2.1复制
BS EN 12845-2004+A2-2009 固定式消防系统.自动喷水系统设计,安装和维修

BRITISH STANDARD
BS EN 12845:2004 +A2:2009
Incorporating Corrigendum August 2009
Fixed firefighting systems — Automatic sprinkler systems — Design, installation and maintenance
1 2 3 4 4.1 4.2 4.3 4.4 4.4.1 4.4.2 4.4.3 4.4.4 5 5.1 5.1.1 5.1.2 5.2 5.3 5.4 5.5 6 6.1 6.2 6.2.1 6.2.2 6.2.3 6.3 6.3.1 6.3.2 7 7.1 7.2 7.2.1 7.2.2 7.2.3 7.3 7.3.1 7.3.2
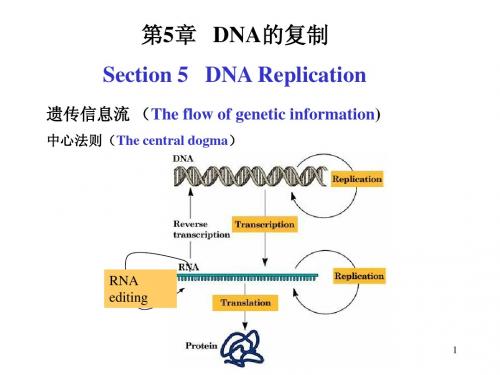
介绍 Introduction ................................................................................................................................................. 10
This British Standard was published under the authority of the Standards Policy and Strategy Committee on 16 November 2004
Amendments/corrigenda issued since publication Date 31 July 2009 Comments Implementation of CEN amendment A2:2009 Correction to National foreword
克隆质粒的复制子以及在宿主菌的拷贝数

克隆质粒的复制子以及在宿主菌的拷贝数质粒,其复制子以及在E. coli宿主内的拷贝数质粒类型复制子拷贝数pUC18 pMB1* 500-700pKS(-) ColE1 300-500pBBR1系列载体 pBBR1 30-40pBAD322,pET等系列载体 pMB1-rop 15-20pACYC系列载体 p15A 10-15pSC101衍生载体 pSC101 5-10~15 (pir宿主菌如BW25141) pKD4, pR6KMCS R6K ~250 (pir-116宿主菌如BUN20) pJB866 RK2-trfA (oriV) mediumopBADTcTypeG pMB1 120 copies at 37C pECBAC1 oriS 1~2Genome One copy拷贝数是指每个细胞中有多少个质粒DNA的分子,质粒的拷贝数直接决定了:1. 从菌液中获得质粒DNA的量。
拷贝数和量成正比关系2. 以之为载体时转化子的数目。
拷贝数的高低和转化子的多少成正相关,是否线性不确切3. 筛选用抗生素的浓度。
拷贝数的高低和抗生素的使用浓度成正相关,如100,g/ml的氨苄青霉素筛选DH10B/pKS(-),也可以用50,g/ml;50,g/ml筛选DH10B/pBAD322;15,g/ml筛选整合至基因组上的抗性基因。
当菌株的筛选和培养出现非预期的情形,适当地调整抗生素的浓度就非常重要。
TaKaRa MiniBEST Plasmid Purification Kit Ver.2.0 (Code:DV801A)提到4 ml的过夜培养的菌液中可纯化得到20 ,g质粒,以此推断,1.5ml 过夜培养含高拷贝质粒如pKS(-)的菌液中可得到多于5 ,g质粒,如溶于50 ,l TE/ddHO,则浓度约100ng/,l。
可参照此标准,来根据2不同拷贝数的质粒来推算本实验需要多少菌体、所提取质粒的量,再通过电泳后与分子量标准的比较来进一步估算出的DNA浓度。
原核生物及真核生物DNA复制

真核生物DNA聚合酶及有关蛋白
表 真核生物五种DNA聚合酶
DNA聚合 酶
位置
功能α核 引发 Nhomakorabeaδ
核 合成
ε
核 修复
βγ
核 线粒体 修复 复制
相对活性 80% 分子量 300K
170-230K
250K
亚基
3’→5’ 外切
催化核心(180K) 催化核心
催化核
两个引物酶(60,50K) (125K)
心
一个未知
