发育生物学第七章胚轴形成
8 胚轴形成

几个概念
分化:从单个全能的受精卵产生各种类型细胞 的发育过程 定型:在分化前,产生一些隐蔽的变化,使细胞朝 特定方向发展,这一过程 自主特化,条件特化 诱导:胚胎一个区域对另一区域发生影响并使 后者沿一条新途径分化的作用
1.图式形成(pattern plan): 胚胎细胞形成不同组织,器官,构成有序空间结构 的过程 2.胚轴:前—后 背—腹轴 至少一种主要的
Nieuwkoop中心
原肠期间新形成的中胚层是由其下方的植物极预定内 胚层细胞对动物极外胚层诱导产生的(产生的是背部 中胚层或者腹侧中胚层要看植物极内胚层细胞的背- 腹极性决定) Nieuwkoop中心是由于精子入卵产生的兼有动物极和 植物极细胞质的特殊区域,该区域含有背部中胚层诱 导信号
第一节:果蝇胚轴形成第二节:两栖类胚轴形成
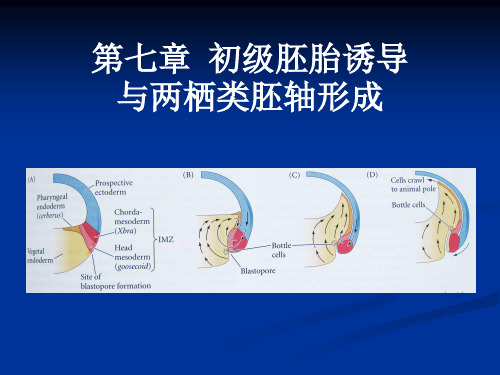
两栖类是调整型发育的典型模式。 两栖类的背-腹轴和前-后轴是受精时卵质的 重排分布而决定的。受精时细胞质的剧烈运动; 灰色新月;胚孔;背唇;原肠运动的启动
一、组织者和Nieuwkoop中心
组织者 蝾螈受精卵结扎试验说明的问题: 1蝾螈早期卵裂球的细胞核有全能性 2由灰色新月区产生的背唇启动了原肠运动 3两栖类早期原肠除了背唇具有自主分化能力外,其他组织发育的命运并 未决定,但到了原肠晚期所有组织发育命运都已经决定 组织者的作用: 1启动原肠作用 2组织者能发育成背部中胚层(脊索中胚层) 3组织者诱导外胚层形成背神经管 4组织者能诱导其周围的中胚层分化为侧板中胚层而不是腹侧中胚层
发育生物学1—7章 课后习题答案

《发育生物学》课后习题答案绪论1、发育生物学的定义,研究对象和研究任务?答:定义:是应用现代生物学的技术研究生物发育机制的科学。
研究对象:主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老死亡,即生物个体发育中生命现象发展的机制。
同时还研究生物种群系统发生的机制。
2、多细胞个体发育的两大功能?答:1.产生细胞多样性并使各种细胞在本世代有机体中有严格的时空特异性;2.保证世代交替和生命的连续。
3、书中所讲爪蟾个体发育中的一系列概念?答:受精:精子和卵子融合的过程称为受精。
卵裂:受精后受精卵立即开始一系列迅速的有丝分裂,分裂成许多小细胞即分裂球,这个过程称为卵裂。
囊胚:卵裂后期,由分裂球聚集构成的圆球形囊泡状胚胎称为囊胚。
图式形成:胚胎细胞形成不同组织,器官和构成有序空间结构的过程胚轴:指从胚胎前端到后端之间的前后轴和背侧到腹侧之间的背腹轴4、模式生物的共性特征?答:a.其生理特征能够代表生物界的某一大类群;b.容易获得并易于在实验室内饲养繁殖;c.容易进行试验操作,特别是遗传学分析。
5、所讲每种发育生物学模式生物的特点,优势及其应用?答:a.两粞类——非洲爪蟾取卵方便,可常年取卵,卵母细胞体积大、数量多,易于显微操作。
应用:最早使用的模式生物,卵子和胚胎对早期发育生物学的发展有举足轻重的作用。
b.鱼类——斑马鱼受精卵较大,发育前期无色素表达,性成熟周期短、遗传背景清楚。
优势:a,世代周期短;b,胚胎透明,易于观察。
应用:大规模遗传突变筛选。
c.鸟类——鸡胚胎发育过程与哺乳动物更加接近,且鸡胚在体外发育相对于哺乳动物更容易进行试验研究。
应用:研究肢、体节等器官发育机制。
d.哺乳动物——小鼠特点及优势:繁殖快、饲养管理费用低,胚胎发育过程与人接近,遗传学背景较清楚。
应用:作为很多人类疾病的动物模型。
e.无脊椎动物果蝇:繁殖迅速,染色体巨大且易于进行基因定位。
酵母:单细胞动物,容易控制其生长,能方便的控制单倍体和二倍体间的相互转换,与哺乳动物编码蛋白的基因有高度同源性。
7 胚轴形成

在卵子发生过程中,这些母源效应基因的mRNA 由滋养细胞合成转运至卵子,定位于卵子的一定 区域。这些mRNA在受精后立即翻译编码蛋白因 子,激活或抑制一些合子基因的表达,调控果蝇 胚轴的形成。 这些母源效应基因的蛋白质产物又称为形态发生 素(morphogen)。
形态发生素(morphogen)?
