改性SF样品的ATR ̄FTIR光谱图ꎬ图3(a)中ꎬ1620㊁1510㊁1230cm-1处的吸收峰分别对应于丝素蛋白分子结构中酰胺Ⅰ㊁酰胺Ⅱ㊁和酰胺Ⅲ的C O伸缩振动峰[21 ̄22]ꎮ除上述特征峰之外ꎬ1373cm-1和1328cm-1处出现对应于CMC的 COO 对称伸缩振动和 OH的面内弯曲振动吸收带ꎬ在1158cm-1附近存在较强氧化壳聚糖C O C 桥式 不对称伸缩吸收峰ꎮ1739cm-1附近出现新的吸收峰ꎬ这是由于引入CMC后 COOH基团的特征吸收峰ꎬ此峰在SF的红91 第6期杨㊀晟等:季铵盐∕两性壳聚糖改性真丝织物及其协同增效作用图2㊀SF和不同改性SF样品的SEM图Fig.2㊀SEMimagesofSFanddifferentmodifiedSFsamples图3㊀SF和不同改性SF样品的ATR ̄FTIR和XRD谱图Fig.3㊀ATR ̄FTIRandXRDspectrumofSFanddifferentmodifiedSFsamples外曲线中没有出现ꎬ说明了SF与CMC发生了酰胺交联反应[23]ꎮSF∕GTA和SF∕CMC∕GTA在1483cm-1处所均出现了新的吸收峰ꎬ这应归因于CH3 N+的伸缩振动[24]ꎬ另外ꎬ相对于SF∕GTAꎬ1483cm-1特征峰在SF∕CMC∕GTA样品中更为明显ꎬ由此也说明CMC的引入有助于后续键接GTA基团ꎮ图3(b)中GTA和SF∕CMC∕GTA∕N(SF∕CMC改性织物与GTA未发生热处理的吸附织物)都在928cm-1处(C O C拉伸振动)出现环氧环的特征峰[25]ꎬ但此峰在SF和SF∕CMC∕GTA曲线中没有出现ꎬ说明环氧环与氨基发生反应ꎬGTA成功键接到真丝织物上ꎮ图3(c)为 02 现代纺织技术第31卷SF和不同改性SF样品的XRD谱图ꎬ2θ=20.8ʎ处为蚕丝的明显特征衍射峰ꎬ是蛋白质分子的β ̄折叠[23]ꎬ9.38ʎ和20.8ʎ归属于丝素蛋白SilkⅡꎬ24.5ʎ和29.8ʎ归属于丝素蛋白SilkⅠ[26]ꎮSF∕CMC㊁SF∕GTA㊁SF∕CMC∕GTA谱图中上述4个特征衍射峰的位置无明显变化ꎬ表明CMC和GTA的引入对真丝织物的晶体结构无明显影响ꎬ该结果与SEM图像结论一致ꎮSF∕GTA在2θ=20.8ʎ处的衍射强度略大于其他样品ꎬ这是由于GTA小分子更容易进入纤维内部ꎬ更多地破坏了其非晶区ꎮXPS可以分析样品的化学组成和各元素的化学状态ꎮ图4(a)为SF和SF∕CMC∕GTA的XPS全谱ꎬ两者均在结合能531㊁399eV和284eV处分别出现了O1s㊁N1s㊁C1s的信号峰ꎮ图4(b)和图4(c)分别为SF和SF∕CMC∕GTA的高分辨N1s分峰拟合谱图ꎬ前者在结合能399eV处出现了 CONH 的信号峰ꎮ不同的是ꎬ后者在结合能402eV处出现了新的信号峰ꎬ归属于反应引入的GTA基团中的CH3 N+[27]ꎮ图4(d)和图4(e)分别为SF和SF∕CMC∕GTA的C1s高分辨率拟合谱图ꎮSF的C1s峰被分峰为3个峰ꎬ其对应的峰分别为287.38eV( C O)㊁285eV(C N)和283eV(C C)ꎬ同时ꎬSF∕CMC∕GTA的C1s峰分别出现在287.49eV( C O)㊁285eV(C N)和283eV(C C)[28 ̄29]ꎮ在改性真丝织物中 C O峰的位置发生了偏移ꎬ这是由于CMC与真丝织物发生的酰胺化反应所导致的[30]ꎬ并且C N峰面积稍大ꎬ这应该与引入的CMC和GTA中均有C N键有关ꎮ㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀12 第6期杨㊀晟等:季铵盐∕两性壳聚糖改性真丝织物及其协同增效作用图4㊀SF和SF∕CMC∕GTA的XPS谱图Fig.4㊀XPSspectrumofSFandSF∕CMC∕GTA图5(a)㊁图5(b)和图5(c)分别为SF和不同改性SF样品的应力 ̄应变曲线㊁热重曲线(TG)和热重微分曲线(DTG)ꎮ图5(a)应力应变曲线所示ꎬSF的拉伸断裂强度和断裂伸长率分别为67.2MPa㊁25.1%ꎮ而SF∕CMC㊁SF∕GTA㊁SF∕CMC∕GTA改性真丝织物的拉伸断裂强度分别为63.5㊁57.8㊁62.7MPaꎬ断裂伸长率为23.1%~26.3%ꎬ由此可以看出不同真丝织物的拉伸强度相近ꎬ表明CMC和GTA的接枝对真丝织物的力学性能无明显影响ꎮ该现象明显优于寇爱静[31]㊁Ferrero等[32]的类似工作ꎬ以甲基丙烯酸羟基乙酯与三乙二醇二甲基丙烯酸酯在二甲基亚砜等有机溶剂的作用下对真丝织物进行接枝处理ꎬ但改性后织物的断裂强度和断裂伸长率均下降5%~15%ꎬ导致其力学性能严重恶化ꎮ图5(b)中的TG曲线所示ꎬSF在低于100ħ范围内有失重峰ꎬ该现象是由织物中水分的蒸发所导致ꎬ此外ꎬ在200~700ħ有1个失重阶段ꎬ在300ħ左右达到最大分解速率ꎬ这一阶段可归因于丝素蛋白分子间侧链和主链遭到破坏引起的分解失重[33]ꎮ同时从图5(c)DTG中可以观察到ꎬSF㊁SF∕CMC㊁SF∕GTA㊁SF∕CMC∕GTA的主失重峰依次是326㊁324㊁329㊁327ħꎮ各样品的TG㊁DTG曲线无明显变化ꎬ真丝织物经CMC㊁GTA处理前后的热稳定性不变ꎮ图6为SF和不同改性SF样品的Zeta电位测试曲线ꎬ表面电荷特性可通过Zeta电位进一步表征ꎮ如图6所示ꎬ在pH为3~10的范围内ꎬSF表面的平均电荷为-26.3mVꎬSF∕CMC和SF∕GTA的平均表面电荷分别为-22.8㊁-9.5mVꎬ这说明经GTA改性后的真丝织物表面电负性远小于未改性真丝织物ꎬ这归因于接枝在真丝织物表面GTA所带的正电荷氨基ꎬ提高了SF表面阳离子的数目ꎬ增强了其表图5㊀SF和不同改性SF样品的应力 ̄应变曲线㊁TG曲线和DTG曲线Fig.5㊀Stress ̄straincurvesꎬTGcurvesandDTGcurvesofSFanddifferentmodifiedSFsamples22 现代纺织技术第31卷面的正电性ꎮ而SF∕CMC∕GTA的Zeta电位得到最大幅度提升ꎬ平均电荷达到1.5mVꎬ在酸性㊁中性和碱性条件下均为正值ꎬ得益于高效的串联化学改性方法ꎬ使得真丝织物带电性能有较大改变ꎬ为抗菌活性和染色性能的改善提供了强有力的保障ꎮ2.3㊀改性真丝织物的性能分析图7(a)㊁图7(b)和图7(c)分别为SF和不同改性SF样品对E.coli㊁S.aureus两种菌的抗菌率统计结果及抗菌实验的光学图像ꎮ如图7(a)所示ꎬSF∕CMC㊁SF∕GTA对E.coli的抗菌率为49.5%㊁95 1%ꎬSF∕CMC∕GTA的抗菌率达到99.9%ꎮ图7(b)中ꎬSF∕CMC㊁SF∕GTA对S.aureus的抗菌率62.1%㊁98.8%ꎬ而SF∕CMC∕GTA的抗菌率达到99.9%ꎮ结果表明:CMC的加入对于真丝织物抗菌性能的提高有积极作用ꎬ且当CMC和GTA通过串联化学改性共同作用在真丝织物表面时ꎬ其抗菌效果得到最大程度的提升ꎮ从图7(c)抗菌实验的光学图像可以更直观地观察到真丝织物在改性前后的抗菌效果差别ꎬ改性真丝织物的菌落数目明显少于未改性织物ꎮ本研究中ꎬCMC本身就具有一定抗菌特性ꎬ并且通过反应将CMC接枝在真丝织物表面可提供更多的反应位点ꎬ将更多的GTA接枝到真丝织物上ꎮ改性真丝织物的抗菌性归因于其表面所带的大量正电荷ꎬ可以与带负电的细菌细胞膜结合ꎬ从而破坏其内部结构ꎬ达到抗菌的效果ꎮ图6㊀SF和不同改性SF样品的Zeta电位曲线Fig.6㊀ZetapotentialcurvesofSFanddifferentmodifiedSFsamples㊀㊀㊀㊀㊀㊀图7㊀SF和不同改性SF样品的抗菌性能Fig.7㊀AntibacterialpropertiesofSFanddifferentmodifiedSFsamples32 第6期杨㊀晟等:季铵盐∕两性壳聚糖改性真丝织物及其协同增效作用㊀㊀透气性㊁吸水性是衡量织物舒适性的重要指标ꎮ图8(a)和图8(b)分别为SF和不同改性SF样品的水蒸气透气率和吸水率ꎮ如图8(a)所示ꎬSF的水蒸气透气率为266g∕(m2 d)ꎬSF∕CMC㊁SF∕GTA㊁SF∕CMC∕GTA的水蒸气透气率分别为263㊁285㊁272g∕(m2 d)ꎮ图8(b)所示ꎬSF的吸水率为266%ꎬSF∕CMC㊁SF∕GTA㊁SF∕CMC∕GTA的吸水率分别为254%㊁328%㊁326%ꎮ相比于SFꎬSF∕GTA㊁SF∕CMC∕GTA都有较高的水蒸气透气率和吸水率ꎬ这是由于强亲水性GTA的引入改善了真丝织物的亲水性ꎬ吸湿㊁保湿性都得到一定程度的提高ꎮ图9(a)㊁图9(b)分别为SF和不同改性SF样品染色后残液吸光度的测试结果和上染率的统计结果ꎮ由图9(a)可知ꎬ染液的特征波长在516nm和540nm处ꎬ后通过测试真丝织物的染色残液在该波长的吸光度并按照式(4)可得到上染率ꎮ通过图9(b)所示ꎬSF的上染率为1.15%ꎬSF∕CMC上染率为3.15%ꎬ略高于SFꎬ这是由于接枝在真丝织物表面的CMC会电解出一部分氨基正电荷ꎬ对真丝织物染色有一定的积极作用ꎮ而SF∕GTA㊁SF∕CMC∕GTA的上染率分别为50.6%㊁63.7%ꎬ远高于SF的上染率ꎮ从图9(c)中真丝织物染色前后的光学图像可以更直观的发现ꎬ相比于SFꎬ改性真丝织物在染色后颜色更深ꎬ上染率提升幅度更大ꎮ综上所述ꎬ当CMC与GTA共同作用在真丝织物表面时ꎬ相比于SFꎬSF∕CMC∕GTA上染率提高了近55倍ꎬ染色性能得到显著改善ꎮ本工作中使用的活性染料属阴离子染料ꎬGTA对真丝织物的改性过程实际上是对氨基的阳离子性改性ꎬ得益于改性真丝织物表面携带的大量季铵盐基团ꎬ可增强表面正电性ꎮ通过强电荷吸引力提高阴离子染料的上染率ꎬ从而提升真丝织物的染色性能ꎮ㊀㊀㊀㊀图8㊀SF和不同改性SF样品的透气率和吸水率Fig.8㊀AirpermeabilityandwaterabsorbabilityofSFanddifferentmodifiedSFsamples㊀㊀㊀㊀42 现代纺织技术第31卷图9㊀SF和不同改性SF样品的染色性能Fig.9㊀DyeingpropertiesofSFanddifferentmodifiedSFsamples3㊀结㊀论本文采用简单㊁高效的串联化学改性方法对SF进行功能改性ꎬ探索CMC和GTA的对真丝织物协同改性的可行性ꎮ分析改性前后真丝织物的形貌㊁结构和物化性能变化ꎬ并对比测试其抗菌和染色性能ꎬ主要结论如下:a)ATR ̄FTIR㊁XPS测试证明了水溶性羧化壳聚糖和2ꎬ3 ̄环氧丙基三甲基氯化铵对真丝织物串联改性的成功实施ꎬSEM㊁XRD㊁拉伸测试和TG等测试显示ꎬ串联改性对SF的微观形貌㊁力学性能和热稳定性能影响较小ꎮZeta电位测试表明串联改性可将SF表面电荷由-26.3mV增至1.5mVꎮb)抗菌测试显示ꎬSF∕CMC对E.coli和S.aureus的抗菌率分别为49.1%和61.3%ꎬSF∕CMC∕GTA对两种菌的抗菌率均达到99.9%ꎬ证明协同改性可以显著提高SF抗菌活性ꎮc)染色实验表明协同改性可显著改善织物染色性能ꎬSF∕CMC∕GTA的上染率相对SF提高了近55倍ꎮ参考文献:[1]张勇ꎬ陆浩杰ꎬ梁晓平ꎬ等.蚕丝基智能纤维及织物:潜力㊁现状与未来展望[J].物理化学学报ꎬ2022ꎬ38(9):64 ̄79.ZHANGYongꎬLUHaojieꎬLIANGXiaopingꎬetal.Silkmaterialsforintelligentfibersandtextiles:Potentialꎬprogressandfutureperspective[J].ActaPhysico ̄ChimicaSinicaꎬ2022ꎬ38(9):64 ̄79.[2]张炜ꎬ毛庆楷ꎬ朱鹏ꎬ等.乙醇∕水体系中改性蚕丝织物的活性染料染色动力学和热力学[J].纺织学报ꎬ2020ꎬ41(6):86 ̄92.ZHANGWeiꎬMAOQingkaiꎬZHUPengꎬetal.Kineticandthermodynamicofreactivedyestudyonsilkfabricmodificationinethanol∕watersystem[J].JournalofTextileResearchꎬ2020ꎬ41(6):86 ̄92.[3]李佳ꎬ王勃翔ꎬ霍雨心ꎬ等.纳米改性制备温敏响应性柞蚕丝织物[J].丝绸ꎬ2022ꎬ59(10):20 ̄26.LIJiaꎬWANGBoxiangꎬHUOYuxinꎬetal.Thermo ̄responsivetussahsilkfabricmodifiedwithnanotechnology[J].JournalofSilkꎬ2022ꎬ59(10):20 ̄26. [4]LIUJLꎬLIANGJYꎬDINGJNꎬetal.Microfiberpollution:Anongoingmajorenvironmentalissuerelatedtothesustainabledevelopmentoftextileandclothingindustry[J].EnvironmentꎬDevelopmentandSustainabilityꎬ2021ꎬ23(8):11240 ̄11256.[5]CHENLZꎬCAROFꎬCorbettCJ.Estimatingtheenvironmentalandeconomicimpactsofwidespreadadoptionofpotentialtechnologysolutionstoreducewateruseandpollution:ApplicationtoChina'stextileindustry[J].EnvironmentalImpactAssessmentReviewꎬ2019ꎬ79:106293. [6]鲁秀国ꎬ官伟ꎬ陈晶.壳聚糖化学改性吸附水中重金属的研究进展[J].化工新型材料ꎬ2022ꎬ50(12):254.LUXiuguoꎬGUANWeiꎬCHENJing.Researchprogressonadsorptionofheavymetalsinwaterbychemicalmodificationofchitosan[J].NewChemicalMaterialsꎬ2022ꎬ50(12):254.[7]汤薇ꎬ董静ꎬ赵金荣ꎬ等.壳聚糖改性及改性壳聚糖应用研究进展[J].济南大学学报(自然科学版)ꎬ2023ꎬ37(1):84 ̄93.TANGWeiꎬDONGJingꎬZHAOJinrongꎬetal.Researchprocessinchitosanmodificationandapplicationofmodifiedchitosan[J].JournalofUniversityofJinan(ScienceandTechnology)ꎬ2023ꎬ37(1):84 ̄93.[8]张雨航ꎬ孙润军ꎬ魏亮ꎬ等.壳聚糖季铵盐溶液对涤∕棉织物的抗菌整理[J].纺织高校基础科学学报ꎬ2022ꎬ35(4):68 ̄73.52第6期杨㊀晟等:季铵盐∕两性壳聚糖改性真丝织物及其协同增效作用ZHANGYuhangꎬSUNRunjunꎬWEILiangꎬetal.Antibacterialfinishingofpolyestercottonfabricwithchitosanquaternaryammoniumsaltsolution[J].BasicSciencesJournalofTextileUniversitiesꎬ2022ꎬ35(4):68 ̄73.[9]OBEIDATWMꎬGHARAIBEHSFꎬJARADATA.Thein ̄fluenceofdrugssolubilitiesandchitosan ̄TPPformulationparametersonthemeanhydrodynamicdiametersanddrugsentrapmentefficienciesintochitosan ̄TPPnanoparticles[J].AAPSPharmSciTechꎬ2022ꎬ23(7):262. [10]郑宏飞ꎬ汪泳ꎬ汪庆ꎬ等.壳聚糖在二元离子液体中的溶解性及结构[J].高分子材料科学与工程ꎬ2020ꎬ36(12):82 ̄89.ZHENGHongfeiꎬWANGYongꎬWANGQingꎬetal.Solubilityandstructureofchitosaninbinaryionicliquids[J].PolymerMaterialsScience&Engineeringꎬ2020ꎬ36(12):82 ̄89.[11]FERREROFꎬPERIOLATTOMꎬBURELLISꎬetal.Silkgraftingwithchitosanandcrosslinkingagents[J].FibersandPolymersꎬ2010ꎬ11(2):185 ̄192.[12]DAVARPANAHSꎬMAHMOODINMꎬARAMIMꎬetal.Environmentallyfriendlysurfacemodificationofsilkfiber:Chitosangraftinganddyeing[J].AppliedSurfaceScienceꎬ2009ꎬ255:4171 ̄4176.[13]SHARIATINIAZ.Carboxymethylchitosan:Propertiesandbiomedicalapplications[J].InternationalJournalofBiologicalMacromoleculesꎬ2018ꎬ120:1406 ̄1419. [14]吴沥豪ꎬ陈功ꎬ任康ꎬ等.羧甲基壳聚糖基生物医用材料降解代谢行为的研究进展[J].高分子通报ꎬ2023ꎬ36(2):148 ̄157.WULihaoꎬCHENGongꎬRENKangꎬetal.Researchprogressoninvivodegradationandmetabolismassessmentofcarboxymethylchitosan ̄basedbiomaterials[J].PolymerBulletinꎬ2023ꎬ36(2):148 ̄157.[15]ZHANGZTꎬCHENLꎬJIJMꎬetal.Antibacterialpropertiesofcottonfabricstreatedwithchitosan[J].TextileResearchJournalꎬ2003ꎬ73(12):1103 ̄1106. [16]SAHARIAHPꎬMássonM.Antimicrobialchitosanandchitosanderivatives:Areviewofthestructure ̄activityrelationship[J].Biomacromoleculesꎬ2017ꎬ18(11):3846 ̄3868.[17]张伟.经壳聚糖季铵盐处理后真丝(绸)结构与性能的研究[D].苏州:苏州大学ꎬ2007:20 ̄35.ZHANGWei.ResearchontheStructureandPropertiesofSilkFiber(Fabric)TreatedwithHTCC[D].Suzhou:SoochowUniversityꎬ2007:20 ̄35.[18]LIMSHꎬHUDSONSM.Synthesisandantimicrobialactivityofawater ̄solublechitosanderivativewithafiber ̄reactivegroup[J].CarbohydrateResearchꎬ2004ꎬ339(2):313 ̄319.[19]WUYꎬYANGSꎬFUFYꎬetal.Aminoacid ̄mediatedloadingofAgNPsandtannicacidontocottonfabrics:Increasedantibacterialactivityanddecreasedcytotoxicity[J].AppliedSurfaceScienceꎬ2022ꎬ576:151821. [20]YANGWQꎬLVLLꎬLIXKꎬetal.Quaternizedsilknanofibrilsforelectricitygenerationfrommoistureandionrectification[J].ACSAanoꎬ2020ꎬ14(8):10600 ̄10607. [21]LUQꎬHUXꎬWANGXQꎬetal.Water ̄insolublesilkfilmswithsilkIstructure[J].ActaBiomaterialiaꎬ2010ꎬ6(4):1380 ̄1387.[22]WANGPꎬZHANGMYꎬQUJHꎬetal.Antibacterialcottonfabricpreparedbya"graftingto"strategyusingaQACcopolymer[J].Celluloseꎬ2022ꎬ29(6):3569 ̄3581. [23]郑宏飞ꎬ汪瑞琪ꎬ汪庆ꎬ等.氧化壳聚糖改性抗菌蚕丝织物的制备及其性能[J].纺织学报ꎬ2020ꎬ41(5):121 ̄128.ZHENGHongfeiꎬWANGRuiqiꎬWANGQingꎬetal.Preparationandpropertiesofantibacterialsilkfabricmodifiedwithoxidizedchitosan[J].JournalofTextileResearchꎬ2020ꎬ41(5):121 ̄128.[24]HUXꎬKAPLANDꎬCEBEP.Determiningbeta ̄sheetcrystallinityinfibrousproteinsbythermalanalysisandinfraredspectroscopy[J].Macromoleculesꎬ2006ꎬ39(18):6161 ̄6170.[25]YEJLꎬMASQꎬWANGBBꎬetal.High ̄performancebio ̄basedepoxiesfromferulicacidandfurfurylalcohol:Synthesisandproperties[J].GreenChemistryꎬ2021ꎬ23(4):1772 ̄1781.[26]LUYHꎬLINHꎬCHENYYꎬetal.StructureandperformanceofBombyxmorisilkmodifiedwithnano ̄TiO2andchitosan[J].FibersandPolymersꎬ2007ꎬ8(1):1 ̄6.[27]DUANPPꎬXUQBꎬZHANGXJꎬetal.Naturallyoccurringbetainegraftedoncottonfabricforachievingantibacterialandanti ̄proteinadsorptionfunctions[J].Celluloseꎬ2020ꎬ27(11):6603 ̄6615.[28]YANXJꎬZHUXWꎬRUANYTꎬetal.Biomimeticꎬdopamine ̄modifiedsuperhydrophobiccottonfabricforoil ̄waterseparation[J].Celluloseꎬ2020ꎬ27(13):7873 ̄7885.[29]SHENLꎬDAIJJ.Improvementofhydrophobicpropertiesofsilkandcottonbyhexafluoropropeneplasmatreatment[J].AppliedSurfaceScienceꎬ2007ꎬ253(11):5051 ̄5055.[30]LIGHꎬLIUHꎬLITDꎬetal.Surfacemodificationandfunctionalizationofsilkfibroinfibers∕fabrictowardhighperformanceapplications[J].MaterialsScienceandEngineeringꎬ2012ꎬ32(4):627 ̄636.[31]寇爱静ꎬ林欢ꎬ柳守婷等.饲喂法制备碳纳米材料改性蚕丝的导热性能[J].材料科学与工程学报ꎬ2021ꎬ3962 现代纺织技术第31卷(2):317 ̄321.KOUAijingꎬLINHaunꎬLIUShoutingꎬetal.Thermalconductivityofmodifiedsilkpreparedbyfeedingcarbonnanomaterials[J].JournalofMaterialsScienceandEngineeringꎬ2021ꎬ39(2):317 ̄321.[32]FERREROFꎬPERIOLATTOMꎬLURASCHIM.Silkgraftingwithmethacrylicmonomers:Processoptimizationandcompa ̄rison[J].JournalofAppliedPolymerScience.ꎬ2007ꎬ103(6):4039 ̄4046.[33]于海洋ꎬ王昉ꎬ刘其春ꎬ等.新型丝素蛋白膜的结构和热分解动力学机理[J].物理化学学报ꎬ2017ꎬ33(2):344 ̄355.YUHaiyangꎬWANGFangꎬLIUQichunꎬetal.Structureandkineticsofthermaldecompositionmechanismofnovelsilkfibroinfilms[J].ActaPhysico ̄ChimicaSinicaꎬ2017ꎬ33(2):344 ̄355.Silkfabricmodifiedbyquaternaryammoniumsalt∕amphotericchitosananditssynergisticeffectYANGSheng1ꎬXUZhaomei2ꎬMATingfang2ꎬFUFeiya1ꎬLIUXiangdong1ꎬYAOJuming1(1.SchoolofMaterialsScience&EngineeringꎬZhejiangSci ̄TechUniversityꎬHangzhou310018ꎬChinaꎻ2.HangzhouWENSLISilkDigitalPrintingCo.ꎬLtd.ꎬHangzhou310020ꎬChina)Abstract:Withexcellentwearability biocompatibilityandbiodegradability silkiswidelyusedintextile biomedicine foodandotherfields.Chemicalmodificationisanimportanttechnicalmeanstofurtherexpandtheuseofsilk.However theproteinpropertiesofsilkhavehighrequirementsonchemicalrawmaterialsandreactionpaths soitisdifficultfortraditionalsinglesubstancemodificationmethodstoachievegreenandefficientmodificationatthesametime.Inthispaper anewmethodoftandemchemicalmodificationofrealsilkfabric SF usingwater ̄solublecarboxylatedchitosan CMC and2 3 ̄epoxy ̄propyltrimethylammoniumchloride GTA isproposedforthefirsttime.Themorphology structureandphysicochemicalpropertiesofSF SF∕CMC SF∕GTAandSF∕CMC∕GTAsampleswerecomparedandanalyzedbymeansofscanningelectronmicroscopy fouriertransforminfrared X ̄raydiffraction X ̄rayphotoelectronspectroscopy tensiletestandthermogravimetricanalysis.TheresultsshowthatalthoughtheamidebondformedbyCMCreactionininfraredspectroscopycoincideswithSFcharacteristicpeak theshiftofcarbonylCbindingenergyandtheincreaseofC NpeakareainX ̄rayphotoelectronspectroscopyprovethatthetandemchemicalmodificationcanbesuccessfullyimplementedat80ħinaqueoussolvent.TheintroductionofCMCprovidesmorereactionsitesforGTA andalargenumberofhydroxylgroupsformingafterthereactionofGTAcanformrichhydrogenbondswithCMCtohelpitstabilize.Withthechemicalmodificationintandem asmallamountoffibrillaappearedonthesurfaceofthefabricfiber.X ̄raydiffractionspectraalsoshowedthatthemodificationreactionwasmainlyintheamorphousregionofthesurface andhadlittleeffectontheSFcrystalstructure.ComparedwithSF thetensilestrength 62.7MPa andthermaldecompositiontemperature 327ħ ofSF∕CMC∕GTAhavenoobviouschanges whichshowsthattheseriesmodificationmethodusedinthisworkismildandeffective andthemodifiedfabricstillmaintainsgoodmechanicalpropertiesandthermalstability.ThedifferenceisthatcomparedwithSF thepermeabilityandwaterabsorptionofSF∕CMC∕GTAincreasedsignificantly reaching272g∕ m2 d and326% respectively whichshouldberelatedtothestronghydrophilicgroupsintroducedbyCMCandGTA.Inaddition theZetapotentialofSF∕CMC∕GTA 1.5mV wassignificantlyincreasedcomparedwiththatofSF -26.3mV provingthehighefficiencyoftandemmodification.ComparedwiththesinglemodifiedsampleSF∕CMCandSF∕GTA thetandem ̄modifiedsampleSF∕CMC∕GTAhadaquitehighantibacterialrate above99.9% bothagainstS.aureusandE.coli.ThedyeingexperimentshowedthatthedyeingrateofSF∕CMC∕GTAwas55timeshigherthanthatofSF whichisexpectedtobeappliedinthefieldofnon ̄washingprintinganddyeing.Thispaperprovidesscientificdatafortheapplicationoftandemreactioninfunctionalmodificationofrealsilkfabricsandanewtechnicalpathforgreensilkdyeingandfinishing.Keywords:silkfabric tandemreaction synergisticeffect antibacterialproperty dyeingproperty 72 第6期杨㊀晟等:季铵盐∕两性壳聚糖改性真丝织物及其协同增效作用。
性连锁平衡致死雄蚕品种培育的实践与追求