原核生物及真核生物DNA复制
9、单链结合蛋白(SSBP-single-strand binding protein):稳定已被解开的DNA 单链,阻止复性和保护单链不被核酸酶 降解。
原核生物及真核生物DNA复制
(三)DNA的复制过程(大肠杆菌为例)
双链的解开
RNA引物的合成
DNA链的延伸
切除RNA引物,填补缺口,连接相邻的
5、 DNA聚合酶:以DNA为模板的DNA合成酶 ●以四种脱氧核苷酸三磷酸为底物 ●反应需要有模板的指导 ●反应需要有3-OH存在 ●DNA链的合成方向为5 3
原核生物及真核生物DNA复制
原核生物中的DNA聚合酶(大肠杆菌)
性质
聚合酶Ⅰ 聚合酶Ⅱ 聚合酶 Ⅲ
3' 5 '外切活性 +
+
+
5' 3 '外切活性 +
在DNA复制时,合成方向与复制叉移动的方向一致并 连续合成的链为前导链;合成方向与复制叉移动的 方向相反,形成许多不连续的片段,最后再连成一 条完整的DNA链为原滞核生后物及链真核。生物DNA复制
在DNA复制过程中,前导链能连续合成, 而滞后链只能是断续的合成53 的多 个短片段,这些不连续的小片段称为冈 崎片段。
复制名词解释

复制名词解释复制(Copyleft)指在版权保护法下,为了保护作品免受严苛的版权限制,作者使用特定的版权声明,赋予其他人以进一步复制、分发、修改和使用作品的自由,而非限制他人的权利。
这种授权方式鼓励分享和合作,允许他人共享及改进原创作品。
复制的概念最早由理查德·斯托曼于1984年提出,他是一个自由软件运动的活跃参与者。
复制的授权方式主要用于自由软件和开源软件的发展和推广。
自由软件是指用户可以自由地运行、复制、分发、研究和改进软件的计算机程序,无论其用于商业目的还是非商业目的,而开源软件则是强调让软件的源代码对所有人开放,允许任何人进行修改和分发。
复制授权方式使得自由软件和开源软件能够更广泛地传播和发展,为广大用户提供了更多选择和自由。
在复制授权方式中,作者使用特定的版权声明,明确表明其作品允许他人自由地复制、分发、修改和使用,同时要求他们以相同的授权方式继续传播。
这种要求被称为“共建共享”。
使用和修改作品的人只需遵守这一要求,不需要再征得作者的许可或支付版权费用。
复制的授权方式有多种形式,最常见的是使用知识共享许可协议。
知识共享是一个非营利组织,提供了一系列灵活的授权协议,适用于不同类型的作品和不同的使用要求。
知识共享许可协议允许作者为自己的作品选择特定的授权方式,并提供了相应的版权声明文本,以便他人了解、遵守和传播。
除了知识共享许可协议外,复制的授权方式还有GNU通用公共许可协议(GPL)和开源通用许可协议(OSL)等。
这些协议在保护作品的自由使用和修改的同时,也规定了一些限制,如要求保留原始版权声明、提供源代码、使用相同的授权方式等。
总之,复制授权方式是一种通过特定的版权声明,允许他人自由复制、分发、修改和使用作品的授权方式。
它鼓励分享和合作,推动自由软件和开源软件的发展和推广。
复制授权方式主要通过知识共享许可协议等形式实现,其中要求共建共享是一项重要的原则。
2.1 减数分裂和受精作用 课件【新教材】2020-2021学年高一生物人教版(2019)必修二

数目
4n
染色体
核 DNA
染色单体
2n
n
0
G1 S G2 减Ⅰ前 减Ⅰ中 减Ⅰ后 减Ⅰ末 减Ⅱ中减Ⅱ后减Ⅱ末 减Ⅱ前
四、减数分裂过程中DNA、染色体和染色单体数的变化规律
5、染色体/核DNA数目变化曲线
染色体/核DNA 变化原因? 染色体复制
1
着丝粒分裂,姐妹染色单 体分开,形成子染色体
变化原因?
1/2
二、精子的形成过程 6、减数第二次分裂前期:
次 级 精 母 细 胞
减Ⅱ前期
减数分裂Ⅰ与减数分裂Ⅱ之间通常没有间期,
或者间期很短;这期间的
。
特点:
染色体散乱分布在纺锤体中央, 该过程无同源染色体;
注意:减数分裂Ⅱ的各时期变化除无同 源染色体之外,其他均与有丝分裂一样;
二、精子的形成过程 7、减数第二次分裂中期:
细胞分裂,同源染色体
每个细胞中 染色单体数
4n
分离,进入两个子细胞
变化原因?
着丝粒分裂,姐妹染色单
体分开,形成子染色体
2n
染色体复制
变化原因?
n
变化原因?
时期
间期 减I前 减I中 减I后 减I末 减II前 减II中 减II后 减II末
四、减数分裂过程中DNA、染色体和染色单体数的变化规律
3、每个细胞中的DNA数目变化曲线
4n
2n
细胞分裂,同源染色体 分离,进入两个子细胞
变化原因?
着丝粒分裂,姐 妹染色单体分开, 形成子染色体
变化原因?