Torso系统基因的失活会导致胚胎不分节的部分,即前 端原头区和后端尾节,缺失
tor基因编码一种跨膜酪氨酸激酶受体 (receptor tyrosine kinase,RTK),在整个 合胞体胚胎的表面表达。 其NH2-基端位于细胞膜外,-COOH基端位 于细胞膜内。当胚胎前、后端细胞外存在 某种信号分子(配体)时可使TOR特异性 活化,最终导致胚胎前、后末端细胞命运 的特化。torso-like (tsl)基因可能编码这 一配体。
NANOS蛋白和CAUDAL蛋白浓度梯度 后端系统包括约10个基因,这些基因的突 变都会导致胚胎腹部的缺失。在这一系统 中起核心作用的是nanos(nos)基因。 后端系统在控制图式形成中起的作用与前 端系统有相似之处,但发挥作用的方式与 前端系统不同。
后端系统并不像BCD蛋白那样起指导性的 作用,不能直接调节合子基因的表达,而 是通过抑制一种转录因子的翻译来进行调 节。 在果蝇卵子发生过程中,nos mRNA定位于 卵子后极。nos基因的编码产物NANOS (NOS)蛋白活性从后向前弥散形成一种 浓度梯度。NOS蛋白的功能是在胚胎后端 区域抑制母性hb mRNA的翻译。
浓度梯度建立位 置信息的模型
图式建成依赖于细胞对位置信息的译读
法国国旗模式
每个细胞位置信息: 获得位置值(positional value),这个值决定它的位置 (两端的边界) 细胞译读位置信息
发育生物学期末复习题集

1.定义:发育生物学是一门研究生物体从精子和卵子发生、受精、发育、生长到衰老、死亡规律的科学。
2. 发育生物学研究的主要任务:生物体发育的遗传程序及调控机制。
3. 发展基础:胚胎学、遗传学、细胞生物学, 是多学科相互渗透的结果。
第一章生殖细胞的发生一、选择题:()1.精子的顶体来自精子细胞的。
A.生发泡;B.高尔基体;C.线粒体;D.疏松泡。
()2.在精子发生过程中,由同一个精原细胞产生的生精细胞A.发育不同步;B.位于生精小管的同一位置;C.称为连接复合体;D.组成一个滤泡。
()3.以下生精细胞中,哪一类是干细胞?A.A1型精原细胞;B.B型精原细胞;C.中间型精原细胞;D.精子细胞。
()4.在昆虫卵母细胞中,mRNA和蛋白质是由。
A.卵母细胞自身合成的;B.滤泡细胞合成的;C.抚育细胞合成的;D.卵室以外的细胞合成的。
()5.一个初级卵母细胞经过两次成熟分裂,形成几个卵子?A.一个;B.二个;C.三个;D.四个。
()6.在一个昆虫的卵室中,含有 A.一个卵母细胞;B.二个卵母细胞;C.16个抚育细胞; D.15个滤泡细胞。
()7.人类出生时,卵巢中的卵母细胞处于 A.偶线期;B.粗线期;C.双线期;D.细线期。
二、判断题:()1.在滋卵型卵子发生过程中,抚育细胞为卵母细胞的发育提供了许多RNA、蛋白质和核糖体等。
()2.两栖类卵子外方的卵黄膜中含有ZP1、ZP2和ZP3等蛋白质。
()3.哺乳动物精子的中心粒位于尾部。
三、填空题:1.生精细胞的整个发育过程离不开细胞。
2.顶体中含有多种。
3.在精子发生过程中,当精子细胞的核浓缩之后,基因就停止。
1.MPF由两种成分组成,分别是和。
四、名词解释:1.配子发生2.精子形成3.顶体4.支持细胞5.透明带第二章受精的机制精卵识别精子与滤泡细胞、ZP和卵质膜在3个水平上独立的准确的相互作用。