L U X n j, e rn Z n r n WA I i-u HE K - o g, HU Xi- o g, NG n - in CHE h Yo g qa g, NS i
维普资讯
委童遍袁
・
g3 卷 g2 2 0 年5 l s 期 07 f
Vo .8 No2 Ma , 0 7 1 . y 2 0 3
6 ・
Bu ltn o e i u r l i fS rc hu e e
性连锁平衡致死雄蚕品种培 育的实践与追求
期 间一 方面开展 性连锁 平衡致 死基 因转 移到 现 行 优 良家 蚕 品种 的方法 研究 ,另一 方 面是利用 弓进 f 品种 的雄 与 我 国 当时 优 良家 蚕 品种 雌 配成 杂交 组
合 , 中筛选较优 良的杂交组合到农村试养 , 4× 从 夏
收稿 日期 : 0 7 0— 0 2 0 - 13 基金项 目: 浙江省和国家科技 成果转化 资金: S F 2 3 0 16 O E N 130 1 作者简介: 柳新菊(9 1 )女, 1 7~ , 河南人 , 大专。主要从 事家蚕 育种研究 。E m i l x@ as r - a :uj za.g li o
广应 用 。俄 罗斯科 学 院斯 特隆 尼科夫 院 士等利 用性 状 标记 和辐 射诱 变技术 ,成功 地育 成 了家 蚕性 连锁 平 衡致死 系 。该 系统 自交 , 一半雌性 和雄性在 胚胎 有 期死亡, 另一 半能正 常孵化 , 品系得 以保 持 。平 衡 使 致死 品 系的雄与常规 品系的雌交 配 的后代 只有 雄蚕
柳新 菊 , 何克 荣 , 祝新 荣 , 王永 强 ,陈 诗
“苏雄×荧晓”蚕品种的创新与发展