细胞分裂,子染色 体进入两个子细胞
变化原因?
n
时期
间期 减I前 减I中 减I后 减I末 减II前 减II中 减II后 减II末
第四章DNA复制

4. DNA ligase: 连接冈崎片断。
4: 终止与分离 Termination and Segregation
终止
•引起终止的序列称为终止位点(ter site) 两个复制叉在Ori C 约1800的对面相遇 •Tus 蛋白: 终止位点结合蛋白, 可阻止Dna B解旋
大肠杆菌复制终点定位偏 离复制叉实际相遇点
• 就DNA复制而言,引物酶(dna G基因产物)合成RNA 链,它提供引发末端
DNA聚合酶需要3‘-OH 端来起始复制
有多种方法可以提供DNA聚合酶起始DNA合成的3‘OH端
大肠杆菌中发现的两类引发反应:
引发需要解旋酶、SSB和引物酶
1、ori C系统
2、ØX系统: 引发体(primosome)
Segregation
•拓扑异构酶IV(Topoisomerase IV):
一种II型拓扑异构酶,功能是使子链
分离,DNA分配到两个子细胞中。
第四节
真核生物DNA的复制
离体实验体系 起始 端粒的复制
1: 离体实验体系
• 酵母(Sacharomyces cervisiae) • 猿猴病毒40(SV40) • 非洲爪蟾(Xenopus laevis)的卵中提出的无 细胞提取物
5 大肠杆菌DNA聚合酶(DNA Polymerase)
硫氧还蛋白 拇指
DNA聚合酶的一般结构 “右手”结构: 拇指(thumb) 手指(finger) 手掌(palm)
手指
扭曲的DNA
DNA位于手掌上 由拇指和手指形 成的槽中
噬菌体T7 DNA聚合酶与 DNA复合体的晶体结构
核酸外切酶结构域 拇指结构域:与DNA结 合并在前进中起重要作 用
快速复制粘贴数据

快速复制粘贴数据在日常工作和学习中,我们经常需要复制和粘贴大量的数据。
为了提高工作效率,快速复制粘贴数据成为一个非常重要的技巧。
本文将介绍几种快速复制粘贴数据的方法,以及在使用过程中需要注意的问题。
一、使用鼠标快速复制粘贴数据在Windows系统中,使用鼠标进行快速复制粘贴数据是最常见的操作方式。
具体步骤如下:1. 选中要复制的数据:将鼠标光标移动至数据的起始位置,按住鼠标左键并拖动至结束位置,即可选中要复制的数据。
2. 复制选中的数据:松开鼠标左键后,按下Ctrl键,并同时按下C 键,即可将选中的数据复制到剪贴板中。
3. 粘贴数据:将鼠标光标移动到要粘贴的位置,按下Ctrl键,并同时按下V键,即可将剪贴板中的数据粘贴到该位置。
二、使用快捷键快速复制粘贴数据除了使用鼠标操作外,还可以通过快捷键来实现快速复制粘贴数据的功能。
以下是一些常用的快捷键:1. 复制数据:选中要复制的数据后,按下Ctrl键,并同时按下C键。
2. 粘贴数据:将鼠标光标移动到要粘贴的位置,按下Ctrl键,并同时按下V键。
快捷键的使用相较于鼠标操作更加方便快捷,可以大大提高工作效率。
三、使用文本编辑器快速复制粘贴数据对于需要处理大量数据的情况,我们可以使用文本编辑器来进行快速复制粘贴。
以下是一些常见的文本编辑器操作技巧:1. 复制多行数据:将鼠标光标移动到要复制的起始行,按住Shift键,并同时按下向下方向键,直到选中所有要复制的行。
然后按下Ctrl 键,并同时按下C键,即可将选中的多行数据复制到剪贴板中。
2. 粘贴多行数据:将鼠标光标移动到要粘贴的位置,按下Ctrl键,并同时按下V键,即可将剪贴板中的多行数据粘贴到该位置。
文本编辑器不仅可以处理文字数据,还可以处理数字、代码等各种格式的数据,非常适合进行批量处理。
四、注意事项在使用快速复制粘贴数据的过程中,需要注意以下问题,以确保数据的准确性和完整性:1. 数据的格式:在复制粘贴数据时,需要确保数据的格式保持一致,避免出现错位或格式错乱的情况。
复制数据的四种方法

复制数据的四种方法四种复制数据的方法在数字化时代,数据的复制变得越来越常见和重要。
有时我们需要将数据从一个地方复制到另一个地方,以备份、共享或分析。
以下是四种常见的数据复制方法。
方法一:拷贝粘贴这是最简单和最常见的数据复制方法之一。
只需选中要复制的数据,然后使用鼠标右键或键盘快捷键复制(Ctrl+C),再将光标移动到目标位置,使用鼠标右键或键盘快捷键粘贴(Ctrl+V)。
这种方法适用于小量数据的复制,如单个文件、文件夹或少量文本。
方法二:文件复制当需要复制大量数据或文件时,可以使用文件复制方法。
首先,选中要复制的文件或文件夹,然后使用鼠标右键或键盘快捷键复制(Ctrl+C)。
接下来,打开目标位置,使用鼠标右键或键盘快捷键粘贴(Ctrl+V)。
这种方法适用于复制大量文件或文件夹,如备份整个硬盘或移动大型项目文件。
方法三:数据传输当需要将数据从一个设备传输到另一个设备时,可以使用数据传输方法。
这种方法可以通过有线或无线方式进行。
有线传输通常使用USB、HDMI或网络连接,而无线传输通常使用蓝牙、Wi-Fi或云存储服务。
例如,可以使用USB数据线将照片从手机传输到电脑,或使用云存储服务将文件从电脑传输到手机。
方法四:数据备份数据备份是一种重要的数据复制方法,用于保护数据免受数据丢失或损坏的风险。