顶体反应:精子顶体水解酶与卵子的ZP蛋白相互作用,使精卵相互黏附,同时进行膜的合。
发育生物学

第二类组织者分泌性蛋白因子和WNT抑制因子
咽鳃区内胚层和头部中胚层形成的内-中胚层 组织构成胚孔背唇的前缘。这些细胞不仅能 诱导最前端的头部结构,而且还能够阻滞 WNT信号传导途径。WNT家族的成员之一 WNT8与BMP4相似,能够抑制神经诱导。 组织者分泌的CERBERUS(CBR)和内中胚 层分泌的FRZB和DICKKOPF(DCK)蛋白 因子可以阻止WNT信号传导途径。
背部中胚层诱 导形成组织者 的可能机制
gcd基因的激活也同时受到定位于植物半球和 Nieuwkoop中心的TGF-β家族蛋白产物 (Vg1、VegT和Nodal相关蛋白)的协同作用。 β-CATENIN与VG1、VEGT信号相互作用的 结果形成Nodal-相关蛋白从背侧到腹侧的浓 度梯度。Nodal-相关蛋白的浓度梯度使中胚 层细胞分化。含有大量Nodal相关蛋白的分化 为组织者,较少的分化为侧板中胚层,不含 Nodal相关蛋白的分化成腹侧中胚层。
BMP4是最重要的上皮分化和腹侧化诱导因 子。对于神经发生而言,它是抑制因子或抗 神经化因子,其功能与组织者正好相反。 组织者分泌的NGN、CHD、Nodal相关蛋白 -3和FST对BMP4均有抑制作用。因此组织 者的功能是分泌具有抑制作用的可溶性蛋白 因子,而不是直接诱导中枢神经系统的发生。
爪蟾卵经紫外线照射 后引起背部结构的缺 失,可以由注射NGN 蛋白而挽救。NGN对 背部结构的诱导呈剂 量依赖性。
后端化因子 RA的作用
中侧轴特化因子
脊椎动物中侧轴形成的关键事件都是原肠 胚左侧侧板中胚层表达的nodal-相关基因调 控。 爪蟾的nodal-相关基因-1(Xnr-1)的表达区 域仅限于原肠胚左侧,Xnr-1激活pitx-2在胚 胎左侧专一性表达。VG1蛋白可以增加胚胎 左右对称的特化,异位表达VG1可导致胚胎 左右逆转。
发育生物学第七章- 分节基因与胚胎体节的形成

8. 分节基因与胚胎体节的形成分节基因的功能是将早期胚胎沿前后轴分为一系列重复的体节原基。
根据分节基因的突变表型及作用方式可分为3类:缺口基因(gap gene ,间隙基因),成对控制基因和体节极性基因。
这三类基因的调控是逐级进行的。
BICOID (BCD )HUNCHBACK (HB )NANOS (NOS )CAUDAL (CDL )gap genepair -rule genesegment gene homeotic gene首先,母体效应基因控制gap 基因的活化;其次gap 基因之间互相调节彼此的转录且共同调节成对控制基因的表达;然后,成对控制基因之间相互作用,把胚体分隔为一系列重复的体节,并且进一步控制体节极性基因的表达。
gap 基因和成对控制基因再共同调控同源异型基因的表达。
最终,每个体节原基都具有其独特的基因表达组合,从而决定每个体节的特征。
9.gap 基因gap 基因表达区域的宽度约相当于3个体节,表达区之间可有部分重叠。
最初,gap 基因通常在整个胚胎中都有微弱表达,随着卵裂的进行逐渐变成一些不连续的区域。
gap 基因的表达最初由母体效应基因启动,其表达图式的维持可能依赖于gap 基因之间的相互作用。