技成 果 的转化 应 用 。苏 豪 国 际集 团于 20 02年 联 合 苏州 大学 和浒关 蚕 种场 合 作 开发 荧 光 茧色 判性 “ 苏 雄 X荧 晓” 品种 、 蚕 规模 化 生 产优 质 雄蚕 茧 、 制 高 缫 品位 雄蚕 丝 , 破 了传 统 的茧丝 生产模 式 , 打 是符合 茧
丝绸 的发 展方 向 , 走优 质 、 品之路 的一个 重要举 是 精 措, 是重 塑江苏 辉煌 的一个 新亮 点 , 依次来 推动 江 并 苏 茧丝 行 业 新 的 发 展 。“ 雄 ×荧 晓 ” 品 种 于 苏 蚕 20 0 8年 1 月份 通过 了江 苏 省桑 蚕 品种 审 定 委员 会 1
种“ 菁松 x皓月 ” 相仿 , 质量指标达 到生产 6 A级生 丝
雄蚕 茧单 丝纤 度 细是生 产 高品位 细条 纹规格 生
茧 为黄荧 光 、 茧为 紫荧光 。 雌
1 生产 雄 蚕 茧 的 四种 途 径
1 1 饲 养家蚕 限性 品种 .
“ 苏雄 X 晓 ” 品种 在 农 村 饲 养 、 茧 收 、 荧 蚕 蚕 烘 与 现行 蚕品种 一样无 须特 殊 的操 作 , 村容 易推广 。 农
丝 厂煮 茧前采 用特 制 的 荧光 灯 , 即可 进行 雌 雄 蚕茧
的审定 。
从 目前 四种途 径实 际运 行分 析 , 养优 质 荧光 饲 茧色 判性 “ 雄 ×荧 晓 ” 品种 是 批 量提 供 优 质 雄 苏 蚕
蚕茧 成本最 低 、 方法最 捷径 的有效 途径 。
2 “ 雄 X荧 晓 ” 品种 的 特 色 与创 新 苏 蚕
现行 蚕 品 种 的蚕 茧在 自然光 下 呈 白色 , 在特 但 定波 长 的紫外 光照射 下能 发 出黄色 、 白色 、 色等多 紫 种荧 光色 。而本 品种 蚕 茧 的荧 光 色 只有 两种 , 其雄
热点专题05 生物变异与育种专题-2023年高考生物临考热点梳理课件

α射线、γ射线、微重 力、激光等处理、再筛 选(2)化学:秋水仙 素、硫酸二乙脂处理,
培养,培育出单倍 体植株(2)将单倍 体植株幼苗经一定 浓度的秋水仙素处
用一定浓度的秋水仙 素处理萌发的种子或
幼苗
基因与运载体结合→ 目的基因导入受体细 胞→目的基因的检测 与表达→筛选出符合
再筛选
理获得纯合子
典型例题4 平衡致死体系
【典型例题4】.雄蚕食桑少、产丝率比雌蚕高20%左右,但普通蚕(图1)难以通过外形判断其性别, 故研究者制作了一套雌蚕和雄蚕的平衡致死系统(图2),利用二者互相交配或者与其他野生型蚕交配 ,最终实现只养雄蚕。图3表示某特殊蚕的染色体和基因组成。下列叙述错误的是( D ) A.l1和l2基因相对于野生型基因而言,是隐性基因 B.图2中雄蚕与野生雌蚕交配,后代雌蚕全部死亡 C.图2中雌雄蚕互相交配,其子代雄蚕的染色体组成与图2中的雄蚕相同 D.如果用图3中雌蚕替代图2中雌蚕,不能实现与图2中雌蚕相同的功能
要求的新品种
优点
①“集优”:使分散 在同一物种不同品种 中的多个优良性状集 中于同一个体上②操
作简便
提改产高良生变某新异些基频性因率状的,(途大是径幅唯)度一明(显子加缩代速短为育育纯 种种合进年子程限),植果营物实养茎、物杆种质粗子含壮都量,比提叶较高片大、,①改杂目造交育的生不种性物亲周强②和期克的, 短服障可远碍定缘③向
个性状常优于双亲,这种现象称为杂种优势。获得具有杂种优势的杂合种子是提高水稻产量
的重要途径。
(1)中国是最早种植水稻的国家,已有七千年以上历史。我国南方主要种植籼稻,北方主要种
植粳稻。籼稻和粳稻是由共同的祖先在不同生态环境中,经过长期的
,进化形成的。
拟南芥温敏雄性不育突变体atms1的获得及表型分析

