分子生物学的研究方法基因定点诱变课件
合集下载
第七章 定点诱变

3. 重叠延伸PCR诱变法 Overlap extension
对于两个具有部分重叠序列的 DNA 片段,在 经过变性和复性后,两个 DNA 片段之间通过 同源序列形成部分杂合的双链,在 DNA 聚合 酶作用下,杂合双链可互为引物和模板,引 导 DNA 的合成,从而形成杂合 DNA 双链
该方法需要4个引物,
再用通用引物扩增诱变的全长的DNA
5’
3’
3’
5’
4. 重叠延伸剪接法
Splicing by overlap extension, SOE
可用于将两个 DNA 片段在所期望的位点进行连接。 核心:设计一对融合寡核苷酸引物
包括两个步骤:设计一个寡聚体引物,其序列包含两个 部分,“引发”和“重叠”部分,引发部分在 3’ 末端, 起引物作用;重叠部分在 5’ 末端。与待融合的 DNA 片 段序列互补。
(二)位点选择诱变 Altered Sites in vitro Mutagenesis System 使用了2个寡核苷酸引物 一个是用来引入突变的突变引物 一个是用来选择用的
选择性引物可用来恢复有缺陷的抗生素抗 性基因,同时可用来选择引入突变的DNA链
1.将目的片段克隆到载体 2.得到重组质粒 3.模板DNA碱变性,与突变引物、氨苄青霉素抗性 基因修复引物(amproligo)和四环素抗性基因敲除 引物(tetsoligo)同时退火 4.用T4 DNA聚合酶和T4 DNA连接酶合成突变链 5.转化大肠杆菌ES1301菌株(mutS),用氨苄青霉 素筛选 6.纯化质粒,转化大肠杆菌JM109,用氨苄青霉素 筛选 7.必要的话,影印一份平板,鉴定四环素敏感的克 隆子,从而获得突变体 8.再做一轮诱变,修复四环素抗性基因并敲除氨苄 青霉素抗性基因
《定点诱变技术》课件

《定点诱变技术》PPT课 件
本课程将介绍定点诱变技术的原理、实验步骤、应用案例和未来展望,以及 它对人类社会的影响和挑战。
概述
什么是定点诱变技术?
定点诱变技术是一种精准编辑基因术?
传统的基因编辑技术往往是非特异性的,不能达到精准编辑的效果,而定点诱变技术可以 避免对非目标区域的影响。
实验步骤
1
实验前的准备工作
首先需要构建引导RNA分子、购买
细胞培养与转染
2
Cas9核酸酶,以及筛选细胞线等。这 些都需要提前准备。
将引物与Cas9,利用转染技术导入到
目标细胞中,完成复合物的构建。
3
PCR扩增和测序
利用PCR扩增技术检测目标基因组位
置是否发生了修饰,并对其进行测序,
数据分析和结果解读
定点诱变技术的应用领域
定点诱变技术可以应用于基础科研、农业生产、医疗诊断等领域,为人类社会带来了诸多 益处。
基本原理
基于CRISPR-Cas9技术实 现的定点诱变
CRISPR-Cas9技术可以在基因 组中精准识别目标区域,并将 核酸酶Cas9导入到目标位置, 再通过引导RNA分子的作用, 完成对基因组上的目标位点的 修饰。
定点诱变技术可能会带来一系 列的伦理、法律和社会问题, 例如导致人们撕裂不和,社会 不公等问题。
定点诱变技术未来发展 的瓶颈和解决方案
目前,定点诱变技术在某些肿 瘤细胞和人类胚胎细胞中并没 有得到广泛应用,需要进一步 研究,探索更为高效、精准、 安全的定点诱变技术。
4
验证修饰深度和准确性。
对PCR扩增和测序的数据进行分析, 解读实验结果,得出结论,为下一步
的研究提供指导。
应用案例
定点诱变技术在基因 修复中的应用
本课程将介绍定点诱变技术的原理、实验步骤、应用案例和未来展望,以及 它对人类社会的影响和挑战。
概述
什么是定点诱变技术?
定点诱变技术是一种精准编辑基因术?