数据备份可以通过多种方式进行,如本地备份、外部存储设备备份或云备份。
本地备份通常将数据复制到另一个硬盘或服务器,而外部存储设备备份通常将数据复制到移动硬盘或U盘。
云备份是将数据复制到云存储服务提供商的服务器上,以便在需要时可以从任何设备访问数据。
总结数据复制是在数字化时代中不可或缺的操作。
通过拷贝粘贴、文件复制、数据传输和数据备份等方法,我们可以轻松地复制和移动数据。
无论是小量数据还是大量数据,都可以选择适合的方法进行复制。
然而,在进行数据复制时,我们也应该注意数据的安全性和隐私保护,避免数据泄露和不必要的风险。
复制临床发热症状动物模型的研究进展

复制临床发热症状动物模型的研究进展复制临床发热症状动物模型的研究进展发热是多种疾病进展过程中的一种非常常见的疾病症状,特别是感染性疾病,发热过高或持久可引起体力消耗,严重者可危及生命。
体温的升高与体内病变存在着依赖关系,可以说,发热常为疾病发生与发展的信号。
近年来,学者们用不同的方法成功复制了发热动物模型,主要通过化学药品、生物制剂和综合诱导方法进行,现就目前制造动物发热模型的方法综述如下。
1.发热动物模型评价标准发热模型的评价指标主要为动物体温升高。
常通过计算体温变化值AT(AT=T2-T0)和体温反应指数[TRI,指发热曲线与基线之间的面积(clC.hr),它代表发热的高度和持续的时间]进行评价。
动物的体温.通常采用温度计插入肛门内测量其体内温度的方式获得,且动物应处于安静状态。
一般认为模型动物造模前,3次的平均体温与动物造模后体温有差异,为造模成功。
2.发热模型的造模方法及药品2.1化学药品造模法本法是将模型动物通过化学药品的作用,使其体温高于正常体温的造模方法。
2.1.1 2,4---硝基苯酚(DNP)DNP是一种离子载体,能够消除线粒体的部分跨膜质子浓度梯度.而质子浓度梯度是绝大部分制造ATP的能量来源,这从而妨碍了ATP的产生,其中浪费的大量能量即通过热量的方式散失。
故给予实验动物一定量的DNP可以复制发热动物病理模型。
何汝帮等[311)2 1.5 mL/kg的剂量给家兔背部皮下注射2%的DNP溶液,30 min后家兔体温显著升高,l h后达到峰值,较之对照组体温升高1.58°C。
陈敬然等在大鼠皮下以160 mg/kg的剂量注射DNP溶液,将温度升高到0.6℃以上作为造模成功的标准。
陈睿等按0.2 mL/100 g的剂量给大鼠背部皮下注射1.0%的DNP溶液,从而得到了合适的动物发热模型。
2.1.2甲状腺片甲状腺片主要成分为甲状腺激素.是影响机体能量代谢的主要物质,能加快细胞内氧化速度,产热增加从而造成发热。
DNA的复制
10
细菌DNA的复制 (Bacterial DNA replication) 的复制 细菌
1. 实验系统 实验系统(Experimental systems) (1) 质粒 ) ):结构简单 (2)噬菌体 )噬菌体φX174 (5kb):结构简单,超螺旋环型 ):结构简单, DNA,完全依赖细菌细胞的复制子来完成自身的复制, ,完全依赖细菌细胞的复制子来完成自身的复制, 为研究Ecoli染色体的复制提供模型。 染色体的复制提供模型。 为研究 染色体的复制提供模型
15
(4) 终止与分离 终止与分离(Termination and Segregation) 1)环状的DNA两个复制叉在 ℃相遇,靠基因 的 )环状的 两个复制叉在180℃相遇,靠基因tus的 两个复制叉在 产物(一种 解旋酶的抑制剂) 产物(一种DnaB解旋酶的抑制剂)抑制了 解旋酶的抑制剂 抑制了DnaB解 解 旋酶的活性,反应停止。 旋酶的活性,反应停止。 2)依靠拓扑异构酶IV的解联作用,把两条子链分配到 )依靠拓扑异构酶 的解联作用 的解联作用, 2个子细胞中。 个子细胞中。 个子细胞中
5. DNA合成的高保真性 The high fidelity of DNA replication 合成的高保真性
8
6. DNA复制的拓扑性质 复制的拓扑性质 (Topological Character of DNA replication) 拓扑异构酶 (Topoisomerases) 拓扑异构酶 拓扑异构酶是一类可以调控DNA分子超螺旋水平的酶。 拓扑异构酶是一类可以调控 分子超螺旋水平的酶。 分子超螺旋水平的酶 I型拓扑异构酶 :在DNA链上的其中一股产生一个切 型拓扑异构酶 链上的其中一股产生一个切 口,使一条链得以穿越,每次改变±1个连接数。 (下 使一条链得以穿越,每次改变± 个连接数。 个连接数 图) II型拓扑异构酶 :是DNA的旋转酶,由ATP提供能量, 型拓扑异构酶 的旋转酶, 提供能量, 的旋转酶 提供能量 向闭环DNA分子中引入负超螺旋,从而抵消了 分子中引入负超螺旋, 向闭环 分子中引入负超螺旋 从而抵消了DNA复 复 制过程中产生的正超螺旋。其作用方式是在DNA两条链 制过程中产生的正超螺旋。其作用方式是在 两条链 同时切开, 得以穿过, 同时切开,使另一条双链 DNA得以穿过,每次改变±2 得以穿过 每次改变± 个连接数。 个连接数。
0202-复制、剪切与粘贴

Excel版本介绍