21 0 1年 l O月
上洛戈
耀( 自然 科 学 舨)
Vo 7 No 5 l + l
Oc . 2 l t Ol
J U N LO H N H I N RIY N T R LSIN E O R A FS A G A WE S ( A U A E C ) U T C
Ab t a t sr c :An a in e e au e A a i o s h l n l t r e mu a ta in e e a u e s n o y mb e ttmp r t r r b d p i t a i a mae se l t n mb e t tmp r t r -e s r s a i -
fw ini c n mp ime t o a e t l t p n h rwh n a e sg f a ti ar n sc mp r d wi wi y e a t e e mbin e e au e wa e o 2 ℃ . i h d— e ttmp r t r s b lw 3
a l的不 育 表 型 是 由单 个 隐 性 核 基 因控 制 的 . t ms
卫
关键 词 : 拟南芥 ; 温敏雄性不 育; 花粉 ; 药; 花 绒毡层
中 图 分 类 号 : 4 Q9 文献标志码 : A 文 章 编 号 : 0 72 6 (0 1 0 -6 10 1o -8 1 2 1 )50 8 -6
mu h t i n rt a h to i — p c h n e n t a fw l t e.a d t e a ms a e u d v l p n x i i d mu h r t r e h n h d y n h t l tp t m e eo me t e h b t c ea d d t a e
雄蚕丝结构与性能初探

J n 01 No 01 研 究 与 技 术 a 2 0
雄蚕丝结构与性能初探
陈玉梅 ,蔡再生 ,丁志用 ,姚 建洲 ,杨 斌
( . 华大 学 生 态 纺 织 教 育 部 重 点 实 验 室 , 上 海 2 1 2 ; 2 江 苏 民星 茧 丝 绸 股 份 有 限公 司 ,江 苏 盐 城 2 4 0 ) 1东 060 . 2 2 0
关 键 词 : 雄 蚕 丝 ; 结 构 ; 性 能
中图分类号:T 1 2 3 80.3
文献标识码:B
文章编号:1 0 — 0 3( 0 0 卜0 2 — 4 0 1 7 0 2 1 )0 0 3 0
P e i i a yEx l r to fS r cur sa dPr p risf rM a eS l r lm n r p o ain o tu t e n o e te o l i k
Key wor : ae s l ds M l i k;S r t r ; o e te tuc u e Pr p ri s
现 行 蚕 茧 在 烘 茧 、煮 茧 、 缫 丝 时 都 是 雌 雄 混 合 , 只 能 缫 制 雌 雄 复 合 丝 ,很 难 批 量 生 产 出 6 级 高 品位 生 A
a d1 se t t c r. o ae t oma i , l i a ih rcy tlnt, r nain d ge , e t n - e s u t e C mp rdwi n r l l maesl h shg e r s l i oi tt e re h a 3h r u h sk k ai y e o
小麦温光敏雄性不育研究进展

式: 核质 互 作 雄性 不 育 利用 ( MS 、 学 杀雄 剂 利用 ( ) 度条 件 下 , C )化 或 温 不育 系表 现 雄 性不 育 , 用 于生 产 可
( H 和 光温 敏核 雄 性不 育 利用 。植 物雄 性 不 育是 杂 交 种 子 ; 在 另 一 光 照 和 ( 温 度 条 件 下 表 现 可 C A) 而 或1 指植 物雌 性 器官 功 能正 常 而雄 性 器官 丧 失其 受 精 能 育 .可 进 行该 材 料 的繁 殖 。从 而达 到 一 系两 用 的 目
落后 于上述 两种 作物 。 光 照 对 其 育 性 没 有 一 定 的 影 响 。 如 B Y 系 列 、 N A 在 小 麦杂 种优 势 利用 的三 种方 式 中 ,核 质 互作 3 1 、 3 44 ; ) 光 互 作 雄 不 育 , 育 性 的 表达 0 7A 3 1[ ( 温 - 3  ̄ 其
米杂 种优 势 利用 方 面取 得 了重 大 突破 。但小 麦 作 为 影 响 , 度对 其 育性 没 有一 定 的影 响 , A 1] 2温 温 如 3 [ () 3 ; 重要 的农 作 物之 一 ,其 杂 种优 势 的机 制 及利 用 研究 度 敏感 型 雄 性不 育 , 育性 的表达 受 温度 高低 影 响 , 其
摘要 : 小麦 温 光敏 雄 性 不 育是 杂种优 势利 用的 重要 部 分 。 而小 麦温光敏 雄 性 不育 受遗 传 和环 境 等
因子 影响 , 传机 理较 为 复杂 , 遗 因此 , 讨 小麦 温光敏 雄 性 不育机 制及 不 育基 因麦温 光敏 雄 性 不育 类型 、 生理 生化研 究 、 遗传 学研 究及分 子 生物 学等 方
4 一 4
专 题论述211 0 . 02
periodical__yyyhjswxb__yyyh2005__0504pdf__050415

程度+ !+ ,+ 4- 蛹虫草对常压耐缺氧作用 - - 7%8 雄性小鼠, 随机分 组, 每组 !" 只, 小鼠分别 #$ 蛹 虫 草 或 冬 虫 夏 草 水 提 液 后 !* (#), 将动物置于 ,*" (3 广口瓶中, 密封 ( 瓶中预先放入 !" 2 无水氯化钙, 上面覆盖二层滤纸) 观察小鼠存活时间+ !+ ,+ 1- 抗肿瘤作用- - 按照该虫草浓度对正常细胞无不良影 响, 但能完全抑制肿瘤细胞增长为有效标准+ !+ ,+ 9- 蛹虫草对异丙肾上原素诱发小鼠心肌耗氧量增加的保 护作用- - 7%8 雄性小鼠, 随机分组, 每组 !" 只, 小鼠分别 #$ 蛹虫草水提液, 冬虫夏草水提液, %& 注射生理盐水, 同时立即 于背部皮下注射盐酸异丙肾上腺素 ( 天丰药厂, ! (2 : , (3)!* (2 ;2
收稿日期: "##%(#/(#*! ! 接稿日期: "##%(#)(+* !通讯作者! 17II>LH7:P@:8 96K=7I( 3(J9@B:O97?=76Z N=@:98>:>, N7J)
+, ", *! 蛹虫草增强戊巴比妥钠的催眠作用 ! ! X1- 雄性小 鼠, 随机分为 1\,冬虫夏草组组 ( $ 8 R8 _ + , @H ) 和蛹虫草两个 剂量组 ( ", $ 8 R8 _ + , $, # 8 R8 _ + , @H) , 每组 +# 只, 给药容积为 "# JV R8 _ + ,1\ @H 给予等容积的生理盐水, 小鼠给药后 +$ J@:, @H 戊巴比妥钠 ( "$ J8 R8 _ + ) , 之后观察动物的睡眠时间, 睡眠 指标以动物翻正反射消失 + J@: 为入睡, +, ", %! 对二甲苯诱发小鼠耳部炎症的影响 ! ! X1- 雄性小
2023-2024学年辽宁省丹东市凤城市高考冲刺模拟生物试题含解析

2024年高考生物模拟试卷请考生注意:1.请用2B铅笔将选择题答案涂填在答题纸相应位置上,请用0.5毫米及以上黑色字迹的钢笔或签字笔将主观题的答案写在答题纸相应的答题区内。
写在试题卷、草稿纸上均无效。
2.答题前,认真阅读答题纸上的《注意事项》,按规定答题。
一、选择题:(共6小题,每小题6分,共36分。
每小题只有一个选项符合题目要求)1.物质X通过细胞膜的运输速率与膜内外浓度差成正比,且不具有载体饱和效应。
据此推断物质X进出细胞的方式为()A.自由扩散B.主动运输C.协助扩散D.胞吞和胞吐2.流感病毒是一类RNA病毒,外壳上的血凝素(HA) 和神经氨酸酶(NA)的作用是让病毒入侵细胞和已经在细胞内复制、组装好的病毒顺利出细胞。
研究人员提取某流感病毒的HA和NA注入小鼠体内,最终从小鼠体内分离得到能与该流感病毒特异性结合的抗体。
下列相关叙述错误的是()A.实验小鼠体内不仅能分离得到相应抗体,还有相应的效应T细胞B.可用荧光标记小鼠体内分离到的抗体检测人体内是否有该种流感病毒C.若指导HA和NA合成的流感病毒RNA发生了变化,则记忆细胞无法识别该病毒D.可通过注射HA和NA的抑制剂治疗被流感病毒感染的患者3.细胞学说揭示了生物界具有()A.差异性B.多样性C.特异性D.统一性4.下图为人体体液物质交换示意图,其中丁表示某类细胞的细胞内液。
下列叙述正确的是()A.甲的渗透压升高则会导致组织水肿B.甲中可含有抗体、激素和神经递质C.丁可表示淋巴细胞的细胞内液D.乙、丙是毛细血管壁细胞的内环境5.下列关于生物膜结构和功能的叙述错误的是()A.生物膜的元素组成相似,结构基本相同B.生物膜成分的更新可通过囊泡实现C.在线粒体和叶绿体的内膜上均可发生能量转换D.人体内性激素的合成与性腺细胞的内质网有关6.下列有关细胞结构与功能的叙述,不正确的是()A.原核细胞的DNA都是游离状态的DNAB.大分子物质都是以胞吞或胞吐方式通过生物膜的C.细胞骨架与细胞运动、分裂、分化以及信息传递等生命活动有关D.细胞间进行信息交流并不都需要细胞膜上的受体蛋白参与二、综合题:本大题共4小题7.(9分)河套地区玉米间作套种小麦,亩产达1000公斤以上,称为吨粮田。
【精编精校卷】2022届江西省七校高三上学期第一次联考生物试题(解析版)

A.细胞膜塑形蛋白是在核糖体上形成的
A.β-分泌酶起催化作用时消耗水分子
B.一个β-AP分子中至少含有39个肽键
C.β-AP寡聚合物可能引起大脑功能异常
D.用双缩脲试剂检测β-AP会产生紫色反应
【答案】B
【解析】
【分析】分析题图:图示表示APP形成β-AP的过程,该过程是在病理状态下进行的,由题图知APP形成β-AP的过程中需要β-分泌酶和y-分泌酶的催化作用,β-AP分子是由前体蛋白APP中的第597位氨基酸到635位氨基酸形成的、其含有的氨基酸数=635-597+1=39个,而β-AP分子沉积是Alzheimer型老年痴呆的主要病理特征。
2、溶酶体含有多种水解酶,能分解衰老、损伤的细胞器,吞噬并杀死侵入细胞的病毒或病菌。
【详解】A、细胞膜塑形蛋白的合成场所是核糖体,A正确;
B、构成“分子垃圾袋”的主要化膜相似,主要成分是磷脂和蛋白质,B正确;
C、根据题干信息“将来自细胞区室表面旧的或受损的蛋白质带到了内部回收利用工厂,在那里将废物降解,使组件获得重新利用”可知,“回收利用工厂”可能是溶酶体,“组件”可能是氨基酸,C正确;
B.构成“分子垃圾袋”的主要化膜相似
C.“回收利用工厂”可能是溶酶体,“组件”可能是氨基酸
D.囊泡将来自细胞区室表而旧的或受损的蛋白质带到了内部回收利用工厂的过程体现了生物膜的选择透过性
【答案】D
【解析】
【分析】1、细胞膜的主要成分是磷脂和蛋白质,其结构特点是具有一定的流动性,功能特性是选择透过性。
蚕业专利选载