传统的基因编辑技术往往是非特异性的,不能达到精准编辑的效果,而定点诱变技术可以 避免对非目标区域的影响。
实验步骤
1
实验前的准备工作
首先需要构建引导RNA分子、购买
细胞培养与转染
2
Cas9核酸酶,以及筛选细胞线等。这 些都需要提前准备。
将引物与Cas9,利用转染技术导入到
目标细胞中,完成复合物的构建。
3
PCR扩增和测序
利用PCR扩增技术检测目标基因组位
置是否发生了修饰,并对其进行测序,
数据分析和结果解读
定点诱变技术的应用领域
定点诱变技术可以应用于基础科研、农业生产、医疗诊断等领域,为人类社会带来了诸多 益处。
基本原理
基于CRISPR-Cas9技术实 现的定点诱变
CRISPR-Cas9技术可以在基因 组中精准识别目标区域,并将 核酸酶Cas9导入到目标位置, 再通过引导RNA分子的作用, 完成对基因组上的目标位点的 修饰。
定点诱变技术可能会带来一系 列的伦理、法律和社会问题, 例如导致人们撕裂不和,社会 不公等问题。
定点诱变技术未来发展 的瓶颈和解决方案
目前,定点诱变技术在某些肿 瘤细胞和人类胚胎细胞中并没 有得到广泛应用,需要进一步 研究,探索更为高效、精准、 安全的定点诱变技术。
4
验证修饰深度和准确性。
对PCR扩增和测序的数据进行分析, 解读实验结果,得出结论,为下一步
的研究提供指导。
应用案例
定点诱变技术在基因 修复中的应用
分子生物学研究方法和技术(共92张PPT)

结构生物学(Structural Biology)
生物大分子的高级三维结构与功能的统一
生物大分子之间的互作
分子发育生物学(Molecular Developing Biology)
基因表达,基因互作
器官发生 胚胎形成
个体发育
分子细胞生物学 (Molecular Cell Biology)
个体
细胞分化
分子生物学是研究核酸、蛋白质等生物大分子 的形态、结构特征及其重要性、规律性和相互关 系的科学,是人类从分子水平上真正揭示生物世 界的奥秘,由被动地适应自然界转向主动地改造 和重组自然界的基础学科。
分子生物学与生物化学之间的关系与区别
分子生物学—研究生物大分子的结构、功能及信 息传递过程,从分子水平研究生命现象。
(二)核蛋白的解聚、变性蛋白的去除
核酸与蛋白质的结合力主要是正负静电吸力 (核酸与碱性蛋白的结合)、氢键和非极性的范 德华力。
分离核酸最困难的是将与核酸紧密结合的蛋 白质分开,同时避免核酸降解。
常用方法:
1. 加入浓盐溶液(如NaCl)
核酸-蛋白质加入NaCl后,破坏静电吸力,使
氢键破坏,核蛋白解聚;
2. RNA酶(RNAase)抑制剂
RNAase分布广泛,极易污染样品,而且耐高温、耐酸、耐 碱,不宜失活。
(1) 皂土(bentonite ) 作用机制:皂土带负电荷,能吸附RNase,使其失活。
(2) DEPC(二乙基焦碳酸盐) (C2H5OCOOCOOC2H5)
粘性液体,很强的核酸酶抑制剂。
基因的概念
基因的结构
结
基因工程
基因的复制
构
细胞工程
基因的表达 基因的重组
生 物 学
分子生物学--定点诱变与蛋白质工程课件

❖ 枯草杆菌蛋白酶可作为洗涤剂的添加剂,但由于其只能水解 苯丙氨酸(Phe)羧基所形成的肽键,底物作用范围过窄而限 制了洗涤剂的高效性,若用带正电荷的赖氨酸(Lys)取代位 于活性中心166位的甘氨酸(Gly),所获得的突变酶不仅能水 解苯丙氨酸(Phe)羧基所形成的肽键,而且可以水解酸性氨 基酸谷氨酸(Glu)所形成的肽键,使其底物作用范围拓宽, 因而可能成为最高效的洗涤剂添加酶,这是定点诱变改变蛋 白质生物学活性的成功例子。
❖ N端-----------笨丙-天冬-----------谷-甘-------------- C端 多肽链
三、定点诱变的原理
定点诱变原理示意图
❖ The Nobel Prize in Chemistry 1993
"for contributions to the developments of methods within DNA-based chemistry"
2.诱变过程
1)合成含有目的 基因的正链DNA; 2)合成含有特殊 突变碱基的引物; 3)制备异源双链 DNA(右图); 4)富集和转化双 链DNA分子; 5)筛选突变体并鉴定
(二)Kunkel定点诱变法