第1段:Excel 2作为Windows Excel的原始版本于1987年问世。
因为当时Windows还不是很游行,所以这个版本只包括了一个Windows的运行版本,它是一种特殊的版本。
,足以使Excel 顺利运行。
根据现在的标准看,这个版本过于粗糙。
第3段: Excel 4 在1992年投入市场。
这个版本在市场上产生了不小的反响。
第2段: Excel 3 1990年底发布。
这个版本无论从外观还是性能上都做了重大改进。
它增加了工具栏、绘图功能、工作表大纲、加载宏支持、3D图表、工作组编辑等。
第9段: Excel 2003 (Excel 11)是在2003年秋季发布的。
第8段: Excel XP (Excel 10)在2001年6月发布的。
第5段: Excel 95 (Excel 7)是在1995年夏天推出的。
第7段: Excel 2000(Excel 9)在1999年6月发布。
它提供的新功能不多,但是它的最大改进是把HTML作为可选的文件格式,它仍然把持标准的二进制文件格式。
第4段: Excel 5 在1994年初投入市场,这个版本推出了更多的新功能,包括多页面工作簿和新的VBA宏语言。
第6段: Excel 97 (Excel 8)它提供了前所未有的升级功能。
期刊发表复制比多少

期刊发表复制比多少期刊发表复制比是指同一篇论文在不同期刊上发表的次数之比。
在学术界,期刊发表复制比是一个重要的指标,可以反映出一篇论文的影响力和重要性。
通常情况下,一篇论文在不同期刊上发表的次数越多,说明该论文的研究成果越受到学术界的认可和重视。
期刊发表复制比的计算方法很简单,就是将一篇论文在不同期刊上的发表次数相加,然后除以该论文在第一期刊上的发表次数。
例如,如果一篇论文在A期刊上发表了两次,在B期刊上发表了一次,那么这篇论文的发表复制比就是(2+1)/1=3。
通常情况下,期刊发表复制比大于1的论文被认为是高度重复发表的,而期刊发表复制比小于1的论文则被认为是低度重复发表的。
期刊发表复制比的高低可以反映出一篇论文的学术影响力和质量。
高度重复发表的论文往往是具有较高学术价值和影响力的论文,因为它们被多个期刊接受和发表,说明该研究成果得到了广泛的认可和重视。
而低度重复发表的论文则可能存在学术不端行为,比如抄袭、剽窃等,其学术质量和影响力可能较低。
此外,期刊发表复制比还可以反映出期刊的学术水平和影响力。
如果一篇论文在多个期刊上发表的次数较多,那么这些期刊往往是具有较高学术声誉和影响力的期刊,因为它们能够吸引到高质量的学术论文。
相反,如果一篇论文在多个期刊上发表的次数较少,那么这些期刊可能是学术水平和影响力较低的期刊,因为它们难以吸引到高质量的学术论文。
综上所述,期刊发表复制比是一个重要的指标,可以反映出论文和期刊的学术影响力和质量。
研究人员和期刊编辑应该重视期刊发表复制比,避免低度重复发表,提高学术论文的质量和影响力。
同时,学术界也应该加强对期刊发表复制比的监管和评估,提高期刊的学术水平和影响力。
复制粘贴有学问

复制粘贴有学问每天我们使用windows的时候,复制粘贴恐怕是用得最多的操作了。
别小看这个简单的复制拷贝操作,里面还有不少技巧学问呢!首先我们来看看,在复制拷贝到粘贴完成的过程里,windows实际上进行了哪些内在的步骤:1、选择复制内容,当你还没点击“复制”选项的时候,windows只是把你选择的内容以高亮状态条标记出来;2、选定内容并点击复制后,windows开始把你选择的内容写入物理内存(剪贴板),如果内容太多超出了当前可用的物理内存容量,就调用硬盘空间即虚拟内存作记录;(注意windows自带的剪贴板功能只能记录一次选定的内容,如果你想选定多段不同的内容汇总到剪贴板里,再分批次处理,要安装专门的剪贴板辅助工具程序)3、当你在目标程序位置中点击“粘贴”,windows就从刚才记录内容的内存中读取相关数据,并显示在屏幕上你当前工作的窗口里。
注意在你进行保存操作以前,粘贴只是一个“读取—显示”的操作而已,如果你现在不存盘关机,工作结果会丢失;别忘了要正式保存你的工作到磁盘上。
好了,从上面叙述的过程,我们可以清楚地看到,复制粘贴的关键是选定的内容在内存中的处理。
这一方面涉及到windows对内存的管理运用;另一方面也涉及到不同的工具程序,对在内存中剪贴板里保存的数据采用什么方式来识别,这点会导致不同的粘贴效果,所以控制得不好的话,实际的粘贴结果和你期待的可能会截然不同。
比如windows的记事本程序,只能处理64K字节以内的数据,当你试图粘贴超过该数值的数据时候,系统会提示“内存不足,XX操作无法完成”或是“XX文件无法被保存”,这是windows记事本程序本身的功能不完善造成的,你必须使用写字板或是WPS等能处理大数据的文本程序。
在复制粘贴操作中,HTML格式是一个至关重要的问题。
比如你在浏览一个网页,尤其是象硬件报价那样带有表格才方便查看的网页,如果你希望获得的粘贴结果和原来网页上一模一样也就是保持HTML 文件的风格,就不能用记事本或是写字板这些不支持HTML格式的程序来进行粘贴处理,即使它的字节数没超过64K。
两个Excel复制小技巧,包你事倍功半!