gap 基因hunckback(hb),kruppel(Kr),giant(gt),tailess(tll),knirps(kni),huckebein(hkb),empty spiracles(ems)缺口基因蛋白对even-skipped 基因表达的调控。
-1070 bp-1550 bp10. 成对控制基因成对控制基因表达区域以2个体节为单位且具有周期性,在相互间隔的一个副体节中表达。
这些基因的功能是把gap 基因确定的区域进一步分为体节。
成对控制基因的表达是胚胎出现分节的最早标志。
成对控制基因在13次卵裂时开始表达,沿前后轴形成一系列斑马纹状的条带,正好把胚胎分为预定的体节。
发育生物学胚轴的特化与体轴的建立

• 母体基因:在卵子发生过程中表达,并在 卵子发生及早期胚胎发育中具有特定功能 的基因 • 合子基因(zygotic gene) 受精卵在 胚胎发育过程中表达的基因称为合子基因 ,受母体基因产物的激活。合子基因的表 达是胚胎模式形成的开端。合子基因分为 若干组,表达顺序有先有后,它们之间有 着严密的连锁关系,协调有序地调控着胚 胎发育的模式形成。
一、果蝇前 – 后轴的形成
1. 果蝇前后极性的产生
• 果蝇的胚胎,幼虫、成体的前后极性均来源 于卵子的极性。 • 调节胚胎前 – 后轴的形成有4个重要基因:
BICOID(BCD)、HUNCHBACK(HB) 调节胚胎 前端结构的形成; NANOS(NOS)、CAUDAL(CDL)调节胚胎后端 结构的形成。
Torso系统基因的失活会导致胚胎不分节的部分,即前 端原头区和后端尾节,缺失
卵膜表面Torso的激活决定胚胎AP轴的两个端点
Torso蛋白为酪氨酸激酶类受体。 未受精前,torso已均匀地分 布在卵的质膜上。但其配体 torsolike定位在两端的卵黄膜 (vitelline membrane)上,不能与 torso结合。 受精时,torsolike得以释放, torsolike与torso结合, torso活化, 启动信号传导。 Torsolike蛋白的存在量很低, 受精后其扩散距离有限。其突变 体类似torso突变体。
2.果蝇前 – 后轴的形成方式
bcd mRNA由滋养细胞合成,后转移至卵细胞中并定 位于卵细胞的前极。
bcd mRNA在 受精后迅速翻 译,形成BCD 蛋白从前到后 的梯度。突变 型的BCD均匀 分布,不能形 成前后浓度梯 度
Bicoid基因提供A-P轴线形态素梯度
Bicoid编码一 种转录因子。 其突变体缺失 头胸结构,原 头区由尾区取 代。
胚轴形成

bcd mRNA由滋养细胞合成,后转移至卵细胞中并定 位于卵细胞的前极。
bicoid基因 对前端结构 的发育是必 需的 。
母源性基因bicoid mRNA在卵子中的分 布以及受精后biocoid 蛋白的浓度梯度。
随着BCD蛋白在胚胎中 的扩散,这种蛋白质也 开始降解——它有着大 约30分钟的半衰期。这 种降解对于建立起前后 浓度梯度是非常重要的。
第一节 果蝇胚轴的形成
• 胚轴指胚胎的前-后轴(anterior -posterior axes)和背 – 腹 轴(dorsal -ventral axis)。
爪蟾胚胎的前后轴、背腹轴和左右轴(中侧轴),互成垂直角度。