❖ 体外DNA合成往往是不完全的,所以部分合 成的DNA分子必须通过蔗糖密度梯度离心除 去,获得纯化的突变DNA。
❖ 理论上来说,DNA是半保留复制的,应用寡 核苷酸定点诱变时,所形成的噬菌体中携带 突变基因的应为一半。但实际上,由于技术 上的原因通常只有1-5%的噬菌斑含有突变基 因的噬菌体。
❖ 因此,为了获得更多含有突变噬菌体的噬菌 斑,必须提高突变体的比率。目前已有多种 改良的寡核苷酸定点诱变方法,此处将 Kunkel 1985年建立的方法做以简单介绍。
❖ N端-----------笨丙-天冬-----------谷-甘-------------- C端 多肽链
三、定点诱变的原理
定点诱变原理示意图
❖ The Nobel Prize in Chemistry 1993
"for contributions to the developments of methods within DNA-based chemistry"
2.诱变过程
1)合成含有目的 基因的正链DNA; 2)合成含有特殊 突变碱基的引物; 3)制备异源双链 DNA(右图); 4)富集和转化双 链DNA分子; 5)筛选突变体并鉴定
(二)Kunkel定点诱变法
❖ 体外DNA合成往往是不完全的,所以部分合 成的DNA分子必须通过蔗糖密度梯度离心除 去,获得纯化的突变DNA。
❖ 理论上来说,DNA是半保留复制的,应用寡 核苷酸定点诱变时,所形成的噬菌体中携带 突变基因的应为一半。但实际上,由于技术 上的原因通常只有1-5%的噬菌斑含有突变基 因的噬菌体。
❖ 因此,为了获得更多含有突变噬菌体的噬菌 斑,必须提高突变体的比率。目前已有多种 改良的寡核苷酸定点诱变方法,此处将 Kunkel 1985年建立的方法做以简单介绍。
基因工程第七章DNA定点诱变

• 3’端所形成的杂交体足以引导DNA合成。如果
错配核苷酸太靠近3’端,3’端将不能形成稳定的 杂交体,易被外切活性降解。所以3’端需有79bp完全配对;
• 为便于筛选,应选用可形成稳定杂交体而长
度最短的诱变寡核苷酸。一般17-19bp, 错配在 中央,使得完全配对的杂交体与错配杂交体 之间的热稳定性差异足够大。
位点选择定点诱变法
三、Transformer SiteDirected mutagenesis
转化子诱变法
Synthesize second strand
Digest DNA (primary digestion)
Transform E.coli mutS to propagate plasmids
ATG ATG
ATG ATG
BamHI
ATG
BamHI
PCR ATG
ATG
EcoRI
PCR
ATG
EcoRI
重叠延伸
BamHI
ATG
PCR EcoRI
第二节 嵌套缺失
第二节 嵌套缺失
1. 外切核酸酶III的消化
Exonuclease III 5‘ 3‘
5‘ 3‘
5‘ 3‘
2. BAL 31的消化
DNA ligase
U
U
U
U
U
2.原理
Insert target DNA
转化 E.coli CJ236
U
U
U U
U
U
Isolate phagemid ss DNA 与突变寡核苷酸退火
转化E.coli MV1190 (dut+/ung+)
UU
基因定点诱变
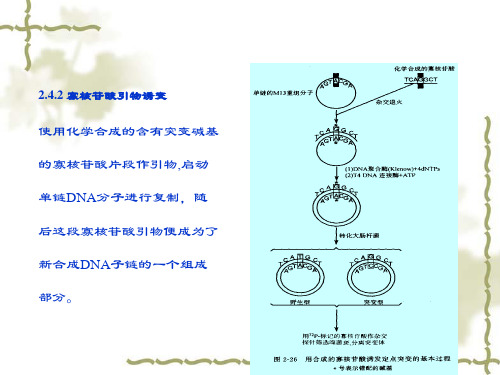
2.4.2 寡核苷酸引物诱变
使用化学合成的含有突变碱基 的寡核苷酸片段作引物,启动 单链DNA分子进行复制,随 后这段寡核苷酸引物便成为了 新合成DNA子链的一个组成 部分。
寡核苷酸引物诱变过程
正链DNA合成 突变引物合成 异源双链DNA分子制备 闭环异源双链DNA分子富集 转化 突变体筛选
寡核苷酸引物诱变法的局限性
异源双链DNA分子并非真正异源 突变体子代中突变碱基被错配修 复体系修复
提高寡核苷酸引物诱变突变效率的方法
(1) Kunkel 定点诱变法: 1985年由
(2) 硫代磷酸诱变法: 已知有些限制酶不能切割 硫代磷酸DNA分子.在异源双链DNA分子 制备时,加入硫代核苷酸,使之掺入突变 链中.然后用前述酶切割,并用外切酶局 部消化后,进行聚合反应,从而产生具有 定点突变的异源双链DNA
2.4.3 PCR诱变
(1) 重组PCR定点诱变:克服寡 核苷酸引物诱变能力仅限于5’ 端.