两个Excel复制小技巧,包你事倍功半!
使用Exce的方法哦!今天我就教大家两个小技巧,让你的工作效率翻番哦~
1、Ctrl+D:向下复制
*包含所复制的单元格:一个或多个单元格,连续或者非连续区域都有效。
2、Ctrl+R:向右复制
*包含所复制的单元格:一次只能复制一个或者一列,且会将原单元格的格式复制。
*不包含所复制的单元格:一个或多个单元格,只对连续区域有效。
需要注意的是,这个快捷键对非连续性区域无效。
gstreamer手册(简体中文版)

2.1.2. “一个目标”媒体播放器/媒体库
典型的 MPEG 播放器可以播放 MPEG 视频和音频,多数的播放器实现了完整的 底层构造来达到他们的唯一目标:播放。没有一套有效的机制可以提供对于音频 和视频数据过滤和效果处理,更没有制定在视频或音频数据中添加滤波器或特殊 效果的任何规定。
组件(Elements)。对于如何编写组件(Elements)的话题在GStreamer Plugin Writer's Guide中有详细的说明。
3.2. 箱柜(Bins)和管道(pipelines)
箱柜(Bins)是一个可以装载组件(element)的容器。管道(pipelines)是箱柜 (Bins)的一个特殊的子类型,管道(pipelines)可以操作包含在它自身内部的所 有组件(element)。因为箱柜(Bins)本身又是组件(element)的子集,所以你能够 象操作普通组件(element)一样的操作一个箱柜(Bins), 通过这种方法可以降低 你的应用程序的复杂度。你可以改变一个箱柜(Bins)的状态来改变箱柜(Bins) 内部所有组件(element)的状态。箱柜(Bins)可以发送总线消息(bus messages) 给它的子集组件(element)(这些消息包括:错误消息(error messages),卷标消 息(tag messages),EOS消息(EOS messages))。
1.2. 谁需要读这个手册?
本手册是从一个程序开发人员的角度来描述GStreamer的: 它叙述了如何利用 GStreamer的开发库以及工具来编写一个基于GStreamer的应用程序。对于想学习" 如何编写插件"的朋友们, 我们建议你去参考<<插件编写指南(Plugin Writers Guide)>>。
实训2.1 创建、复制并删除文件夹_计算机操作与应用实训(第2版)_[共2页]
![实训2.1 创建、复制并删除文件夹_计算机操作与应用实训(第2版)_[共2页]](https://img.taocdn.com/s3/m/0573a61008a1284ac950437e.png)
计算机操作与应用实训(第2版)
8
图1-18 “保存方案”对话框图1-19 “高级”选项卡
实训2 管理个人文件
计算机系统中的大部分数据都要以文件的形式存储在磁盘上,只有学会管理文件资源才能轻松使用这些数据。
“资源管理器”是Windows XP提供的专用文件管理工具,利用它能够方便地管理个人文件。
实训2.1 创建、复制并删除文件夹
在D盘根目录下创建“netpic”文件夹,并在该文件夹下创建“背景图片”文件夹和“卡通图片”文件夹。
将位于D盘“个人文件”文件夹中“桌面背景”文件夹中的所有图片复制到“背景图片”文件夹中,彻底删除“桌面背景”文件夹。
【操作步骤】
(1)打开“资源管理器”程序窗口。
右击“开始”按钮,在弹出的快捷菜单中选择“资源管理器”命令,如图1-20所示,打开“资源管理器”程序窗口,如图1-21所示。
(2)创建新文件夹。
①将D盘根目录设置为当前文件夹。
单击左窗格中“LENOVO(D:)”盘符,将其打开,在右窗格中显示“D”盘根目录下的文件及文件夹内容,如图1-22所示。
②右击右窗格中的任意空白处,在弹出的快捷菜单中选择“新建→文件夹”命令,创建的新文件夹将出现在右窗格文件列表底部,其默认文件名为“新建文件夹”,并且该名字处于编辑状态,为该文件夹输入一个新名字“netpic”,然后按<Enter>键或者单击该名字方框外的任意位置。
图1-20 右击“开始”按钮弹出的快捷菜单。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
DNA聚合酶I ( 109kD)