• 目前已筛选到与胚胎前后轴和背腹轴形成有关的约50个母 体效应基因(maternal effect gene)和120个合子基因 (zygotic gene)。 • 在果蝇最初的发育中,由母体效应基因构建的位置信息网 络,激活合子基因的表达,控制果蝇躯体模式的建立。 • 一、 果蝇胚胎的极性 果蝇的卵、胚胎、幼虫和成体都具有明确的前-后轴和 背-腹轴。 • 果蝇躯体模式的形成是沿前-后轴和背-腹轴进行的。果蝇 胚胎和幼虫沿前-后轴可分为头节、3个胸节和8个腹节, 两末端又分化出前面的原头(acron)和尾端的尾节 (telson);沿背腹轴分化为背部外胚层、腹侧外胚层、 中胚层和羊浆膜。
• 表皮细胞:细胞与细胞间紧密连接成管状或片层状结构,
局部或整个结构一起动。
• 间质细胞: 细胞与细胞间松散相连,每个细胞为一个行动 单元
外包的表皮细胞
内移的间质细胞
三、海胆的原肠作用
1. 初级间质细胞内移 囊胚期植物极一侧变厚称为植物极板,中央的一簇小细胞 脱离表面单层细胞进入囊胚腔,称为初级间质细胞,来源于卵 裂期的小裂球。初级间质细胞沿囊胚腔内表面运动,最终占据 囊胚腔预定腹侧面,最终形成幼虫碳酸钙骨针的轴。
07 第七章 胚胎诱导及组织、器官形成

B 蛙囊胚背面
C 同一囊胚的腹面
E 在胚胎腹面注入
β-catenin后产
生的双体轴胚胎
2.中胚层形成的诱导
在囊胚赤道板有一条宽的环状的细胞带, 被植物极部位的信号因子所决定而发育为中 胚层组织。
将美西螈的中期囊胚分 为动物半球顶部帽状的 Ⅰ区、下部的环形的Ⅱ 区,植物半球的Ⅲ区。 如果将Ⅰ、Ⅱ区分离培 养,则只形成未分化的 非典型表皮,以后逐渐 退化。说明在中期囊胚 Ⅱ区尚未获得中胚层的 分化能力。
第一节 初级胚胎诱导
(primary embryonic induction)
初级胚胎诱导是指脊索中胚层诱导其表 面覆盖的外胚层发育为神经板的现象。
(一)组织者的发现及在初级胚胎诱导中的作用
❖将一种浅色蝾螈原肠胚的胚孔背唇 移植到深色蝾螈同龄胚胎腹侧。在宿 主原有的神经板相对的腹侧面形成了 另一个次级神经板。
❖其中最重要的一个背部 化决定子是转录因子βCATENIN。
❖在非洲爪蟾中,βcatenin在细胞质转动的过 程中开始在背部区域聚集, 在囊胚期只存在于背侧的 细胞中。
❖β-catenin是背轴形成的 关键。用β-catenin的反义 寡聚核苷酸处理会使胚胎 缺失背部结构。而在胚胎 的腹侧注入β-catenin会产 生第二个体轴。
❖晚期原肠胚胚孔背唇诱导躯干和尾部结构的形成,称为躯 干组织者和躯尾组织者
将原肠晚期原肠顶不同部位的移植块放到早 期原肠胚的囊胚腔中,会诱导不同的结构形成.
(二)初级诱导的过程及体轴决定的机制
初级胚胎诱导导致的胚胎模式形成过程包括四个 阶段:
1、Nieuwkoop中心的形成与背腹决定 2、中胚层形成的诱导 3、背部化与头尾化诱导 4、神经形成的诱导
发育生物学体轴与胚层
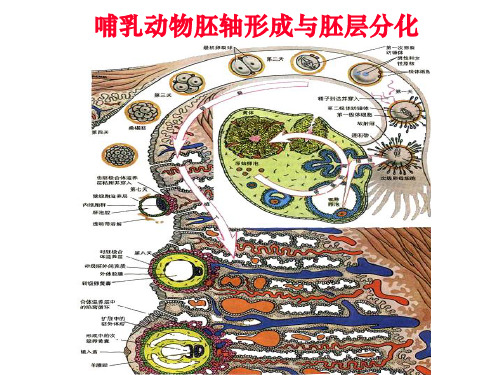
哺乳动物卵裂球的compaction: 发生第三次卵裂后不久
紧密化Compaction: 8细胞分裂后不久,卵裂球突然挤在一起, 卵裂球之间的接触面增大,形成一个紧密的细胞球体.
人类的原肠作用
上胚层和下胚层的形成:在原肠作用开始时,内细胞团分裂为两层.与囊胚 腔接触的一层为下胚层,将用于形成yolk sac;另一层为上胚层.上胚层细胞 间的缝隙将合并、扩大成为羊膜腔,腔中的液体可防止胚胎脱水和保护胚胎受 振荡.上胚层将发育为胚胎的本体.