特点:经3轮PCR反应,一对互补 的带有突变碱基的内侧引物和 2个外侧引物
(2) 大引物诱变:核心是第一 轮PCR产物为第二轮PCR引 物
返回目录
返回第二章
使用化学合成的含有突变碱基 的寡核苷酸片段作引物,启动 单链DNA分子进行复制,随 后这段寡核苷酸引物便成为了 新合成DNA子链的一个组成 部分。
寡核苷酸引物诱变过程
正链DNA合成 突变引物合成 异源双链DNA分子制备 闭环异源双链DNA分子富集 转化 突变体筛选
寡核苷酸引物诱变法的局限性
异源双链DNA分子并非真正异源 突变体子代中突变碱基被错配修 复体系修复
提高寡核苷酸引物诱变突变效率的方法
(1) Kunkel 定点诱变法: 1985年由
(2) 硫代磷酸诱变法: 已知有些限制酶不能切割 硫代磷酸DNA分子.在异源双链DNA分子 制备时,加入硫代核苷酸,使之掺入突变 链中.然后用前述酶切割,并用外切酶局 部消化后,进行聚合反应,从而产生具有 定点突变的异源双链DNA
2.4.3 PCR诱变
(1) 重组PCR定点诱变:克服寡 核苷酸引物诱变能力仅限于5’ 端.
特点:经3轮PCR反应,一对互补 的带有突变碱基的内侧引物和 2个外侧引物
(2) 大引物诱变:核心是第一 轮PCR产物为第二轮PCR引 物
返回目录
返回第二章
《分子生物学研究法》PPT课件

解决办法:
1 用T4 DNA聚合酶除去PCR产物3’端多余的 核苷酸,成为平端后连接。
2 给平端载体的两个3’端各加上一个T,使 其与3’A突出的PCR产物互补,然后连接。
PCR产物与载体的A-T连接
mRNA
差 别 显 示
利 用
PCR
的
定
点
引物1或引物2的 5’端有不与模板
突
互补的突变。
变
利用PCR的定点突变
在循环温度的设定中,最重要的是退火温度。 退火温度较低可提高扩增效率,但产物的特异性较差; 退火温度较高可提高产物的特异性,但扩增效率较低。
嵌套PCR
( nested PCR)
嵌套PCR主要是为了提 高扩增产物的特异性。
嵌套PCR提高产物特异性的原理
单用第一对引物扩增,得一特异性产物和可能存在的非特异性产物。
循环条件的设定
变性---------→退火----------→链延伸
循环条件举例
循环 首轮循环 中间循环 末轮循环
变性 94℃ 5 min 94℃ 1 min 94℃ 1 min
退火 50℃ 2 min 50℃ 2 min 50℃ 2 min
链延伸 72℃ 3 min 72℃ 3 min 72℃10min
荧光定量PCR
1995年美国PerKin Elmer公司研制成功具有 革 命 性 意 义 的 荧 光 定 量 PCR 技 术 , 它 融 合 了 PCR高灵敏性、DNA杂交的高特异性和光谱技 术的高精确定量等优点。荧光定量PCR是直接 探测PCR过程中荧光信号的变化以获得定量的 结 果 , 不 需 要 PCR 后 处 理 或 电 泳 检 测 , 完 全 闭 管操作,在整个过程中,仅有加入样品的一次开 盖。
1 用T4 DNA聚合酶除去PCR产物3’端多余的 核苷酸,成为平端后连接。
2 给平端载体的两个3’端各加上一个T,使 其与3’A突出的PCR产物互补,然后连接。
PCR产物与载体的A-T连接
mRNA
差 别 显 示
利 用
PCR
的
定
点
引物1或引物2的 5’端有不与模板
突
互补的突变。
变
利用PCR的定点突变
在循环温度的设定中,最重要的是退火温度。 退火温度较低可提高扩增效率,但产物的特异性较差; 退火温度较高可提高产物的特异性,但扩增效率较低。
嵌套PCR
( nested PCR)
嵌套PCR主要是为了提 高扩增产物的特异性。
嵌套PCR提高产物特异性的原理
单用第一对引物扩增,得一特异性产物和可能存在的非特异性产物。
循环条件的设定
变性---------→退火----------→链延伸
循环条件举例
循环 首轮循环 中间循环 末轮循环
变性 94℃ 5 min 94℃ 1 min 94℃ 1 min
退火 50℃ 2 min 50℃ 2 min 50℃ 2 min
链延伸 72℃ 3 min 72℃ 3 min 72℃10min
荧光定量PCR