功能:校读错误,填补空隙,切除引物
生命科学与技术学院 16 分子生物学(胡凯) 2015-2-10
DNA聚合酶Ⅲ
主要的合成酶 分子量约为140kD,是由10种亚基组 成的不对称二聚体。
生命科学与技术学院
6. 反平行DNA链和不连续复制
前导链—在DNA复制时,合成方向与复制 叉移动的方向一致并连续合成的链。
随后链—在DNA复制时,合成方向与复制 叉移动的方向相反,形成许多不连续的 片段,最后再连成一条完整的DNA链。 半不连续复制—在DNA复制时,领头链是 连续合成的,而随后链的合成是不连续的, 这种复制方式称为半不连续复制。
生命科学与技术学院 26 分子生物学(胡凯) 2015-2-10
冈崎片段 在DNA复制过程中,领头链能连续合成,而随后链只 能是断续的合成53 的多个短片段,这些不连续 的小片段以其发现者的名字命名为冈崎片段。
冈崎片段:真核生物中100-200个核苷酸(核小体的 DNA单位)。原核生物中1000-2000个核苷酸(相当 于一个顺反子)。
生命科学与技术学院
9
分子生物学(胡凯) 2015-2-10
3. DNA复制的起始
基本概念: ●复制子:基因组能独立进行复制的单位。
● 起点:控制复制起始,是含有 100 ~ 200 个碱
基对的一段DNA。细胞内存在着能识别起点的 特殊蛋白质(大肠杆菌中称为Dna A蛋白)。
●原核生物:只有一个复制子 ● 真核生物:多个复制子,多个起点,形成多
生命科学与技术学院
6
分子生物学(胡凯) 2015-2-10
2. 复制要求高度缠绕的DNA解螺旋
环状DNA的复制引入缠绕
当两条子链的复制 沿 双 链 DNA 螺 旋 进 行 时,会引起分子的整 个非复制部分过度缠 绕,形成正超螺旋。 这种超螺旋必须被某 种机制克服。
生命科学与技术学院
7
分子生物学(胡凯) 2015-2-10
SSB
拓扑异构酶 oriC(245bp的DNA组分)
稳定解开的单链
理顺DNA链 E.coli的复制起始点
生命科学与技术学院
33
分子生物学(胡凯) 2015-2-10
① 互相缠绕的双链母本DNA,复制从特定的位置(复制原 点Ori or O)开始,该位置常是富含A、T区段。
生命科学与技术学院
34
分子生物学(胡凯) 2015-2-10
分子生物学(胡凯) 2015-2-10
19
聚合酶活 性位点
模 板
引物
3 ′ → 5 ′外 切酶位点
生命科学与技术学院
20
分子生物学(胡凯) 2015-2-10
核酸外切酶活性
5´→3´外切酶活性: 切除引物 切除突变的片段
3´→5´外切酶活性: 切除错配的核苷酸
3'
5' 3'
C T T C A G G A
生命科学与技术学院
14
分子生物学(胡凯) 2015-2-10
原核生物的DNA聚合酶
1958年A.Kornberg在大肠杆菌中发现
DNA聚合酶I:校读错误,填补空隙, 切除引物。 DNA聚合酶II:在无polI及pol III的情 况下才起作用,其真正的功能也未完 全清楚。 DNA聚合酶III:主要的合成酶,多种亚 基组成的聚合体,作用同酶I。
2. 复制要求高度缠绕的DNA解螺旋
拓扑异构酶І:使DNA一条链发生断裂和再连接。 作用是松解负超螺旋,反应不需要能量。主要 集中在活性转录区,同转录有关。
拓扑异构酶 Π :使 DNA两条链发生断裂和再连 接。当引入负超螺旋时需要由 ATP提供能量, 同复制有关。
二者共同控制DNA的拓扑结构。
生命科学与技术学院
29
分子生物学(胡凯) 2015-2-10
• 引物合成酶(引发酶):此酶以DNA为模板合成一 段RNA,这段RNA作为合成DNA的引物(Primer)。实 质是以DNA为模板的RNA聚合酶。 DNA连接酶(1967年发现):若双链DNA中一条链有
切口,一端是3’-OH,另一端是5‘-磷酸基,连接 酶可催化这两端形成磷酸二酯键,而使切口连接。 但是它不能将两条游离的DNA单链连接起来。
DNA聚合酶Ⅲ >1(多亚基酶)
5’ 3’聚合活性
3‘ 5’外切活性
(保护DNA复制的 忠实性fidelity)
+ 中
+
+ 很低
+
+ 很高
+
5‘ 3’外切活性
+
-
-
主要是对DNA损伤的 修复;以及在DNA复 制时切除RNA引物并 填补其留下的空隙。
生命科学与技术学院
修复紫外光引起 的DNA损伤