原肠作用前的胚泡,示内细胞团和滋养层细胞.
注射E-Cadherin抗体的胚胎不能够致密化. 在4细胞期激活 蛋白激酶C导致compaction发生.因此,Compaction可能始于蛋 白激酶C的活化,它引起细胞骨架的重排,在膜上均匀分布的ECadherin重新定位在胞间相交处.
哺乳动物囊胚细胞命运的早期分化-位置决定论
16细胞期桑椹胚morula:位于内部的少数细胞产生的子细胞将组 成内细胞团inner cell mass,而位于外部的细胞产生的子细胞大多 构成滋养外胚层trophoblast.
哺乳动物胚轴形成与胚层分化
Vertebrate embryos show differences in form before gastrulation
After gastrulation, rather similar at
Phylotypic
stage 种系型 期: head structure, neural tube, notochord, somites
发育生物学教学课件07-2 胚层的形成

25
周围神经系统的发生
神经褶顶缘的神经外胚层细胞迁移至神经管的背外侧,称神经嵴 (neural crest)。是周围神经系统的原基,将分化为脑神经节、脊神 经节、 植物神经节和外周神经与分别迁移各器官分化为嗜铬细胞、 甲状腺的滤泡旁细胞和皮肤的黑色素细胞等。
神经嵴
神经节
神经背根 脊神经
26
② 表面(普通)外胚层:
20
(四)胚层分化与胚体外形建立 (第
4~8周)
21
1、外胚层的分化 differentiation of ectoderm
① 神经外胚层:neural ectoderm
神经板(neural plate)。
(脊索背侧的) 细胞增生 外胚层 脊索诱导下
神经板
neural crest
神经嵴
PNS
神经褶 神经沟 神经褶
Disc (二) 相关结构的发生
2
(一) Bilaminar germ disc formation
第2周,Inner cell mass: cell division, proliferation & differentiation形成embryonic disc 上胚层(epiblast or Primitive ectoderm layer ), 下胚层(hypoblast )。称为二胚层胚盘,是人体发育的原基。
上胚层 下胚层
4
(二) 相关结构的发生
二胚层时期(第2周)
滋养层细胞向外增殖,形成合体滋养层(Syncytiotrophoblast),
深面一层立方形细胞,边界明显称细胞滋养层 (Cytotrophoblast)。
羊膜腔 epiblast与极端滋养层之间的腔
胚胎模式形成

In the embryonic development of animals, the initial pattern formation mainly involves the hypocotyl ( embryonic axes ) formation and a series of related cell differentiation process. Hypocotyl of embryonic front - rear axle ( anterior posterior axes ) and dorsal, ventral axis ( dorsal ventral axis ). 在动物胚胎发育中,最初的图式形成主要涉及胚轴 在动物胚胎发育中,最初的图式形成主要涉及胚轴 (embryonic axes)形成及其一系列相关的细胞分化过程。 axes)形成及其一系列相关的细胞分化过程。 胚轴指胚胎的前-后轴(anterior 胚轴指胚胎的前-后轴(anterior -posterior axes)和 axes)和 背 – 腹轴(dorsal -ventral axis)。 腹轴(dorsal axis) 胚轴的形成是在一系列基因的多层次、网络性调控下完成 的。
第二节 两栖类胚轴形成
Section second amphibian hypocotyl formation
Amphibian is the adjustment of embryonic development of the typical model. Amphibian embryo dorsal abdominal shaft
果蝇沿前 后轴、背 腹轴和中 侧轴建立 形体模式。
二、果蝇前 – 后轴的形成 In two, before the formation of Drosophila melanogaster, rear axle
07第七章 初级胚胎诱两栖类胚轴形成