华 书店购买 , 也可汇款给华德公同志邮购 . 邮购每本 外加2 元 邮挂费( . 5 邮购2 本以上者赠送《 中国蚕桑书
录》本 ) 1 。 地址: 山东省高密市人民政府办公室 邮编 :6 0 2 10 5
电话 : 5 6 2 1 0 0 1 5 6 7 9 8 0 3 — 3 2 或 0 4 4 5 3 3
维普资讯
委素遍 窳
-
g3 卷 第2 2 0 年5 7 期 06 月
Vo .7 No2 Ma , 0 6 1 . y 2 0 3
7 ・ 0
Bult f eiutr l i o rc l e en S u
蚕业专利选载
专 利名称 : 家蚕低 制种成 本单 交制种 方 法
单独饲养一部分蚕用于专门试验 , 大田附近和怀 对 疑地块必须进行试验 ,确保安全后方可采摘桑叶 : 四是严禁使用不明成分的农药 。
2 做好 蔟 中保护 , 少不 结茧蚕 . 4 减
关键 之一 。一是要稀放上蔟 ,确保有足够的营茧位
置; 二是要通风干燥 , 营茧期 间温度控制在 2%, 6 相
体 . 成 广 食 性 蚕 育 种 索 材 , 生 物 反 应器 用广 形 对
方法 . 它利用高孵化率非减数分裂孤雌生殖 . 繁殖
家蚕杂交种的雌性亲本 。 用家蚕限性卵色品种 繁杂 交种的雄性亲本。由于非减数分裂孤雌生殖产生全 雌子代。而限性卵色品种在卵期可根据卵色分辨雌
雄, 可以挑选 出雄卵 , 制成全雄子代。形成杂交种双 亲一亲 只养雌性 , 另一亲只养雄 亲, 然后 把这 2亲
号 利申请号: 260 4 0 10 7 . 0
公开号: N 4 l3 C 12 l0
致死品种 s 作索材 。 c h ②用“ 回交导入” 世代定向 和“ 选育 ” 相结合的方法 。 获得遗传基因稳定 、 经济性状
雄蚕新品种“川山×平30”的育成