1995年美国PerKin Elmer公司研制成功具有 革 命 性 意 义 的 荧 光 定 量 PCR 技 术 , 它 融 合 了 PCR高灵敏性、DNA杂交的高特异性和光谱技 术的高精确定量等优点。荧光定量PCR是直接 探测PCR过程中荧光信号的变化以获得定量的 结 果 , 不 需 要 PCR 后 处 理 或 电 泳 检 测 , 完 全 闭 管操作,在整个过程中,仅有加入样品的一次开 盖。
《基因的定点突变》课件

基因定点突变技术是基因工程技 术的重要组成部分,广泛应用于 生物医学、农业、工业等领域。
基因定点突变的意义
基因定点突变有助于深入了解基因的 结构和功能,揭示生命活动的本质和 规律。
通过基因定点突变技术,可以实现对 特定基因的精确调控,为疾病治疗、 新药研发、生物育种等领域提供有力 支持。
基因定点突变的分类
筛选方法包括抗生素筛选、PCR鉴定 等,根据实验需求选择合适的方法。
04 基因定点突变的实验结果分析
突变基因的鉴定
01
02
03
突变基因的识别
通过DNA测序技术,对突 变基因进行精确的识别和 定位。
突变类型的分类
根据突变碱基的数量,将 突变基因分为点突变、插 入和删除突变等类型。
突变位点的统计
统计突变位点的分布和频 率,分析突变热点和区域 。
根据突变的方式,基因定点突变可分为碱基替换突变和插入/ 缺失突变。
根据突变的目的,基因定点突变可分为正向突变和反向突变 。正向突变是指将野生型基因突变为功能变异型基因,而反 向突变是指将功能变异型基因突变为野生型基因。
02 基因定点突变的方法
逆转录酶法
总结词
逆转录酶法是一种利用逆转录酶将RNA转化为cDNA的方法,适用于基因的定 点突变。
03
引物长度一般在1530bp之间,以保证足够 的特异性。
04
引物设计还需考虑突变 后可能的遗传信息改变 ,如密码子改变等。
合成突变基因
在成功设计引物后, 需通过DNA合成仪 合成突变基因。
合成后的突变基因需 进行质量检测,确保 其准确性。
合成过程中需确保突 变位点的准确性,避 免其他位点的突变。
锌指核酸酶法
总结词
基因定点突变的意义
基因定点突变有助于深入了解基因的 结构和功能,揭示生命活动的本质和 规律。
通过基因定点突变技术,可以实现对 特定基因的精确调控,为疾病治疗、 新药研发、生物育种等领域提供有力 支持。
基因定点突变的分类
筛选方法包括抗生素筛选、PCR鉴定 等,根据实验需求选择合适的方法。
04 基因定点突变的实验结果分析
突变基因的鉴定
01
02
03
突变基因的识别
通过DNA测序技术,对突 变基因进行精确的识别和 定位。
突变类型的分类
根据突变碱基的数量,将 突变基因分为点突变、插 入和删除突变等类型。
突变位点的统计
统计突变位点的分布和频 率,分析突变热点和区域 。
根据突变的方式,基因定点突变可分为碱基替换突变和插入/ 缺失突变。
根据突变的目的,基因定点突变可分为正向突变和反向突变 。正向突变是指将野生型基因突变为功能变异型基因,而反 向突变是指将功能变异型基因突变为野生型基因。
02 基因定点突变的方法
逆转录酶法
总结词
逆转录酶法是一种利用逆转录酶将RNA转化为cDNA的方法,适用于基因的定 点突变。
03
引物长度一般在1530bp之间,以保证足够 的特异性。
04
引物设计还需考虑突变 后可能的遗传信息改变 ,如密码子改变等。
合成突变基因
在成功设计引物后, 需通过DNA合成仪 合成突变基因。
合成后的突变基因需 进行质量检测,确保 其准确性。
合成过程中需确保突 变位点的准确性,避 免其他位点的突变。
锌指核酸酶法
总结词
分子生物学原理--基因表达调控 PPT课件

• 无阻遏物时,O区开放让酶通过并转录 下游的结构基因
• 有阻遏物时酶就不能通过
2021/3/28
分子生物学原理
二、操纵子的结构与功能
• 阻遏物基因 : 产生阻遏物,位于离操纵 子较远的上游区。
• 负调控:起调控作用的蛋白质分子抑制 转录
关闭的基因由代谢底物开放(诱导)-----阻遏物失活
开放的基因由代谢底物关闭(阻遏)-----阻遏物激活
i 基 因 产物 加 通常开放 合成代谢 上代谢终产物 由 代谢 终 产
物关闭
2021/3/28
分子生物学原理
三、乳糖操纵子和色氨酸操纵子
2021/3/28
分子生物学原理
乳糖操纵子
• 操纵子的三个结构基因为-半乳糖苷酶、 -半乳糖苷通透酶和-半乳糖苷乙酰转移 酶。
• 在无乳糖时,阻遏蛋白与O区结合,阻 止RNA聚合酶的转录
RNA聚合酶II的转录因子