DNA 复制的主 要聚合酶,还 具有3’-5‘ 外切酶的校对 功能,提高DNA 复制的保真性
单链结合蛋白:稳定已被解开的 DNA单链,阻 止复性和保护单链不被核酸酶降解。
生命科学与技术学院
13
分子生物学(胡凯) 2015-2-10
5. 新合成链的延长
聚合反应与聚合酶
DNA聚合酶:以DNA为模板的DNA合成酶。其特点为: (1)以四种脱氧核苷酸三磷酸为底物; (2)反应需要有模板的指导; (3)反应需要有3-OH存在; (4)DNA链的合成方向为5 3 (5)DNA聚合酶的反应可以利用 DNA双链作为模板和引物,亦可以单链 DNA 作为模板和引物. (6)DNA的体外聚合必须加入少量的 DNA才能进行。DNA在提取过程中易形 成切口(nick)或缺口(gap).则加入的DNA一条链作为模板而另一条链可作 为引物。
生命科学与技术学院
1
分子生物学(胡凯) 2015-2-10
主要内容 目 录
1
双链DNA的半保留复制 DNA复制的起始 复制的延长
5 6
完整的DNA复制系统
真核染色体的复制
2
3
7 未来实际的应用
4 反平行DNA链的不连续复制
生命科学与技术学院
2
分子生物学(胡凯) 2015-2-10
聚合反应机理:
17
分子生物学(胡凯) 2015-2-10
前导链
合成 可滑动的 夹持器亚基
后随链
合成
夹持 器装置
催化亚基
催化亚基
生命科学与技术学院
亚基保持核心酶 的二聚体结构
3´→5´外切酶亚基
18 分子生物学(胡凯) 2015-2-10
DNA聚合酶Ⅰ 亚基数目 1(单体酶)
DNA聚合酶Ⅱ >1(多亚基酶 )
5'
P 引物 P
5'
O G
3'
C 模 3'
5'
O A T
3'
板 链 5'
3'
OH
P
P
P
5' 5' O O 3' 3'
T T
A
OH OH
生命科学与技术学院 3 分子生物学(胡凯) 2015-2-10
1.双链DNA的半保留复制
• 半保留复制 DNA复制时每个子代DNA分子的一条链来自亲 代DNA,另一条链是新合成的,这种复制方式 称半保留复制。
大肠杆菌DNA复制错配率约10-9~10-10。其 染色体中有4.5×106bp,平均每1 000~10 000 个细胞经过一次分裂才会插入一个不正确的碱基。
生命科学与技术学院 22 分子生物学(胡凯) 2015-2-10
生命科学与技术学院
23
分子生物学(胡凯) 2015-2-10
小结-维持DNA复制准确性的因素: 内因: ① 按碱基配对原则 (错配率10-4~10-5) ② DNA聚合酶的作用 (错配率10-4~10-5) • 对碱基的识别作用----选择正确的碱基参 入到引物末端 • 对底物的识别作用----先识别引物最后 一个碱基是否正确, 后识别参入的dNTP是否正确 • 校正阅读---- 3′→ 5′外切酶的作用 ③ RNA引物最终被切除, 提高了复制准确性 ④复制完成后对错配碱基进行修复的酶系统 外因: • 四种dNTP要平衡; • Mn2+和Mg2+的比例、浓度(酶活)
生命科学与技术学院
11
分子生物学(胡凯) 2015-2-10
3. DNA复制的起始
从头起始与共价延伸: 1. 生物大多是在 DNA 分子特定的新的位置起 始,合成从RNA引物开始。
2. D- 环式复制方式(线粒体双链环状 DNA : 两条链的复制起点不同位置,且复制不同步) 3.共价延伸起始也叫单向滚环式复制(噬菌体 X174DNA—单链环状),环状分子产生一个 多联体。
生命科学与技术学院
4
分子生物学(胡凯) 2015-2-10
半保留复制 实验依据
1958年 M.Messelson 等 用实验加以证实
双螺旋结构是 半保留复制的 分子基础
生命科学与技术学院
5
分子生物学(胡凯) 2015-2-10
半保留复制的意义
复制的这种 方式可保证亲代 的遗传特征完整 无误的传递给子 代,体现了遗传 的保守性。
G A A G T C C G G C G
21
5'
生命科学与技术学院
分子生物学(胡凯) 2015-2-10
DNA聚合酶的“校对”(proofreading)作用 复制的保真性 ( fidelity ) 主要依赖 3 种机制:
• 聚合酶对碱基的选择; • 3′→ 5′方向的外切核酸酶活性起校正作用; • 复制后错配现象的特异性修复机制。