资助项目:国家现代农业技术体系四川蚕桑创新团队项目(编号sccxtd-2020-17)作者简介:袁桂阳(1963-),男,本科高级农艺师,从事蚕桑生产科研推广工作。
*通讯作者为曹宁宁。
文献信息:袁桂阳,祝新荣,王少伯,等.雄蚕新品种“川山×平30”的育成[J].四川蚕业,2020,48(1):27-30.雄蚕新品种“川山×平30”的育成袁桂阳1祝新荣2王少伯1曾光远3吴钢4曹宁宁1*王永强2邵云华3龚大刚5蒋佳4文廷勇1(1四川省南充蚕种场,四川南充637000,2浙江省农业科学院蚕桑研究所,浙江杭州310021,3杭州市蚕桑技术推广总站(杭州蚕种场),浙江杭州310000,4四川省蚕业管理总站,四川成都610041,5四川省阆中蚕种场,四川阆中637400)摘要:以四川省南充蚕种场保存的七个优良品种与从浙江省农业科学院蚕桑研究所引进的性连锁平衡致死系雄蚕品种“平30”进行杂交组配,筛选出了一对综合经济性状优良的杂交组合川山×平30。
川山×平30在四川省家蚕新品种鉴定中表现良好,综合性状稳定、雄蚕率高、发育齐一、产量稳定、饲养及丝质成绩优良,尤其在雄蚕率、发育整齐度、健康性及茧丝质等重要指标中表现优异,达到雄蚕品种的生产实用化要求,是一对春秋兼用的雄蚕新品种。
2018年3月通过四川省家蚕品种审定委员会审定,适宜在四川蚕区春、秋季饲养。
关键词:雄蚕新品种川山伊平30平衡致死系杂交组配雄蚕品种具有强健好养、叶丝转化率高、茧丝质量优良等优点,因而“专养雄蚕技术”被人们认为是继利用一代杂交种后,蚕业科技上的第二次重大革命。
家蚕性连锁平衡致死系是实现专养雄蚕的一条重要途径,性连锁平衡致死系在其雄蚕的Z 染色体上带有2个非等位的胚胎期隐性纯合致死突变基因l 1和l 2,即雄蚕的性染色体结构为Z l 1+Z l 2+,而其雌蚕的W 染色体上则易位有这2个致死基因的正常型等位基因,因而该系统内的雌雄交配,能完整地保留平衡致死基因。
桑蚕国家基金1999-2011
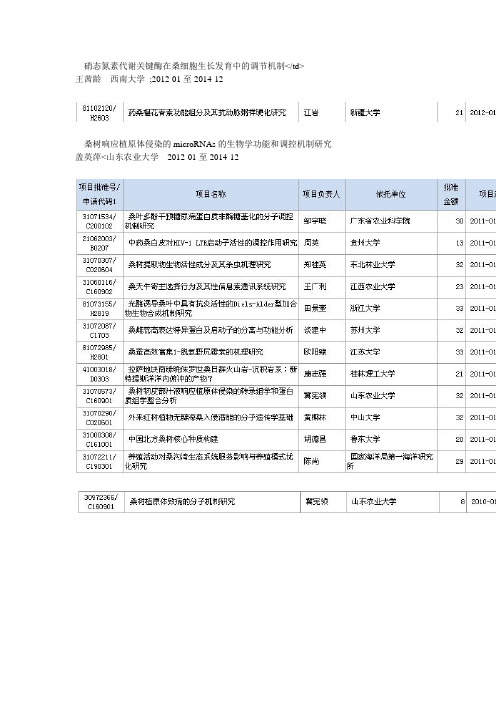
硝态氮素代谢关键酶在桑细胞生长发育中的调节机制</td>王茜龄西南大学;2012-01至2014-12桑树响应植原体侵染的microRNAs的生物学功能和调控机制研究盖英萍<山东农业大学2012-01至2014-12负责人:鲁成金额:9万申请时间:1998学科代码:养蚕学(C1703)项目批准号:39870603申请单位:西南大学研究类型:应用基础研究查看摘要收藏负责人:朱良均金额:11万申请时间:1997学科代码:养蚕学(C1703)项目批准号:39770573申请单位:浙江大学研究类型:应用基础研究查看摘要收藏负责人:鲁成金额:21万申请时间:2002学科代码:养蚕学(C1703)项目批准号:30271005申请单位:西南大学研究类型:基础研究查看摘要收藏负责人:向仲怀金额:18万申请时间:2001学科代码:养蚕学(C1703)项目批准号:30170719申请单位:西南大学研究类型:基础研究查看摘要收藏负责人:宋方洲金额:13万申请时间:1997学科代码:养蚕学(C1703)项目批准号:39770574申请单位:西南大学研究类型:基础研究查看摘要收藏负责人:周泽扬参与人:周泽扬, 朱勇, 夏庆友, 冯丽春金额:8.5万申请时间:1995学科代码:养蚕学(C1703)项目批准号:39570557申请单位:西南大学研究类型:基础研究关键词:家蚕RAPD 基因定位查看摘要收藏负责人:纪涛参与人:纪涛, 韩秀文, 宋永哲, 苏凡, 滕瑛, 王国祯, 王东, 马中飞, 李瑶, 金欣金额:5万申请时间:1992学科代码:结构化学(B0301)项目批准号:29273137申请单位:中国科学院大连化学物理研究所研究类型:应用基础研究查看摘要收藏负责人:孙京臣参与人:孙京臣, 严会超, 李茵茵, 刘颖斐, 龙虎, 黄曾慰, 戴兴红, 谭佩婵申请单位:华南农业大学研究类型:应用基础研究关键词:BmCPV;冷冻电子断层扫描;电子显微镜;入侵;机制查看摘要收藏负责人:黄自然参与人:黄自然, 方菲芳, 潘庆中, 谭炳安, 黄玉莲, 陈风珍, 徐飞金额:3万申请时间:1989学科代码:养蚕学(C1703)项目批准号:38970583申请单位:华南农业大学研究类型:应用基础研究关键词:抗菌肽;家蚕;桑树.收藏负责人:王永强参与人:王永强, 孟智启, 翁宏飚, 何秀玲, 柳新菊, 刘培刚金额:62万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172271申请单位:浙江省农业科学院研究类型:应用基础研究关键词:遗传纯合系;杂种优势;基因芯片;家蚕查看摘要收藏负责人:孟艳参与人:孟艳, 高俊山, 魏国清, 张军, 王敬, 王文静, 赵小明, 陈莉金额:62万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172270申请单位:安徽农业大学研究类型:基础研究关键词:家蚕;β-呋喃果糖苷酶;基因表达;昆虫食性;生物碱毒性查看摘要收藏负责人:鲁成参与人:鲁成, 陈鹏, 张春冬, 彭丽娜, 李方方, 黄茂华, 陈雪梅申请单位:西南大学研究类型:基础研究关键词:家蚕;细胞凋亡;bcl2基因家族;基因调控查看摘要收藏负责人:李泰参与人:李泰, 向仲怀, 祝顺琴, 柯晓雪, 谈娟, 李婷, 郝向伟, 郑春琴, 张奎, 陈毅彪金额:58万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172268申请单位:西南大学研究类型:基础研究关键词:家蚕;中肠干细胞;识别;鉴定;功能分析查看摘要收藏负责人:程道军参与人:程道军, 赵萍, 魏玲, 聂红毅, 张辉洁, 孟勐, 王永虎, 刘勇金额:62万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172267申请单位:西南大学研究类型:应用基础研究关键词:家蚕;眠性主基因;定位克隆;功能;分子作用机制查看摘要收藏负责人:沈兴家参与人:沈兴家, 宋菲, 王欣, 黄勇, 唐顺明, 郝碧芳, 张谦瑾, 王力刚, 侯启瑞金额:62万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172266申请单位:江苏科技大学研究类型:应用基础研究关键词:非编码RNA,microRNA,家蚕,丝素基因,转录后调控查看摘要收藏负责人:冯启理参与人:冯启理, 邓惠敏, 张丽丽, 章佳玲, 饶中臣, 吴琼芳, 李勇, 王辉申请单位:华南师范大学研究类型:基础研究关键词:家蚕;变态发育;蜕皮激素;翅原基表皮蛋白;转录因子查看摘要收藏负责人:徐世清参与人:徐世清, 司马杨虎, 陈息林, 甘丽萍, 杨德印, 袁红霞, 殷为民, 季明明, 梁辉, 王文栋金额:60万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172264申请单位:苏州大学研究类型:应用基础研究关键词:家蚕;滞育诱导;生物钟信号途径;基因发掘;温度授时查看摘要收藏负责人:孙京臣参与人:孙京臣, 何键, 刘吉平, 李鲲鹏, 钟杨生, 李茵茵, 徐亮亮, 粟硕, 梁智升, 李征金额:60万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172263申请单位:华南农业大学研究类型:应用基础研究关键词:家蚕质型多角体病毒;冷冻电子显微镜;结构;入侵机理;BiFC查看摘要收藏负责人:鲁兴萌参与人:鲁兴萌, 李明乾, 蒋振东, 马焕艳, 孟祥坤, 孙文静, 何欣怡金额:60万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31172262申请单位:浙江大学研究类型:应用基础研究关键词:家蚕传染性软化病病毒;内部核糖体进入位点;翻译机制;反向遗传学;侵染机制查看摘要收藏负责人:李治参与人:李治, 许平, 张怡, 张小燕, 韦伟, 林海静, 张玺, 王丽君, 姜春宝, 王钰金额:23万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31101770申请单位:重庆师范大学研究类型:基础研究关键词:极管蛋白;孢壁蛋白;相互作用;鉴定;家蚕微孢子虫查看摘要收藏负责人:王茜龄参与人:王茜龄, 余茂德, 赵爱春, 李镇刚, 陈雪娇, 黎其友, 张琼予, 吕蕊花金额:23万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31101769申请单位:西南大学研究类型:应用基础研究关键词:桑树;硝态氮代谢;RT-PCR; 调控机制查看摘要收藏负责人:林英参与人:林英, 程道军, 杨从文, 邹勇, 费纪涛, 罗娟, 沈关望金额:23万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31101768申请单位:西南大学研究类型:基础研究关键词:家蚕;卵黄原蛋白;卵黄原蛋白受体;机制查看摘要收藏负责人:唐旭东参与人:唐旭东, 许平震, 陶美林, 沈中元, 朱峰, 管蕊, 陈大睿金额:22万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31101767申请单位:江苏科技大学研究类型:基础研究关键词:SUMO;家蚕;类泛素化;杆状病毒查看摘要收藏负责人:郝碧芳参与人:郝碧芳, 黄金山, 汪生鹏, 张美蓉, 王欣, 王猛, 陈爱春金额:23万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31101766申请单位:江苏科技大学研究类型:基础研究关键词:BmNPV;ORF126;亚型;RGD;表达查看摘要收藏负责人:杨婉莹参与人:杨婉莹, 温硕洋, 许俊峰, 叶慧, 程玉, 刘洋, 贾浩伟金额:23万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31101765申请单位:华南农业大学研究类型:基础研究关键词:LPS;BmMLs;BmToll9;抗菌肽;自噬查看摘要收藏负责人:桂仲争参与人:桂仲争, 桂仲争金额:4万申请时间:2011学科代码:养蚕学(C1703)项目批准号:31110303037申请单位:中国农业科学院蚕业研究所研究类型:基础研究收藏负责人:代方银参与人:代方银, 董占鹏, 童晓玲, 胡海, 乔梁, 陈鹏, 曹存, 陈璐诗, 李元浩, 吴松原金额:35万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072088申请单位:西南大学研究类型:基础研究关键词:家蚕;E群突变;定位克隆;Hox基因;躯体模式查看摘要收藏负责人:程廷才参与人:程廷才, 段军, 牛维环, 胡忠雷, 谢小东, 张兴坦, 刘玟杉, 祝慧敏金额:20万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31001034申请单位:重庆大学研究类型:应用基础研究关键词:家蚕;黑胸败血芽孢杆菌;毒素蛋白;结合蛋白;相互作用查看摘要收藏负责人:柴春利参与人:柴春利, 冯丽春, 徐立, 左伟东, 张春冬, 彭丽娜, 张军, 吕军金额:20万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31001035申请单位:西南大学研究类型:基础研究关键词:家蚕、黄红茧、色素、Rc、基因调控查看摘要收藏负责人:范琦参与人:范琦, 王林美, 赵振军, 叶博, 岳冬梅, 张波金额:33万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072081申请单位:辽宁省农业科学院研究类型:基础研究关键词:柞蚕;杆状病毒;细胞凋亡;细胞凋亡抑制蛋白查看摘要收藏负责人:贡成良参与人:贡成良, 薛仁宇, 韩宏岩, 张鹏杰, 梁聪, 薛高旭, 钟金风, 张晓丽, 刘林, 彭景琨金额:33万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072085申请单位:苏州大学研究类型:基础研究关键词:家蚕;Bmovo;卵巢发育;丝蛋白合成;转基因查看摘要收藏负责人:李兵参与人:李兵, 沈卫德, 虞晓华, 许西奎, 王燕红, 王举梅, 林超申请单位:苏州大学研究类型:应用基础研究关键词:家蚕;乙酰胆碱酯酶基因;定点突变;转基因;抗药性查看摘要收藏负责人:李田参与人:李田, 党晓群, 林立鹏, 李治, 黄为, 贾立丽, 王营, 邓远洪, 陈洁金额:20万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31001036申请单位:西南大学研究类型:基础研究关键词:家蚕微孢子虫;Ricin;进化;功能;靶蛋白查看摘要收藏负责人:刘彦群参与人:刘彦群, 武松, 李群, 徐亮, 刘莹, 马楠, 吉尚志, 胡紫菱金额:8万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072082申请单位:沈阳农业大学研究类型:基础研究关键词:柞蚕;滞育解除;KK-42结合蛋白;表达谱;RNAi查看摘要收藏负责人:潘国庆参与人:潘国庆, 黄兰香, 耿莉娜, 罗洁, 胡怀翠, 郭广清, 杨东林, 贺元莉金额:33万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072089申请单位:西南大学研究类型:应用基础研究关键词:家蚕微孢子虫;serpin;;病原与宿主互作;免疫调控查看摘要收藏负责人:时连根参与人:时连根, 金洁, 顾燕燕, 杨东, 黄海山, 陈小攀, 蒋学, 张亚萍, 洪德志申请单位:浙江大学研究类型:基础研究关键词:家蚕;脂动激素受体;信号转导;构效机制查看摘要收藏负责人:谈建中参与人:谈建中, 孙丙耀, 郑必平, 王海玲, 周丽丽, 王廷良金额:32万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072087申请单位:苏州大学研究类型:应用基础研究关键词:桑;雌蕊特征蛋白;特异启动子;差异蛋白质组学;分子克隆查看摘要收藏负责人:王永强参与人:王永强, 孟智启, 陈金娥, 祝新荣, 何秀玲, 柳新菊, 刘培刚金额:33万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072084申请单位:浙江省农业科学院研究类型:应用基础研究关键词:性连锁平衡致死基因;胚胎;蛋白质组;家蚕查看摘要收藏负责人:魏兆军参与人:魏兆军, 李玉萍, 朱绪伟, 田科巍, 汪爱民, 邱娜莎, 潘文娟金额:34万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072091申请单位:合肥工业大学研究类型:基础研究关键词:柞蚕;PBAN;受体;信号转导查看摘要收藏负责人:许金山参与人:许金山, 唐发辉, 张小燕, 和七一, 李治, 王敏, 张玺, 林海静, 王丽君金额:21万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31001037申请单位:重庆师范大学研究类型:基础研究关键词:MITE; 甲基化;微孢子虫;转座子展示;活性查看摘要收藏负责人:赵巧玲参与人:赵巧玲, 陈安利, 张月华, 付凡, 陈蔚, 张婷婷金额:32万申请时间:2010学科代码:养蚕学(C1703)项目批准号:31072083申请单位:中国农业科学院蚕业研究所研究类型:基础研究关键词:家蚕;"明"死卵突变体;基因定位;基因克隆查看摘要收藏负责人:桂仲争参与人:桂仲争, 汪生鹏, 颜辉, 朱雅红, 邹凤鸣, 娄德帅, 吴风瑶金额:30万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30972144申请单位:中国农业科学院蚕业研究所研究类型:基础研究关键词:家蚕;组织蛋白酶D;表达调控;分子机制查看摘要收藏负责人:郭锡杰参与人:郭锡杰, 吴萍, 裘智勇, 刘挺, 侯成香, 赵盼, 王秀金额:30万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30972143申请单位:江苏科技大学研究类型:应用基础研究关键词:家蚕防御质型多角体病毒基因查看摘要收藏负责人:李木旺参与人:李木旺, 侯成香, 孙平江, 郭秋红, 钱荷英, 王修业, 赵云坡, 杨晓博, 梅兴林, 张蕊金额:30万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30972145申请单位:中国农业科学院蚕业研究所研究类型:基础研究关键词:家蚕;茧丝质量;数量性状定位;分子连锁图;单核苷酸多态性查看摘要收藏负责人:童晓玲参与人:童晓玲, 代方银, 康丽芳, 杨微, 胡海, 时桂芹, 陈军金额:20万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30901053申请单位:重庆大学研究类型:基础研究关键词:家蚕;翅型突变;定位克隆;分子机制;基因功能查看摘要收藏负责人:王根洪参与人:王根洪, 徐汉福, 胡忠雷, 牛维环, 解敏敏, 蒋亮, 蒋礼金额:19万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30901054申请单位:重庆大学研究类型:应用基础研究关键词:家蚕;核型多角体病毒(NPV);滞育;piggyBac转座子;RNA干扰查看摘要收藏负责人:缪云根参与人:缪云根, 江丽军, 李兴华, 杨华军, 王丹金额:30万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30972141申请单位:浙江大学研究类型:基础研究关键词:家蚕,糖蛋白,蛋白糖酰化途径,糖苷查看摘要收藏负责人:钟伯雄参与人:钟伯雄, 梁建设, 杨惠娟, 李季生, 庄兰芳, 李建营, 范伟, 危浩金额:30万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30972142申请单位:浙江大学研究类型:应用基础研究关键词:家蚕;组织特异型启动子;启动子陷阱;DNA片段删除法查看摘要收藏负责人:潘敏慧参与人:潘敏慧, 鲁成, 张金叶, 李娜, 田甜, 成传刚金额:30万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30972146申请单位:西南大学研究类型:应用基础研究关键词:家蚕卵巢细胞系;家蚕核型多角体病毒;侵染机制;基因功能查看摘要收藏负责人:赵萍参与人:赵萍, 侯勇, 黄璐琳, 谭祥, 王凌燕, 董照明, 李游山, 衣启营金额:30万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30972147申请单位:西南大学研究类型:应用基础研究关键词:家蚕;丝氨酸蛋白酶抑制剂;基因鉴定;表达特征;生理功能查看摘要收藏负责人:周泽扬参与人:周泽扬, 万永继, 潘国庆, 许金山, 王林玲, 李治, 党晓群, 耿莉娜, 向恒, 李田金额:180万申请时间:2009学科代码:养蚕学(C1703)项目批准号:30930067申请单位:西南大学研究类型:基础研究关键词:家蚕微孢子虫;孢壁蛋白;定位;侵染机制;宿主与病原互作查看摘要收藏负责人:张国政参与人:张国政, 韦亚东, 夏定国, 张业顺, 方瑷, 李翠玲, 王艳敏, 任晓俊金额:30万申请时间:2008学科代码:养蚕学(C1703)项目批准号:30871829申请单位:中国农业科学院蚕业研究所研究类型:应用基础研究关键词:家蚕第二白卵功能基因克隆查看摘要收藏负责人:孙京臣参与人:孙京臣, 严会超, 李茵茵, 刘颖斐, 龙虎, 黄曾慰, 戴兴红, 谭佩婵金额:30万申请时间:2008学科代码:养蚕学(C1703)项目批准号:30871828申请单位:华南农业大学研究类型:应用基础研究关键词:BmCPV;冷冻电子断层扫描;电子显微镜;入侵;机制查看摘要收藏负责人:吴小锋参与人:吴小锋, 曹翠平, 姚慧鹏, 兰丽盼, 何芳青, 郭爱芹, 张有做金额:30万申请时间:2008学科代码:养蚕学(C1703)项目批准号:30871827申请单位:浙江大学研究类型:基础研究关键词:家蚕BmNPV;多角体;蛋白质固定查看摘要收藏负责人:姚勤参与人:姚勤, 郭忠建, 何远清, 陆健, 陈慧卿, 孙晨, 程劲松, 张俊红, 李晓刚, 吕萌金额:30万申请时间:2008学科代码:养蚕学(C1703)项目批准号:30871826申请单位:江苏大学研究类型:基础研究关键词:浓核病毒;中国株;非结构蛋白;病毒复制;家蚕查看摘要收藏负责人:何宁佳参与人:何宁佳, 唐云明, 李斌, 王子龙, 刘碧朗, 刘仕平, 梁九波, 占智高, 唐亮金额:25万申请时间:2008学科代码:养蚕学(C1703)项目批准号:30871825申请单位:西南大学研究类型:基础研究关键词:家蚕;表皮蛋白;几丁质;串联质谱;基因芯片查看摘要收藏负责人:查幸福参与人:查幸福, 向仲怀, 申鸿, 段军, 许平震, 郭靖怡, 李东金额:21万申请时间:2008学科代码:养蚕学(C1703)项目批准号:30800804申请单位:西南大学研究类型:基础研究关键词:家蚕;连锁图;突变;定位查看摘要收藏负责人:刘彦群参与人:刘彦群, 夏润玺, 王欢, 李喜升, 朱绪伟, 马积彪, 亓芳, 叶青雷, 贾利, 田书音金额:20万申请时间:2008学科代码:养蚕学(C1703)项目批准号:30800803申请单位:沈阳农业大学研究类型:基础研究关键词:柞蚕;mtDNA控制区;遗传多样性;遗传结构;系统进化查看摘要收藏负责人:林健荣参与人:林健荣, 廖富蘋, 王叶元, 刘伟强, 林碧敏, 刘奎, 范兰芬, 文凤云, 孔庆明金额:33万申请时间:2007学科代码:养蚕学(C1703)项目批准号:30771634申请单位:华南农业大学研究类型:基础研究关键词:家蚕;表皮蛋白基因;表达调控机理查看摘要收藏负责人:朱良均参与人:朱良均, 闵思佳, 林蓉, 张海萍, 丰强, 高欣金额:30万申请时间:2007学科代码:养蚕学(C1703)项目批准号:30771633申请单位:浙江大学研究类型:应用基础研究关键词:蚕丝蛋白;化纤;功能纤维材料;粘接复合;机理查看摘要收藏负责人:徐世清参与人:徐世清, 司马杨虎, 陈息林, 杨德印, 王更先, 郑必平, 王英, 朱晓苏, 宋艳金额:30万申请时间:2007学科代码:养蚕学(C1703)项目批准号:30771632申请单位:苏州大学研究类型:应用基础研究关键词:家蚕;滞育;生物钟;基因互作;分子机制查看摘要收藏负责人:汪生鹏参与人:汪生鹏, 沈兴家, 唐顺明, 汪伟, 王兴科, 朱晨金额:8万申请时间:2007学科代码:养蚕学(C1703)项目批准号:30771631申请单位:中国农业科学院蚕业研究所研究类型:应用基础研究关键词:蚕丝蛋白基因;时期特异性表达;反式因子;蜕皮激素受体;酵母单杂交技术查看摘要收藏负责人:鲁成参与人:鲁成, 潘敏慧, 余全友, 付雍, 洪开丽, 许湲, 王先燕金额:30万申请时间:2007学科代码:养蚕学(C1703)项目批准号:30771630申请单位:西南大学研究类型:应用基础研究关键词:家蚕;原代培养;细胞系;基因表达谱;丝腺查看摘要收藏负责人:代方银参与人:代方银, 沈以红, 童晓玲, 谭端, 胡海, 黄飞飞, 邓琼珍金额:30万申请时间:2006学科代码:养蚕学(C1703)项目批准号:30671591申请单位:西南大学研究类型:应用基础研究关键词:家蚕;新突变;基因定位;分子标记;连锁图谱查看摘要收藏负责人:贡成良参与人:贡成良, 潘中华, 周文林, 张梅, 张鹏杰, 贾海芳, 刘波, 诸戌娴, 刘炜彬, 杨贺金额:26万申请时间:2006学科代码:养蚕学(C1703)项目批准号:30671590申请单位:苏州大学研究类型:应用基础研究关键词:转基因家蚕;egt;蝎毒素基因;蛹期控制查看摘要收藏负责人:楼程富参与人:楼程富, 陆小平, 潘刚, 叶志毅, 岳万福, 沈飞英金额:24万申请时间:2006学科代码:养蚕学(C1703)项目批准号:30671589申请单位:浙江大学研究类型:应用基础研究关键词:桑树;抗冻蛋白;转基因;功能验证;低温应答查看摘要收藏负责人:刘吉平参与人:刘吉平, 陈芳艳, 黄志君, 郝娟, 李敬霞, 王星, 张平, 张莉金额:27万申请时间:2006学科代码:养蚕学(C1703)项目批准号:30671588申请单位:华南农业大学研究类型:基础研究关键词:桑蚕;微孢子虫;垂直传播;分子进化;系统发育查看摘要收藏负责人:张泽参与人:张泽, 曹致琦, 段彪, 余泉友, 黄飞飞金额:30万申请时间:2006学科代码:养蚕学(C1703)项目批准号:30671587申请单位:西南大学研究类型:基础研究关键词:家蚕;野桑蚕;抗菌肽;分子进化;分子群体遗传查看摘要收藏负责人:潘国庆参与人:潘国庆, 李继祥, 李艳红, 吴正理, 杨柳, 刘铁, 李田, 谢俪, 庞敏金额:21万申请时间:2006学科代码:养蚕学(C1703)项目批准号:30600452申请单位:西南大学研究类型:应用基础研究关键词:家蚕微孢子虫;孢壁蛋白;侵染;免疫定位;糖基化位点查看摘要收藏负责人:程道军金额:27万申请时间:2005学科代码:养蚕学(C1703)项目批准号:30571407申请单位:西南大学研究类型:应用基础研究查看摘要收藏负责人:时连根金额:24万申请时间:2005学科代码:养蚕学(C1703)项目批准号:30571406申请单位:浙江大学研究类型:基础研究查看摘要收藏负责人:司马杨虎金额:23万申请时间:2005学科代码:养蚕学(C1703)项目批准号:30571405申请单位:苏州大学研究类型:应用基础研究查看摘要收藏负责人:贡成良金额:25万申请时间:2005学科代码:养蚕学(C1703)项目批准号:30571404申请单位:苏州大学研究类型:应用基础研究查看摘要收藏负责人:魏兆军金额:23万申请时间:2005学科代码:养蚕学(C1703)项目批准号:30500374申请单位:合肥工业大学研究类型:基础研究查看摘要收藏负责人:赵萍金额:21万申请时间:2004学科代码:养蚕学(C1703)项目批准号:30471313申请单位:西南大学研究类型:应用基础研究查看摘要收藏负责人:鲁成金额:23万申请时间:2004学科代码:养蚕学(C1703)项目批准号:30471312申请单位:西南大学研究类型:应用基础研究查看摘要收藏负责人:鲁兴萌金额:20万申请时间:2004学科代码:养蚕学(C1703)项目批准号:30471311申请单位:浙江大学研究类型:应用基础研究查看摘要收藏负责人:苗雪霞金额:19万申请时间:2004学科代码:养蚕学(C1703)项目批准号:30471310申请单位:中国科学院上海生命科学研究院研究类型:基础研究查看摘要收藏负责人:沈卫德金额:20万申请时间:2004学科代码:养蚕学(C1703)项目批准号:30471309申请单位:苏州大学研究类型:应用基础研究查看摘要收藏负责人:李春峰金额:21万申请时间:2004学科代码:养蚕学(C1703)项目批准号:30400328申请单位:西南大学研究类型:基础研究查看摘要收藏负责人:李木旺金额:19万申请时间:2004学科代码:养蚕学(C1703)项目批准号:30400327申请单位:中国农业科学院蚕业研究所研究类型:基础研究查看摘要收藏负责人:刘朝良金额:16万申请时间:2003学科代码:养蚕学(C1703)项目批准号:30371088申请单位:安徽农业大学研究类型:应用研究查看摘要收藏负责人:孟智启金额:20万申请时间:2003学科代码:养蚕学(C1703)项目批准号:30371087申请单位:浙江省农业科学院研究类型:应用研究查看摘要收藏负责人:徐世清金额:19万申请时间:2003学科代码:养蚕学(C1703)项目批准号:30371086申请单位:苏州大学研究类型:应用研究查看摘要收藏负责人:周泽扬金额:22万申请时间:2003学科代码:养蚕学(C1703)项目批准号:30371085申请单位:西南大学研究类型:基础研究查看摘要收藏负责人:向仲怀金额:145万申请时间:2003学科代码:养蚕学(C1703)项目批准号:30330460申请单位:西南大学研究类型:应用研究查看摘要收藏负责人:李斌金额:20万申请时间:2003学科代码:养蚕学(C1703)项目批准号:30300262申请单位:西南大学研究类型:基础研究查看摘要收藏负责人:张志芳金额:19万申请时间:2002学科代码:养蚕学(C1703)项目批准号:30271007申请单位:中国农业科学院蚕业研究所研究类型:基础研究查看摘要收藏负责人:万永继金额:20万申请时间:2002学科代码:养蚕学(C1703)项目批准号:30271006申请单位:西南大学研究类型:应用基础研究查看摘要收藏负责人:鲁成金额:21万申请时间:2002学科代码:养蚕学(C1703)项目批准号:30271005申请单位:西南大学研究类型:基础研究查看摘要收藏负责人:钟伯雄金额:19万申请时间:2002学科代码:养蚕学(C1703)项目批准号:30271004申请单位:浙江大学研究类型:基础研究。
雄蚕种质量标准与研究