转录因子 TFIIA TFIIB TFIID TFIIE TFIIF TFII-I
功能 稳定 TFIID 促进 RNA pol II 结合 辨认 TATA ATPase 解旋酶 促进 TFIID 结合
2021/3/28
分子生物学原理
转录前起始复合物
• TFIID是唯一与 DNA特异位点即 TATA盒结合的转 录因子。
2021/3/28
分子生物学原理
四、cAMP对转录的调控
• 培养基中有葡萄糖时:葡萄糖代谢引起 细胞内cAMP水平下降,乳糖操纵子基因 关闭。
• 培养基中葡萄糖不足时: cAMP水平升 高, cAMP-CAP复合物生成, cAMP使 CAP变构,而与CAP位点结合,促进乳 糖操纵子基因的转录,以便细胞利用乳 糖。
• 有阻遏物时酶就不能通过
2021/3/28
分子生物学原理
二、操纵子的结构与功能
• 阻遏物基因 : 产生阻遏物,位于离操纵 子较远的上游区。
• 负调控:起调控作用的蛋白质分子抑制 转录
关闭的基因由代谢底物开放(诱导)-----阻遏物失活
开放的基因由代谢底物关闭(阻遏)-----阻遏物激活
i 基 因 产物 加 通常开放 合成代谢 上代谢终产物 由 代谢 终 产
物关闭
2021/3/28
分子生物学原理
三、乳糖操纵子和色氨酸操纵子
2021/3/28
分子生物学原理
乳糖操纵子
• 操纵子的三个结构基因为-半乳糖苷酶、 -半乳糖苷通透酶和-半乳糖苷乙酰转移 酶。
• 在无乳糖时,阻遏蛋白与O区结合,阻 止RNA聚合酶的转录
RNA聚合酶II的转录因子
转录因子 TFIIA TFIIB TFIID TFIIE TFIIF TFII-I
功能 稳定 TFIID 促进 RNA pol II 结合 辨认 TATA ATPase 解旋酶 促进 TFIID 结合
2021/3/28
分子生物学原理
转录前起始复合物
• TFIID是唯一与 DNA特异位点即 TATA盒结合的转 录因子。
2021/3/28
分子生物学原理
四、cAMP对转录的调控
• 培养基中有葡萄糖时:葡萄糖代谢引起 细胞内cAMP水平下降,乳糖操纵子基因 关闭。
• 培养基中葡萄糖不足时: cAMP水平升 高, cAMP-CAP复合物生成, cAMP使 CAP变构,而与CAP位点结合,促进乳 糖操纵子基因的转录,以便细胞利用乳 糖。
基因工程8-DNA诱变ppt课件

error-prone PCR的实质:通过改变PCR反应条件来调整PCR 反应中突变的频率,降低聚合酶固有的突变序列倾向性,提高 突变谱的多样性,使得错误碱基随机地以一定的频率掺入到扩 增的基因中,从而得到随机突变的DNA群体,最后用合适的 载体克隆突变基因。
易错PCR采用的方法:
• 增加MgCl2浓度到7mM,稳定非互补的碱基配对; • 加入0.5mM的MnCl2,Mn 2+能降低聚合酶对模板的特异性; • 增加聚合酶量到5U,促使在错配碱基处继续延伸反应; • 限定4种碱基中的一种,通常为正常浓度的1-10%,在缺乏正 确核苷酸时,DNA聚合酶经短暂停顿后,会插入另外3种可用 核苷酸的一种; • 3种为正常浓度的正常碱基,第四种为次黄嘌呤dITP(可与C、 T、A配对); • 增加dCTP和dTTP的浓度到1mM,促进错误掺入; • 使用突变DNA聚合酶,如Mutazyme。
在体外用诱变引物合成的杂合双链DNA在导入正常ung+ dut+菌 株后,只有带有突变位点的新合成链和作模板进一步复制,而 野生型不能复制。这样能够生长的细胞就带有突变位点。
(二)位点选择诱变
(altered sites in vitro mutagenesis)
位点选择诱变法使用了2个寡核苷酸引物,一个是用来引入突变 的引物,另一个是用来选择用的。选择性引物可用来恢复有缺陷 的抗生素抗性基因,同时可用来选择引入突变的DNA链。
特点:可正向选择突变链,使用高保真的T4DNA聚合酶合成 DNA,可以使用双良DNA作模板,可进行多轮筛选。但需特定载 体,即含有一个有缺陷的抗生素抗性基因。
(三)转化子诱变
(transformer site-directed mutagenesis)
易错PCR采用的方法:
• 增加MgCl2浓度到7mM,稳定非互补的碱基配对; • 加入0.5mM的MnCl2,Mn 2+能降低聚合酶对模板的特异性; • 增加聚合酶量到5U,促使在错配碱基处继续延伸反应; • 限定4种碱基中的一种,通常为正常浓度的1-10%,在缺乏正 确核苷酸时,DNA聚合酶经短暂停顿后,会插入另外3种可用 核苷酸的一种; • 3种为正常浓度的正常碱基,第四种为次黄嘌呤dITP(可与C、 T、A配对); • 增加dCTP和dTTP的浓度到1mM,促进错误掺入; • 使用突变DNA聚合酶,如Mutazyme。
在体外用诱变引物合成的杂合双链DNA在导入正常ung+ dut+菌 株后,只有带有突变位点的新合成链和作模板进一步复制,而 野生型不能复制。这样能够生长的细胞就带有突变位点。
(二)位点选择诱变
(altered sites in vitro mutagenesis)
位点选择诱变法使用了2个寡核苷酸引物,一个是用来引入突变 的引物,另一个是用来选择用的。选择性引物可用来恢复有缺陷 的抗生素抗性基因,同时可用来选择引入突变的DNA链。
特点:可正向选择突变链,使用高保真的T4DNA聚合酶合成 DNA,可以使用双良DNA作模板,可进行多轮筛选。但需特定载 体,即含有一个有缺陷的抗生素抗性基因。
(三)转化子诱变
(transformer site-directed mutagenesis)
05分子生物学研究法99页PPT

PCR三步曲
变性 90~97℃ 退火 45~55℃ 延伸 72℃
变变 90变95变
70变75变
变变
PCR
变1) 将mRNA反转录为双链DNA. (2) 特点: A.稳定者组织mRNA中所含的全部或绝大部 分遗传信息.
名称
检测对象
探针
检测原理 处理过程
用途
Southern blot
DNA
标记单链核 酸
核酸复性中 碱基配对专
一性
琼脂糖电泳 后转膜
基因检测 (拷贝数)
Northern blot
RNA
标记单链核 酸
核酸复性中 碱基配对专
一性
变性琼脂糖 电泳后转膜
基因表达的 检测(表达
量)
Hale Waihona Puke Western blot蛋白
抗体
加标准分子量DNA Marker,测定分子量 加标准浓度的DNA,测定浓度
琼脂糖凝胶电泳技术要点
1.不同浓度凝胶分辨DNA片段能力不一样 (p33) (1)低: 不利于小片段的分辨(扩散) (2)高: 不利于大片段的分辨(迁移不动) (3) 一般选1.0%
2. 凝胶要用缓冲液配( TBE 或 TAE ) 3. EB强致癌,带手套操作; 4. agarose (琼脂糖)---电泳
3.SDS-PAGE测定的是单体蛋白分子量;(多亚基不行) 4、SDS-PAGE不适于:
(1)电荷异常蛋白:组蛋白(+电多) (2)构象异常的 蛋白 (3)带有较大辅基的蛋白--糖蛋白,脂蛋白。
PCR技术
1、原理 2、应用 3、技术要点
PCR技术原理
5’
3’
3’
5’
循环过程(32 次左右) 1.解链:94 ℃, 30 s 2.引物结合: ? ℃ ,30 s 3. 延伸:72 ℃,?Min
分子生物学-研究方法(上)PPT课件

.
5
基因工程技术区别于其它技术的根本特征:具 有跨越天然物种屏障、把来自任何生物的基因 置于毫无亲缘关系的新的寄主生物细胞之中的 能力。
1962年Arber 发现限制性核酸内切酶, 1967年Gellert发现了 DNA 连接酶。
.
6
酶类
重组DNA实验中常见的主要工具酶
功能
限制性核酸内切酶 DNA连接酶
分离DNA片段的大小范围(bp) 50 000~1 000 20 000~1 000 6 000~300 1 000~100 500~25 50~1
凝胶的分辨能力同凝胶类型和浓度有关,凝胶
浓度的高低影响凝胶介质孔隙的大小,浓度越高,
孔隙越小,其分辨能力就越强。反之,浓度降低,
孔隙就增大,其分辨能力也就随之减弱。
.
42
(3)PCR的应用
• 反向PCR:扩增已知序列旁侧的未知DNA序列
• 锚定PCR: 用于测定基因结构
• 不对称PCR: 用于扩增某一单链模板
• RT-PCR(逆转录-PCR): 逆转录联合PCR以扩增cDNA
• 复合PCR: 用多对引物同时扩增几条DNA片段
• 易错PCR:引入错配碱基,导致随机突变
• 荧光定量PCR: 用于估计模板DNA的浓度
• 免疫PCR:扩增抗原-抗体复合物上的DNA分子
• 原位PCR:在组织或细胞标本片上直接进行PCR
• 微生物PCR: 用于微生物的鉴定和分类
.