含Macho重分配到b4.2,结果b4.2发育出肌肉
镶嵌型胚胎:早期胚胎大部分细胞通过自主分 化决定,由此发育的胚胎,传统上称为镶嵌型 胚胎
二、条件特化Conditional specification
条件特化是细胞通过与其他细胞相互作用完成他们 各自发育命运的能力。亦即,细胞通过与邻近细胞 相互作用逐渐限制他们的发育命运,并只能朝一定 方向分化。发育命运的特化取决于邻近细胞分泌的 旁泌因子。
(2) GSK-3活性受Dsh(Disheveled) 和GBP(GSK-3 binding protein)抑制
A、精子入卵点定向微管, Kinesin运送GBP植物极 B、植物极皮质处的Dsh联上 GBP被Kinesin沿微管运送, 一旦到达精子入卵点对 侧植物极(未来胚胎背 侧),Dsh和DBP即与微 管分开 C、 在最背侧植物极Dsh和 DBP抑制GSK-3,β-Cat积 累于此,腹面植物极中, β-Cat降解
第一节 细胞定型commitment
细胞分化(cell differentiation) :从单个受精卵产 生各种类型细胞的发育过程。分化细胞不仅形态各 异, 而且功能各异,它们都有自己特异的蛋白质产物
部分已分化细胞的类型、特征产物及其功能
细胞类型
角质细胞 红细胞 眼晶体细胞 B-cell T-cell 黑素细胞 胰岛细胞 间质细胞 软骨细胞 成骨细胞 肌肉细胞 肝细胞 神经元 鸡输卵管管状细胞 昆虫滤泡细胞
(Regional specificity of mesoderm induction)
3、腹面和侧面 植物极细胞诱 导中段中胚层 (肌肉、间充 质)和腹面中 胚层(间充质、 血液、前肾) 的形成 4、最背面植物 极细胞诱导胚 轴中胚层成分 (脊索、体节) 的形成
胚轴发育成什么

胚轴发育成什么
胚芽发育成茎和叶。
胚轴是种子植物胚的组成部分之一,为子叶着生点与胚根之间的轴体。
种子萌发后,由子叶到第1片真叶之间的部分,称为“上胚轴”;子叶与根之间的一部分,称为“下胚轴”。
胚轴发育为连接茎和根的部分。
扩展资料
胚已有初步的分化,在双子叶植物分化为胚芽、胚轴、胚根和子叶四部分(如大豆和蚕豆);在单子叶植物分化为胚芽、胚轴、胚根和胚乳(如玉米)。
胚芽位于胚的顶端,是未来植物茎叶系统的原始体,将来发育成为植物的地上部分。
胚轴位于胚芽和胚根之间,并与子叶相连。
胚根位于胚轴之下,呈圆锥状,是种子内主根的雏型,将来可发育成植物的主根,也是最先从破例的种子皮中长出来,并形成植株的根系。
子叶是胚的叶,或者说,是暂时的叶,一般为1或2片,位于胚轴的侧方。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
胚轴的形成是在一系列基因多层次羊浆膜
2. 果蝇前后极性的产生
BICOID(BCD)HUNCHBACK(HB)NANOS(NOS)CAUDAL(CDL)
gap gene
pair-rule gene segment gene homeotic gene
Torso系统基因的失活会导致胚胎不分节的部分,即前端原头区和后端尾节,缺失
末端系统:Torso信号途径
•tor基因编码一种跨膜酪氨酸激酶受体(receptor tyrosine kinase,RTK),在整个合胞体胚胎的表面表达。
•其NH2-基端位于细胞膜外,COOH基端位于细胞膜内。
•当胚胎前、后端细胞外存在某种信号分子(配体)时可使TOR 特异性活化,最终导致胚胎前、后末端细胞命运的特化。
torso-like(tsl)基因可能编码这一配体。
末端系统的作用方式与前两个系统
完全不同
tor
因
因均编码转录调节因子,可
进一步调节其他基因(如
hb,ftz,kr
Torso信号传导途径TOLL激活后可激发一系列细胞内信号,并涉及到信号传导途径中其他母体效应基
因,如pelle,tube基
因产物的作用,最终
使CACTUS降解,DL 蛋白释放进而进入细胞核中。
7. 卵子发生过程中体轴的极化和
滤泡细胞背腹极性的产生
滋养细胞合成mRNA, rRNA,甚至是完整的核糖体,
并通过细胞间桥的胼合体,单向转运到卵母细胞里。
果蝇卵子
发生过程
中前后和
背腹轴的
特化。
滤泡细胞
的相互作
用可引起
卵母细胞
前后和背
腹轴的特
化。
Max-Planck-Institut für
Entwicklungsbiologie
Tübingen, Federal Republic of
Germany
1942
Princeton University
Princeton, NJ, USA
1947
Eric F. Wieschaus
Edward B. Lewis Christiane Nüsslein-Volhard。