雄蚕种质量标准与研究
雄蚕是蚕丝生产中不可或缺的重要资源,其种质量标准的制定和研究对于提高蚕丝产量和质量具有重要意义。
一、雄蚕种质量标准1.外形标准:雄蚕体长应在2.5-3.5厘米之间,体重应在0.5-1.5克之间,身体呈圆柱形,头部稍微扁平,触角长而细。
2.生长发育标准:雄蚕的生长发育应该健康,幼虫期应在25-30天之间,蛹期应在8-10天之间,成虫期应在5-7天之间。
3.生殖能力标准:雄蚕的生殖能力是评价其种质量的重要指标之一。
雄蚕应该具有良好的交配能力和精液质量,能够产生高质量的种子。
4.遗传标准:雄蚕的遗传标准是评价其种质量的重要指标之一。
雄蚕应该具有优良的遗传基因,能够传递良好的性状和性能。
二、雄蚕种质量研究1.遗传改良:通过选择和育种,选出具有优良遗传基因的雄蚕,进行交配和繁殖,以提高蚕丝产量和质量。
2.繁殖技术:通过优化繁殖技术,提高雄蚕的繁殖效率和生殖能力,以提高种质量和蚕丝产量。
3.疾病防治:对雄蚕的疾病进行防治,保证其健康生长和繁殖,以提高种质量和蚕丝产量。
4.营养管理:通过合理的饲养和营养管理,提高雄蚕的生长发育和生殖能力,以提高种质量和蚕丝产量。
总之,雄蚕种质量标准的制定和研究对于提高蚕丝产量和质量具有重要意义,需要通过遗传改良、繁殖技术、疾病防治和营养管理等方面的努力,不断提高雄蚕的种质量和生产效益。
加强桑蚕饲养技术分析与病害有效防控研究

加强桑蚕饲养技术分析与病害有效防控研究【摘要】桑蚕产业是中国重要的农业产业之一,加强桑蚕饲养技术与病害防控研究对其发展至关重要。
本文从桑蚕饲养技术分析和病害防控技术研究入手,探讨了如何提升桑蚕饲养效率和有效防控病害,提出了疫情防治策略。
研究结果表明,加强桑蚕饲养技术与病害防控对提高桑蚕产量和质量具有重要意义。
未来,应加强对桑蚕饲养技术和病害防控的研究,提高产业可持续发展能力。
通过本研究,桑蚕养殖业能够更加稳定和可持续地发展,为农民增收和经济发展做出贡献。
【关键词】桑蚕饲养技术、病害防控、效率提升、病害防治、疫情防治、研究成果、未来研究方向、重要性、桑蚕、饲养、防控、研究、疫情、技术、分析、措施、策略。
1. 引言1.1 研究背景桑蚕饲养技术一直是桑蚕产业链中至关重要的环节,直接关系到桑蚕的生长发育和产量质量。
随着农业生产技术的不断进步和科学研究的深入,桑蚕饲养技术也在不断完善和提高。
当前桑蚕饲养技术依然存在一些问题和挑战,例如养殖成本较高、养蚕效率不高、病害防控不到位等。
加强桑蚕饲养技术分析与病害有效防控研究,对于提升桑蚕产业的发展水平,保障养蚕农民的经济收入,具有重要的现实意义和深远影响。
在当前经济社会发展的背景下,桑蚕产业已成为我国农业产业结构调整的重要组成部分。
随着桑蚕产业的不断发展壮大,桑蚕饲养技术和病害防控技术的研究和应用也更加引人关注。
对桑蚕饲养技术进行深入分析,研究病害的有效防控措施,提升桑蚕饲养效率,成为当前桑蚕产业发展中急需解决的问题之一。
1.2 研究目的研究目的是为了深入探讨加强桑蚕饲养技术与病害有效防控研究的重要性,进一步提高桑蚕养殖的效率和质量,保障桑蚕产业的可持续发展。
通过对桑蚕饲养技术进行分析和总结,找出存在的问题和瓶颈,并提出改进措施和方法,以提升桑蚕的生长速度和产量,提高产品的质量和市场竞争力。
通过对桑蚕病害的研究和防控措施的探讨,促进桑蚕疫情的有效防控,降低损失,保障养殖稳定生产。
2022-2023学年天津市宁河区芦台一中高一5月月考生物试题