43
➢ 设计引物应遵循以下原则:
①引物长度:15-30bp,常为20nt左右。 ②引物扩增跨度:以200-500bp为宜。 ③引物碱基:G+C含量以40-60%为宜。避免5个以上 相同碱基的成串排列。 ④引物内部应避免出现出现二级结构,避免两条引 物间互补。 ⑤引物3ˊ末端碱基,应与模板单链严格配对。 ⑥引物中具有或能够加上合适的酶切位点。 ⑦特异性:与数据库中的其它序列无明显同源性。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
(1) Kunkel 定点诱变法: 1985年由 T.A.Kunkel发明
(2) ung:尿嘧啶脱糖苷酶
分子生物学的研究方法基因定点诱变
4
(2) 硫代磷酸诱变法: 已知有些限制 酶不能切割硫代磷酸DNA分 子.在异源双链DNA分子制备 时,加入硫代核苷酸,使之掺 入突变链中.然后用前述酶切 割,并用外切酶局部消化后, 进行聚合反应,从而产生具有 定点突变的异源
5
2.4.3 PCR诱变
(1) 重组PCR定点诱 变:克服寡核苷酸引 物诱变能力仅限于5’ 端.
特点:经3轮PCR反 应,一对互补的带有 突变碱基的内侧引 物和2个外侧引物
分子生物学的研究方法基因定点诱变
6
(2) 大引物诱变:核心 是第一轮PCR产物为 第二轮PCR引物
分子生物学的研究方法基因定点诱变
2
寡核苷酸引物诱变过程
正链DNA合成 突变引物合成 异源双链DNA分子制备 闭环异源双链DNA分子富集 转化 突变体筛选
寡核苷酸引物诱变法的局限性
异源双链DNA分子并非真正异源
突变体子代中突变碱基被错配修 复体系修复
分子生物学的研究方法基因定点诱变
3
提高寡核苷酸引物诱变突变效率的方法
No Image
分子生物学的研究方法基因定点诱变
7
返回目录
返回第二章
分子生物学的研究方法基因定点诱变
8
此课件下载可自行编辑修改,供参考! 感谢您的支持,我们努力做得更好!
分子生物学的研究方法基因定点诱变
9
返回目第录 四节 基因定点诱变
2.4.1 盒式诱变
利用一段人工合成 的具有突变序列的 寡核苷酸片段取代 野生型基因中的相 应序列.
分子生物学的研究方法基因定点诱变
返回第二章
1
2.4.2 寡核苷酸引物诱变
使用化学合成的含有突变碱基 的寡核苷酸片段作引物,启动 单链DNA分子进行复制,随 后这段寡核苷酸引物便成为了 新合成DNA子链的一个组成 部分。
(2) ung:尿嘧啶脱糖苷酶
分子生物学的研究方法基因定点诱变
4
(2) 硫代磷酸诱变法: 已知有些限制 酶不能切割硫代磷酸DNA分 子.在异源双链DNA分子制备 时,加入硫代核苷酸,使之掺 入突变链中.然后用前述酶切 割,并用外切酶局部消化后, 进行聚合反应,从而产生具有 定点突变的异源
5
2.4.3 PCR诱变
(1) 重组PCR定点诱 变:克服寡核苷酸引 物诱变能力仅限于5’ 端.
特点:经3轮PCR反 应,一对互补的带有 突变碱基的内侧引 物和2个外侧引物
分子生物学的研究方法基因定点诱变
6
(2) 大引物诱变:核心 是第一轮PCR产物为 第二轮PCR引物
分子生物学的研究方法基因定点诱变
2
寡核苷酸引物诱变过程
正链DNA合成 突变引物合成 异源双链DNA分子制备 闭环异源双链DNA分子富集 转化 突变体筛选
寡核苷酸引物诱变法的局限性
异源双链DNA分子并非真正异源
突变体子代中突变碱基被错配修 复体系修复
分子生物学的研究方法基因定点诱变
3
提高寡核苷酸引物诱变突变效率的方法
No Image
分子生物学的研究方法基因定点诱变
7
返回目录
返回第二章
分子生物学的研究方法基因定点诱变
8
此课件下载可自行编辑修改,供参考! 感谢您的支持,我们努力做得更好!
分子生物学的研究方法基因定点诱变
9
返回目第录 四节 基因定点诱变
2.4.1 盒式诱变
利用一段人工合成 的具有突变序列的 寡核苷酸片段取代 野生型基因中的相 应序列.
分子生物学的研究方法基因定点诱变
返回第二章
1
2.4.2 寡核苷酸引物诱变
使用化学合成的含有突变碱基 的寡核苷酸片段作引物,启动 单链DNA分子进行复制,随 后这段寡核苷酸引物便成为了 新合成DNA子链的一个组成 部分。