2022-2023学年天津市宁河区芦台一中高一5月月考生物试题1.孟德尔采用的假说—演绎法是现代科学研究中常用的一种方法,下列属于孟德尔在发现基因分离定律时的“演绎”过程的是()A.生物的性状是由遗传因子决定的B.由F 2出现了“3∶1”推测,生物体产生配子时成对遗传因子彼此分离C.若F 1产生配子时成对遗传因子分离,则测交后代会出现两种性状,比例接近1∶1 D.若F 1产生配子时成对遗传因子分离,则F 2中三种基因型比例接近1∶2∶12.一般人对苯硫脲感觉有苦味,由显性基因B控制;也有人对其无味觉,叫味盲,由隐性基因b控制。
统计味盲家族,若三对夫妇的子女味盲的概率各是25%、50%和100%,则这三对夫妇的遗传因子组成最可能是( )①BB×BB ②bb×bb ③BB×bb ④Bb×Bb ⑤Bb×bb ⑥BB×BbA.①②③B.④⑤⑥C.④⑤②D.④②⑤3.玉米甜和非甜是一对相对性状,随机取非甜玉米和甜玉米进行间行种植,其中一定能够判断甜和非甜的显隐性关系的是()A.B.C.D.4.小麦的高秆对矮杆为显性,抗病对不抗病为显性,用纯种的高秆抗病和矮杆不抗病杂交得到F1,F1自交得到F2,在F2的矮杆抗病植株中能稳定遗传的植株占比为()A.1/3 B.9/16 C.1/16 D.3/165.生物学是一门实验科学,选择合适的实验材料是实验成功与否的先决条件。
下列有关实验选材的叙述错误的是()A.豌豆有明显的相对性状,自花传粉、闭花授粉,可用于研究伴性遗传B.果蝇繁殖能力强,子代数目多,易于做数学统计分析C.山柳菊有时进行无性生殖,有时进行有性生殖,不适合做遗传学实验的材料D.洋葱根尖分生区细胞分裂旺盛,利于观察有丝分裂过程中染色体形态的变化6.某男孩的基因型为X A Y A,正常情况下,其A基因不可能来自于A.外祖父B.外祖母C.祖父D.祖母7.家蚕(性别决定方式为ZW型)刚孵化时形如蚂蚁,故称为蚁蚕。
家蚕性染色体和性别决定

家蚕性染色体和性别决定刘增虎;李涛;杨海;刘敏;董占鹏【摘要】家蚕是重要的经济昆虫和鳞翅目昆虫的典型代表,由于生产上专养雄蚕和农业害虫防治有极好的发展前景,对其性别决定机制的研究极为迫切.对家蚕的性染色体及其结构特征和性别调控相关基因的研究现状进行了综述.%Silkworm is an important economic insect and a typical representative of Lepidoptera. It is stringent and important to research the mechanism of silkworm sex determination due to its excellent prospects in exclusive male sericulture and agriculture pest control. The current research of silkworm sex chromosome, structural feature and genes relate to sex regulation were reviewed.【期刊名称】《安徽农业科学》【年(卷),期】2012(040)034【总页数】5页(P16627-16631)【关键词】家蚕;性染色体;性别决定【作者】刘增虎;李涛;杨海;刘敏;董占鹏【作者单位】云南省农业科学院蚕桑蜜蜂研究所,云南蒙自 661101;云南省农业科学院蚕桑蜜蜂研究所,云南蒙自 661101;云南省农业科学院蚕桑蜜蜂研究所,云南蒙自 661101;云南省农业科学院蚕桑蜜蜂研究所,云南蒙自 661101;云南省农业科学院蚕桑蜜蜂研究所,云南蒙自 661101【正文语种】中文【中图分类】S882.3昆虫性别决定是经过长期进化选择产生的结果,是昆虫生命活动的基本特征,影响个体发育的许多方面(如胚胎发生、性别分化、生殖等生理活动过程),并且最终表现为形态学、生理学以及行为特征上的二态化现象[1]。
蚕桑生物工程研究进展

蚕桑生物工程研究进展
孟智启;计东风
【期刊名称】《蚕桑通报》
【年(卷),期】1989(020)001
【摘要】蚕桑生物工程是当代蚕业科学研究的新领域。
现已取得某些振奋人心的
成果并展现出灿烂前景。
本文将国内外近年来蚕桑生物工程研究的进展作一概述。
一、桑蚕生物工程研究现状及展望蚕作为一种经济昆虫为人类利用巳有数千年历史。
随着研究成果的推广应用,蚕的生产水平有了很大程度的提高。
但由于生物的生理
平衡特性,依靠传统的研究手段和技术体系,已不可能再大幅度提高其生产水平和利
用价值。
因此各蚕业发达国家都相继开展了生物工程在桑蚕中的应用研究。
【总页数】5页(P1-5)
【作者】孟智启;计东风
【作者单位】不详;不详
【正文语种】中文
【中图分类】S881
【相关文献】
1.煤层气生物工程研究进展 [J], 苏现波;夏大平;赵伟仲;伏海蛟;郭红光;何环;鲍园;
李丹;魏国琴
2.微生物谷氨酰胺转氨酶的来源与生物工程技术研究进展 [J], 张秀江;权淑静;向凌云;刘丽;解复红;冯菲;胡虹
3.以新冠肺炎疫情为案例的问题导向式课程思政教学——以“生物工程研究进展”
为例 [J], 姜良珍;李建;万燕;李云成;向达兵;文永平;刘达玉
4.以新冠肺炎疫情为案例的问题导向式课程思政教学——以"生物工程研究进展"为例 [J], 姜良珍;李建;万燕;李云成;向达兵;文永平;刘达玉
5.基于土壤生物工程的小流域侵蚀沟治理研究进展 [J], 冷瑜;曹丽娜;张振兴;孙伟;杨海军;林晨鹭;高玉福
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
vr t setni h j n rvne i w sas u gs d hen w s k om vr t s r ̄ igojcs o ai i xes ni Z el gpo ic , t a l sge t t e i w r ai i edn betfr ee o n a o e l eeb
一
2 黄衍锋
秋种春养优越性和推广前景的探讨 . 蚕桑通
报 .0 l3 ( )4 2 O ,24 :5—4 7
表述 。笔者认 为在 把握 新蚕 品种选 育 的前
3 靳远祥 ,陈玉银 等 . 浙江省现行夏秋用桑蚕品种比轻
试验 蚕桑通报 . ̄03 ( :6 2 2o . l)2 — 8 t 中 4 楼 黎静 .沈玉丽等 夏秋蚕品种比较试验与调查
目 蚕韭 . .2 :4 2 ()4
提下 ,通过大家的努力,对 目前培育的蚕 品种
进行不断的完善,一定 能培育 出适合 目前 蚕桑
T eN w S k om V r tsB edn betn Z ei P oic C a Ⅲ f e uu dRs r h e i w r ai i reigO jc i hj  ̄ rv e ho l ee a n St lr e a h e i  ̄c ec I tu ,脚 nit ste Aae yo d 砌 出 Si c,H r o 101 hn) cdm 占 f “ c ne a e  ̄ u302 ,Ci a
维普资讯
廉海燕等 : 利用温敏致死性状专养雄蚕 的研 究进展
‘ 5‘
的 F代, l 在高温干燥 (  ̄、0 H 条件催 3 C 6%R ) 0 青, 赤蚁雌蚕的孵化率很低 , 只有 5 %左右 , 伴性品系 s ,与普通品种新九雌体杂交的 c h F 代新九 x c , 1 h 同样具有伴性温敏致死性。 s 在正常 温湿 度 (5 、 0 H)条件下催 青 . 2℃ 8 %R 雌雄个体正常孵化 ,在高温干燥条件下催青 时, 雌蚕的孵化率极度下降 , 而雄蚕则基本不 受影响, 杂交种的雌雄性 比得到控制 】 , 由 此开创了实现雄蚕饲育的新途径。 11 温敏性表达的时期 . 伴性 赤 蚁胚 胎发 育对 高温 、干燥环 境反 应敏感的时期主要为后期 “。 l 胚胎对高温 最敏感的时期 为气管显现期 ( , 对干燥最 己 ) , 敏感的时期为转青期 ( ,,这两个时期雌 己 ) 雄个体表现了显著的反应差异 ,因此分别将 己 ,己 , 为高 温敏感 期 和干燥敏感 期 。 、 称 12 催青 处理 条件 . 从高温 敏感 期 ( ,起 用 3℃ 、 0 R 己 ) 0 6% H 催青, 就可达 到基 本 抑制雌 蚕孵化 的效果 , 在 己3 ~己 期 间用 3  ̄ 8%R 0C、 0 H保护 , 己 ~ 孵化期 改用 3 ̄ 6%R 0C、0 H保护 , 也能有 效地 抑制 雌蚕孵 化 。
维普资讯
蚕 桑 通报 B l t f eiu ue3 ( ) ul i o r h r 3 1 4—7 en S c
利 用 温敏 致 死 性状 专养 雄 蚕 的 研究 进展
颜 海燕 扬 明观 钟伯雄
( 浙江大学动物科学学 院蚕蜂科学系 , 杭州 30 2 109
Ab ta t t Ss g e td te [ e r n e o re ign w ik r v rei n o uai n J s aite sl c I Wa u g se h mp t c fb e d n e slwom ait a d p p lr i gt0 ev reis ' a s e z 1 frp o t n o es rc l r n u ty i h rs n a e .A ̄ odn o te p ee tst t n a ik r o rmoi ft e lut e i d sr n t ep ee tp p r o h u r ig t h rs n i i f lwo m ua o s
tedfee tslw r raig e vrn na O dt n n es n . h i rn i om e rn n io me tlC n io sa d s ao s k l
Ke (d i w r y w 1 s Sl om N w vr t r dn bet r k e a e B e i ojc iy e g
摘
要 伴性赤蚁 ( ) 有催青期高温于燥 敏感性 , 3 、 具 用 相对湿度 6 %条件催青时雌卵孵化 0
率很j 雄卵正常孵化 。利用 品种作 父本 与普通蚕品种杂交 , F 代 的蚕卵仍具有高温催青敏 氐. 其 1 感性 . 这是控制桑蚕性别实现雄蚕 饲养的一条新途径 。
簧键词 桑蚕 雄蚕 胚胎 温敏性
1 伴性赤蚁蚕胚胎期的温敏性表现
12 9 年华南农业大学潘庆 中等发 现正 9 常黑蚁品种雌体与伴性赤蚁 ( ) 雄体杂交
生产和市场需要的桑蚕新 品种 。 参 考 文 献
1 沈永 泉,沈坤元 等 . 谈春养夏秋蚕种稳定蚕桑生产
要课题之一。19 年潘庆中等发现伴 92
性赤蚁 (c s h)具 有胚胎期 温敏 致死 性 “ .
的桑蚕新 品种研究 ,也只是从浙江省农科院蚕
桑所这 一侧面初 浅地 分 析探讨今 后 的蚕 品种 选 育方 向,众所周知浙江大学蚕蜂科 学系 、湖 州市蚕研 所在 蚕 品种选育 上也 有很 多技术 积 累 ,育 成 了不 少 品种 ,有 的 已在生 产上推广 应用 ,有 的正 在生 产上进行 试养 ,在此不一
雄蚕与雌蚕相 比,具有体质强健 、饲育 容易 、食桑量 少 、出丝 率高 、纤 度细 、丝质 好等优点 , 适于缫制高品位生丝, 具有较高的 经济价值 I } t o据统计专养雄蚕 比雌雄蚕混 , 2 养, 产丝量可增加 1%, 5 提高经济效益 1% 0 左右。因此在蚕业生产上实现专养雄蚕是 2 l 世纪 家蚕育 种 的主要 方 向之一 ,进一 步加 强
雄蚕 品种 的选育 和推广 也是蚕 业科 学基础研
由于这个特性可实现专养雄蚕,因而广大学 者迅速 开展 了相关 的研究 ,对温 敏致死 的表 达特性 、生理机制及育种上 的利用做了大量 的探讨 , 前 已初步育成伴性温敏实用品种, 目 研 究表明这是 实现专养雄蚕 的一条较好途
径。